Embed Size (px)
Citation preview

Chapter 11Trends of the Herbs Ecological Evolution
Sergei N. Sheremet’ev and Yuri V. Gamalei
Abstract The results of analytic research show that the evolution of leaf structureand water balance are completely coincident to global changes of planet climateand hydrology. Taxonomical diversity of herbs and herbaceous biomes is thefunction of paleoclimate variability and plant adaptogenesis to it. Two globaltrends of ecological evolution contrast differing by the composition of herbaceousadaptive types is the next: (a) the line of herbs of chilling plains with dominationthe group of plant species with C3 apoplastic syndrome formed under cold climateinfluence, and (b) the line of herbs of hot plains with domination of plant specieswith C4 apoplastic syndrome. Both trends include the monocots and dicots, andboth are the results of climate changes in Cenozoic. C3 herbs of chilling plains andthe steppe and meadow phytocoenosis formed by them arise as the answer totemperature decrease in great areas of high latitudes. The apoplastic syndrome(transfer from symplastic transport of assimilates suppressed by cold to theirapoplastic transport) is the diagnostic test for this group of herbs. C4 herbs of hotplains and the savanna, desert and solontchak plant vegetation are the adaptiveanswer to aridization of low latitude areas. C4 syndrome (compensation of stomataclosure by the mechanism of CO2 concentration in the leaf tissues) is a special signof this group of herbs. Both types of herbaceous biomes come to change forestbiomes which were strongly decreased in both areas, at low and high latitudes.This tendency is continued in parallels with climate tendency to continentdesiccation and cooling.
S. N. Sheremet’ev (&) � Y. V. GamaleiKomarov Botanical Institute of the Russian Academy of Sciences,Prof. Popov Street 2, St. Petersburg 197376, Russiae-mail: [email protected]
P. Pontarotti (ed.), Evolutionary Biology: Mechanisms and Trends,DOI: 10.1007/978-3-642-30425-5_11, � Springer-Verlag Berlin Heidelberg 2012
189

11.1 Introduction
Last years have appeared and intensively developed databases on the Internet,containing the information on plant taxa age—time of their earliest appearance infossil record. There are data for many families, and in some cases for genera ofangiosperms. The fullest and informative database on the Internet, in our opinion,is the Paleobiology database (PBDB) (http://paleodb.org/cgi-bin/bridge.pl).Possibility to establish age of herbs genera which were earlier objects of researchesof the structural–functional organization (Gamalei 2000) and water relations(Sheremet’ev 2005), has allowed to track changes of structural–functionalproperties of plants in time and to compare this time trends with some majorpaleoclimate changes. Being engaged in this work, we understood neverthelesssome vagueness of such constructions in connection with incomplete clarificationof genera average functioning parameters, and also the data about genera age.However this question, in our opinion, is interesting and demands the preliminaryanalysis and discussion at least.
Global climatic changes, as a rule, are considered in connection with theirinfluence on large biosphere reorganizations and biotic events (Briggs andCrowther 1997; Culver and Rawson 2000; Semikhatov and Chumakov 2004, andothers). Maybe that these changes have left a trace in less considerable manifes-tations, such, as structure and functioning of plant leaves.
11.2 Geological Ages of Some Herbs Generaand Their Functioning
Representatives of the genera whose ages are known (Table 11.1) were the objectsof comparative structural and functional studies (Gamalei 2000; Sheremet’ev2005). Assuming that structural and functional characters have remainedunchanged from the moment when the given taxon arose (Gamalei et al. 2008),one can use the data on the studied genera to construct the time series of the plantstructure and water relations parameters reflecting the accessibility of water for theplants and to trace their relationships with drastic global climatic changes. Weselected the following characters: the transpiration rate, water content of theleaves, partial volume of intercellular spaces in the mesophyll, and water satura-tion deficit (Fig. 11.1). Correlations of these characters with the environmentalmoisture vector have been confirmed by comparative studies along the contem-porary gradients of soil moisture (Sheremet’ev 2005).
The curves of the mean values of the studied characters plotted as functions ofthe genus ages show interrelated variation patterns (Fig. 11.1). The similarity ofthe trends of transpiration rate and water content of the leaves is undoubted; bothparameters increased during the Cretaceous, peaked during the Paleocene, beganto decrease in the late Eocene, and reached the minimum in the Oligocene. Next
190 S. N. Sheremet’ev and Y. V. Gamalei
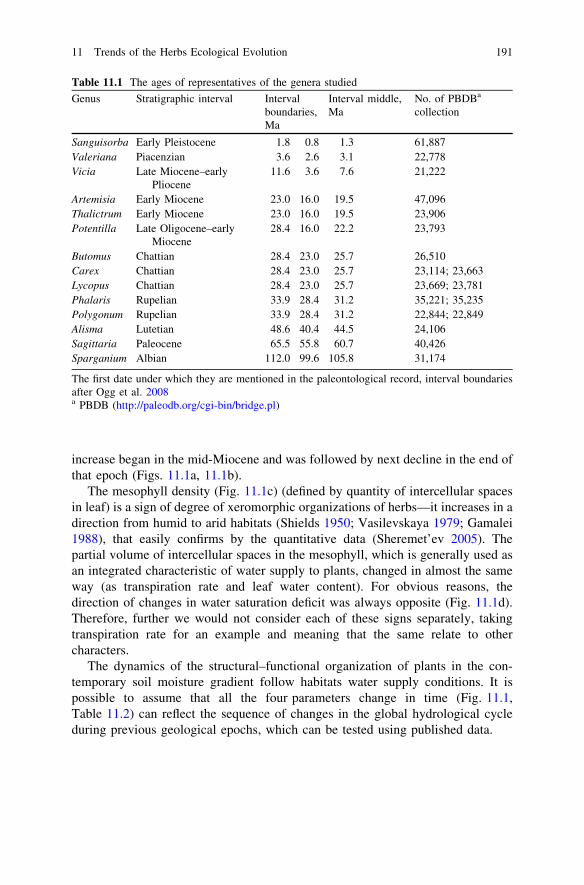
increase began in the mid-Miocene and was followed by next decline in the end ofthat epoch (Figs. 11.1a, 11.1b).
The mesophyll density (Fig. 11.1c) (defined by quantity of intercellular spacesin leaf) is a sign of degree of xeromorphic organizations of herbs—it increases in adirection from humid to arid habitats (Shields 1950; Vasilevskaya 1979; Gamalei1988), that easily confirms by the quantitative data (Sheremet’ev 2005). Thepartial volume of intercellular spaces in the mesophyll, which is generally used asan integrated characteristic of water supply to plants, changed in almost the sameway (as transpiration rate and leaf water content). For obvious reasons, thedirection of changes in water saturation deficit was always opposite (Fig. 11.1d).Therefore, further we would not consider each of these signs separately, takingtranspiration rate for an example and meaning that the same relate to othercharacters.
The dynamics of the structural–functional organization of plants in the con-temporary soil moisture gradient follow habitats water supply conditions. It ispossible to assume that all the four parameters change in time (Fig. 11.1,Table 11.2) can reflect the sequence of changes in the global hydrological cycleduring previous geological epochs, which can be tested using published data.
Table 11.1 The ages of representatives of the genera studied
Genus Stratigraphic interval Intervalboundaries,Ma
Interval middle,Ma
No. of PBDBa
collection
Sanguisorba Early Pleistocene 1.8 0.8 1.3 61,887Valeriana Piacenzian 3.6 2.6 3.1 22,778Vicia Late Miocene–early
Pliocene11.6 3.6 7.6 21,222
Artemisia Early Miocene 23.0 16.0 19.5 47,096Thalictrum Early Miocene 23.0 16.0 19.5 23,906Potentilla Late Oligocene–early
Miocene28.4 16.0 22.2 23,793
Butomus Chattian 28.4 23.0 25.7 26,510Carex Chattian 28.4 23.0 25.7 23,114; 23,663Lycopus Chattian 28.4 23.0 25.7 23,669; 23,781Phalaris Rupelian 33.9 28.4 31.2 35,221; 35,235Polygonum Rupelian 33.9 28.4 31.2 22,844; 22,849Alisma Lutetian 48.6 40.4 44.5 24,106Sagittaria Paleocene 65.5 55.8 60.7 40,426Sparganium Albian 112.0 99.6 105.8 31,174
The first date under which they are mentioned in the paleontological record, interval boundariesafter Ogg et al. 2008a PBDB (http://paleodb.org/cgi-bin/bridge.pl)
11 Trends of the Herbs Ecological Evolution 191
11.3 Late Cretaceous–Cenozoic Climateand Plants Functioning
11.3.1 Late Cretaceous–Eocene
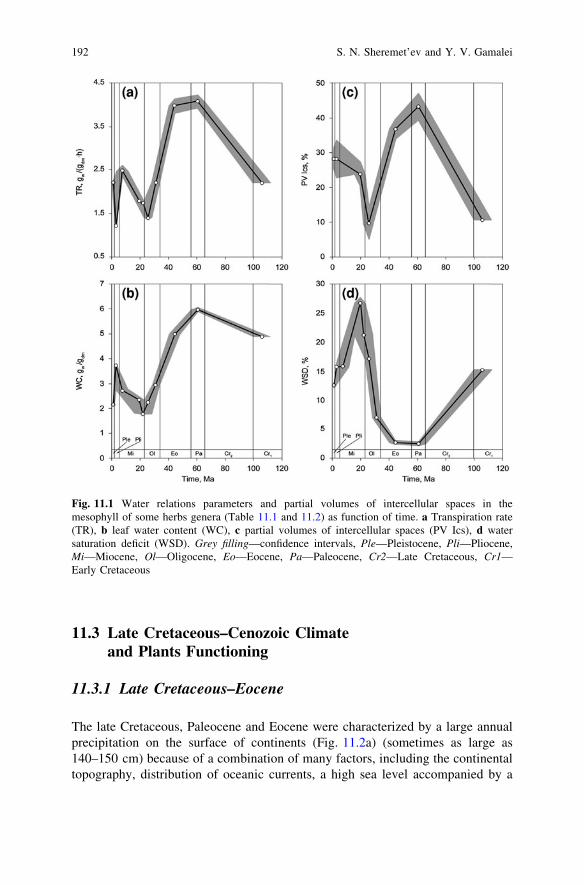
The late Cretaceous, Paleocene and Eocene were characterized by a large annualprecipitation on the surface of continents (Fig. 11.2a) (sometimes as large as140–150 cm) because of a combination of many factors, including the continentaltopography, distribution of oceanic currents, a high sea level accompanied by a
Fig. 11.1 Water relations parameters and partial volumes of intercellular spaces in themesophyll of some herbs genera (Table 11.1 and 11.2) as function of time. a Transpiration rate(TR), b leaf water content (WC), c partial volumes of intercellular spaces (PV Ics), d watersaturation deficit (WSD). Grey filling—confidence intervals, Ple—Pleistocene, Pli—Pliocene,Mi—Miocene, Ol—Oligocene, Eo—Eocene, Pa—Paleocene, Cr2—Late Cretaceous, Cr1—Early Cretaceous
192 S. N. Sheremet’ev and Y. V. Gamalei

Tab
le11
.2T
rans
pira
tion
rate
(TR
,gw
ater
/gdry
mas
s�h),
wat
ersa
tura
tion
defi
cit(
WS
D,%
),le
afw
ater
cont
ent(
LW
C,g
wat
er/g
dry
mas
s)fo
rre
pres
enta
tive
sof
this
stud
y
Gen
usA
ge,
Ma
TR
WS
DL
WC
n� X
SxC
Ln
� XSx
CL
n� X
SxC
L
Sang
uiso
rba
1.3
620
2.21
0.90
0.07
620
12.6
4.7
0.37
620
2.15
0.38
0.03
Val
eria
na3.
122
81.
220.
450.
0622
615
.83.
80.
5022
83.
720.
490.
06V
icia
7.6
290
2.48
1.13
0.13
288
15.8
4.4
0.51
290
2.72
0.49
0.06
Art
emis
ia,
Tha
lict
rum
19.5
1,12
11.
791.
260.
071,
057
26.7
15.9
0.96
1,36
02.
350.
740.
04P
oten
till
a22
.292
01.
740.
930.
0692
021
.26.
50.
4292
01.
790.
300.
02B
utom
us,
Car
ex,
Lyc
opus
25.7
2,00
61.
400.
830.
041,
954
17.2
10.5
0.47
2,00
62.
241.
280.
06P
hala
ris,
Pol
ygon
um31
.257
02.
211.
200.
1057
07
2.1
0.17
570
2.94
0.85
0.07
Ali
sma
44.5
138
3.98
1.01
0.17
138
2.7
1.8
0.29
138
4.98
0.56
0.09
Sagi
ttar
ia60
.713
84.
080.
930.
1613
82.
51.
70.
2913
85.
970.
600.
10Sp
arga
nium
105.
813
82.
200.
680.
1113
815
.24.
60.
7713
84.
890.
470.
08
n—nu
mbe
rof
obse
rvat
ions
,� X
—av
erag
es,
Sx—
stan
dard
devi
atio
ns,
CL
—co
nfide
nce
leve
ls(P
=0.
95)
11 Trends of the Herbs Ecological Evolution 193
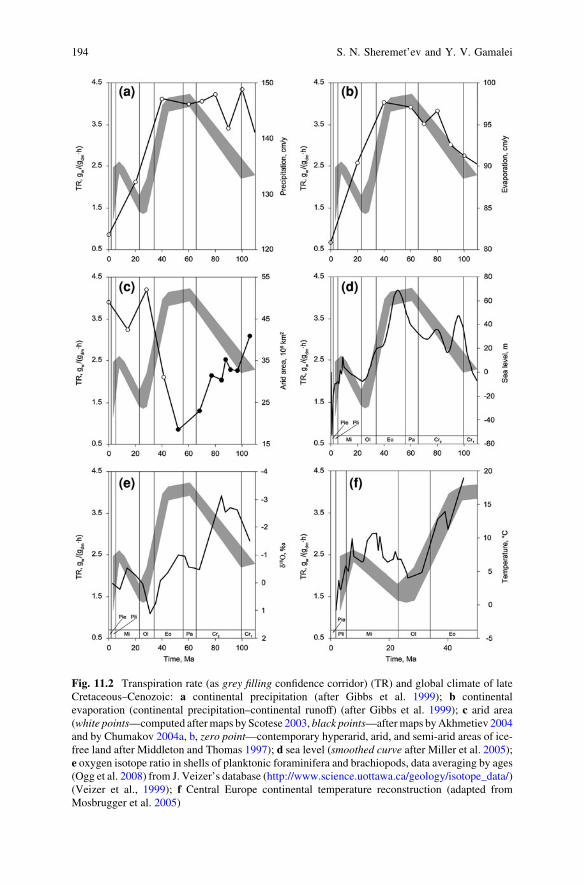
Fig. 11.2 Transpiration rate (as grey filling confidence corridor) (TR) and global climate of lateCretaceous–Cenozoic: a continental precipitation (after Gibbs et al. 1999); b continentalevaporation (continental precipitation–continental runoff) (after Gibbs et al. 1999); c arid area(white points—computed after maps by Scotese 2003, black points—after maps by Akhmetiev 2004and by Chumakov 2004a, b, zero point—contemporary hyperarid, arid, and semi-arid areas of ice-free land after Middleton and Thomas 1997); d sea level (smoothed curve after Miller et al. 2005);e oxygen isotope ratio in shells of planktonic foraminifera and brachiopods, data averaging by ages(Ogg et al. 2008) from J. Veizer’s database (http://www.science.uottawa.ca/geology/isotope_data/)(Veizer et al., 1999); f Central Europe continental temperature reconstruction (adapted fromMosbrugger et al. 2005)
194 S. N. Sheremet’ev and Y. V. Gamalei
high temperature of the ocean and land surface, etc (Gibbs et al. 1999). Thisgeological time is characterized as a ‘‘warm biosphere’’ (or a «warm mode» –Frakes et al. 2005) exceptionally favorable for plants (Chumakov 1993, 1997,2004a, b; Akhmetiev 2004). The thermal optimum was accompanied by largeatmospheric precipitation and a comparatively high atmospheric carbon dioxideconcentration (Tajika 1999; Berner and Kothavala 2001; Beerling and Royer 2002;Hansen and Wallmann 2003; Wallmann 2004; Berner 2006; Royer 2006, andothers), however considerable territories of a land have been occupied by arid belts(Chumakov 1997, 2004a, b; Zharkov et al. 2004; Scotese 2003) (Fig. 11.2c).
Often late Cretaceous–Eocene describes as «greenhouse world» (Beerling andWoodward 2001; Zachos et al. 2001; Retallack 2004; Miller et al. 2005; Pearsonet al. 2007). For example, in Eocene (Lutetian) woods in the conditions of enoughwarm and damp climates in high latitudes behind polar circle (70–80�N) grew(Jahren 2007). On this background, a number of cold events (geologically short-term ones—up to 100 thousand years: Miller et al. 2005) are detected (we meanrelatively cold events; temperatures were, possibly, on some degrees abovemodern). Cretaceous climate cooling up to end of Maastrichtian and then in middleEocene up to Oligocene beginning, nevertheless, were continued (Beerling andWoodward 2001; Zachos et al. 2001, and others) (Fig. 11.2e).
The estimation of the structural and functional parameters of plants of the lateCretaceous–Eocene as typical of mesophytes (and maybe hydrophytes) is
0.0
0.2
0.4
0.6
0.8
1.0
0 20 40 60 80 100 120
Time, Ma
Evo
lutio
n ra
te, f
amili
es/M
a
0
5
10
15
20
25
Are
a, ×
106 k
m2
Evolution rate
Area
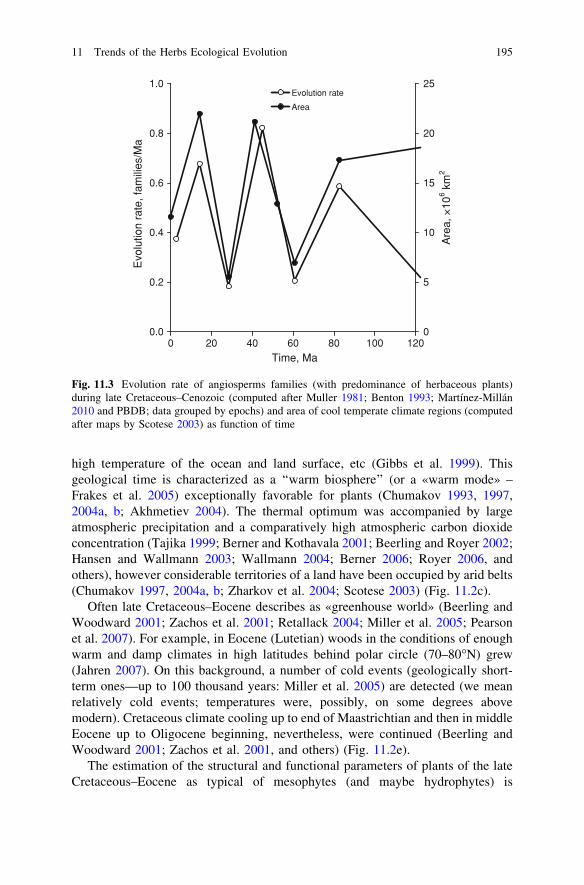
Fig. 11.3 Evolution rate of angiosperms families (with predominance of herbaceous plants)during late Cretaceous–Cenozoic (computed after Muller 1981; Benton 1993; Martínez-Millán2010 and PBDB; data grouped by epochs) and area of cool temperate climate regions (computedafter maps by Scotese 2003) as function of time
11 Trends of the Herbs Ecological Evolution 195

suggested (loose mesophyll promotes high transpiration rate, to accumulation of aconsiderable quantity of water in leaves and to low water saturation deficit)(Fig. 11.1). It is confirmed by the coincidence of the curves of the transpirationrate and global continental precipitation (Fig. 11.2a), and especially waterevaporation from the continent surface (Fig. 11.2b) in this time.
During the Cretaceous–early Eocene, the arid areas considerably decreased(Fig. 11.2c), that corresponds to assumptions about high activity of a hydrologicalcycle in this time (Figs. 11.2a, 11.2b), high sea level (Fig. 11.2d), increasing totalarea of temperate (warm and cool) regions (Scotese 2003), herbs as mesophytes,and hydrophytes functioning (Fig. 11.1).
In the beginning of Eocene, the forest vegetation prevailed in the world andonly in the end of this epoch have appeared and started to develop grasslands inEurasia (Bredenkamp et al. 2002). Therefore, the Paleocene–Eocene beginning ofherbs advancement in an area with cool and warm moderate climates could occuras a part of woody (or shrubby) communities. Perhaps, development and evolutionof herb vegetation mainly was connected with cool temperate climate. Evolutionrate of angiosperm families with predominating of herbs (as number of familiesappeared in some geological epoch divided by duration of this epoch) close cor-responding with Cenozoic dynamics of this climate (Fig. 11.3). Paleocene andEocene dynamics of cool temperate climate was determined by antiphase changesof warm temperate climate, but the next (Oligocene–Pleistocene) cool temperateclimate changes was connected with antiphase dynamics of arid climate. Wideexpansion of herbs ecosystems (as biome) became the general tendency only in aMiocene (see below).
11.3.2 Oligocene
A slow decrease in temperature after the early Eocene climatic optimum wasfollowed by a drastic climatic cooling and large-scale glaciation of the Antarctica(Kennett 1977; Lear et al. 2000; Zachos et al. 2001; DeConto and Pollard 2003a, b;Pollard and DeConto 2003, 2005). Somewhat later, glaciation spread overGreenland (later Eocene–early Oligocene: Eldrett et al. 2007). After that, sym-metric glaciation of both poles occurred (Tripati et al. 2005; Moran et al. 2006).The temperature gradient between the equatorial zone and the poles increased(Nikolaev et al. 1998). The transition from the ‘‘warm biosphere’’ to the ‘‘coldbiosphere’’ was begun (Akhmetiev 2004).
The Oligocene climatic cooling (Figs. 11.2e, 11.2f), which is considered one ofthe most important events in the climatic record (DeConto and Pollard 2003a, b),was accompanied by a decrease in the atmospheric carbon dioxide concentration(Pagani et al. 2005), decrease in precipitation (Fig. 11.2a) and evaporation(Fig. 11.2b), growing aridity of continents (Fig. 11.2c), and lowering the sea level(Fig. 11.2d). Evolution rate of herbaceous plants was decreased up to minimallevel (Fig. 11.3) perhaps as a result of climate cooling, and in consequence of the
196 S. N. Sheremet’ev and Y. V. Gamalei
Tab
le11
.3T
hefi
rst
appe
aran
cean
dex
pans
ion
ofhe
rbbi
omes
ondi
ffer
ent
cont
inen
ts
Fir
stap
pear
ance
Exp
ansi
onR
efer
ence
C3/C
4C
omm
ent
Nor
thA
mer
ica
Ear
lyO
ligo
cene
(des
ert
herb
ecos
yste
ms)
Lat
eM
ioce
neR
etal
lack
(199
7,20
04)
Not
spec
ified
Pal
eoso
ls
Ear
lyO
ligo
cene
Lat
eO
ligo
cene
orea
rly
Mio
cene
Str
ömbe
rg(2
005)
C3
Col
orad
o,N
ebra
ska,
Wyo
min
g,M
onta
na,
Idah
o.P
hyto
lith
asse
mbl
ages
Lat
eO
ligo
cene
–ea
rly
Mio
cene
Lat
eM
ioce
neR
etal
lack
(199
8)N
otsp
ecifi
edP
aleo
sols
Lat
eO
ligo
cene
-ea
rly
Mio
cene
Str
ömbe
rg(2
002,
2004
)C
3N
ebra
ska.
Phy
toli
thas
sem
blag
es
Ear
lyM
ioce
ne(s
hort
-gra
sspr
airi
e)
Lat
eM
ioce
neR
etal
lack
(199
7,20
04)
Not
spec
ified
Pal
eoso
ls
Ear
lyM
ioce
neL
ate
Mio
cene
Mac
Fad
den
(199
7,20
00)
C3
and
C4
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
s.C
3he
rbbi
omes
befo
reex
pans
ion
ofC
4he
rbbi
omes
inth
ela
teM
ioce
neE
arly
Mio
cene
Lat
eM
ioce
neJa
nis
etal
.(2
000,
2004
);Ja
nis
(200
7)N
otsp
ecifi
edD
iscu
ssio
nof
the
lite
rary
data
Ear
lyM
ioce
neL
ate
Mio
cene
–ear
lyP
lioc
ene
Fox
and
Koc
h(2
003,
2004
)C
4G
reat
plai
ns.
Pal
eoso
ls.
Car
bon
isot
opes
Ear
lyM
ioce
neS
tröm
berg
(200
6)C
3C
olor
ado,
Neb
rask
a,W
yom
ing,
Mon
tana
,Id
aho.
Phy
toli
thas
sem
blag
esM
iddl
eM
ioce
neF
oxan
dK
och
(200
3)C
3G
reat
plai
ns.
Pal
eoso
ls.
Car
bon
isot
opes
Mid
dle
Mio
cene
–lat
eM
ioce
neL
ate
Mio
cene
–ear
lyP
lioc
ene
Kel
logg
(200
1)C
4D
iscu
ssio
nof
the
lite
rary
data
Lat
eM
ioce
neL
ate
Mio
cene
Cer
ling
etal
.(1
997)
C4
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
s.
(con
tinu
ed)
11 Trends of the Herbs Ecological Evolution 197
Tab
le11
.3(c
onti
nued
)
Fir
stap
pear
ance
Exp
ansi
onR
efer
ence
C3/C
4C
omm
ent
Lat
eM
ioce
neP
lioc
ene
Axe
lrod
(198
5)N
otsp
ecifi
edP
aleo
bota
nyda
taL
ate
Mio
cene
Pli
ocen
e–P
leis
toce
neT
rave
rse
(200
7)N
otsp
ecifi
edP
aleo
paly
nolo
gyL
ate
Mio
cene
And
erso
n(2
006)
C4
Dis
cuss
ion
ofth
eli
tera
ryda
taL
ate
Mio
cene
Mac
Fad
den
(200
5)C
4T
ooth
mor
phol
ogy
and
carb
onis
otop
esof
toot
hen
amel
offo
ssil
hors
esL
ate
Mio
cene
Mac
Fad
den
and
Cer
ling
(199
4)C
4C
arbo
nis
otop
esof
toot
hen
amel
offo
ssil
herb
ivor
em
amm
als
Lat
eM
ioce
ne(t
allg
rass
prai
rie)
Ret
alla
cket
al.
(200
2)N
otsp
ecifi
edP
aleo
sols
Lat
eM
ioce
neW
ang
etal
.(1
994)
C4
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
sL
ate
Mio
cene
–ear
lyP
lioc
ene
Cer
ling
etal
.(1
998)
C4
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
sE
uras
iaL
ate
Eoc
ene,
Oli
goce
neB
rede
nkam
pet
al.
(200
2)N
otsp
ecifi
edD
iscu
ssio
nof
the
lite
rary
data
Ear
lyM
ioce
neL
ate
Mio
cene
Str
ömbe
rget
al.
(200
7)C
3T
urke
y,G
reec
e,Ir
an.
Phy
toli
ths.
Car
bon
isot
opes
ofpa
leos
ols
and
ofto
oth
enam
els
ofm
amm
als
Mid
dle
Mio
cene
Lat
eM
ioce
neH
oorn
etal
.(2
000)
C3.
C4—
part
iall
yC
entr
alN
epal
.P
aleo
paly
nolo
gyL
ate
Mio
cene
Lat
eM
ioce
neC
erli
nget
al.
(199
7)C
4P
akis
tan.
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
sL
ate
Mio
cene
Lat
eM
ioce
neJa
nis
etal
.(2
000,
2004
);Ja
nis
(200
7)N
otsp
ecifi
edD
iscu
ssio
nof
the
lite
rary
data
Lat
eM
ioce
neL
ate
Mio
cene
Mor
gan
etal
.19
94C
4P
akis
tan.
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
sL
ate
Mio
cene
–ear
lyP
lioc
ene
Cer
ling
etal
.(1
998)
C4
Pak
ista
n.C
arbo
nis
otop
esof
toot
hen
amel
offo
ssil
herb
ivor
em
amm
als (con
tinu
ed)
198 S. N. Sheremet’ev and Y. V. Gamalei
Tab
le11
.3(c
onti
nued
)
Fir
stap
pear
ance
Exp
ansi
onR
efer
ence
C3/C
4C
omm
ent
Thr
eeba
sic
epis
odes
ofec
osys
tem
deve
lopm
ent
(see
com
men
t)
Wan
get
al.
(200
6)N
otsp
ecifi
edC
hina
.L
oess
Pla
teau
.P
aleo
paly
nolo
gy.
Lat
eM
ioce
ne(6
.2–5
.8M
a)–s
tepp
e,la
teM
ioce
ne–e
arly
Pli
ocen
e(5
.8–
4.2
Ma)
expa
nsio
nof
fore
stve
geta
tion
,ea
rly
Pli
ocen
e–la
teP
lioc
ene
(4.2
–2.4
Ma)
—st
eppe
Lat
eM
ioce
neE
arly
Pli
ocen
eQ
uade
etal
.(1
989)
C4
Pak
ista
n.P
aleo
sols
.C
arbo
nis
otop
esL
ate
Mio
cene
Ear
lyP
lioc
ene
Ret
alla
ck(1
998)
Not
spec
ified
Pak
ista
n.P
aleo
sols
Lat
eM
ioce
neM
erce
ron
etal
.(2
004)
C3
Afg
hani
stan
.T
ooth
mor
phol
ogy
offo
ssil
mam
mal
sP
lioc
ene
Pli
ocen
eD
ing
and
Yan
g(2
000)
C4
Chi
na.
Loe
ssP
late
au.
Pal
eoso
ls.
Car
bon
isot
opes
Pli
ocen
eP
lioc
ene
Wan
gan
dD
eng
(200
5)C
4C
hina
.T
ibet
anP
late
au.
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
sP
lioc
ene–
Ple
isto
cene
Tra
vers
e(2
007)
Not
spec
ified
Pal
eopa
lyno
logy
Thr
eeep
isod
esof
herb
ecos
yste
mex
pans
ion
(see
com
men
t)
Zhi
shen
get
al.
(200
5)C
4C
hina
.L
oess
Pla
teau
.P
aleo
sols
.C
arbo
nis
otop
es.
Pli
ocen
e(2
.9–2
.7M
a),
Ple
isto
cene
(1.3
–0.9
Ma)
,P
leis
toce
ne–u
okow
ey(0
.6–0
Ma)
Sout
hA
mer
ica
Lat
eO
ligo
cene
Lat
eM
ioce
neJa
nis
etal
.(2
000,
2004
);Ja
nis(
2007
)N
otsp
ecifi
edD
iscu
ssio
nof
the
lite
rary
data
Lat
eO
ligo
cene
Lat
eM
ioce
neM
acF
adde
n(2
000)
C3
and
C4
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
s.C
3he
rbbi
omes
befo
reex
pans
ion
ofC
4he
rbbi
omes
inth
ela
teM
ioce
ne
(con
tinu
ed)
11 Trends of the Herbs Ecological Evolution 199
Tab
le11
.3(c
onti
nued
)
Fir
stap
pear
ance
Exp
ansi
onR
efer
ence
C3/C
4C
omm
ent
Ear
lyM
ioce
neL
ate
Mio
cene
Mac
Fad
den
(199
7)C
3an
dC
4C
arbo
nis
otop
esof
toot
hen
amel
offo
ssil
herb
ivor
em
amm
als.
C3
herb
biom
esbe
fore
expa
nsio
nof
C4
herb
biom
esin
the
late
Mio
cene
Mid
dle
Mio
cene
–lat
eM
ioce
neL
ate
Mio
cene
–ear
lyP
lioc
ene
Kel
logg
(200
1)C
4D
iscu
ssio
nof
the
lite
rary
data
Lat
eM
ioce
neL
ate
Mio
cene
Cer
ling
etal
.(1
997)
C4
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
sL
ate
Mio
cene
Lat
eM
ioce
neL
ator
reet
al.
(199
7)C
4A
rgen
tina
.C
arbo
nis
otop
esof
toot
hen
amel
offo
ssil
herb
ivor
em
amm
als
and
ofpa
leos
olca
rbon
ates
Lat
eM
ioce
neM
acF
adde
net
al.
(199
6)C
4A
rgen
tina
.C
arbo
nis
otop
esof
toot
hen
amel
offo
ssil
herb
ivor
em
amm
als
Lat
eM
ioce
ne–e
arly
Pli
ocen
eC
erli
nget
al.
(199
8)C
4C
arbo
nis
otop
esof
toot
hen
amel
offo
ssil
herb
ivor
em
amm
als
Afr
ica
Oli
goce
neO
ligo
cene
Bre
denk
amp
etal
.(2
002)
Not
spec
ified
Dis
cuss
ion
ofth
eli
tera
ryda
taE
arly
Mio
cene
Lat
eM
ioce
neR
etal
lack
(199
8,20
01)
Not
spec
ified
Eas
tA
fric
a.K
enya
.P
aleo
sols
Ear
lyM
ioce
neC
oppe
nsan
dP
ickf
ord
(200
2)N
otsp
ecifi
edE
ast
Afr
ica.
Uga
nda.
Pal
eobo
tany
Mid
dle
Mio
cene
Lat
eM
ioce
neM
orga
net
al.
(199
4)C
4E
ast
Afr
ica.
Ken
ya.
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
sM
iddl
eM
ioce
neL
ate
Mio
cene
Jaco
bs(2
004)
C4
Tro
pica
lAfr
ica
sava
nna.
Pal
eopa
lyno
logy
and
carb
onis
otop
esM
iddl
eM
ioce
neL
ate
Mio
cene
–ear
lyP
lioc
ene
Jani
set
al.
(200
0,20
04);
Jani
s(2
007)
Not
spec
ified
Dis
cuss
ion
ofth
eli
tera
ryda
ta
(con
tinu
ed)
200 S. N. Sheremet’ev and Y. V. Gamalei
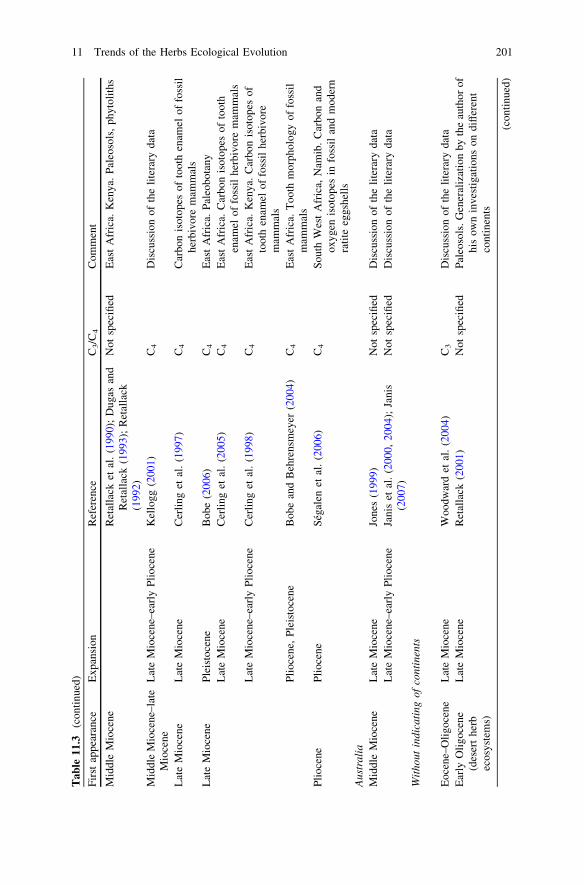
Tab
le11
.3(c
onti
nued
)
Fir
stap
pear
ance
Exp
ansi
onR
efer
ence
C3/C
4C
omm
ent
Mid
dle
Mio
cene
Ret
alla
cket
al.
(199
0);
Dug
asan
dR
etal
lack
(199
3);
Ret
alla
ck(1
992)
Not
spec
ified
Eas
tA
fric
a.K
enya
.P
aleo
sols
,ph
ytol
iths
Mid
dle
Mio
cene
–lat
eM
ioce
neL
ate
Mio
cene
–ear
lyP
lioc
ene
Kel
logg
(200
1)C
4D
iscu
ssio
nof
the
lite
rary
data
Lat
eM
ioce
neL
ate
Mio
cene
Cer
ling
etal
.(1
997)
C4
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
sL
ate
Mio
cene
Ple
isto
cene
Bob
e(2
006)
C4
Eas
tA
fric
a.P
aleo
bota
nyL
ate
Mio
cene
Cer
ling
etal
.(2
005)
C4
Eas
tA
fric
a.C
arbo
nis
otop
esof
toot
hen
amel
offo
ssil
herb
ivor
em
amm
als
Lat
eM
ioce
ne–e
arly
Pli
ocen
eC
erli
nget
al.
(199
8)C
4E
ast
Afr
ica.
Ken
ya.
Car
bon
isot
opes
ofto
oth
enam
elof
foss
ilhe
rbiv
ore
mam
mal
sP
lioc
ene,
Ple
isto
cene
Bob
ean
dB
ehre
nsm
eyer
(200
4)C
4E
ast
Afr
ica.
Too
thm
orph
olog
yof
foss
ilm
amm
als
Pli
ocen
eP
lioc
ene
Ség
alen
etal
.(2
006)
C4
Sou
thW
est
Afr
ica,
Nam
ib.
Car
bon
and
oxyg
enis
otop
esin
foss
ilan
dm
oder
nra
tite
eggs
hell
sA
ustr
alia
Mid
dle
Mio
cene
Lat
eM
ioce
neJo
nes
(199
9)N
otsp
ecifi
edD
iscu
ssio
nof
the
lite
rary
data
Lat
eM
ioce
ne–e
arly
Pli
ocen
eJa
nis
etal
.(2
000,
2004
);Ja
nis
(200
7)N
otsp
ecifi
edD
iscu
ssio
nof
the
lite
rary
data
Wit
hout
indi
cati
ngof
cont
inen
ts
Eoc
ene–
Oli
goce
neL
ate
Mio
cene
Woo
dwar
det
al.
(200
4)C
3D
iscu
ssio
nof
the
lite
rary
data
Ear
lyO
ligo
cene
(des
ert
herb
ecos
yste
ms)
Lat
eM
ioce
neR
etal
lack
(200
1)N
otsp
ecifi
edP
aleo
sols
.Gen
eral
izat
ion
byth
eau
thor
ofhi
sow
nin
vest
igat
ions
ondi
ffer
ent
cont
inen
ts
(con
tinu
ed)
11 Trends of the Herbs Ecological Evolution 201
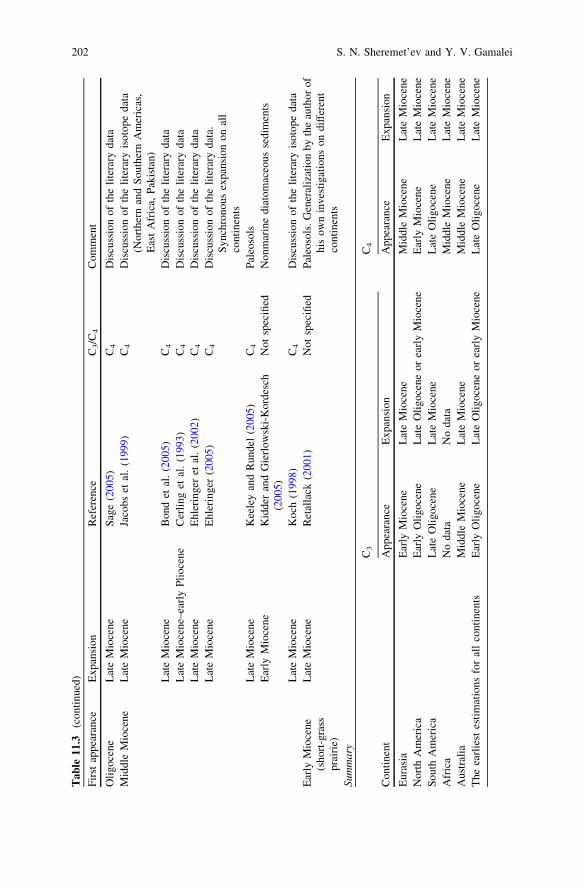
Tab
le11
.3(c
onti
nued
)
Fir
stap
pear
ance
Exp
ansi
onR
efer
ence
C3/C
4C
omm
ent
Oli
goce
neL
ate
Mio
cene
Sag
e(2
005)
C4
Dis
cuss
ion
ofth
eli
tera
ryda
taM
iddl
eM
ioce
neL
ate
Mio
cene
Jaco
bset
al.
(199
9)C
4D
iscu
ssio
nof
the
lite
rary
isot
ope
data
(Nor
ther
nan
dS
outh
ern
Am
eric
as,
Eas
tA
fric
a,P
akis
tan)
Lat
eM
ioce
neB
ond
etal
.(2
005)
C4
Dis
cuss
ion
ofth
eli
tera
ryda
taL
ate
Mio
cene
–ear
lyP
lioc
ene
Cer
ling
etal
.(1
993)
C4
Dis
cuss
ion
ofth
eli
tera
ryda
taL
ate
Mio
cene
Ehl
erin
ger
etal
.(2
002)
C4
Dis
cuss
ion
ofth
eli
tera
ryda
taL
ate
Mio
cene
Ehl
erin
ger
(200
5)C
4D
iscu
ssio
nof
the
lite
rary
data
.S
ynch
rono
usex
pans
ion
onal
lco
ntin
ents
Lat
eM
ioce
neK
eele
yan
dR
unde
l(2
005)
C4
Pal
eoso
lsE
arly
Mio
cene
Kid
der
and
Gie
rlow
ski-
Kor
desc
h(2
005)
Not
spec
ified
Non
mar
ine
diat
omac
eous
sedi
men
ts
Lat
eM
ioce
neK
och
(199
8)C
4D
iscu
ssio
nof
the
lite
rary
isot
ope
data
Ear
lyM
ioce
ne(s
hort
-gra
sspr
airi
e)
Lat
eM
ioce
neR
etal
lack
(200
1)N
otsp
ecifi
edP
aleo
sols
.Gen
eral
izat
ion
byth
eau
thor
ofhi
sow
nin
vest
igat
ions
ondi
ffer
ent
cont
inen
tsSu
mm
ary
C3
C4
Con
tine
ntA
ppea
ranc
eE
xpan
sion
App
eara
nce
Exp
ansi
on
Eur
asia
Ear
lyM
ioce
neL
ate
Mio
cene
Mid
dle
Mio
cene
Lat
eM
ioce
neN
orth
Am
eric
aE
arly
Oli
goce
neL
ate
Oli
goce
neor
earl
yM
ioce
neE
arly
Mio
cene
Lat
eM
ioce
neS
outh
Am
eric
aL
ate
Oli
goce
neL
ate
Mio
cene
Lat
eO
ligo
cene
Lat
eM
ioce
neA
fric
aN
oda
taN
oda
taM
iddl
eM
ioce
neL
ate
Mio
cene
Aus
tral
iaM
iddl
eM
ioce
neL
ate
Mio
cene
Mid
dle
Mio
cene
Lat
eM
ioce
neT
heea
rlie
stes
tim
atio
nsfo
ral
lco
ntin
ents
Ear
lyO
ligo
cene
Lat
eO
ligo
cene
orea
rly
Mio
cene
Lat
eO
ligo
cene
Lat
eM
ioce
ne
202 S. N. Sheremet’ev and Y. V. Gamalei

Earth aridization, on the other hand. Herbs structural and functional changes weresuch that correspond to mesoxeromorphic and even xeromorphic plant organiza-tion: representatives of these ones had a low water evaporation rate (Fig. 11.1a),lower water content (Fig. 11.1b) of the leaves, denser mesophyll (Fig. 11.1c), andstronger water saturation deficit (Fig. 11.1d).
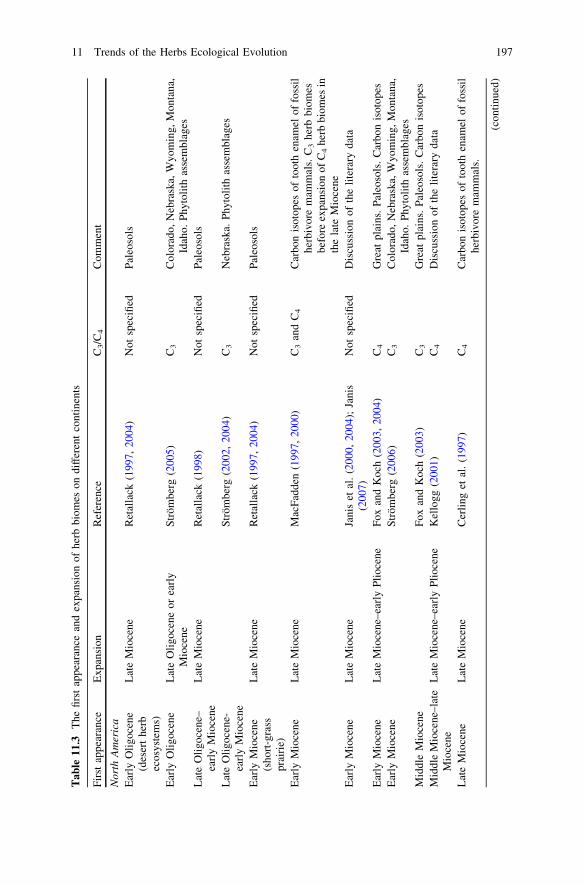
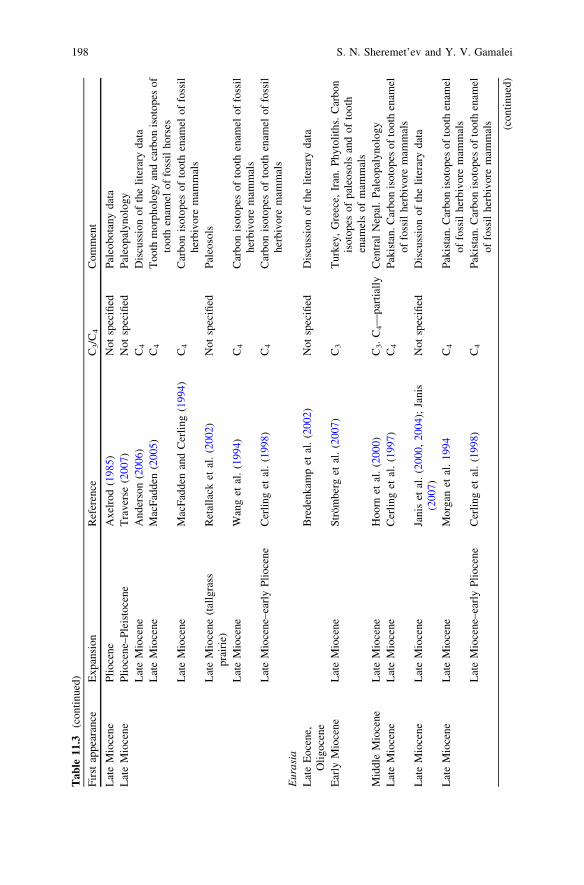
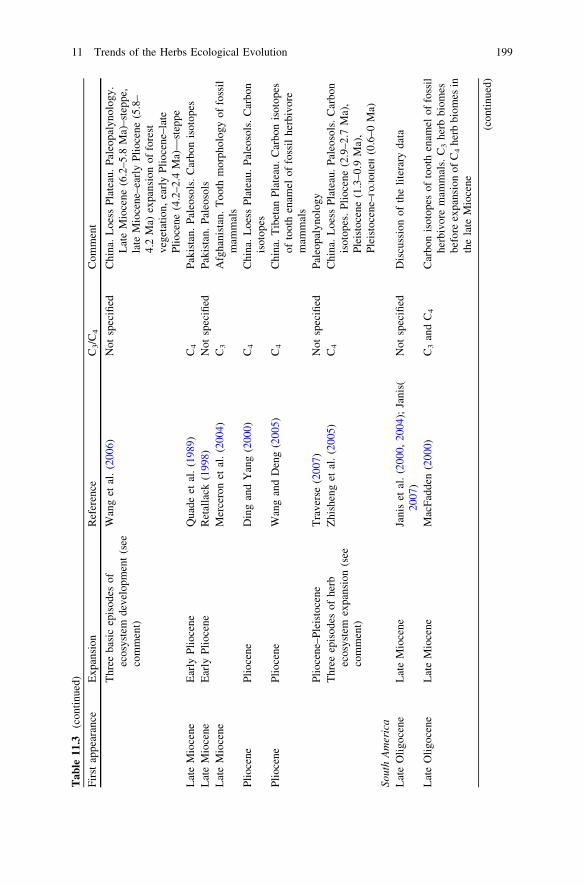
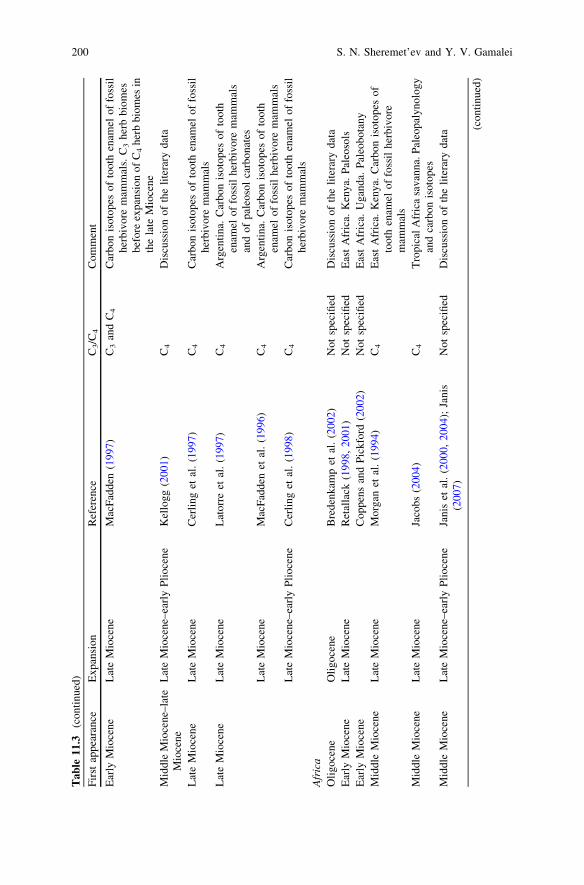
During Oligocene still there were no open grassland ecosystems similar modernones (see Table 11.3). In the early Oligocene of North America only firstappearance of desert herbs ecosystems (which landscape domination neverthelesswas not fixed) was pointed out (Retallack 1997, 2004; see also: Strömberg 2002,2004, 2005).
11.3.3 Miocene–Holocene
In the late Oligocene, climatic cooling was replaced by warming, but only for ashort time. In the early Miocene, the trend toward cooling became predominantagain (Zachos et al. 2001). It was interrupted by a mid-Miocene (17–14 Ma)climate optimum (see for example Shevenell et al. 2004), which became the lastone during the Cenozoic era. The structural and functional characteristics of plantsbegan changing toward mesomorphic organization again; however, this trendstopped soon. Another cooling, which began 14 Ma ago (Shevenell et al. 2004)(and continues until now), restored the structural and functional characteristicstypical of the Oligocene (Figs. 11.1 and 11.2).
Analyses have demonstrated that the time trends of the characters of plantleaves structure and water relations entirely corresponds to the simultaneousdirections of global climatic and hydrological changes (Fig. 11.2). Two obviousassumptions if the found parallels do not turn out artefacts are arising.
The first assumption that taxon (species or genus) once having appeared with acertain set of properties (or with certain norms of reactions and limits of a variationof structural and functional characters), keeps this set invariable throughout all itshistory. Considerable changes of the structural and functional organization can beconnected only with the origin of a new taxon. Perhaps, the invariance of themorphological organization of a taxon (it is base of morphological taxonomy) alsogo with an invariance of its structural and functional organization (Gamalei et al.2008). Otherwise, it would be impossible to explain why taxon-specific charactersso precisely reflect the specificity of the epoch when the given taxon was formed.The constant characters of each taxon in the varying environment explain thevariability of their geographic ranges, which reflects the pattern of subsequentdevelopment of the ecological niche where the taxon has been formed.
The second assumption that process of adaptogenesis is possible only in time oforigination and formation of a new taxon follows from the first assumption. Whenall characters have formed an equilibrium system, the taxon has been formed andadaptogenesis is completed. The further adaptive possibilities of it taxon (in timesof areal expansions or migrations) are limited by system developed in process of
11 Trends of the Herbs Ecological Evolution 203

adaptogenesis structural and functional properties. They are not related to addi-tional adaptive transformations (otherwise, a new taxon appears). The maximumpermissible quantitative deviations are those that do not disturb systemic rela-tionships between characters. There is a variety of the ways and mechanisms ofmigration. An intrazonal network is one of them.
11.4 Origin and Expansion of Grassland Biomes
There are definitions of biomes as large clusters of plant species that are charac-terized by the same life-form, whereas traditional definitions of biomes have alsoincluded either geographic or climatic descriptors (Woodward et al. 2004). In thefirst case, only two herbs biomes are discovering-savanna (mainly tropical her-baceous vegetation with woody canopy up to 30 %) and grassland (mainly tem-perate herbaceous vegetation with woody or/and shrubby canopy less than 10 %)(Woodward et al. 2004). On the other hand, in one of the traditional classificationthere are 14 biomes and 4 biomes of grassy vegetation between them are finding(Olson et al. 2001).
Tropical herbaceous biome is constituted by South American (llanos inVenezuela and Colombia, cerrado in Brazil), African savannas and Australiangrasslands (Allaby, 2006). Temperate grassland biome is constituted by Eurasiansteppes, North American prairies, South American pampas and Patagonian steppe,South African veld (Walter 1985).
By mode of carbon dioxide assimilation all herbs on two major groups aredivided: species with C3 and C4 photosynthesis.
Group of C3 herbs has been compelled to adapt for cold conditions. Thisadaptation were transferring an efflux of an assimilates into apoplast pathway andthan to guard a photosynthesis against suppression by cold because of effluxabsence into symplast pathway (Gamalei 2004; Gamalei et al. 2008). The coldcaused rigidity of actomyosin cytoskeleton, blocking development of plasmodes-mata and symplast. Barrier properties of a tonoplast have been as a result brokenand assimilates began to exude into apoplast. Gathering and return of assimilatesinto symplast—function of specialized «transfer cells». This function compensatesan absence of plasmodesmata and symplast pathway between mesophyll andphloem. Thus, perhaps in the late Eocene–early Oligocene was originated a groupof apoplast dicotyledonous (approximately 20,000 species from 21 families),which test sign-transfer cells (Gamalei 2004).
C4 herbs do not have transfer cells. They test sign-Kranz anatomy and plastiddimorphism. About 7,000 C4 species from 18 families are counted. This groupwas formed at the same time or a bit later (perhaps in the Oligocene, see Tippleand Pagani 2007), but in another climatic zone and in another composition oflimiting conditions of photosynthesis. In this composition an aridity, high tem-perature, and related salinization rather than cold dominates than xylem transportsuppresses and promotes to stomata closing. In the Cenozoic Era the CO2
204 S. N. Sheremet’ev and Y. V. Gamalei

concentration in the atmosphere is gradually decreasing (Berner 2006) than pho-tosynthesis sharply limits in such conditions. Dysfunction of stomata apparatus ofleaves still more strengthens carbon dioxide starvation. Mechanisms of CO2
concentration inside leaves became necessary. Kranz anatomy and cooperativephotosynthesis (inside 1 or 2 cells, see Voznesenskaya et al. 2006) have been as aresult generated. These characters for apoplastic halophytes and xerophytes arepeculiar. These plants reacted to the factor of CO2 concentration decrease becauseit was combined with not less stressful factors for photosynthesis such as waterdeficit and salinization. Closed stomata because of water deficiency—strong bar-rier to influx of carbon dioxide into the leaves (under whatever CO2 concentrationin the atmosphere).
Stomata apparatus—attribute of xylem transport, its functioning is subordinatedto it in a greater degree, than to photosynthesis. Stomata closing during the hotperiod of day is supervised by water relations and is inevitable reaction to envi-ronment water deficit. Therefore, the climate aridization which peaked in theOligocene and has led to occurrence of hot plains with a high aridity index couldbe the climatic factor that led to origination of C4 herbs. Sometimes C4 species canbe met in other conditions, but these are isolated instances which become at oncesubjects of special publications (see Gamalei et al. 1992). Distribution of C4
photosynthesis mainly in tropics (Ehleringer 2005)—argument that aridity factor ismore important for origination and formation of C4 syndrome. If the reason wasdecrease of carbon dioxide in atmosphere, C4 syndrome undoubtedly became thegeneral property of all land plants. Parallel and independent occurrence of C4
photosynthesis in several families (Kellogg 1998) does not leave doubt that C4
photosynthesis is one of the manifestations of arid adaptogenesis.Thus, approximately at the same time on last big wave of taxonogenesis
two phylogenetic branches of herbs are originated, one (C3) as the answer to aclimate cooling, another (C4) as the answer to climate aridization. Cold openplains (meadows, steppes, tundra, high mountains) occupy C3 species, hot and dryhabitats (savannas, saline soils, deserts) inhabit C4 species.
There are assumptions that origination of C4 photosynthesis occurred in theOligocene (25–33 million years ago) (Kellogg 1998; Sage 2003, 2005, and others),and its wide expansion took place in a late Miocene, 5–7 million years ago (seeTable 11.3). C4 herbs, especially grasses, extend within tropics in grasslandbiomes of Northern and Southern Americas, Africa, Australia (Kovalev 2000;Kellogg 2001). These are plants with pronounced adaptive possibilities and highrates of photosynthesis and growth (Gamalei et al. 1992). In the modern worldwhere C4 taxa are presented less than 2 % of higher plant species, their contri-bution to global primary production constitutes about 25 % (Still et al. 2003;Ehleringer 2005). Approximately, half of 10,000 species of grasses and sedgeshave C4 photosynthesis, whereas among about 2,000 dicotyledonous species thisway of a metabolism is fixed (Ehleringer et al. 2002).
Occurrence of temperate herbs biomes with C3 photosynthesis took place in themiddle or late Eocene or early Oligocene (Zherikhin 1994; Retallack 1997, 2001;Jacobs et al. 1999; Bredenkamp et al. 2002) (Table 11.3). Wide expansion of these
11 Trends of the Herbs Ecological Evolution 205

biomes began in the early Miocene (Retallack 1997, 2001, 2004; Jacobs et al.1999; Willis and McElwain 2002) and was combined with intensive radiation ofgrazing mammalian herbivores (Zherikhin 1994; Cerling et al. 1998; Janis et al.2000, 2004; Kemp 2005; MacFadden 2005; Janis 2007) (Table 11.3).
Tropical and paratropical megathermal rainforests that covered more than 50 %of continents in the late Paleocene–early Eocene (Morley 2007), in the modernworld almost completely are replaced by herbaceous vegetation (Ehleringer 2005).Replacement of forests by herbaceous vegetation was probably not unidirectional,but, as shown in a number of works, proceeded by pulsations (Zhisheng et al.2005; Wang et al. 2006). Such replacement could result from increasing of cli-matic zonality and global cooling (Willis and McElwain 2002), destruction offorests by fires (Bond et al. 2005), changes of carbon dioxide concentration inatmosphere (Ehleringer et al. 2002). This tendency becomes stronger as conse-quence of anthropogenic influence.
11.5 Conclusion
Results of the analytical research have shown that dynamics of leaves structure andwater relations of plants corresponds to conceptions about global climate changesand a planet hydrology.
Taxonomical diversity of herbs and herbaceous biomes is the function ofpaleoclimate variability and plant adaptogenesis to it. Two global trends of eco-logical evolution contrast differing by the composition of herbaceous adaptivetypes is the next: (a) the line of herbs of chilling plains with domination the groupof plant species with C3 apoplastic syndrome formed under cold climate influence,and (b) the line of herbs of hot plains with domination of plant species with C4
apoplastic syndrome.Both trends include the monocots and dicots, and both are the results of climate
changes in Cenozoic. C3 herbs of chilling plains and the steppe and meadowphytocoenosis formed by them arise as the answer to temperature decrease in greatareas of high latitudes. The apoplastic syndrome (transfer from symplastic trans-port of assimilates supressed by cold to their apoplastic transport) is the diagnostictest for this group of herbs. C4 herbs of hot plains and the savanna, desert andsolontchak plant vegetation are the adaptive answer to aridization of low latitudeareas. C4 syndrome (compensation of stomata closure by the mechanism of CO2
concentration in the leaf tissues) is a special sign of this group of herbs.Formation of these adaptive types and expansion of the biomes formed by them
occurred almost simultaneously. Both groups derived from primitive apoplasticherbs. Both groups have appeared in the Oligocene which is characterized byclimate cooling and aridization, as a result they are separated territorially. The firstgroup acclimatized to conditions of cold open territories and obtained adaptivefeatures corresponding to the cold environment (transfer cells that compensateabsence of plasmodesmata). The second group adapted for the arid and salted open
206 S. N. Sheremet’ev and Y. V. Gamalei

spaces with excess of heat and water deficit, that compel stomata be closed in theafternoon and accordingly to get special mechanisms of supplying carbon dioxide(carbon dioxide concentrating—C4 syndrome, or its time deposition—CAM).
Problem of the first—how to provide efflux of an assimilates in cold temperatureconditions at which symplastic transport is impossible. The problem of the second—how to provide photosynthesis by CO2 in hot conditions when stomata is closed andwhen xylem transport is suppressed. For overcoming both problems adaptivemechanisms were discovered but in different groups of taxa. Physiologically, thesemechanisms are incompatible, alternative; and taxonomically these groups of theherbs which have acclimatized to conditions of cold and hot plains are separated.
Both types of herbs biomes began to replace the forest biomes whose areal inthe late Paleogene–Neogene began to reduce both in low and in high latitudes.This tendency, probably, proceeds in parallel with the climatic tendency ofaridization and cooling of continents.
Constancy of taxa characters in combination with inconstancy of climate—thereason of areal mobility. Territorial niche drifts which are connected with globaland local variability of a climate explain migration of taxa after completion offormation period. There is a variety of the ways and mechanisms of migration. Anintrazonal network is one of them.
Acknowledgments Funding for this work was provided by a grant from the Russian Foundationfor Basic Research (10-04-01165-a). We would like to thank Pierre Pontarotti and Marie-HélèneRome for the invitation to contribute to the 15th evolutionary biology meeting at Marseille wherethis work was presented.
References
Akhmetiev MA (2004) Globe climate in Palaeocene and Eocene according to data ofpaleobotany. In: Semikhatov MA, Chumakov NM (eds) Climate in the epochs of majorbiosphere transformations. Nauka, Moscow, pp 10–43 (In Russian)
Allaby M (2006) Biomes of the world: grasslands. Chelsea House, New YorkAnderson RC (2006) Evolution and origin of the Central Grassland of North America: climate,
fire, and mammalian grazers. J Torrey Bot Soc 133(4):626–647Axelrod DI (1985) Rise of the grassland biome, Central North America. Bot Rev 51(2):163–201Beerling DJ, Royer DL (2002) Fossil plants as indicators of the phanerozoic global carbon cycle.
Annu Rev Earth Planet Sci 30:527–556Beerling DJ, Woodward FI (2001) Vegetation and the terrestrial carbon cycle: modelling the first
400 million years. Cambridge Univ Press, CambridgeBenton MJ (1993) The fossil record 2. Chapman & Hall, LondonBerner RA, Kothavala Z (2001) GEOCARB III: a revised model of atmospheric CO2 over
Phanerozoic time. Amer J Sci 301(2):182–204Berner RA (2006) GEOCARBSULF: a combined model for Phanerozoic atmospheric O2 and
CO2. Geoch et Cosmoch Acta 70(23):5653–5666Bobe R (2006) The evolution of arid ecosystems in eastern Africa. J Arid Environ 66(3):564–584Bobe R, Behrensmeyer AK (2004) The expansion of grassland ecosystems in Africa in relation to
mammalian evolution and the origin of the genus Homo. Palaeogeogr PalaeoclimatolPalaeoecol 207(3–4):399–420
11 Trends of the Herbs Ecological Evolution 207

Bond WJ, Woodward FI, Midgley GF (2005) The global distribution of ecosystems in a worldwithout fire. New Phytol 165(2):525–538
Bredenkamp GJ, Spada F, Kazmierczak E (2002) On the origin of northern and southernhemisphere grasslands. Plant Ecol 163(2):209–229
Briggs DEG, Crowther PR (eds) (1997) Palaeobiology: a synthesis. Blackwell Science Ltd,Oxford
Cerling TE, Ehleringer JR, Harris JM (1998) Carbon dioxide starvation, the development of C4
ecosystems, and mammalian evolution. Phil Trans R Soc Lond B Biol Sci 353(1365):159–171Cerling TE, Harris JM, Leakey MG (2005) Environmentally driven dietary adaptations in African
mammals. In: Ehleringer JR, Cerling TE, Dearing MD (eds) A history of atmospheric CO2
and its effects on plants, animals, and ecosystems. Ecological studies 177, Springer,New York, 258–272
Cerling TE, Harris JM, MacFadden BJ, Leakey MG, Quade J, Eisenmann V, Ehleringer JR(1997) Global vegetation change through the Miocene/Pliocene boundary. Nature 389(6647):153–158
Cerling TE, Wang Y, Quade J (1993) Expansion of C4 ecosystems as an indicator of globalecological change in the late Miocene. Nature 361(6410):344–345
Chumakov NM (1993) Problems of paleoclimate in investigations of biosphere evolution. In:Rozanov AYu (ed) Problems of biosphere evolution before anthropogenic, Nauka, Moscow,pp 106–122 (In Russian)
Chumakov NM (1997) Warm biosphere. Nature 5:66–78 (In Russian)Chumakov NM (2004a) The general review of late Mesozoic climate and events. In: Semikhatov
MA, Chumakov NM (eds) Climate in the epochs of major biosphere transformations. Nauka,Moscow, pp 44–51 (In Russian)
Chumakov NM (2004b) Climate zonality and climate of the Cretaceous. In: Semikhatov MA,Chumakov NM (eds) Climate in the epochs of major biosphere transformations. Nauka,Moscow, pp 105–123 (In Russian)
Coppens Y, Pickford M (2002) Early Miocene grassland ecosystem at Bukwa, Mount Elgon.Uganda Comptes Rendus Palevol 1(4):213–219
Culver SJ, Rawson PF (eds) (2000) Biotic response to global change: the last 145 million years.Cambridge Univ Press, Cambridge
DeConto RM, Pollard D (2003a) Rapid Cenozoic glaciation of Antarctica induced by decliningatmospheric CO2. Nature 421(6920):245–249
DeConto RM, Pollard D (2003b) A coupled climate-ice sheet modeling approach to the earlyCenozoic history of the Antarctic ice sheet. Palaeogeogr Palaeoclimatol Palaeoecol 198(1–2):39–52
Ding ZL, Yang SL (2000) C3/C4 vegetation evolution over the last 7.0 Myr in the Chinese LoessPlateau: evidence from pedogenic carbonate d13C. Palaeogeogr Palaeoclimatol Palaeoecol160(3):291–299
Dugas DP, Retallack GJ (1993) Middle Miocene fossil grasses from Fort Ternan. Kenya J Paleont67(1):113–128
Ehleringer JR (2005) The influence of atmospheric CO2, temperature, and water on theabundance of C3/C4 taxa. In: Ehleringer JR, Cerling TE, Dearing MD (eds) A history ofatmospheric CO2 and its effects on plants, animals, and ecosystems. Ecological studies 177,Springer, New York, pp 214–231
Ehleringer JR, Cerling TE, Dearing MD (2002) Atmospheric CO2 as a global change driverinfluencing plant-animal interactions. Integr Comp Biol 42(3):424–430
Ehleringer JR, Cerling TE, Helliker BR (1997) C4 photosynthesis, atmospheric CO2, and climate.Oecologia 112(3):285–299
Eldrett JS, Harding IC, Wilson PA, Butler E, Roberts AP (2007) Continental ice in Greenlandduring the Eocene and Oligocene. Nature 446(7132):176–179
Fox DL, Koch PL (2003) Tertiary history of C4 biomass in the great plains. USA Geology31(9):809–812
208 S. N. Sheremet’ev and Y. V. Gamalei

Fox DL, Koch PL (2004) Carbon and oxygen isotopic variability in Neogene paleosol carbonates:constraints on the evolution of the C4-grasslands of the great plains, USA. PalaeogeogrPalaeoclimatol Palaeoecol 207(3–4):305–329
Frakes LA, Francis JE, Syktus JI (2005) Climate modes of the Phanerozoic: The history of theEarth’s climate over the past 600 million years. Cambridge Univ Press, New York
Gamalei YuV (1988) Structure of plants of Trans-Altai Gobi. In: Gamalei YuV et al (eds) Desertsof Trans-Altai Gobi. Nauka, Leningrad, pp 44–107 (In Russian)
Gamalei YuV (2000) Structural-functional variety of species-a basis of a variety of floras andvegetation types. In: Yurtsev BA (ed) Comparative floristics on a boundary of III millennium.Komarov Botanical Institute, St Petersburg, pp 350–374 (In Russian)
Gamalei YuV (2004) Transport system of vascular plants. St. Petersburg Univ. Press, St.Petersburg, p 422 (In Russian)
Gamalei YuV, Glagoleva TA, Kolchevsky KG, Chulanovskaya MV (1992) Ecology andevolution of types of C4 syndrome in connection with phylogeny of families Chenopodiaceaeand Poaceae. Bot J 77(2):1–12 (In Russian)
Gamalei YuV, Pakhomova MV, Sheremet’ev SN (2008) Dicotyledonous of Cretaceous,Paleogene, and Neogene. Adaptogenesis of the terminal phloem. J Gen Biol 69(3):220–237(In Russian)
Gibbs MT, Bluth GJS, Fawcett PJ, Kump LP (1999) Global chemical erosion over the last250 My: variations due to changes in paleogeography, paleoclimate, and paleogeology. AmerJ Sci 299(7–9):611–651
Hansen KW, Wallmann K (2003) Cretaceous and Cenozoic evolution of seawater composition,atmospheric O2 and CO2: a model perspective. Amer J Sci 303(2):94–148
Hoorn C, Ohja T, Quade J (2000) Palynological evidence for vegetation development andclimatic change in the Sub-Himalayan zone (Neogene, Central Nepal). PalaeogeogrPalaeoclimatol Palaeoecol 163(3–4):133–161
Jacobs BF (2004) Palaeobotanical studies from tropical Africa: relevance to the evolution offorest, woodland and savannah biomes. Phil Trans R Soc Lond B 359(1450):1573–1583
Jacobs BF, Kingston JD, Jacobs LL (1999) The origin of grass dominated ecosystems. Ann MoBot Gard 86(2):590–643
Jahren AH (2007) The Arctic forest of the middle Eocene. Annu Rev Earth Planet Sci 35:509–540
Janis CM, Damuth J, Theodor JM (2000) Miocene ungulates and terrestrial primary productivity:Where have all the browsers gone? PNAS 97(14):7899–7904
Janis CM, Damuth J, Theodor JM (2004) The species richness of Miocene browsers, andimplications for habitat type and primary productivity in the North American Grasslandbiome. Palaeogeogr Palaeoclimatol Palaeoecol 207(3–4):371–398
Janis CM (2007) An evolutionary history of browsing and grazing ungulates. In: Gordon IJ, PrinsHHT (eds) The Ecology of browsing and grazing. Ecological studies 195, Springer, Berlin,p 21–45
Jones RN (1999) The biogeography of the grasses and lowland grasslands of South-EasternAustralia. In: Jones RN (ed) The great plains crash: proceedings of a conference on victorianlowland grasslands and grassy woodlands. Adv Nat Conserv 2:11–18
Keeley JE, Rundel PW (2005) Fire and the Miocene expansion of C4 grasslands. Ecol Lett8(7):683–690
Kellogg E (1998) Phylogenetic aspects of the evolution of C4 photosynthesis. In: Sage RF,Monson RK (eds) C4 plant biology. Academic Press, San Diego, pp 411–444
Kellogg EA (2001) Evolutionary history of the grasses. Plant Physiol 125(3):1198–1205Kemp TS (2005) The origin and evolution of mammals. Oxford Univ Press, OxfordKennett JP (1977) Cenozoic evolution of Antarctic glaciation, the circum-Antarctic Ocean, and
their impact on global paleoceanography. J Geophys Research 82(C27):3843–3860Kidder DL, Gierlowski-Kordesch EH (2005) Impact of grassland radiation on the nonmarine
silica cycle and Miocene diatomite. Palaios 20(2):198–206
11 Trends of the Herbs Ecological Evolution 209

Koch PL (1998) Isotopic reconstruction of past continental environments. Annu Rev Earth PlanetSci 26:573–613
Kovalev OV (2000) Evolution of C4 syndrome of the angiosperm’s photosynthesis. Bot J85(11):7–20 (In Russian)
Latorre C, Quade J, McIntosh WC (1997) The expansion of C4 grasses and global change in thelate Miocene: stable isotope evidence from the Americas. Earth Planet Sci Lett 146(1–2):83–96
Lear CH, Elderfield H, Wilson PA (2000) Cenozoic deep-sea temperatures and global icevolumes from Mg/Ca in benthic foraminiferal calcite. Science 287(5451):269–272
MacFadden BJ (1997) Origin and evolution of the grazing guild in new world terrestrialmammals. Trends Ecol Evol 12(5):182–187
MacFadden BJ (2000) Cenozoic mammalian herbivores from the Americas: reconstructingancient diets and terrestrial communities. Annu Rev Ecol Syst 31:33–59
MacFadden BJ (2005) Terrestrial mammalian herbivore response to declining levels ofatmospheric CO2 during the cenozoic: evidence from North American fossil horses (familyEquidae). In: Ehleringer JR, Cerling TE, Dearing MD (eds) A history of atmospheric CO2 andits effects on plants, animals, and ecosystems. Ecological studies 177, Springer, New York,pp 273–292
MacFadden BJ, Cerling TE (1994) Fossil horses, carbon isotopes and global change. Trends EcolEvol 9(12):481–486
MacFadden BJ, Cerling TE, Prado J (1996) Cenozoic terrestrial ecosystem evolution inArgentina: evidence from carbon isotopes of fossil mammal teeth. Palaios 11(4):319–327
Martínez-Millán M (2010) Fossil record and age of the Asteridae. Bot Rev 76(1):83–135Merceron G, Blondel C, Brunet M et al (2004) The late Miocene paleoenvironment of
Afghanistan as inferred from dental microwear in artiodactyls. Palaeogeogr PalaeoclimatolPalaeoecol 207(1–2):143–163
Middleton N, Thomas D (1997) World atlas of desertification, 2nd edn. Arnold, London 182 pMiller KG, Kominz MA, Browning JV et al (2005) The Phanerozoic record of global sea-level
change. Science 310(5752):1293–1298Moran K, Backman J, Brinkhuis H et al (2006) The Cenozoic palaeoenvironment of the Arctic
Ocean. Nature 441(7093):601–605Morgan ME, Kingston JD, Marino BD (1994) Carbon isotopic evidence for the emergence of C4
plants in the Neogene from Pakistan and Kenya. Nature 367(6459):162–165Morley RJ (2007) Cretaceous and Tertiary climate change and the past distribution of
megathermal rainforests. In: Bush MB, Flenley JR (eds) Tropical rainforest responses toclimatic change. Springer, Berlin, pp 1–31
Mosbrugger V, Utescher T, Dilcher DL (2005) Cenozoic continental climatic evolution ofCentral Europe. PNAS 102(42):14964–14969
Muller J (1981) Fossil pollen record of extant angiosperms. Bot Rev 47(1):1–142Nikolaev SD, Oskina NS, Blyum NS, Bubenshchikova NV (1998) Neogene–Quaternary
variations of the ‘Pole–Equator’ temperature gradient of the surface oceanic waters in theNorth Atlantic and North Pacific. Glob Planet Change 18(3–4):85–111
Ogg JG, Ogg G, Gradstein FM (2008) The concise geologic time scale. Cambridge Univ Press,New York
Olson DM, Dinerstein E, Wikramanayake ED et al (2001) Terrestrial ecoregions of the world: anew map of life on earth. Bioscience 51(11):933–938
Pagani M, Zachos JC, Freeman KH, Tipple B, Bohaty S (2005) Marked decline in atmosphericcarbon dioxide concentrations during the Paleogene. Science 309(5734):600–603
Pearson PN, van Dongen BE, Nicholas CJ et al (2007) Stable warm tropical climate through theEocene epoch. Geology 35(3):211–214
Pollard D, DeConto RM (2003) Antarctic ice and sediment flux in the Oligocene simulated by aclimate-ice sheet-sediment model. Palaeogeogr Palaeoclimatol Palaeoecol 198(1–2):53–67
Pollard D, DeConto RM (2005) Hysteresis in Cenozoic Antarctic ice-sheet variations. GlobPlanet Change 45(1–3):9–21
210 S. N. Sheremet’ev and Y. V. Gamalei

Quade J, Cerling TE, Bowman JR (1989) Development of Asian monsoon revealed by markedecological shift during the latest Miocene in northern Pakistan. Nature 342(6246):163–166
Retallack GJ (1992) Middle Miocene fossil plants from Fort Ternan (Kenya) and evolution ofAfrican grasslands. Paleobiology 18(4):383–400
Retallack GJ (1997) Neogene expansion of the North American prairie. Palaios 12(4):380–390Retallack GJ (1998) Grassland ecosystems as a biological force in dusty dry regions. Busacca AJ
(ed) Dust aerosols. Loess soils and global change (Conference proceedings, Seattle),Washington State University, College of Agriculture and Home Economics, Pullman,Washington, pp 171–174
Retallack GJ (2001) Cenozoic expansion of grasslands and climatic cooling. J Geology109(4):407–426
Retallack GJ (2004) Late Oligocene bunch grassland and early Miocene sod grassland paleosolsfrom central Oregon, USA. Palaeogeogr Palaeoclimatol Palaeoecol 207(3–4):203–237
Retallack GJ, Dugas DP, Bestland EA (1990) Fossil soils and grasses of a middle Miocene EastAfrican grassland. Science 247(4948):1325–1328
Retallack GJ, Tanaka S, Tate T (2002) Late Miocene advent of tall grassland paleosols in Oregon.Palaeogeogr Palaeoclimatol Palaeoecol 183(3–4):329–354
Royer DL (2006) CO2-forced climate thresholds during the Phanerozoic. Geochim CosmochimActa 70(23):5665–5675
Sage RF (2003) The evolution of C4 photosynthesis. New Phytol 161(2):341–370Sage RF (2005) Atmospheric CO2, environmental stress, and the evolution of C4 photosynthesis.
In: Ehleringer JR, Cerling TE, Dearing MD (eds) A history of atmospheric CO2 and its effectson plants, animals, and ecosystems. Ecological studies 177, Springer, New York, pp 185–213
Scotese CR (2003) PALEOMAP Project. (http://www.scotese.com)Ségalen L, Renard M, Lee-Thorp JA et al (2006) Neogene climate change and emergence of C4
grasses in the Namib, southwestern Africa, as reflected in ratite 13C and 18O. Earth Planet SciLett 244(3–4):725–734
Semikhatov MA, Chumakov NM (eds) (2004) Climate in the epoches of major biospherictransformations (transactions of the Geological Institute of the Russian Academy of Sciences,issue 550). Nauka, Moscow, p 299 (in Russian)
Sheremet’ev SN (2005) Herbs on the soil moisture gradient (water relations and the structural-functional organization). KMK, Moscow, p 271 (In Russian)
Shevenell AE, Kennett JP, Lea DW (2004) Middle Miocene Southern Ocean cooling andAntarctic cryosphere expansion. Science 305(5691):1766–1770
Shields LM (1950) Leaf xeromorphy as related to physiological and structural influences. BotRev 16(8):399–447
Still CJ, Berry JA, Collatz GJ, DeFries RS (2003) Global distribution of C3 and C4 vegetation:carbon cycle implications. Glob Biogeochem Cycles 17(1):6.1–6.14
Strömberg CAE (2002) The origin and spread of grass-dominated ecosystems in the late tertiaryof North America: preliminary results concerning the evolution of hypsodonty. PalaeogeogrPalaeoclimatol Palaeoecol 177(1–2):59–75
Strömberg CAE (2004) Using phytolith assemblages to reconstruct the origin and spread of grass-dominated habitats in the great plains of North America during the late Eocene to earlyMiocene. Palaeogeogr Palaeoclimatol Palaeoecol 207(3–4):239–275
Strömberg CAE (2005) Decoupled taxonomic radiation and ecological expansion of open-habitatgrasses in the Cenozoic of North America. PNAS 102(34):11980–11984
Strömberg CAE (2006) Evolution of hypsodonty in equids: testing a hypothesis of adaptation.Paleobiology 32(2):236–258
Strömberg CAE, Werdelin L, Friis EM, Saraç G (2007) The spread of grass-dominated habitats inTurkey and surrounding areas during the Cenozoic: Phytolith evidence. PalaeogeogrPalaeoclimatol Palaeoecol 250(1–4):18–49
Tajika E (1999) Carbon cycle and climate change during the Cretaceous inferred from abiogeochemical carbon cycle model. Island Arc 8(2):293–303
11 Trends of the Herbs Ecological Evolution 211

Tipple BJ, Pagani M (2007) The early origins of terrestrial C4 photosynthesis. Annu Rev EarthPlanet Sci 35:435–461
Traverse A (2007) Paleopalynology. Springer, Dordrecht 813 pTripati A, Backman J, Elderfield H, Ferretti P (2005) Eocene bipolar glaciation associated with
global carbon cycle changes. Nature 436(7049):341–346Vasilevskaya VK (1979) Development of ecological anatomy in the USSR. Bot J 64(5):654–664
(In Russian)Veizer J, Ala D, Azmy K et al (1999) 87Sr/86Sr, d18O and d13C evolution of phanerozoic
seawater. Chem Geol 161(1–3):59–88 (web update 2004). (http://www.science.uottawa.ca/-geology/isotope_data)
Voznesenskaya EV, Franceschi VR, Chuong SDX, Edwards GE (2006) Functional character-ization of phosphoenolpyruvate carboxykinase-type C4 leaf anatomy: immuno-, cytochemicaland ultrastructural analyses. Ann Bot 98(1):77–91
Wallmann K (2004) Impact of atmospheric CO2 and galactic cosmic radiation on Phanerozoicclimate change and the marine d18O record. Geochem Geophys Geosyst 5(6):1–29
Walter H (1985) Vegetation of the earth and ecological systems of the geo-biosphere (3rdedition)., 3rd edn. Springer-Verlag, New York pp 318
Wang L, Lü HY, Wu NQ et al (2006) Palynological evidence for late Miocene–Pliocenevegetation evolution recorded in the red clay sequence of the central Chinese Loess Plateauand implication for palaeoenvironmental change. Palaeogeogr Palaeoclimatol Palaeoecol241(1):118–128
Wang Y, Cerling TE, MacFadden BJ (1994) Fossil horses and carbon isotopes: new evidence forCenozoic dietary, habitat, and ecosystem changes in North America. Palaeogeogr Palaeo-climatol Palaeoecol 107(3–4):269–279
Wang Y, Deng T (2005) A 25 m.y. isotopic record of paleodiet and environmental change fromfossil mammals and paleosols from the NE margin of the Tibetan Plateau. Earth Planet SciLett 236(1–2):322–338
Willis KJ, McElwain JC (2002) The evolution of plants. Oxford Univ Press, Oxford 378 pWoodward FI, Lomas MR, Kelly CK (2004) Global climate and the distribution of plant biomes.
Phil Trans R Soc Lond B 359(1450):1465–1476Zachos J, Pagani M, Sloan L, Thomas E, Billups K (2001) Trends, rhythms, and aberrations in
global climate 65 ma to present. Science 292(5517):686–693Zharkov MA, Murdmaa IO, Filatova NI (2004) Paleogeographical reorganizations and
sedimentation of the cretaceous period. In: Semikhatov MA, Chumakov NM (eds) Climatein the epochs of major biosphere transformations. Nauka, Moscow, pp 52–87 (In Russian)
Zherikhin VV (1994) Genesis of herbs biomes. In: Rozanov AYu, Semikhatov MA (eds)Ecosystems reorganizations and biosphere evolution, issue 1, Nedra, Moscow, pp 132–137(In Russian)
Zhisheng A, Yongsong H, Welguo L et al (2005) Multiple expansions of C4 plant biomass in EastAsia since 7 Ma coupled with strengthened monsoon circulation. Geology 33(9):705–708
212 S. N. Sheremet’ev and Y. V. Gamalei





























