Embed Size (px)
Citation preview

Protoplasma (1997) 198:85-92 PROTOPLASMA �9 Springer-Verlag 1997 Printed in Austria
Vacuole motility and tubule-forming activity in Pisolithus tinctorius hyphae are modified by environmental conditions
Geoffrey J. Hyde* and Anne E. Ashford
School of Biological Science, The University of New South Wales, Sydney, New South Wales
Received October 9, 1996 Accepted March 5, 1997
Summary, Motile tubular vacuole systems have been visualised using DIC optics in living hyphae of Pisolithus tinctorius without loading of any fluorescent tracer. Adding new medium, with or with- out the tracer CFDA, alters the motility of this system and increases the number of tubules. This response has been shown in individual hyphal tip cells and quantified in populations of tip cells. Vacuoles with motile tubules are also demonstrated in more basal cells of the hyphae, within 600 gm of the growing hyphal front. The vacuoles in these cells show more limited motility, but similarly respond to addi- tion of new medium by increased motility and tubular activity. This demonstration that the vacuole system in more mature regions is both motile and interconnected as in the tips, and similarly responds to changes in external conditions, supports the hypothesis that the vac- uole system may play a role in long-distance transport. Vacuoles in the most mature cells, more than 600 am behind the hyphal growth zone are not motile. They do not respond to these stimuli and remain spherical and isolated. There are many explanations for this and the present lack of response does not exclude the transport hypothesis. The findings further support the concept that tubular vacuole systems are equivalent to animal endosomal/lysosomal systems and have implications for their motility, especially their plasticity in response to external stimuli, such as fluorescent tracers.
Keywords: Tubular vacuoles; Pisolithus tinctorius; Endosomes; 6- Carboxyfluorescein; Long-distance transport.
Abbreviations: CFDA 6-carboxyfluorescein diacetate; DIC differ- ential interference contrast; MMN modified Melin-Norkrans medi- um; SEM standard error of the mean.
Introduction
The endosomal / lysosomal pa thway is known to play a central role in molecular sorting, process ing and
*Correspondence and reprints: School of Biological Science, The University of New South Wales, Sydney, NSW 2052, Australia.
b reakdown of constitutive and ingested molecules in
animal cells (van Deurs et al. 1989). It has recently
been speculated that the motile vacuolar system in the
hyphal apex of f i lamentous fungi may play a similar
endocyt ic role, and may also be part of a vacuolar
pa thway for transport of molecules between the apical
and basal cells of the hypha (Shepherd et al. 1993a,
Ashford and Orlovich 1994). A key c o m m o n factor o f
the animal and fungal systems is that exchange of
compar tmenta l content can occur via narrow, sporad-
ical ly-forming membranous tubules, and not just by
vesicle traffic (Swanson et al. 1987a, Hopkins et al.
1990, Knapp and Swanson 1990, Shepherd et al.
1993b). In animal cells, the f requency of tubule for-
mat ion and general motil i ty o f the endosomal system
vary in response to changes in internal pH, endocyt ic
load, and uptake of f luorochromes and toxic com-
pounds (Swanson e ta l . 1987a, b; Heuser 1989;
Knapp and Swanson 1990). These responses may be
part o f how these cells adjust to changing environ- mental conditions. Tubule- forming fungal vacuole
systems might similarly be expected to respond to
changes in internal and external condit ions and a study of this phenomenon may help us to determine if
the vacuolar system plays a role in the response of
fungi to environmental fluctuations, such as those that
occur during growth and survival in soil. Also, it
should further test the comparabi l i ty o f these fungal
vacuole systems to animal endosomal / lysosomal
systems. Finally, proper evaluat ion o f the effects o f 6- carboxyf luorescein diacetate (CFDA) is called for,
since to date nearly all the data relating to vacuoles in

86 G. J. Hyde and A. E. Ashford: Vacuole motility in Pisolithus tinctorius
l iv ing hyphae (and inc identa l ly also an imal endo-
somes) have resul ted from the use of f luorochromes.
We have used DIC and f luorescence optics to study
how exposure to C F D A and "control" solut ions
affects the mot i le vacuolar system in apical cells of
the bas id iomycete fungus Pisolithus tinctorius. This
work fol lows some pre l iminary observat ions which
indicated that use of differential in terference contrast
(DIC) optics would permi t v isua l i sa t ion of the mot i le
vacuole system in non loaded cells (Shepherd et al.
1993b). We have also examined vacuolar activity in
the previous ly uns tudied basal cells of P. tinctorius
hyphae. We now present more comprehens ive data
which show that all major types of m o v e m e n t seen in
apical vacuoles of CFDA- loaded cells also occur in
non loaded cells. In both loaded and non loaded basal
cells, the vacuolar sys tem differs f rom that of apical
cells, but still exhibits tubule format ion and other
moti le activities which suggest some role in long-
dis tance transport. These results clearly establish that
mot i le activities of the P. tinctorius vacuole system
are nei ther artifacts of f luorochrome loading nor
restricted to the hyphal apex. Al though the mot i le
vacuole system occurs in non loaded hyphae, expo-
sure to C F D A solut ion under appropriate condi t ions
does nevertheless t rans ient ly increase its moti le
activities. The effects are not, however, due to CFDA,
since they can also be brought about by exposure of
hyphae to control solut ions wi thout the f luorochrome.
Material and methods Fungal material, loading of vacuoles, and fluorescence microscopy
The basidiomycete fungus Pisolithus tinctorius (Pers.) Coker and Couch, strain DI-15, isolated by Grenville et al. (1986) was cultured on modified Melin-Norkrans (MMN) agar as previously described (Shepherd et al. 1993b).
Loading hyphal vacuoles with fluorochrome
Hyphal vacuoles were loaded with 6-carboxyfluorescein diaeetate (CFDA; Molecular Probes, Eugene, OR) in one of two ways. In the first method, a wedge of MMN agar was cut from the colony margin and was placed in an embryo cup containing 20 btg of CFDA per ml in water purified by reverse osmosis (hereafter referred to as water) for 15-30 min, then rinsed several times over a period of 20 rain in water, or in some instances, liquid MMN medium. The rinsed wedge was then squashed under a coverslip in water, and left for 30-40 min to allow the hyphae to "recover" before observation. In the second method, a wedge cut from the colony margin was squashed directly onto a slide with water (or in some instances, liquid MMN) and left for 30-40 min. Solution containing 20 gg of CFDA per ml in water was then drawn under the coverslip to replace the water. Some colo- nies that were loaded with CFDA by the first method were mounted directly on slides without coverslips, for viewing with a • 0.9
N.A. Zeiss water-immersion objective on a Zeiss Axiophot micro- scope.
Nonloaded hyphae
Hyphae were prepared for observation as in the second CFDA-load- ing method described above, but without the introduction of CFDA under the coverslip. They were observed either (1) directly after mounting, (2) at the end of a 30-40 min "recovery" period following mounting, or (3) after a fresh solution of water had been introduced under the coverslip, following the recovery period.
Microscopy and videomicroscopy
Most CFDA-loaded hyphae were observed using a X63, 1.4 N.A. objective on a Zeiss Axiophot microscope fitted with DIC and epi- fluorescence optics. For observation of CFDA fluorescence a fluo- rescein filter block (BP 450--490, FT 510 and BP 515-585) was used. Vacuolar activity was recorded using a real-time digital imaging set- up comprising an ImagePoint camera (Photometrics, Tucson, AZ), a PCI-compatible LG3 framegrabber (Scion Corp., Frederick, MD), a Scion version of NIH Image (public domain image analysis soft- ware), and a Macintosh 9500 computer. Using this set-up, sequences of images of vacuolar activity could be captured at 25 frames per second. Individual images from these sequences were arranged using CorelDraw 5 and printed on a Phaser 440 dye-sublimation printer (Tektronix, Wilsonville, OR).
Estimation of vacuolar activity
To estimate the relative extent and activity of the vacuolar system in the apical or basal cells of loaded and nonloaded hyphae two methods were used. For apical cells, a portion of the colony margin was systematically scanned and the percentage of apical cells con- taining at least one narrow, longitudinally-oriented vacuolar tubule was calculated. A narrow tubule was one which was less than half the width of the hypha for most of the tubule's length. For basal cells, a region of the colony approximately 200-600 ~m back from the mar- gin was scanned, and the percentage of hyphae with motile "large" vacuoles was calculated. A "large" vacuole was one more than half, and usually nearly all, the width of the hypha for most of its length. The definition of large vacuolar "motility" is given under Results. When scoring basal cells before and after a treatment, the same region of the colony was scanned. The mean percentages and their SEM (of three colony wedges, at least 50 hyphae counted per wedge) were calculated using the arcsine transformation (Zar 1974).
Results
Types of vacuolar motility in loaded and nonloaded
hyphae
Apical ceils
Our observat ions of the vacuolar sys tem in the apical
cells of CF D A - l oa de d hyphae of P. tinctorius agree
with those previous ly reported for these cells of this
o rganism (Shepherd et al. 1993b). Most of the various
types of mot i l i ty were apparent, inc lud ing the rapid
extens ion and fus ion of tubules (Fig. 1 A - D ) and
peris tals is- l ike waves along the length of exis t ing
tubules (Fig. 2 B). The r ight -hand tubule shown in
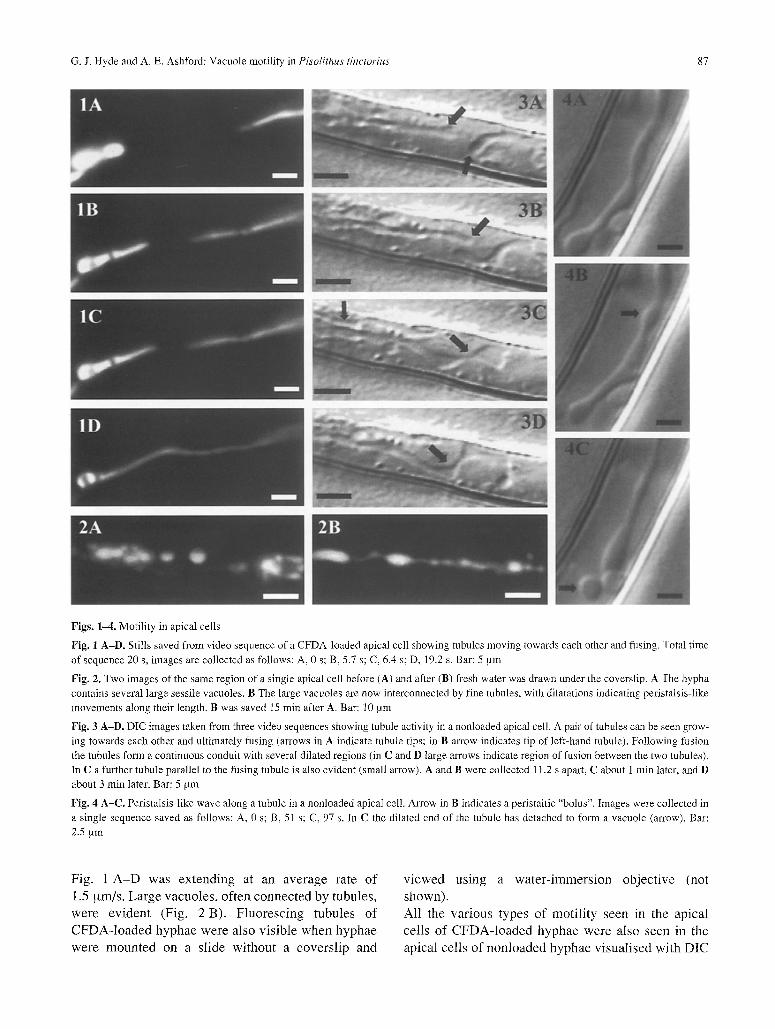
G. J. Hyde and A. E. Ashford: Vacuole motility in Pisolithus tinctorius 87
Figs. 1-4. Motility in apical cells
Fig. 1 A-D. Stills saved from video sequence of a CFDA-loaded apical cell showing tubules moving towards each other and fusing. Total time of sequence 20 s, images are collected as follows: A, 0 s; B, 5.7 s; C, 6.4 s; D, 19.2 s. Bar: 5 am
Fig. 2. Two images of the same region of a single apical cell before (A) and after (B) fresh water was drawn under the coverslip. A The hypha contains several large sessile vacuoles. B The large vacuoles are now interconnected by fine tubules, with dilatations indicating peristalsis-like movements along their length. B was saved 15 min after A. Bar: 10 btm
Fig. 3 A-D. Die images taken from three video sequences showing tubule activity in a nonloaded apical cell. A pair of tubules can be seen grow- ing towards each other and ultimately fusing (arrows in A indicate tubule tips; in B arrow indicates tip of left-hand tubule). Following fusion the tubules form a continuous conduit with several dilated regions (in C and D large arrows indicate region of fusion between the two tubules). In C a further tubule parallel to the fusing tubule is also evident (small arrow). A and B were collected 11.2 s apart, C about 1 min later, and D about 3 min later. Bar: 5 ~tm
Fig. 4 A-C. Peristalsis-like wave along a tubule in a nonloaded apical cell. Arrow in B indicates a peristaltic "bolus". Images were collected in a single sequence saved as follows: A, 0 s; B, 51 s; C, 97 s. In C the dilated end of the tubule has detached to form a vacuole (arrow). Bar: 2.5 ,am
Fig . 1 A - D w a s e x t e n d i n g at a n a v e r a g e r a t e o f
1.5 g m / s . L a r g e v a c u o l e s , o f t e n c o n n e c t e d b y t u b u l e s ,
w e r e e v i d e n t (F ig . 2 B ) . F l u o r e s c i n g t u b u l e s o f
C F D A - l o a d e d h y p h a e w e r e a l so v i s i b l e w h e n h y p h a e
w e r e m o u n t e d o n a s l i de w i t h o u t a c o v e r s l i p a n d
v i e w e d u s i n g a w a t e r - i m m e r s i o n o b j e c t i v e ( n o t
s h o w n ) .
A l l t h e v a r i o u s t y p e s o f m o t i l i t y s e e n in t h e a p i c a l
ce l l s o f C F D A - l o a d e d h y p h a e w e r e a l so s e e n in the
a p i c a l c e l l s o f n o n l o a d e d h y p h a e v i s u a l i s e d w i t h D I C
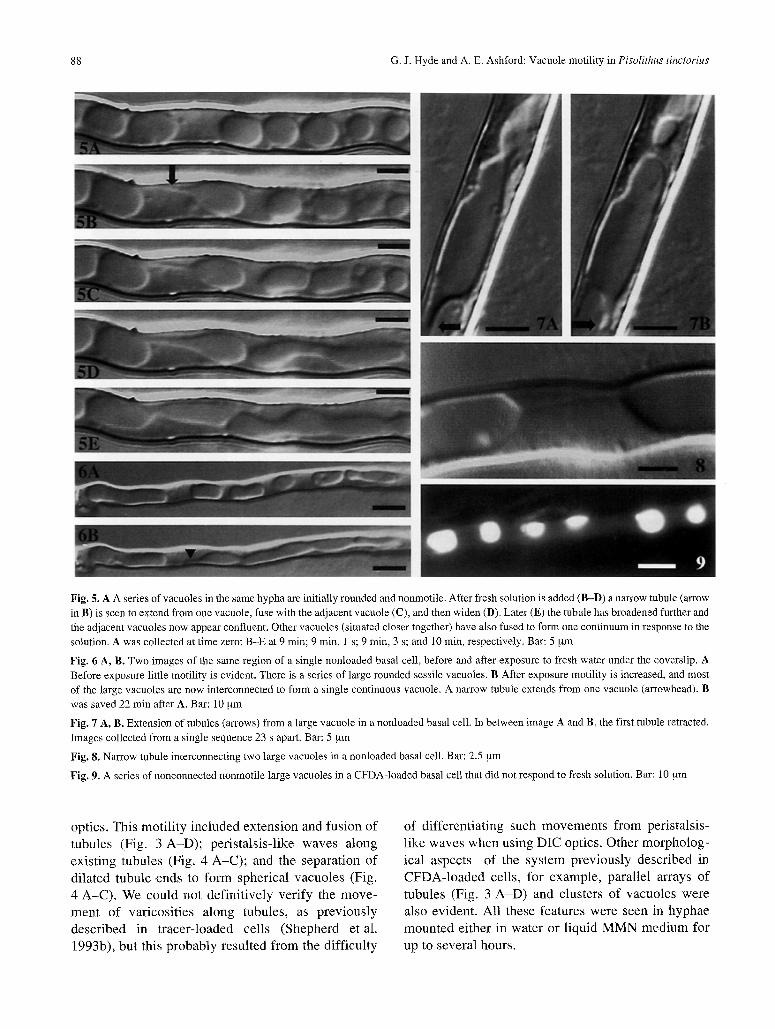
88 G.J. Hyde and A. E. Ashford: Vacuole motility in Pisolithus tinctorius
Fig. 5. A A series of vacuoles in the same hypha are initially rounded and nonmotile. After fresh solution is added (B-D) a narrow tubule (arrow in B) is seen to extend from one vacuole, fuse with the adjacent vacuole (C), and then widen (D). Later (E) the tubule has broadened further and the adjacent vacuoles now appear confluent. Other vacuoles (situated closer together) have also fused to form one continuum in response to the solution. A was collected at time zero; B-E at 9 min; 9 min, 1 s; 9 min, 3 s; and 10 min, respectively. Bar: 5 gm
Fig. 6 A, B. Two images of the same region of a single nonloaded basal cell, before and after exposure to fresh water under the coverslip. A Before exposure little motility is evident. There is a series of large rounded sessile vacuoles. B After exposure motility is increased, and most of the large vacuoles are now interconnected to form a single continuous vacuole. A narrow tubule extends from one vacuole (arrowhead). B was saved 22 min after A. Bar: 10 gm
Fig. 7 A, B. Extension of tubules (arrows) from a large vacuole in a nonloaded basal cell. In between image A and B, the first tubule retracted. Images collected from a single sequence 23 s apart. Bar: 5 ~xm
Fig. 8. Narrow tubule interconnecting two large vacuoles in a nonloaded basal cell. Bar: 2.5 gm
Fig. 9. A series of nonconnected nonmotile large vacuoles in a CFDA-loaded basal cell that did not respond to fresh solution. Bar: 10 ~tm
opt ics . Th i s m o t i l i t y i n c l u d e d e x t e n s i o n and f u s i o n o f
t ubu l e s (Fig. 3 A - D ) ; pe r i s t a l s i s - l i ke w a v e s a long
e x i s t i n g t ubu l e s (Fig. 4 A - C ) ; and the s e p a r a t i o n o f
d i l a ted t u b u l e - e n d s to f o r m spher ica l v a c u o l e s (Fig.
4 A - C ) . W e c ou ld n o t d e f i n i t i v e l y ve r i fy the m o v e -
m e n t o f va r i cos i t i e s a l ong tubu le s , as p r e v i o u s l y
d e s c r i b e d in t r a ce r - l oaded cel ls ( S h e p h e r d et al.
1993b) , b u t this p r o b a b l y r e su l t ed f r o m the d i f f i cu l ty
of d i f f e r en t i a t i ng such m o v e m e n t s f r o m per i s ta l s i s -
l ike w a v e s w h e n u s i n g D I C opt ics . O the r m o r p h o l o g -
ica l aspects o f the s y s t e m p r e v i o u s l y d e s c r i b e d in
C F D A - l o a d e d cel ls , for e x a m p l e , pa ra l l e l a r rays of
t ubu l e s (Fig. 3 A - D ) and c lus te r s o f v a c u o l e s were
also ev iden t . A l l these fea tures were s een in h y p h a e
m o u n t e d e i ther in wa te r or l i qu id M M N m e d i u m for
up to severa l hours .
G. J. Hyde and A. E. Ashford: Vacuole motility in Pisolithus tinctorius
Basal cells
In basal cells, at a distance between 200 and 600 gm behind the colony margin, the vacuolar elements are typically much broader, most often being the full width of the cell (Figs. 5-8). While these vacuoles were sometimes quiescent (Figs. 5 A, 6 A, and 9), in other cases various types of motility were evident. These included the extension of narrow tubules from the large vacuoles (Figs. 5 B, C, 6 B, and 7 A, B) and the interconnection of large vacuoles by narrow tubules (Figs. 5 C and 8). Often the originally narrow connection expanded to a much broader one, thus converting a series of vacuoles into one long large element (Fig. 5 C-E). Other movements of the vac- uoles themselves ranged from "ruffling" of the vacuo- lar edges to rhythmic fluctuations involving the whole vacuole. All types of motility observed were seen in both loaded and nonloaded cells.
Effects of exposure to fresh water or CFDA solution
Apical cells
Exposing apical cells to fresh CFDA solution typical- ly resulted in an obvious and dramatic increase in activity of the vacuolar system. This effect, however, does not appear to be due to CFDA itself, since a similar effect was caused by exposing loaded (Fig. 2 A, B) or nonloaded hyphae to fresh water alone. Further evidence that CFDA was not a stimulant to vacuolar activity was provided by a comparison of motility in loaded and nonloaded colonies that had been left to recover for 30-40 rain. We calculated the mean percentage of apical cells in loaded and non-
89
loaded colonies that contained at least one narrow longitudinal tubule (Table 1). There were no signifi- cant differences between treatments.
Basal cells
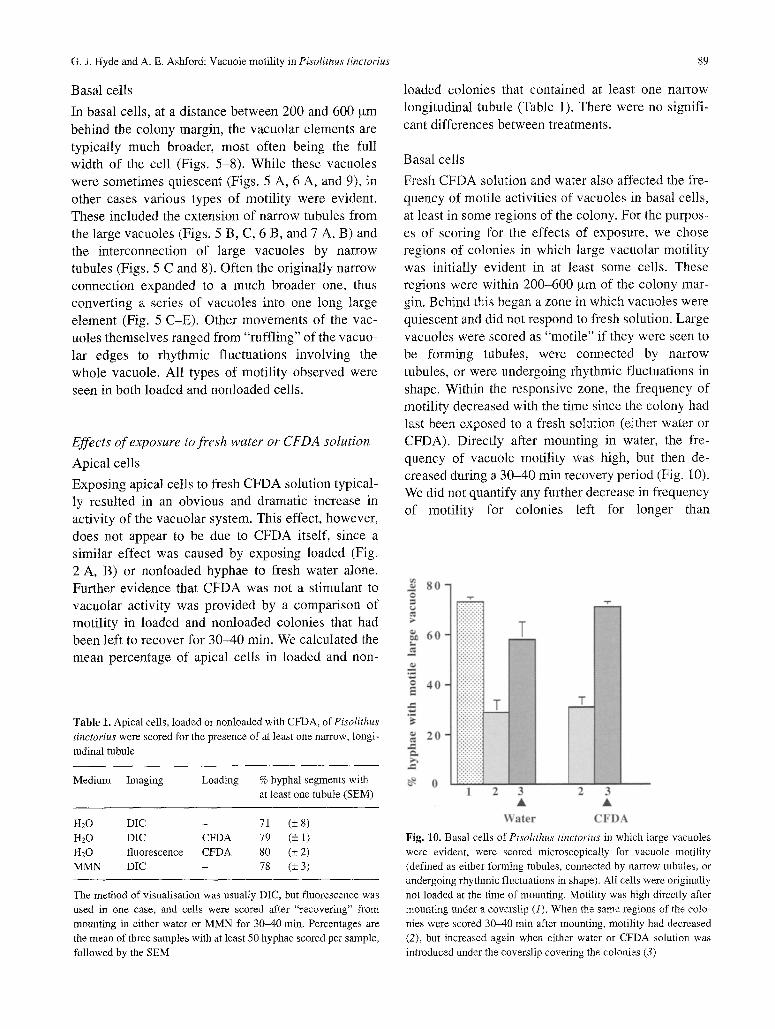
Fresh CFDA solution and water also affected the fre- quency of motile activities of vacuoles in basal cells, at least in some regions of the colony. For the purpos- es of scoring for the effects of exposure, we chose regions of colonies in which large vacuolar motility was initially evident in at least some cells. These regions were within 200-600 ,urn of the colony mar- gin. Behind this began a zone in which vacuoles were quiescent and did not respond to fresh solution. Large vacuoles were scored as "motile" if they were seen to be forming tubules, were connected by narrow tubules, or were undergoing rhythmic fluctuations in shape. Within the responsive zone, the frequency of motility decreased with the time since the colony had last been exposed to a fresh solution (either water or CFDA). Directly after mounting in water, the fre- quency of vacuole motility was high, but then de- creased during a 30-40 min recovery period (Fig. 10). We did not quantify any further decrease in frequency of motility for colonies left for longer than
Table 1. Apical cells, loaded or nonloaded with CFDA, of Pisolithus
tinctorius were scored for the presence of at least one narrow, longi- tudinal tubule
Medium Imaging Loading % hyphal segments with at least one tubule (SEM)
H20 DIC - 71 (+ 8) H20 DIC CFDA 79 (_+ 1) H20 fluorescence CFDA 80 (+ 2) MMN DIC - 78 (_+ 3)
The method of visualisation was usually DIC, but fluorescence was used in one case, and cells were scored after "recovering" from mounting in either water or MMN for 30-40 min, Percentages are the mean of three samples with at least 50 hyphae scored per sample, followed by the SEM
Fig. 10. Basal cells of Pisolithus tinctorius in which large vacuoles were evident, were scored microscopically for vacuole motility (defined as either forming tubules, connected by narrow tubules, or undergoing rhythmic fluctuations in shape). All cells were originally not loaded at the time of mounting. Motility was high directly after mounting under a coverslip (1). When the same regions of the colo- nies were scored 30-40 min after mounting, motility had decreased (2), but increased again when either water or CFDA solution was introduced under the coverslip covering the colonies (3)

90 G.J. Hyde and A. E. Ashford: Vacuole motility in Pisolithus tinctorius
30-40 min, but we have observed that the vacuolar system retains considerable motility even if colonies are kept on a coverslip-covered slide for many hours. When a colony that had been scored after a 30-40 min recovery period was then exposed, under the coverslip, to fresh water, and rescored, frequency of motility increased to a level nearly as high as that seen after mounting (Fig. 10). Increased frequency of motility within a defined region was also brought about by exposure, under the coverslip, to CFDA solution (Fig. 10). The stimulatory effect of fresh water or CFDA solu- tion upon both loaded and nonloaded vacuoles of basal cells was also observed in individually identi- fied cells (Figs. 5 and 6). The types of motility shown by vacuoles that had been stimulated to increased activity were the same as those exhibited by vacuoles that were already motile prior to exposure to fresh solution. The responses to exposure often occurred rapidly, as soon as cells could be observed after expo- sure (within a minute), while on other occasions they developed more gradually over a period of several minutes. Some quiescent vacuoles, in both the responsive and nonresponsive zones of the colony, remained inactive after exposure to fresh solution. This was not due to a lack of contact between these vacuoles and the fresh solution because (1) in cells exposed to CFDA, many large vacuoles became highly fluorescent but remained nonmotile (Fig. 9); (2) some cells would respond to exposure while other adjacent cells would not.
Discussion
The results show that vacuoles in basal hyphal cells exhibit many of the motile activities previously demonstrated only for apical cells, and that vacuoles of both cell types can increase their motile activity and interconnectedness in response to changing envi- ronmental conditions. These new findings indicate first that vacuoles take a much more active role in maintaining cellular equilibrium than was formerly assumed; and second, they attest to the potential of the vacuolar system to operate as a long-distance transport pathway. We further argue that the motile activities of P. tinctorius vacuoles are unaffected by loading with the fluorescent probe CFDA, and con- sider the significance of this finding in relationship to the effects of fluorochrome loading in animal endoso- mal/lysosomal systems.
Vacuolar motility in P. tinctorius is not an artefact
of CFDA- loading
Most aspects of tubular vacuole behaviour in apical hyphal cells described by Shepherd et al. (1993a, b), such as tubule extension, fusion, peristalsis-like waves, and separation of dilated tubule ends to form small vacuoles, were also observed with DIC optics in nonloaded cells. Thus, the tubular form, interconnect- edness, pleiomorphic behaviour and overall motility of the vacuolar system in cells of P. tinctorius are not a consequence of loading the system with fluorescent tracers. It was more difficult to assess whether the vacuolar system had an equivalent morphological complexity in nonloaded hyphae to that demonstrated with CFDA, since DIC is not well-suited to visualis- ing extensive three-dimensional arrays. Nevertheless, there were no obvious differences in the appearance of vacuoles in apical cells in loaded and nonloaded P. tinctorius colonies, nor were there differences in the number of hyphae with at least one narrow tubule. Our results also show that vacuolar motility is not a consequence of either UV irradiation or mounting the hyphae under a coverslip. Confirmation that the motile tubular vacuole system is constitutive in fungal hyphae and not induced by fluorochromes has significant implications for the study of motility in endosomal/lysosomal systems. Fungal vacuoles provide a valuable model for com- parison with the animal systems because they are relatively large and they exhibit, more than any other tubule-forming system, an equivalent range of motile activities. The key papers describing the morphology and motility of endosomes/lysosomes in living ani- mal cells have all relied upon loading with various fluorochromes. However, different fluorochromes affect the system differently and give different results. For example, lysosomes of macrophages are tubulo- reticular when loaded with Lucifer Yellow, but spher- ical with Acridine Orange (Knapp and Swanson 1990). The possibility that tubules per se might be an artefact of tracer-loading is, however, strongly argued against by their widespread occurrence and associa- tion with vacuoles or vesicles in animal, plant, and fungal cells that have been freeze-substituted (e.g., Hoch and Howard 1980, McCully and Canny 1985, Robinson et al. 1986, Roberson and Fuller 1988). Likewise, the results of the present paper support the idea that the dynamic behaviour of endosomal/lyso- somal tubular systems seen in tracer-loaded cells is not induced by the methods used to study them.

G. J. Hyde and A. E. Ashford: Vacuole motility in Pisolithus tinctorius 91
Increased vacuolar motility: survival in a changing environment
Soil-dwelling hyphae such as P. tinctorius constantly encounter new environmental conditions either by virtue of their growth or as a result of changes in the environment itself. Changing levels of soil nutrients, water potential, pH and toxic compounds will lead to variations in hyphal growth rate and viability. The results provide evidence that altered vacuolar activity is an early hyphal response to small environmental changes. Specifically, vacuolar motility increased transiently when hyphae were exposed to fresh solu- tions. We have not defined what was responsible for the altered vacuole motility, although it would seem most likely that it involves the washing-away of some "inhibitory" factor that accumulates around the hypha. Recent work on animal cell endosomal/lyso- somal systems has indicated that their tubule-forming activity increases with endocytic lead, variations in pH, and osmotic pressure (Heuser 1989, Knapp and Swanson 1990). It is likely that greater vacuole (or lysosomal) activity in fungal and animal cells enables the cell more rapidly to sequester compounds entering the cytoplasm from outside, to neutralize the effects of changing pH, or to deal with an increase in the level of metabolic by-products. This first evidence of the ability of the vacuolar system to respond to altered external conditions strengthens the argument that it is analogous to the endosomal/lysosomal systems of animal cells. It also illustrates how mistaken was the old conception of the vacuolar system as little more than a passive dump site. In the maintenance of cellu- lar equilibrium vacuoles and their animal analogues clearly play a primary and active role.
Support for a role for the fungal vacuolar system in long-distance transport
The observations that vacuoles in the more basal cells of P. tinctorius show motility and may be inter- connected over several microns are particularly important in supporting a role for vacuoles in long- distance transport in fungal hyphae. Previous work on the activity of the vacuole system in P. tinctorius has shown that vacuole-derived tubules can interconnect the apical and penultimate cells by penetrating the small gap in the septum that separates these cells (Shepherd et al. 1993a). This has raised speculation that the vacuole system might be one hyphal transport pathway by which nutrients may travel, in this mycorrhizal fungus, between foraging hyphae in the
soil and the root/fungus interface (Shepherd et al. 1993a). A comparable transport function has recently been proposed for the vacuolar-tubular network of chickpea trichomes, in which adjacent cells are inter- connected by vacuolar protrusions through plasmod- esmata (Lazzaro and Thomson 1996). For long- distance transport to occur in hyphae, vacuolar inter- connections must be present in all cells along the pathway between the penultimate cells and those at the interface with the root epidermis. We have docu- mented evidence that for at least some distance behind the penultimate cell, vacuoles within cells are sporadically interconnected and thus perhaps serve as a functional continuum over time. We do not at this stage think our inability to stimulate vacuolar motili- ty in cells that lie a long way from the penultimate cell disproves the long-distance transport model. It is like- ly that any long-distance transport system will only operate fully when hyphae are actively transferring nutrients from the mycorrhizal interface to the hyphal tip, which did not occur under the growth conditions of this study. Further work is needed to examine vac- uolar activity in mycorrhizal fungi growing in associ- ation with plant roots.
Acknowledgements The work was supported by an Australian Research Council grant awarded to AEA and a Research Fellowship awarded to GJH. The authors are grateful to Dr. Candy Briggs for her assistance. We would also like to thank Drs. Lara Perasso and Louise Cole for stim- ulating discussions.
References Ashford AE, Orlovich DA (1994) Vacuole transport, phosphorus and
endosomes in the growing tips of fungal hyphae. In: Stephenson AG, Kao T-H (eds) Pollen-pistil interactions and pollen tube growth. American Society of Plant Physiologists, Rockville, MD, pp 135-149 (Current topics of plant physiology, vol 12)
Grenville DJ, Peterson RL, Ashford AE (1986) Synthesis in growth pouches of mycorrhizae between Eucalyptus pilularis and sever- al strains of Pisolithus tinctorius. Aust J Bot 34:95-102
Heuser J (1989) Changes in lysosome shape and distribution corre- lated with changes in cytoplasmic pH. J Cell BioI 108:855-864
Hoch HC, Howard RJ (1980) Ultrastructure of freeze substituted hyphae of the basidiomycete Laetisaria arvalis. Protoplasma 103:281-297
Hopkins CR, Gibson A, Shipman M, Miller K (1990) Movement of internalized ligand-receptor complexes along a continuous endo- somal reticulum. Nature 346:335-339
Knapp P, Swanson J (l 990) Plasticity of the tubular lysosomal com- partment in macrophages. J Cell Sci 95 :433439
Lazzaro MD, Thomson WW (1996) The vacuolar-tubular continuum

92
in living trichomes of chickpea (Citer arietinurn) provides a rapid means of solute delivery from base to tip. Protoplasma 193:181-190
McCully ME, Canny MJ (1985) The stabilization of labile configu- rations of plant cytoplasm by freeze-substitution. J Microsc 139: 27-33
Roberson RW, Fuller MS (1988) Ultrastructural aspects of the hyphal tip of Sclerotium rolfsii preserved by freeze substitution. Protoplasma 146:143-149
Robinson JM, Okada T, Castellot JJ Jr, Karnovsky MJ (1986) Unusual lysosomes in aortic smooth muscle cells: presence in living and rapidly frozen cells. J Cell Biol 102:1615-1622
Shepherd VA, Orlovich DA, Ashford AE (1993a) Cell-to-cell trans- port via motile tubules in growing hyphae of a fungus. J Cell Sci 105:1171-1178
G, J. Hyde and A. E. Ashford: Vacuole motility in Pisolithus tinctorius
- - (1993b) A dynamic continuum of pleiomorphic tubules and vacuoles in growing hyphae of a fungus. J Cell Sci 104: 495- 5O7
Swanson J, Burke E, Silverstein SC (1987a) Tubular lysosomes accompany stimulated pinocytosis in macrophages. J Cell Biol 104:1217-1222 Bushnell A, Silverstein SC (1987b) Tubular lysosome morpho- logy and distribution within macrophages depend on the integri- ty of cytoplasmic microtubules. Proc Nail Acad Sci USA 84: 1921-1925
van Deurs B, Petersen OW, Olsnes S, Sandvig K (1989) The ways of endocytosis. Int Rev Cytol 117:131-177
Zar JH (1974) Biostatistical analysis. Prentice Hall, Englewood Cliffs, NJ






















