Embed Size (px)
Citation preview

Yeast 15, 1609–1618 (1999)
Transformation of Candida albicans by Electroporation
MARIANNE D. DE BACKER1*, DIRK MAES1, SANDY VANDONINCK1, MARC LOGGHE2,ROLAND CONTRERAS2 AND WALTER H. M. L. LUYTEN1
1Department of Advanced Biotechnologies, Janssen Research Foundation, Turnhoutseweg 30, B2340 Beerse, Belgium2Department of Fundamental and Applied Molecular Biology, University of Gent, K.L. Ledeganckstraat 35,B9000 Gent, Belgium
In contrast to a variety of other yeasts, Candida albicans has proved difficult to transform with high efficiency.Lithium acetate transformation is fast and simple but provides a very low efficiency of DNA transfer (50–100transformants/�g DNA), while spheroplast transformation, although more efficient (�300 transformants/�gintegrative DNA and 103–104 transformants/�g replicative DNA), is complicated and time-consuming. In this studywe applied various yeast transformation techniques to C. albicans and selected an electroporation procedure forfurther optimization. Transformation efficiencies of up to 300 transformants/�g were obtained for an integrativeplasmid and up to 4500 transformants/�g for a CARS-carrying plasmid. This reasonably high transformationefficiency, combined with the ease and speed of electroporation in comparison to alternative techniques, make it thepreferred method for transformation of C. albicans. Copyright � 1999 John Wiley & Sons, Ltd.
— Candida albicans; transformation; electroporation; homologous recombination; integrative vector;episomal vector
*Correspondence to: M. D. De Backer, Department of Ad-vanced Biotechnologies, Janssen Research Foundation, Turn-houtseweg 30, B2340 Beerse, Belgium. Tel: +32-14-60.38.81;fax: +32-14-60.61.11; e-mail: [email protected]/grant sponsor: Vlaams Instituut voor de Bevorderingvan het Wetenschappelijk-Technologisch Onderzoek in de In-
INTRODUCTION
Candida (C.) albicans is an important opportunis-tic pathogen of humans and belongs to the genusCandida, a group of yeasts united only by the factthat none has a known natural sexual cycle. ManyCandida species are diploid or aneuploid andtheir genetics and molecular biology have there-fore proved difficult to study. A prerequisite formolecular biological manipulation is, of course,a reliable and efficient means for introducingexogenous DNA into the cell. Lithium acetatetransformation (Ito et al., 1983) and spheroplasttransformation (Hinnen et al., 1978), both widelyused methods, suffer from significant limitations,especially when applied to C. albicans. Lithiumacetate transformation is fast and simple butgives low transformation efficiencies (50–100transformants/�g DNA) for C. albicans (Sanglard
dustrie (IWT), Belgium; Contract/grant number: 960192.
CCC 0749–503X/99/151609–10$17.50Copyright � 1999 John Wiley & Sons, Ltd.
et al., 1996). Spheroplast transformation,although more efficient (�300 transformants/�gintegrative DNA and 103–104 transformants/�greplicative DNA; Herreros et al., 1992), islaborious and time-consuming.
Development of alternative transformationmethods which do not rely on spheroplasts maysimplify genetic engineering in Candida. Whenusing the spheroplast PEG procedure, the riskremains of generating protoplast fusions, withresulting polypoidy. By far the fastest and simplestmethod of transformation is electroporation.Unlike the lithium acetate procedure, however,electroporation saturates at low DNA levels(Ausubel et al., 1992). Successful high-efficiencyelectroporation has been described for a variety ofyeasts, such as S. cerevisiae (Hill, 1989; Delorme,1989; Becker and Guarente, 1991; Manivasakamand Schiestl, 1993; Simon, 1993), Hansenula poly-morpha (Faber et al., 1993), C. maltosa (Kasuskeet al., 1992; Becher and Oliver, 1995) and C. utilis(Kondo et al., 1995).
However, C. albicans, unlike many other yeasts,has proved difficult to transform by electropora-
tion with high efficiency (Brown et al., 1996;Received 4 May 1999Accepted 12 July 1999

1610 M. D. DE BACKER ET AL.
Thompson et al., 1998). The most recent reportsdescribe transformation efficiencies for C. albicansof �250 transformants/�g with linearized integra-tive DNA and �3 transformants/�g with circularintegrative DNA (Thompson et al., 1998).
We compared various known yeast transforma-tion procedures and optimized one electroporationprocedure for C. albicans to yield transformationefficiencies of �250 transformants/�g for integra-tive circular DNA and �3000 transformants/�gfor a CARS-carrying plasmid. These transforma-tion efficiencies are comparable to those obtainedwith the more laborious spheroplast method andsuffice, for example, for efficient screening ofC. albicans libraries.
Electroporation thus offers an excellent alterna-tive to other, more time-consuming, transforma-tion methods. Due to its ease, speed and efficiencyin comparison to alternative techniques, it maywell become the preferred method for transforma-tion of C. albicans.
MATERIALS AND METHODS
Strains and plasmidsC. albicans CAI-4 (�ura3::imm434/�ura3::
imm434) was kindly provided by Dr WilliamFonzi, University of California, Irvine, USA(Fonzi and Irwin, 1993). The episomal vectorpRM2 used for transformation originates frompRM1 (provided by Professor C. Nombela,University of Madrid, Spain) by removal of the C.albicans LEU2 gene. The integrative vectorpGAL1PNiST-1 used for transformation haspGEM11Zf(+) (Promega Inc.) as a backbonewith an origin of replication for maintenance inEscherichia coli and a �-lactamase gene conferringresistance of E. coli to ampicillin, and contains a C.albicans URA3 gene (EMBL Accession No.X14196), a C. albicans galactokinase 1 promoter(EMBL Accession No. Y13580), a NotI–stuffer–SfiI cassette and a S. cerevisiae cytochrome ctranscription terminator (EMBL Accession No.M37696; nucleotides 588–857).
MediaYPD medium contains 2% peptone (Difco), 1%
yeast extract (Difco) and 2% glucose (Sigma);Synthetic selective medium contains 0·67% YeastNitrogen Base without amino acids (Difco), 0·2%Ura dropout powder (Bufferad, Inc) and 2%glucose.
Copyright � 1999 John Wiley & Sons, Ltd.
Nucleic acid isolationsPlasmid DNA was isolated using the Qiagen
tip-100 procedure (Qiagen). Genomic DNA of C.albicans was isolated using the Nucleon MiY Yeastkit (Amersham Life Sciences) and the concen-tration of genomic DNA was estimated by resolv-ing a sample on a 0·7% agarose gel in 0·5�TBEand comparing the intensity of the band withthat of a molecular weight marker (MWM II,Boehringer).
Lithium acetate transformationCAI-4 was transformed using the lithium acetate
method adapted from Gietz et al. (1995) andmodified as described by Sanglard et al. (1996).
Spheroplast transformationCAI-4 was transformed using the spheroplast
transformation method described by Beggs (1978)or a modified spheroplast transformation method(Logghe M., in preparation).
DNA-mediated transformation by electroporation
Growth and harvesting of yeast cellsFive �l of cells from a glycerol stock (�25�106
cells) were inoculated into 100 ml YPD supple-mented with 400 �l 0·1 uridine. The resultingculture was grown overnight at 30�C with vigorous(300 rpm) shaking. The OD600 was measured andcells were harvested at an optical density of �1·3.Cells were collected by centrifugation at 3500 rpmfor 10 min at ambient temperature and the result-ing cell pellet resuspended in 25 ml of 100 mlithium acetate/10 m dithiothreitol/TE and incu-bated at ambient temperature for 1 h. Cells werecollected by centrifugation at 3500 rpm for 5 minat 4�C and the resulting pellet was resuspended in25 ml ice-cold, high-quality water (MilliQ water,Millipore). This step was repeated once more andthe pellet was resuspended in 10 ml ice-cold 1 sorbitol. The mixture was centrifuged at 3500 rpmfor 5 min at 4�C. The supernatant was carefullyremoved and the pellet resuspended in 100 �l of1 sorbitol. This cell suspension was kept on icetill used.
ElectroporationFor each transformation, 40 �l cells was trans-
ferred to an electroporation cuvette (Eurogentec),DNA was added [typically 1 �g (in a maximum
Yeast 15, 1609–1618 (1999)

1611TRANSFORMATION OF CANDIDA ALBICANS BY ELECTROPORATION
volume of 2 �l) circular plasmid DNA] to the cellsuspension and the mixture was incubated on icefor 5 min.� Protocol 1 (Thompson et al., 1998): The cuvette
(0·2 cm) was placed in the electroporation cham-ber (Easyject Plus, Eurogentec) and an electricpulse of 1·5 kV, 25 �F, 201 � was applied.
� Protocol 2 (Equibio twin pulse, Eurogentec):The cuvette (0·2 cm) was placed in the electro-poration chamber (Easyject Plus, Eurogentec)and a first high-voltage electric pulse of 2·0 kV,25 �F, 99 � was applied, followed by a low-voltage second pulse of 0·15 kV, 2100 �F, 99 �.
For each set of experiments, an aliquot of cellsuspension was electroporated without prioraddition of DNA to control for possible contami-nation. Whenever this control yielded colonies,experiments were not taken into consideration forfurther analysis. After electroporation, the cuvettewas removed from the electroporation apparatus,1 ml ice-cold 1 sorbitol was added immediatelyand the contents of the cuvette were mixed byinversion.
PlatingTransformation mixes were plated on selective
synthetic medium containing 1 sorbitol (AcrosOrganics Belgium) and 2% agar (Difco) usingsterile 0·4 cm glass beads (Glaverbel, Belgium) andincubated for 3–5 days at 30�C.
Preparation of C. albicans spheroplasts prior toelectroporation
Cells were grown as described above andharvested by centrifugation at 3500 rpm for10 min at ambient temperature. Candida albicansspheroplasts were prepared as described athttp://www.invitrogen.com/pdf manuals/pichspherman.pdf in section 3·4. Spheroplasts were gentlyresuspended in 100 �l 1 sorbitol, placed on iceand used immediately for electroporation.
Southern blot analysisGenomic DNA (200 ng) was digested overnight
with EcoRI, treated with RNAse to removeresidual RNA and incubated for 20 min at 65�C toinactivate the enzymes. Electrophoresis and blot-ting were performed as described by Sambrooket al. (1989).
A 1·6 kb URA-blaster cassette-specific probefragment was amplified from pMB7 (Fonzi andIrwin, 1993) using primers P3 (5� CAGCAATT
Copyright � 1999 John Wiley & Sons, Ltd.
CTCGTGAATCATCG 3�) and P5 (5� TGATCAATTCCTGAGCAACAAC 3�). Amplification wasperformed in 30 cycles, each consisting of (a) 1 minat 94�C, (b) 1 min at 57�C, and (c) 2 min at 72�C. Inthe reaction mixture 5 units of Taq polymerase(Boehringer) were used, and the final concen-trations were 0·2 � of each primer, 2 m MgCl2and 200 � dNTPs. The URA-blaster cassette frag-ment (1·6 kb) was labelled with [32P]dCTP usingthe Rediprime� labelling system (Amersham). Theblot was hybridized overnight at 65�C in 5�SSPE, 5� Denhardt’s solution, 0·5% SDS, 100 �g/ml denatured herring sperm DNA (Clontech) and106 cpm/ml of probe. Post-hybridization, mem-branes were washed in 2� SSC, 0·1% SDS for 1 hat ambient temperature and in 1� SSC, 0·1% SDSfor 1 h at 65�C and the blot was exposed overnightto Fuji PhosphorImaging screen at ambient tem-perature. The screen was scanned using a STORMPhosphorImager (Molecular Dynamics) and theresulting image was visualized using ImageQuantsoftware (Molecular Dynamics).
Data analysisMeans and standard deviations were calculated
using Excel (Microsoft Office 97). Statisticalanalysis was performed by means of a two-tailedStudent’s t-test using Excel.
RESULTS
Transformation of C. albicans bynon-electroporation methods
The C. albicans ura3 mutant, CAI-4, was trans-formed with circular DNA from either an integra-tive (pGAL1PNiST-1) or an episomal (pRM2)vector using either the lithium acetate method, thespheroplast method or a modified spheroplastmethod (Logghe et al., in preparation). Thelithium acetate method is based on the observationthat alkali cations make yeast competent to takeup DNA. After incubation of the yeast in bufferedlithium acetate, the transforming DNA is intro-duced with high molecular weight carrier DNA.Addition of PEG and a heat shock trigger DNAuptake. For the spheroplast method, cells areprepared for transformation by enzymatic diges-tion of the cell wall, followed by several gentleisotonic washes to remove the digestive enzyme.DNA is mixed with the resulting spheroplasts andtreated with PEG to promote DNA uptake. Afterremoval of the PEG, spheroplasts and DNA are
Yeast 15, 1609–1618 (1999)

1612 M. D. DE BACKER ET AL.
resuspended in osmotically stabilizing regenerationmedium. Unlike S. cerevisiae spheroplasts, C.albicans spheroplasts can regenerate their cell wallon the surface of osmotically supported mediaand do not require agar overlay techniques (Plaet al., 1996). Resulting transformation efficiencies(number of transformants/�g) were calculatedand are shown in Table 1. In our hands, thelithium acetate method gives on average �1·3transformants/�g with integrative plasmid DNA(pGAL1PNiST-1; Figure 1) and �68transformants/�g with replicative plasmid DNA(pRM2; Figure 1). The spheroplast method gives amuch higher transformation efficiency of �400 or600 transformants/ �g for integrative and replica-tive plasmids, respectively. With the modifiedspheroplast method, again high numbers of trans-formants were obtained: �2000 transformants/�gwith integrative DNA (pGAL1PNiST-1) and�8000 transformants/�g with replicative DNA(pRM2).
The pGAL1PNiST-1 plasmid can integrate atthe homologous galactokinase 1 promoter site inthe Candida genome. Plasmid pRM2 contains twoCandida autonomous replicating sequences (ARS)(Herreros et al., 1992; Negredo et al., 1997) whichallow for (unstable) autonomous replication inC. albicans.
All transformants were re-streaked onto selec-tive medium in order to determine the fractionof abortive colonies. All transformants obtainedusing the lithium acetate procedure were able togrow upon re-streaking. For the modified sphero-
Copyright � 1999 John Wiley & Sons, Ltd.
plast method (Logghe et al., in preparation) 70–90% of the colonies were found to be abortive.
Table 1. Efficiency of three different (non-electroporation) transformation procedures for C. albi-cans CAI-4 using either integrative (pGAL1PNiST-1) orepisomal (pRM2) DNA. Results are means�SD forthree or more experiments.
VectorTransformation
method
Transformationefficiency(# tr./�g)
pGAL1PNiST-1 Lithium acetate 1·3�1·5 (n=3)Spheroplast 428�214 (n=3)Modified
spheroplast2015�205 (n=3)
pRM2 Lithium acetate 68�43 (n=3)Spheroplast 611�67 (n=4)Modified
spheroplast8050�763 (n=3)
Figure 1. Plasmid maps from pGAL1PNiST-1 (A) and pRM2(B). CaURA3, C. albicans URA3 gene; Pgal1, C. albicansgalactokinase 1 promoter; CYC T, S. cerevisiae cytochrome cterminator; ORI, origin of replication for E. coli; Apr, ampicil-lin resistance gene for E. coli; CARS 1/2 and 3, C. albicansautonomously replicating sequences.
Transformation of C. albicans by electroporationFor transformation by electroporation, yeast
cells are concentrated from a log-phase cultureusing multiple washes with sterile water to removeextracellular ions. The concentrated cells are sus-
pended in 1 sorbitol for osmotic stabilization.Yeast 15, 1609–1618 (1999)
1613TRANSFORMATION OF CANDIDA ALBICANS BY ELECTROPORATION
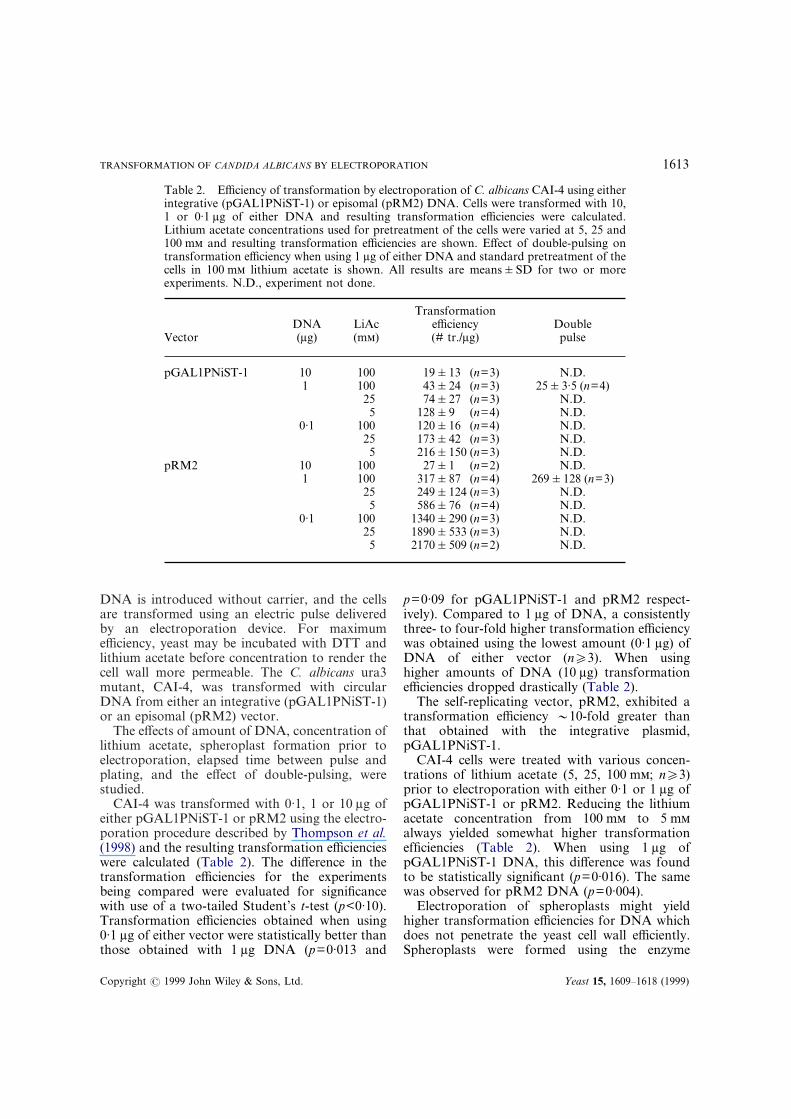
Table 2. Efficiency of transformation by electroporation of C. albicans CAI-4 using eitherintegrative (pGAL1PNiST-1) or episomal (pRM2) DNA. Cells were transformed with 10,1 or 0·1 �g of either DNA and resulting transformation efficiencies were calculated.Lithium acetate concentrations used for pretreatment of the cells were varied at 5, 25 and100 m and resulting transformation efficiencies are shown. Effect of double-pulsing ontransformation efficiency when using 1 �g of either DNA and standard pretreatment of thecells in 100 m lithium acetate is shown. All results are means�SD for two or moreexperiments. N.D., experiment not done.
VectorDNA(�g)
LiAc(m)
Transformationefficiency(# tr./�g)
Doublepulse
pGAL1PNiST-1 10 100 19�13 (n=3) N.D.1 100 43�24 (n=3) 25�3·5 (n=4)
25 74�27 (n=3) N.D.5 128�9 (n=4) N.D.
0·1 100 120�16 (n=4) N.D.25 173�42 (n=3) N.D.5 216�150 (n=3) N.D.
pRM2 10 100 27�1 (n=2) N.D.1 100 317�87 (n=4) 269�128 (n=3)
25 249�124 (n=3) N.D.5 586�76 (n=4) N.D.
0·1 100 1340�290 (n=3) N.D.25 1890�533 (n=3) N.D.
5 2170�509 (n=2) N.D.
DNA is introduced without carrier, and the cellsare transformed using an electric pulse deliveredby an electroporation device. For maximumefficiency, yeast may be incubated with DTT andlithium acetate before concentration to render thecell wall more permeable. The C. albicans ura3mutant, CAI-4, was transformed with circularDNA from either an integrative (pGAL1PNiST-1)or an episomal (pRM2) vector.
The effects of amount of DNA, concentration oflithium acetate, spheroplast formation prior toelectroporation, elapsed time between pulse andplating, and the effect of double-pulsing, werestudied.
CAI-4 was transformed with 0·1, 1 or 10 �g ofeither pGAL1PNiST-1 or pRM2 using the electro-poration procedure described by Thompson et al.(1998) and the resulting transformation efficiencieswere calculated (Table 2). The difference in thetransformation efficiencies for the experimentsbeing compared were evaluated for significancewith use of a two-tailed Student’s t-test (p<0·10).Transformation efficiencies obtained when using0·1 �g of either vector were statistically better thanthose obtained with 1 �g DNA (p=0·013 and
Copyright � 1999 John Wiley & Sons, Ltd.
p=0·09 for pGAL1PNiST-1 and pRM2 respect-ively). Compared to 1 �g of DNA, a consistentlythree- to four-fold higher transformation efficiencywas obtained using the lowest amount (0·1 �g) ofDNA of either vector (n�3). When usinghigher amounts of DNA (10 �g) transformationefficiencies dropped drastically (Table 2).
The self-replicating vector, pRM2, exhibited atransformation efficiency �10-fold greater thanthat obtained with the integrative plasmid,pGAL1PNiST-1.
CAI-4 cells were treated with various concen-trations of lithium acetate (5, 25, 100 m; n�3)prior to electroporation with either 0·1 or 1 �g ofpGAL1PNiST-1 or pRM2. Reducing the lithiumacetate concentration from 100 m to 5 malways yielded somewhat higher transformationefficiencies (Table 2). When using 1 �g ofpGAL1PNiST-1 DNA, this difference was foundto be statistically significant (p=0·016). The samewas observed for pRM2 DNA (p=0·004).
Electroporation of spheroplasts might yieldhigher transformation efficiencies for DNA whichdoes not penetrate the yeast cell wall efficiently.Spheroplasts were formed using the enzyme
Yeast 15, 1609–1618 (1999)
1614 M. D. DE BACKER ET AL.
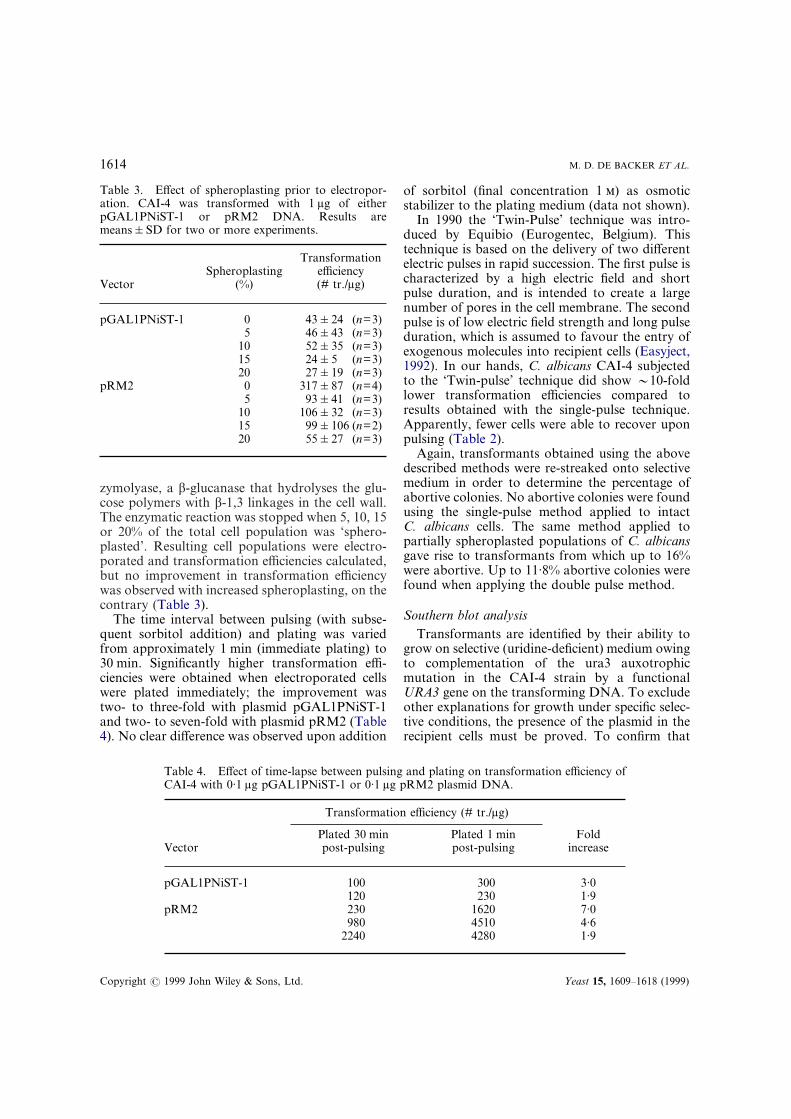
zymolyase, a �-glucanase that hydrolyses the glu-cose polymers with �-1,3 linkages in the cell wall.The enzymatic reaction was stopped when 5, 10, 15or 20% of the total cell population was ‘sphero-plasted’. Resulting cell populations were electro-porated and transformation efficiencies calculated,but no improvement in transformation efficiencywas observed with increased spheroplasting, on thecontrary (Table 3).
The time interval between pulsing (with subse-quent sorbitol addition) and plating was variedfrom approximately 1 min (immediate plating) to30 min. Significantly higher transformation effi-ciencies were obtained when electroporated cellswere plated immediately; the improvement wastwo- to three-fold with plasmid pGAL1PNiST-1and two- to seven-fold with plasmid pRM2 (Table4). No clear difference was observed upon addition
Copyright � 1999 John Wiley & Sons, Ltd.
of sorbitol (final concentration 1 ) as osmoticstabilizer to the plating medium (data not shown).
In 1990 the ‘Twin-Pulse’ technique was intro-duced by Equibio (Eurogentec, Belgium). Thistechnique is based on the delivery of two differentelectric pulses in rapid succession. The first pulse ischaracterized by a high electric field and shortpulse duration, and is intended to create a largenumber of pores in the cell membrane. The secondpulse is of low electric field strength and long pulseduration, which is assumed to favour the entry ofexogenous molecules into recipient cells (Easyject,1992). In our hands, C. albicans CAI-4 subjectedto the ‘Twin-pulse’ technique did show �10-foldlower transformation efficiencies compared toresults obtained with the single-pulse technique.Apparently, fewer cells were able to recover uponpulsing (Table 2).
Again, transformants obtained using the abovedescribed methods were re-streaked onto selectivemedium in order to determine the percentage ofabortive colonies. No abortive colonies were foundusing the single-pulse method applied to intactC. albicans cells. The same method applied topartially spheroplasted populations of C. albicansgave rise to transformants from which up to 16%were abortive. Up to 11·8% abortive colonies werefound when applying the double pulse method.
Table 3. Effect of spheroplasting prior to electropor-ation. CAI-4 was transformed with 1 �g of eitherpGAL1PNiST-1 or pRM2 DNA. Results aremeans�SD for two or more experiments.
VectorSpheroplasting
(%)
Transformationefficiency(# tr./�g)
pGAL1PNiST-1 0 43�24 (n=3)5 46�43 (n=3)
10 52�35 (n=3)15 24�5 (n=3)20 27�19 (n=3)
pRM2 0 317�87 (n=4)5 93�41 (n=3)
10 106�32 (n=3)15 99�106 (n=2)20 55�27 (n=3)
Table 4. Effect of time-lapse between pulsing and plating on transformation efficiency ofCAI-4 with 0·1 �g pGAL1PNiST-1 or 0·1 �g pRM2 plasmid DNA.
Vector
Transformation efficiency (# tr./�g)
Foldincrease
Plated 30 minpost-pulsing
Plated 1 minpost-pulsing
pGAL1PNiST-1 100 300 3·0120 230 1·9
pRM2 230 1620 7·0980 4510 4·6
2240 4280 1·9
Southern blot analysisTransformants are identified by their ability to
grow on selective (uridine-deficient) medium owingto complementation of the ura3 auxotrophicmutation in the CAI-4 strain by a functionalURA3 gene on the transforming DNA. To excludeother explanations for growth under specific selec-tive conditions, the presence of the plasmid in therecipient cells must be proved. To confirm that
Yeast 15, 1609–1618 (1999)
1615TRANSFORMATION OF CANDIDA ALBICANS BY ELECTROPORATION
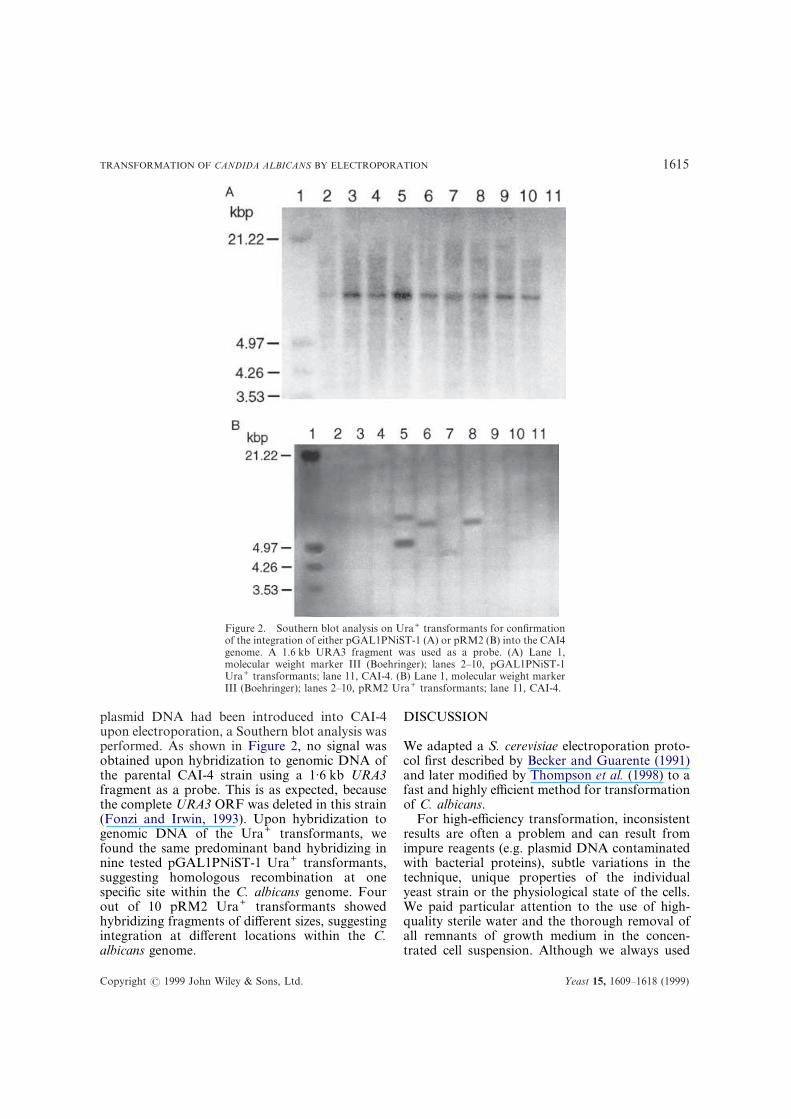
plasmid DNA had been introduced into CAI-4upon electroporation, a Southern blot analysis wasperformed. As shown in Figure 2, no signal wasobtained upon hybridization to genomic DNA ofthe parental CAI-4 strain using a 1·6 kb URA3fragment as a probe. This is as expected, becausethe complete URA3 ORF was deleted in this strain(Fonzi and Irwin, 1993). Upon hybridization togenomic DNA of the Ura+ transformants, wefound the same predominant band hybridizing innine tested pGAL1PNiST-1 Ura+ transformants,suggesting homologous recombination at onespecific site within the C. albicans genome. Fourout of 10 pRM2 Ura+ transformants showedhybridizing fragments of different sizes, suggestingintegration at different locations within the C.
Figure 2. Southern blot analysis on Ura+ transformants for confirmationof the integration of either pGAL1PNiST-1 (A) or pRM2 (B) into the CAI4genome. A 1.6 kb URA3 fragment was used as a probe. (A) Lane 1,molecular weight marker III (Boehringer); lanes 2–10, pGAL1PNiST-1Ura+ transformants; lane 11, CAI-4. (B) Lane 1, molecular weight markerIII (Boehringer); lanes 2–10, pRM2 Ura+ transformants; lane 11, CAI-4.
albicans genome.
Copyright � 1999 John Wiley & Sons, Ltd.
DISCUSSION
We adapted a S. cerevisiae electroporation proto-col first described by Becker and Guarente (1991)and later modified by Thompson et al. (1998) to afast and highly efficient method for transformationof C. albicans.
For high-efficiency transformation, inconsistentresults are often a problem and can result fromimpure reagents (e.g. plasmid DNA contaminatedwith bacterial proteins), subtle variations in thetechnique, unique properties of the individualyeast strain or the physiological state of the cells.We paid particular attention to the use of high-quality sterile water and the thorough removal ofall remnants of growth medium in the concen-
trated cell suspension. Although we always usedYeast 15, 1609–1618 (1999)

1616 M. D. DE BACKER ET AL.
the same C. albicans strain, the same plasmidpreparations and the same batches of reagents forall experiments, still some variation in the resultscan be seen from one experiment to another (seetables). Despite this variation, clear trends towardshigher or lower transformation efficiencies couldbe observed upon changing certain parameters.
For C. maltosa it has been shown that self-replicating vectors exhibit a transformation effi-ciency that is 100- to 1000-fold greater than thatobtained with integrative plasmids (Becher andOliver, 1995), reflecting the need for additionalsteps (e.g. homologous recombination) in case ofthe latter. We observed only a 10-fold greatertransformation efficiency when using pRM2 com-pared to pGAL1PNiST-1 upon transformation ofC. albicans CAI-4. This suggests that homologousrecombination is far more frequent or efficient inC. albicans than in C. maltosa.
With both plasmids, the transformation effi-ciency was increased three- to four-fold when usinglower amounts of DNA (0·1 �g instead of 1). Thissuggests that at higher DNA concentrations thecapacity for DNA uptake is limiting.
Lowering the concentration of lithium acetatefor pretreatment of the cells to as low as 5 myielded a statistically significant improvement intransformation efficiency when using 1 �g DNAfor electroporation. Lithium acetate is supposed torender the cell wall more permeable but highconcentrations give toxic effects.
Enzymatic removal of the cell wall prior toelectroporation lead to a lower recovery of cellsafter pulsing. Spheroplasts can be envisaged to bemore sensitive to pulsing compared to intact cells.
A clear and very significant improvement intransformation efficiency was observed when cellswere plated immediately after pulsing. Incubationof the cells for 30 min in cold osmotic mediumprior to plating clearly reduced the transformationefficiency; apparently, more cells fail to regeneratetheir cell wall after a prolonged incubation inosmotic stabilizing solution prior to plating ontoselective medium. It is unclear whether this is dueto inadequate stabilization of the cell membrane,but allowing the cells to ‘recover’ in osmotic solu-tion is clearly counterproductive. C. albicans cellsmay just recover more efficiently on solid medium.
A combination of using 0·1 �g DNA for trans-formation and the immediate plating resulted inmaximum transformation efficiencies of �300transformants/�g for pGAL1PNiST-1 and �4500transformants/�g for pRM2.
Copyright � 1999 John Wiley & Sons, Ltd.
The pGAL1PNiST-1 vector contains the galacto-kinase 1 promoter region which is identical to partof a resident gene of the CAI-4 genome and thusprobably promotes homologous recombinationat this site. Southern blot analysis on Ura+ trans-formants indeed showed that the integrative vectorpGAL1PNiST-1 integrates at the same location inthe genome for all transformants analysed.
pRM2 contains multiple CARSs but still fails tostay episomal for a full 100%; integration haspreviously been reported to occur at a frequency of15% (Negredo et al., 1997). This results in theoccurrence of small (transformants carryingpRM2 as an episomal molecule) and large (trans-forming pRM2 DNA integrated into the genome)colonies (Herreros et al., 1992). Southern blotanalysis on pRM2 Ura+ transformants showedthat integration occurred and this at a frequencyhigher than expected.
It has previously been shown that electroporatedcells remain osmotically sensitive for a consider-able time; Becher and Oliver (1995) obtainedtransformants when high concentrations of DNAwere added in sorbitol 10 s after pulsing. We foundthat the presence of sorbitol in the medium at thetime of plating (�1–2 min post-pulsing) is nolonger mandatory. This is in contrast to what wasfound for S. cerevisiae, where �15-fold highertransformation efficiencies were found when sorbi-tol was added to the plating medium (Becker andGuarente, 1991). Schmitt and Erdos (1968)showed that C. albicans is highly resistant to theplasmolytic effects of glucose and sucrose andothers also found that C. albicans is indeed veryresilient to high external osmotic pressure (OddsF., personal communication).
Double-pulse electroporation did not result inany improvement in C. albicans transformationefficiency; on the contrary, transformation efficien-cies were found to be generally about 10-foldlower. For C. maltosa, this technique was alsofound not to be superior to the standardsingle-pulse technique (Becher and Oliver, 1995).
In conclusion, the electroporation procedure wedescribe here for C. albicans CAI-4 yields transfor-mation efficiencies comparable to those obtainedwhen using the more laborious and time-consuming spheroplast procedures. Electropor-ation is fast and in addition gives rise tosignificantly fewer abortive colonies comparedto the spheroplast transformation methods.Although the lithium acetate transformationmethod is less time-consuming as well, this method
Yeast 15, 1609–1618 (1999)

1617TRANSFORMATION OF CANDIDA ALBICANS BY ELECTROPORATION
has been shown to be particularly inefficient forC. albicans (�10- to 100-fold lower transforma-tion efficiencies compared to electroporation).
ACKNOWLEDGEMENTS
We would like to thank Dr William Fonzi(University of California, Irvine, CA) for supply-ing us with the CAI-4 strain. We thank FerminSimons and Inge Loonen (both at JanssenResearch Foundation) for technical support andFrank Odds (JRF) for advice. We thank GertVerheyen (Janson Research Foundation) forgraphical support. This work was supported inpart by the Vlaams Instituut voor de Bevorderingvan het Wetenschappelijk-Technologisch Onder-zoek in de Industrie (IWT, Belgium, Grant No.960192).
ABBREVIATIONS
Deoxyribonucleoside triphosphates (dNTPs)Dithiothreitol (DTT)Orotidine-5�-phosphate decarboxylase (URA3)Polyethylene glycol (PEG)Sodium chloride/sodium citrate (SSC). . . (SSPE) Sodium chloride/sodium phosphateEDTASodium dodecyl sulphate (SDS)Standard deviation (SD)Tris acetate EDTA (TAE). . . (TBE) Tris borate EDTATris EDTA (TE)Candida autonomously replicating sequence(CARS)
REFERENCES
Ausubel, F. M., Brent, R., Kingston, R. E., Moore,D. D., Seidman, J. G., Smith, J. A. and Struhl, K.(eds) (1990). Current Protocols in Molecular Biology,Vol. 1, Unit 13.7. Wiley, New York.
Becher, D. and Oliver, S. G. (1995). Electroporationprotocols for microorganisms. In Nickoloff, J. A.(Ed.), Methods in Molecular Biology, Vol. 47.Humana, Totowa, NJ.
Becker, D. M. and Guarente, L. (1991). High-efficiencytransformation of yeast by electroporation. MethodsEnzymol. 194, 182–187.
Beggs, J. D. (1978). Transformation of yeast by a repli-cating hybrid plasmid. Nature 275(5676), 104–109.
Brown, D. H. Jr, Slobodkin, I. V. and Kumamoto, C. A.(1996). Stable transformation and regulated expres-
sion of an inducible reporter construct in CandidaCopyright � 1999 John Wiley & Sons, Ltd.
albicans using restriction enzyme-mediated integra-tion. Mol. Gen. Genet. 251(1), 75–80.
Delorme, E. (1989). Transformation of Saccharomycescerevisiae by electroporation. Appl. Environ. Micro-biol. 55, 2242–2246.
Easyject (1992). User’s Manual 2, 1–46.Faber, K. N., Haima, P., Harder, W. and Veenhuis, M.,
Ab G. (1993). Highly-efficient electrotransformationof the yeast Hansenula polymorpha. Curr. Genet. 25,305–310.
Fonzi, W. A. and Irwin, M. Y. (1993). Isogenic strainconstruction and gene mapping in Candida albicans.Genetics 134, 717–728.
Gietz, R. D., Schiestl, R. H., Willems, A. R. and Woods,R. A. (1995). Studies on the transformation of intactyeast cells by the LiAc/SS-DNA/PEG procedure.Yeast 11, 355–360.
Herreros, E., Garcia-Saez, M. I., Nombela, C. andSanchez, M. (1992). A reorganized Candida albicansDNA sequence promoting homologous non-integrative genetic transformation. Mol. Microbiol.6(23), 3567–3574.
Hill, D. E. (1989). Integrative transformation of yeastusing electroporation. Nucleic Acids Res. 17(19), 8011.
Hinnen, A., Hicks, J. B. and Fink, G. R. (1978).Transformation of yeast. Proc. Natl Acad. Sci. U S A75, 1929.
Invitrogen (1997). Pichia Spheroplast Module: AManual for the Preparation and Transformation ofPichia pastoris Spheroplasts, Version 1.0.
Ito, H., Fukuda, Y., Murata, K. and Kimura, A. (1983).Transformation of intact yeast cells treated with alkalications. J. Bacteriol. 153, 163–168.
Kasuske, A., Wedler, H., Schulze, S. and Becher, D.(1992). Efficient electropulse transformation of intactCandida maltosa cells by different homologous vectorplasmids. Yeast 8, 691–697.
Kondo, K., Saito, T., Kajiwara, S., Takagi, M. andMisawa, N. (1995). A transformation system for theyeast Candida utilis: use of a modified endogenousribosomal protein gene as a drug-resistant marker andribosomal DNA as an integration target for vectorDNA. J. Bacteriol. 177, 7171–7177.
Manivasakam, P. and Schiestl, R. H. (1993). Highefficiency transformation of Saccharomyces cerevisiaeby electroporation. Nucleic Acids Res. 21(18), 4414–4415.
Negredo, A., Monteoliva, L., Gil, C., Pla, J. andNombela, C. (1997). Cloning, analysis and one-stepdisruption of the ARG5,6 gene of Candida albicans.Microbiology 143, 297–302.
Pla, J., Gil, C., Monteoliva, L., Navarro-Garcia, F.,Sanchez, M. and Nombela, C. (1996). UnderstandingCandida albicans at the molecular level. Yeast 12(16),1677–1702.
Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989).Molecular Cloning. Cold Spring Harbor Laboratory
Press, New York.Yeast 15, 1609–1618 (1999)

1618 M. D. DE BACKER ET AL.
Sanglard, D., Ischer, F., Monod, M. and Bille, J. (1996).Susceptibilities of Candida albicans multidrug trans-porter mutants to various antifungal agents andother metabolic inhibitors. Antimicrobiol. AgentsChemother. 40, 2300–2305.
Schmitt, J. A. and Erdos, G. W. (1968). Plasmolyticeffects of glucose and sucrose on cells of Candidaalbicans. Mycologia 60(5), 1107–1112.
Copyright � 1999 John Wiley & Sons, Ltd.
Simon, J. R. (1993). Transformation of intact yeast cellsby electroporation. Methods Enzymol. 217, 478–483.
Thompson, J. R., Register, E., Curotto, J., Kurtz, M.and Kelly, R. (1998). An improved protocol for thepreparation of yeast cells for transformation byelectroporation. Yeast 14, 565–571.
Yeast 15, 1609–1618 (1999)


![PENGHAMBATAN CAJUPUTS CANDY TERHADAP VIABILITAS KHAMIR Candida albicans SECARA IN VITRO [Inhibition of Cajuputs Candy Toward the Viability of Candida albicans by using In Vitro Assay]](https://img.pdfslide.net/doc/110x75/635c4cc7a3fa66b45c0dd761/penghambatan-cajuputs-candy-terhadap-viabilitas-khamir-candida-albicans-secara-in.jpg)


























![[Characterization of azole resistance mechanisms in Chilean clinical isolates of Candida albicans]](https://img.pdfslide.net/doc/110x75/634a291cdedec593d70ede27/characterization-of-azole-resistance-mechanisms-in-chilean-clinical-isolates-of.jpg)