Embed Size (px)
Citation preview

THE JOURNAL OF GENE MEDICINE R E S E A R C H A R T I C L EJ Gene Med 2008; 10: 1019–1031.Published online 25 July 2008 in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/jgm.1222
Tumor targeting of baculovirus displaying alymphatic homing peptide
Anna R. Makela1*Juulia Enback2
Johanna P. Laakkonen1
Maija Vihinen-Ranta1
Pirjo Laakkonen2
Christian Oker-Blom1
1NanoScience Center, Department ofBiological and Environmental Science,University of Jyvaskyla, Finland2Molecular Cancer Biology Programand Institute of Biomedicine,Biomedicum Helsinki, University ofHelsinki, Finland
*Correspondence to:Anna R. Makela, NanoScienceCenter, Department of Biologicaland Environmental Science, PO Box35, FIN-40014 University ofJyvaskyla, Finland.E-mail: [email protected]
Received: 28 February 2008Revised: 19 April 2008Accepted: 8 May 2008
Abstract
Background Tumor-associated cells and vasculature express attractivemolecular markers for site-specific vector targeting. To attain tumor-selectivetropism, we recently developed a baculovirus vector displaying the lymphatichoming peptide LyP-1, originally identified by ex vivo/in vivo screening ofphage display libraries, on the viral envelope by fusion to the transmembraneanchor of vesicular stomatitis virus G-protein.
Methods In the present study, we explored the specificity and kinetics ofviral binding and internalization as well as in vivo tumor homing of the LyP-1 displaying virus to elucidate the applicability of baculovirus for targetedtherapies.
Results We demonstrated that the LyP-1 peptide contributes to saturablebinding of baculovirus in human MDA-MB-435 and HepG2 carcinoma cellsand escalates the kinetics of viral internalization leading to earlier nuclearaccumulation and enhanced transgene expression. The LyP-1 displayingvirus also showed stronger competitiveness against transduction with wild-type baculovirus, suggesting involvement of a specific receptor in cellularattachment and entry. Following intravenous injections, the modified virusaccumulated within the human MDA-MB-435 and MDA-MB-231 carcinomaxenografts in mice with higher specificity and efficiency than the control virus.Targeting of the modified virus was more specific in the MDA-MB-435 than inthe MDA-MB-231 xenografts as demonstrated by higher tumor accumulationand lower distribution in nontarget organs. No apparent cytotoxicity wasassociated with the surface modification.
Conclusions This first demonstration of in vivo tumor targeting of asystemically administered, tropism-modified baculoviral vector highlights thepotential of baculovirus-mediated targeted therapies. Copyright 2008 JohnWiley & Sons, Ltd.
Keywords baculovirus; display; peptide; targeting; gene therapy
Introduction
Characterization of the molecular diversity at the surface of disease-associatedcells and identification of specific ligands for these markers provide a basisfor the development of targeted therapies. Recently, in vivo targeting oftherapeutic entities to specific cells by vector engineering has become amajor focus of cancer therapy research. Many of these vectors are modi-fied viruses with unique benefits and shortcomings. Baculovirus, an insect
Copyright 2008 John Wiley & Sons, Ltd.

1020 A. R. Makela et al.
pathogen, represents a multifunctional, biotechnologicaltool with the potential for biomedical applications includ-ing gene therapy and vaccination [1]. An exceptionaladvantage of baculovirus is its molecular flexibility, allow-ing modification of the vector phenotype for tissue andcell targeting as well as for resistance to innate immu-nity [2]. Moreover, the virus is regarded as safe withnegligible pathogenic potential in humans, has a largetransgene insertion capacity, induces no cytotoxic effectseven at high virus concentrations, is able to mediatetransient and stable transgene expression, and facilitateslarge-scale production to high vector titers for clinicalapplications.
Previous studies highlight the complexity of baculovir-us-mammalian cell interactions both in vitro and in vivo.A number of dividing and nondividing primary as wellas established cell lines are susceptible to baculovirustransduction in culture [3,4]. Furthermore, gene deliv-ery to diverse tissues in vivo has been conducted withnative or pseudotyped viruses following administrationvia diverse routes [5–19]. The precise endocytic mech-anism, pathway, and participating molecules enablingbaculovirus binding and entry to mammalian targetcells are incompletely characterized. The virus has beensuggested to employ ubiquitous cell surface molecules,heparan sulphate proteoglycans or phospholipids, forcellular attachment [20,21], and clathrin-mediated endo-cytosis together with macropinocytosis for internalization[22,23]. We are currently investigating the preference,circumstances, and the relative extent to which these, oralternative, pathways are exploited (J. P. Laakkonen et al.,manuscript submitted). Additionally, recent studies havefocused on elucidating the general behaviour and biodis-tribution of baculovirus in vivo to better comprehend thekinetics of viral administration and to optimize the viraldose [5,9,16,18,19]. Understanding these mechanisms indetail is crucial for improving the performance of currentbaculoviral vectors.
To restrict vector homing and transgene expression onlyto the cells of interest, effort has been devoted to improvethe specificity of baculoviral vectors via incorporationof specific targeting mechanisms, both transductional[24–29] and transcriptional [14,30]. Despite the absenceof pre-existing anti-baculovirus humoral immunity inhumans [31], the usage of baculovirus for systemicgene delivery in vivo is compromised by the activationof innate immune responses, the major barrier beingviral inactivation by the complement system. Partialprotection from the complement has been accomplishedin vitro by applying various inhibitory agents [5,15,32,33]and in vivo using a baculovirus displaying human decay-accelerating factor on the viral envelope [34]. Therefore,the development of targeted, innate-immunity resistantbaculovirus vectors is necessary to overcome theselimitations, permitting the full potential of baculoviralvectors to be realized.
Tumor vasculature has received increased attentionas a target for potential anti-cancer therapies. Severalpeptides that recognize tumor-specific vascular signatures
and selectively target to tumor blood vessels, lymphaticvessels and/or tumor cells have been identified byin vivo biopanning of phage display libraries [35,36].The LyP-1 is a cyclic, nine amino-acid peptide originallyisolated by ex vivo/in vivo screening of phage displaylibraries using human MDA-MB-435 carcinoma xenografts[37]. The peptide specifically homes to tumor-associatedlymphatic vessels, is internalized by certain tumor cellsand tumor lymphatic endothelial cells, and is capableof delivering therapeutic agent-like payloads such as T7-phage, fluorescein and quantum dots to the target cellsin vivo [37,38].
Via envelope display of a battery of tumor-homingpeptides with internalizing properties, namely LyP-1, F3,CGKRK and RGD, we have previously shown that it ispossible to alter both the efficiency and the specificity ofbaculovirus-mediated gene delivery in vitro [24,25]. Inthe view of these recent data, the evidence of sequence-specific as opposed to charge-mediated enhancement ofviral binding was the strongest for the LyP-1 peptide[24]. In the present study, we further investigated thespecificity and kinetics of viral binding and internalizationas well as in vivo tumor homing of the LyP-1 displayingvirus to elucidate the applicability of baculovirus fortargeted therapies. To the best of our knowledge, thisis the first demonstration of in vivo tumor targetingusing an intravenously administered, tropism-modifiedbaculovirus vector.
Materials and methods
Baculoviruses
The wild-type baculovirus utilized was Autographacalifornica nucleopolyhedrovirus (AcMNPV) strain E2.Construction of the recombinant viruses Ac-luc, AcLyP-1-luc, AcF3-luc, AcCGKRK-luc and AcCL-luc was describedpreviously [24,25]. Ac-EGFP and AcLyP-1-EGFP wereconstructed as described for Ac-luc and AcLyP-1-luc [24]with the exception of cloning the 1329 bp sequenceof cytomegalovirus promoter/enhancer and enhancedgreen fluorescent protein (EGFP) coding region underthe P10 promoter of pFastBac Dual (Invitrogen, Carlsbad,CA, USA) following amplification by polymerase chainreaction with 5′-AA AAA GGT ACC TAG TTA TTA ATAGTA ATC AAT TAC GGG G-3′ forward and 5′-ATA CGAGGT ACC TTA CTT GTA CAG CTC GTC CAT GCC GAG-3′ reverse primers containing KpnI-restriction sites usingpEGFP-C1 (Clontech, BD Biosciences, Mountain View,CA, USA) as a template. Viral titers were determinedby end-point dilution assay either from unconcentrated,concentrated in phosphate-buffered saline (PBS) (7000 gfor 7 h at 4 ◦C) or sucrose gradient purified baculovirusstocks using standard protocols.
Cell lines and tumors
Spodoptera frugiperda (Sf9; ATCC CRL-1711), humanMDA-MB-435 melanoma/breast carcinoma and human
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm

Tumor targeting of baculovirus 1021
HepG2 hepatocarcinoma (ATCC HB-8065) cell lineswere cultured as described previously [24]. HumanMDA-MB-231 breast carcinoma cells (ATCC HTB-26)were maintained in RPMI 1640 supplemented with10% inactivated fetal bovine serum, 2 mM L-glutamine,1% penicillin-streptomycin (all from Invitrogen) in ahumidified 5% CO2 atmosphere at 37 ◦C. Athymic, 4–6-week-old female BALB/c nu/nu mice from TaconicEurope were maintained under SPF-conditions in atemperature and humidity controlled environment. Miceunder anesthesia were injected with 2.5 × 106 tumor cellsinto the mammary fat pad to induce tumors and usedfor in vivo targeting experiments at 8–12 weeks post-injection. All animal studies were conducted accordingto the guidelines of the Provincial Government ofSouthern Finland and the protocol was approved bythe Experimental Animal Committee of the Universityof Helsinki.
Binding assays
Equilibrium binding of each virus to MDA-MB-435 andHepG2 cells was titrated by incubating cells with increas-ing concentrations of virus for 2 h at 4 ◦C in suspensionto reach equilibrium. All the steps throughout the exper-iment were performed at 4 ◦C. Unbound virus particleswere removed by extensive washes with 1.5% bovineserum albumin (BSA)-PBS. Bound virus was detectedby incubation with mouse monoclonal GP64 B12D5antibody for 1 h followed by washes with 1.5% BSA-PBS and incubation with Alexa Fluor488-conjugatedsecondary antibody (Molecular Probes, Eugene, OR,USA) for 30 min. Fluorescence of cells was anal-ysed with a FACSCalibur flow cytometer (BecktonDickinson, Heidelberg, Germany) using CellQuest soft-ware.
Transduction experiments
MDA-MB-435 and HepG2 cells were plated and allowedto grow to subconfluency for 1–2 days at 37 ◦C. Viraltransduction experiments were generally performed for24 h at 37 ◦C using 200 plaque forming units (PFU)/cellof baculovirus. Cells were harvested and monitored forluciferase activity as described previously [24] or for EGFPexpression by flow cytometry.
Competitive binding/transduction assays were per-formed on HepG2 cells for 2 h in complete growthmedium at 37 ◦C followed by thorough washes and furtherincubation up to 24 h p.t. at 37 ◦C. The virus dilutions, aconstant amount (200 PFU/cell) of a recombinant virusand increasing concentrations (0, 100, 200, 500, 1000,1500 and 2000 PFU/cell) of the competitor wild-type bac-ulovirus, were prepared in complete growth medium priorto addition to the cells.
Blocking of endocytosis was performed by addingammonium chloride onto cells at different time intervals
of synchronized viral binding and transduction. Viruspreparations were diluted with ice-cold medium andallowed to adsorb to HepG2 cells for 1 h at 4 ◦C. Thevirus-containing medium was replaced with warm (37 ◦C)medium followed by addition of ammonium chlorideto a final concentration of 10 mM at fixed times. Toexclude the possibility that ammonium chloride quenchesthe EGFP fluorescence, 10 mM ammonium chloride wasadded to the transduced control cells 1 h (37 ◦C) prior toharvesting.
Viral internalization studies and immunofluorescencelabeling. For all binding and internalization experiments,MDA-MB-435 and HepG2 cells were allowed to attachto glass cover slips and grown to subconfluency for1–2 days at 37 ◦C. Viruses (200 PFU/cell in PBS) wereallowed to bind to cells for 1 h at 4 ◦C to avoid virusinternalization. Unbound virus was removed by washeswith PBS at 4 ◦C followed by incubation at 37 ◦C incomplete growth medium for set times and fixation with3% or 4% paraformaldehyde in PBS for 15 min or 20 minat room temperature (RT), respectively. Viruses werelabeled with mouse monoclonal VP39 antibody or rabbitpolyclonal AcMNPV [39] primary antibody.
For quantification of virus internalization kinetics, theviruses were labeled without permeabilization with rabbitpolyclonal AcMNPV primary antibody [39] for 1 h andwith goat anti-rabbit Alexa Fluor546 conjugate (MolecularProbes) for 30 min at RT. Subsequent to washes with1.5% BSA-PBS and permeabilization with 0.1% TritonX-100 in PBS, the virus was similarly labeled with rabbitpolyclonal AcMNPV primary antibody and goat anti-rabbit Alexa Fluor488 conjugate. Thus, cell membrane-associated virus was labeled with both Alexa Fluor546 andAlexa Fluor488 conjugates and was observed as yellow byconfocal microscopy when the red and green channelswere merged, and internalized/cytoplasmic virus wasobserved as green signal. The cover slips were mountedon microscope slides in Mowiol (Calbiochem, Darmstadt,Germany) supplemented with DABCO (25 mg/ml; Sigma-Aldrich, Saint Louis, MO, USA).
For cellular localization studies with endosomalmarkers, baculovirus-transduced cells were double-immunolabeled with rabbit polyclonal AcMNPV andmouse monoclonal lysosome-associated membrane pro-tein 2 (LAMP-2) antibodies (Developmental StudiesHybridoma Bank, Iowa City, IA, USA) for 1 h at RT.Primary antibodies were visualized with goat anti-rabbit Alexa Fluor488 and anti-mouse Alexa Fluor546conjugates. Cells were embedded in ProLong Goldantifade reagent supplemented with 4′,6′-diamidino-2-phenylindole hydrochloride (DAPI) (Molecular Probes).
Confocal microscopy and imageanalysis
Images were acquired with a laser scanning confo-cal microscope (LSM510, Zeiss Axiovert 100M, Jena,
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm

1022 A. R. Makela et al.
Germany; or Olympus FluoView 1000, Olympus Opti-cal Co., Tokyo, Japan) using appropriate excitationand emission settings together with the multitrackingmode. The pixel resolution was adjusted to 100 nm/pixel(512 × 512 pixels/image). The images shown are singleconfocal optical slices of z-stacks approximately 0.8 µmin thickness and were linearly adjusted in brightness andcontrast to the entire field.
For quantification of viral internalization kinetics,25 images (z-stacks) representing a minimum of 40cells were analysed for each virus and time pointwith ImageJ software (NIH Image, Bethesda MD,USA) using the Colocalization Threshold plug-in (TonyCollins, Wright Cell Imaging Facility, WCIF, Toronto,Canada), which is based on the Costes ColocalizationQuantification algorithm [40]. Percentage values ofcolocalized fluorescence intensity above threshold foreach channel were used to determine the relative quantityof internalized virus. Colocalization of the red signal withthe green signal remained constant (approximately 100%)during the time course of viral transduction whereas, dueto virus internalization, the colocalization of the greensignal with the red signal decreased as a function of time.For quantification of viral nuclear-accumulation, imagesfrom 25–35 cells were collected and analysed by WCIFImageJ. The relative standard deviations of the nuclearfluorescence intensities were calculated.
Cytotoxicity assay
Cytotoxicity of the recombinant baculoviruses wasdetermined by CellTiter 96 Aqueous One Solution CellProliferation Assay (MTT assay; Promega, Madison,WI, USA) according to the manufacturer’s instructions.The measurements were performed with a multilabelcounter Wallac 1420 Victor2 (Wallac Oy, Turku, Finland)at an absorbance of 492 nm using software version2.00. Viability of the baculovirus transduced cells wasdetermined by comparing the absorbance values to thoseof mock-transduced control cells set to 100%.
In vivo targeting assays
As a preclinical model, nude mice bearing tumorxenografts derived from MDA-MB-435 or MDA-MB-231cells were used. Viruses (5 × 108 PFU) were intravenouslyinjected via the tail vein and allowed to circulate for24 h. The mice were perfused through the heart with10 ml of PBS followed by 10 ml of 4% paraformaldehyde(PFA) in PBS. Tumors and organs were removed, PFA-fixed, and soaked in 30% sucrose (w/v) overnight.Tissues were frozen in OCT embedding medium (Tissue-Tek, Sakura Finetek USA, Torrance, CA, USA). Forhistological analyses, 5–10 µm sections were cut. Todetect the biodistribution of the baculovirus, tumors andorgans were labeled using a rabbit polyclonal AcMNPVantibody [39] and goat anti-rabbit Alexa Fluor594
conjugate (Molecular Probes). Nuclei were counterstainedwith DAPI (Vector Laboratories, Burlingame, CA, USA).Blood vessels were visualized by staining tissue sectionswith monoclonal antibodies against CD31 (rat anti-mouse or Syrian hamster anti-mouse antibodies fromBD Biosciences Pharmingen, San Diego, CA, USA).Goat anti-mouse VEGFR-3 (R&D Systems, Minneapolis,MN, USA) was used to stain VEGFR-3 in the tumorsections. A polyclonal rabbit anti-mouse LYVE-1 antibodywas used to visualize the lymphatic vessels. Theprimary antibodies were detected with the appropriateAlexa Fluor488 or 594-conjugated secondary antibodies(Molecular Probes). The sections were examined underan inverted fluorescence microscope (Zeiss Axioplan 2).The intensity of baculovirus-specific fluorescence from thedissected tumors was analysed with ImageJ software.
Statistical analysis
The data were analysed using the unpaired Student’st-test with a two-tailed p-value, and statistical signifi-cance was determined relative to the positive controlsamples (∗∗p < 0.05 and ∗∗∗p < 0.001). The mean flu-orescence/luminescence and expression-% values werenormalized relative to the positive control samples.
Results
LyP-1 peptide contributes to saturablebinding of baculovirus
The LyP-1 lymphatic homing peptide was displayed onthe baculovirus envelope by fusion the transmembrane-cytoplasmic domains of vesicular stomatitis virus G-protein (VSVG) through a polyalanine linker [24]. TheLyP-1-displaying recombinant viruses, namely AcLyP-1-luc [24] and AcLyP-1-EGFP, were further equipped withexpression cassettes encoding either firefly luciferase (luc)or EGFP, respectively, enabling detection of transgeneexpression in both insect and mammalian cells. Thesuccessful incorporation and the functionality of the LyP-1-VSVG fusion protein in virus particles was previouslyverified by western blotting and binding/transductionstudies [24]. The baculoviruses Ac-luc [24] and Ac-EGFPwith wild-type envelope phenotypes served as controls.Importantly, the ratio of total particle number versusthe amount of infectious virus particles was similarwith the control and modified viruses as determined byimmunoblotting of equal PFUs with VP39 [24] and GP64antibodies (data not shown).
Cellular binding of unmodified baculovirus is non-saturable, of low affinity, and mediated by ubiquitouscell surface molecules, heparan sulphate proteoglycansand/or phospholipids [20,21]. To determine whetherthe LyP-1 peptide could direct the surface-modified bac-ulovirus, AcLyP-1-luc, to a specific, saturable receptor dur-ing the cellular attachment, the equilibrium binding was
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm

Tumor targeting of baculovirus 1023
determined in human MDA-MB-435 and HepG2 cells byflow cytometry using progressively increasing virus con-centrations. To prevent the endocytic entry of baculovirusduring binding [23,41,42], the cells were exposed tolow temperature (4 ◦C), instead of the physiological tem-perature (37 ◦C) and sodium azide that, being an ionicmetabolic inhibitor, might interfere with the electrostaticinteractions between the virus and the target cells [20].In general, viral binding increased in a concentration-dependent fashion. In both cell lines, binding of thecontrol virus, Ac-luc [24], possessing the wild-type phe-notype displayed the highest efficiency at approximately5000 PFU/cell under these conditions (Figure 1A) and lin-early decreased thereafter (data not shown). The bindingof AcLyP-1-luc, however, reached the highest efficiencywith significantly lower virus concentrations, in the range750–1000 PFU/cell, and remained essentially constantthereafter (Figure 1A). Independent of the viral concen-tration, binding of the AcLyP-1-luc to both MDA-MB-435and HepG2 cell lines was constantly more efficient com-pared to the Ac-luc, and no significant difference in viralbinding between the cell lines was observed (Figure 1A).It is noteworthy, however, that the obtained binding effi-ciencies are regulated in part by the assay temperature(4 ◦C) and the absolute saturation levels of viral bindinglikely are lower at 37 ◦C [20]. In addition to saturabilityand higher efficiency, the pattern of AcLyP-1-luc binding to
target cells clearly exhibited a more scattered and uniformappearance whereas the Ac-luc formed large aggregatesat the cell surface (Figure 1B). Despite higher efficiency[24], the binding of baculoviruses displaying the F3 andCGKRK tumor-homing peptides by fusion to the trans-membrane anchor of VSVG [24] was similar to that ofAc-luc with respect to saturability (data not shown).
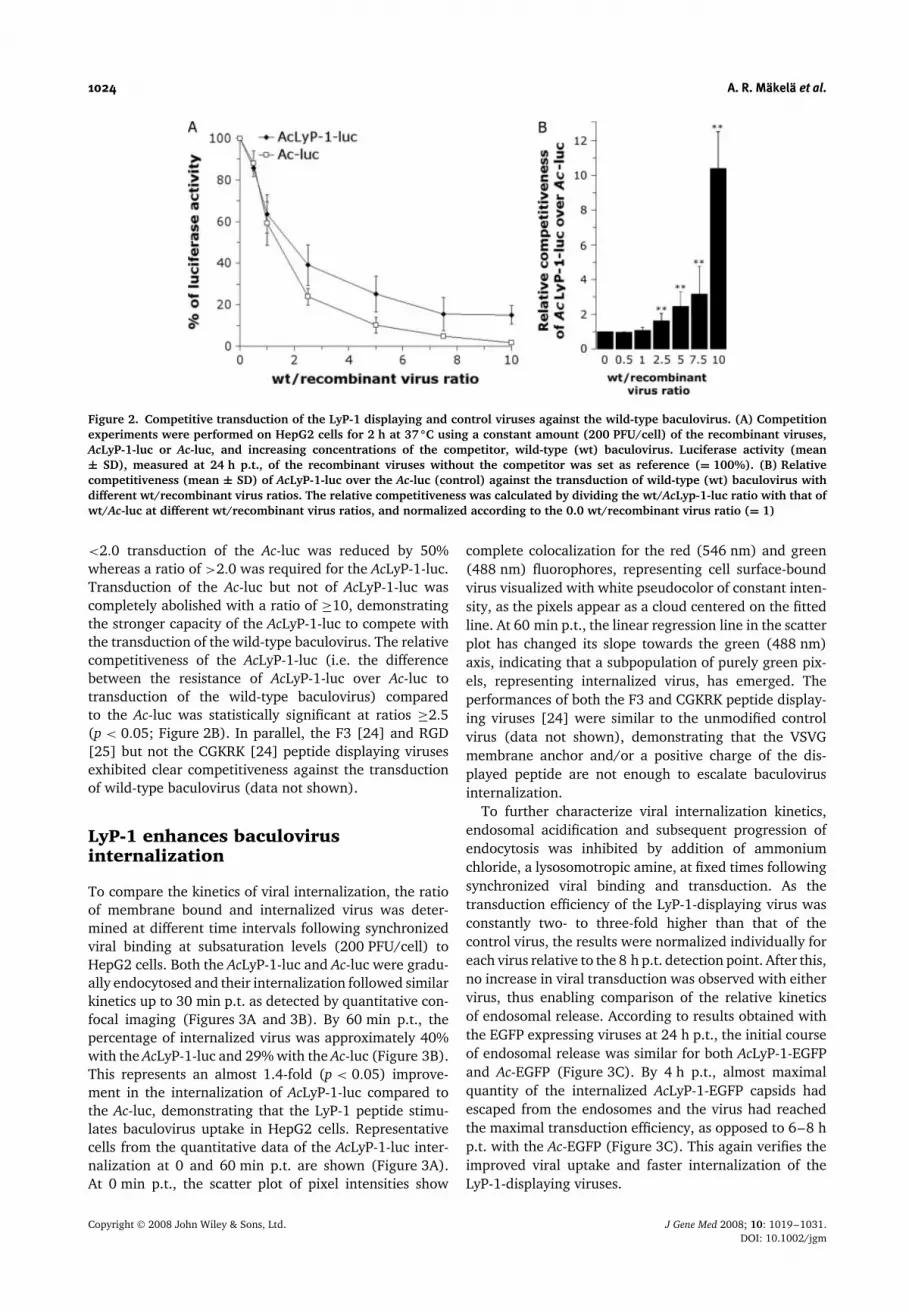
LyP-1 displaying baculovirus exhibitsstronger competitiveness againsttransduction with wild-type virus
We previously showed in competition studies thatthe soluble LyP-1 peptide is able to inhibit both thebinding and transduction of the AcLyP-1-luc with ahigh efficacy and an apparent specificity [24]. Tofurther compare the specificity and efficiency of viralbinding and internalization, competitive transductionswere performed for 2 h on HepG2 cells using a constantquantity of the AcLyP-1-luc or Ac-luc and increasingconcentrations of the competitor, a wild-type baculovirus.The transduction efficiency was measured by luciferaseactivity at 24 h p.t. The control virus Ac-luc wasdisplaced more rapidly from the target cells than AcLyP-1-luc by increasing the amount of the competitor virus(Figure 2A). With a wild-type/recombinant virus ratio of
Figure 1. Equilibrium and pattern of viral binding. (A) The equilibrium of AcLyP-1-luc and Ac-luc binding was titrated on theMDA-MB-435 and HepG2 cells at 4 ◦C using increasing concentrations of each virus. Bound viruses were detected with the mousemonoclonal GP64 primary and Alexa Fluor488 secondary antibodies using flow cytometry. Fluorescence intensities (mean ± SD)are shown. (B) Confocal microscopic analysis of the AcLyP-1-luc and Ac-luc (200 PFU/cell) binding to MDA-MB-435 and HepG2 cellsat 4 ◦C. Viruses were labeled with the mouse monoclonal VP39 primary and Alexa Fluor488 secondary antibodies. Pseudocoloredfluorescence images with intensity increasing from blue to white are shown to visualize the bound virus particles and differentialinterference contrast (DIC) images to outline cellular boundaries. Scale bar = 10 µm
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm
1024 A. R. Makela et al.
Figure 2. Competitive transduction of the LyP-1 displaying and control viruses against the wild-type baculovirus. (A) Competitionexperiments were performed on HepG2 cells for 2 h at 37 ◦C using a constant amount (200 PFU/cell) of the recombinant viruses,AcLyP-1-luc or Ac-luc, and increasing concentrations of the competitor, wild-type (wt) baculovirus. Luciferase activity (mean± SD), measured at 24 h p.t., of the recombinant viruses without the competitor was set as reference (= 100%). (B) Relativecompetitiveness (mean ± SD) of AcLyP-1-luc over the Ac-luc (control) against the transduction of wild-type (wt) baculovirus withdifferent wt/recombinant virus ratios. The relative competitiveness was calculated by dividing the wt/AcLyp-1-luc ratio with that ofwt/Ac-luc at different wt/recombinant virus ratios, and normalized according to the 0.0 wt/recombinant virus ratio (= 1)
<2.0 transduction of the Ac-luc was reduced by 50%whereas a ratio of >2.0 was required for the AcLyP-1-luc.Transduction of the Ac-luc but not of AcLyP-1-luc wascompletely abolished with a ratio of ≥10, demonstratingthe stronger capacity of the AcLyP-1-luc to compete withthe transduction of the wild-type baculovirus. The relativecompetitiveness of the AcLyP-1-luc (i.e. the differencebetween the resistance of AcLyP-1-luc over Ac-luc totransduction of the wild-type baculovirus) comparedto the Ac-luc was statistically significant at ratios ≥2.5(p < 0.05; Figure 2B). In parallel, the F3 [24] and RGD[25] but not the CGKRK [24] peptide displaying virusesexhibited clear competitiveness against the transductionof wild-type baculovirus (data not shown).
LyP-1 enhances baculovirusinternalization
To compare the kinetics of viral internalization, the ratioof membrane bound and internalized virus was deter-mined at different time intervals following synchronizedviral binding at subsaturation levels (200 PFU/cell) toHepG2 cells. Both the AcLyP-1-luc and Ac-luc were gradu-ally endocytosed and their internalization followed similarkinetics up to 30 min p.t. as detected by quantitative con-focal imaging (Figures 3A and 3B). By 60 min p.t., thepercentage of internalized virus was approximately 40%with the AcLyP-1-luc and 29% with the Ac-luc (Figure 3B).This represents an almost 1.4-fold (p < 0.05) improve-ment in the internalization of AcLyP-1-luc compared tothe Ac-luc, demonstrating that the LyP-1 peptide stimu-lates baculovirus uptake in HepG2 cells. Representativecells from the quantitative data of the AcLyP-1-luc inter-nalization at 0 and 60 min p.t. are shown (Figure 3A).At 0 min p.t., the scatter plot of pixel intensities show
complete colocalization for the red (546 nm) and green(488 nm) fluorophores, representing cell surface-boundvirus visualized with white pseudocolor of constant inten-sity, as the pixels appear as a cloud centered on the fittedline. At 60 min p.t., the linear regression line in the scatterplot has changed its slope towards the green (488 nm)axis, indicating that a subpopulation of purely green pix-els, representing internalized virus, has emerged. Theperformances of both the F3 and CGKRK peptide display-ing viruses [24] were similar to the unmodified controlvirus (data not shown), demonstrating that the VSVGmembrane anchor and/or a positive charge of the dis-played peptide are not enough to escalate baculovirusinternalization.
To further characterize viral internalization kinetics,endosomal acidification and subsequent progression ofendocytosis was inhibited by addition of ammoniumchloride, a lysosomotropic amine, at fixed times followingsynchronized viral binding and transduction. As thetransduction efficiency of the LyP-1-displaying virus wasconstantly two- to three-fold higher than that of thecontrol virus, the results were normalized individually foreach virus relative to the 8 h p.t. detection point. After this,no increase in viral transduction was observed with eithervirus, thus enabling comparison of the relative kineticsof endosomal release. According to results obtained withthe EGFP expressing viruses at 24 h p.t., the initial courseof endosomal release was similar for both AcLyP-1-EGFPand Ac-EGFP (Figure 3C). By 4 h p.t., almost maximalquantity of the internalized AcLyP-1-EGFP capsids hadescaped from the endosomes and the virus had reachedthe maximal transduction efficiency, as opposed to 6–8 hp.t. with the Ac-EGFP (Figure 3C). This again verifies theimproved viral uptake and faster internalization of theLyP-1-displaying viruses.
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm

Tumor targeting of baculovirus 1025
Figure 3. Kinetics of viral internalization in HepG2 cells. (A, B) Following the synchronized binding of AcLyP-1-luc and Ac-luc(200 PFU/cell), the percentage of internalized virus (mean ± SD) was determined by quantitative confocal imaging at 0, 15, 30and 60 min p.t. Representative cells of the AcLyP-1-luc internalization at 0 and 60 min p.t. are shown in (A). The viruses weredetected using a rabbit polyclonal AcMNPV primary antibody. Cell membrane associated virus was labeled with both Alexa Fluor488and 546 secondary antibody conjugates and colocalized pixels are shown with white pseudocolor of constant intensity, whereasinternalized/cytoplasmic virus was labeled solely with the Alexa Fluor488 conjugate and can be detected as a green signal. Scatterplot of pixel intensities and Differential interference contrast (DIC) images are shown. The colors in the scatter plot indicate thefrequency with which particular intensity combinations are observed, with blue being the lowest and white the highest. Scalebar = 10 µm. (C) The rate of endosomal escape was investigated by adding ammonium chloride (10 mM) at the indicated timeintervals following synchronized viral binding and transduction (200 PFU/cell). At 24 h p.t., cells were analysed for the EGFPexpression. The mean fluorescence (mean ± SD) of EGFP expressing cells was determined relative to the highest expression level.The results from two independent assays, each performed in triplicate, are shown. (D) Quantitative analysis of nuclear fluorescenceintensity of viral capsids (200 PFU/cell) in HepG2 cells at 8 h p.t. by confocal microscopy. The viruses were detected using therabbit polyclonal AcMNPV primary and Alexa Fluor488 secondary antibodies. Relative fluorescence intensities (mean ± SD) areshown. (E) Viral transduction efficiency of HepG2 cells measured by luciferase activity at 8 h p.t. The results are shown as meansof the luciferase activity (relative light units, RLU) (mean ± SD) of two individual experiments each performed in triplicate pervariable
In HepG2 cells, nuclear localization of the wild-typebaculovirus capsid has been demonstrated to be the mostabundant at 8 h p.t. decreasing linearly thereafter over10–24 h p.t. [43]. The intensity of nucleus-associatedlabeling of the AcLyP-1-luc capsids in HepG2 cells at8 h p.t. was approximately half (p < 0.05; Figure 3D),whereas the transgene expression level was two-foldhigher than that of the Ac-luc (p < 0.05; Figure 3E).These results indicate an earlier nuclear accumulation,subsequent capsid dissociation and genomic release bythe AcLyP-1-luc.
LyP-1 displaying baculovirusaccumulates to tumor tissue
To evaluate tumor-homing of the AcLyP-1-luc followingsystemic administration of the virus, nude mice bear-ing tumor xenografts derived from the MDA-MB-435(Figure 4A) or MDA-MB-231 (Figure 4B) cells were usedas a preclinical model. The viruses (5 × 108 PFU) were
injected intravenously and allowed to circulate for 24 hbefore the examination. Histological analysis of dissectedtumors and organs from the treated animals revealed clearbaculovirus-specific fluorescence in the tumors from miceinjected with the AcLyP-1-luc, whereas only negligible flu-orescence was detectable in the tumors from the controlvirus-injected mice (Figure 4). No difference in the tumortargeting between the F3 or CGKRK [2] peptide display-ing viruses and the control virus was observed (data notshown). Semiquantitative fluorescence intensity analysisshowed that, compared to the control virus, the concentra-tion of AcLyP-1-luc was approximately 7.3- and 5.2-foldhigher in the MDA-MB-435 (Figure 4A) and MDA-MB-231 (Figure 4B) tumors, respectively. These results showthat the AcLyP-1-luc is targeted to these tumors withimproved efficiency and specificity. As the LyP-1 peptiderecognizes both the tumor cells and tumor-associated lym-phatic vessels, tumor sections were double-labeled withbaculovirus antibodies and with antibodies raised againstthe endothelial hyaluronic acid receptor-1 (LYVE-1) or
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm

1026 A. R. Makela et al.
Figure 4. Tumor accumulation of the LyP-1 displaying baculovirus. Mice bearing MDA-MB-435 (A) or MDA-MB-231 (B) xenografttumors were intravenously injected with 5 × 108 PFU of AcLyP-1-luc and Ac-luc (control) and allowed to circulate for 24 h.Tumor-associated virus was labeled with rabbit polyclonal AcMNPV primary antibody (red) and the cell nuclei were visualizedwith DAPI (blue). Magnification: (A) ×400; (B) ×200. Bar graphs show the semiquantitative analysis of virus-related fluorescenceintensity from the dissected tumors. Normalized fluorescence values determined relative to the control virus, Ac-luc (= 1), from arepresentative experiment are shown
vascular endothelial growth factor receptor-3 (VEGFR-3),both expressed in the endothelium of the lymphatic sys-tem. No apparent colocalization of the virus with thesemarkers or with the blood vessel marker, CD31, wasobserved (data not shown).
Inspection of different organs from the MDA-MB-435tumor-bearing mice showed accumulation of both theAcLyP-1-luc and Ac-luc in liver (Figure 5) and spleen(data not shown) whereas weaker labeling was observedin kidneys (Figure 5) and lungs (data not shown) andthe lowest amounts in the heart or brain (data notshown). The systemic biodistribution of the virusesdisplayed a similar pattern in both tumor-models. Theoverall specificity and efficiency of the AcLyP-1-luc was,however, significantly higher in the MDA-MB-435 tumorsas demonstrated by the higher tumor accumulation andweaker baculovirus-specific fluorescence in nontargetorgans.
Baculovirus transduction ofMDA-MB-435 cells is impeded duringthe early steps of viral entry
We previously reported that the MDA-MB-435 cellsare weakly permissive to baculovirus transduction asjudged by the transgene expression level [24]. Todetermine the mechanism underlying this transductionalblock, viral internalization, intracellular localization andtransgene expression were studied. Both the Ac-luc andAcLyP-1-luc appeared to form cytoplasmic aggregates
Figure 5. Biodistribution of the LyP-1-displaying and con-trol baculoviruses after systemic administration. MDA-MB-435tumor-bearing mice were intravenously injected with 5 × 108
PFU of AcLyP-1-luc and Ac-luc (control) and allowed to circulatefor 24 h. The distributed virus particles from different organswere labeled with a rabbit polyclonal AcMNPV primary anti-body (red) and cell nuclei were visualized with DAPI (blue).Magnification: ×400
peripheral to the plasma membrane in MDA-MB-435cells at 6 h p.t. as visualized by confocal microscopy(Figure 6). At this time point, only few AcLyP-1-luc capsids had reached the nucleus, whereas noapparent nuclear localization of the Ac-luc was detected.No considerable increase in the nuclear accumulationof the viruses was observed at later time points
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm

Tumor targeting of baculovirus 1027
(24–48 h p.t.; data not shown). In HepG2 controlcells, intranuclear localization of viral capsids was clearat 6 h p.t. with both the Ac-luc (Figure 6) and theAcLyP-1-luc (data not shown). The late endosomal andlysosomal marker LAMP-2, used to visualize vesicularcompartmentalization, showed partial colocalization withboth viruses in the HepG2 but not in the MDA-MB-435cells.
In HepG2 cells transgene, luciferase [24] or EGFP(not shown), expression by both virus phenotypes wasdetectable starting at 4 h p.t. and peaked approximatelyat 34 h p.t. In the MDA-MB-435 cells, maximal expressionlevels were achieved at approximately 48 h p.t. and thepercentage of transduced cells even at 1000 PFU/cellwas generally less than 1%. Treatment of the MDA-MB-435 cells with nocodazole, a microtubule depolymerizingagent, or with sodium butyrate, a histone deacetylaseinhibitor, had no enhancing effect on baculovirustransduction (not shown).
Figure 6. Cellular localization of baculoviruses in theMDA-MB-435 and HepG2 cells at 6 h p.t. The cells were trans-duced with AcLyP-1-luc and Ac-luc (200 PFU/cell) followed bylabeling with the rabbit polyclonal AcMNPV antibody and goatanti-rabbit Alexa Fluor488 conjugate (green). Late endosomesand lysosomes were labeled with the mouse monoclonal lyso-some-associated membrane protein 2 (LAMP-2) antibody andgoat anti-mouse Alexa Fluor546 conjugate (red). Nuclei werevisualized using ProLong Gold antifade reagent supplementedwith DAPI (blue). Scale bar = 10 µm
Table 1. MTT cytotoxicity assay presenting percentage of livingcells at 24 h p.t. with 200 PFU/cell of recombinant baculoviruses(mean ± SEM, performed in triplicate)
Cell line Control AcLyP-1-luc Ac-luc
MDA-MB-435 100.0 ± 13.3 86.42 ± 8.88 88.61 ± 5.33HepG2 100.0 ± 4.83 93.52 ± 3.69 99.45 ± 7.34
Display of the LyP-1 peptide on thebaculovirus envelope contributes to nosignificant cytotoxicity of the virus
Soluble LyP-1 peptide induces concentration-dependentcell death in cultured MDA-MB-435 cells upon bind-ing and internalization, and inhibits tumor growth inmice with xenografted tumors following systemic admin-istration [38]. To investigate whether the baculovirus-displayed LyP-1 peptide caused cell death, cytotoxicity ofthe recombinant viruses was determined by the MTT assayin living MDA-MB-435 and HepG2 cells at 24 h p.t. Nosignificant cytotoxicity was associated with transductionof the AcLyP-1-luc or the control virus, Ac-luc (Table 1).Correspondingly, transduction with the F3 [2], CGKRK[2] or RGD [25] peptide displaying viruses was nontoxic(not shown).
Discussion
Potential gene therapeutic applications of baculoviruswould significantly benefit from selective vector homingand entry to target cells in vivo as opposed to thebroad tropism that baculovirus naturally possesses. Thismotivates the development of vectors for improvedselectivity, efficiency and stability. Towards these aims,we pursued tumor-specific targeting of baculovirus byrestricting viral tropism via display of the lymphatichoming peptide LyP-1 on the viral envelope. Inaddition to improved performance in vitro, this vectorselectively accumulated to xenografted human MDA-MB-435 and MDA-MB-231 tumors in mice after systemicadministration, highlighting the potential of baculovirusin targeted therapies in vivo.
The LyP-1 peptide specifically homes to tumor-associated lymphatic vessels, and is specifically inter-nalized by certain tumor cells and tumor lymphaticendothelial cells by an unidentified cell surface recep-tor [37,38]. We have previously shown that the LyP-1peptide intensifies cellular attachment and transductionof baculovirus, and viral binding to MDA-MB-435 cells isinhibited by the soluble peptide with a high magnitudeand an apparent specificity [24]. In the present study, wereport a saturable binding of the LyP-1 displaying virus tothe MDA-MB-435 and HepG2 cells reaching the highestconstant efficiency at significantly lower concentrationsthan the control virus. As the ratio of physical versus infec-tious particles was similar with both viruses, these resultssuggest increased affinity of viral binding and saturation
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm

1028 A. R. Makela et al.
of receptor attachment sites for the LyP-1 displaying virus.By contrast, after reaching maximal efficiency, the bind-ing of the control virus linearly decreased, demonstratingthat the number of available binding sites and/or spacebecame restrictive and the further attachment above thesethreshold levels promoted viral release. This supports theearlier findings that binding of unmodified baculovirus tomammalian cells is nonsaturable and mediated throughlow-affinity ubiquitous cell surface motifs [20,21]. Thisprocess is rather slow (Kd of approximately 10−4 –10−6)compared to receptor-mediated binding (Kd of approx-imately 10−7 –10−11) [20,21]. In general, baculovirusbinding to the HepG2 cells is moderately efficient concen-trating on certain areas at the plasma membrane [23].In the present study, the overall efficacy and patternof viral binding in the MDA-MB-435 cells were similarto those in HepG2 cells. Accordingly, viral cluster for-mation at the cell surface was clearly visible with thecontrol virus in both cell lines, whereas binding of theLyP-1 displaying virus was highly abundant and exhib-ited more uniform and dispersed appearance. Althoughbinding of the fluorescein-conjugated LyP-1 is increasedto serum-starved MDA-MB-435 cells [38], starvation hadno significant effect on binding or subsequent gene deliv-ery of either virus to the MDA-MB-435 or HepG2 cells(Makela et al., unpublished results). As this feature of thesoluble LyP-1 peptide was not directly translated into theLyP-1 displaying baculovirus differing greatly in size andcomplexity from fluorescein, these results likely reflectthe involvement of more than one receptor in the cellularbinding of the virus. Together, these findings suggest thatthe LyP-1 peptide directs accumulation of baculovirus atspecific cell surface receptors.
The principles of baculovirus-mammalian cell interac-tions have been characterized [20–23,41–44] but thereceptor molecule(s) and the specific details underly-ing the mode and kinetics of viral entry are yet to bedefined. Because baculovirus surface modifications likelycontribute not only to viral binding, but also to the earlyevents of viral uptake competitive transduction exper-iments were performed. The fact that over a ten-foldexcess of the wild-type virus was not sufficient to com-pletely displace the LyP-1 displaying virus from the targetcells suggests that these viruses only partially competefor the same motif, and hence the modified virus exploitsalternative receptors for viral binding and internalization.The escalated viral uptake, earlier nuclear accumulationand higher transgene expression further support this anal-ysis. These results show that this tropism-modified virusexhibits subtle but biologically significant differences inthe mode and kinetics of viral binding and uptake, whichare mainly conferred by the nature of the displayed ligand.As we constantly used subsaturation quantities of virus, itis improbable that the amount of bound/internalized viruswould have affected the degree of internalization kinetics.Together, these results reflect the possibility of construct-ing a baculovirus vector capable of both targeting andmodulating the kinetics of viral entry.
The binding and internalization of baculovirus inmammalian cells have generally been considered uni-versal whereas endosomal escape, cytoplasmic traffickingand nuclear translocation represent critical steps dur-ing viral entry limiting the efficacy of gene transduction[11,20,21,23,41,42,44–46]. In the baculovirus permis-sive HepG2 cells, nuclear localization of virus capsidsis detectable starting at 3-4 h p.t. [23,43] in tandemwith the transgene expression [23,24] and is indepen-dent of mitotic disintegration [43]. In the MDA-MB-435cells, the progression of baculovirus entry was blockedat the periphery of the cytoplasm as demonstrated bythe aggregation of internalized viruses near the plasmamembrane and severely impaired nuclear accumulationof viral capsids. In addition, treatment with nocoda-zole, a microtubule depolymerizing agent known toenhance baculovirus transduction following endosomalrelease [41,44], or with sodium butyrate, a histonedeacetylase inhibitor mediating transcriptional activationof baculovirus-encoded transgenes [47], was unable torescue the transductional block. These results indicatethat the baculovirus transduction in the MDA-MB-435cells is impeded during the early steps of virus entry andlikely results from inefficient viral uptake and subsequentcytoplasmic release prior to nuclear entry and transgeneexpression.
Soluble LyP-1 peptide induces concentration-dependentcell death in the cultured MDA-MB-435 cells, and inhibitstumor growth in xenograft-bearing mice following sys-temic administration [38]. This proapoptotic/cytotoxicactivity is dependent on the efficient cellular bindingand internalization of the peptide [38]. The baculovirus-displayed LyP-1, however, caused no significant cyto-toxicity upon viral binding and internalization in theMDA-MB-435 or in HepG2 control cells. In addition to thecompromised viral uptake by the MDA-MB-435 cells, theapplied virus dose and the peptide copy number on theviral surface together may not correspond to the effectivedose or alignment of the soluble peptide. Alternatively,the antitumor activity of LyP-1 may be dependent on otherfactors such as solubility and/or successful transport to thetarget cell nuclei. As the baculovirus envelope, togetherwith the displayed peptides, is left behind following theendosomal escape, the viral nucleocapsid could providea functional vector platform for multimeric display anddelivery of LyP-1 to the target cell cytoplasm and nucleifor inhibition of tumor cell growth.
After demonstrating that the LyP-1 promotes bac-ulovirus performance in vitro, tumor homing of the viruswas evaluated following intravenous administrations. Asthe LyP-1 peptide was initially isolated in a screen usinghuman MDA-MB-435 xenografts [37], the same model, inaddition to the MDA-MB-231 breast carcinoma xenograftmodel, was applied to preserve the molecular character-istics of the original tumor. Because the MDA-MB-435cells are virtually resistant to baculovirus transduction,tumor-accumulation and systemic biodistribution of theinjected viruses were assessed by immunolabeling of thevirus particles to reliably compare the distributed virus
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm

Tumor targeting of baculovirus 1029
dose avoiding potential underestimations associated withmonitoring transgene expression. In comparison with thecontrol virus, the LyP-1 displaying vector injected into thetail vein of tumor-bearing mice was clearly confined tothe grafted tumors. As the control virus-derived fluores-cence in the tumor was regionally too low to be accuratelydistinguished from the background, tumor accumulationof the modified virus probably was at least ten- to 20-foldhigher than that of the control vector. Importantly, the F3and CGKRK peptide displaying viruses did not accumu-late into the MDA-MB-435 or MDA-MB-231 xenografts,indicating that the VSVG membrane anchor, a positivecharge or a general tumor-homing ability of the displayedpeptide, is not sufficient to target baculovirus to thesetumors. This further supports the specificity of the LyP-1-displaying virus. The LyP-1 virus-specific labeling wasdistributed over large regions of the tumor in a heteroge-neous pattern with focal areas showing stronger labelingat the tumor periphery. Some labeling was also observedin single cells within the tumor mass. The virus was par-ticularly concentrated to vessel-like structures resemblinglymphatic vessels. However, upon closer inspection withmarkers expressed in the endothelium of the lymphaticsystem, no apparent colocalization of the virus with theLYVE-1 or VEGFR-3 could be detected. In parallel, virus-specific fluorescence was absent from the vessels positivefor the blood vessel marker, CD31. As the viruses wereallowed to circulate for 24 h after injection, the viral par-ticles apparently had infiltrated within the tumor massthrough the leaky tumor vessels permeable to small-sizedparticles, and therefore colocalization of the virus withthe vascular markers was not observed. In general, thesystemic biodistribution of the viruses displayed a simi-lar pattern in both the MDA-MB-435 and MDA-MB-231tumor models. However, stronger tumor-associated flu-orescence and weaker labeling of the nontarget organsimplies that the specificity and efficiency of tumor hom-ing with the LyP-1 displaying virus was significantlyhigher in the MDA-MB-435 xenografts. This was antic-ipated because the LyP-1 peptide was originally identifiedusing the MDA-MB-435 tumors [37]. In nontarget organs,baculovirus-specific fluorescence was detected in liver andspleen with diminished amounts in kidneys and lungs, andnegligible amounts in brain and heart. This is consistentwith an earlier study where systemic delivery of phenotyp-ically unmodified baculovirus into complement deficientNeuro2a tumor-bearing A/J mice resulted in transgeneexpression primarily in liver, spleen and kidneys withvery low expression in lungs and heart [16]. Additionally,another study showed expression of GFP in liver, spleen,lung, heart, kidney and brains of BALB/c mice follow-ing intravenous administration of the baculovirus [48].Recently, single photon emission computed tomography-assisted biodistribution analysis of an avidin-displayingbaculovirus suggested association of the lymphatic sys-tem with systemic trafficking of the virus in a rat model[18]. Whether intrinsic to baculovirus or conferred bythe displayed avidin, this finding may nevertheless haveimplications on the development of baculovirus vectors
exploiting the lymphatic system for systemic, targetedtrafficking.
The LyP-1 peptide has been attached to different pay-loads varying in size, structure and chemical composition,such as T7 phage via a genetic fusion to the capsid pro-tein 10b, or quantum dots and fluorescein by chemicalcoupling [37,38,49]. Following systemic administration,these complexes have successfully been delivered tothe relevant vascular target sites in xenografted tumors[37,38,49]. With this type of study, baculovirus can beincluded within the therapeutic entities targetable by theLyP-1, highlighting the adaptation potential of the pep-tide to diverse systems. Being, on average, 256 × 68 nmin size (T. O. Ihalainen et al., unpublished data) and pos-sessing a phospholipid bilayer envelope derived from theinsect cell host, baculovirus represents by far the largestand the most complex cargo of LyP-1. Thus, by applyinga targeting ligand with high specificity and affinity, suchas LyP-1, specific cellular binding and tissue targeting ofbaculovirus appear to be feasible in the presence of themajor envelope glycoprotein, GP64, despite its ability toindiscriminately mediate viral binding and entry [50].Thereby, large-scale production of baculovirus vectors forclinical applications remains uncompromised, which is anexceptional advantage over most other virus vectors and,for the time being, over GP64-null baculoviruses [50].The targeting efficiency and specificity of the virus couldfurther be enforced by displaying multiple copies of thepeptide by fusion to both heterologous and endogenousmembrane anchors.
The present study emphasizes that differences in thestructures of the surface displayed targeting peptides havea detectable influence on the mode and kinetics of bindingand internalization of baculovirus vectors. Moreover, thisfirst demonstration of targeting baculovirus vehicles toxenografted tumors in vivo enhances the potential ofbaculovirus-mediated cancer therapies. Nevertheless, theeventual implementation of baculovirus in the clinicrequires further improvement of the vector in severalareas. The virus, administered systemically, should evadeinnate immune responses and inflammatory reactions,selectively target tumor tissue avoiding sequestrationby off-targets, and conduct specific cancer inhibition.These vectors could be specifically suitable for transient,acute expression of factors (e.g. proapoptotic secretedcytokines) that jeopardize the survival of proliferatingtarget cells without the need of transducing each cellwithin the tumor mass. Additionally, a viral vectormixture containing a cocktail of several surface andgenetic modifications may be required to address theheterogeneity of the disease.
Acknowledgements
We are grateful to Sharon Braunagel and Max Summers (TexasA&M University, TX, USA), as well as Loy Volkman (Universityof California at Berkeley, CA, USA) for providing baculovirus-specific antibodies. Teemu O. Ihalainen and Eila Korhonen(University of Jyvaskyla, Finland) are acknowledged for their
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm

1030 A. R. Makela et al.
excellent technical assistance. This study was supported bygrants from the Finnish Cultural Foundation (A.R.M.), theFinnish Konkordia Fund (A.R.M.), the Orion-Farmos ResearchFoundation (A.R.M.), the K. Albin Johansson Foundation(A.R.M. and J.P.L.), the Emil Aaltonen Foundation (A.R.M.and J.P.L.), the Finnish Foundation for Research on ViralDiseases (A.R.M. and J.P.L.), Finnish Cancer Foundations (P.L.)and the Academy of Finland (P.L. and M.V-R.). J.E. has beensupported by the Helsinki Biomedical Graduate School and J.P.Lby the National Graduate School of Informational and StructuralBiology.
References
1. Makela AR, Oker-Blom C. The baculovirus display technol-ogy – an evolving instrument for molecular screening and drugdelivery. Comb Chem High Throughput Screen 2008; 11: 86–98.
2. Makela AR, Oker-Blom C. Baculovirus display: a multifunctionaltechnology for gene delivery and eukaryotic librarydevelopment. Adv Virus Res 2006; 68: 91–112.
3. Kost TA, Condreay JP, Jarvis DL. Baculovirus as versatile vectorsfor protein expression in insect and mammalian cells. NatBiotechnol 2005; 23: 567–575.
4. Hu YC. Baculovirus vectors for gene therapy. Adv Virus Res 2006;68: 287–320.
5. Tani H, Limn CK, Yap CC, et al. In vitro and in vivo gene deliveryby recombinant baculoviruses. J Virol 2003; 77: 9799–9808.
6. Sarkis C, Serguera C, Petres S, et al. Efficient transduction ofneural cells in vitro and in vivo by a baculovirus-derived vector.Proc Natl Acad Sci USA 2000; 97: 14638–14643.
7. Li Y, Wang X, Guo H, Wang S. Axonal transport of recombinantbaculovirus vectors. Mol Ther 2004; 10: 1121–1129.
8. Pieroni L, Maione D, La Monica N. In vivo gene transfer inmouse skeletal muscle mediated by baculovirus vectors. HumGene Ther 2001; 12: 871–881.
9. Lehtolainen P, Tyynela K, Kannasto J, Airenne KJ, Yla-Herttuala S. Baculoviruses exhibit restricted cell type specificityin rat brain: a comparison of baculovirus- and adenovirus-mediated intracerebral gene transfer in vivo. Gene Ther 2002; 9:1693–1699.
10. Haeseleer F, Imanishi Y, Saperstein DA, Palczewski K. Genetransfer mediated by recombinant baculovirus into mouse eye.Invest Ophthalmol Vis Sci 2001; 42: 3294–3300.
11. Kaikkonen MU, Raty JK, Airenne KJ, Wirth T, Heikura T, Yla-Herttuala S. Truncated vesicular stomatitis virus G proteinimproves baculovirus transduction efficiency in vitro and in vivo.Gene Ther 2006; 13: 304–312.
12. Sandig V, Hofmann C, Steinert S, Jennings G, Schlag P,Strauss M. Gene transfer into hepatocytes and human liver tissueby baculovirus vectors. Hum Gene Ther 1996; 7: 1937–1945.
13. Ong ST, Li F, Du J, Tan YW, Wang S. Hybrid cytomegalovirusenhancer-h1 promoter-based plasmid and baculovirus vectorsmediate effective RNA interference. Hum Gene Ther 2005; 16:1404–1412.
14. Wang CY, Li F, Yang Y, Guo HY, Wu CX, Wang S. Recombinantbaculovirus containing the diphtheria toxin A gene for malignantglioma therapy. Cancer Res 2006; 66: 5798–5806.
15. Hoare J, Waddington S, Thomas HC, Coutelle C, McGarvey MJ.Complement inhibition rescued mice allowing observationof transgene expression following intraportal delivery ofbaculovirus in mice. J Gene Med 2005; 7: 325–333.
16. Kircheis R, Wightman L, Schreiber A, et al. Polyethylen-imine/DNA complexes shielded by transferrin target geneexpression to tumors after systemic application. Gene Ther 2001;8: 28–40.
17. Airenne KJ, Hiltunen MO, Turunen MP. Baculovirus-mediatedperiadventitial gene transfer to rabbit carotid artery. Gene Ther2000; 7: 1499–1504.
18. Raty JK, Liimatainen T, Huhtala T, et al. SPECT/CT imaging ofbaculovirus biodistribution in rat. Gene Ther 2007; 14: 930–938.
19. Raty JK, Liimatainen T, Wirth T, et al. Magnetic resonanceimaging of viral particle biodistribution in vivo. Gene Ther 2006;13: 1440–1446.
20. Duisit G, Saleun S, Douthe S, Barsoum J, Chadeuf G, Moullier P.Baculovirus vector requires electrostatic interactions includingheparan sulfate for efficient gene transfer in mammalian cells. JGene Med 1999; 1: 93–102.
21. Tani H, Nishijima M, Ushijima H, Miyamura T, Matsuura Y.Characterization of cell-surface determinants important forbaculovirus infection. Virology 2001; 279: 343–353.
22. Long G, Pan X, Kormelink R, Vlak JM. Functional entry ofbaculovirus into insect and mammalian cells is dependent onclathrin-mediated endocytosis. J Virol 2006; 80: 8830–8833.
23. Matilainen H, Rinne J, Gilbert L, Marjomaki V, Reunanen H,Oker-Blom C. Baculovirus entry into human hepatoma cells.J Virol 2005; 79: 15452–15459.
24. Makela AR, Matilainen H, White DJ, Ruoslahti E, Oker-Blom C.Enhanced baculovirus-mediated transduction of human cancercells by tumor-homing peptides. J Virol 2006; 80: 6603–6611.
25. Matilainen H, Makela AR, Riikonen R, et al. RGD motifs on thesurface of baculovirus enhance transduction of human lungcarcinoma cells. J Biotechnol 2006; 125: 114–126.
26. Kitagawa Y, Tani H, Limn CK, Matsunaga TM, Moriishi K,Matsuura Y. Ligand-directed gene targeting to mammalian cellsby pseudotype baculoviruses. J Virol 2005; 79: 3639–3652.
27. Raty JK, Airenne KJ, Marttila AT, et al. Enhanced gene deliveryby avidin-displaying baculovirus. Mol Ther 2004; 9: 282–291.
28. Luz-Madrigal A, Clapp C, Aranda J, Vaca L. In vivo transcrip-tional targeting into the retinal vasculature using recombinantbaculovirus carrying the human flt-1 promoter. Virol J 2007; 4:88.
29. Kim YK, Choi JY, Yoo MK, et al. Receptor-mediated genedelivery by folate-PEG-baculovirus in vitro. J Biotechnol 2007;131: 353–361.
30. Mahonen AJ, Airenne KJ, Purola S, et al. Post-transcriptionalregulatory element boosts baculovirus-mediated geneexpression in vertebrate cells. J Biotechnol 2007; 131: 1–8.
31. Strauss R, Huser A, Ni S, et al. Baculovirus-based vaccinationvectors allow for efficient induction of immune responsesagainst Plasmodium falciparum circumsporozoite protein. MolTher 2007; 15: 193–202.
32. Hofmann C, Strauss M. Baculovirus-mediated gene transfer inthe presence of human serum or blood facilitated by inhibitionof the complement system. Gene Ther 1998; 5: 531–536.
33. Hofmann C, Huser A, Lehnert W, Strauss M. Protection ofbaculovirus-vectors against complement-mediated inactivationby recombinant soluble complement receptor type 1. Biol Chem1999; 380: 393–395.
34. Huser A, Rudolph M, Hofmann C. Incorporation of decay-accelerating factor into the baculovirus envelope generatescomplement-resistant gene transfer vectors. Nat Biotechnol2001; 19: 451–455.
35. Enback J, Laakkonen P. Tumour-homing peptides: tools fortargeting, imaging and destruction. Biochem Soc Trans 2007;35: 780–783.
36. Ruoslahti E, Duza T, Zhang L. Vascular homing peptideswith cell-penetrating properties. Curr Pharm Des 2005; 11:3655–3660.
37. Laakkonen P, Porkka K, Hoffman JA, Ruoslahti E. A tumor-homing peptide with a targeting specificity related to lymphaticvessels. Nat Med 2002; 8: 751–755.
38. Laakkonen P, Akerman ME, Biliran H, et al. Antitumor activityof a homing peptide that targets tumor lymphatics and tumorcells. Proc Natl Acad Sci USA 2004; 101: 9381–9386.
39. Braunagel SC, Summers MD. Autographa californica nuclearpolyhedrosis virus, PDV, and ECV viral envelopes andnucleocapsids: structural proteins, antigens, lipid and fatty acidprofiles. Virology 1994; 202: 315–328.
40. Costes SV, Daelemans D, Cho EH, Dobbin Z, Pavlakis G,Lockett S. Automatic and quantitative measurement of protein-protein colocalization in live cells. Biophys J 2004; 86:3993–4003.
41. van Loo ND, Fortunati E, Ehlert E, Rabelink M, Grosveld F,Scholte BJ. Baculovirus infection of nondividing mammaliancells: mechanisms of entry and nuclear transport of capsids. JVirol 2001; 75: 961–970.
42. Kukkonen SP, Airenne KJ, Marjomaki V, et al. Baculoviruscapsid display: a novel tool for transduction imaging. Mol Ther2003; 8: 853–862.
43. Laakkonen JP, Kaikkonen MU, Ronkainen PH, et al. Baculovirus-mediated immediate-early gene expression and nuclear reorga-nization in human cells. Cell Microbiol 2007; 10: 667–681.
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm

Tumor targeting of baculovirus 1031
44. Salminen M, Airenne KJ, Rinnankoski R, et al. Improvement innuclear entry and transgene expression of baculoviruses bydisintegration of microtubules in human hepatocytes. J Virol2005; 79: 2720–2728.
45. Boyce FM, Bucher NL. Baculovirus-mediated gene transfer intomammalian cells. Proc Natl Acad Sci USA 1996; 93: 2348–2352.
46. Boublik Y, Di Bonito P, Jones IM. Eukaryotic virus display:engineering the major surface glycoprotein of the Autographacalifornica nuclear polyhedrosis virus (AcNPV) for thepresentation of foreign proteins on the virus surface.Biotechnology (NY) 1995; 13: 1079–1084.
47. Condreay JP, Witherspoon SM, Clay WC, Kost TA. Transient andstable gene expression in mammalian cells transduced with a
recombinant baculovirus vector. Proc Natl Acad Sci USA 1999;96: 127–132.
48. Kim YK, Park IK, Jiang HL, et al. Regulation of transductionefficiency by pegylation of baculovirus vector in vitro and in vivo.J Biotechnol 2006; 125: 104–109.
49. Akerman ME, Chan WC, Laakkonen P, Bhatia SN, Ruoslahti E.Nanocrystal targeting in vivo. Proc Natl Acad Sci USA 2002; 99:12617–12621.
50. Zhou J, Blissard GW. Display of heterologous proteins ongp64null baculovirus virions and enhanced budding mediatedby a VSV G-stem construct. J Virol 2007; 82: 1368–1377.
Copyright 2008 John Wiley & Sons, Ltd. J Gene Med 2008; 10: 1019–1031.DOI: 10.1002/jgm





























