Embed Size (px)
Citation preview

WOLIATA SODO UNIVERSITY
SCHOOL OF VETERINARY MEDICINE
TRYPANOCIDAL DRUG AND DEVELOPMENT OF DRUG RESISTANCE IN
AFRICAN ANIMAL TRYPANOSOMOSIS
A paper submitted for the course Senior Seminar on Animal Health (EMPH 504)
BY: Elias Kalel
Advisor: Dr.Asefa Asmare (PhD)
April, 2015
Woliata Sodo, Ethiopia

i
TABLE OFCONTENTS PAGE
LIST OF TABLES III
LIST OF FIGURES IV
LIST OF ABBREVATIONS V
ACKNOWLEDGEMENT VI
SUMMARY VII
1. INTRODUCTION 1
2. AFRICAN TRYPANOSOMES 3
2.1. Morphology and characterization ............................................................................ 3
2.2. Life cycle ..................................................................................................................... 4
2.3. The vector of African trypanosomosis ..................................................................... 5
2.4. Host ............................................................................................................................. 6
2.5. Risk factors of trypanosomosis ................................................................................. 7
2.5.1. Animal factor 7
2.5.2. Environmental factor 8
2.6. Trypanosomosis control methods ............................................................................. 8
2.6.1. Sequential aerosol technique 8
2.6.2. Stationary attractive devices (traps and targets) 9
2.6.3. Live bait technique 9
2.6.4. Sterile insect technique 10
3. TRYPANOCIDAL DRUGS 11
3.1.Prophylactic treatments ........................................................................................... 11
3.2.Curative treatments .................................................................................................. 12
3.3.Mechanism of action of current used drugs ........................................................... 12
3.3.1. Diminazene aceturate (Berenil®) 12
3.3.2. Isometamidium and homidium 12
3.3.3.Suramin (Polysulphonated naphtyl urea) 13
3.3.4. Melarsomine hydrochloride (Cymelarsan®
) 13
4. TRYPANOCIDAL DRUG RESISTANCE 14

ii
4.1. Definition .................................................................................................................. 14
4.2. Causes of trypanocidal drug resistance ................................................................. 14
4.3. Types of trypanocidal drug resistance ................................................................... 15
4.4. Cross and multiple drug resistance ........................................................................ 16
4.5. Mechanism of drug resistance ................................................................................ 17
4.6. Current situation of resistance ............................................................................... 18
4.7. Impact of drug resistance ........................................................................................ 19
4.7.1. Factor influencing development of resistance to Trypanocidal drugs 19
4.8. Detection of drug resistance .................................................................................... 20
4.8.1.Field methods 20
4.8.2. Drug sensitivity studies in experimental animals 21
4.8.3. In vitro assays 23
4.8.4. Xenodiagnosis 23
4.8.5. Serological techniques 24
4.8.6. Molecular techniques 24
4.9. Measures to combat drug resistance in the field................................................... 25
4.9.1. Use of the correct dose 25
4.9.2. Changes of drugs 26
4.9.3. Sanative treatment 26
4.9.4. High dose and repeat treatment regimen 26
4.9.5. Use of combined drugs 27
4.9.6. Beware of fake drugs 27
4.10. Quality assurance of trypanocidal drugs ............................................................. 27
5. FUTURE PROSPECT IN TREATMENT FOR TRYPANOSOMOSIS 28
6. CONCLUSSION AND RECOMMENDATIONS 29
7. REFERENCES 30

iii
LIST OF TABLES
Page
Table 1: Drug resistant trypanosomes in African countries ------------------------------------- 15
Table 2: Cross-resistance between trypanocidal drugs ------------------------------------------ 17

iv
LIST OF FIGURES
Page
Figure 1. Schematic representation of the digenetic life cycle of Trypanosoma brucei in the
mammalian host and in the tsetse fly vector --------------------------------------------- 5
Figure 2. The structures of the four most commonly used drug in the chemotherapy and
chemoprophylaxis of animal trypanosomosis Africa ---------------------------------- 11
Figure 3. African countries with reported resistance to trypanocidal drugs ---------------- 15
Figure 4. Some factors influencing the development of resistance trypanocidal drugs ------20

v
LIST OF ABBREVATIONS
AT African Trypanosomosis
DIM Diminazene aceturate
GPI Glycosyl-Phosphatidyl-Inositol
HAT Human African Trypanosomosis
Ig M Immunoglobulin M
ILRAD International Laboratory for Research on Animal Diseases
ISMM Isometamidium Chloride
Square kilometres
PCR Polymerase Chain Reaction
RFLP Restriction Fragment Length Polymorphism
USD US Dollars
VSG Variant Surface Glycoprotein
SSA Sub-Saharan Africa
HOM Homidium Salts
BCT Phase Contrast Buffy Coat' Technique
i.m Intra-muscular
PCV Packed cell volume
ED Effective dose
DIIT Drug incubation infectivity test
DIGIT Drug incubation Glossina Infectivity Test
bp Base pairs
SMT Standard Mouse Test

vi
ACKNOWLEDGEMENT
I would like to express my heartfelt thanks to Dr. Asefa Asmare for his valuable advice, over
all guidance and unreserved effort he made to supply with essential materials as well as to
correct this paper. In fact, this is a good opportunity to express my warm appreciation for his
exemplary professional quality.
Furthermore, I sincerely address my appreciation to Dr, Eyob Eshetu, Dr Taju Nugusse and
My father Kalel Mohammed, My mother Ahado Badhaso, My uncle Addus Mohammed and
My Brother Mohammed Xaha for their direct and indirect contribution in preparation of this
seminar paper.

vii
SUMMARY
The general features of trypanosomosis, drugs for the treatment and drug resistance in African
trypanosomoses are briefly reviewed in this paper and measures to combat drug resistance
especially at field level are also suggested. Trypanosomosis is the most serious animal health
problem in Sub-Saharan Africa and prevents the keeping of animals over 10 millions square
kilometers of potentially productive land making 50 to 70 million cattle at risk. To prevent
and treat trypanosomosis trypanocidal drugs belonging to different chemical families are used
quite intensively in veterinary medicine. Prophylactic and curative treatment of animal
trypanosomosis is carried out using three main drugs known as homidium salts, diminazene
aciturate and isometamidium chloride. About thirty five million doses of trypanocidal drugs
are used annually in the treatment of animal trypanosomosis in Africa. Most of these drugs are
very old and have been utilized for a long period of time. Hence, treatment of trypanosomosis
is complicated by development of drug resistance. Trypanocidal drug resistance has been
widely distributed in Africa and was reported in 17 countries. Like bacterial resistances to
antibiotics, resistance to trypanocides is associated with genetic modification that transforms a
susceptible population of trypanosome into resistant ones. The role of pressure selection on
the development of drug resistant trypanosomes cannot also be undermined. Uncontrolled
infections of drug-resistant trypanosomes will have a severe impact on both survival and
productivity of animals. It is very unlikely that new trypanocidal drugs will be released into
the market in the near future because these diseases are found in marginalized communities of
the developing countries. Therefore, it is essential to maintain the efficacy of the currently
available drugs through proper utilization of trypanocidal drugs such as use of correct dose,
change of drugs, sanative treatment, high dose, use of combined drugs and quality assurance
of trypanocidal drugs.

1
1. INTRODUCTION
African animal trypanosomosis (nagana) is a group of diseases of ruminants, camels, equines,
swine and carnivores caused by different trypanosome species. The major pathogenic species in
African cattle are T. congolense, T. vivax, and, to a lesser extent, T. brucei (Taylor and Authie,
2004). Although hosts acquire infection principally via the bite of infected tsetse flies, other
haematophagous insects like Tabanids and Stomoxys species also transmit trypanosomosis
mechanically (Eisler et al., 2004).
Trypanosomosis in humans, also known as sleeping sickness, is caused by two sub-species of
Trypanosoma brucei:T. b. gambiense, which is the agent of a late on set, chronic form that is
endemic in Western and Central Africa, and T. b. rhodesiense which is responsible for an early-
onset, acute disease found in Eastern and Southern Africa (Solano et al., 2003). Human African
trypanosomosis represents a major public health threat in Africa and together with nagana, the
animal form of African trypanosomosis, is considered a main obstacle for development of rural
regions of the continent (Simarro et al., 2008). Estimates put the number of cattle at risk from
trypanosomosis between 50-70 million animals (Geerts and Holmes, 1998).The impact of the
tsetse-associated disease extends in sub-Saharan Africa over 10 million (a third of the
continent). Trypanosomosis in Africa costs livestock producers an estimated 1340 million USD
each year (Radostits et al., 2007). If lost potential in livestock and crop production are considered,
then trypanosomosis is costing Africa an estimated five billion USD per year (ILRAD,1994).
To prevent and treat trypanosomosis different trypanocidal drugs are used. The frequency with
which treatment has to be applied is often to economically unacceptable levels. About 35 million
doses of drugs are used in Africa each year, with about 50-70 million animals at risk from
trypanosomosis (Geerts et al., 2001). Since most of trypanocidal drugs have been in use for more
than half a century, they can cause the appearance of the drug resistant strain of trypanosomes.
Until new trypanocidal drugs are available, it is of outmost importance that measures are taken to
avoid or delay the development of resistance so that the efficacy of the currently available drugs is
maintained.

2
Shared characteristics of the different species of African trypanosomes include the ability to
produce almost unlimited antigenic variation of their variant surface glycoprotein (VSG) and to
induce a predominantly T cell-independent antibody response to the VSG, profound
immunosuppression, polyclonal B cell activation and persistent hypo-complementemia in infected
mammalian hosts (Pan et al., 2006). Infection of the mammal host leads to cycle of parasitemia
associated with new VSGs. Each new VSG initially elicits a strong immunoglobulin M (IgM)
anti-VSG response which leads to phagocytosis of the trypanosomes, predominantly by
macrophages of the liver (Naessens, 2006). The present day methods for the control of African
trypanosomosis, namely, systemic case detection and treatment, tsetse control, do not more than
limit the disease although both these approaches have been shown to be effective where they have
been vigorously applied (Delespaux et al., 2008).
Therefore, the general objective of this seminar paper is:
►To review the drug and drug resistance of African animal trypanosomosis
Specific objectives:
►To illustrate causes of trypanocidal drug resistance
►To discuss types of trypanocidal drug resistance
►To highlight mechanism of development of trypanocidal drug resistance
► To show current situation of trypanocidal drug resistance
► To indicate impact of trypanocidal drug resistance
► To elaborate methods of detection of trypanocidal drug resistance

3
2. AFRICAN TRYPANOSOMES
Trypanosomes belong to the phylum Sarcomastigophora, order Kinetoplastida, family
Trypanosomatidae, and genus Trypanosoma. They are unicellular flagellated protozoan
haemoparasites characterized by one nucleus and one flagellum, either free or attached to the
parasites body by means of an undulating membrane (Bourn et al., 2005). Trypanosomes also
usually contain a small, compact kinetoplast, a disc-shaped DNA-containing organelle, situated
within a large mitochondrion (Brun et al., 2009). Within the subgenus, Nannomonas there are
three major trypanosome species; T. brucei, T. equiperdum, and T. evansi. T. brucei can further be
divided into three subspecies, of which T. brucei rhodesiense and T. brucei gambiense are the
causative agents of the debilitating sleeping sickness disease. T. b. brucei and T.b. congolense
(belonging to the subgenus, Nannomonas) and T. vivax (belonging to the subgenus Duttonella),
are associated with trypanosomosis of domestic (Hoare, 1972).
2.1. Morphology and characterization
The trypanosome species differ in morphological characteristics as described by (Maudlin et al.,
2004). All these trypanosome species have a size range of 15-55 μm and typically live in the
blood, lymph, and tissues of their hosts. Bloodstream forms of trypanosomes are covered by a
protective surface coat consisting of variant surface glycoproteins (VSG) linked in turn to the
plasma membrane surface by means of glycosyl-phosphatidyl-inositol (GPI) anchors (Vickerman,
1985).
African trypanosomes are characterized on the basis of their size, shape, position of the nucleus,
size, and location of the kinetoplast, host range, and geographical distribution. Generally they are
elongated, spindle-shape organisms with a single flagellum (Morrison et al., 2009). The flagellum
originates from the basal body near the kinetoplast and runs the length of the trypanosome. The
pellicle, the layer bordering the cytoplasm, while maintaining a definite shape, is flexible enough
to permit a certain degree of body movement. The pellicle and the cytoplasm are pinched up into a
thin sheet of tissue along the length of the body forming the undulating membrane (Soltys and
Woop, 1997).

4
2.2. Life cycle
Trypanosomes are pleomorphic, single-celled parasites with a two-host life cycle: mammalian and
arthropod (Brun et al., 2009). With the exception of T. equiperdum and T. evansi, the majority of
trypanosome species undergo a developmental phase in insect vectors, the tsetse fly (Vickerman,
1985). The cycle (Figure 1) starts when blood from a trypanosome infected animal is ingested by
the tsetse fly. It is within the insect vector that trypanosomes undergo a chain of events involving
differentiation, multiplication and biochemical alterations, such as swapping their energy
metabolism from glucose (in bloodstream forms) to proline (in procyclic forms), before migrating
to the salivary glands, where they progress into infective metacyclic forms by re-gaining their
VSG coat and are then ready to be inoculated into a new host during the next blood meal
(Vickerman, 1985). Trypanosoma brucei group of trypanosomes migrate from the gut to the
proventriculus, to the pharynx and eventually to the salivary glands; the cycle for T. congolense
stops at the hypopharynx, and the salivary glands are not invaded; the entire cycle for T.vivax
occurs in the proboscis. The animal-infective form in the tsetse salivary gland is referred to as the
metacyclic form (Delespaux et al., 2008).
Once inoculated in a new host, trypanosomes quickly lose their surface coat, transform into the
long slender trypomastigotes and proliferate by binary fission at the site of the bite for a few days,
leading to an inflammatory chancre. The parasites, then, spread to the draining lymph nodes and
the bloodstream (first or early haemolymphatic stage of infection), through which they reach other
organs such as the spleen, liver, heart and endocrine system (Delespaux et al., 2008). The T.
brucei group of trypanosomes (T.b.brucei, T. b. gambiense, T. b. rhodesiense) and T. evansi
mostly invade tissues (humoral) whereas, T. congolense and to a lesser extent T. vivax and T. cruzi
predominantly restrict themselves to the blood circulation (haemic) (Morrison et al., 2009).

5
Figure 1. Schematic representation of the digenetic life cycle of Trypanosoma brucei in the
mammalian host and in the tsetse fly vector
Source: Blum et al., 2008
2.3. The vector of African trypanosomosis
Tsetse flies, the vectors for African trypanosomosis (AT), belong to the family Glossinidae, order
Diptera the two winged flies. There are 31 recognized Glossina species and sub-species, divided
into three groups (morsitans, palpalis and fusca) which have been given sub-generic status
(Solano et al., 2010). Recently, comparative gene sequence analysis and geometric wing
morphometry have been proposed to help in the Glossina group identification (Patterson and
Schofield, 2005). The morsitans group that includes G.morsitans morsitans, G. m. submorsitans,
G. pallidipes, G. longipalis and G. austeni is found mainly in the savannah ecosystems. They are
the most important vectors of bovine trypanosomosis (Leak, 1999). The palpalis group is found
mainly in the riverine galleries of West and Central Africa but sometimes extends into savannah
regions between the river systems (Hendrickx et al., 2004). The palpalis fly species are less
mobile than the morsitans group, often relying on sight rather than smell to locate their hosts
(Leak, 1999). In West Africa, important bovine trypanosomosis vectors among the palpalis group
include G. palpalis palpalis, G. p. gambiensis and G. tachinoides (Solano et al., 2010). The fusca
group flies settle mainly in forests and are therefore less important vectors of bovine
trypanosomosis. Glossina longipennis and G. brevipalpis found in the drier areas of Kenya are

6
exceptions among the fusca group, as they have been demonstrated to transmit trypanosomes
(Makumi et al., 2000).
Adult Glossina species are dull in appearance, varying in colour from light yellowish brown to
dark blackish brown (Leak, 1999). In some species the abdomen may have alternate darker and
lighter bands. The smallest species is 6-8 mm long and the largest 10-14 mm (Jordan, 1986). The
adult female produces a single egg, which hatches to first stage larva in the uterus. After a period
of development and moulting, a third stage larva is deposited on the ground. Females produce one
full grown larva every 8-10 days which pupates in light clay or sandy soil. The adult fly will
emerge after a puparial period that varies according to temperature but may be around 30 days at
240C. Consequently, tsetse flies have a very low rate of reproduction, closer to that of a small
mammal than to most insects. This reproductive method of tsetse flies is known as adenotropic
viviparity (Leak, 1999). Females are receptive to males as soon as they start seeking food and are
often mated soon after taking their first blood meal (Leak, 1999). They are usually mated once
with viable sperms remaining stored in the spermathecae throughout the life of the female from
where the sperms get nourished by secretions from alayer of cells surrounding the cuticular lining
of the lumen of each spermathecae. In certain instances, mating may happen more than once. Male
flies may not mate soon after emergence from the pupa and they are not fully fertile until they are
a few days old. Other than tsetse flies, other haematophagous insects like tabanids and stomoxys
species also transmit trypanosomosis mechanically as has been demonstrated by (Desquesnes and
Dia (2003).
2.4. Host
Trypanosomosis is known to affect a number of mammalian vertebrates, either as African animal
trypanosomosis (AAT) and or as human African trypanosomosis (HAT) (Leak, 1999).
Trypanosoma vivax, T. congolense and T. brucei, for example, affect various ungulates including
cattle, sheep, goats, horses, pigs and camels (Maudlin et al., 2004). Other animals like dogs, cats
and the wild carnidae are also affected (Hoare, 1972). Trypanosoma evansi, principally a parasite
of camels and equines, also infects other animals like water buffaloes, sheep, goats, cattle and
deers. Trypanosoma vivax and T. evansi by virtue of their transmission by haematophagous biting
flies occur in sub-Saharan Africa, Asia, Central and North and Southern America (Hoare, 1972).

7
The silvatic cycle that involves wild animals is known to greatly influence the epidemiology of
trypanosomosis since wild animals serve as reservoirs for both human and animal trypanosomosis
(Taylor and Authie, 2004). Trypanosomosis maintains large areas of Africa (so-called ʻfly beltsʼ)
free of livestock, and it is presumed that wildlife have developed an evolutionary immuno
tolerance to these parasites with which they have cohabitated for millennia (Reichard, 2002).
2.5. Risk factors of trypanosomosis
2.5.1. Animal factor
A number of factors contribute to the severity of disease in its various host. The animal and
environmental factors,play important roles in modulating the severity of the disease (Taylor and
Authie, 2004). For animal factor exotic animals (dairy cattle) are more severely affected by
trypanosomosis than the local genotypes, which exhibit a range of breed and individual animal
susceptibility. The West African taurine breeds, like the N'Dama, Baoule and their crosses with
zebu (Dieteren et al., 1998), and certain zebu cattle in East Africa (Njogu et al., 1985) can survive
and remain productive under trypanosomosis risk. This phenomenon is called trypanotolerance
and involves the ability of the animals to control parasitaemia, maintain weight and resist anaemia
(Murray et al., 1982). Established through experimental work that animals within the
trypanotolerant breeds with previous exposure to trypanosomosis suffer less severe
trypanosomosis effects as compared to those without previous exposure (naive animals). Suckling
calves are also known not to suffer from serious attacks of trypanosomosis, possibly because of
the influence of maternal antibodies in their systems (Dwinger et al., 1992). There is also
evidence from studies that tsetse get attracted mostly to larger cattle from which they feed rather
than smaller ones (Torr and Mangwiro, 2000).
According to Torr and Mangwiro (2000) that a large ox was bitten ~10 times more often by tsetse
than a calf. Within herd differences have also been registeredwhere ~75% of the tsetse feed from
~25% of the herd (Torr et al., 2001). It would be expected that animals with concomitant
infections with other parasites like haemonchus species would develop serious disease when
infected by trypanosomes, particularly T. congolense, as was reported in the Gambia (Kaufmann
et al., 1992).

8
2.5.2. Environmental factor
The environment allows for the interaction between the Glossina species, vertebrate hosts and the
trypanosomes in order for trypanososmosis to be produced. In West Africa, tsetse habitats have
been sub-divided along distinct North-South climatic gradients, with predominantly riverine tsetse
species in the North and a mixture in the South (Hendrickx et al., 2004). In the North, arid
conditions prevent fly spread and riparian vegetation constitutes suitable niches for the localized,
well-demarcated pockets of tsetse populations. Outside these favourable microclimates, tsetse
hardly survives and it would appear that no links exist between pockets, except occasionally and
in spatially limited neighbouring areas during the rainy seasons. In the intermediary band, climatic
conditions and vegetation become gradually more suitable. Distinct fly pockets tend to merge and
tsetse distribution patterns become more linear along main streams. Tsetse populations still remain
concentrated in pockets during the dry season, but disperse during the rainy season over large
parts of the river systems, including important tributaries and savannah buffers (Bouyer et al.,
2006). In the humid South, there are no climatic limitations to fly distribution and flies are present
along river sytems and even the surrounding humid woodlands and forests. Due to increasing
human population and consequently the opening up of more land for crops, the morsitans group is
disappearing in most places of West Africa (Djiteye et al., 1997).
2.6. Trypanosomosis control methods
Controlling the vector remains theoretically the most desirable way of containing trypanosomosis
(Leak, 1999). It is a strategy that has worked well in many areas where multiple drug resistance
has been reported before (Fox et al., 1993). Vector control methods available include:
2.6.1. Sequential aerosol technique
The sequential aerosol technique involves the ultra-low volume spraying of non-residual
insecticides 10-15 metres above tree canopy by fixed wing aircraft or helicopter (in more difficult
terrain) in 5-6 subsequent spraying cycles, separated by 16-18 days depending on temperature
(Allsopp and Hursey, 2004). The goal is to kill all adult tsetse flies in the first spraying cycle and
then kill all emerging flies in the subsequent cycles before they start reproducing. It remains a
perfect method if done under global positioning system navigation, especially for effective area-

9
wide tsetse suppression (the efficacy of this technique still needs to be assessed in dense humid
forest ecosystems) or even eradication (in open savannah-type ecosystems) (Allsopp and Hursey,
2004). The disadvantage with the method is that insecticides sprayed may also kill non-target
insects.
2.6.2. Stationary attractive devices (traps and targets)
Stationary attractive devices attract and either kill the flies through tarsal contact with insecticides
embedded in the fabric or the flies are guided and trapped in a non-return cage (Reichard, 2002).
This technique is suitable for deployment by local farmer communities to treat small areas, but the
high target densities required against certain species and in certain dense habitats make the use of
these devices over large areas uneconomical (Kappmeier et al., 2007). The major disadvantage of
Stationary attractive devices is that the active ingredient gets washed off by rain water, hence
compromising its efficacy. Increasing the concentration of insecticide to 0.6-0.8% allows the
Stationary attractive devices to remain deployed even during the wet season, retaining tsetse
mortality rates of > 90% for about 300 days (Torr et al., 1992). Blue fabrics lose colour and
become inefficient after a short time, depending on the dye used and method of fixation. Theft of
the targets, bush fires and maintenance problems are some further problems associated with use of
Stationary attractive devices (Leak, 1999).
2.6.3. Live bait technique
This method involves the application of insecticide onto cattle as pour-ons, sprays or dips so that
tsetse flies attempting to feed on the treated cattle get killed on picking up a lethal deposit of the
insecticide through their tarsi and pre-tarsi. Unlike Stationary attractive devices, the live bait
technique is less prone to theft and does not suffer from maintenance problems. Because of its
added advantage of also controlling ticks, the use of live baits is appreciated as a private good and
can easily be adopted by the rural farming community (Torr et al., 2002). Other disadvantages
with the live bait treatment schemes are the high treatment frequency, the high cost of the
insecticides, insecticide residues in cattle dung, motivation and participation of farmers and the
potential development of resistance to the insecticides in ticks and insects with a high
reproductive rate (Vale and Grant, 2002). It is also thought that the use of insecticides on live

10
animals has the profound effect of interfering with the enzootic stability that is manifested in
young indigenous cattle when exposed to tick challenge (Bourn et al., 2005). Since the attachment
sites of ticks and the feeding sites of tsetse differ, treating only legs and bellies of older cattle will
help, as >95% of tsetse feed come from on adult cattle (Torr et al., 2001). This approach will
more effectively control tsetse without threatening the enzootic stability in calves.
2.6.4. Sterile insect technique
The Sterile insect technique is used if the objective is tsetse eradication. The introduction of the
Sterile insect technique helped eradicate the fly from this island in 1996 in a campaign that had
been commenced two years earlier (Reichard, 2002). As a prerequisite, tsetse density has to be
suppressed through the widespread application of insecticide treated stationary attractive devices,
live baits or fly trapping to a point where the sterile insect technique is considered feasible. In
Zanzibar, a sterile insect plant producing 70,000 irradiated pupae weekly was constructed that
made the release of over 7.8 million sterile male flies possible. Dispersal of the irradiated males
over time was done to achieve an estimated ratio of 50 sterile males for every 1 wild male in order
to overwhelm the residual wild tsetse population (Reichard, 2002). The released sterile males in
the target area do out-compete the wild male population for wild females (Vreysen, 2005). Mating
of the sterile males with virgin, native females results in no offspring. With each generation, the
ratio of sterile to wild insects will increase, making this technique more and more efficient with
lower wild female population densities (inversely-density dependent). The Sterile insect technique
is non-intrusive to the environment, has no adverse effects on non-target organisms, is species-
specific and can easily be integrated with biological control methods such as parasitoids, predators
and pathogens (Leak, 1999). There is no threat of resistance development to the effects of sterile
males, provided that adequate quality assurance is assured during the production process and that
the sterile insects cannot get established in released areas as is the case with other biological
control programmes (Vreysen, 2001). In addition, the sterile insect technique necessitates efficient
release and monitoring methods, which have to be applied on an area-wide basis (Vreysen, 2005).
Since the irradiated tsetse are fully capable of developing and transmitting mature trypanosomes
of all three main species pathogenic to cattle (Moloo and Kutuza, 1984), the sterilized tsetse are
fed on either uninfected blood meals or blood-meals are medicated with trypanocidal drugs before

11
the sterilized insects are fed. Sterilized tsetse are less likely to become infected (at least with
Nannomonas and Trypanozoon parasites) after they have taken an uninfected blood meal and
trypanocidal drugs in medicated meals helps reduce the establishment of infections in subsequent
meals.
3. TRYPANOCIDAL DRUGS
In the 37 African countries where animal trypanosomosis is endemic, trypanocides are used for
the control of the disease (Geerts and Holmes, 1998). Drugs have proven sustainably and
sufficiently attractive to the livestock keepers. Three compounds - isometamidium
chloride(ISMM), homidium salts (homidium bromide (Ethidium®) and homidium chloride
(Novidium®)) and diminazene aceturate (DIM) have been and are still in use more than 50 years
since they were released in the market (Holmes et al., 2004).
Figure 2. The structures of the four most commonly used drug in the chemotherapy and
chemoprophylaxis of animal trypanosomosis Africa
Source: Anene et al., 2001
3.1.Prophylactic treatments
Prophylactic treatments target all animals in a herd or a particular group of valuable or ʻat-riskʼ
animals (Holmes et al., 2004). Isometamidium administered intramuscularly at a dose rate of 0.5-
1mg/kg b.w. provides up to 3 months (a range between 2‒22 weeks) protection against pathogenic
trypanosomes of cattle and against T. vivax especially in small stock, T. brucei in equidae and T.
evansi in camels (Geerts et al., 2001). Isometamidium is given either as routine block
treatment(pre-determined intervals) or as strategic block treatment (when challenge reaches a
predetermined threshold). It is recommended that once a year, additional to ISMM, the animals

12
are separately treated with DIM in order to delay the development of resistance, following the
concept of `sanative pair' (Whiteside, 1962).
3.2.Curative treatments
Diminazene aceturate and HOM salts are the main therapeutic drugs used in the management of
clinical trypanosomosis in animals (Holmes et al., 2004). Diminazene aceturate is administered
i.m. at a dose rate of 3.5‒7mg/kg bw the lowest dose being effectiveagainst T. congolense and T.
vivax and the highest dose against T. brucei(Peregrine, 1994). At these dose rates, DIM, in
addition to its curative uses, alsooffers short term protection of up to 2 weeks (Geerts et al., 2001).
Homidium on the otherhand is administered at a dose rate of 1mg/kg b.w. in cattle. In low tsetse
challenge areas, aprophylactic effect of HOM salts has also been observed (Peregrine, 1994).
3.3.Mechanism of action of current used drugs
3.3.1. Diminazene aceturate (Berenil®)
Diamidines are dicationic molecules, which bind to the minor groove of DNA at AT-rich sites.
They exert their biological activity by primarily binding to DNA and then inhibiting one or more
of the DNA dependent enzymes (such as topoisomerases or nucleases) or by directly impeding the
transcription process. Berenil is the only comercial diamidine for treatment of animal
trypanosomosis at this moment, but new diamidines are being investigated as future therapeutic
options. Two diamidine compounds (DB 75 and DB 867) present comparable efficacy at lower
doses than the standard drug diminazene and could be considered as potential clinical candidates
against T. evansi infection (Gilling et al.,2011).
The selectivity of diamidines is primarily due to its selective accumulation in the pathogen rather
than in the host cells. Diminazene aceturate do not cross the blood–brain barrier. The new
diamidine DB75 (furamidine) does not have sufficient penetration capacity into the central
nervous system, but a related compound (DB820, azafuramidine) seems to completely cure a
murine late-stage model of sleeping sickness (Soeiro et al., 2005).
3.3.2. Isometamidium and homidium

13
Isometamidium chloride (Samorin®, Trypamidium®, Veridium®) has been prophylactically or
therapeutically used in the field for livestock suffering from trypanosomosis for several decades. It
was first synthesized by coupling homidium (Ethidium®) with p-aminophenyldiazonium chloride,
this is, by coupling homidium with a part of the diminazene aceturate molecule. It has been
proposed that the main mechanism of action of isometamidium is the cleavage of kDNA-
topoisomerase complexes, causing the desegregation of the minicircle network within the
kinetoplast (Shapiro and Englund, 1990). But Kaminsky et al.(1997) showed that dyskinetoplastic
trypanosomes are at least as sensitive to isometamidium as kinetoplastic lines.
3.3.3.Suramin (Polysulphonated naphtyl urea)
The molecular structures of the dyes provided a starting point for the synthesis of suramin, which
has been used as a trypanocidal drug since 1916 and is still in clinical use although unfortunately
its large scale production for animal use was stopped in the middle of 90’s. Suramin do not cross
the blood brain barrier. Inhibition of several enzymes by Suramin (among others, dihydrofolate
reductase, fumarase, glycerol-3-phosphate dehydrogenase, hexokinase, reverse transcriptase, RNA
polymerase and kinases) has been described (Hagos et al., 2010). Deprivation from cholesterol
and phospholipids by inhibition of the uptake of low density lipoproteins has been proposed as the
main mechanism of action for this drug.
3.3.4. Melarsomine hydrochloride (Cymelarsan®
)
Melarsoprol is an arsenical compound that contains the trivalent arsenic element with a markedly
reactive arsenoxide group. The presence of arsenoxide confers the physiochemical ability of lipid
solubility that allows passage across the blood brain barrier. The veterinary arsenical trypanocide
melarsamine hydrochloride (Cymelarsan®) is a conjugation of melarsen oxide and two equivalents
of cysteamine. It is used mainly to treat T. evansi infections in domestic animals.

14
4. TRYPANOCIDAL DRUG RESISTANCE
4.1. Definition
Drug resistance is the heritable loss of sensitivity of a micro-organism to a drug to which it was
sensitive before. Resistance to the commonly used animal trypanocides has emerged in sub-
Saharan Africa and interfer with effective veterinary management of trypanosomosis.
4.2. Causes of trypanocidal drug resistance
Trypanocidal drug resistance is caused by the exposure of trypanosomes to sub-therapeutic drug
concentrations, resulting from under-dosing and the irrational use of drugs and the lack of proper
diagnosis (Whiteside, 1962). The prolonged and frequent use of trypanocides in high tsetse
challenge areas, even when used at the right doses, is also likely to cause resistance (Geerts and
Holmes, 1998). Furthermore, poor quality drugs have been finding their way on to the market in
some cases, products with no trypanocidal activity have been also identified and in other
situations compound with reduced activity have been marketed. Such products are not only less
effective when used by farmers, but also greatly increase the risk of drug resistance development.
Reduction in drugs accumulation by the target cell or organism and diminished drug activity in
immune-suppressed animals can contribute to the emergence of drug resistance. Thus, drug
resistance can arise either as a consequence of changes in drug concentration at the target site or
alteration in the target, or both. There is experimental evidence that drug-resistant trypanosome
clones accumulate fewer drugs than their sensitive counterparts (Anene et al.,2001).

15
Figure 3. African countries with reported resistance to trypanocidal drugs. *Resistance to
trypanocidal drugs has been reported in animal trypanosomes in that country;
Source: Delespaux et al., 2008
4.3. Types of trypanocidal drug resistance
Two types of resistance against trypanocidal drugs are recognized: single drug resistance and
multiple drug resistance. In single drug resistance, trypanosomosis control still could be achieved
by using one of the drug pairs in which resistance has not developed through the application of the
sanative pair principle (Geerts and Holmes, 1998). However, the second drug should be used with
caution in order to avoid resistance development against it as well. Multiple drug resistance is
resistance concurrently to two or more drugs, making sanative drug pairs ineffective. Multiple
drug resistance can only be counteracted by intervening at the level of the vector (Fox et al.,
1993).
Table 1: Drug resistant trypanosomes in African countries
Country
Trypanosome
species
Animal
Examined
Resistance
% of
Resistance
Resist to

16
D= diminazene; H = homidium bromide (ethidium); I =isometamidium
Source:Geerts and Holmes, 1998
So far, resistance to one or more of the three more commonly used trypanocidal drugs used in
cattle has been reported in at least 17 countries in sub-Saharan Africa (Burkina Faso, Chad, Ivory
coast, Ethiopia, Kenya, Mali, Somalia, Sudan, Tanzania, Uganda, Zimbabwe, Zambia,
Mozambique, Cameroon, Nigeria, Guinea and Central African Republic) (Fig. 3) (Delespaux et
al., 2008). In eight of the 17 countries, multiple resistances have been reported. This is probably
an underestimation of the true situation,because in several countries surveys for resistance have
not yet been carried out or cases of resistance have not been published. Table 1: shows the
Trypanosoma species and type of drugs in which their resistance has been reported (Geerts and
Holmes, 1998).
4.4. Cross and multiple drug resistance
With development mechanisms of drug resistance to one compound, trypanosomes may show of
resistance to compound of the same series and may also to those of other of series thus, in the
phenanthridium series, resistance to pyrithidium bromide leads to resistance to isometamidium
and homidium. There is also cross resistance between quinapyramine and phenanthridium series.
So sometimes, one strain of trypanosome may be resistant to many drugs (multiple drug
resistant), a situation that constitutes a particularly grave threat to livestock production and health
in Africa. In contrast, a number of experimental drug sensitivity studies with suramin and
diminazene in vitro and in rodents have demonstrated that acquisition of resistance to suramin
does not confer resistance to diminazene, suggesting that cross resistance may not exist between
Isolates
Ethiopia T. congolense 12 12 100 D
Burkina faso T. congolense 12 9 75 I
Kenya T. congolense 7 2 29 I
Kenya/Somalia T. vivax 7 6 86 I
Nigeria T. vivax 19 12 63 D, H, I
T. brucei 12 2 17 D, I
Sudan T. congolense,
T.vivax, T.brucei
12 5 42 H
Uganda T. brucei 36 1 12 D, I
Zimbabwe T. congolense 14 6 43 D
17
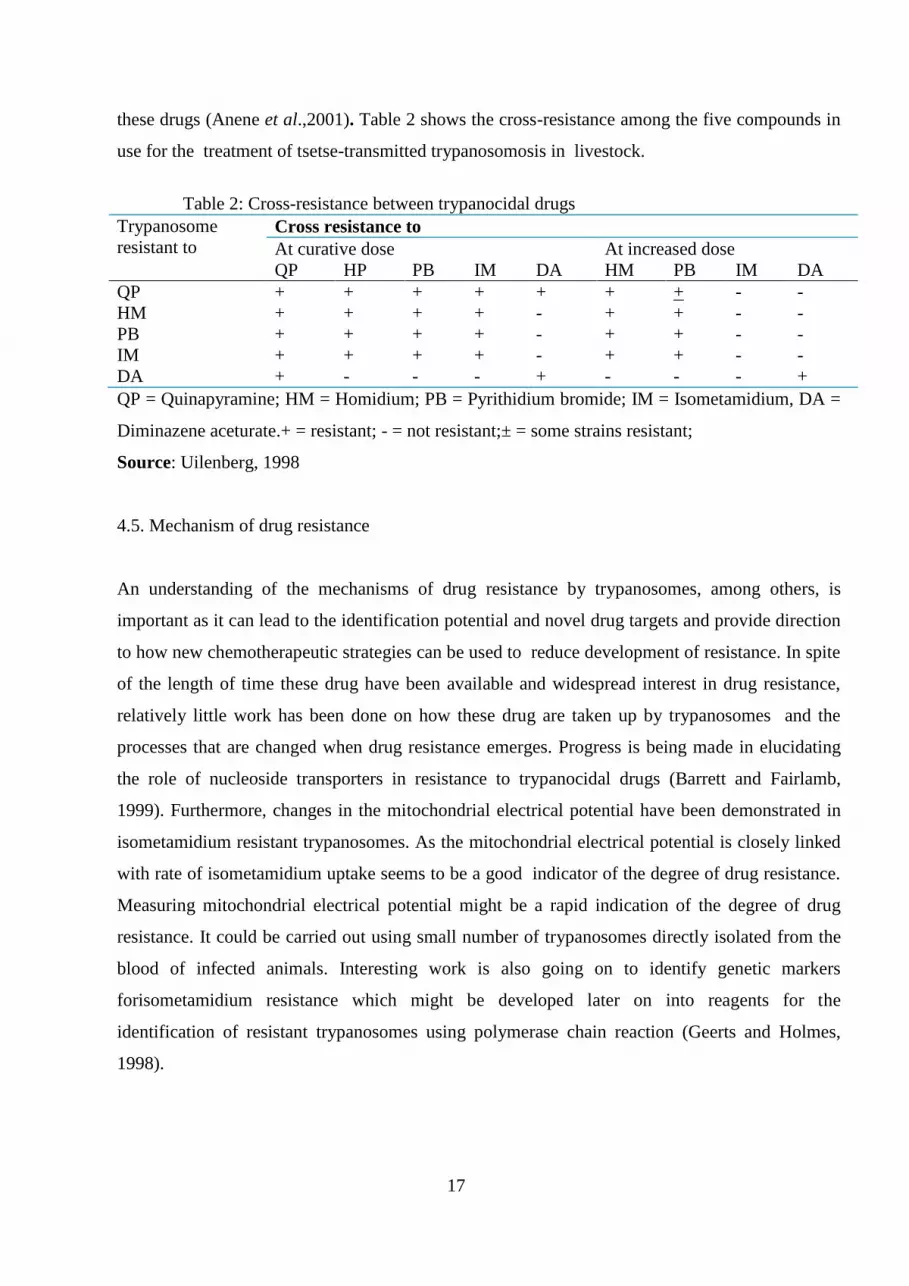
these drugs (Anene et al.,2001). Table 2 shows the cross-resistance among the five compounds in
use for the treatment of tsetse-transmitted trypanosomosis in livestock.
Table 2: Cross-resistance between trypanocidal drugs
Trypanosome
resistant to
Cross resistance to
At curative dose At increased dose
QP HP PB IM DA HM PB IM DA
QP + + + + + + + - -
HM + + + + - + + - -
PB + + + + - + + - -
IM + + + + - + + - -
DA + - - - + - - - +
QP = Quinapyramine; HM = Homidium; PB = Pyrithidium bromide; IM = Isometamidium, DA =
Diminazene aceturate.+ = resistant; - = not resistant;± = some strains resistant;
Source: Uilenberg, 1998
4.5. Mechanism of drug resistance
An understanding of the mechanisms of drug resistance by trypanosomes, among others, is
important as it can lead to the identification potential and novel drug targets and provide direction
to how new chemotherapeutic strategies can be used to reduce development of resistance. In spite
of the length of time these drug have been available and widespread interest in drug resistance,
relatively little work has been done on how these drug are taken up by trypanosomes and the
processes that are changed when drug resistance emerges. Progress is being made in elucidating
the role of nucleoside transporters in resistance to trypanocidal drugs (Barrett and Fairlamb,
1999). Furthermore, changes in the mitochondrial electrical potential have been demonstrated in
isometamidium resistant trypanosomes. As the mitochondrial electrical potential is closely linked
with rate of isometamidium uptake seems to be a good indicator of the degree of drug resistance.
Measuring mitochondrial electrical potential might be a rapid indication of the degree of drug
resistance. It could be carried out using small number of trypanosomes directly isolated from the
blood of infected animals. Interesting work is also going on to identify genetic markers
forisometamidium resistance which might be developed later on into reagents for the
identification of resistant trypanosomes using polymerase chain reaction (Geerts and Holmes,
1998).

18
In addition, alterations of the transporters can cause development of drug resistance (Carter et al.,
1995). There are numerous reports of resistance to Berenil in different countries and in several
Trypanosoma species. In any case, resistance seems to be limited to highly endemic areas where
the use of this drug is very common. Barret et al. (1995) demonstrated that resistance to
diminazene aceturate in T. equiperdum was due to lack of activity of P2 aminopurine transporter,
required to translocate the drug across the cell membrane. The role of the P2-type purine
transporter in the uptake of arsenical diamidines, pentamidine and DA by T. brucei, T. evansi and
T. equiperdum, and the consequences of inhibition, knocking down or silencing this gene have
been extensively described and reviewed in the literature (De Koning et al., 2004)
These transporters have been cloned and expressed in yeasts to demonstrate their role in resistance
(Matovu et al., 2003). But P2 is not the only significant transporter for melaminophenyl arsenicals
expressed in bloodstream trypanosomes. Some theories of heavy metal resistance in protozoa
proposed the implication of aqua-glyceroporins (Gourbal et al., 2004), thiamine transporters
(Schweingruber, (2004) or high affinity pentamidine transporter (HAPT1) (Matovu et al., 2003) ,
but it seems to be that these molecules do not play an important role in vivo.
4.6. Current situation of resistance
Currently, there are close to 20 African countries in which resistance has already been reported
(Delespaux et al., 2008). In addition to the 13 countries mentioned by Geerts and Holmes, (1998),
resistance has been reported in Mozambique (Jamal, 2005), Mali, Guinea (Diall et al., 2003),
Cameroon (Mamoudou et al., 2008) and recently in Ghana and Benin (Allegye-Cudjoe, 2009). It
is suspected that in several other African countries, resistance is present but is yet to be
demonstrated (Delespaux et al., 2008). Large scale surveys have been conducted in 13 African
countries including Kenya (Mdachi, 1999), Uganda, Tanzania (Eisler et al., 2000), Ethiopia
(Tewelde et al., 2004), Zambia (Shinyangwe et al., 2004), Zimbabwe (Joshua et al., 1995),
Cameroon (Mamodou et al., 2008), Nigeria (Geerts et al, 2001), Burkina Faso(McDermott et al,
2003), Mali, Guinea ( Grace, 2005) Ghana and Benin (Allegye-Cudjoe, 2009) demonstrating area-
wide resistance in at least one regionof these countries.Confirmed reports about resistance in the
cotton zone of West Africa were first made particularly in Burkina Faso in the early 1980s (Pinder
and Authie, 1984).These authors described stocks of T. congolense isolated from cattle in the

19
Samorogouan area in 1982 which were resistant to isometamidium. Later, tests in mice showed
that certain T.congolense strains from the same area were also resistant to diminazene aceturate
indicating existence of multiple drug resistant T. congolense (Authie, 1984). Recent studies on
resistance in the same area of Burkina Faso (McDemott et al., 2003) and in other areas of West
Africa underline that the problem is present and is expanding (Allegye-Cudjoe, 2009).
4.7. Impact of drug resistance
It is essential to assess not only the distribution of drug resistance, but also the constraint it
imposes on effective control. To date, few studies have accurately assessed the impact of drug-
resistant trypanosomes on livestock productivity, although it is generally assumed that
uncontrolled infections will have a severe impact on both survival and productivity of animal. A
useful recent study to assess the impact of drug-resistant trypanosomes on the productivity of the
local cattle was carried out in the Ghibe valley, Ethiopia, where a high prevalence of multiple drug
resistance was reported. In the study, incidence of abortion was increased and the financial and
economic returns were also affected (Anene et al., 2001).
4.7.1. Factor influencing development of resistance to Trypanocidal drugs
The exposure of trypanosomes to sub-therapeutic concentrations of trypanocidal drugs, the
treatment frequency and the degree of drug exposure of the parasite population are important
factors influencing the development of drug resistance (Geerts and Holmes 1998). Furthermore,
some trypanocidal drugs such as ethidium are well-known mutagenic compounds and might
induce mutations, the most resistant of which might be selected under drug pressure (Holmes et
al., 2004). The phenomenon of cross-resistance has now been clearly demonstrated.
Quinapyramine usage has been shown to induce resistance to isometamidium, homidium and
diminazene (Uilenberg, 1998). Research on drug resistance in Plasmodium has shown also that
the genetic structure of a parasite population (clonal or panmictic) is an important parameter
influenced by the transmission intensity, and this in turn might influence the rate of development
of drug resistance (Holmes et al., 2004).

20
Figure 4. Some factors influencing the development of resistance to trypanocidal drugs;
Source:Geerts et al.,2001
4.8. Detection of drug resistance
4.8.1.Field methods
Eisler et al. (2000) proposed a method for the assessment of prevalence of resistance to
isometamidium chloride by monitoring cattle populations under natural challenge in the field.
Briefly, two groups consisting of 30 to 80 cattle each are used. One group is treated with 1 mg/kg
b.w. ISMM and the other is used as untreated control. The two groups then are exposed to natural
challenge and tested for the presence of trypanosomes using the phase contrast buffy coat
technique (BCT) (Murray et al., 1977) every two weeks for two to three months. A comparison
through survival analysis curves is made on the data of new trypanosome infections between the
group of cattle treated with ISMM and the untreated control group (Eisler et al., 2000; Tewelde et
al., 2004). If >25% of the ISMM treated cattle become infected within 8 weeks of exposure, drug
resistance is strongly suspected (Mdachi, 1999; Eisler et al., 2000; Tewelde et al., 2004).
Several epidemiological studies to map field trypanocidal drug resistance, based on the protocol
by Eisler et al. (2000). McDermott et al. (2003) working in the Kenedougou Province of Burkina
Faso, Shinyangwe et al. (2004) working in Eastern Zambia, Tewelde et al. (2004) in Ethiopia,

21
Grace (2005) in Guinea and South-Eastern Mali and Allegye-Cudjoe (2009) in Ghana and in
Benin are some examples. An abbreviated version of the original 8‒12 week-protocol by Eisler et
al. (2000) was validated in the cotton zone of West Africa and found effective and reliable for use
not by researchers but by the national veterinary services. This involves a 4 week long follow-up
(Diall et al., 2003) period in order to reduce costs and still generate data within a very short time.
The abbreviated protocol is effective in areas where trypanomosis risk is high (prevalence is
>10%) as has been demonstrated in the cotton zone of West Africa (Diall et al., 2003; Grace,
2005). Rowlands et al. (1994) developed a model to distinguish new and recurrent infections to
determine if the high infection rates observed in cattle in the Ghibe valley, south-west Ethiopia,
following treatment of T. congolense infections with diminazene aceturate were due to the tsetse
challenge or if they rather were a relapse of infections following treatment. An infection was
defined as new if it was preceded by two previous months in which monthly collected samples
had packed cell volumes (PCV) of ≥ 26% and in which trypanosomes were not detected.
4.8.2. Drug sensitivity studies in experimental animals
Tests in ruminants
Neither the single-dose nor the multiple-dose tests in mice are able to predict accurately the
curative doses of trypanocidal drugs needed to clear trypanosome populations from infected cattle
(Eisler et al., 2001). The test in ruminants should hence be used to just determine whether or not
drugs are principally efficacious at recommended curative doses in cattle infected with a particular
trypanosome populations. The test in calves may further be used for investigations on drug
resistance in T. vivax, which is usually not infective for mice. A group of cattle or small
ruminants, preferably of a breed native to the area and without prior exposure to tsetse or
trypanosomosis are used (Eisler et al., 2001). They should also be negative for anti-trypanosomal
antibodies as determined by the indirect fluorescent antibody test or ELISA (Luckins and Mehlitz,
1978) if these tests are available. Specific detailed protocols on this are as contained in Eisler et
al. (2001). Duereduction in drugs accumulation by the target cell or organism and diminished
drug activity in immune-suppressed animals can contribute to the emergence of drug resistance.
Thus, drug resistance can arise either as a consequence of changes in drug concentration at the
target site or alteration in the target, or both. There is experimental evidence that drug-resistant
trypanosome clones accumulate fewer drugs than their sensitive counterparts (Anene et al.,

22
2001).to individual variation in the response to trypanocidal drug treatment among ruminants
inoculated with the same T. congolense isolate (Peregrine et al., 1991), it is advisable to use a
minimum of three and preferably six animals. However, economic considerations may often
preclude the use of more than a single animal per stabilate for drug-sensitivity testing.The
experimental animals must be kept in a fly-proof stable or in a non-tsetse infested area to
eliminate the risk of reinfection during the study. A breakthrough infection, indicative that one of
the inoculated trypanosome populations was drug-resistant can be inoculated into a group of
calves and mice to determine the level of drug resistance. A variation of this method also exists
whereby blood from a group of infected cattle is pooled and inoculated into a single recipient calf
which is monitored and later, if parasitaemic, treated with trypanocide at the recommended dose.
This technique is appropriate where laboratory facilities are limited but only allows for a
qualitative assessment of resistance. Further constraints of the technique are that not all
trypanosome populations might grow equally well and that sensitive isolates might overgrow
resistant ones when inoculated together (Sones et al., 1989); this however, is not a consistent
observation (Burudi et al., 1994). A useful indication of the level of resistance can be obtained
from studies in ruminants by recording the length of time between treatment and detection of
break through populations of trypanosomes. The shorter the period, the greater the level of
resistance (Ainanshe et al.,1992).
Tests in mice
Either single-dose or multi-dose tests are conducted in mice to provide information on resistant
trypanosome isolates from a given area, as described in the protocol by Eisler et al. (2001). After
expansion of an isolate in a donor mouse, experimental mice are inoculated with the test
trypanosome isolate and treated with a trypanocidal drug. Tail blood wet smears are checked 2‒3
times per week for parasites for a period of up to 60 days. The ED50 and ED95 (effective dose
that gives temporary clearance of the parasite in 50% or 95% of the animals, respectively) can be
calculated as can the CD50 and CD95 (curative dose that gives complete cure in 50 and 95% of
the animals, respectively). Sones et al. (1988) used a group of five mice, which allowed an easy
calculation of ED80 and CD80 values (one out of five mice not cleared or cured). Knoppe et al.
(2006), using the standard mouse test (SMT), screened a number of T. congolense isolates
collected in the Kenedougou Province of Burkina Faso against isometamidium chloride at a dose

23
of 0, 0.25, 1.0, 5.0, 10, 15 or 20 mg/kg b. w. and found the method very sensitive but labour
intensive.
There are however several disadvantages with this method. Firstly, most T. vivax isolates, and also
some T. congolense isolates, do not grow in mice (Holmes et al., 2004). Secondly, although there
is a reasonable correlation between drug sensitivity between mice and cattle, higher doses of drugs
must be used in mice (normally ten times higher) in order to obtain results comparable to those
from cattle, because of the vast different metabolic size (Sones et al., 1988). Thus, the curative
dose for ruminants cannot be extrapolated from the assay results in mice (Sones et al., 1988).
Thirdly, a danger further exists of selecting against particular trypanosome species, particularly in
mixed infections. Fourthly, precise assessment of resistance requires a large number of mice per
isolate. Finally, it takes as long as 60 days to evaluate the drug sensitivity of an isolate.
4.8.3. In vitro assays
In vitro assays use blood stream or metacyclic forms instead of procyclic forms. This technique
has been used to detect resistance in T. brucei and T. congolense (Clausen et al., 2000). It takes up
to 40 to 50 days of in vitro incubation to generate metacyclic trypanosomes (Gray and Peregrine,
1993). The advantage with this technique is that large numbers of isolates can be examined and
tests with metacyclic trypanosomes correlate well with field observations. However, in vitro
cultivation of bloodstream forms is only possible using pre-adapted lines and not isolates directly
from naturally infected animals (Hirumi et al., 1993). A simplified axenic culture system has been
developed by these authors, but further research is still necessary to study the correlation with
field data. A potential problem associated with this lengthy time adaptation is the possible
selection against trypanosomes that possess the phenotype of the original population. Further, in
vitro assays are quite expensive and require good laboratory facilities and well-trained staff.The
drug incubation infectivity test (DIIT) combining both in vivo and in vitro techniques issuitable
for drug sensitivity testing of T. b. brucei, T. evansi and T. vivax (Kaminsky et al., 1990). It was
modified by Sutherland et al. (1991) and it proved suitable for T. congolense.
4.8.4. Xenodiagnosis

24
Xenodiagnosis is the feeding of a clean susceptible vector species on a suspected case of
trypanosomosis, after which it is either dissected and examined for the presence of infection, or
allowed to feed on a clean animal which then is itself examined for the development of infection.
A modification of this approach, the drug incubation glossina infectivity test (DIGIT), in which
trypanosomes are exposed to the trypanocidal drugs in vitro for a short time and thereafter are fed
to tsetse flies to check whether or not they develop into metacyclic forms was successfully
validated and proved sensitive for detecting drug resistance (Clausen et al., 1999). This technique
distinguishes resistant from sensitive isolates and does not require experimental animals.
However, it does require a ready supply of teneral tsetse flies from an artificially reared colony.
4.8.5. Serological techniques
Although not frequently used, enzyme linked immuno-sorbent assay (ELISA) has proved valuable
in diagnosing isometamidium resistance (Eisler et al., 1996). The use of ELISA in the detection of
ISMM in the serum of cattle can becombined with field block treatment studies or for individual
treatment of ruminants to detect resistant trypanosomes (Eisler et al., 1996). The presence of
trypanosomes in animals with an ISMM serum concentration > 0.4ng/ml suggests that parasites
are resistant (Eisler et al., 1997). Similar drug-ELISAs have been developed for the detection of
sub-nanogramme amounts of homidium and diminazene (Holmes et al., 2004). A closely related
technique to drug-ELISAs is the mitochondrial electrical potential (MEP) which determines the
rate of ISMM accumulation in the trypanosome kinetoplast (Wilkes et al., 1997).
4.8.6. Molecular techniques
i. Polymerase chain reaction
Because of the problems associated with the low sensitivity of the parasitological techniques
(Paris et al., 1982) and the long follow-up time of study animals (Eisler et al., 2001), polymerase
chain reaction (PCR) with high sensitivity and specificity is a good solution to these problems.
Gall et al. (2004) used this method in Burkina Faso and found it four times more sensitive
compared to the field parasitological techniques.
ii. PCR-RFLP

25
Molecular methods for the diagnosis of ISMM resistance were recently developed (Afework et
al., 2006). The first method enables discrimination between ISMM-ISMMsensitive and ISMM-
resistant strains of T. congolense by PCR-RFLP (Delespaux et al., 2005). This test is based on the
polymorphism observed in the 381 bp fragment (in sensitive strains) or the 384 bp fragment (in
resistant strains) of a putative gene presenting some homologies with transporter. The second
method has been developed to distinguish ISMM-resistant from ISMM-sensitive strains of T.
brucei (Afework et al., 2006). This PCR-RFLP test is based on the polymorphism of the 677 bp
fragment of the TbAT1 gene. The same set of six point mutations could confer resistance to the
melarsenoxide cysteamine cymelarsan (an arsenical diamidine) and to ISMM (diamidine
compound) and the detection of one of these six mutations could enable reliable identification of
sensitivity or resistance to ISMM (Maser et al., 2003).
4.9. Measures to combat drug resistance in the field
As it was mentioned above, drug resistance in trypanosomes is likely to occur under certain
circumstances such as: i) under large-scale drug use; ii) by using inadequate dosing; and iii) by
using correct dosing with drugs that are slowly eliminated from the body. Furthermore, some
trypanocidal drugs are well-known mutagenic compounds and might induce mutations, the most
resistant of which are certainly selected under drug pressure. Taking into account of these factors
different measures can be proposed in order to reduce the chance of drug resistance. Of these the
most important measures are use of the correct dose, changing of drugs, sanative treatment,
increased dosage, repeative treatment and use of combined drugs. In addition to these, care must
be taken to avoid fake drugs and good quality assurance must be implemented (Geerts and
Holmes, 1998).
4.9.1. Use of the correct dose
Under dosing is one of major causes of resistance development. Sub-therapeutic drug
concentrations exert a strong selective pressure for the emergence of resistant clones that pre-exist
in the trypanosome population. Unfortunately, under dosing occurs very frequently. Farmers have
the tendency to underestimate the weight of their animals when they have to treat them since

26
farmers or unskilled persons in many countries of Africa are administering drugs due to absence
of strict rules about the utilization of veterinary drugs (Geerts and Holmes, 1998).
4.9.2. Changes of drugs
Changing drugs or alterative use of drugs in different time may reduce the chance of drug
resistance. For example one group of chemical can be used for prophylactic purpose and the other
can be applied for curative (Uilenberg, 1998).
4.9.3. Sanative treatment
The concepts of sanative treatment is the use of a pair of trypanocides which are chemically
unrelated and therefore, unlikely to induce cross resistance (Rowlands et al., 1994) Diminazene
and homidium, or diminazene and isometamidium can be used in the field as sanative
combinations. These pairs when strategically employed can be used to maintain herd productivity
in the field without the development of resistance to either of the compounds (Anene et al.,2001).
4.9.4. High dose and repeat treatment regimen
High dose treatment offers the best opportunity for eliminating infections with trypanosomes
which express high degree of resistance to drugs. However, it must be appreciated that the scope
for increased drug dosage is highly dependent on the relationship between the maximal tolerated
dose and the minimal dose required to treat cure (the therapeutic index). This is a major limitation
to high dose treatment with trypanocides as the margin of safety of most of them is usually quite
narrow, trypanocidal drug toxicity being quite common. So this technique is helpful in the
utilization of drugs with wide safety of margin. Studies on the efficacy of repeat treatments of T.
congolense infections with diminazene aceturate indicate that such regimen may be useful
especially if administered at 48 or 96 hour intervals. This tends to support the suggestion that the
efficacy of trypanocides depends not only on the concentration of the drug to which the parasites
are exposed, but also on the length of exposure. But this may not be true for all trypanocidal drugs
(Anene et al.,2001).

27
4.9.5. Use of combined drugs
The rationale for the use of two or more of existing drugs in combinations to increase therapeutic
activity, decrease clinical toxicity and potentially reducing the frequency of the emergence of
drug resistance (Anene et al.,2001).
4.9.6. Beware of fake drugs
Fake trypanocides may be sold in Africa. The drugs are either fake in their composition or are
faked by dilution of the original products or substitutions by an ordinary component apparently a
like (Nere powder for diminazine (berenil), coffee or charcoal for ethidium, potassium
permanganate for isometamidium. For isometamidium, one must pay attention to the information
given on the packages (the examples of a shell found on a fake product labelled “for veterinary
use” and in general carefully check the logo of firms. Use will known products and be regular
customer to trust worthysupply service (Leeflang, 1978).
4.10. Quality assurance of trypanocidal drugs
In recent years, a further issue has arisen associated with the liberalization of veterinary drug
supply and market. The growing problem of poor quality drugs finding their way on to the market
in some cases, products with no trypanocidal activity have been identified and in other situations
compound with reduced activity have been marketed. Such products are not less effective when
used by farmers, but also greatly increase the risk of drug resistance developing (especially when
under dosing also allows the survival for the heterozygote resistant trypanosomes). Unfortunately,
quality control on pharmaceutical products used in the developing world is frequently inadequate
and there is already considerable evidence that the problem is widespread for a variety of
pharmaceutical products (Shakoor et al., 1997).

28
5. FUTURE PROSPECT IN TREATMENT FOR TRYPANOSOMOSIS
Trypanosomosis is controlled either by controlling the vector or by controlling the parasite, or a
combination of both. Over the years, a large arsenal of vector-control tools has been developed.
Nevertheless, the control of animal trypanosomosis in often poor rural communities has and will
continue to rely heavily on the use of trypanocidal drugs. However, trypanocidal drugs resistance
has been reported in 17 African countries. In addition to this, some authors discuss the possible
role of inaappropriate usage of trypanocides in livestock in the development of resistance
observed in human trypanosomosis (Barrett, 2001). Trypanosomes have the capacity for antigenic
variation, which is the basis of their ability to escape the host immune response, and because of
this, prospects for the development of a vaccine against trypanosomosis have been considered
poor. Tonin et al., (2011) assessed the use of diminazene aceturate in association with vitamin E
and sodium selenite in rats, concluding that results in terms of longevity, hematocrit reduction,
leukocytes and lymphocytes number and lipid peroxidation were improved using this combinative
therapy compared to the use of single diminazene aceturate, although sodium selenite showed
better protective action than vitamin E. There is a progressive interest in the utilization of
antioxidants in the prevention and treatment of this disease. T. evansi pathogenic mechanisms
include oxidation of the erythrocytes inducing oxidative stress due to free radical generation
(Habila et al., 2012). Pallavi et al. (2010) investigated the potential of heat shock protein 90
inhibitors as drugs for the treatment of trypanosoma infection in animals. This protein regulates
cell cycle progression and signal transduction.There is an urgent need for new drugs for the
chemotherapy of trypanosomosis. Progress has been made in the identification and
characterization of novel drug targets for rational chemotherapy and inhibitors of trypanosomatid
glycosomal enzymes, trypanothione reductase, ornithine decarboxylase, S-adenosylmethionine
decarboxylase, cysteine proteases and of the purine and sterol biosynthetic pathways

29
6. CONCLUSSION AND RECOMMENDATIONS
Despite limited number of trypanocidal drugs, they are more widely used than means to control
the disease. Drug resistance poses a potential treat to control measures. The exact mechanism how
trypanosomal parasite develop resistant and the factors responsible for the development of drug
resistance are yet to be further established. In addition, it is very unlikely that new trypanocidal
drugs will be released into the market in the near future. Therefore, in order to continue to tackle
the economic lose posed by trypanosomosis by using typanocidal drugs, among others strict drug
quality assurance of trypanocidal drugs coupled with proper utilization of currently available
drugs should be implemented. In addition the latest contol method by Sterile insect technique and
Live bait technique because this two control have advantage over other control such as; Sequential
aerosol technique and Stationary attractive devices (traps and targets).
In line with the above conclusion, the following recommendation are forwarded:
The quality assurance of trypanocidal should be used in overall African country.
The vector control method with Live bite technique shuld be should be used because of its
bivalent advantage.
Using of cattle that trypanotolarent in area with high tsetse fly infestation should be
better.for those people live in that area.
The great attention should be given to applying the latest tsetse control method by Sterile
insect techique.

30
7. REFERENCES
Afework, Y., Maser, P., Etschmann, B., von Samson-Himmelstjerna, G., Zessin, K.-H, Clausen,
P.H. (2006): Rapid identification of isometamidium-resistant stocks of Trypanosoma b.
brucei by PCR-RFLP. Parasitol. Res. 99 (3): 253-261.
Ainanshe, O.A., Jennings, F.W., Holmes, P.H. (1992): Isolation of drug-resistant strains of
Trypanosomacongolense from the lower Shabelle region of southern Somalia. Trop.
Anim. Health Prod. 24: 65-73.
Allegye-Cudjoe, E. (2009): Field detection and evaluation of trypanocidal drug resistance in
Ghana and Benin. MSc. dissertation, University of Edinburgh, UK. pp 60.
Allsopp, R., Hursey, B.H. (2004): Insecticidal control of tsetse. CABI International Wallingford,
UK. Pp, 491-507.
Anene, B.M., Onah, D.N., and Y. Nawa, (2001): Drug resistance in pathogenic African
trypanosomes: what hopes for the future? Veterinary Parasitology, 96: 83-100.
Authie, E. (1984): Mise en evidence dune resistance aux trypanocidesparmi des souches de T.
congolenserecemmentisolees au Burkina. Rev. Elev. Med. Vet. Pays. Trop. 37: 219-235.
Barrett M.P. (2001): Veterinary link to drug resistance in human African trypanosomiasis?
Lancet; 358: 603–604.
Barrett, M.P. and A.H. Fairlamb, (1999): The biochemical basis th of arsenical-diamidine cross-
resistance in African trypanosomes. Parasitology Today, 15: 136-140.
Blum, J., Brun, R., Chappuis, F., and Burri, C. (2008): Human African trypanosomiasis. Lancet,
375: 148-159.
Bourn, D., Grant, I., Shaw, A., and Torr, S. (2005): Cheap and safe tsetse control for livestock
production and mixed farming in Africa. Aspects App. Biol. 75: 1-12
Bouyer, J., Guerrini, L., Desquesnes, M., de la Rocque, S., Cuisance, D. (2006): Mapping
African animal trypanosomosis risk from the sky. Vet. Res. 37: 633–645.
Burudi, E.M., Peregrine, A.S., Majiwa, P.A., Mbiuki, S.M., Murphy, N.B. (1994): Response of
diminanzene-resistant and diminanzene-susceptible Trypanosomacongolense to
treatment with diminazene when occurring as a mixed infection in goats. Ann. Trop.
Med. Parasitol. 88: 595-606.

31
Carter, N.S., Berger B.J., Fairlamb A.H. (1995): Uptake of diamidine drugs by the P2
nucleoside transporter in melarsen-sensitive and resistant Trypanosomabruceibrucei.
Journal of Biological Chemistry.270: 28153–28157.
Clausen, P.H., Leendertz, F.H., Blackenburg, A., Tiertjen, U., Mehlitz, D., Sidibe, I., Bauer, B.
(1999): A drug incubation glossina infectivity test (DIGIT) to assess the susceptibility of
Trypanosomacongolense bloodstream forms to trypanocidal drugs (xenodiagnosis). Acta,
Trop. 72: 111-117.
Clausen, P.H., Pellman, C., Scheer, A., Tietjen, U., Schares, G., Bauer, B., Peregrine, A.S.,
Mehlitz, D. (2000): Application of in vitro methods for the detection of drug resistance in
trypanosome field isolates, I.C.P.T.V. 2: 9-12.
De Koning H.P., Anderson LF, Stewart M, Burchmore RJS, Wallace LJM, Barrett MP.( 2004):
The trypanocidediminazeneaceturate is accumulated predominantly through the TbAT1
purine transporter: additional insights on diamidine resistance in African trypanosomes.
Antimicrobial Agents and Chemotherapy, 48: 1515–1519.
Delespaux, V., D. Geysen, P.V. Bossche and S. Geerts, International Health, (2008): Molecular
tools for the rapid detection of drug resistance in animal trypanosomes. Trends in
Parasitology, 24: 236-241
Delespaux, V., Geysen, D., Majiwa, P.A., Geerts, S. (2005): Identification of a genetic marker
for isometamidium chloride resistance in Trypanosomacongolense. Int. J. Parasitol, 35:
235–243.
Desquesnes, M., Dia, M.L. (2003a): Mechanical transmission of Trypanosomacongolense in
cattle by the African tabanidAtylotusagrestis. Exp. Parasitol, 105 (3-4): 226-231.
Diall, O., Clausen, P.-H, Diallo, M.B., Muenstermann, S., Bocoum, Z., Diarra, B., Barry,A.M.,
Bengaly, Z., Affognon, H., Randolph, T., McDermot, J. (2003): Field characterization of
trypanocide resistance in the cotton zone of West Africa. Paper abstract. In: Proceedings
of the International Symposium for Veterinary Epidemiology and Economics (ISVEE)
held in Vina del Mar, Chile, 17th - 23rd November 2003, Pp 1013-1015.
Dieteren, G.D.M., Authie, E., Wissocq, N., Murray, M. (1998): Trypanotolerance, an option for
sustainable livestock production in areas at risk from trypanosomosis. Rev. Sci. Tech.
Off. Int. Epiz. 17 (1): 154-175.

32
Djiteye, A., Moloo, S.K., Foua, B.I.K., Toure, M., Boire, S., Bengaly, S., Coulibaly, E., Diarra,
M., Traore, D., Ouattara, I., Coulibaly, Z. (1997): Reactualisation des donneessur la
repartition des glossines au Mali. Rev. Elev. Med. Vet. Pays Trop. 50: 126-132.
Dwinger R.H., Grieve, A.S., Snow, W.F., Rawlings, P., Jabang, B., Williams, D.J (1992):
Maternal antibodies in N©Dama calves kept under natural trypanosomiasis risk in The
Gambia. Parasite Immunol. 14 (3): 351-354.
Eisler, M., Dwinger, R., Majiwa, D., and Picozzi, K. (2004): Diagnosis and Epidemiology of
African Animal trypanosomiasis. In: Maudlin, I., Holmes, P., and Miles, M. (ed.): The
Trypanosomiases. UK: CABI, CAB International. Pp. 253-267.
Eisler, M.C., Brandt, J., Bauer, B., Clausen, P.-H., Delespaux, V., Holmes, P.H., Ilemobade, A.,
Machila, N., Mbwambo, H., McDermott, J., Mehlitz, D., Murilla, G., Ndung'u, J.M.,
Peregrine, A.S., Sidibe, I., Sinyangwe, L., Geerts, S. (2001): Standardised tests in mice
and cattle for the detection of drug resistance in tsetse-transmitted trypanosomes of
African domestic cattle. Vet. Parasitol, 97: 171-182.
Eisler, M.C., Elliot, C.T., Holmes, P.H. (1996): A simple competitive enzyme immunoassay for
the detection of trypanocidal drug isometamidium. Therapeut.Drug Monitor, 18: 73-79.
Eisler, M.C., Gault, E.A., Moloo, S.K., Holmes, P.H., Peregrine, A.S. (1997): Concentrations of
isometamidium in the sera of cattle challenged with drug-resistant
Trypanosomacongolense. Acta Trop, 63: 89–100.
Eisler, M.C., McDermott, J.J., Mdachi, R., Murilla, G.A., Shinyangwe, L., Mubanga, J., Machila
N, Mbwambo, H., Coleman, P.G., Clausen, P.-H., Bauer, B., Sidibé, I., Geerts, S.,
Holmes, P.H., Peregrine, A.S. (2000): Rapid method for the assessment of trypanocidal
drug resistance in the field. In: Proceedings of the 9th symposium of the International
Society for Veterinary Epidemiology and Economics (ISVEE) held in Breckenridge,
Colorado, USA, August 6th – 11th 2000, Paper 353: 1-3.
Fox, R.G.R., Mbando, S.O., Fox, M.S., Wilson, A. (1993): Effect on herd health and
productivity of controlling tsetse and trypanosomiasis by applying deltamethrin to cattle.
Trop. Anim. Health Prod, 25: 203-214.
Gall, Y., Woitag, T., Bauer, B., Sidibe, I., McDermott, J.J., Mehlitz, D., Clausen, P.-H. (2004):
Trypanocidal failure suggested by PCR results in cattle field samples. Acta Trop, 92: 7-
16.

33
Geerts S, Holmes PH. (1998): Drug management and parasite resistance in bovine
trypanosomiasis in Africa. PAAT Technical Scientific Series No.1.
Geerts, S., P.H. Holmes, O. Diall and M.C. Eisler.(2001): African bovine trypanosomiasis: the
problem of drug resistance. Trends in Parasitology, 17: 25-28.
Gilling water K, Gutierrez C, Bridges A, Wu H, Deborggraeve S, Ekangu RA, Kumar A, Ismail
M, Boykin D, Brun R. Efficacy study of novel diamidine compounds in a Trypanosoma
evansi goat model. PLoS ONE (2011), 6 (6): e20836. doi:10.1371/journal.pone.0020836.
Gourbal B, Sonuc N, Bhattacharjee H, Legare D, Sundar S, Ouellette M, Rosen BP,
Mukhopadhyay R. (2004): Drug uptake and modulation of drug resistance in Leishmania
by an aquaglyceroporin. Journal of Biological Chemistry, 279: 31010-31017.
Grace, D (2005): The epidemiology and control of trypanosomosis in areas of high risk of
trypanocide resistance in West Africa. PhD Thesis, the Freie Universität Berlin, pp 195.
Gray, M.A., Peregrine, A.S. (1993): An in vitro assay for drug sensitivity of Trypanosoma
congolense using in vitro derived metacyclic trypanosomes. Acta Trop, 54: 291-300.
Habila N, Inuwa MH, Aimola IA, Udeh MU, Haruna E.( 2012): Pathogenic mechanisms of
Trypanosoma evansi infections. Research in Veterinary Science, 93: 13-17.
Hagos A, Goddeeris BM, Yilkal K, Alemu T, Fikru R, Yacob HT, Feseha G, Claes,F. (2010):
Efficacy of Cymelarsan® and Diminasan® against Trypanosoma equiperdum infections
in mice and horses. Veterinary Parasitology, 171: 200-206.
Hendrickx, G., de la Rocque, S., Mattioli, R. C. (2004): Long-term tsese and trypanosomiasis
management options in West Africa. PAAT technical series No. 6.
Hirumi, H., Hirumi, K., Peregrine, A.S. (1993): Axenic culture of Trypanosoma congolense:
application to the detection of sensitivity levels of bloodstream trypanomastigotes to
diminazeneaceturate, homidium chloride, isometamidium chloride and
quinapyraminesulphate. J. Protozool. Res. 1: 1-12.
Hoare, A. (1972): The Salivaria. In: Aderem, H. (4th ed.): The Trypanosomes of mammals.
Oxford and Edinburgh: Blackwell Scientific Publications. Pp. 40-609.
Holmes, P.H., Eisler, M.C., Geerts, S. (2004): Current chemotherapy of animal trypanosomiasis.
In: The trypanosomiases. Maulidn, I., Holmes, P.H., Miles M.A. (eds.). CABI
International Wallingford, UK.pp 431-444.
ILRAD, (1994): International Laboratory for Research on Animal Diseases: Trypanosomiasis,
International Laboratory for Research on Animal Diseases Reports, Nairobi, pp: 21-29.

34
Jamal, S.A. (2005): The susceptibility of Trypanosoma congolense isolated in Zambezia
Province (Mozambique) to isometamidium chloride, homidium chloride and diminazene
aceturate. MSc. Thesis, University of Pretoria, pp 67.
Jordan, A.M. (1986): Trypanosomiasis control and African rural development. Longman INC.,
New York, USA, pp 44-45.
Joshua, R.A., Obwolo, M.J., Bwangamoi, O., Mandebvu, E. (1995): Resistance to
diminazineaceturate by Trypanosoma congolense from cattle in the Zambezi Valley of
Zimbabwe. Vet. Parasitol, 60: 1-6.
Kaminsky R, Schmid C, Lun ZR. (1997): Susceptibility of dyskinetoplastic Trypanosoma evansi
and T. equiperdum to isometamidium chloride.Parasitology Research, 83: 816–818.
Kaminsky, R., Gumm, I.D., Zweygarth, E., Chuma, F. (1990): A drug incubation infectivity test
(DIIT) for assessing resistance in trypanosomes. Vet. Parasitol, 34: 335-343.
Kappmeier, G.K., Potgieter, F.T., Vreysen, M.J.B. (2007): A strategy for area-wide control
campaign with an SIT component to establish a tsetse-free South Africa. In: Verysen,
M.J.B., Hendrichs, J and Robinson, A.S (eds.), Area-wide control of insect pests: From
Research to Field Implementation, Springer, Dordrecht, The Netherlands. IAEA pp, 309-
323.
Kaufmann J., Dwinger R.H., Hallebeek A., van Dijk B., Pfister, K. (1992): The interaction of
Trypanosomacongolense and Haemonchuscontortus infections in trypanotolerantN'Dama
cattle. Vet. Parasitol, 43: 157-170.
Knoppe, N.K., Bauer, B., McDermott, J.J., Peregrine, A.S., Mehlitz, D., Clausen, P.-H. (2006):
Isometamidium sensitivity of Trypanosoma congolense stocks from cattle in West Africa
tested in mice and the drug incubation infectivity test. Acta Trop, 97: 108- 116.
Leak, S.G.A. (1999): Tsetse Biology and Ecology: Their role in the Epidemiology and control of
trypanosomosis. CABI publishing, Wallingford, UK, pp. 568.
Leeflang, P., (1978): Bovine Trypanosomiasis in Northern Nigeria; A contributions to the
epidemiology, host specificity and drug –sensitivity of Trypanosomavivax . PhD thesis
.University of Utrecht, The Netherlands, pp:.388-396.
Luckins, A.G., Mehlitz, D. (1978): Evaluation of an indirect fluorescent antibody test (IFAT),
enzyme linked immunosorbent assay (ELISA) and quantification of bovine
immunoglobulins in the diagnosis of bovine trypanosomiasis. Trop. Anim. Health Prod,
10: 149-159.

35
Makumi, J.N., Stevenson, P., Green, C.H. (2000): Control of Glossina longipennis (Diptera:
Glossinidae) by insecticide-treated targets at Galana ranch, Kenya, and confirmation of
the role of G. longipennis as a vector of cattle trypanosomiasis. Bull. Entomol. Res, 90:
397-406.
Mamoudou, A., Delespaux, V., Chepnda, V., Hachimou, Z., Andrikaye, J.P., Zoli, A., Geerts, S.
(2008): Assessment of the occurrence of trypanocidal drug resistance in trypanosomes of
naturally infected cattle in the Adamaoua region of Cameroon using the standard mouse
test and molecular tools. Acta Trop, 106 (2): 115-118.
Maser, P., Luscher, A., Kaminsky, R. (2003): Drug transport and drug resistance in African
trypanosomes. Drug Resist.Updat, 6: 281-290.
Matovu E, Stewart ML, Geiser F, Brun R, Mäser P, Wallace LJM, Burchmore RJ, Enyaru JCK,
Barrett MP, Kaminsky R, Seebeck T, De Koning HP. (2003): Mechanisms of arsenical
and diamidine uptake and resistance in Trypanosomabrucei. Eukaryotic Cell, 2: 1003-
1008.
Maudlin, I., Holmes, P., and Miles, M. (2004): Trypanosomes. In: Rovina, K. (ed.): The
trypanosomiasis. UK. CABI International Wallingford, Pp. 1-25.
McDermott, J., Woitag, T., Sidibe, I., Bauer, B., Diarra, B., Ouedraogo, D., Kamuanga, M.,
Peregrine, A., Eisler, M., Zessin.K.-H., Mehlitz, D., Clausen, P.-H. (2003): Field studies
of drug-resistant cattle trypanosomes in Kenedougou Province, Burkina Faso. Acta Trop,
86: 93-103.
Mdachi, R.E. (1999): Epidemiological studies into the impact of trypanocidal drug resisatnce on
the control of trypanosomiasis in coastal Kenya. Ph.D Thesis, University of Glasgow.pp,
352.
Moloo, S.K., Kutuza, S.B. (1984): Vectorial capacity of gamma-irradiated sterile male Glossina
morsitans centralis, G. austeni and G. tachinoides for pathogenic Trypanosoma species.
Insect Sci. Appl, 5: 411-414.
Morrison, L., Marcello, L., and McCulloch, R. (2009): Antigenic variation in the African
trypanosome: Molecular mechanisms and phenotypic complexity. Cell Microbiol,
11(12): 1724-1734.
Murray, M., Morrison, W. I., Whitelaw, D.D. (1982): Host susceptibility to African
trypanosomiasis: trypanotolerance. Advanc.Parasitol, 21: 1-68.

36
Murray, M., Murray P.K., McIntyre, W.I.M. (1977): An improved parasitological technique for
the diagnosis of African trypanosomiasis. Trans. Royal. S. Trop. Med. Hyg. 71: 325-326.
Naessens, J. (2006): Bovine trypanotolerance: a natural ability to prevent severe anaemia and
haemophagocytic syndrome. Int J Parasitol, 36: 521–528.
Njogu, A.R., Dolan, R.B., Wilson, A.J., Sayer, P.D. (1985): Trypanotolerance in East African
Oromiyaa Boran cattle. Vet. Rec, 117: 632-636.
Pallavi R, Roy N, Nageshan RK, Talukdar P, Pavithra SR, Reddy R, Venketesh S, Kumar R,
Gupta AK, Singh RK, YadavSCh, Tatu U. (2010): Heat Shock Protein 90 as a Drug
Target against Protozoan Infections: biochemical characterization of HSP90 from
Plasmodium falciparum and Trypanosoma evansi and evaluation of its inhibitor as a
candidate drug. Journal of Biological Chemistry, 285: 37964-37975.
Pan, W., Ogunremi, O., Wei, G., Shi, M., and Tabel, H. (2006): CR3 (CD11b/CD18) is the
major macrophage receptor for IgM antibody-mediated phagocytosis of African
trypanosomes: diverse effect on subsequent synthesis of tumor necrosis factor alpha and
nitric oxide. Microbes Infect, 8: 1209-1218.
Paris, J., Murray, M., McOdimba, F. (1982): A comparative evaluation of the parasitological
techniques currently available for the diagnosis of African trypanosomosis in cattle. Acta
Trop, 39: 307-316.
Patterson, J. S., Schofield. C. J. (2005): Preliminary study of wing morphometry in relation to
tsetse population genetics. S. Afr. J. Sci. 101: 1-3.
Peregrine A.S. (1994): Chemotherapy and delivery systems: haemo-parasites. Vet. Parasitol, 54:
223-248.
Peregrine, A.S., Knowles, G., Ibitayo, A.I., Scott, J.R., Moloo, S.K., Murphy, N.B. (1991):
Variation in resistance to isometamidium chloride and diminazene aceturate by clones
derived from a stock of Trypanosoma congolense. Parasitol, 102: 93-100.
Pinder, M., Authie, E. (1984): The appearance of isometamidium resistant Trypanosoma
congolense in West Africa. Acta Trop, 41: 247-252.
Radostits, O.M., Gay, C.C., Cliff, K.W., Hinch, P.H and P.D. Constable, (2007). Veterinary
medicine: Textbook of the disease of cattle, sheep, goat and horses. ed. London:
Sounders, Pp. 1534-1537.
Reichard, R.E. (2002): Area-wide biological control of disease vectors and agents affecting
wildlife. Rev. Sci. Tech. Off. Int. Epiz. 21 (1): 179-185.

37
Rowlands, G.J., W. Mulatu, E. Authie, G.D. D'Ieteren, S.G.A. Leak and S.M. Nagda. (1994):
Effect of Trypanosomiasis on growth and mortality of young East African zebu’s cattle
Schweingruber AE.(2004): The melaminophenyl arsenicals melarsoprol and melarsen oxide
interfere with thiamine metabolism in the fission yeast Schizosaccharo mycespombe.
Antimicrobial Agents and Chemotherapy, 48: 3268-3271.
Shakoor, O., R.B. Taylor and R.H. Behrens, (1997): Assessment of the incidence substandard
drug in developing Countries. Tropical Medicine and International Health, 2: 839- 845.
Shapiro, TA., and Englund, PT. (1990): Selective cleavage of kinetoplast DNA minicircles
promoted by anti-trypanosomal drugs. Proceedings of the National Academy of Sciences
USA, 87: 950-954.
Shinyangwe, L., Delespaux, V., Brandt, J., Geerts, S., Mubanga, J., Machila, N., Holmes, P.H.,
Eisler., M.C. (2004): Trypanocidal drug resistance in eastern province of Zambia. Vet.
Parasitol, 119: 125-135.
Simarro, P., Jannin, J., and Cattand, P. (2008): Eliminating human African trypanosomiasis:
Where do we stand and what comes next? PLoS Med, 5 (2):174-180.
Soeiro MNC, De Souza EM, Stephens CE, Boykin DW. (2005): Aromatic diamidines as
antiparasitic agents. Expert Opinion on Investigational Drugs, 14: 957-972.
Solano, P., Kone, A., Garcia, A., Sane, B., Michel, V., Michel, J., Coulibaly, B., Jamonneau, V.,
Kaba, D., Dupont, S., and Fournet, F. (2003): Role of patient travel in transmission of
human African trypanosomiasis in a highly endemic area of the Ivory Coast. Med Trop
(Mars), 63: 577-582.
Solano, P., Ravel, S., de Meeus, T. (2010): How can tsetse population genetics contribute to
African trypanosomiasis control? Trends Parasitol, 26(5): 255-263.
Soltys, M., and Woop, T. (1997): Trypanosomes producing disease in livestock in Africa. In:
Kreier, J. (ed.): Parasitic Protozoa. Vol. 1. New York: Academic Press, Pp 27-36.
Sones, K.R., Holmes P.H., Urquhart, G. M. (1989).Interference between drug-resistant and drug-
sensitive stocks of Trypanosoma congolense in goats. Res. Vet. Sci. 47: 75–77. Sones, K.
(2001): Pharmarceutical companies: partners or enemies? I.C.P.T.V, 3: 19-21.
Sones, K.R., Njogu, A.R., Holmes, P.H. (1988): Assessment of sensitivity of T. congolense to
ismetamidium chloride: a comparison of tests using cattle and mice. Acta Trop, 45: 153-
164.

38
Sutherland, I.A., Mounsey, A., Holmes, P.H. (1991): Effect of isometamidium on Trypanosoma
congolense infectivity. Vet. Parasitol, 39: 13-17.
Taylor, K., and Authie, E. (2004): Pathogenesis of Animal trypanosomiasis. In:Maulidn, I.,
Holmes, P., and Miles M. (ed.): The Trypanosomiases. UK: CABI International
Wallingford. Pp, 331-353.
Tewelde, N., Abebe, G., Eisler, M., McDermot, J., Greiner, M., Afework, Y., Kyule, M.,
Munstermann, S., Zessin, K-H., Clausen, P-H. (2004): Application of field methods to
assess isometamidium resistance of trypanosomes in cattle in Western Ethiopia. Acta
Trop, 90: 163-170.
Torr, S.J., Eisler, M., Coleman, P., Morton, H., Machila, N. (2002): Integrated control of ticks
and tsetse. A report for the Department for International Development Advisory and
support services Contract, managed by NR International Limited – Project ZV, 01510.
Torr, S.J., Holloway, M.T.P., Vale, G.A. (1992): Improved persistence of insecticide deposits on
targets for controlling Glossina pallidipes (Diptera: Glossinadae). Bull. Entomol. Res, 82:
525-533.
Torr, S.J., Mangwiro, T.N.C. (2000): Interactions between cattle and biting flies: effects on the
feeding rate of tsetse. Med. Vet. Entomol, 14: 400-409.
Torr, S.J., Wilson, P.J., Schonfield, S., Mangwiro, T.N.C., Aber, S., White, B.N. (2001):
Application of DNA markers to identify the individual-specific host of tsetse feeding on
cattle. Med. Vet. Entomol, 15: 78-86.
Uilenberg, G., (1998): A Field Guide for Diagnosis,Treatment and Prevention of African Animal
Trypanasomosis. Adopted from the original edition of boyt. W. P. FAO, Rome. Pp, 43-
135.
Vale, G.A., Grant, I.F. (2002): Modelled impact of insecticide-contaminated dung on the
abundance and distribution of dung fauna. Bull. Entomol. Res, 92: 251-263.
Vickerman, K. (1985): Developmental cycles and biology of pathogenic trypanosomes. Br Med
Bull, 41: 105-14.
Vreysen, M.J.B. (2001): Principles of area-wide integrated tsetse fly control using the sterile
insect technique. Med. Trop, 61: 397-411.
Vreysen, M.J.B. (2005): Monitoring sterile and wild insects in area-wide integrated pest
management programmemes. A. S. pp, 325-361.

39
Whiteside, E.F. (1962): Interactions between drugs, trypanosomes and cattle in the field. In :
Goodwin, L.G and Nimmo-Smith, R.H (eds) Drugs, Parasites and Hosts. J and A.
Churchill, London, pp, 116-141.
Wilkes, J.M., Mulugeta, W., Wells, C., Peregrine, A.S. (1997): Modulation of mitochondrial
electrical potential: a candidate mechanism for drug resistance in African trypanosomes.
Biochem. J, 326(3): 755–761.





























