Embed Size (px)
Citation preview

Accepted Manuscript
Title: Use of a live attenuated Salmonella enterica serovar Typhimurium
vaccine on farrow-to-finish pig farms
Author: L. De Ridder, D. Maes, J. Dewulf, P. Butaye, F. Pasmans, F. Boyen, F.
Haesebrouck, Y. Van der Stede
PII: S1090-0233(14)00381-5
DOI: http://dx.doi.org/doi: 10.1016/j.tvjl.2014.09.012
Reference: YTVJL 4277
To appear in: The Veterinary Journal
Accepted date: 13-9-2014
Please cite this article as: L. De Ridder, D. Maes, J. Dewulf, P. Butaye, F. Pasmans, F. Boyen, F.
Haesebrouck, Y. Van der Stede, Use of a live attenuated Salmonella enterica serovar
Typhimurium vaccine on farrow-to-finish pig farms, The Veterinary Journal (2014),
http://dx.doi.org/doi: 10.1016/j.tvjl.2014.09.012.
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service
to our customers we are providing this early version of the manuscript. The manuscript will
undergo copyediting, typesetting, and review of the resulting proof before it is published in its
final form. Please note that during the production process errors may be discovered which could
affect the content, and all legal disclaimers that apply to the journal pertain.

Use of a live attenuated Salmonella enterica serovar Typhimurium vaccine on farrow-to-1
finish pig farms 2 3
4
L. De Ridder a,b
, D. Maes b,
*, J. Dewulf b, P. Butaye
a,c, F. Pasmans
c, F. Boyen
c, F. 5
Haesebrouck c, Y. Van der Stede
a,d 6
7 a Unit of Co-ordination Veterinary Diagnose-Epidemiology and Risk Analysis, CODA-8
CERVA, Groeselenberg 99, 1180 Ukkel, Belgium 9 b Department of Obstetrics, Reproduction and Herd health, Faculty of Veterinary Medicine, 10
Ghent University, Salisburylaan 133, 9820 Merelbeke, Belgium 11 c Department of Pathology, Bacteriology and Avian diseases, Faculty of Veterinary Medicine, 12
Ghent University, Salisburylaan 133, 9820 Merelbeke, Belgium 13 d Department of Virology, Parasitology and Immunology, Faculty of Veterinary Medicine, 14
Ghent University, Salisburylaan 133, 9820 Merelbeke, Belgium 15
16
17
18
19
* Corresponding author. Tel.: +32 9 2647542. 20
E-mail address: [email protected] (D. Maes). 21
22
Page 1 of 22

23
Highlights 24
Salmonella enterica infection in pigs is economically important and poses a risk for 25
salmonellosis in humans. 26
The usefulness of an attenuated S. enterica serovar Typhimurium (S. Typhimurium) 27
strain to control S. enterica infections was investigated in three farrow-to-finish pig 28
herds infected with S. Typhimurium. 29
Within each herd, 120 piglets were orally vaccinated at 3 and 24 days of age, and 120 30
piglets served as unvaccinated controls. At specific time points, faeces, ileocaecal 31
lymph nodes caecal contents were examined for S. enterica by isolation and serum 32
was analysed for antibodies against S. enterica by ELISA. 33
All pigs were weighed at suckling and slaughter age to obtain complementary data on 34
the daily weight gain. 35
Prior to slaughter, there were significantly less S. enterica-excreting pigs, a 36
significantly lower S. enterica-specific mean antibody titre and a significantly higher 37
mean daily weight gain in vaccinated animals compared to unvaccinated controls. 38
In two herds, there were significantly lower proportions of S. enterica-positive 39
ileocaecal lymph nodes and caecal contents at slaughter between the vaccinated and 40
control groups, but this difference was not significant across all three herds. 41
S. enterica with the same auxotrophic characteristics and genotype as the vaccine 42
strain was isolated from several samples of faeces, ileocaecal lymph node and caecal 43
contents from vaccinated pigs. 44
These findings indicate that vaccination with an attenuated S. Typhimurium strain 45
reduces S. enterica shedding in S. enterica problem herds, but without consistent 46
Page 2 of 22

reduction of S. enterica loads and with possible persistence of the vaccine strain in 47
tissues. 48
49
Abstract 50
Salmonella enterica infection in pigs is economically important and poses a zoonotic 51
risk. In this study, the efficacy of an attenuated S. enterica serovar Typhimurium strain was 52
evaluated in three farrow-to-finish pig herds. In each herd, 120 piglets were vaccinated orally 53
at 3 and 24 days of age, while 120 piglets served as unvaccinated controls. Faeces, ileocaecal 54
lymph nodes and caecal contents were examined for S. Typhimurium by isolation and serum 55
was analysed for antibodies against S. Typhimurium by ELISA. All pigs were weighed at pre-56
weaning and slaughter to determine daily weight gain. In vaccinated pigs prior to slaughter, 57
significantly fewer animals excreted S. enterica, there was a significantly lower S. enterica-58
specific mean antibody titre and there was a significantly higher mean daily weight gain 59
compared to unvaccinated controls. In two herds, there were significantly lower proportions 60
of S. enterica positive ileocaecal lymph nodes and caecal contents at slaughter between the 61
vaccinated and control groups, but this difference was not significant across all three herds. S. 62
enterica with the same auxotrophic characteristics and genotype as the vaccine strain was 63
isolated from several samples of faeces, ileocaecal lymph nodes and caecal contents from 64
vaccinated pigs. These findings indicate that vaccination with an attenuated S. Typhimurium 65
strain reduces S. enterica shedding, but the reduction is not consistent and the vaccine strain 66
may persist in tissues. 67
68
Keywords: Salmonella enterica serovar Typhimurium; Pigs; Live attenuated vaccine; Field 69
trial70
Page 3 of 22

Introduction 71
The largest number of reported food-borne outbreaks in the EU is currently caused by 72
Salmonella enterica (26.6% of all outbreaks) (EFSA, 2013). In 2011, approximately 56.8% of 73
human salmonellosis cases were related to pigs and their products (EFSA, 2013). Although 74
most infected pigs do not exhibit clinical signs, S. enterica infections may reduce weight gain 75
(Boyen et al., 2009; Davis et al., 2010). Biosecurity measures may help to decrease levels of 76
S. enterica on pig farms (Lo Fo Wong et al., 2004a; Baptista et al., 2010), but they are often 77
insufficient as stand-alone measures. 78
79
Most vaccination studies against S. enterica in pigs have reported beneficial effects 80
(Springer et al., 2001; Roesler et al., 2004; Lindner et al., 2007; Selke et al., 2007; Rösler et 81
al., 2010; Leyman et al., 2012; Farzan and Friendship, 2010; Arguello et al., 2013; De Ridder 82
et al., 2013). However, most of these studies were conducted with relatively small numbers of 83
pre-weaning (Rösler et al., 2010) or weaned piglets (Springer et al., 2001; Roesler et al., 2004; 84
Selke et al., 2007; Leyman et al., 2012; De Ridder et al., 2013) and used a challenge infection 85
protocol. Their relevance for field conditions needs to be verified with large numbers of 86
finisher pigs (Lindner et al., 2007; Farzan and Friendship, 2010; Arguello et al., 2013). 87
88
Currently, the only S. enterica vaccine available commercially for use in pigs in the EU 89
is a live S. enterica serovar Typhimurium vaccine (Salmoporc, IDT Biologika), authorised 90
only in Germany and Poland. This vaccine has been evaluated on a large scale in sows and 91
weaned piglets at 3 and 6 weeks of age (Lindner et al., 2007). In a small scale experiment, 92
administration of two doses of this vaccine to pre-weaning piglets at 3 and 24 days of age 93
significantly reduced organ colonisation and faecal excretion (Rösler et al., 2010). This early 94
vaccination scheme at 3 and 24 days of age minimises labour, but its efficacy needs to be 95
Page 4 of 22

confirmed by large scale studies monitoring pigs to slaughter age. In addition, the possible 96
interference of this vaccination protocol with S. enterica serosurveillance requires 97
investigation. In the present study, we evaluated S. enterica-specific antibody titres in serum, 98
the presence of S. enterica in faeces and tissues, and weight gain of pigs after oral 99
immunisation with this commercially available live attenuated S. Typhimurium vaccine at 3 100
and 24 days of age. 101
102
Materials and methods 103
Herd selection and experimental animals 104
Three pig herds (A, B and C) were selected based on the following inclusion criteria: (1) 105
a high S. enterica-specific antibody level in finishing pigs as determined by the Belgian 106
national Salmonella Surveillance Programme, in accordance with EU Regulation number 107
2160/2003 (Van der Stede et al., 2008); (2) being a closed farrow-to-finish herd; (3) the 108
feasibility of monitoring piglets from birth to slaughter; and (4) agreement by the farmer to 109
apply the same management practices during the study as used previously, with the exception 110
of the use of antimicrobial agents, which was not allowed from 5 days before vaccination 111
until 5 days after vaccination. 112
113
The numbers of sows in herds A, B and C were 200, 180 and 300, and they practised a 114
3, 1 and 2 week batch production system, respectively. The three herds had a conventional 115
health status and did not experience substantial specific clinical disease problems. Pigs in all 116
three herds were vaccinated against Mycoplasma hyopneumoniae and pigs in herd B were 117
vaccinated against porcine circovirus type 2. Anthelmintic treatment was applied every 5 to 6 118
weeks in the fattening unit. Fully slatted floors were present in the nursery and fattening units 119
of all three herds. The pens held 25-30 pigs in the nursery units and 10-15 pigs in the 120
Page 5 of 22

fattening units. Weaned piglets received non-medicated liquid feed and fattening pigs 121
received liquid feed (herd A) or meal (herds B and C). 122
123
On each farm, 20 pregnant sows (Topigs20) were selected 3 days after farrowing, and 124
their piglets were randomly allocated at litter level to a vaccinated or unvaccinated group. All 125
pre-weaning piglets from each sow were included in the study and monitored until slaughter 126
age. The sows from the piglets in each group had the same parity distribution. The study was 127
approved by the ethical committee of the Faculty of Veterinary Medicine, Ghent University, 128
Belgium (EC 2011/196, date of approval January 2012). 129
130
Vaccination and sampling design 131
Details of the study design are summarised in Fig. 1. In each herd, ~120 piglets from 10 132
litters were vaccinated orally at 3 and 24 days of age with an attenuated histidine-adenine 133
auxotrophic S. Typhimurium strain (Salmoporc, IDT Biologika) (V, vaccination group), while 134
120 piglets from 10 other litters were unvaccinated (C, control group). Transfer of piglets 135
from one sow to another was not allowed after 3 days of age. All piglets were weaned at 24 136
days of age. From weaning until slaughter age, the pigs in the V and C groups were kept 137
separately on the farm, during transportation to the abattoir and in the lairage. 138
139
Within each herd, rectal faeces and blood were collected serially from 30 randomly 140
selected pigs per group. Blood was collected into plain tubes (Sarstedt 15 mL). At slaughter, 141
caecal contents and ileocaecal lymph nodes were collected from the same 30 pigs, while 142
ileocaecal lymph nodes were collected from an additional 40 randomly selected slaughter 143
pigs. All pigs were weighed to calculate the daily weight gain (DWG) between 3 days and 29 144
weeks of age. 145
Page 6 of 22

146
Bacteriology 147
S. enterica was isolated from ~20 g faecal samples (rectal and caecal contents) using the 148
ISO 6579 Annex D method (ISO, 2007). Samples were diluted 1:10 in buffered peptone water 149
(BPW, Bio-Rad) and incubated for 16-20 h at 37 °C. A quantity of 100 μL from every pre-150
enrichment solution was spotted onto one modified semi-solid Rappaport-Vassiliadis plate 151
(MSRV, Bio-Rad) and incubated for 46-50 h at 41 °C. A loopful from the edge of a typical 152
migration zone on each MSRV plate was streaked onto a xylose lysine deoxycholate agar 153
plate (XLD, Bio-Rad) and a brilliant green agar plate (BGA, Lab M). Both plates were 154
incubated at 37 °C for 21-27 h. When S. enterica presumptive colonies were present on both 155
the XLD and BGA plates, one colony was inoculated in triple sugar iron agar (TSI, Bio-Rad), 156
sorbitol agar (Becton Dickinson) and lysine decarboxylase broth (Oxoid), and incubated for 157
18-24 h at 37 °C. 158
159
Ileocaecal lymph nodes (~10-13 g) were flamed briefly to decontaminate the surface, 160
then sliced, diluted 1:10 in BPW and homogenised with a stomacher blender (BagMixer, 161
Interscience) for 1 min. Samples were then processed as for faecal samples. 162
163
Isolates from vaccinated pigs were tested using the S. enterica Diagnostic Kit, produced 164
by the manufacturer of the vaccine (IDT Biologika). This kit distinguishes wild type S. 165
enterica strains from the vaccine strain through the use of two fluid media (A and B). Since 166
the vaccine strain is histidine-adenine auxotrophic, it only grows in medium B, which 167
contains histidine and adenine, whereas wild type strains grow in both media. In addition, 168
isolates were typed by multiple locus variable number tandem repeat analysis (MLVA) 169
(Larsson et al., 2009) and pulsed field gel electrophoresis (PFGE) (Peters et al., 2009). 170
Page 7 of 22

171
Serology 172
After coagulation, blood samples were centrifuged for 5 min at 1500 g to collect serum, 173
which was diluted 20-fold and analysed for S. enterica-specific antibodies with a commercial 174
ELISA kit based on lipopolysaccharide (LPS) O-antigens of serogroups B, C1 and D 175
(HerdChek Swine Salmonella, IDEXX Laboratories). Results are expressed as a sample to 176
positive ratio (S:P); samples with S:P ratios ≥ 0.25 (OD% ≥ 10) were defined as positive. 177
178
Statistical analysis 179
The number of positive faecal samples was compared between vaccinated and control 180
animals by means of generalised estimating equations (GEE; Proc Genmod in SAS 9.2). A 181
logit link function, a binomial distribution and an exchangeable correlation structure were 182
used; correlation is the same among any two observations from the same animal. Mortality 183
and isolation data at slaughter were analysed using the Pearson χ2 test in SAS 9.2. In these 184
analyses, samples containing the vaccine strain were considered to be negative for S. enterica. 185
186
The differences in S:P ratios between vaccinated and control animals were analysed by 187
repeated measures analysis of variance (ANOVA) using the Proc Mixed procedure with 188
lsmeans and slice options in SAS 9.2. Time and group (vaccinated and control) were included 189
in the model as fixed effects, and the herd as a random effect. An unstructured covariance 190
structure was selected, based on the lowest Akaike Information Criterion (AIC) value. To 191
adjust for the multiple comparisons in this procedure, the Ryan-Einot-Gabriel-Welch 192
(REGWQ) test was used. P values < 0.05 were considered to be significant (two-sided tests). 193
194
Page 8 of 22

The DWG between 3 days and 29 weeks of age for each pig (excluding pigs that died 195
before slaughter) was normally distributed and was compared between the vaccinated and 196
control groups using the Proc Mixed procedure with the lsmeans option and taking into 197
account the herd effect (SAS 9.2). 198
199
Results 200
Some animals in both groups were lost for follow-up due to mortality (runts, physical 201
abnormalities or idiopathic causes) or lost ear tags (Fig. 1). There was no significant 202
difference in mortality between groups V (50/347, 14.4%) and C (59/362, 16.3%; P = 0.90). 203
204
Bacteriology of faecal samples collected on-farm 205
The proportions of S. enterica positive faecal samples collected on-farm are shown for 206
the three sampling occasions in Fig. 2. Some data are missing, due to an empty rectum or lost 207
ear tags (Fig. 1). Across all herds, the proportions of pigs excreting S. enterica did not differ 208
significantly between groups V and C at 10 weeks of age (9/99, 9.1% vs. 11/98, 11.2%, 209
respectively; P = 0.46) and 16 weeks of age (9/89, 10.1% vs. 11/82, 13.4%, respectively; 210
P = 0.50). However, 2 weeks prior to slaughter (at 29 weeks of age), this proportion was 211
significantly different between V (7/79, 8.9%) and C (22/79, 27.8%; P < 0.01). 212
213
When analysing each herd separately, the proportion of pigs excreting S. enterica was 214
also lower in herd B (1/29, 3.4% for V vs. 12/29, 41.4% for C; P < 0.01) and herd C (0/22, 215
0.0% for V vs. 6/22, 27.3% for C; P < 0.01), whereas this was not the case in herd A (6/28, 216
21.4% for V vs. 4/28, 14.3% for C; P = 0.49). 217
218
Bacteriology of faecal samples collected at the abattoir 219
Page 9 of 22

The proportions of S. enterica positive samples from ileocaecal lymph nodes and caecal 220
contents collected at the abattoir are presented in Table 1; some data are missing, due to lost 221
ear tags (Fig. 1). Over all herds, there were no significant differences in the proportions of 222
samples positive by isolation for S. enterica between groups V and C for ileocaecal lymph 223
nodes (73/201, 36.3% vs. 72/180, 40.0%, respectively; P = 0.46) and caecal contents (37/121, 224
30.6% vs. 49/115, 42.6%, respectively; P = 0.06). 225
226
When analysing each herd separately, significant differences were found in the 227
proportions of samples positive by isolation for S. enterica between groups V and C in two 228
herds: (1) in lymph nodes from herd B (37/64, 57.8% for V vs. 47/61, 77.0% for C; P < 0.03); 229
and (2) in caecal contents from herd C (9/49, 18.4% for V vs. 24/44, 54.5% for C; P < 0.01). 230
231
Detection of the vaccine strain of Salmonella enterica 232
The proportions of samples putatively positive for the S. enterica vaccine strain in each 233
herd are presented in Table 2. Using the Salmonella Diagnostic Kit, 3/99 (3.0%), 2/89 (2.2%) 234
and 3/79 (3.8%) isolates from faecal samples of vaccinated pigs were histidine-adenine 235
auxotrophic at 10, 16 and 29 weeks of age, respectively. Histidine-adenine auxotrophic 236
isolates were detected in 10/121 (8.3%) samples of caecal contents and 4/201 (2.0%) samples 237
of lymph nodes. All histidine-adenine auxotrophic isolates belonged to the same MLVA type 238
and had the same PFGE profile as the vaccine strain. 239
240
Serology 241
S. enterica-specific antibody levels are shown for each group, herd and sampling 242
occasion in Fig. 2. At 3 days of age, prior to vaccination, mean ± (standard deviation, SD) S:P 243
ratios across all herds were comparable between groups V (1.50 ± 0.87; n = 120) and C 244
Page 10 of 22

(1.53 ± 0.79; n = 119; P = 0.73). In herd A, there was a significant difference in mean (± SD) 245
S:P ratios between groups V (2.11 ± 0.68; n = 40) and C (1.75 ± 0.58; n = 40; P < 0.03). 246
247
S:P ratios in groups V and C were three- to five-fold lower at 10 weeks of age than at 3 248
days of age (P < 0.001). The mean (± SD) S:P ratio was significantly higher in group V 249
(0.49 ± 0.51; n = 100) than group C (0.31 ± 0.46; n = 102; P < 0.01). This was mainly due to 250
the significant differences in mean (± SD) S:P ratios between groups V and C in herds A (V: 251
0.43 ± 0.33; n = 35 vs. C: 0.18 ± 0.21; n = 36; P < 0.02) and C (V: 0.71 ± 0.70; n = 33 vs. C: 252
0.09 ± 0.21; n = 37; P < 0.01). In herd B, the mean (± SD) S:P ratio was 0.32 ± 0.36 (n = 32) 253
for V and 0.66 ± 0.66 (n = 29) for C (P < 0.01). 254
255
At 29 weeks of age, the mean (± SD) S:P ratio across all herds was significantly higher 256
in group C (1.07 ± 0.55; n = 89 for V vs. 1.52 ± 0.68; n = 87 for C; P < 0.01). This outcome 257
was mainly attributable to the results obtained in herds A (0.94 ± 0.39 for V vs. 1.53 ± 0.55 258
for C; P < 0.01) and C (0.86 ± 0.54 for V vs. 1.69 ± 0.71 for C; P < 0.01). On farm B, there 259
was no significant difference in mean (± SD) S:P ratios between groups V (1.39 ± 0.58) and C 260
(1.34 ± 0.75; P = 0.71). 261
262
Daily weight gain 263
Over all three herds, a significantly higher DWG (P < 0.01) was observed in group V 264
(n = 289; DWG 546.8 g, 95% confidence interval, CI, 537.7-555.8 g) compared to group C 265
(n = 302; DWG, 509.3 g, 95% CI 499.4-519.2 g; Table 3). This result was consistent in each 266
herd; differences in mean DWG between groups V and C were 29.6 g in herd A, 37.2 g in 267
herd B and 38.9 g in herd C (Table 3; P < 0.01). 268
269
Page 11 of 22

Discussion 270
In this study, a live vaccine administered orally to piglets at 3 and 24 days of age 271
resulted in lower S. enterica excretion in faeces and lower serum antibody levels in vaccinated 272
pigs than in control animals prior to slaughter. An increased DWG in vaccinated animals was 273
also evident. Apart from these beneficial effects, some limitations of the vaccine under field 274
conditions were found. 275
276
Although there was considerable variation among the three herds, significantly fewer 277
vaccinated pigs overall excreted S. enterica prior to slaughter compared to control pigs. This 278
is important, since finishing pigs may introduce S. enterica into the abattoir environment 279
(Letellier et al., 2009). In the vaccinated group, there was no significant difference in the 280
proportion of pigs excreting S. Typhimurium at 10 and 16 weeks, whereas at 29 weeks the 281
percentage was significantly lower; a possible explanation is that the level of infection with S. 282
Typhimurium in the herds included in the study was higher at 29 weeks than at 10 and 16 283
weeks, allowing more opportunity for improvement due to vaccination. A significantly lower 284
S. enterica-specific antibody level was observed before slaughter in vaccinated animals across 285
all herds, suggesting decreased S. enterica infection pressure at this stage of growth (Lo Fo 286
Wong et al., 2004b; Baptista et al., 2009). 287
288
The variable results among the three herds might be due to several reasons. Under field 289
conditions, pigs are infected at different points in time, with a herd-dependent and even batch-290
dependent variability in both infection pressure and host response (Beloeil et al., 2003; Lo Fo 291
Wong et al., 2004b; Rostagno et al., 2012). The presence of herd-specific S. enterica strains 292
might have affected vaccination (Van Parys et al., 2013). The degree of separation of V and C 293
groups was most stringent in herd C (separated rooms), followed by herd A (separated pens, 294
Page 12 of 22

vaccinated pens grouped together) and herd B (separated pens, vaccinated pens dispersed 295
among unvaccinated ones). Antibody titres and faecal excretion of S. enterica in herd C were 296
significantly reduced in vaccinated pigs at finishing, in contrast with herds A and B. This 297
suggests that the best vaccination results will be obtained when all pigs in a herd (or 298
shed/barn) are vaccinated (Arguello et al., 2013). Finally, different maternal antibody levels in 299
piglets at the time of vaccination might have contributed to herd variation. 300
301
Piglets were vaccinated at 3 and 24 days of age to induce protection against possible 302
infection in the nursery period. However, most pigs seroconverted to S. enterica at the start of 303
the fattening period, similar to other studies (Beloeil et al., 2003). Nevertheless, vaccination at 304
an early age has some practical advantages, since piglets are normally handled at this time and 305
can be picked up easily to apply the vaccine orally. 306
307
Rösler et al. (2010) did not demonstrate a negative effect of maternal antibodies on S. 308
enterica vaccination using the same vaccine in sucking pigs. However, in the present study, 309
the herd with the highest maternal S. enterica-specific antibody levels in pigs at first 310
vaccination was the only herd without significantly decreased S. enterica excretion in the 311
vaccinated pigs at finishing stage. Thus, maternal immunity in this herd might have interfered 312
with oral vaccination. 313
314
In a previous study, the vaccine strain could not be detected in the lymph nodes of 315
slaughter pigs that had been vaccinated at 21 and 42 days of age (Lindner et al., 2007). 316
Therefore, persistence of the vaccine strain was not expected. However, the present study 317
indicated that S. enterica isolates similar to the vaccine strain could be detected in faeces 318
throughout the fattening period in all three herds and in the lymph nodes and caecal contents 319
Page 13 of 22

at slaughter. This finding might have important implications for bacteriological monitoring 320
programmes. It is uncertain whether the histidine-adenine auxotrophic S. Typhimurium 321
vaccine strain used in the present study is pathogenic for human beings. 322
323
The DWG was significantly higher in vaccinated pigs, suggesting that vaccination is 324
associated with a higher feed conversion ratio; however, the feed conversion ratio could not 325
be determined in the present study, since feed intake could not be recorded reliably. 326
327
Conclusions 328
The early vaccination scheme with a current commercial live attenuated S. 329
Typhimurium vaccine in pig herds with a high level of S. enterica infection reduced faecal 330
excretion and was associated with lower S. enterica-specific antibody titres at slaughter age 331
when compared to unvaccinated pigs. This vaccine could be a useful tool in the control of S. 332
enterica infection. However, the effects were variable between herds. Furthermore, there were 333
indications that the vaccine strain could persist until slaughter age. 334
335
Conflict of interest statement 336
None of the authors of this paper has a financial or personal relationship with other 337
people or organisations that could inappropriately influence or bias the content of the paper. 338
339
Acknowledgements 340
The authors thank the Federal Public Service of Health, Food Chain Safety and 341
Environment for funding this project (RT-09/05) and the participating farmers for their 342
cooperation. The assistance of veterinary colleagues and students in sampling, and the help of 343
Page 14 of 22

H. Vander Veken, M. Van Hessche and A. Lucchina in performing the diagnostic kit test, 344
MLVA and PFGE are gratefully acknowledged. 345
346
References 347
Arguello, H., Carvajal, A., Naharro, G., Rubio, P., 2013. Evaluation of protection conferred 348
by a Salmonella Typhimurium inactivated vaccine in Salmonella-infected finishing pig 349
farms. Comparative Immunology, Microbiology and Infectious Diseases, 36, 489-498. 350
351
Baptista, F.M., Alban, L., Ersbøll, A.K., Nielsen, L.R., 2009. Factors affecting persistence of 352
high Salmonella serology in Danish pig herds. Preventive Veterinary Medicine 92, 301-353
308. 354
355
Baptista, F.M., Alban, L., Nielsen, L.R., Domingos, I., Pomba, C., Almeida, V., 2010. Use of 356
herd information for predicting Salmonella status in pig herds. Zoonoses and Public 357
Health 57, 49-59. 358
359
Beloeil, P.A., Chauvin, C., Proux, K., Rose, N., Queguiner, S., Eveno, E., Houdayer, C., 360
Rose, V., Fravalo, P., Madec, F., 2003. Longitudinal serological responses to 361
Salmonella enterica of growing pigs in a subclinically infected herd. Preventive 362
Veterinary Medicine 60, 207-226. 363
364
Boyen, F., Pasmans, F., Van Immerseel, F., Donne, E., Morgan, E., Ducatelle, R., 365
Haesebrouck, F., 2009. Porcine in vitro and in vivo models to assess the virulence of 366
Salmonella enterica serovar Typhimurium for pigs. Laboratory Animals 43, 46-52. 367
368
Davis, B.L., Fraser, J.N., Burkey, T.E., Skjolaas, K.A., Dritz, S.S., Johnson, B.J., Minton, 369
J.E., 2010. Oral inoculation with Salmonella enterica serovar Typhimurium or 370
Choleraesuis promotes divergent responses in the somatotropic growth axis of swine. 371
Journal of Animal Science 88, 1642-1648. 372
373
De Ridder, L., Maes, D., Dewulf, J., Pasmans, F., Boyen, F., Haesebrouck, F., Meroc, E., 374
Butaye, P., Van der Stede, Y., 2013. Evaluation of three intervention strategies to 375
reduce the transmission of Salmonella Typhimurium in pigs. The Veterinary Journal 376
197, 613-618. 377
378
EFSA (European Food Safety Authority), 2013. The European Union summary report on 379
trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2011. 380
EFSA Journal 11, 3129. 381
382
Farzan, A., Friendship, R.M., 2010. A clinical field trial to evaluate the efficacy of 383
vaccination in controlling Salmonella infection and the association of Salmonella-384
shedding and weight gain in pigs. Canadian Journal of Veterinary Research 74, 258-385
263. 386
387
Page 15 of 22

ISO (International Organization of Standardization), 2007. Detection of Salmonella spp. in 388
animal feces and in environmental samples from the primary production stage. ISO 389
6579:2002/Amd 1:2007, Annex D. 390
391
Larsson, J.T., Torpdahl, M., Petersen, R.F., Sorensen, G., Lindstedt, B.A., Nielsen, E.M., 392
2009. Development of a new nomenclature for Salmonella Typhimurium multilocus 393
variable number of tandem repeats analysis (MLVA). Eurosurveillance 14, 1-5. 394
395
Letellier, A., Beauchamp, G., Guevremont, E., D'Allaire, S., Hurnik, D., Quessy, S., 2009. 396
Risk factors at slaughter associated with presence of Salmonella on hog carcasses in 397
Canada. Journal of Food Protection 72, 2326-2331. 398
399
Leyman, B., Boyen, F., Verbrugghe, E., Parys, A.V., Haesebrouck, F., Pasmans, F., 2012. 400
Vaccination of pigs reduces Salmonella Typhimurium numbers in a model mimicking 401
pre-slaughter stress. The Veterinary Journal 194, 250-252. 402
403
Lindner, T., Springer, S., Selbitz, H.J., 2007. The use of a Salmonella Typhimurium live 404
vaccine to control Salmonella Typhimurium in fattening pigs in field and effects on 405
serological surveillance. Proceedings of the 7th International Safepork Symposium on 406
Epidemiology and Control of Foodborne Pathogens in Pork, Verona, Italy, pp. 237-239. 407
408
Lo Fo Wong, D.M.A., Dahl, J., Stege, H., van der Wolf, P.J., Leontides, L., von Altrock, A., 409
Thorberg, B.M., 2004a. Herd-level risk factors for subclinical Salmonella infection in 410
European finishing-pig herds. Preventive Veterinary Medicine 62, 253-266. 411
412
Lo Fo Wong, D.M.A., Dahl, J., Wingstrand, A., van der Wolf, P.J., von Altrock, A., 413
Thorberg, B.M., 2004b. A European longitudinal study in Salmonella seronegative- and 414
seropositive-classified finishing pig herds. Epidemiology and Infection 132, 903-914. 415
416
Peters, T.M., 2009. Pulsed-field gel electrophoresis for molecular epidemiology of food 417
pathogens. In: Caugant, D.A. (Ed.). Molecular Epidemiology of Microorganisms: 418
Methods in Molecular Biology. Humana Press, New York, USA, pp. 59-70. 419
420
Roesler, U., Marg, H., Schröder, I., Mauer, S., Arnold, T., Lehmann, J., Truyen, U., Hensel, 421
A., 2004. Oral vaccination of pigs with an invasive gyrA-cpxA-rpoB Salmonella 422
Typhimurium mutant. Vaccine 23, 595-603. 423
424
Rösler, U., Stief, M., Leffler, M., Truyen, U., Lehmann, J., Szabo, I., Springer, S., Lindner, 425
T., Selbitz, H.J., 2010. Persistenz, ausscheidung und wirksamkeit der attenuierten 426
Salmonella Typhimurium vakzine Salmoporc® bei saugferkeln. Der Praktischer Tierarzt 427
91, 59-65. 428
429
Rostagno, M.H., Hurd, H.S., McKean, J.D., 2012. Variation of bacteriologic and serologic 430
Salmonella enterica prevalence between cohorts within finishing swine production 431
farms. Food Research International 45, 867-870. 432
433
Selke, M., Meens, J., Springer, S., Frank, R., Gerlach, G.F., 2007. Immunization of pigs to 434
prevent disease in humans: Construction and protective efficacy of a Salmonella 435
enterica serovar Typhimurium live negative-marker vaccine. Infection and Immunity 436
75, 2476-2483. 437
Page 16 of 22

438
Springer, S., Lindner, T., Steinbach, G., Selbitz, H.J., 2001. Investigation of the efficacy of a 439
genetically-stabile live Salmonella Typhimurium vaccine for use in swine. Berliner und 440
Münchener Tierärztliche Wochenschrift 114, 342-345. 441
442
Van der Stede, Y., Bollaerts, K., Abrahantes Cortinãs, J., Aerts, M., Imberechts, H., Mintiens, 443
K., 2008. Assignment of Salmonella risk farms by serological surveillance at pre-444
harvest level: An impossible task!? Proceedings of the 20th Congress of the 445
International Pig Veterinary Society, Durban, South Africa, pp. 175. 446
447
Van Parys, A., Boyen, F., Leyman, B., Verbrugghe, E., Maes, D., Haesebrouck, F., Pasmans, 448
F., 2013. Induction of seroconversion and persistence of Salmonella Typhimurium in 449
pigs are strain dependent. Comparative Immunology, Microbiology and Infectious 450
Diseases 36, 465-471. 451
452
453
Figure legends 454
455
Fig.1. Flowchart of the study design in the three farrow-to-finish herds. In total, 456
approximately 700 pigs were selected; half of them were vaccinated, whereas the other half 457
were unvaccinated and used as controls. Treatment pigs were vaccinated orally at 3 and 24 458
days of age with an attenuated Salmonella enterica serovar Typhimurium strain (Salmoporc, 459
IDT Biologika). Blood and faeces were collected from 90 vaccinated and 90 control pigs. 460
Sera were analysed by ELISA (HerdChek Swine Salmonella, IDEXX Laboratories). 461
Salmonella isolation (ISO 6579 Annex D) was performed on faeces. At slaughter, ileocaecal 462
lymph nodes (210 vaccinated and 210 control pigs) and caecal content (105 vaccinated and 463
105 control pigs) were collected for Salmonella isolation (ISO 6579 Annex D. All study pigs 464
were weighed at 3 days and 29 weeks of age to calculate daily weight gain (DWG). * Extra 465
data were gathered initially to account for possible mortality losses later. ° Some data were 466
missing due to identification (loss of ear tags) or sampling (empty rectum) problems. 467
468
Fig. 2. Pigs excreting Salmonella enterica (%) (A, C, E) and S. enterica-specific antibody 469
levels in blood (mean S:P ratio) (B, D, F) per group and herd at 3 days of age, 10 weeks of 470
Page 17 of 22

age, 16 weeks of age and 29 weeks of age (2 weeks prior to slaughter). Error bars represent 471
the 95% confidence interval. Asterisks (*) below the graphs indicate a significant difference 472
(P < 0.05) between the control (black) and vaccinated (grey) groups in herd A (A, B), herd B 473
(C, D) and herd C (E, F). 474
475
Page 18 of 22
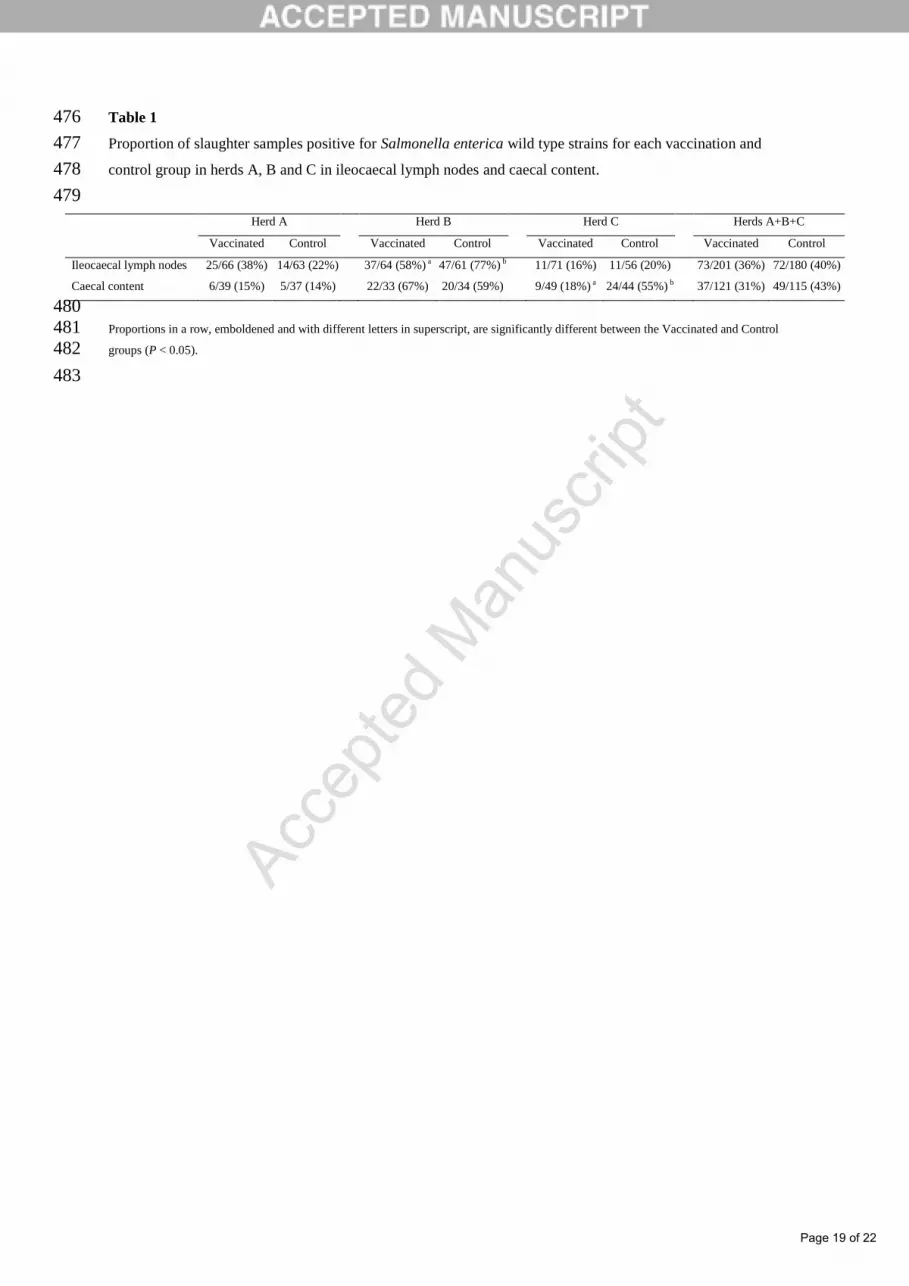
Table 1 476
Proportion of slaughter samples positive for Salmonella enterica wild type strains for each vaccination and 477
control group in herds A, B and C in ileocaecal lymph nodes and caecal content. 478
479
Herd A Herd B Herd C Herds A+B+C
Vaccinated Control Vaccinated Control Vaccinated Control Vaccinated Control
Ileocaecal lymph nodes 25/66 (38%) 14/63 (22%) 37/64 (58%) a 47/61 (77%) b 11/71 (16%) 11/56 (20%) 73/201 (36%) 72/180 (40%)
Caecal content 6/39 (15%) 5/37 (14%) 22/33 (67%) 20/34 (59%) 9/49 (18%) a 24/44 (55%) b 37/121 (31%) 49/115 (43%)
480
Proportions in a row, emboldened and with different letters in superscript, are significantly different between the Vaccinated and Control 481
groups (P < 0.05). 482
483
Page 19 of 22

Table 2 484
Proportion of samples from vaccinated pigs, which possibly contain the Salmonella enterica serovar 485
Typhimurium vaccine strain in faeces at 10, 16 and 29 weeks of age, and in ileocaecal lymph nodes 486
and caecal contents at slaughter. 487
488
Herd
Faeces Ileocaecal lymph
nodes Caecal contents
10 weeks 16 weeks 29 weeks
A 2/35 (5.7%) 0/32 (0.0%) 0/28 (0.0%) 1/66 (1.5%) 2/39 (5.1%)
B 1/30 (3.3%) 1/28 (3.6%) 0/29 (0.0%) 0/64 (0.0%) 2/33 (6.1%)
C 0/34 (0.0%) 1/29 (3.4%) 3/22 (13.6%) 3/71 (4.2%) 6/49 (12.2%)
A+B+C 3/99 (3.0%) 2/89 (2.2%) 3/79 (3.8%) 4/201 (2.0%) 10/121 (8.3%)
489
Page 20 of 22

Table 3 490
Daily weight gain (DWG; g) between 3 days and 29 weeks of age in pigs in the Vaccinated and 491
Control groups for each of the three farrow-to-finish pig herds and over all herds. 492
493
Herd Mean DWG (95% confidence interval) Difference DWG (P)
Vaccinated Control
A 593.0 (578.8-607.3) 563.4 (549.0-577.9) 29.6 (< 0.01)
B 504.3 (488.9-519.8) 467.1 (452.5-481.7) 37.2 (< 0.01)
C 535.6 (520.9-550.4) 496.7 (482.3-511.0) 38.9 (< 0.01)
A+B+C 546.8 (537.7-555.8) 509.3 (499.4-519.2) 37.5 (< 0.01)
494
495
Page 21 of 22

496
Page 22 of 22





























