Embed Size (px)
Citation preview

RESEARCH
Transgenic Brassica juncea Plants Expressing MsrA1, a SyntheticCationic Antimicrobial Peptide, Exhibit Resistance to FungalPhytopathogens
Anjana Rustagi • Deepak Kumar • Shashi Shekhar •
Mohd Aslam Yusuf • Santosh Misra •
Neera Bhalla Sarin
� Springer Science+Business Media New York 2014
Abstract Cationic antimicrobial peptides (CAPs) have
shown potential against broad spectrum of phytopathogens.
Synthetic versions with desirable properties have been
modeled on these natural peptides. MsrA1 is a synthetic
chimera of cecropin A and melittin CAPs with antimicro-
bial properties. We generated transgenic Brassica juncea
plants expressing the msrA1 gene aimed at conferring
fungal resistance. Five independent transgenic lines were
evaluated for resistance to Alternaria brassicae and Scle-
rotinia sclerotiorum, two of the most devastating patho-
gens of B. juncea crops. In vitro assays showed inhibition
by MsrA1 of Alternaria hyphae growth by 44–62 %. As
assessed by the number and size of lesions and time taken
for complete leaf necrosis, the Alternaria infection was
delayed and restricted in the transgenic plants with the
protection varying from 69 to 85 % in different transgenic
lines. In case of S. sclerotiorum infection, the lesions were
more severe and spread profusely in untransformed control
compared with transgenic plants. The sclerotia formed in
the stem of untransformed control plants were significantly
more in number and larger in size than those present in the
transgenic plants where disease protection of 56–71.5 %
was obtained. We discuss the potential of engineering
broad spectrum biotic stress tolerance by transgenic
expression of CAPs in crop plants.
Keywords Biotic stress � Brassica juncea � Cationic
antimicrobial peptides � MsrA1 � Transgenic plants
Introduction
Despite significant advances in agricultural practices, an
estimated 13.6 % of the world population (*925 million
people) remains hungry. The Asia and Pacific regions con-
tribute *63 % to these hunger statistics [1]. Besides the loss
of arable lands to urban developments, stresses imposed by
various abiotic and biotic factors on crops remain the major
impediment for the agricultural productivity to catch up with
the global demand. Bacteria, fungi, and viruses are the main
phytopathogens affecting crops leading to significant losses
and have been responsible for scripting some of the most
devastating famines in the human history [2, 3]. Plant disease
control by chemical pesticides, apart from being costly, is
increasingly being scrutinized for health and environmental
concerns [2]. Conventional approaches to breed disease-
resistant crop varieties, though significant, are time con-
suming and have fallen short to tackle the vast array of dis-
eases. Thus, the focus has shifted to developing resistance in
plants against a broad spectrum of pathogens by engineering
plants expressing transgenes that can combat these
microorganisms.
Antimicrobial peptides (AMPs) have increasingly
gained attention as candidates for disease protection in
plants. AMPs are usually 12–50-amino acid-long peptides
Anjana Rustagi and Deepak Kumar contributed equally to this work.
A. Rustagi � D. Kumar � S. Shekhar � M. A. Yusuf �N. B. Sarin (&)
School of Life Sciences, Jawaharlal Nehru University,
New Delhi 110067, India
e-mail: [email protected]
Present Address:
A. Rustagi
Department of Botany, Ramjas College, University of Delhi,
Delhi 110007, India
S. Misra
Department of Biochemistry and Microbiology, University of
Victoria, Victoria, BC V8W3P6, Canada
123
Mol Biotechnol
DOI 10.1007/s12033-013-9727-8

which are components of innate defense mechanisms in
organisms ranging from microbes to plants and animals [4].
More than 900 AMPs have been reported, which have
either been identified from natural sources or have been
artificially synthesized. The cationic AMPs (CAPs) con-
stitute the largest group, and the term is often inter-
changeably used for the AMPs in the scientific literature.
The CAPs usually acquire an a-helical or a b-sheet struc-
ture and have been found to be active against Gram-posi-
tive and -negative bacteria, fungi, protozoa, and viruses [5].
Most of the AMPs have a common mechanism of action
that targets the differences between host and target cell
membranes and lead to lethal membrane dysfunction by
making pores through it [6]. Besides, they have also been
reported to interfere with cell division, macromolecular
synthesis, and cell wall formation [7]. The transgenic
expression of AMPs has delivered encouraging results in
conferring specific or broad spectrum disease resistance in
plants such as tobacco [8–11], rice [12], potato [13, 14],
banana [15], tomato [16], hybrid poplar [17], and grapevine
[18].
Cecropin A, exhibiting antibacterial and antifungal
activities (isolated from Hyalophora cecropia), and melit-
tin (the major lytic component in the venom of honeybee)
are well-known CAPs. A chimeric peptide, CEMA, com-
bining the bioactive portions of cecropin A and melittin
was engineered by Hancock et al. [19]. In a previous study
by Osusky et al. [13], an MsrA1 peptide was designed by
adding a hexapeptide at the N-terminus of CEMA. The
MsrA1 peptide was predicted, by molecular modeling, to
acquire an a-helical structure and maintain a positively
charged N-terminus. The msrA1 gene when expressed in
transgenic tobacco and potato plants conferred broad-
spectrum resistance to phytopathogens [8, 13].
Brassica juncea (Indian mustard) is an important oilseed
crop grown in many countries around the world and has
been suggested as an alternative crop to canola because of
its higher heat and water-stress tolerance [20, 21]. In India,
mustard contributes 28.6 % to the total oilseed production
and has been projected to provide for 41 % (14 million
tons) of the country’s demand by the year 2020 [22].
However, the crop productivity over the years has been
seriously affected by fungal pathogens, especially, Alter-
naria brassicae and Sclerotinia sclerotiorum [23, 24]. The
fungicide application to counter these pathogens has not
been a very effective and economical proposition, and
therefore, better combat strategies relying on generation of
transgenic plants armored with defense proteins are being
explored [23].
In view of the protective action of MsrA1 in tobacco and
potato, we assessed the efficacy of MsrA1 in conferring
disease-resistance traits to the economically important
oilseed crop of B. juncea. In the present work, we describe
the generation of transgenic B. juncea plants expressing the
msrA1 gene and evaluation of their resistance to A.
brassicae and S. sclerotiorum. The results suggest a high
degree of protection of B. juncea plants against these
fungal pathogens. We discuss our results in the light of
current efforts to generate biotic stress-tolerant transgenic
plants and the economic and environmental impacts that
these approaches could have.
Materials and Methods
Binary Vector for Expression of msrA1 Gene
The construction of pSAI4 plasmid used for the expression
of msrA1 gene has been described by Osusky et al. [13].
The T-DNA portion of this expression construct is shown
in Fig. 1a. The pSAI4 plasmid was transformed into the
Agrobacterium tumefacienes strain GV3101 following the
standard freeze–thaw procedure [25].
Plant Transformation and Regeneration
The B. juncea cv. Varuna seeds used for transformation
were procured from the Indian Agricultural Research
Institute, New Delhi. The seeds were surface sterilized by
treatment with a detergent (Extran) for 5 min, 70 % etha-
nol for 1 min, and HgCl2 (0.05 %) for 10 min followed by
three washes with autoclaved distilled water under sterile
conditions. The sterilized seeds were germinated on semi-
solid Murashige and Skoog’s (MS) basal medium [26].
One-cm-long hypocotyl explants were chopped from
5-day-old in vitro grown seedlings and used for Agrobac-
terium-mediated transformation [27]. In brief, the hypo-
cotyl explants were precultured in liquid MS B1N1 (MS
medium supplemented with 1 mg/l each of BAP and NAA)
at 25 �C for 18–24 h with gentle shaking followed by
coinfection with pSAI4 harboring Agrobacterium at a
bacterial density of 0.5 OD for 30 min. After cocultivation
for 18–24 h with gentle shaking (100 rpm), the explants
were washed twice with MS B1N1 containing augmentin
(200 mg/l) and subsequently plated on semi-solid MS
B1N1 containing AgNO3 (3.4 mg/l), augmentin (200 mg/l),
and kanamycin (50 mg/l). The plates were kept in culture
room maintained at 22 ± 2 �C and 14-h light/10-h dark
photoperiod. The shoots obtained (after 15–20 days) from
the explants were subcultured on the above medium and
finally rooted on MS I2 (MS medium supplemented with
2 mg/l IBA). The well-established control and putative
transgenic plantlets obtained in vitro were hardened and
transferred to the glass house to grow under 25 ± 2 �C
temperature, 16-h light/8-h dark photoperiod, and 70 %
relative humidity. The seeds obtained from the T0 plants
Mol Biotechnol
123

were grown to get the T1 plants for performing the fungal
infection experiments.
DNA Extraction and PCR Analysis of the Transgenic
Plants
Genomic DNA was isolated from the leaves of untrans-
formed control and transgenic plants following the protocol
of Murray and Thompson [28]. The preliminary screening
of the transgenic plants for successful transformation was
performed by polymerase chain reaction (PCR) analysis
using the neomycin phosphotranferase (npt II) and msrA1
gene-specific primers mentioned below:
npt II
Forward primer: 50-GGAGCGGCGATACCGTAAAGC-30
Reverse primer: 50-GAGGCTATTCGGCTATGACTG-30
msrA1
Forward primer: 50-TTACTTAGTTAGCTTCAGCGC-30
Reverse primer: 50-ATGGCTCTAGAGCATATGAAA-30
The PCR conditions for both the set of primers were
identical and included an initial denaturation at 94 �C for
a
b
d
e
c
Fig. 1 Generation and molecular analysis of transgenic Brassica
juncea plants expressing msrA1 gene. (a) T-DNA portion of the
pSAI4 vector construct used for plant transformation. The position of
XbaI restriction endonuclease site used for Southern analysis of the
plants is shown. The amino acid and nucleotide sequence of MsrA1 is
also shown. PCR analysis to screen the transgenic plants for the
presence of transgene was done using msrA1 gene specific (b) and
nptII primers (c). The Southern blot analysis of the transgenic plants
was done after digestion of genomic DNA with XbaI (d) and the
expression of the transgene was confirmed by RT-PCR using the
msrA1 specific primers (e). M DNA molecular weight marker, UT
untransformed control plant, M1–M8 independent transgenic lines
expressing msrA1 gene
Mol Biotechnol
123

5 min, followed by 35 cycles of 94 �C for 30 s, 58 �C for
30 s, 72 �C for 30 s, and a final extension at 72 �C for
5 min. An aliquot of the PCR product was electrophoresed
on a 1.5 % agarose gel and visualized by ethidium bromide
staining.
Southern Blot Analysis
Ten micrograms of genomic DNA from untransformed
control and transgenic plants was digested with XbaI
restriction endonuclease, electrophoresed on a 0.8 % aga-
rose gel and transferred to a Hybond-N? membrane
(Amersham Biosciences, UK) using the capillary transfer
method [29]. The membrane was prehybridized for 2 h at
65 �C in a buffer containing 0.5 M sodium phosphate
buffer, pH 7.2, 1 mM EDTA, and 7 % SDS. Thereafter,
denatured radiolabeled probe (100 bp msrA1 amplicon
obtained by PCR amplification of pSAI4 plasmid and
labeled with [a-32P] dCTP using Random primer DNA
labeling kit (Amersham Biosciences, UK) as per the
manufacturer’s instructions) was added to the prehybrid-
ization buffer and incubated overnight at 65 �C. The
membrane was washed (10 min per wash) sequentially in
39 SSC, 0.1 % SDS; 0.59 SSC, 0.1 % SDS; 0.29 SSC;
0.1 % SDS with constant agitation at 65 �C. The hybrid-
ization signals were captured using phosphorimaging (FLA
5000 imaging system, Fujiflim).
RNA Extraction and Reverse Transcriptase-PCR
Analysis
Total RNA was isolated from the leaves using TRIzol
reagent (Invitrogen, CA, USA) and treated with RNase-free
DNase I to remove any contaminating genomic DNA. RT-
PCR amplification was carried out using a kit (AccuScript,
Stratagene, USA) as per the manufacturer’s recommenda-
tions. Total RNA (1 lg) was reverse transcribed using
MMLV reverse transcriptase. The primers for amplification
of the msrA1 gene were as described above. The primers
for actin (used as an internal control) were 50-AG
TAAGGTGACCTTGCAATTACTTTAGACTTCACCG-30
and 50-AAAGGCTAGCGTTGAAGATGCCTCTGCCGA
C-30. The RT-PCR products were visualized by electro-
phoresis on a 1.5 % agarose gel.
Evaluation of Resistance Against Alternaria brassicae
In Vitro Antifungal Assay
The antifungal activity of MsrA1 protein was assessed
using a plate assay. Alternaria brassicae (Accession No.
ITCC 5097) procured from the Division of Plant Pathol-
ogy, Indian Agricultural Research Institute, New Delhi was
cultured on potato dextrose agar (PDA) medium at
22 ± 1 �C, and the spore suspension was prepared as
described by Sharma et al. [30]. Total protein was extracted
from the leaves of the untransformed control and msrA1-
expressing transgenic plants using protein extraction buffer
(50 mM sodium phosphate buffer, pH 7.5, containing plant
protease inhibitor cocktail from G-Biosciences, USA).
Wells were punched out in agar plates and filled with 50 ll
of A. brassicae spore suspension (106 spores/ml) and
incubated overnight at 30 �C. After the incubation, the
wells were filled with 50 ll (200 lg protein) of one of the
following: untransformed control leaf extract, untrans-
formed control leaf extract treated with proteinase K
(Promega; 100 lg/ml for 2 h at 37 �C), transgenic leaf
extract, or transgenic leaf extract treated with proteinase K
(likewise as in the case of untransformed control). The
plates were further incubated, and the radial growth of A.
brassicae was recorded every 24 h for 5 days. The per-
centage inhibition of hyphal growth was calculated using
the method of Mondal et al. [23].
Inhibition of hyphal growth %ð Þ¼ Diameter of the fungal colony around the wellð
treated with the transgenic leaf extract/diameter
of the fungal colony around the well treated
with the control leaf extract�100
The antifungal activity against A. brassicae was also
checked using the synthetic MsrA1 peptide. The peptide
(amino acid sequence shown in Fig. 1a) was custom syn-
thesized by a commercial source (Link Biotech, India). A
small agar plug containing the fungus was placed in the
center of a petri dish filled with PDA. When the fungal
growth reached to about 45 mm in diameter, sterile filter
paper disks were placed along the circumference of the
growing mycelia. Specific amounts of the peptide were
pipetted onto the disks in 20 ll volumes (sterile water
served as the control). Further growth of the fungus was
monitored by incubating for another 24 h, and the plates
were photographed.
In Vivo Plant Bioassay
The A. brassicae spore suspension was filtered through
cheesecloth to remove the mycelium debris. The spores
were washed twice with sterile distilled water and resus-
pended to a count of 100 spores/ml. For in vivo plant
bioassay, the fifth leaf (from the top) of the 45-day-old
plants was painted with the spore suspension using a
painting brush. The inoculated plants were covered with
transparent polythene bag to maintain artificial epiphytotic
and humid conditions conducive to infection. The data on
lesions per leaf and individual lesion diameter were
Mol Biotechnol
123

recorded up to 15 days of inoculation. Percentage disease
protection was calculated as described by Mondal et al.
[23].
Disease protection %ð Þ¼ No: of lesions per leaf in untransformed control plantð½�No: of lesions per leaf in transgenicÞ=No: of lesions per leaf in untransformed control plantÞ��100
Evaluation of Resistance Against Sclerotinia
sclerotiorum
Fungal culture of S. sclerotiorum (Accession No. ITCC
6583) obtained from the Division of Plant Pathology,
Indian Agricultural Research Institute, New Delhi was
maintained in the lab following the protocol described by
Jensen et al. [31]. The 45-day-old plants were inoculated
with the fungus by placing a 20-mm mycelial disk at the
third internode (axillary position) of the main stem. The
site of inoculation was covered with parafilm, and the
plants were enclosed under transparent polythene bags to
provide high humid conditions. Disease incidence was
assessed by recording the average size of the lesion on
stem, days to stem breaking, and the number of sclerotia
formed inside the main stem. The percentage disease pro-
tection was calculated as
Disease protection %ð Þ¼ Average size of lesions in untransformed control plantð½� Average size of lesions in transgenicÞ=Average size of lesions in untransformed control plantÞ��100
Statistical Analysis
The infection experiments were carried out three inde-
pendent times, in triplicates with three plants used for each
of the untransformed control and transgenic line. The data
are presented as average ± standard deviation. Results
were analyzed by the Student’s t test. Significance was
defined as P \ 0.05.
Results
Generation and Molecular Analysis of Transgenic
Brassica juncea Plants Expressing msrA1 Gene
Transgenic B. juncea plants expressing the msrA1 gene
were generated by Agrobacterium-mediated transforma-
tion. The plasmid, pSAI4, contains msrA1 gene cloned
between an enhanced CaMV 35S promoter with an AMV
RNA4 translation-enhancing element and NOS termination
sequence (Fig. 1a). The neomycin phosphotransferase II
(npt II) gene present in the plasmid was used for kana-
mycin screening of the transgenic plants. Preliminary
screening of the T0 transgenic B. juncea plants selected on
kanamycin was done by PCR amplification of the msrA1
and npt II genes. In eight independent T0 transgenic lines,
an amplicon of *100 bp was found with the msrA1 gene-
specific primers and of *700 bp with npt II-sspecific
primers (Fig. 1b, c). These fragments were not amplified in
the case of untransformed control plants. Out of these
seven surviving PCR positive lines, five lines (M1, M3,
M5, M6, and M7) were found to be Southern positive.
Figure 1d shows the signals obtained in Southern blot
analysis in the T1 generation of these lines. As the genomic
DNA was digested with XbaI endonuclease—which has a
single restriction site in the T-DNA of the construct—it is
evident from Fig. 1d that a single copy of msrA1 was
integrated in M6 and M7, while two copies of the transgene
were present in each of the lines M1, M3, and M5. The
expression level of the msrA1 gene in the transgenic lines
was assessed by RT-PCR analysis and deduced densito-
metrically after normalizing for the actin levels. Figure 1e
shows the presence of *100 bp band corresponding to the
amplified msrA1 cDNA in all the confirmed T1 transgenic
lines. All the transgenic plants were morphologically
similar to the untransformed control plants and showed no
visible signs of impaired growth or physiology. The plants
from these five T1 transgenic lines as well as the untrans-
formed control plants were used for protection assays
against the fungal pathogens.
Resistance of Transgenic B. juncea Plants Expressing
msrA1 Gene to A. brassicae
In Vitro Antifungal Assay
The efficacy of MsrA1 against A. brassicae was assayed
using an in vitro assay for inhibition of hyphal growth as
described in the materials and methods. The growth of
hyphae around the wells treated with the protein extract
from the transgenic leaves was greatly inhibited compared
to those treated with protein extract from the untrans-
formed control leaves or either of the extracts after pro-
teinase K treatment (Fig. 2a). The hyphal growth inhibition
observed with the extract from different transgenic lines
was calculated with reference to the growth around the
wells treated with untransformed control extract and is
represented as percentage inhibition in Fig. 2b. The per-
centage inhibition in the different transgenic lines varied
between 44 and 62 % over the untransformed control. To
further ascertain the antifungal activity of MsrA1 peptide
against A. brassicae, another in vitro assay was done using
Mol Biotechnol
123
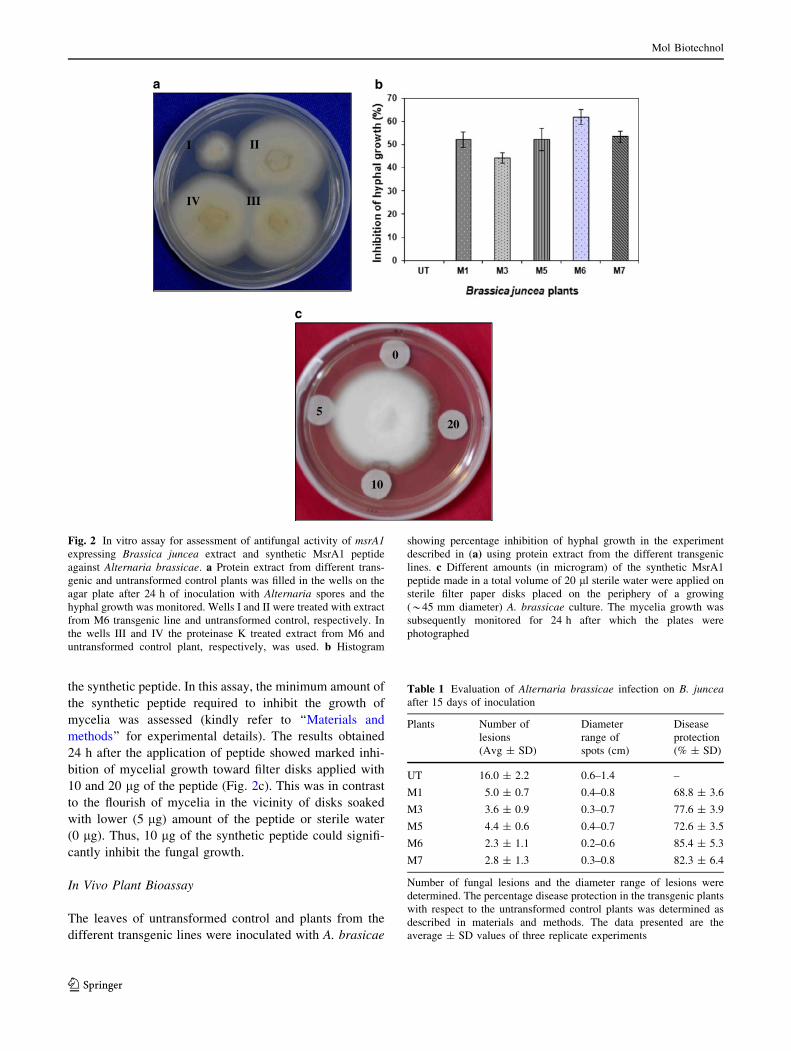
the synthetic peptide. In this assay, the minimum amount of
the synthetic peptide required to inhibit the growth of
mycelia was assessed (kindly refer to ‘‘Materials and
methods’’ for experimental details). The results obtained
24 h after the application of peptide showed marked inhi-
bition of mycelial growth toward filter disks applied with
10 and 20 lg of the peptide (Fig. 2c). This was in contrast
to the flourish of mycelia in the vicinity of disks soaked
with lower (5 lg) amount of the peptide or sterile water
(0 lg). Thus, 10 lg of the synthetic peptide could signifi-
cantly inhibit the fungal growth.
In Vivo Plant Bioassay
The leaves of untransformed control and plants from the
different transgenic lines were inoculated with A. brasicae
a b
II
IV III
c
0
5
10
20
I
Fig. 2 In vitro assay for assessment of antifungal activity of msrA1
expressing Brassica juncea extract and synthetic MsrA1 peptide
against Alternaria brassicae. a Protein extract from different trans-
genic and untransformed control plants was filled in the wells on the
agar plate after 24 h of inoculation with Alternaria spores and the
hyphal growth was monitored. Wells I and II were treated with extract
from M6 transgenic line and untransformed control, respectively. In
the wells III and IV the proteinase K treated extract from M6 and
untransformed control plant, respectively, was used. b Histogram
showing percentage inhibition of hyphal growth in the experiment
described in (a) using protein extract from the different transgenic
lines. c Different amounts (in microgram) of the synthetic MsrA1
peptide made in a total volume of 20 ll sterile water were applied on
sterile filter paper disks placed on the periphery of a growing
(*45 mm diameter) A. brassicae culture. The mycelia growth was
subsequently monitored for 24 h after which the plates were
photographed
Table 1 Evaluation of Alternaria brassicae infection on B. juncea
after 15 days of inoculation
Plants Number of
lesions
(Avg ± SD)
Diameter
range of
spots (cm)
Disease
protection
(% ± SD)
UT 16.0 ± 2.2 0.6–1.4 –
M1 5.0 ± 0.7 0.4–0.8 68.8 ± 3.6
M3 3.6 ± 0.9 0.3–0.7 77.6 ± 3.9
M5 4.4 ± 0.6 0.4–0.7 72.6 ± 3.5
M6 2.3 ± 1.1 0.2–0.6 85.4 ± 5.3
M7 2.8 ± 1.3 0.3–0.8 82.3 ± 6.4
Number of fungal lesions and the diameter range of lesions were
determined. The percentage disease protection in the transgenic plants
with respect to the untransformed control plants was determined as
described in materials and methods. The data presented are the
average ± SD values of three replicate experiments
Mol Biotechnol
123

spores, and the initiation and progression of the disease
lesions were followed in the subsequent days. After
15 days, the number of characteristic lesions on the leaves
of transgenic plants was much less (\5 lesions/leaf) in
comparison to that on the leaves of the untransformed
control plants ([14 lesions/leaf) (Table 1). The size of the
lesions ranged between 0.6 and 1.4 cm in diameter in the
case of untransformed control plants as against 0.2–0.8 cm
in the different transgenic lines (Table 1). In the untrans-
formed control plants, the lesions characteristically mani-
fested as concentric rings that kept on increasing in size
until they merged with each other forming an enlarged
necrotic zone which eventually covered the entire leaf and
dried it up within 30 days of inoculation (Fig. 3a, c). This
process of the spread of infection took more than 50 days
in the case of transgenic plants (Fig. 3b, d). The percentage
disease protection in the different transgenic lines com-
pared with the untransformed control plants varied between
68.8 and 85 % (Table 1).
Resistance of Transgenic B. juncea Plants Expressing
msrA1 Gene to S. sclerotiorum
The untransformed control and transgenic B. juncea plants
were infected with S. sclerotiorum as described in the
materials and methods section. The relative severity of the
infection in the plants was compared by determining the
average size of lesion, days to stem breaking, and the
number of sclerotia formed per main stem at the end of
season. The stages in the progression of Sclerotinia
ba
dc
Fig. 3 Phenotype of Brassica
juncea leaves after infection
with Alternaria brassicae. Leaf
from untransformed control
(a) and representative M6
transgenic line (b) infected with
the fungus after 15 days of
inoculation. Leaf from
untransformed control (c) and
M6 transgenic line (d) after
30 days of fungal inoculation
Mol Biotechnol
123

infection in the plants are shown in Fig. 4a–h. Whitish
mycelia were observed, which resulted in soft rot lesions
on the stem. The lesions were significantly larger (average
size 7.4 cm) on the stem of untransformed control plants in
comparison to those on the transgenic plants (average sizes
ranging between 2.1 and 3.3 cm for different lines) as seen
in Fig. 4c, d (Table 2). The mycelial expansion toward
noninfected portions of stem was slower in the transgenic
compared with the untransformed control plants (Fig. 4e,
f). The days taken to stem breaking were much less
(31.4 days) in the untransformed control plants, while the
breaking was delayed (until 41.7–65.3 days) in the differ-
ent transgenic lines (Table 2). The number of sclerotia
formed inside the main stem of the untransformed control
plants was also significantly higher (21.7) than in the
transgenic plants (3.1–8.4) signifying the resistance of the
transgenic plants to the fungus (Table 2; Fig. 4g, h). The
percentage disease protection in the transgenic plants with
respect to the untransformed control varied from 56 to
71.5 %. for the different lines (Table 2).
Discussion
Significant yield losses because of varied stress factors
dampen the productivity potential of agriculture, the world
over. Deteriorating environmental conditions, rendering of
large tracts of lands infertile over the years, losses caused
by pest infestation, and development of resistance in dis-
ease causing pathogens are the major challenges that need
to be surmounted by the agriculture community. Fungal
diseases are rated as the most or the second-most important
ba c d
e f g h
UT UT
UT UT
M6
M6
M6
M6
Fig. 4 Stages in the development and progression of Sclerotinia
sclerotiorum infection on Brassica juncea stem. a, c, e Different
stages in the infection of untransformed (UT) control plants. b, d, f
Different stages in the infection of plants of M6 transgenic line (M6)
expressing the msrA1 gene. g, h Sclerotia bodies formed inside the
main stem of UT and M6 plants
Mol Biotechnol
123

factor contributing to yield loss in some of the economi-
cally important crops [32]. B. juncea is an important oil-
seed crop in many countries of the world. It ranks second,
next to ground nut, in contributing to the Indian oilseed
economy and is expected to supply for the increasing
edible oil demands in the decades to come [22]. Yield
losses due to fungal diseases alternaria leaf spot caused by
A. brassicae and soft stem rot caused by S. sclerotiorum are
a serious problem in the cultivation of this crop [23, 33].
These diseases are difficult to control once they set in and
have been reported to lead to complete crop failure in their
most devastating forms [34, 35]. The conventional strate-
gies of tackling the diseases with fungicides and selecting
for the disease-resistant genotypes through breeding are
fraught with concerns of development of resistant fungal
strains, cost, time, and uncertainty.
In view of the above, transgenic approaches for gener-
ation of fungus-resistant plants is increasingly being
explored as a viable alternative. The strategies for confer-
ring fungal resistance either involve the production of
transgenic plants with antifungal molecules like proteins
and toxins or generation of a hypersensitive response
through R (resistance) genes or by manipulating genes of
the systemic acquired resistance pathway. Some of the
proteins used have been pathogenesis-related proteins,
ribosome-inactivating proteins, small cystein-rich proteins,
lipid transfer proteins, storage albumins, polygalacturonase
inhibitor proteins, chitinases, antiviral proteins, and non-
plant antifungal proteins [32].
In the present article, we explored the prospects of
engineering fungal resistance in B. juncea plants by
transgenic expression of MsrA1, a synthetic cationic anti-
microbial peptide. CAPs from microbial sources often have
significant phytotoxicity that limits their direct use for plant
protection. MsrA1 was previously designed by modifying
CEMA which is a chimeric CAP with bioactive portions of
the much investigated cecropin A and melittin proteins.
The change introduced in the MsrA1 was aimed at cur-
tailing the high antimicrobial activity of CEMA to ward off
the unwarranted toxicity effects on the expressing host
plants [13]. We generated five independent transgenic lines
with 1–2 copies of the integrated transgenes. The integrated
gene was transcriptionally active in all these lines. The
expressions of the gene in the lines M1, M3, and M5 (with
two copies of the transgene) were lower than those in M6
and M7 lines which had single transgene copy. This could
be because copy number might have a negative effect on
the transgene expression due to silencing effects [36]. It is,
therefore, mostly desired to have single-copy transgenic
lines. The observation that despite having two copies of the
gene the M5 line showed high number of days to stem
breaking for S. sclerotiorum infection (in comparison with
the single copy carrying M7 line) which could be due to the
reasons that we cannot authoritatively comment upon
based on the present data. The variation in the expression
(and the consequent biologic effect) in independent trans-
formants could be due to several factors including site of
integration on the chromosome, promoter methylation,
posttranscriptional gene silencing mediated by the pro-
duction of aberrant transcripts, etc. [37]. No unintended
deleterious effects on morphology or physiology of the
transgenic plants were evident. In the previous evaluation
of this gene, Osusky et al. [13] found the disease ‘‘’lesion-
mimick’’ phenotype in a particular potato cultivar, while
this abnormality was not seen in another variety used in
their study as well as in tobacco plants expressing the gene
under the control of pathogen-responsive win3.12T pro-
moter in a subsequent study reported from the same group
[8]. This underlines the importance of assessing the effects
of transgenesis in different species and cultivars on a case-
to-case basis.
The transgenic B. juncea plants were found to be
resistant to both the fungal pathogens tested. In the in vitro
assay, protein extract from the transgenic plants inhibited
the hyphal growth of A. brassicae, while the extract from
the untransformed control or the proteinase K treated
transgenic extract could not do so, suggesting that the
MsrA1 protein present in the transgenic plants was active
against the pathogen. Although, the spread of this fungus in
the untransformed control plants was much faster (the
leaves necrosed in 30 days post infection) as seen in the
in vivo bioassay, it was restricted in the transgenic plants
and the infected leaves got necrosed not before 50 days
after infection. Similarly, protection was also observed
against S. sclerotiorum. The spread of fungus from the site
of infection toward uninfected parts was faster and prolific
in case of the untransformed control in comparison with the
transgenic plants. Also, the number of sclerotia formed
Table 2 Evaluation of Sclerotinia sclerotiorum infection on B.
juncea
Plants Size of
lesion (cm)
(Avg ± SD)
Days to stem
breaking
(Avg ± SD)
No. of Sclerotia
in main stem
(Avg ± SD)
Disease
protection
(% ± SD)
UT 7.4 ± 0.9 31.4 ± 3.6 21.7 ± 1.5 –
M1 3.3 ± 1.1 44.5 ± 1.7 7.6 ± 1.5 56.0 ± 3.5
M3 2.7 ± 1.1 41.7 ± 1.5 8.4 ± 2.5 63.0 ± 2.9
M5 2.4 ± 1.4 58.6 ± 2.9 6.7 ± 1.7 68.1 ± 3.2
M6 2.1 ± 1.4 65.3 ± 2.0 3.1 ± 1.5 71.5 ± 1.6
M7 2.8 ± 1.3 53.4 ± 2.0 5.9 ± 2.0 62.2 ± 2.3
The average size of lesion, days to stem breaking, and the number of
sclerotia formed in the main stem were determined. The percentage
disease protection in the transgenic plants with respect to the
untransformed control plants was determined as described in mate-
rials and methods. The data presented are the average ± SD values of
three replicate experiments
Mol Biotechnol
123

inside the main stem was much less in the transgenic
plants. The restricted spread of these two fungi is expected
to translate into higher yields in the transgenic B. juncea
harvest under field conditions.
In conclusion, our work demonstrates the generation of
an economically important oilseed crop having broad
spectrum resistance against fungal pathogens by transgenic
expression of an antimicrobial peptide. This could poten-
tially reduce the yield loss caused by these pathogens and
also decrease the dependence on fungicides that are
increasingly being wished-off for environmental, health,
and economic reasons.
Acknowledgments This work was funded by Grant (No. BT/
PR9616/AGR/02/458/2007) from the Department of Biotechnology,
India to N.B.S. A.R. and D.K. were recipients of Senior Research
Fellowship from the Council of Science and Industrial Research,
India. M.A.Y. is a U.G.C. Dr. D.S. Kothari Postdoctoral Fellow. The
authors thank Dr. Pratibha Sharma, Division of Plant Pathology,
Indian Agricultural Research Institute, New Delhi, for her suggestions
in fungal infection experiments.
References
1. Food and Agriculture Organization (2010) The state of food
insecurity in the world. http://www.fao.org/docrep/013/i1683e/
i1683e.pdf.
2. Agrios, G. N. (2005). Plant Pathology. London: Academic Press.
3. Ali, G. S., & Reddy, A. S. N. (2000). Inhibition of fungal and
bacterial plant pathogens by synthetic peptides: In vitro growth
inhibition, interaction between peptides and inhibition of disease
progression. Molecular Plant–Microbe Interactions, 13, 847–859.
4. Brown, K. L., & Hancock, R. E. W. (2006). Cationic host defense
(antimicrobial) peptides. Current Opinion in Immunology, 18,
24–30.
5. Hancock, R. E. W., & Lehrer, R. (1998). Cationic peptides: A
new source of antibiotics. Trends in Biotechnology, 16, 82–88.
6. Yount, N. Y., & Yeaman, M. R. (2005). Immunocontinuum:
Perspectives in antimicrobial peptide mechanisms of action and
resistance. Protein and Peptide Letters, 12, 49–67.
7. Hancock, R. E. W. (2005). Mechanisms of action of newer
antibiotics for Gram-positive pathogens. Lancet Infectious Dis-
eases, 5, 209–218.
8. Yevtushenko, D. P., Romero, R., Forward, B. S., Hancock, R. E.,
Kay, W. W., & Misra, S. (2005). Pathogen-induced expression of
a cecropin A melittin antimicrobial peptide gene confers anti-
fungal resistance in transgenic tobacco. Journal of Experimental
Botany, 56, 1685–1695.
9. Koo, J. C., Chun, H. J., Park, H. C., Kim, M. C., Koo, Y. D., Koo,
S. C., et al. (2002). Over-expression of a seed specific hevein-like
antimicrobial peptide from Pharbitis nil enhances resistance to a
fungal pathogen in transgenic tobacco plants. Plant Molecular
Biology, 50, 441–452.
10. Jaynes, J. M., Nagpala, P., Destefanobeltran, L., Huang, J. H.,
Kim, J. H., Denny, T., et al. (2002). Expression of a cecropin-B
lytic peptide analog in transgenic tobacco confers enhanced
resistance to bacterial wilt caused by Pseudomonas solanacea-
rum. Plant Science, 89, 43–53.
11. Huang, Y., Nordeen, R. O., Di, M., Owens, L. D., & McBeath, J.
H. (1997). Expression of an engineered cecropin gene cassette in
transgenic tobacco plants confers resistance to Pseudomonas sy-
ringae pv. tabaci. Phytopathology, 87, 494–499.
12. Sharma, A., Sharma, R., Imamura, M., Yamakawa, M., & Machii,
H. (2000). Transgenic expression of cecropin B, an antibacterial
peptide from Bombyx mori, confers enhanced resistance to bac-
terial leaf blight in rice. FEBS Letters, 484, 7–11.
13. Osusky, M., Zhou, G., Osuska, L., Hancock, R. E., Kay, W. W.,
& Misra, S. (2000). Transgenic plants expressing cationic peptide
chimeras exhibit broad-spectrum resistance to phytopathogens.
Nature Biotechnology, 18, 1162–1166.
14. Gao, A. G., Hakimi, S. M., Mittanck, C. A., Wu, Y., Woerner, B.
M., Stark, D. M., et al. (2000). Fungal pathogen protection in
potato by expression of a plant defensin peptide. Nature Bio-
technology, 18, 1307–1310.
15. Chakrabarti, A., Ganapathi, T. R., Mukherjee, P. K., & Bapat, V. A.
(2003). MSI-99, a magainin analogue, imparts enhanced disease
resistance in transgenic tobacco and banana. Planta, 216, 587–596.
16. Alan, A. R., Blowers, A., & Earle, E. D. (2004). Expression of a
magainin-type antimicrobial peptide gene (MSI-99) in tomato
enhances resistance to bacterial speck disease. Plant Cell
Reports, 22, 388–396.
17. Mentag, R., Luckevich, M., Morency, M. J., & Se0guin, A.
(2003). Bacterial disease resistance of transgenic hybrid poplar
expressing the synthetic antimicrobial peptide D4E1. Tree
Physiology, 23, 405–411.
18. Vidal, J. R., Kikkert, J. R., Malnoy, M. A., Wallace, P. G.,
Barnard, J., & Reisch, B. I. (2006). Evaluation of transgenic
‘Chardonnay’ (Vitis vinifera) containing magainin genes for
resistance to crown gall and powdery mildew. Transgenic
Research, 15, 69–82.
19. Hancock, R. E. W., Brown, M. H. & Piers, K. (1992) Cationic
peptides and method of preparation. US Patent application serial
No. 07/913, 492, filed August 21, 1992.
20. Niknam, S. R., & Turner, D. W. (1999). A single drought event,
at different stages of development, has different effects on the
final yield of Brassica napus cv. Monty and B. juncea line
397-23-2-3-3. In G. Shea, (Ed.), Oilseed Crop Updates (pp.
14–15). Northam: Agriculture Western Australia.
21. Wright, P. R., Morgan, J. M., & Jessop, R. S. (1996). Compar-
ative adaptation of canola (Brassica napus) and Indian mustard
(B. juncea) to soil water deficits: Plant water relations and
growth. Field Crop Research, 49, 51–64.
22. Shekhawat, K., Rathore, S. S., Premi, O. P., Kandpal, B. K., &
Chauhan, J. S. (2012). Advances in agronomic management of
Indian mustard (Brassica juncea (L.) Czernj. Cosson): An over-
view. International Journal Of Agronomy, 2012, 1–14.
23. Mondal, K. K., Bhattacharya, R. C., Koundal, K. R., & Chat-
terjee, S. C. (2007). Transgenic Indian mustard (Brassica juncea)
expressing tomato glucanase leads to arrested growth of Alter-
naria brassicae. Plant Cell Reports, 26, 247–252.
24. Ghasolia, R. P., Shivpuri, A., & Bhargava, A. K. (2004). Scle-
rotinia rot of Indian mustard (Brassica juncea) in Rajasthan.
Indian Phytopathology, 57, 76–79.
25. Hofgen, R., & Willmitzer, L. (1988). Storage of competent cells for
Agrobacterium transformation. Nucleic Acids Research, 16, 9877.
26. Murashige, T., & Skoog, F. (1962). A revised medium for rapid
growth and bio-assay with tobacco tissue cultures. Physiologia
Plantarum, 15, 473–497.
27. Yusuf, M. A., & Sarin, N. B. (2007). Antioxidant value addition
in human diets: genetic transformation of Brassica juncea with
gamma-TMT gene for increased alpha-tocopherol content.
Transgenic Research, 16, 109–113.
28. Murray, H. G., & Thompson, W. F. (1980). Rapid isolation of
high molecular weight DNA. Nucleic Acids Research, 8,
4321–4325.
Mol Biotechnol
123

29. Sambrook, J., Fritsch, E. F., & Maniatis, T. (1989). Molecular
cloning. A laboratory manual. New York: Cold Spring Harbor
Laboratory Press, Cold Spring Harbor.
30. Sharma, N., Rahman, M. H., Strelkov, S., Thiagarajah, M.,
Bansal, V. K., & Kav, N. N. V. (2007). Proteome-level changes
in two Brassica napus lines exhibiting differential responses to
the fungal pathogen Alternaria brassicae. Plant Science, 172,
95–110.
31. Jensen, B. D., Finckh, M. R., Munk, L., & Hauser, T. P. (2008).
Susceptibility of wild carrot (Daucus carota ssp. carota) to
Sclerotinia sclerotiorum. European Journal of Plant Pathology,
122, 359–367.
32. Grover, A., & Gowthaman, R. (2003). Strategies for development
of fungus-resistant transgenic plants. Current Science, 84,
330–340.
33. Ghasolia, R. P., & Shivpuri, A. (2007). Management of Scle-
rotinia rot of Indian mustard through cultural practices. Journal of
Mycology and Plant Pathology, 37, 244–247.
34. Kang, I. S., & Chahal, S. S. (2000). Prevalence and incidence of
white rot of mustard incited by Sclerotinia sclerotiorum in Pun-
jab. Plant Disease Research, 15, 232–233.
35. Sharma, S., Yadav, J. L., & Sharma, G. R. (2001). Effect of
various agronomic practices on the incidence of white rot of
Indian mustard caused by Sclerotinia sclerotiorum. Journal of
Mycology and Plant Pathology, 31, 83–84.
36. Finnegan, J., & McElroy, D. (1994). Transgene inactivation:
Plants fight back. Biotechnology, 12, 883–888.
37. Day, C. D., Lee, E., Kobayashi, J., Holappa, L. D., Albert, H., &
Ow, D. W. (2000). Transgene integration into the same chro-
mosome location can produce alleles that express at a predictable
level, or alleles that are differentially silenced. Genes & Devel-
opment, 14, 2869–2880.
Mol Biotechnol
123





























