Embed Size (px)
Citation preview

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA
ECOLOGIA, MANEJO E CONSERVAÇÃO DE Pentaclethra macroloba (Willd.) Kuntze (Fabaceae) EM FLORESTA DE VÁRZEA
ESTUARINA DO RIO AMAZONAS
ADELSON ROCHA DANTAS
Manaus, Amazonas Fevereiro, 2021

I
ADELSON ROCHA DANTAS
ECOLOGIA, MANEJO E CONSERVAÇÃO DE Pentaclethra macroloba (Willd.) Kuntze (Fabaceae) EM FLORESTA DE VÁRZEA
ESTUARINA DO RIO AMAZONAS
Orientadora: Dra. Maria Teresa Fernandez Piedade Coorientador: Dr. Marcelino Carneiro Guedes
Tese apresentada ao Programa de Pós-graduação em Ecologia do Instituto Nacional de Pesquisas da Amazônia como parte dos requisitos para obtenção do título de Doutor em Biologia (Ecologia).
Manaus, Amazonas Fevereiro, 2021

II

III

IV
©SEDAB/INPA - Ficha Catalográfica Automática gerada com dados fornecidos pelo(a) autor(a)
Bibliotecário responsável: Jorge Luiz Cativo Alauzo - CRB11/908
D192e Dantas, Adelson Rocha Ecologia, manejo e conservação de Pentaclethra macroloba (Willd.) Kuntze (Fabaceae) em floresta de várzea estuarina do Rio Amazonas / Adelson Rocha Dantas; orientadora Dra. Maria Teresa Fernandez Piedade; coorientador Dr. Marcelino Carneiro Guedes. --Manaus:[s.l], 2021. 147 f.
Tese (Doutorado - Programa de Pós Graduação em Ecologia) -- Coordenação do Programa de Pós-Graduação, INPA, 2021.
1. Adaptações morfológicas. 2. Fenologia. 3. Visitantes florais. 4. Produção de sementes. 5. História natural. I. Piedade, Dra. Maria Teresa Fernandez, orient. II. Guedes, Dr. Marcelino Carneiro, coorient. III. Título. CDD: 598

V
Sinopse:
Estudou-se um amplo espectro da história natural de Pentaclethra macroloba e suas relações com o ambiente de várzea do estuário amazônico. Foram avaliados a estrutura morfológica, distribuição geográfica, fenologia, demografia e padrões de crescimento desta espécie, para entender a sua dominância na Região Amazônica e subsidiar planos de conservação e manejo desta árvore de múltiplos usos não madeireiros.
Palavras-chave: 1. Pracaxi. 2. Hiperdominante. 3. Adaptação. 4. Inundação diária. 5. Anéis de crescimento. 6. Espécie pioneira. 7. Longa vida. 9. Dispersão. 10. Predação de sementes. 11. Fenologia.

VI
Agradecimentos
A Deus por me fortalecer e nunca me deixar desistir dos meus objetivos.
À Dra. Maitê por ter aceitado me orientar, por sempre me incentivar, por compartilhar
seu conhecimento e por ter me acolhido muito bem no INPA e no grupo MAUA. Além
de orientadora, ela é uma grande amiga que quero levar para a vida.
Ao Dr. Marcelino pela amizade e por sempre estar presente na minha vida científica.
À coordenadora do projeto Kamukaia III, Dra. Ana Cláudia, pelo apoio financeiro e
pela amizade.
À Embrapa Amapá por dar suporte à realização desta tese.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico pela bolsa de
doutorado.
Ao Grupo Ecologia, Monitoramento e Uso Sustentável de Áreas Úmidas
(MAUA/PELD/CNPq/FAPEAM) pelo suporte à análise de dados desta pesquisa.
Ao Programa de Pós-graduação em Ecologia, em nome da coordenadora Dra.
Noemia Ishikawa, pela oportunidade de cursar o doutorado neste prestigiado
programa.
Ao professor Dr. Jochen Schöngart pela maravilhosa disciplina “Dendroecologia de
Florestas Tropicais” e por ter compartilhado seu conhecimento no capítulo quatro
desta tese.
Aos professores do Programa de Pós-graduação em Ecologia por todo conhecimento
compartilhado com os alunos.
À turma de mestrado de 2016 pelos momentos de aprendizagem e pela amizade.
Aos técnicos da Embrapa Amapá (Paulo André, Enoque Lima, Jonas Viana e Adjalma
Souza) pelo auxílio em campo.
Aos estudantes de Engenharia Florestal da UEAP (Marcos, Isabelly e Gabriel), IMMES
(Tailon, Gabriel, Jony e Pedro) e da UnB (Cinthia) por darem suporte ao inventário de
100% dos pracaxizeiros na APA da Fazendinha.
À Caroline Vasconcelos por sempre me acolher muito bem em sua casa quando eu
precisava ir à Manaus.
Aos mestres em biodiversidade tropical (UNIFAP) Felipe e Hugo pela ajuda na
instalação das cercas coletoras de sementes (nesta atividade, agradeço também ao
mestrando em Ciências Ambientais – UNIFAP- Ediglei) e na instalação das 30 bandas
dendrométricas nos pracaxizeiros do Campo Experimental do Mazagão.

VII
Aos colegas da sala de estudo da pós-graduação (Ana Carolina, Aldine, Hugo, Felipe,
Haroldo e Rafael) da Embrapa Amapá pelos momentos de descontração.
Aos agentes ambientais Nerivan e Hermeson pelo suporte que deram na realização
das coletas de campo na APA da Fazendinha.
Agradeço imensamente a minha mãe Osvaldina, meu pai Ademir, minha irmã Cleidi e
meus irmãos Adenilson e Adriano por sempre me incentivarem nos estudos.
Muito obrigado!

VIII

IX
Ecologia, manejo e conservação de Pentaclethra macroloba (Willd.) Kuntze (Fabaceae) em floresta de várzea estuarina do Rio Amazonas Resumo Pentaclethra macroloba (Willd.) Kuntze (pracaxi) é uma árvore hiperdominante e de múltiplas utilidades. Pouco se conhece sobre a história natural de P. macroloba na Amazônia, o que é preocupante, pois essa espécie é largamente explorada no estuário amazônico para extração do óleo medicinal das suas sementes. Nós estudamos um amplo espectro da autoecologia de P. macroloba para auxiliar em planos de manejo e conservação dos seus recursos e no entendimento de sua dominância na várzea estuarina no nordeste da Amazônia. As árvores adultas têm estruturas morfológicas adaptadas em resposta a inundação diária, como raízes adventícias para aumentar a eficiência respiratória, e lenticelas no tronco que servem como ponto de conexão entre a raiz e a copa para o transporte de oxigênio. Os frutos secos favorecem a ativação do mecanismo de deiscência explosiva, permitindo que as sementes sejam expelidas a longas distâncias. O formato deltoide das sementes permite maior eficiência na dispersão hidrocórica. A plântula é hipógea fanerocotiledonar com um par de cotilédones de reserva, fonte extra de energia para a plântula fugir da inundação. A taxa de germinação foi de 78% e a velocidade foi 0,2 sementes dia-1. O método circular buffer revelou a presença de 123 populações de P. macroloba distribuídas em um raio de 5.699.943 km² na região Neotropical. P. macroloba possui floração altamente sincronizada (Z = 0.70) no período de estiagem (botões florais: βtemperatura =1.30, p <0,01 e flores em antese: βtemperatura =1,84; p <0,001). Frutos imaturos aparecem no período de estiagem (βtemperatura =0,67; p <0,01) e amadurecem no período chuvoso. As sementes são dispersas durante o período de chuvas (βprecipitação =0.0051; p <0.01) e de cheia do Rio Amazonas (βinundação =0.12; p <0.001). Vespas, abelhas e formigas foram os principais visitantes florais. 33 árvores produziram 261 frutos (9 ± 8 frutos) e 380 sementes (54 ± 50 sementes). A taxa de predação foi de 5% (18 sementes), com a maioria das sementes predadas por larvas de mariposa 50%. No período de produção de sementes, o crescimento diamétrico de P. macroloba desacelera (r = -0,76; F = 12,7; p < 0,01); o número de sementes aumenta no período de inundação da floresta (r = 0,90; F = 40,3; p < 0,001). Em uma área de 136,59 ha, foram registradas 2.072 árvores adultas (15 indivíduos.ha-1). Indivíduos regenerantes apresentaram padrão de distribuição diamétrica exponencial negativa e os adultos padrão log-normal. A altura média dos regenerantes foi 0,63 m (± 0,55 m) e dos adultos 12,7 m (± 5,4 m). Regenerantes e adultos apresentam padrão espacial agregado. A análise dos anéis de crescimento de 38 árvores revelou média de idade de 60 anos (± 20 anos), com idade mínima de 28 anos e máxima de 102 anos. Bandas dendrométricas instaladas no tronco de árvores em diferentes topografias, mostraram que na várzea baixa ocorre estagnação do crescimento durante o período chuvoso (R2 = 0,41; t = -2,62; p < 0,01) e de maior cheia do rio Amazonas (R2 = 0,47; t = -3,01; p < 0,01), sendo que formação de anéis de crescimento pode estar associada com a sazonalidade desses fenômenos. A alta adaptação de P. macroloba com a sazonalidade climática e com a maré diária do Rio Amazonas, explicam a sua alta dominância e adaptabilidade no estuário amazônico. O resultado desta pesquisa será fundamental para buscar políticas públicas para o bom manejo e conservação dos pracaxis estuarinos e para fortalecer a cadeia produtiva dos produtos não madeireiros como geradora de renda e de inclusão social das populações ribeirinhas ao longo do Rio Amazonas.

X
Ecology, management, and conservation of Pentaclethra macroloba (Willd.) Kuntze (Fabaceae) in estuarine floodplain forest of the Amazon River Abstract Pentaclethra macroloba (Willd.) Kuntze (pracaxi) is a species of multiple utilities and hyperdominant in the Amazon estuary. Little is known about the natural history of P. macroloba in the Amazon region, which is worrying, as this species is largely exploited in the Amazon estuary to extract the medicinal oil from its seeds. We studied a broad aspect of P. macroloba autoecology to assist in management and conservation plans of its resources and in understanding its dominance in the estuarine floodplain of Northeast Amazonia. Adult individuals have adapted structures in response to daily flood, such as adventitious roots to increase respiratory efficiency and lenticels in the trunk, serving as a connection point for oxygen transport between the root and the aerial part. Dried fruit favor the activation of the explosive dehiscence mechanism, allowing the seeds to reach long distances. The deltoid shape of the seeds allows their fluctuation and more efficient dispersal. The seedlings are phanerocotylar hypogeal with a pair of reserve cotyledons that provide an extra source of energy for the seedlings to escape the flood. Germination rate was 78 % and the speed was 0.2 seeds.day-1. Circular buffer method revealed the presence of 123 subpopulations of P. macroloba distributed in a radius of 5.699.943 km² across the Neotropical region. Flowering was synchronized in the dry season (flower buds: βtemperature =1.30, p <0.01 and flowers in anthesis: βtemperature =1.84; p <0.001). Immature fruits appeared during the dry season (βtemperature =0.67; p <0.01) and ripen in the rainy season. Seeds were dispersed during the rainfall (βrainfall =0.0051; p <0.01) and flooding of the river (βwater =0.12; p <0.001). There was no relationship between leaf change and hydrometeorological variables. The main floral visitors were wasps, bees and ants. 33 trees produced 261 fruits (9 ± 8 fruits) and 380 seeds (54 ± 50 seeds). The predation rate was 5% (18 seeds), with most of the seeds predated by moth larvae (50%). During the period of seed production, the diametric growth of P. macroloba decelerates (r = -0.76; F = 12.7; p < 0.01) and the number of seeds increases during the period of forest flooding (r = 0.90; F = 40.3; p < 0.001). In an area of 136.59 ha, 2,072 adult trees (15 individuals ha-1) were registered. The regenerants showed negative exponential pattern of diameter distribution and adult trees have a log-normal pattern. The mean height of the regenerants was 0.63 m (± 0.55 m) and adults 12.7 m (± 5.4 m). Regenerants and adults had aggregate spatial pattern. The analysis of the growth rings of 38 trees revealed mean age of 60 years (± 20 years), with minimum age of 28 years and maximum of 102 years. Dendrometric bands installed on the tree trunk in different topographies, showed that in the low floodplain growth stagnation occurred during the rainy season (R2 = 0.41; t = -2.62; p < 0.01) and higher water levels of the river (R2 = 0.47; t = -3.01; p <0.01), the formation of growth rings may be associated with the seasonality of these phenomena. The high synchrony of P. macroloba with climatic seasonality and with the daily tide of the Amazon River, explain its high dominance and adaptability in the Amazon estuary. The result of this research will be fundamental to seek public policies for the good management and conservation of estuarine P. macroloba trees and to strengthen the productive chain of non-wood products as a generator of income and social inclusion of riverside populations along the Amazon River.

XI
Sumário
Lista de figuras ........................................................................................................ XIII
Introdução geral .......................................................................................................... 1
Objetivo Geral ............................................................................................................. 6
Objetivos específicos .................................................................................................. 6
Capítulo 1: Morphology, germination, and geographic distribution of Pentaclethra macroloba (Fabaceae): hyperdominant Amazonian tree
Introduction.................................................................................................................. 9
Materials and methods .............................................................................................. 11
Results ...................................................................................................................... 13
Discussion ................................................................................................................. 22
Conclusions ............................................................................................................... 26
References ................................................................................................................ 27
Capítulo 2: Phenological behavior and floral visitors of Pentaclethra macroloba, a hyperdominant tree in the Brazilian Amazon River estuary
Introduction................................................................................................................ 36
Materials and methods .............................................................................................. 39
Results ...................................................................................................................... 44
Discussion ................................................................................................................. 53
Conclusions ............................................................................................................... 59
References ................................................................................................................ 60
Capítulo 3: Produção e predação de frutos e sementes de Pentaclethra macroloba (Fabaceae) em floresta de várzea do estuário do Rio Amazonas
Introdução ................................................................................................................. 72
Material e metódos .................................................................................................... 75
Resultados ................................................................................................................ 78
Discussão .................................................................................................................. 82
Conclusões................................................................................................................ 87
Referências ............................................................................................................... 88
Capítulo 4: Demographic structure and growth pattern of Pentaclethra macroloba (Willd.) Kuntze in the floodplain forest of the Amazon River estuary
Introduction................................................................................................................ 99
Materials and methods ............................................................................................ 101
Results .................................................................................................................... 108
Discussion ............................................................................................................... 115

XII
Conclusions ............................................................................................................. 119
References .............................................................................................................. 120
Síntese .................................................................................................................... 127
Referências bibliográficas ....................................................................................... 128

XIII
Lista de figuras
Capítulo 1
Fig. 1. Morphological characteristics of adult individuals of Pentaclethra macroloba in the Amazon estuary: A. Pentaclethra macroloba tree, B. Straight trunk, C. Protuberant lenticels on the trunk, D. Moss and lichen stain, E. Red phloem and yellow sapwood, F. Base of the trunk, G. Spread of branches at the base of the trunk, H. Adventitious roots and I. Exsudate on the trunk. ............................................................................ 13
Fig. 2. Morphological characteristics of the Pentaclethra macroloba leaf in the Amazon estuary: A. Ripe leaf, B. New leaf, C. Alternate spiral phyllotaxy, D. Petiole channeled with trichomes and E. Pulvinus base stipule.............................................................. 14
Fig. 3. Morphological characteristics of the Pentaclethra macroloba inflorescence in the Amazon estuary: A. Inflorescences in the phase of floral bud and anthesis, B. Immature flower buds, C. Pentamer flower with their guiding lines [gl] of pollinators, D. Gomosepalous calyx and E. Gland [g] in the anther. ................................................ 15
Fig. 4. Morphometric relationships of fruits and seeds of Pentaclethra macroloba in the Amazon estuary. ....................................................................................................... 17
Fig. 5. Morphological characteristics of fruits and seeds of Pentaclethra macroloba in the Amazon estuary: A. Immature fruits, B. Ripe fruit with its valves, C. Ripe seeds, D. Seed embryo and E. Seeds floating in the water of the Amazon river. ...................... 18
Fig. 6. Maturation phases of fruits and seeds of Pentaclethra macroloba in the Amazon estuary. Evolution of shape, size and colour: phase A. 22nd day after flower fertilization; phase B. 32nd day; phase C. 53rd day; phase D. 78th day; phase E. 110th day; phase F. 144th day; and phase G. 198th day of maturation. ............................. 19
Fig. 7. Phases of seedling development of Pentaclethra macroloba in the Amazon estuary: phase A. Primary root (pr) formation; phase B. Opening of the cotyledons (cot) and formation of secondary roots (sr); phase C. Expansion of the epicotyl (epi); phase D. Development of the first leaves (prot) and formation of cataphyll (cat) in the epicotyl; and phase E. Ripe leaflets (fo) and apical bud (ag). .................................................. 20
Fig. 8. Morphological characteristics of Pentaclethra macroloba seedling in the Amazon estuary: A. Phyllotaxy of the subsequent leaves, B. Stipule at the base of the pulvinus and C. Gland at the lateral base of the pulvinus. ......................................... 21
Fig. 9. Dispersion area (left) and number of subpopulations (right) of Pentaclethra macroloba in the Neotropical region (Data source: tropicos.org and splink.cria.org.br). .................................................................................................................................. 22
Capítulo 2
Fig. 1: Fazendinha Environmental Protection Area (APA) in the floodplain forest of the Amazon estuary, Macapá, Brazil. .............................................................................. 39
Fig. 2: Rainfall ( ), average maximum temperature ( ) and average tidal amplitude of the Amazon River ( ) in the floodplain forest of the Amazon estuary, Macapá, Brazil. Source: (a) INMET - National Institute of Meteorology; (b) Brazilian Navy Hydrography Center (Marinha do Brasil). ......................................................... 45

XIV
Fig. 3: Intensity of flower buds ( ) and flowers in anthesis ( ) phenophases of Pentaclethra macroloba, in relation to the water level ( ) in the floodplain forest of the Amazon estuary, Macapá, Brazil. ........................................................................ 46
Fig. 4: Relationship between the proportion of individuals of Pentaclethra macroloba emitting flower buds ( ) and flowers in anthesis ( ), with (a) rainfall, (b) water level and (c) maximum temperature in the floodplain forest of the Amazon estuary, Macapá, Brazil. .......................................................................................................... 47
Fig. 4: Development of inflorescence and floral longevity of Pentaclethra macroloba: Phase A: formation of peduncle and rachis of the inflorescence; Phase B: Appearance of the first flower buds; Phase C: expansion and maturation of flower buds; Phase D: Floral anthesis; and Phase E: floral senescence. ...................................................... 48
Fig. 5: Intensity of immature fruit ( ), ripe fruit ( ) and seeds dispersal ( ) phenophases of Pentaclethra macroloba, in relation to the water level ( ) in the floodplain forest of the Amazon estuary, Macapá, Brazil. .......................................... 49
Fig. 6: Relationship between the proportion of individuals of Pentaclethra macroloba emitting with immature fruits ( ), ripe fruits ( ) and seeds dispersal ( ), with (a) rainfall, (b) water level and (c) maximum temperature in the floodplain forest of the Amazon estuary, Macapá, Brazil. .............................................................................. 51
Fig. 7: Intensity of leaf flush ( ), ripe mature leaf ( ) and leaf fall ( ) phenophases of Pentaclethra macroloba, in relation to the water level ( ) in the floodplain forest of the Amazon estuary, Macapá, Brazil. .......................................... 52
Fig. 8: Foraging activity of the main floral visitors, between the hours of 07:00 to 17:30, during 31 days of observation in Pentaclethra macroloba in the floodplain forest of the Amazon estuary, Macapá, Brazil. .............................................................................. 55
Capítulo 3
Figura 1: Sementes de Pentaclethra macroloba coletada na floresta de várzea do estuário amazônico. Comprimento (L), largura (W) e espessura (T). ........................ 77
Figura 2: Relações entre a produção de sementes de Pentaclethra macroloba com o crescimento diamétrico (a) e a altura de inundação (b) em floresta de várzea do estuário amazônico. .................................................................................................. 80
Figura 3: Variabilidade no comprimento (a), largura (b), espessura (c) e peso fresco (d) de sementes de três populações de Pentaclethra macroloba ao longo do estuário amazônico. ................................................................................................................ 81
Figura 4: Predação por larvas de mariposa (A e B), por roedores (C) e por fungos (D) em sementes de Pentaclethra macroloba em floresta de várzea do estuário amazônico. ................................................................................................................ 82
Capítulo 4
Fig.1 Two populations of Pentaclethra macroloba located in the Fazendinha Environmental Protection Area (APA da Fazendinha) and in the Mazagão Experimental Field (CEM), Amazon estuary region. ................................................ 101

XV
Fig.2 Anatomical structure of the growth rings, formed by bands of marginal parenchyma, of Pentaclethra macroloba in the Amazon estuary. ........................... 106
Fig.3 Diameter (a) and height (b) distribution pattern of the regeneration and adult trees of a population of Pentaclethra macroloba in the Amazon estuary. ................ 109
Fig.4 Spatial distribution pattern of adult trees (population [a, b] and productive individuals [c, d]) and regeneration (parcel 1 [e, f] and parcel 2 [g, h]) of a population of Pentaclethra macroloba in the Amazon estuary. ................................................. 110
Fig.5 Age structure of regeneration and adult trees (reproductive trees red bars) of a population of Pentaclethra macroloba in the Amazon estuary. ............................... 111
Fig.6 Growth patterns of Pentaclethra macroloba during its life trajectory in the Amazon estuary: (a) direct growth, (b) one release, (c) one suppression and (d) multiple events of release and suppression. .................................................................................... 112
Fig.7 Growth patterns of Pentaclethra macroloba during its life trajectory in the Amazon estuary: accumulated diameter (a), relationship between height and diameter (b), and increment in diameter (c), height (d) and biomass (e). Current annual increment (CAI) and mean annual increment (MAI). ......................................................................... 113
Fig.8 Relationships between flood level in the interior of the forest (a), monthly rainfall (b) and mean monthly maximum temperature (c) with the diameter increment of Pentaclethra macroloba in high floodplain (HV) and low floodplain (LV) in the Amazon estuary. ................................................................................................................... 115

1
Introdução geral
As florestas de várzeas são ecossistemas inundáveis da Amazônia que
ocorrem ao longo do Rio Amazonas/Solimões e seus tributários de águas brancas,
com pH próximo à neutralidade e alta carga sedimentar devido à sua origem nas
regiões andinas e pré-andinas, geologicamente mais recentes (Junk et al. 2011). Por
serem áreas de maior fertilidade (Sioli 1968), são também as mais exploradas pelas
populações ribeirinhas, podendo propiciar, se bem manejadas, diversos serviços
ecossistêmicos essenciais (Junk e Piedade 2000). Essas áreas regulam a
biogeoquímica, a ecologia e a hidrologia da bacia amazônica, pois são importantes
fontes de metano e dióxido de carbono (Sawakuchi et al. 2014), desempenhando um
papel importante no balanço de carbono orgânico regional devido às elevadas taxas
de produção de plantas aquáticas (Melack et al. 2009). A heterogeneidade
hidrogeomorfológica das várzeas resulta em um mosaico de paisagens conectadas
por manchas de florestas, lagos, meandros e rios, proporcionando uma grande
diversidade de habitats para o estabelecimento da biota terrestre e aquática (Pinedo-
Vasquez e Sears 2011, Junk et al. 2012, 2018, Wittmann et al. 2013).
Com uma área de cobertura de cerca de 450.000 km² (Wittmann e Junk 2016),
os ecossistemas de várzea ocupam aproximadamente dois terços das áreas alagáveis
amazônicas, sendo cerca de 70% de sua área coberta por florestas (Wittmann et al.
2010b). Na região estuarina do Rio Amazonas e de seus tributários de águas brancas,
as florestas de várzea perfazem aproximadamente 199.281 km² (Junk e Piedade
2010). No estado do Amapá, essas florestas compreendem 4,85% da superfície total
do Estado, sendo, após a floresta de terra firme, o segundo maior ecossistema
florestado da região, em termos de estrutura, diversidade e representatividade
espacial (Rabelo 2005). Sua maior área de abrangência e concentração ocorre,
principalmente, na margem esquerda da orla do Estado (Rabelo 2005). Devido às
inundações diárias, e principalmente àquelas resultantes da inundação anual, as
várzeas estuarinas recebem grande carga de material sedimentar que lhes confere
fertilidade adicional (Latrubesse e Franzinelli 2002, McClain e Naiman 2008, Meade
2008, Wittmann et al. 2010a).
O fenômeno anual de subida e descida do nível das águas, denominado de
pulso de inundação (Junk et al. 1989), ocorre regularmente ao longo dos grandes rios
da Amazônia (Piedade et al. 2015). Na Amazônia Central, o pico da inundação

2
acontece entre os meses de junho e julho, seguido pela descida das águas nos meses
subsequentes, com o pico das secas entre os meses de novembro e janeiro,
caracterizando um pulso de inundação monomodal (Junk et al. 2014). Nessa região,
a fase aquática pode durar até 243 dias e chegar a uma altura de até 6 m no tronco
das árvores (Schöngart et al. 2002). Por outro lado, no estuário amazônico, o pulso
de inundação é diário ou polimodal (Junk et al. 2014), com marés semi-diurnas
(Rabelo 2000), podendo a coluna de água alcançar até 4 m de altura no período
chuvoso na foz do Rio Amazonas (Cunha et al. 2017).
As florestas de várzea são detentoras de uma importante biodiversidade
arbórea conforme apontado por Wittmann et al. (2006), que registraram mais de 900
espécies de árvores tolerantes à inundação nesses ecossistemas da Bacia
Amazônica. Os autores concluem que as florestas de várzea amazônicas são mais
biodiversas dentre as florestas inundáveis do mundo. A diversidade arbórea das
várzeas é utilizada para múltiplos propósitos (Junk 2000, Pinedo-Vasquez e Sears
2011) e sustenta uma indústria pesqueira altamente produtiva (Junk 2000). As árvores
desses ambientes são uma importante fonte de recursos para a maioria da população
rural da Amazônia (Wittmann et al. 2010b), fornecendo uma variedade de produtos
florestais madeireiros e não madeireiros utilizados para diversos fins. Investigando o
uso de 186 espécies arbóreas de floresta de várzea, Wittmann et al. (2010b)
verificaram que do total de espécies amostradas, 135 (72,6%) são utilizadas pela
população ribeirinha, sendo que 49 dessas espécies (36,3%) são usadas para uma
única finalidade e a grande maioria é utilizada para múltiplos propósitos. Dentre as
espécies estudadas, 98 oferecem produtos não madeireiros, uma grande parte dos
quais é utilizada na medicina popular.
Diante da grande biodiversidade que caracteriza a várzea, se fazem
necessários estudos com enfoque conservacionista e de sustentabilidade, para que
os recursos sejam geridos respeitando o ambiente. As pesquisas voltadas para as
espécies de múltiplas utilidades são um grande incentivo para o manejo e
conservação desses ecossistemas, pois várias alternativas de uso podem contribuir
para a renovação cíclica das espécies de interesse. Além disso, a extração de
produtos não madeireiros pode proporcionar uma renda extra para as comunidades
ribeirinhas que vivem da floresta. Isso confere vantagens a favor desse tipo de uso,
pois a obtenção de vários produtos durante todo o ano é mais vantajosa do que o
corte da árvore.

3
O manejo sustentável das florestas de várzea é importante para que se possa
assegurar as suas múltiplas funções, os seus recursos naturais e seus serviços
ecossistêmicos, possibilitando o crescimento sustentável da população humana na
Amazônia (Schöngart 2008). No entanto, é necessário aprofundar as investigações
sobre as espécies arbóreas desse ecossistema, para que se conheça seu potencial e
a sua capacidade reprodutiva, principalmente das espécies que sofrem maiores
pressões antrópicas ou daquelas que possuem grande potencial para a exploração.
Dentre as espécies com elevado potencial não madeireiro para exploração nas
florestas de várzea estuarinas, destaca-se a Pentaclethra macroloba (Willd.) Kuntze,
conhecida popularmente como “pracaxi” (Dantas et al. 2017). A espécie pertence à
família Fabaceae, tem em média 14 m de altura e o diâmetro pode chegar até 59 cm.
Duas características chamam atenção para identificação rápida da árvore em campo,
o tronco com manchas acinzentadas e esverdeadas, devido a proliferação de musgos
e liquens, e a propagação de ramos com folhas bipinadas na base do tronco. A
inflorescência é uma espiga terminal ou subterminal, com flores brancas que medem
de 4 a 5,5 mm de comprimento (Pesce 2009). Os frutos são do tipo vagem, secos,
deiscentes, de coloração verde, tornando-se marrom escuro com a maturação. As
sementes são de coloração marrom escura, opaca, com depressões superficiais,
formando linhas salientes, às vezes reticuladas, próximas à base.
A espécie ocupa a decima posição no rank das árvores hiperdominantes na
região Amazônica (ter Steege et al. 2020) e apresenta alta densidade populacional
nas várzeas do estuário amapaense, sempre ocupando as primeiras posições em
densidade absoluta nas listas florísticas (Santos e Jardim 2006, Queiroz et al. 2007,
Carim e Jardim 2008). Hartshorn (1983) considera a existência de três grandes
populações de P. macroloba distribuídas na América Latina. A primeira população,
amplamente estudada, se concentra na Costa Rica, principalmente na reserva
florestal La Selva, onde domina a floresta pantanosa. A segunda população se
concentra na extensão do Panamá até a Colômbia. A terceira e última população se
encontra no nordeste da Amazônia brasileira, sendo sua história natural pouco
conhecida.
O pracaxi pode ser enquadrado no grupo ecológico das espécies esciófitas
parciais (Maciel et al. 2003), pois suas plântulas toleram sombra na fase inicial de
crescimento (Oberbauer e Strain 1985), porém, necessita de elevado grau de luz solar
da fase jovem até a maturidade (Maciel et al. 2003). Na região Amazônica, P.

4
macroloba pode ser encontrada habitando florestas de terra firme (Condé e Tonini
2013), várzea (Dantas et al. 2017) e igapó (Ferreira e Parolin 2007). Essa espécie
pode repor, com eficiência, o nitrogênio de solos degradados, devido a associação de
suas raízes com bactérias fixadoras de nitrogênio do gênero Frankia (Eaton et al.
2012). Nas florestas de várzea do estuário amazônico, onde o nitrogênio é limitante,
o pracaxi pode ser utilizado em consorcio com o açaizeiro (Euterpe oleracea Mart.),
com possíveis interações benéficas para a produção do fruto do açaí (Braga 2018),
produto economicamente importante na cadeia produtiva local. P. macroloba
apresenta interação mutualísticas com a formiga Paraponera clavata (Fabricius,
1775), na qual a formiga se abriga na planta e encontra alimento no nectário extrafloral
e, em troca, protege a árvore capturando os seus herbívoros (Bennett e Breed 1985).
O macerado da casca do tronco de P. macroloba possui saponinas terpenóides
com ação anti-hemorrágica contra o veneno de víboras do gênero Bothrops (Silva et
al. 2007). Seu grande potencial está no óleo proveniente das sementes que possui
propriedades medicinais. Estudos comprovam a eficácia do óleo para tratar
inflamações na pele, cicatrização de queimaduras e ulceras diabéticas (Simmons et
al. 2015, Banov et al. 2014). Os moradores da Ilha de Cotijuba, em Belém (Pará),
utilizam o óleo para o tratamento de enfermidades como dores musculares,
inflamações, picadas de cobra e até no tratamento do câncer (Crespi e Guerra 2013).
Segundo esses últimos autores, recentemente a coleta de sementes vem se
intensificando nesta ilha, devido ao interesse de multinacionais na utilização do óleo
em seus produtos.
O óleo de pracaxi contém alta concentração de ácidos graxos (principalmente
ácidos behênico, oleico, linoleico e lignocérico) que têm ação emoliente na pele (Costa
et al. 2014). Atualmente, é crescente o uso do óleo de pracaxi nas indústrias de
cosméticos. Universidades da Amazônia, como a Universidade Federal do Pará e a
Universidade Federal do Amazonas, vêm desenvolvendo cosméticos como loções,
cremes e lápis de olho a partir dos ácidos graxos presentes no óleo de pracaxi
(Oliveira et al. 2020). A multinacional francesa L’Oréal é a empresa que mais
patenteou produtos de beleza à base do óleo de pracaxi (Oliveira et al. 2020). Esse
óleo tem grande potencial para se tornar um biocombustível, apresentando um
rendimento bruto acima de 90% para a composição do biodiesel (Lima et al. 2017).
Embora na literatura existam vários trabalhos focando nas propriedades
medicinais do óleo de P. macroloba (Silva et al. 2005, Silva et al. 2007, Teixeira et al.

5
2012, Crespi e Guerra 2013, Shebitz et al. 2013, Banov et al. 2014, Costa et al. 2014),
poucos são os estudos sobre a ecologia da espécie (Hartshorn 1972, Oberbauer e
Strain 1985, Oberbauer et al. 1987, Shebitz e Eaton 2013) e esses são voltados para
a população de P. macroloba da Amarica Central, principalmente na Costa Rica, onde
a espécie forma agrupamento monodominante. Apesar do conhecimento escasso
sobre a ecologia e o ciclo de vida dessa espécie na Amazônia brasileira, sua semente
já é bastante explorada na região, o que demanda traçar com a maior brevidade ações
que propiciem a sustentabilidade de seu manejo e conservação.
Amplos estudos autoecológicos são fundamentais para nortear formas
adequadas de manejo e conservação, e para elucidar as estratégias de que P.
macroloba dispõe para se tornar hiperdominante na Amazônia e tolerar variados
gradientes de inundação nas várzeas da região. Neste estudo foi feita uma detalhada
caracterização morfológica de P. macroloba, desde a germinação das sementes até
a fase adulta da árvore, analisando ainda sua distribuição biogeográfica. Investigamos
o comportamento fenológico reprodutivo e vegetativo, bem como a atividade dos
visitantes florais de P. macroloba. Avaliamos a produção de frutos e sementes e os
principais predadores de sementes. Analisamos a estrutura demográfica de P.
macroloba e seus padrões de crescimento por meio da dendrocronologia. Discutimos
os dados com uma abordagem evolutiva e adaptativa frente às variáveis ambientais,
principalmente o pulso de inundação polimodal da várzea estuarina.

6
Objetivo Geral
Gerar bases científicas sobre a autoecologia de Pentaclethra macroloba, para
subsidiar o manejo e conservação desta espécie, bem como o entendimento de sua
elevada dominância na várzea do estuário amazônico, frente às marés diárias do Rio
Amazonas.
Objetivos específicos
Capítulo 1
1. Avaliar as estratégias adaptativas de P. macroloba, por meio da análise
da sua estrutura morfológica e de seu padrão de dispersão.
2. Determinar sua distribuição geográfica.
3. Definir parâmetros indicativos de maturação de frutos e sementes e a
época mais adequada para sua coleta.
Capítulo 2
1. Determinar o padrão, duração e período de maior intensidade das
fenofases reprodutivas e vegetativas de P. macroloba, relacionando
esses eventos às variáveis climáticas e hidrológicas da várzea estuarina.
2. Identificar os principais agentes polinizadores da espécie.
Capítulo 3
1. Determinar a taxa de predação e produção de sementes.
2. Avaliar os fatores ambientais e os atributos arbóreos relacionados com
a produção de sementes.
Capítulo 4
1. Determinar os padrões demográficos e as taxas de crescimento de P.
macroloba, e analisar os fatores relacionados a esses padrões durante
a trajetória de vida da espécie na várzea estuarina.
2. Determinar a idade mínima, máxima e média da população avaliada,
assim como a idade reprodutiva.

7
Capítulo 1
________________________________________ Dantas, A.R., Guedes, M.C., Vasconcelos, C.C., Isacksson, J.G.L., Pastana, D., Lira-Guedes, A.C & Piedade, M.T.F. Morphology, germination, and geographic distribution of Pentaclethra macroloba (Fabaceae): hyperdominant Amazonian tree. Published in Revista de Biología Tropical, 69(1), 181-196, DOI 10.15517/rbt.v69i1.43446.

8
Morphology, germination, and geographic distribution of Pentaclethra macroloba
(Fabaceae): hyperdominant Amazonian tree
Adelson Rocha Dantas1*, Marcelino Carneiro Guedes2, Caroline da Cruz Vasconcelos3, Jaynna
Gonar Lobo Isacksson4, Dayane Nathália Barbosa Pastana5, Ana Cláudia Lira-Guedes2 & Maria
Teresa Fernandez Piedade1
1. Programa de Pós-graduação em Ecologia, Instituto Nacional de Pesquisas da Amazônia,
Manaus, Brasil; [email protected], [email protected]
2. Departamento de Recursos Florestais, Empresa Brasileira de Pesquisa Agropecuária,
Macapá, Brasil; [email protected], [email protected]
3. Programa de Pós-graduação em Botânica, Instituto Nacional de Pesquisas da Amazônia,
Manaus, Brasil; [email protected]
4. Grupo de Pesquisa em Ecologia Florestal, Instituto de Desenvolvimento Sustentável
Mamirauá, Tefé, Brasil; [email protected]
5. Programa de Pós-graduação em Engenharia Florestal, Universidade Federal de Lavras,
Lavras, Brasil; [email protected]
* Correspondence
Abstract. Introduction: Pentaclethra macroloba is a hyperdominant tree in the Amazon
estuary of great socioeconomic importance for the region because the oil from its seeds is a
powerful herbal medicine. Objective: We aimed to characterize the morphological structure,
the morphological adaptations in response to the daily flooding of the Amazon estuary and the
biogeographic area of P. macroloba. Methods: Detailed description of the external
morphology, from germination to the adult tree, was obtained from individuals located in
floodplain forest, Northeast Amazonia. The occupation area and the geographical extension of
P. macroloba were determined from point of geographical coordinates of botanical samples
deposited in the digital collections of Mobot and SpeciesLink. Results: Adult individuals have
adapted structures in response to daily flooding, such as: adventitious roots to increase

9
respiratory efficiency and lenticels in the trunk, serving as a connection point for oxygen
transport between the root and the aerial part. Dried fruit favours the activation of the explosive
dehiscence mechanism, allowing the seed to be expelled long distance. Deltoid shape of the
seed allows water fluctuation and more efficient dispersal. Seedling is phanerocotylar hypogeal
and with one pair of reserve cotyledons that provide the seedling an extra source of energy to
escape the flood. Germination rate was 78 % and the speed was 0.2 seeds.day-1. Circular buffer
method revealed the presence of 123 subpopulations of P. macroloba distributed in a radius of
5 699 943 km² across the Neotropical region. Conclusions: Much of the morphological
structures of P. macroloba are adaptive and evolutionary responses to the periodically flooded
environment of the Amazon estuary, showing that these environments select the trees, best
adapted, to inhabit the flood. P. macroloba has a wide geographical area denotes the plasticity
of adapting to different environments, which may justify its monodominance in some regions.
Key words: morphological adaptation; tidal flooding; Neotropical tree; adventitious roots;
pracaxi oil; management, conservation.
Some tree species in the Amazon region have great dominance, occurring in a wide area,
others have isolated populations and their distribution is restricted to a specific environment
(ter Steege et al., 2013). The great challenge for many forest ecologists is to understand the
mechanisms and strategies that trees develop over time to colonize the most varied
environments in the Amazon rainforest.
In the Amazon floodplain, the periodically flooded environment can promote speciation
and adaptation processes in the trees that inhabit this environment. Various adaptations in the
external morphology of the plants are observed in response to the low availability of oxygen in
the rhizosphere and the constant erosion of the soil, such as high density of lenticels in the trunk
and the presence of roots and aerial structures: rhizophores, pneumatophores, haustoria and
buttresses (Almeida, Amaral, & Silva, 2004; Parolin, 2012).
In Central Amazonia, flooding in the forest can last up to 243 days and reach a height
of up to 6 m on the tree trunk (Schöngart, Piedade, Ludwigshausen, Horna, & Worbes, 2002).
Trees respond to this long period of flooding reducing metabolic activity, which in many cases
leads to leaf fall in the canopy (De Simone, Junk, & Schmidt, 2003) and formation of annual

10
rings on the trunk (Schöngart et al., 2004). However, in the Eastern Amazon, the knowledge of
the morphological and ecophysiological adaptations of tree species that grow under the regime
of daily flooding and low amplitude of the Amazon estuary (Junk et al., 2011) is still quite
modest.
The floodplains of the Amazon estuary have several endemic and dominant species as
Pentaclethra macroloba. Species of the Fabaceae family, popularly known by name “pracaxi”
(Dantas, Marangon, Guedes, Feliciano, & Lira-Guedes, 2017). This species is hyperdominant
Amazon (ter Steege et al., 2013) and large occurrence in Amazon estuary (Carim, Wittmann,
Piedade, Guimarães, & Tostes, 2016). Among the three populations of P. macroloba in
Neotropical America, little is known about the natural history of the population that lives in the
Amazon Basin (Hartshorn, 1983).
P. macroloba is an important non-timber species in the local socioeconomic, source of
income for several riverside Amazon, due to its multiple uses. The bark of the tree trunk has
triterpenoid saponins that have anti-haemorrhagic action against the venom of snakes of the
Bothrops genus (Silva et al., 2007). The oil extracted from the seeds is a powerful natural
medicine, which is very common riverside Amazon people to use this oil to treat muscle pain
and inflammation. The oil has a high concentration of fatty acids that moisturize the skin (Costa
et al., 2014), which is highly requested by the cosmetics industry as raw material.
Despite the great arboreal biodiversity of the Amazon, information on the basic
morphology of plant species is still lacking, difficult to understand the evolutionary adaptations
of trees to the environment. In addition, studies of plant morphology are important to ensure
the correct identification of trees and natural regeneration (Melo, Mendonça, & Mendes, 2004),
indispensable requirement in biodiversity inventory.
Dispersion patterns of the trees has great contribution to the diversification of the
Amazon rainforest (Gentry, 1981). The biogeographic patterns of trees are the result, in many
cases, of limited dispersion (Wiens, 2011) and reveal characteristics of its ecology and
evolutionary biology (Gaston & Fuller, 2009), important knowledge for conservation strategies
of species that have economic potential, such as P. macroloba.
In this study, we evaluated the morphological adaptations of P. macroloba in response
to the daily flooding of the Amazon estuary. We provide a detailed description of the external
morphology of P. macroloba and evaluate its biogeographic area to assist in the management
and conservation of this species.

11
MATERIALS AND METHODS
Study area: Botanical material was collected from individuals located in two floodplain areas:
Mazagão Experimental Field (CEM) and Fazendinha Environmental Protection Area (APA da
Fazendinha hereafter), State of Amapá, Brazil. CEM belongs to the Brazilian Agricultural
Research Corporation, located in the city of Mazagão (00°02’33” S & 51°15’24” W), it has
55.95 ha of floodplain forest and is bathed by the tributary of the Amazon River called “furo
do Mazagão”. The APA da Fazendinha is located in the city of Macapá (00º03’04.24” S &
51°07’42.72” W), it has 13 659 ha of floodplain forest and is directly bathed by the Amazon
River.
The climatic system of the region is Am (Köppen classification), tropical rainy (Alvares,
Stape, Sentelhas, Gonçalves, & Sparovek, 2013), and in the state of Amapá, the rainy season
begins in December, with the concentration of rainfall in the month of March where the peak
of precipitation is above 60 mm until the month of August. The dry season starts in September
when the monthly rainfall is below 60 mm (Vilhena, Silva, & Freitas, 2018). The soils of the
area are classified as typical Melanic Gleysols Ta Eutrophic, with predominance of silt. Both
the sediment and the soil are formed by smectite, illite, kaolinite, goethite, anatase and quartz
(Pinto, 2014).
The vegetation is classified as dense alluvial ombrophilous forest (IBGE, 2012), with
predominance of species Mora paraensis, Astrocaryum murumuru, P. macroloba , Carapa
guianensis and Virola surinamensis (Carim et al., 2016).
Data collection: Morphological description of the 34 adult trees (30 trees in APA and 4 trees
in CEM) occurred in the field, observing their characteristics from the trunk base towards the
top of the crown. After the description of the tree adults, fertile branches containing flowers,
fruits and seeds were collected with the help of a pruning stick of 10 m and the usual tree
climbing technique according to Laman (1995).
Branches were stored in moistened polyethylene bags and sent to the Embrapa Seed
Laboratory, where the structures were measured and morphological description was conducted,
through a specialized form with botanical terms. After the description, exsiccates were prepared
for deposit in the Embrapa Herbarium Amazônia Oriental - IAN, Belém, Pará (registration
numbers IAN192676 and IAN192677).

12
Morphological structures of the trunk, branches, leaves, flowers, fruits, seeds and
seedlings were recorded using a Canon® camera (Model EOS Rebel T2i). Small structures
were recorded using a Leica stereomicroscope (model EZ4D) with fixed camera.
The morphological stages of fruit and seed maturation were determined monitoring 30
individuals, during the period from April 2018 to March 2019 in the APA da Fazendinha. It
was possible to observe the development of the first fruits, as well as to determine the
maturation time and the morphological changes that occurred over time. The fruits that showed
any significant change in shape or colour in the crown were collected, measured, and
photographed. A chronological sequence of the maturation of fruits and seeds was built.
For the morphometric analysis, 54 fruits and 114 seeds were collected to obtain fresh
mass (g), length (mm), width (mm) and thickness (mm). Fruits and seeds were weighed on
analytical balance (Shimadzu® model AUW22OD, Japan), with accuracy of 0.001 g. The
measures of length, width and thickness were measured with a digital caliper (Carbografite®
model 150, accuracy of 0.01 mm).
To evaluate the germination process and seedling phases, 60 seeds (4 replicates x 15
seeds) were sown in plastic trays (45 x 28 x 7.5 cm), containing sand as substrate sterilized at
100 °C and vermiculite of medium particle size (1:1 ratio). The germination phases and seedling
formation were monitored every two days in a greenhouse, for a period of six months.
The dispersal pattern of the species was analyzed through the geographical coordinates
of the botanical samples deposited in the digital repositories of the Missouri Botanical Garden
(Tropicos, 2019) and SpeciesLink (CRIA, 2019).
Data analysis: All descriptions of the morphological characteristics of P. macroloba
followed a standardized form in botanical terms, based on botanical literature (Barroso, Morim,
Pioxoto, & Ichaso, 2004; Gonçalves & Lorenzi, 2011; Harris & Harris, 2001). Mean ± standard
deviation and coefficient of variation (CV) of the morphometric data were calculated. The
morphometric relationships between fruits and seeds were evaluated using simple linear
regression. The significance of the relationship was assessed by analysis of variance at 0.05 %
probability. The percentage of germinated seeds, average germination time and the germination
speed were determined (Maguire, 1962). Germinated seed was considered when the cotyledons
open emitting the epicotyl and the seedling phase begins with the expansion of the first pair of
leaves (Camargo, Ferraz, Mesquita, Santos, & Brum, 2008).

13
Geographic dispersal map of the species was prepared in the software QGIS 2.18. The
extent of occurrence (EOO), area of occupation (AOO) and number of subpopulations (IUCN,
2017) of P. macroloba were determined using the statistical package ConR (Dauby, 2019). The
number of subpopulations was estimated using the circular buffer method (Rivers, Bachman,
Meagher, Lughadha, & Brummitt, 2010). All statistical analysis were performed using the R
program (R core Team, 2019).
RESULTS
Description of adult trees in the field: P. macroloba (Fig. 1A) has an average height of 13 m
(maximum 37 m) and 26 cm of diameter (maximum 59 cm). The base of the trunk (Fig. 1F)
can be channeled or in buttresses, with the tree stem (Fig. 1B) straight or inclined and cylindrical
shape. Rhytidome (outer bark) has a rough texture and with a mixture of green and gray colors
(presence of lichen stains, Fig. 1D), with abundant elliptical and blackish green lenticels (Fig.
1C). The phloem (inner bark) is reddish and the sapwood is yellowish (Fig. 1E). The tree does
not show exudate when the rhytidome is injured However, one day after, the tree exudes a
colourless and viscous liquid (Fig. 1I). During the flood period of the Amazon estuary, P.
macroloba produces a large number of adventitious roots (Fig. 1H). Individuals often show
spread of branches at the base of the trunk (Fig. 1G).
Fig. 1. Morphological characteristics of adult individuals of Pentaclethra macroloba in the
Amazon estuary: A. Pentaclethra macroloba tree, B. Straight trunk, C. Protuberant lenticels on
the trunk, D. Moss and lichen stain, E. Red phloem and yellow sapwood, F. Base of the trunk,

14
G. Spread of branches at the base of the trunk, H. Adventitious roots and I. Exsudate on the
trunk.
The leaves (21.3 cm ± 4.7 cm) are bipinnate (Fig. 2A), paripinnate, with alternate spiral
phyllotaxy (Fig. 2C) and have 6-15 pairs of leaflets (8.6 cm ± 2.8 cm). When new, the leaves
have a light red colour (Fig. 2B) and when ripe, they have a dark green colour (Fig. 2A). The
leaflets (8.6 cm ± 2.8 cm) are opposite or sub-opposite, containing 28-45 pairs of pinnules. The
pinnules (11.7 mm ± 0.6 mm) are opposite, oblong or linear, with an obtuse/asymmetric base;
acute apex, mucronate and with the entire blade margin. The pinnules are coriaceous, dark green
on the adaxial side, light green abaxial and glabrous on both sides. The petiole (32.6 mm ± 10.6
mm) varies from yellowish green to blackish brown, smooth to slightly rough, channelled and
covered by abundant trichomes (Fig. 2D). The pulvinus (7.5 mm ± 1.4 mm) varies from dark
green to black, rough and with trichomes. The rachis (16.6 cm ± 5.8 cm) varies from yellowish
green to blackish brown colour, channelled and with trichomes similar to the petiole. The
pulvinule (1.7 mm ± 0.3 mm) is rough and varies from yellowish green to blackish brown
colour. The rachilla has yellowish green or blackish brown colour, smooth and with brownish
trichomes. Presence of a pair of linear stipules (Fig. 2E) at the base of the pulvinus.
Fig. 2. Morphological characteristics of the Pentaclethra macroloba leaf in the Amazon
estuary: A. Ripe leaf, B. New leaf, C. Alternate spiral phyllotaxy, D. Petiole channeled with
trichomes and E. Pulvinus base stipule.

15
The inflorescence (17.1 cm ± 3.9 cm) is a terminal spike of white colour, due to the
presence of numerous whitish staminodes (Fig. 3A). A spike can have up to 411 flowers. The
floral bud (4 mm ± 1.1 mm) is initially light green when immature (Fig. 3B), becoming light
green to light brown with maturation. The flower (7.2 mm ± 1.3 mm) is pentamer (Fig. 3C),
calyx gamosepalous (Fig. 3D) and presenting up to 10 staminodes. The stamen has a gland at
the tip of the anther (Fig. 3E). The petals have floral nectar guide that direct the pollinator into
the flower (Fig. 3C).
Fig. 3. Morphological characteristics of the Pentaclethra macroloba inflorescence in the
Amazon estuary: A. Inflorescences in the phase of floral bud and anthesis, B. Immature flower
buds, C. Pentamer flower with their guiding lines [gl] of pollinators, D. Gomosepalous calyx
and E. Gland [g] in the anther.
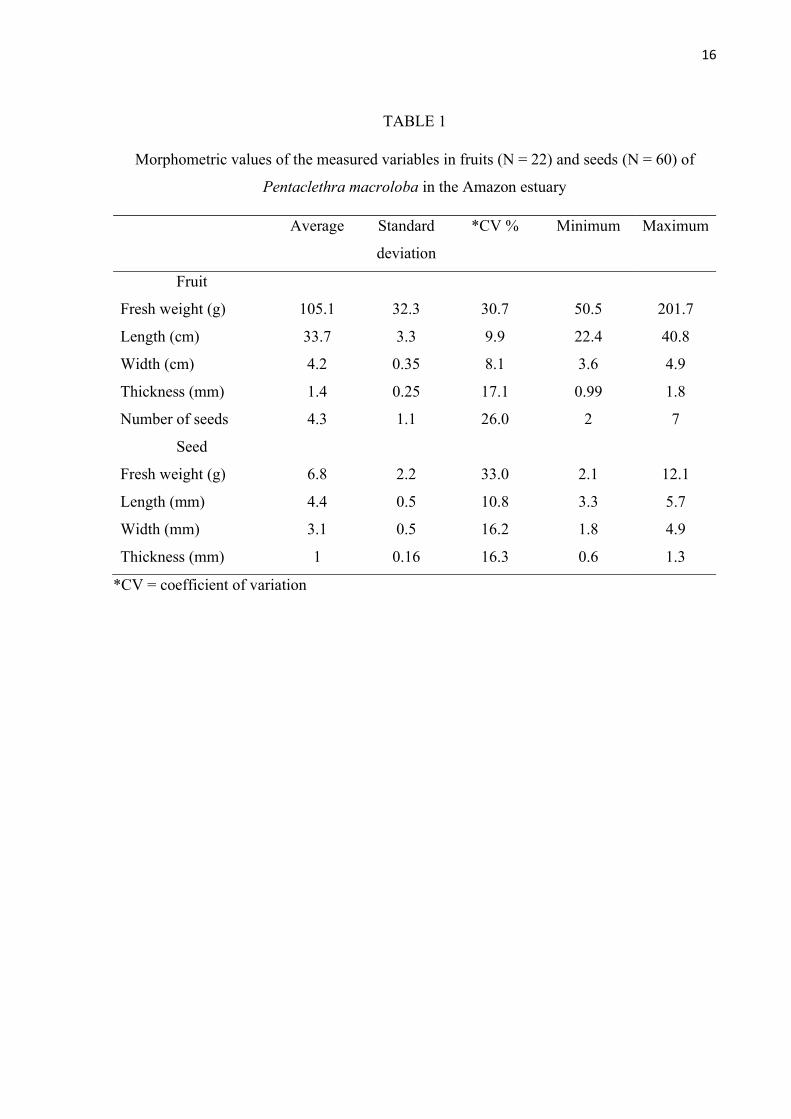
Morphometry and characterization of fruits and seeds: The fruit has an average length of
33.7 cm (± 3.3 cm) and weight of 105.1 g (± 32.3 g) (Table 1). The average number of seeds
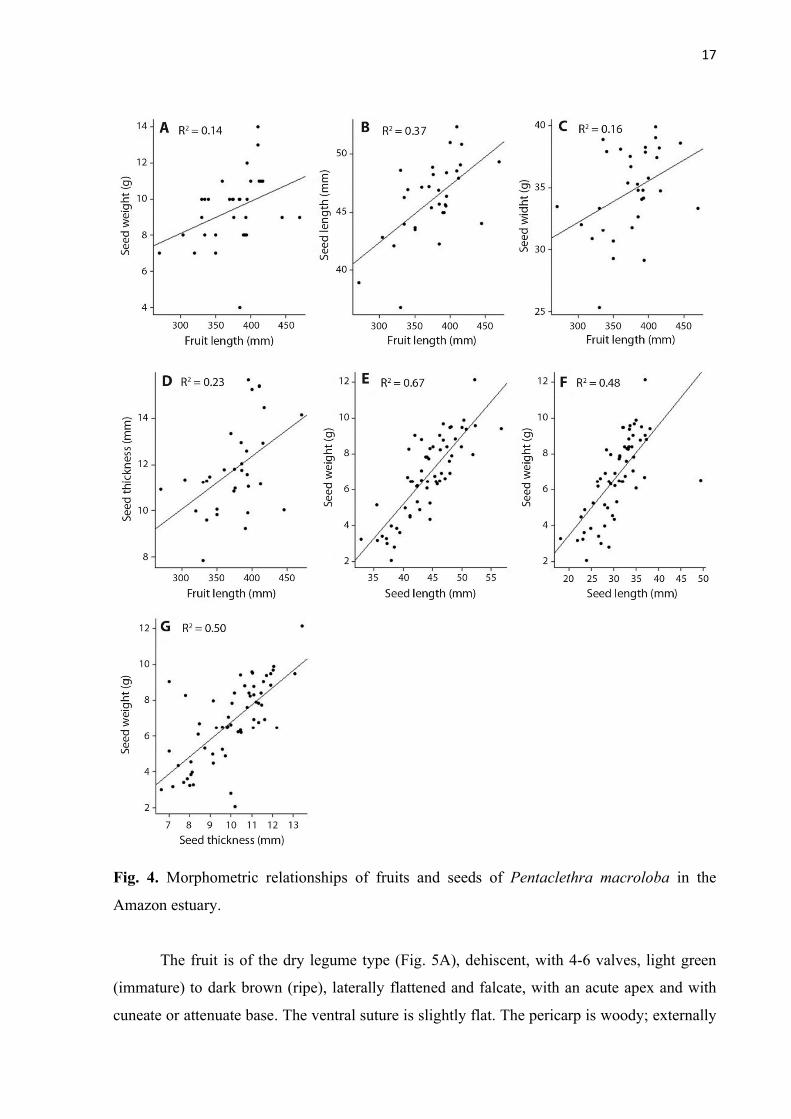
per fruit was four seeds (± 1 seed) (Table 1). Fruit length (mm) had a positive linear relationship
between the measures of fresh weight (F = 5.11; p = 0.031*; R² = 0.14; Fig. 4A), length (F =
17.48; p = 0.0002***; R² = 0.37; Fig. 4B), width (F = 59.2; p = 0.023*; R² = 0.16; Fig. 4C) and
thickness (F = 9.11; p = 0.005**; R² = 0.23; Fig. 4D) of the seeds.
The seed has an average length of 4.4 cm (± 0.48 cm) and weight of 6.8 g (± 2.2 g)
(Table 1). Fresh mass (g) of seeds has a positive linear relationship with its measures of length
(F = 116.23; p < 0.001***; R² = 0.67; Fig. 4E), width (F = 53.01; p < 0.001***; R² = 0.48; Fig.
4F) and thickness (F = 56.48; p < 0.001***; R² = 0.50; Fig. 4G).
16
TABLE 1
Morphometric values of the measured variables in fruits (N = 22) and seeds (N = 60) of
Pentaclethra macroloba in the Amazon estuary
Average Standard
deviation
*CV % Minimum Maximum
Fruit
Fresh weight (g) 105.1 32.3 30.7 50.5 201.7
Length (cm) 33.7 3.3 9.9 22.4 40.8
Width (cm) 4.2 0.35 8.1 3.6 4.9
Thickness (mm) 1.4 0.25 17.1 0.99 1.8
Number of seeds 4.3 1.1 26.0 2 7
Seed
Fresh weight (g) 6.8 2.2 33.0 2.1 12.1
Length (mm) 4.4 0.5 10.8 3.3 5.7
Width (mm) 3.1 0.5 16.2 1.8 4.9
Thickness (mm) 1 0.16 16.3 0.6 1.3
*CV = coefficient of variation
17
Fig. 4. Morphometric relationships of fruits and seeds of Pentaclethra macroloba in the
Amazon estuary.
The fruit is of the dry legume type (Fig. 5A), dehiscent, with 4-6 valves, light green
(immature) to dark brown (ripe), laterally flattened and falcate, with an acute apex and with
cuneate or attenuate base. The ventral suture is slightly flat. The pericarp is woody; externally

18
corrugated, glabrous, rare punctiform or elliptical lenticels; internally reddish brown, slightly
striate, glabrous and with diagonal cracks due to the natural twist for opening and releasing the
seeds.
The seed is deltoid, rhomboid or obovoid (Fig. 5B), with flat and irregular surface, dark
brown, opaque, striate and with thin grooves. The integument (< 1 mm) varies from light brown
to dark brown and opaque. Cotyledons are fleshy, light yellow to whitish colour and with
channels that exude oil of rapid oxidation when exposed to air. The seed embryo has a straight
embryonic axis, with well-developed plumule and radicle (Fig. 5C). The seeds have an air
pocket between the cotyledons, which allows floating in water (Fig. 5D).
Fig. 5. Morphological characteristics of fruits and seeds of Pentaclethra macroloba in the
Amazon estuary: A. Immature fruits, B. Ripe fruit with its valves, C. Ripe seeds, D. Seed
embryo and E. Seeds floating in the water of the Amazon river.
The average time of the fruit and seed maturation was 5 months (± 3 months) and 3
months (± 2 months), respectively. We recorded the formation of up to five fruits per
inflorescence. First fruit arise from the 22nd day after flower fertilization (Fig. 6A). At this
stage, it is still possible to observe the persistence of the calyx and corolla. On the 32nd day,
the perianth disappears, the fruit gains length and falcate shape, presenting light red colour in
the centre and pale green colour at the edges. (Fig. 6B). On the 53rd day, the fruit has a pale
green colour, membranaceous consistency and the first light green seed appears (Fig. 6C). On
the 78th day, the fruit has a light green colour, with slightly blackened edges, slightly woody
consistency and with protuberances (valves) on the surface, due to the development and
expansion of the light beige seeds (Fig. 6 D). On the 110th day, the fruit has a dark green colour,
with blackening from the border to the centre, woody consistency and the seed has beige colour
with light brown tones (Fig. 6E). On the 144th day, the fruit has brown colour, with some parts

19
still green, woody consistency and slightly dry and the seed has light brown colour (Fig. 6F).
On the 198th day, the fruit is ripe, ready to disperse the ripe seeds (Fig. 6G).
Fig. 6. Maturation phases of fruits and seeds of Pentaclethra macroloba in the Amazon estuary.
Evolution of shape, size and colour: phase A. 22nd day after flower fertilization; phase B. 32nd
day; phase C. 53rd day; phase D. 78th day; phase E. 110th day; phase F. 144th day; and phase
G. 198th day of maturation.
Germination and initial seedling development: The first seeds germinated on the 28th day
(44 ± 27 days) after sowing and the seedling phase on the 51st after sowing and germination.
Germination is hypogeal, phanerocotylar and unipolar, with an axis between the cotyledons.
During germination, the seed integument breaks, and the primary root emerges (Fig. 7A). The
cotyledons open forming an acute to right angle, with the epicotyl appearing (Fig. 7 B). The
epicotyl lengthens and the first pair of leaves appearing (opposite or sub-opposite; Fig. 7C),
which soon produces the subsequent leaves (Fig. 7D, Fig. 7E). Germination rate was 78 %, with
average germination time of 54 days and the average germination speed was 0.2 seeds.day-1.
Seedling morphology: Seedling (Fig. 7D) has hypocotyl not elongate; cotyledons with
reserves, asymmetrical, internally dark green and externally brownish green, with longitudinal
grooves becoming rough over time. The epicotyl (17.7 cm ± 3.7 cm) is initially greenish,

20
becoming violet to brownish, smooth to slightly rough, striate longitudinally, with yellow
translucent trichomes and with abundant circular and punctate lenticels. Presence of 11-13
triangular cataphylls (1.7 mm ± 0.6 mm) along the epicotyl.
Fig. 7. Phases of seedling development of Pentaclethra macroloba in the Amazon estuary:
phase A. Primary root (pr) formation; phase B. Opening of the cotyledons (cot) and formation
of secondary roots (sr); phase C. Expansion of the epicotyl (epi); phase D. Development of the
first leaves (prot) and formation of cataphyll (cat) in the epicotyl; and phase E. Ripe leaflets
(fo) and apical bud (ag).
First pair of leaves (15.7 cm ± 4.4 cm) are bipinnate, opposite, sub-opposite or alternate,
often paripinnate and rarely imparipinnate and with 6-12 opposite leaflets. The pinnules (9.2
mm ± 0.4 mm) are sessile, oblong, papyraceous, with obtuse and asymmetric base, acute and
mucronate apex and margin of the entire and ciliate blade (yellowish trichomes). The petiole
(3.8 cm ± 1.2 cm) is brownish green, with trichomes, smooth or rough, with wing and flat or
channelled. Discolour leaf blade, with dark green adaxial face and light green abaxial face. The
rachis (6.6 cm ± 2.4 cm) is brownish-green, smooth to rough, apiculate and with trichomes
similar to the petiole, but more abundant. The pulvinus (3.3 mm ± 0.8 mm) is dark green, rough,
with trichomes equal to that of the petiole and with transverse fissures. Presence of a pair of
stipules at the base of the pulvinus (Fig. 8B). Glands or extrafloral nectary at the base of the
pulvinus (Fig. 8C). The petiole is reduced to pulvinule (0.6 mm ± 0.2 mm).
Subsequent leaves (10.4 cm ± 1.7 cm) are equal to the first leaves, but smaller in size,
with alternate spiral phyllotaxy (Fig. 8A) and have 9 to 16 leaflets. The pinnules (7.4 mm ± 2.1
mm) is equal to the first leaves, but larger. The petiole (3.8 cm ± 1.2 cm) is equal to the first
leaves, but smaller. The rachis (6.6 cm ± 2.3 cm) is equal to the first leaves. The pulvinus (3.2

21
mm ± 0.8 mm) is equal to the first leaves. The petiolule is reduced to pulvinule. The pulvinule
(0.6 mm ± 0.2 mm) is equal to the first leaves. Conduplicate prefoliation, from violet to reddish-
brown colour and with translucent trichomes. The internodes have lenticels similar to the
epicotyl, but more abundant.
Fig. 8. Morphological characteristics of Pentaclethra macroloba seedling in the Amazon
estuary: A. Phyllotaxy of the subsequent leaves, B. Stipule at the base of the pulvinus and C.
Gland at the lateral base of the pulvinus.
Geographic distribution: We analysed 210 points of geographical coordinates of digital
botanical collections. The centre of occurrence of P. macroloba was throughout Latin America,
showing that it is a neotropical species (Fig. 9). Data from digital repositories show the presence
of P. macroloba in Guatemala, Costa Rica, Panama, Colombia, Venezuela, Trinidad and
Tobago, Guyana, French Guyana Suriname, and Brazil. In Brazil, the species is endemic to the
Amazon biome and its dispersion area comprises the states of Acre, Amazonas, Roraima, North
Mato Grosso, Amapá and Pará.

22
The species has 123 subpopulations (Fig. 9) distributed on a radius of 5 699 943 km²
along its extent of occurrence. The area of occupation of a species represents its suitable habitat,
for P. macroloba is 9 990 km².
Fig. 9. Dispersion area (left) and number of subpopulations (right) of Pentaclethra macroloba
in the Neotropical region (Data source: tropicos.org and splink.cria.org.br).
DISCUSSION
The individuals described in this study appear to be medium size compared to
individuals of P. macroloba from Central America, where populations are more robust
(Hartshorn, 1983). In Costa Rica, adult individuals occupy the canopy of the forest, reaching a
height of 25 to 35 m and with diameter reaching up to 130 cm (Flores, 2003). In South America,
in the forests of the department of Chocó (Colombia), the majority of individuals occupy the
sub-canopy forest, with a maximum diameter of 65 cm (Hartshorn, 1983). In general, the
population of P. macroloba located in the Amazon forest is of medium height (Hartshorn,
1983), as presented the individuals in this study with a maximum diameter of 59 cm. This shows
evidence that individuals in the Amazon estuary invest more resources to gain height, as the
maximum height (37 m) found was greater than other studies. According to Parolin (2002), the
trees of the Amazon floodplain forests need to increase the maximum height to escape the flood.
External morphological structures shown by P. macroloba, evidence an adaptive
relationship to the periodically flooded environment of the Amazon estuary. In response to
flooding, adventitious roots are produced during the flood season in the Amazon river (from
January to March influenced by the rainy season), decreasing in the dry season (from September

23
to December). This characteristic presented by P. macroloba is a morphological adaptation to
the periodically flooded environment, because the adventitious roots increase the energetic and
respiratory efficiency of the plant (Haase, Simone, & Junk, 2003) when the rhizosphere has low
oxygen concentration. Individuals of P. macroloba from some regions of Costa Rica also have
adventitious roots in flooded environments (Walter & Bien, 1989).
Reproductive strategies can also be observed in P. macroloba. A characteristic that calls
attention in P. macroloba is the spread of several branches at the base of its trunk (Fig. 1G).
According to Gaddis, Zukin, Dieterich, Braker, and Sork (2014), P. macroloba has a high power
of vegetative propagation (clonal reproduction), even if the tree is down, there are numerous
sprouts of branches on the trunk and roots. This strategy is efficient in periodically flooded
environments, because a faster increase in height prevents the plant from being submerged and
has its productivity limited by the flood (Parolin, 2002).
The species has nectar guides on its petals to increase pollen efficiency. Nectar guides
are floral traits that some superior plants have to attract the attention of effective pollinators
(Leonard, Brent, Papaj, & Dornhaus, 2013). The nectar guide on the P. macroloba petal attracts
and directs pollinators into the corolla, where is the nectar gland at the base of the ovary (Barros,
Pedersoli, Paulino, & Teixeira, 2017), increasing the chances of fertilization of the ovule.
Morphological characteristics presented in the fruits and seeds of P. macroloba are
adaptive evidence in search of greater efficiency in the seed dispersal. The low moisture content
that the fruit has when ripe (dry fruit), favours the activation of the explosive dehiscence
mechanism (the ventral suture and the valves) for the seeds to be expelled (Williamson & Costa,
2000) at a distance of up to 10 m out of the tree canopy (Hartshorn, 1983). The flattened shape
(deltoid), together with the accumulation of air between the cotyledons (Soares, Santos, &
Silva, 2019) and the specific gravity of the seed (Williamson & Costa, 2000), allows the seed
to float in the water of the Amazon estuary, achieving greater efficiency in dispersion.
Positive linear relationship between the length of the fruit and the measures of weight,
length and thickness of the seeds, give clue that P. macroloba invests in resources to produce
seeds with high energy content. This strategy allows the seedlings to survive the adverse
conditions of the flooded environment. According to Primack (1987), bulky seeds have high
reserve content in their cotyledons, allowing the seedling to obtain an extra energy source for a
long period.

24
This study showed that the size of the fruit is a good predictor for selecting seeds with
higher fresh weight, this characteristic is important for the management of oil extraction from
Pracaxi. The experience of riverine people in oil extraction shows that larger seeds yield a
greater amount of oil, compared to smaller seeds (in conversation with some riverine people
from the Bailique community, Amapá, Brazil). This linear trend to produce larger fruits and
have larger seeds, can be related to a more lasting source of nutrients for the plants. Large seeds
have greater amounts of nutritive reserves for the development of the plant and help to supply
the lack of photosynthetic tissue in the early stages of the seedling's life (Parolin, 2002).
The germination rate of P. macroloba shows variability in different flooded
environments in the Amazon. In this study, estuarine floodplain forest, the germination rate
found was 78 %. In the igapó forest of Central Amazonia, Parolin, Ferreira, and Junk (2003)
found a germination rate for P. macroloba of 45 %. The two environments differ, among other
factors, by the nutritional status of the soil. The Amazon estuary is bathed by a river containing
high load of sedimentary and nutritional material, which are deposited in the soil of this
environment. The igapós of the Amazon are of older geological origin and are bathed by rivers
of black water, with a high load of plant material in decomposition, of low nutritional fertility
(Junk et al., 2011).
Hypogeal germination presented by P. macroloba is common among Amazonian
floodplain species (Parolin et al., 2003) and among native Amazonian legumes (Moreira &
Moreira, 1996). This type of germination is related to the size of the seedling. Species with
hypogeal germination have seedlings eight times larger than species of epigeal germination,
both types of germination help the seedlings to survive long periods of flooding (Parolin et al.,
2003). Melo, Franco, Silva, Piedade, & Ferreira (2015) emphasize that the germination of the
type epigeal phanerocotylar is an adaptive strategy for obtaining light and CO2 in a short period
of time.
Seedlings of P. macroloba produce numerous lenticels in the epicotyl, fact also
observed by Parolin (2001) in floodplain seedlings in the Central Amazon. Lenticels are
essential in flooded environments seedlings, due to these structures facilitate the entry of
oxygen and improve the internal diffusion of gas between the plant's organs (Parolin, 2012).
When located above the surface of the water, lenticels are a connection point between the aerial
organs and the root of the tree (Haase & Gudrun, 2010).

25
Seedlings that manifest reserve cotyledons in a flooded environment have great chances
of survival in this type of environment. Cotyledon reserves guarantee rapid growth in height
and the seedling can escape total submersion in its initial growth stages (Maia, Maia, & Parolin,
2005). This strategy is efficient in the Amazon estuary, because most pracaxi seeds germinate
at the end of the flood period (period that the waters recede from the interior of the forest) and
develop in the drought period (dry season from September to December, where river water does
not reach the interior of the forest), being sufficient for the seedling to use the cotyledon reserves
and gain height to escape the flood of next year.
Dispersion area of P. macroloba is quite wide in the Neotropical region, allowing the
species to colonize various habitats. In Costa Rica, the species is found in ancient alluvial soils
of virgin or secondary forests (Hartshorn, 1983), in annual precipitation regime above 4 300
mm and average temperature of 25 °C (Eaton et al., 2011). On the island of Trinidad, the species
inhabits seasonal perennials forests with high content of clay and sand (Greig-Smith, 1952). In
the Amazon region, the species can colonize in flooded and non-flooded environments. Dense
populations of the species can be found in clayey latosolic soils of the dense non-flooded
ombrophilous forest (Condé & Tonini, 2013). In Central Amazonia, P. macroloba is not
dominant, but it tolerates lasting flooding up to 14 m in height and poor soils in the igapó forests
(Ferreira & Parolin, 2007). P. macroloba dominates estuarine floodplain forests (Carim et al.,
2016), areas with high sedimentary load, rich in nutrients and low amplitude of flooding (Junk
et al., 2014).
The circular buffer method detected the presence of 123 subpopulations of P. macroloba
widely distributed in the Neotropical region. Hartshorn (1983) considers the presence of three
large populations of P. macroloba distributed in Latin America. According to the author, the
first population, widely studied, occurs initially in the coastal region of western Nicaragua,
passing through Costa Rica and taking the Western part of Panama. The second population
occurs in the West of the department of Chocó, Colombia and in the humid part adjacent to the
Darien province of Panama. The third population is found in Northeast Venezuela, the island
of Trinidad, the Guianas and the Amazon Basin. The author considers the necessity for further
studies aimed at the population of the Amazon Basin, since this population is little known in
the scientific community.
Considering the Amazon region, P. macroloba presents a pattern “centered in the
Amazon” proposed by Gentry (1981). The pattern centered in the Amazon is characterized by
having a high density of species in the Amazon Basin, while the pattern centered in the Andes

26
has its centre of diversity on the border of Central America with North-western South America
(Antonelli & Sanmartín, 2011). Gentry (1981) explains that the center of endemism for some
species in the Amazon was influenced by changes that occurred in the Pleistocene (theory of
Pleistocene forest refuges). Drastic climate fluctuations in the Pleistocene period caused the
fragmentation and coalescence of tropical forests, resulting in the isolation and speciation of
large forest populations (Connor, 1986). Several theories emerged to explain the diversity and
distribution pattern of tropical species (Antonelli & Sanmartín, 2011), however, some
knowledge gaps remain unanswered, requiring in-depth studies on the topic.
External morphological characteristics of P. macroloba shows evolutionary and
adaptive evidence of the species in response to the dynamics and the varied flooding amplitudes
of the Amazon floodplain forests. The plasticity of adaptation to different habitats may explain
the dominance of this species and its centre of diversification in the Amazon. The result of this
study is essential to assist in management strategies, allowing, among others, a more precise
identification of productive trees to order the collection of seeds. In the scope of public policies,
the results reinforce the valorisation of non-wood forest products, orienting towards the better
use of pracaxi seeds, a non-wood product of local socioeconomic importance and that inhabits
the fragile estuarine floodplains of the Amazon.
ACKNOWLEDGMENTS
We thank Programa de Pós-graduação em Ecologia (PPGEco/INPA), Instituto Nacional de
Pesquisas da Amazônia (INPA), Conselho Nacional de Desenvolvimento Científico e
Tecnológico (CNPq) (doctoral scholarship 142316/2016-4), Empresa Brasileira de Pesquisa
Agropecuária (Embrapa Amapá) and Grupo Ecologia, Monitoramento e Uso Sustentável de
Áreas Úmidas (MAUA/PELD/CNPq/FAPEAM). This work was supported by the Empresa
Brasileira de Pesquisa Agropecuário do Amapá through Kamukaia III project
[02.13.07.007.00.00].
RESUMEN
Morfología, germinación y distribución geográfica de Pentaclethra macroloba (Fabaceae):
árbol amazónico hiperdominante. Introducción: Pentaclethra macroloba es un árbol
hiperdominante en el estuario del Amazonas, de gran importancia socioeconómica para la

27
región, pues el aceite de sus semillas es un poderoso medicamento natural. Objetivo: Nuestro
objetivo fue caracterizar la estructura morfológica, las adaptaciones morfológicas en respuesta
a las inundaciones diarias del estuario del Amazonas y el área biogeográfica de P. macroloba.
Métodos: Una descripción detallada de la morfología externa, desde la germinación hasta el
árbol adulto, se obtuvo de individuos ubicados en el bosque periódicamente inundado, al
noreste de la Amazonia. El área de ocupación y la extensión geográfica de P. macroloba se
determinaron a partir del punto de coordenadas geográficas de muestras botánicas depositadas
en las colecciones digitales de Mobot y SpeciesLink. Resultados: Los individuos adultos tienen
estructuras adaptadas en respuesta a las inundaciones diarias, tales como: raíces adventicias
para aumentar la eficiencia respiratoria y lenticelas en el tronco, que sirven como punto de
conexión para el transporte de oxígeno entre la raíz y la parte aérea. Los frutos secos favorecen
la activación del mecanismo de dehiscencia explosiva, permitiendo que la semilla sea expulsada
a larga distancia. La forma deltoidea de la semilla permite la fluctuación en el agua y una
dispersión más eficiente. La plántula es hipogel fanerocotiledones y con un par de cotiledones
de reserva que proporcionan a la plántula una fuente extra de energía para escapar de la
inundación. La tasa de germinación fue del 78 % y la velocidad fue de 0.2 semillas día-1. El
método Circular buffer reveló la presencia de 123 subpoblaciones de P. macroloba distribuidas
en un radio de 5 699 943 km² a lo largo de la región neotropical. Conclusiones: Gran parte de
las estructuras morfológicas de P. macroloba son respuestas adaptativas y evolutivas al
ambiente periódicamente inundado del estuario del Amazonas, lo que demuestra que estos
ambientes actúan como filtro ambiental seleccionado las especies mejor adaptadas al medio. Su
amplia área geográfica denota la plasticidad de adaptarse a diferentes ambientes, lo que puede
justificar su monodominancia en algunas regiones.
Palabras clave: adaptación morfológica; inundaciones por mareas; árbol neotropical; raíces
adventicias; aceite de pracaxi; manejo, conservación.
REFERENCIAS
Almeida, S.S., Amaral, D.D., & Silva, A.S.L. (2004). Análise florística e estrutura de florestas
de várzea no estuário amazônico. Acta Amazonica, 34(4), 513-524.
Alvares, C.A., Stape, J.L., Sentelhas, P.C., Gonçalves, J.L.M., & Sparovek, G. (2013).
Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift, 22(6), 711-

28
728.
Antonelli, A., & Sanmartín, I. (2011). Why are there so many plant species in the Neotropics?
TAXON, 60(2), 403-414.
Barros, T.C., Pedersoli, G.D., Paulino, J.V., & Teixeira, S.P. (2017). In the interface of
Caesalpinioids and Mimosoids: Comparative floral development elucidates shared
characters in Dimorphandra mollis and Pentaclethra macroloba (Leguminosae).
American Journal of Botany, 104(2), 218-232.
Barroso, G.M., Morim, M.P., Pioxoto, A.L., & Ichaso, C.L.F. (2004). Frutos e sementes:
morfologia aplicada à sistemática de dicotiledôneas. Viçosa, Brasil: Universidade Federal
de Viçosa.
Camargo, J., Ferraz, I., Mesquita, M., Santos, B., & Brum, H. (2008). Guia de propágulos e
plântulas da Amazônia. Manaus, Brasil: Instituto Nacional de Pesquisas da Amazônia.
Carim, M.J.V., Wittmann, F. K., Piedade, M. T. F., Guimarães, J.R.S., & Tostes, L.C.L. (2016).
Composition, diversity, and structure of tidal “Várzea” and “Igapó” floodplain forests in
eastern Amazonia, Brazil. Brazilian Journal of Botany, 40(1), 115-124.
Condé, T.M., & Tonini, H. (2013). Fitossociologia de uma Floresta Ombrófila Densa na
Amazônia Setentrional, Roraima, Brasil. Acta Amazonica, 43(3), 247-260.
Connor, E.F. (1986). The role of Pleistocene forest refugia in the evolution and biogeography
of tropical biotas. Trends in Ecology and Evolution, 1(6), 165-168.
Costa, M.N.F.S., Muniz, M.A.P., Negrão, C.A.B., Costa, C.E.F., Lamarão, M.L.N., Morais, L.,
… Costa, R.M.R. (2014). Characterization of Pentaclethra macroloba oil. Journal of
Thermal Analysis and Calorimetry, 115(3), 2269-2275.
CRIA (Centro de Referência e Informação Ambiental). (2019). speciesLink is a distributed
information system that integrates primary data from biological collections. Retrieved
from http://splink.cria.org.br
Dantas, A.R., Marangon, L.C., Guedes, M.C., Feliciano, A.L.P., & Lira-Guedes, A.C. (2017).
Spatial distribution of a population of Pentaclethra macroloba (Willd.) Kuntze in a
floodplain forest of the Amazon estuary. Revista Árvore, 41(4), 1-11.

29
Dauby, G. (2019). ConR: Computation of Parameters Used in Preliminary Assessment of
Conservation Status (Version 1.3.0, R Package) Retrieved from https://CRAN.R-
project.org/package=ConR.
De Simone, O., Junk, W.J., & Schmidt, W. (2003). Central Amazon floodplain forests: Root
adaptations to prolonged flooding. Russian Journal of Plant Physiology, 50(3), 848-855.
Eaton, W.D., McDonald, S., Roed, M., Vandecar, K.L., Hauge, J.B., & Barry, D. (2011). A
comparison of nutrient dynamics and microbial community characteristics across seasons
and soil types in two different old growth forests in Costa Rica. Tropical Ecology, 52(1),
35-48.
Ferreira, L.V., & Parolin, P. (2007). Tree phenology in Central Amazonian floodplain forests:
effects of water level fluctuation and precipitation at community and population level.
Pesquisas Botânica, 58, 139-155.
Flores, E.M. (2003). Pentaclethra macroloba (Willd.) Kuntze. In J.A Vozzo (Ed.), Tropical
tree: seed manual (pp. 601–604). Washington DC, USA: United States Department of
Agriculture.
Gaddis, K.D., Zukin, H.L., Dieterich, I.A., Braker, E., & Sork, V.L. (2014). Effect of clonal
reproduction on genetic structure in Pentaclethra macroloba (Fabaceae: Mimosoideae)
Keith. Revista de Biología Tropical, 62(2), 443-454.
Gaston, K.J., & Fuller, R.A. (2009). The sizes of species’ geographic ranges. Journal of Applied
Ecology, 46(1), 1-9.
Gentry, A.H. (1981). Distributional patterns and an additional species of the Passiflora vitifolia
complex: Amazonian species diversity due to edaphically differentiated communities.
Plant Systematics and Evolution, 137(1–2), 95-105.
Gonçalves, E.G., & Lorenzi, H. (2011). Morfologia Vegetal: organografia e dicionário
ilustrado de morfologia das plantas vasculares (2nd Ed.). São Paulo, Brazil: Instituto
Plantarum de Estudos da Flora.
Greig-Smith, P. (1952). Ecological observations on degraded and secondary forest in Trinidad,
British West Indies: I. General features of the vegetation. Journal of Ecology, 40(2), 283-
315.
Haase, K., & Gudrun, R. (2010). The morphology and anatomy of tree roots and their aeration

30
strategies. In W.J. Junk, M.T.F. Piedade, F.K. Wittmann, J. Schöngart, & P. Parolin (Eds.),
Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable
Managementoodplain (pp. 141–161). Dordrecht, Netherlands: Springer Dordrecht
Heidelberg London.
Haase, K., Simone, O.D.E., & Junk, W.J. (2003). Internal oxygen transport in cutting from
flood-adapted várzea tree species. Tree Physiology, 23(15), 1069-1076.
Harris, J.G., & Harris, M.W. (2001). Plant identification terminology: an illustrated glossary
(2nd ed.). Utah, USA: Spring Lake Publishing.
Hartshorn, G.S. (1983). Pentaclethra macroloba (Gavilan). In D.H. Janzen (Ed.), Costa Rican
Natural History (pp. 301–303). Chicago, USA: University of Chicago Press.
IBGE - Instituto Brasileiro de Geografia e Estatística. (2012). Manual técnico da vegetação
brasileira : sistema fitogeográfico : inventário das formações florestais e campestres :
técnicas e manejo de coleções botânicas : procedimentos para mapeamentos (2nd Ed.). Rio
de Janeiro, Brasil: IBGE.
IUCN Standards and Petitions Subcommittee. (2017). Guidelines for Using the IUCN Red List
Categories and Criteria. Retrieved 7 June 2019, from
http://www.iucnredlist.org/documents/RedListGuidelines.pdf
Junk, W.J., Piedade, M.T.F., Lourival, R., Wittmann, F., Kandus, P., Lacerda, L.D., …
Agostinho, A.A. (2014). Brazilian wetlands: Their definition, delineation, and
classification for research, sustainable management, and protection. Aquatic
Conservation: Marine and Freshwater Ecosystems, 24(1), 5-22.
Junk, W.J., Piedade, M.T.F., Schöngart, J., Cohn-Haft, M., Adeney, J.M., & Wittmann, F.
(2011). A classification of major naturally-occurring amazonian lowland wetlands.
Wetlands, 31(4), 623-640.
Laman, T.G. (1995). Safety recommendations for climbing rain forest trees with "Single rope
technique". Biotropica, 27(3), 406-409.
Leonard, A.S., Brent, J., Papaj, D.R., & Dornhaus, A. (2013). Floral nectar guide patterns
discourage nectar robbing by bumble bees. PLoS ONE, 8(2), 1-6.
Maguire, J.D. (1962). Speed of germination-aid selection and evaluation for seedling
emergence and vigor. Crop Science, 2(2), 176-177.

31
Maia, L.A., Maia, S., & Parolin, P. (2005). Seedling morphology of non-pioneer trees in Central
Amazonian várzea floodplain forests. Ecotropica, 11(1–2), 1-8.
Melo, M.G.G., Mendonça, M.S., & Mendes, A.M.S. (2004). Análise morfológica de sementes,
germinação e plântulas de jatobá (Hymenaea intermedia Ducke var. adenotricha (Ducke)
Lee & Lang.) (Leguminosae-caesalpinioideae). Acta Amazonica, 34(1), 9-14.
Melo, R.B., Franco, A.C., Silva, C.O., Piedade, M.T.F., & Ferreira, C.S. (2015). Seed
germination and seedling development in response to submergence in tree species of the
Central Amazonian floodplains. AoB PLANTS, 7(Plv041), 1-12.
Moreira, F.M.S., & Moreira, F.W. (1996). Características da germinação de sementes de 64
espécies de leguminosas florestais nativas da Amazônia, em condições de viveiro. Acta
Amazonica, 26(1/2), 3-16.
Parolin, P.I.A., Ferreira, L.V., & Junk, W.J. (2003). Germination characteristics and
establishment of trees from Central Amazonian flood plains. Tropical Ecology, 44(2), 157-
169.
Parolin, P. (2012). Diversity of adaptations to flooding in trees of amazonian floodplains.
Pesquisas Botanica, 63, 7-28.
Parolin, P. (2001). Seed germination and early establishment of 12 tree species from nutrient-
rich and nutrient-poor Central Amazonian floodplains. Aquatic Botany, 70(2), 89-103.
Parolin, P. (2002). Submergence tolerance vs. escape from submergence: two strategies of
seedling establishment in Amazonian floodplains. Environmental and Experimental
Botany, 48(2), 177-186.
Pinto, E.R. (2014). Solos, hidrologia e estrutura populacional de pracuubeiras em florestas de
várzea do estuário amazônico (Master's thesis). Universidade Federal do Amapá, Macapá,
Brazil.
Primack, R.B. (1987). Relationships among flowers, fruits, and seeds. Annual Review of
Ecology and Systematics, 18(1), 409-430.
R core Team. (2019). R: A Language and Environment for Statistical Computing. Vienna,
Austria. Retrieved from http://www.R-project.org/.
Rivers, M.C., Bachman, S., Meagher, T.R., Lughadha, E.N., & Brummitt, N.A. (2010).

32
Subpopulations, locations and fragmentation: Applying IUCN red list criteria to herbarium
specimen data. Biodiversity and Conservation, 19(7), 2071-2085.
Schöngart, J., Junk, W.J., Piedade, M.T.F., Ayres, J.M., Hüttermann, A., & Worbes, M. (2004).
Teleconnection between tree growth in the Amazonian floodplains and the El Niño-
Southern Oscillation effect. Global Change Biology, 10(5), 683-692.
Schöngart, J., Piedade, M.T.F., Ludwigshausen, S., Horna, V., & Worbes, M. (2002).
Phenology and stem-growth periodicity of tree species in Amazonian floodplain forests.
Journal of Tropical Ecology, 18(4), 581-597.
Silva, J.O., Fernandes, R.S., Ticli, F.K., Oliveira, C.Z., Mazzi, M.V., Franco, J.J., … Sampaio,
S.V. (2007). Triterpenoid saponins, new metalloprotease snake venom inhibitors isolated
from Pentaclethra macroloba. Toxicon, 50(2), 283-291.
Soares, R.N., Santos, R.O., & Silva, B.M.S. (2019). Morphological aspects and anatomy of the
fruit, seeds and seedlings of Pentaclethra macroloba (willd.) Kuntze (Fabaceae). Journal
of Seed Science, 41(4), 452-460.
ter Steege, H., Pitman, N.C.A., Sabatier, D., Baraloto, C., Salomão, R.P., Guevara, J.E., …
Silman, M.R. (2013). Hyperdominance in the Amazonian tree flora. Science, 342(6156),
325-334.
Tropicos. (2019). Missouri Botanical Garden. Retrieved 11 February 2019, from
http://www.tropicos.org
Vilhena, J.E.S., Silva, R.B.L., & Freitas, J.L. (2018). Climatologia do Amapá: quase um século
de história. Rio de Janeiro, Brasil: Gramma.
Walter, C.A., & Bien, A. (1989). Aerial root nodules in the tropical legume, Pentaclethra
macroloba. Oecologia, 80(1), 27-31.
Wiens, J.J. (2011). The niche, biogeography and species interactions. Philosophical
Transactions of the Royal Society B: Biological Sciences, 366(1576), 2336-2350.
Williamson, G.B., & Costa, F. (2000). Dispersal of Amazonian Tress: hydrocory in
Pentaclethra macroloba. Biotropica, 32(3), 548-552.

33
Capítulo 2
________________________________________ Dantas, A.R., Guedes, M.C., Lira-Guedes, A.C & Piedade, M.T.F. Phenological behavior and floral visitors of Pentaclethra macroloba, a hyperdominant tree in the Brazilian Amazon River estuary. Published in Trees, DOI 10.1007/s00468-021-02095-x.

34
Phenological behavior and floral visitors of Pentaclethra macroloba, a hyperdominant tree
in the Brazilian Amazon River estuary
Adelson Rocha Dantas1*, Marcelino Carneiro Guedes2, Ana Cláudia Lira-Guedes2, Maria
Teresa Fernandez Piedade1
1 Instituto Nacional de Pesquisas da Amazônia, Programa de Pós-Graduação em Ecologia,
69067-375 Manaus, Brasil;
2 Empresa Brasileira de Pesquisas Agropecuária, Departamento de Recursos Florestais, 68903-
419 Macapá, Brasil
*corresponding author: [email protected]
Abstract
Natural history of hyperdominant tree populations in the Brazilian Amazon Region is still
unknown in plant science. Pentaclethra macroloba (Willd.) Kuntze (Fabaceae) is a
hyperdominant tree species which has multiple-uses and its seeds are extensively explored to
extract medicinal oil. We evaluated the phenological cycles and floral visitors of P. macroloba
and the effect of the daily tide of the Amazon River on the phenophases. Phenophases of flower
bud, anthesis flowers, immature fruit, ripe fruit, seed dispersal, new leaf, mature leaf, and leaf
fall of 29 trees in the Northeast of the Brazilian Amazon were monitored during 28 months.
Hydrometeorological data of rainfall, maximum temperature and flood height on the tree trunk
were obtained. Generalized Linear Models were used to explain the relationship between
phenophases and hydrometeorological parameters. Flowering was synchronized in the dry
season (flower buds: βtemperature =1.30, p <0.01 and flowers in anthesis: βtemperature =1.84; p
<0.001). Immature fruits appeared during the dry season (βtemperature =0.67; p <0.01) and ripe
fruits in the rainy season. Seeds were dispersed during the rainfall (βrainfall =0.0051; p <0.01)

35
and flooding of the river (βwater =0.12; p <0.001). There was no relationship between leaf change
and hydrometeorological variables. The main floral visitors were wasps, bees and ants. The
rainfall seasonality is a key factor that stimulates reproductive events of P. macroloba. Daily
river flooding can be considered a driver that stimulates the tree to disperse its seeds at the peak
of the river flood, where they can be transported the long distances.
Keywords: daily tide, phenophases, pollination, pracaxi oil, rainfall, reproductive strategy.
Declarations
Funding
The study received financial support from Empresa Brasileira de Pesquisa Agropecuária
(Embrapa Amapá)/ Kamukaia III Project (Process Number: 02.13.07.007.00.00) and Conselho
Nacional de Desenvolvimento Científico e Tecnológico (Process Number/ Doctoral
scholarship: 142316/2016-4).
Author Contribution Statement
ARD, MCG, ACLG and MTFP conceived the ideas and designed methodology; ARD collected
the data; ARD, MCG, ACLG and MTFP analyzed the data; ARD led the writing of the
manuscript. All authors contributed critically to the drafts and gave final approval for
publication.
Key message Amazon River tide is an environmental drive that stimulates Pentaclethra
macroloba to disperse its seeds during the flood peak. Reproductive strategies of the species
result in its dominance in the Amazon.

36
Introduction
Phenology study the interactions of seasonal biological events with the abiotic and biotic
factors of the habitat (Lieth 1974). Environmental triggers that stimulate and drive the
phenological patterns of tropical trees are still very much questioned and debated by plant
ecologists (Morellato et al. 2013; Mendoza et al. 2017). The great diversity of phenological
patterns found in tropical forests may be the result of a long evolutionary process of adaptation
to changes in the environment and to dispersing and pollinating agents (van Schaik et al. 1993;
Sakai 2001).
In the flooded forests of the Amazon Region, the elevation and descent of the water
level of the Amazonian rivers is a phenomenon known as “flood pulse” (Junk 2001). Flood
pulse is the main driver of the phenological patterns of flooded forests (Schöngart et al. 2002;
Haugaasen and Peres 2005; Parolin et al. 2010) and it also acts as an environmental filter
selecting species adapted to the constantly flooded environment (Luize et al. 2018). The
floodplain forests of the Amazon estuary are bathed by the Amazon River and comprises an
area of approximately 199,281 km² (Junk and Piedade 2010), between the coastal areas of the
States of Amapá and Pará in Brazil. Different from the Central Amazonia, where the flood pulse
is monomodal and conditions the species to long periods of flooding that can last up to 243
days and reach a mean height of 6 m by the river water mark registered on the tree trunks
(Schöngart et al. 2002), the Amazon estuary is influenced by ocean tides, resulting in two daily
flood cycles (Junk et al. 2014), that can reach a height of up to 4 m in the rainy season (Cunha
et al. 2017).
The flood pulse of the Central Amazon floodplains is the main driver that models
reproductive cycles (Parolin et al. 2010) and phenological patterns of the trees (Worbes 1997;
Schöngart et al. 2002; Haugaasen and Peres 2005; Ferreira and Parolin 2007; Hawes and Peres
2016). However, the influence of the tidal cycles on the phenology of the trees in the Amazon

37
estuary, where the flood pulse is polymodal and of low amplitude (Junk et al. 2014), is still little
known (Cattanio et al. 2004; Dantas et al. 2016).
Sedimentary material of Andean and pre-Andean origin (Wittmann et al. 2010)
deposited inside the forest by the Amazon River, and the annual flood cycles, allows the
establishment of several tree species adapted to the daily flood cycle (Carim et al. 2016). One
of these species is Pentaclethra macroloba (Willd.) Kuntze, locally known as pracaxizeiro,
which is a hyperdominant Fabaceae from the Amazon region (ter Steege et al. 2013) and has
high frequency in the Amazon estuary (Carim et al. 2016). P. macroloba is a pioneer species,
monoecious (Barros et al. 2017), with height of 13 m and diameter of 26 cm means, bipinnate
leaves, inflorescence presents up to 411 flowers, dry legume fruit has explosive dehiscence and
the flattened brown seed has an air pocket between the cotyledons which facilitates hydrocoric
dispersion (Dantas et al. 2021). P. macroloba forms highly aggregate populations (Dantas et al.
2017) and it is adapted to the daily tide of the Amazon River, presenting adventitious roots and
lenticels to increase respiratory efficiency in conditions of low oxygen availability in the
rhizosphere (Dantas et al. 2021).
Hartshorn (1983) considers the existence of three large populations of P. macroloba in
Latin America. The first population, widely studied, is concentrated in Costa Rica, mainly in
the La Selva Forest Reserve, where it has population monodominance. The second population
is concentrated in the extension from Panama to Colombia. The third and last population is
located in the Northeast of the Brazilian Amazon and its natural history is little known.
P. macroloba has great socioeconomic relevance in the Amapá State. Many riverside
women collect the seeds to extract the oil, a traditional knowledge that is passed from generation
to generation. The oil is usually sold at traditional fairs and even for some cosmetic industries.
The oil is also used by riverine forest people as a natural medicine, due to be a powerful healing
agent for wounds and burns (Banov et al. 2014). The bark of the P. macroloba trunk has

38
triterpenoid saponins that act against hemorrhage caused by the venom of Bothrops genus
vipers (Silva et al. 2007).
Multiple uses of P. macroloba and the low knowledge about basic aspects of its ecology
are factors that make the species vulnerable to overexploitation and the exhaustion of its natural
resources, mainly the seeds, in the Amazon estuary. Therefore, phenological studies of P.
macroloba are essential to elucidate the seasonal growth and reproduction rhythms of trees,
related to the abiotic and biotic characteristics of the habitat (Lieth 1974). One of the objectives
of the phenological studies is to evaluate the rhythm of flowering (emission of flowers and
floral development), fruiting (fruit ripening and seed dispersal) and leaf change (emission of
new leaves and leaf senescence) phases (Alencar 1998). What is known about the phenology
of P. macroloba is that it has annual pattern of flowering and fruiting and remains with its
canopy evergreen (Freitas et al. 2003; Vilchez et al. 2007). These few studies are at the
community level and superficial, being that a large knowledge gap remains unanswered about
the interactions of the phenological phases of the species with its habitat.
Plant-pollinator interactions play an important role in diversification of the flowering
patterns of tropical trees (Oliveira and Rech 2018). The periodicity and intensity of the
flowering are factors directly related to attractiveness and availability of floral resources for the
pollinators (Vieira et al. 2012). Many trees show synchronized flowering (Augspurger 1983)
and floral adaptations (Fouks and Wagoner 2019) to attract pollinators. This synchrony is
certainly adaptive, as many tree species, such as P. macroloba, invest enough energy in floral
development, which can last for days, only then to expose their reproductive organs for one or
two days.
We aimed to determine the reproductive and vegetative phenophases pattern and
evaluate the main floral visitors of P. macroloba to assist in the management and conservation
of its natural resources in the Amazon estuary. Due to its high adaptation and dominance in the
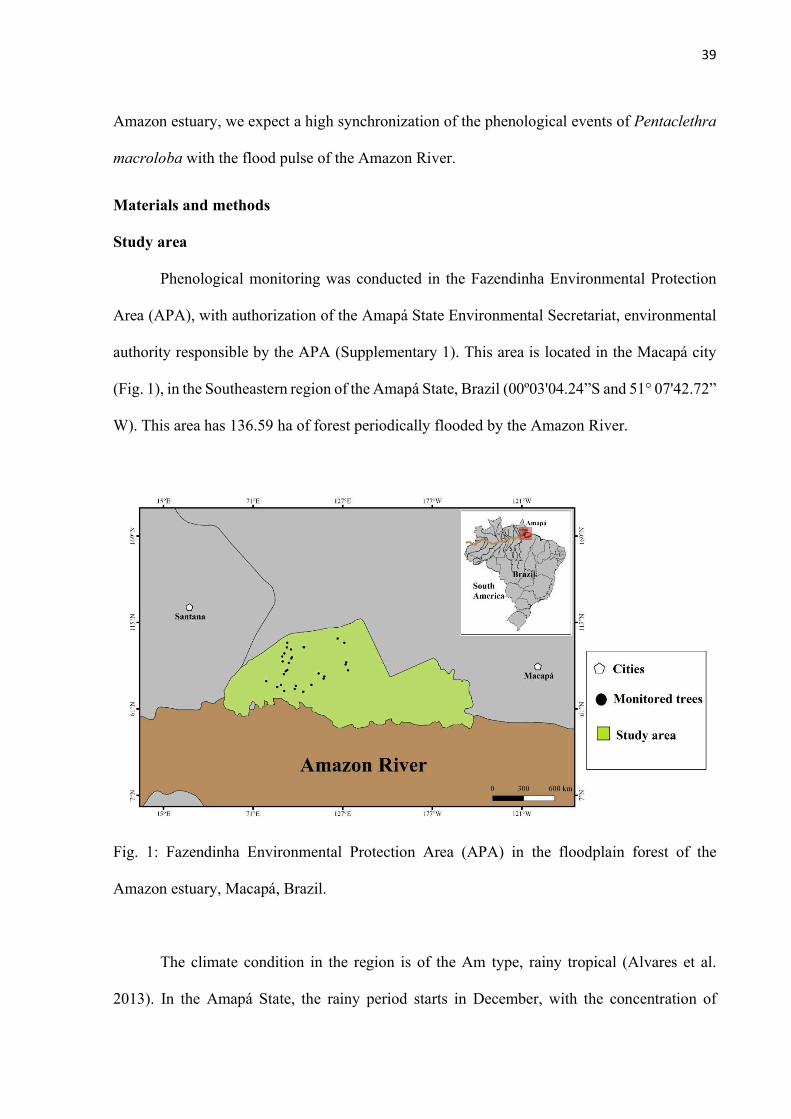
39
Amazon estuary, we expect a high synchronization of the phenological events of Pentaclethra
macroloba with the flood pulse of the Amazon River.
Materials and methods
Study area
Phenological monitoring was conducted in the Fazendinha Environmental Protection
Area (APA), with authorization of the Amapá State Environmental Secretariat, environmental
authority responsible by the APA (Supplementary 1). This area is located in the Macapá city
(Fig. 1), in the Southeastern region of the Amapá State, Brazil (00º03'04.24”S and 51° 07'42.72”
W). This area has 136.59 ha of forest periodically flooded by the Amazon River.
Fig. 1: Fazendinha Environmental Protection Area (APA) in the floodplain forest of the
Amazon estuary, Macapá, Brazil.
The climate condition in the region is of the Am type, rainy tropical (Alvares et al.
2013). In the Amapá State, the rainy period starts in December, with the concentration of

40
rainfall in March, when the peak of rainfall remains above 60 mm/month until August. The
drought period starts in September when the rainfall is below 60 mm monthly. Climatological
data from 1987 to 2016 show that the mean annual rainfall is 2460 mm and the mean maximum
temperature varies from 30º to 32ºC (Vilhena et al. 2018).
The soils of the estuarine floodplain forests are classified as typical Melanic Gleysols
Ta Eutrophic, with predominance of silt. Both the sediment and the soil are formed by smectite,
illite, kaolinite, goethite, anatase and quartz (Pinto 2014).
Vegetation is classified as Dense Alluvial Ombrophilous Forest (IBGE 2012) and
evergreen, with dominance, among trees, of Mora paraensis (Ducke) Ducke, Astrocaryum
murumuru Mart., Pentaclethra macroloba (Willd.) Kuntze, Carapa guianensis Aubl. and
Virola surinamensis (Rol.) Warb. (Carim et al. 2016).
Phenological data and floral visitors
An inventory of all individuals with Diameter at Breast Height DBH ≥ 5 cm conducted
in the study area registered 2,072 trees. Twenty-nine adult and productive trees of different
diametric classes (with diameters between 14 cm to 49 cm measured at a height of 1.30 m from
the ground) and dominant and codominant in the canopy (between 15 m to 30 m of height) were
selected for phenological observations and to record the activity of floral visitors. Morellato et
al. (2010) recommend sampling at least 15 individuals to represent the phenological pattern of
the population. The selection criteria were good phytosanitary aspect and minimum distance of
50 m between individuals. The species was identified and the vouchers were deposited in the
Embrapa Herbarium Amazônia Oriental – IAN (registration numbers IAN192676 and
IAN192677) (Dantas et al. 2021).
Phenological monitoring was conducted from September 2017 to December 2019,
fortnightly, totaling 28 months of observations. In this study, we consider as: flowering – the
emission of the floral bud until the senescence of the flower, fruiting – emission of the fruit

41
until the dispersion of the seed and leaf change – the emission of leaf buds on the branch until
the senescence of the leaf.
The phenophases observed were: 1) flower buds, begins when the species has small
green closed buds and ends with the opening of the corolla; 2) flowers in anthesis, begins with
the opening of the corolla and ends when staminodes lose their white color, changing to a brown
color until their complete senescence; 3) immature fruits, begins when small pods of light green
color appear with shiny surface and ends when the pods acquire dark green color and obtain
over time brown color; 4) ripe fruits, when the fruits have brown color and become dry; 5) seed
dispersal, occurs when the dried fruit releases the seeds - autochoric dispersion; 6) new leaves,
begins when the branches produce small leaves of violet color, changing to a light green color,
and ends when the leaves have dark green color; 7) mature leaves, when the leaves lose their
dark green color and change to brown color until leaf senescence; and 8) leaf fall, disconnection
of the petiole leaf from the branch, causing empty spaces in the crown. The trees were
monitored with the aid of binoculars (60x magnification).
Phenophases were semi-quantified using the methodology proposed by Fournier (1974),
in which the phenological events are grouped into four categories of 25% amplitude. The
frequency of manifestation of the reproductive phenophases was classified as: continual, sub-
annual, annual and supra-annual (Newstrom et al. 1994). The pattern of production and leaf fall
were determined based on three categories: deciduous, brevi-deciduous and evergreen
(Schöngart et al. 2002).
Thirty-one inflorescences of 15 individuals were marked and monitored, from August
to November 2018 every two days, to assess the stages of floral development over time and the
floral longevity of P. macroloba. Ninety flowers were marked to estimate floral longevity.
Floral longevity was considered from floral anthesis to flower senescence (Primack 1985).

42
The behavior and frequency of floral visitors were monitored during August to
November 2018, every two days, in 10 trees. Focal observations were conducted, in each tree,
in the morning (5 hours) and afternoon (5 hours) period (Malucelli et al. 2018). The behavior
of the floral visitors was filmed with camera of high-resolution (Sony HDR-CX405 60x), for a
better analysis of the visitor-flower interactions. Insect floral visitors were captured with the
aid of traps, 1.5 liter polyethylene bottle with inverted funnel, containing a sugary solution
(water + sugar) (PPBio 2019). The traps were placed close of the monitored inflorescences, at
heights of 15 m to 25 m in the tree crowns. The parameters analyzed of the floral visitors were:
frequency of visits, foraging duration and number of visitors per inflorescence. The insects were
identified and deposited in the entomological collection of the National Research Institute for
Amazon.
Hydrometeorological data
Monthly data of maximum temperature (°C) and accumulated precipitation (mm) were
obtained from the Macapá Climatological Station (00°02’07’’ S e 51°05’55’’ W), located about
1 km from the study area (INMET 2019).
The height of the flood was monitored inside the forest together with the phenological
observations. For this, white strings were installed parallel, at a height of 1 m, on the trunk of
each monitored tree. The Amazon river water, rich in sediment, stains the white string and
records a brownish mark, facilitating the measurement of the height that the water reaches with
a tape measure. Every fortnight, the strings were changed for subsequent evaluation. In
addition, tide level data of the Amazon River was obtained from the Porto de Santana
Maregraph Station (00°03’07’’ S e 51°10’1 W), about 6 km from the study area (CHM 2019).
Data analysis
To obtain a single response value, the fortnightly data of each phenophases were
converted to monthly data through the sum of the fortnightly phenological data, divided by the

43
number of observations in the month for each individual. The intensity of each phenophase in
the species was determined using the Fournier formula (Fournier 1974):
% Fournier = ∑ Fournier
4 × N × 100
where: ƩFournier is the sum of the categories of Fournier, divided by the maximum Fournier
that can be reached by all individuals N in the sample.
The relationship of the meteorological and hydrological variables with the phenophases
was evaluated using univariate Generalized Linear Models (GLMs) with quasi-binomial errors
(Newbery et al. 2006; Dantas et al. 2016). We attribute 0 for absence and 1 for presence of
phenophase, and the proportion of individuals manifesting certain phenophases in each month
was calculated (Dantas et al. 2016).
Circular statistics was applied to verify whether the phenological patterns of P.
macroloba are seasonal (Morellato et al. 2010a). For this, the 12 months of the year were
converted into angles, with intervals of 30º, from 0º = January (first month) to 330º = December
(last month). The mean angle 𝑎 and vector r parameters were calculated. The 𝑎 indicates the
period of year when phenophases are most frequently and the vector r is a measure of
concentration around of the mean angle. The Rayleigh test (Z) was applied to assess the
statistical significance of the mean angle and the following hypotheses were tested: H0 = dates
are randomly or uniformly distributed around the year and H1 = dates are not uniformly
distributed around the year. If the alternative hypothesis is accepted (H1), the vector r can be
considered a measure of the degree of phenological seasonality. The vector r has no unity and
can vary from 0 to 1, greater is the phenological seasonality when the value approaches 1
(Morellato et al. 2000).
The Augspurger index of synchrony was calculated aiming to verify the presence or
absence of synchrony in the flowering and fruiting phases (Augspurger 1983). The Augspurger
index of synchrony is calculated using the following formula:

44
Xi =∑ ej
(N − 1). fi; Z =
∑ Xi
N
where: Xi is the synchrony index of individual i; Z is the population synchrony index; ej is the
number of months in which individuals i and j flourished synchronously, with i ≠ j; fi is the
number of months that individual i flowered; N is the total number of individuals in the sample.
The intensity of the synchrony was classified in the following scales: 0 - absence of
synchrony; 0.1 to 0.27 - minimum synchrony; 0.28 to 0.45 - low synchrony; 0.46 to 0.69 -
medium synchrony; 0.70 to 0.99 - high synchrony; and 1 - perfect synchrony (Dantas et al.
2016). Basic commands of the R program were used to perform all statistical analysis (R core
Team 2019), and the circular statistics was performed using the “circular” package (Agostinelli
and Lund 2017).
Results
Hydrometeorology
The highest rainfall values were registered in the months of April 2018 (627 mm) and
May 2019 (total rainfall 481 mm). The drought months were October 2018 (did not rain) and
September 2019 (total rainfall 2.8 mm). The doughiest mean maximum temperature always
coincided with the month of October, reaching 34 ºC (Fig. 2a). The greatest amplitudes of
flooding in the Amazon River (Fig. 2b) were recorded in the months of April 2018 (3.11 m
height) and February 2019 (3.03 m height). When the rainfall in the region increases, the tide
level of the Amazon River also increases (r = 0.65) and the temperature decreases (r = -0.78).
There is a clear distinction between aquatic phase and terrestrial phase (Fig. 3, represented by
bars) in the forest. Aquatic phase begins in January and ends in July, influenced by increased
rainfall (r = 0.65) and tide level of the Amazon River (r = 0.56). Terrestrial phase has duration
of five months (August to December), when the volume of rainfall decreases and the maximum
tidal amplitudes of the Amazon River are below 3 m.
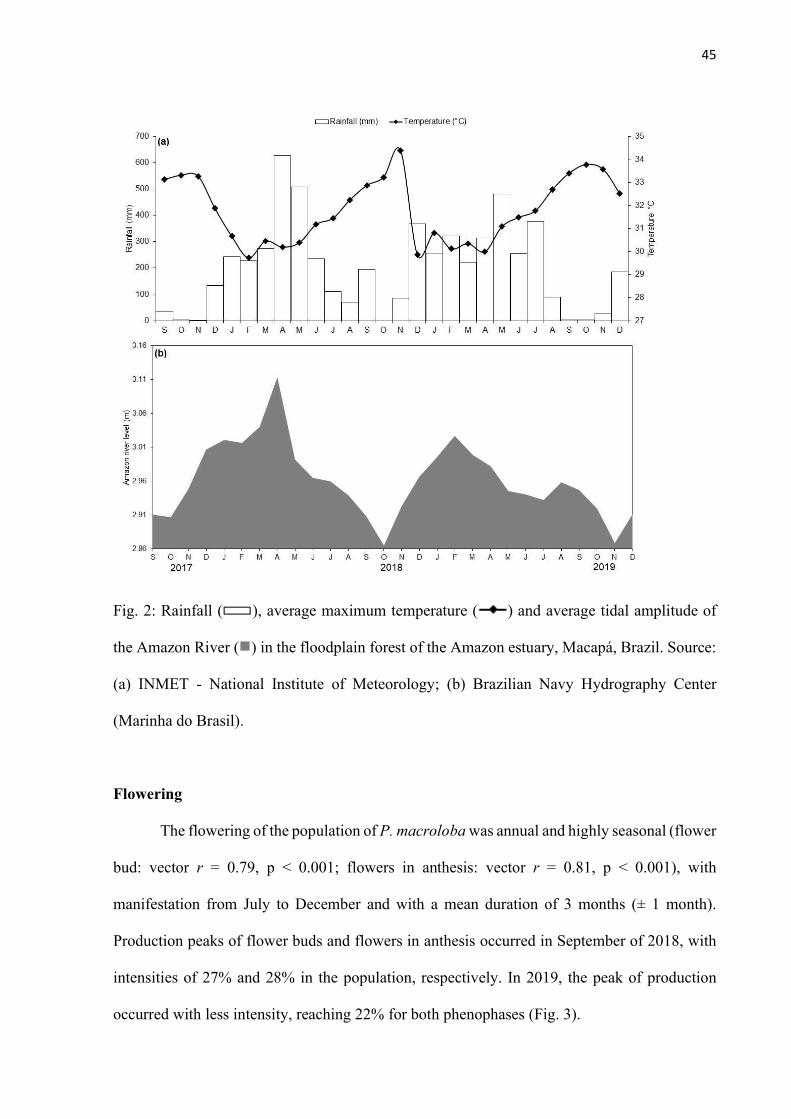
45
Fig. 2: Rainfall ( ), average maximum temperature ( ) and average tidal amplitude of
the Amazon River ( ) in the floodplain forest of the Amazon estuary, Macapá, Brazil. Source:
(a) INMET - National Institute of Meteorology; (b) Brazilian Navy Hydrography Center
(Marinha do Brasil).
Flowering
The flowering of the population of P. macroloba was annual and highly seasonal (flower
bud: vector r = 0.79, p < 0.001; flowers in anthesis: vector r = 0.81, p < 0.001), with
manifestation from July to December and with a mean duration of 3 months (± 1 month).
Production peaks of flower buds and flowers in anthesis occurred in September of 2018, with
intensities of 27% and 28% in the population, respectively. In 2019, the peak of production
occurred with less intensity, reaching 22% for both phenophases (Fig. 3).
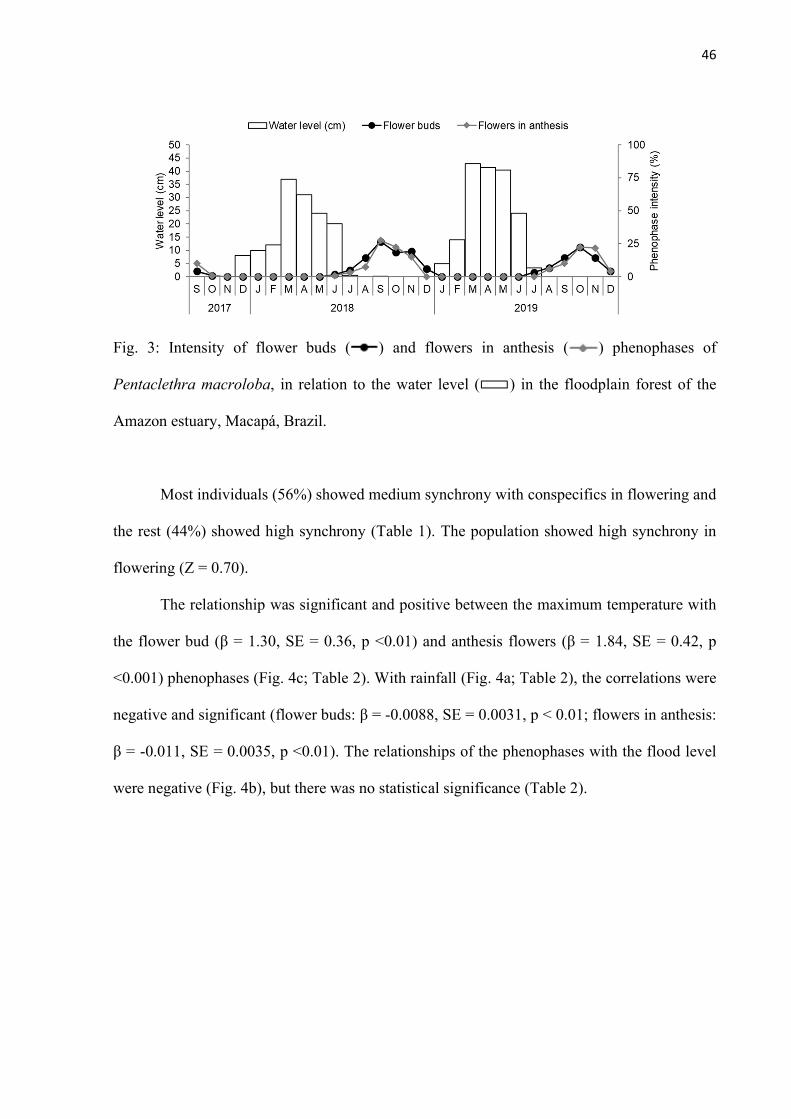
46
Fig. 3: Intensity of flower buds ( ) and flowers in anthesis ( ) phenophases of
Pentaclethra macroloba, in relation to the water level ( ) in the floodplain forest of the
Amazon estuary, Macapá, Brazil.
Most individuals (56%) showed medium synchrony with conspecifics in flowering and
the rest (44%) showed high synchrony (Table 1). The population showed high synchrony in
flowering (Z = 0.70).
The relationship was significant and positive between the maximum temperature with
the flower bud (β = 1.30, SE = 0.36, p <0.01) and anthesis flowers (β = 1.84, SE = 0.42, p
<0.001) phenophases (Fig. 4c; Table 2). With rainfall (Fig. 4a; Table 2), the correlations were
negative and significant (flower buds: β = -0.0088, SE = 0.0031, p < 0.01; flowers in anthesis:
β = -0.011, SE = 0.0035, p <0.01). The relationships of the phenophases with the flood level
were negative (Fig. 4b), but there was no statistical significance (Table 2).
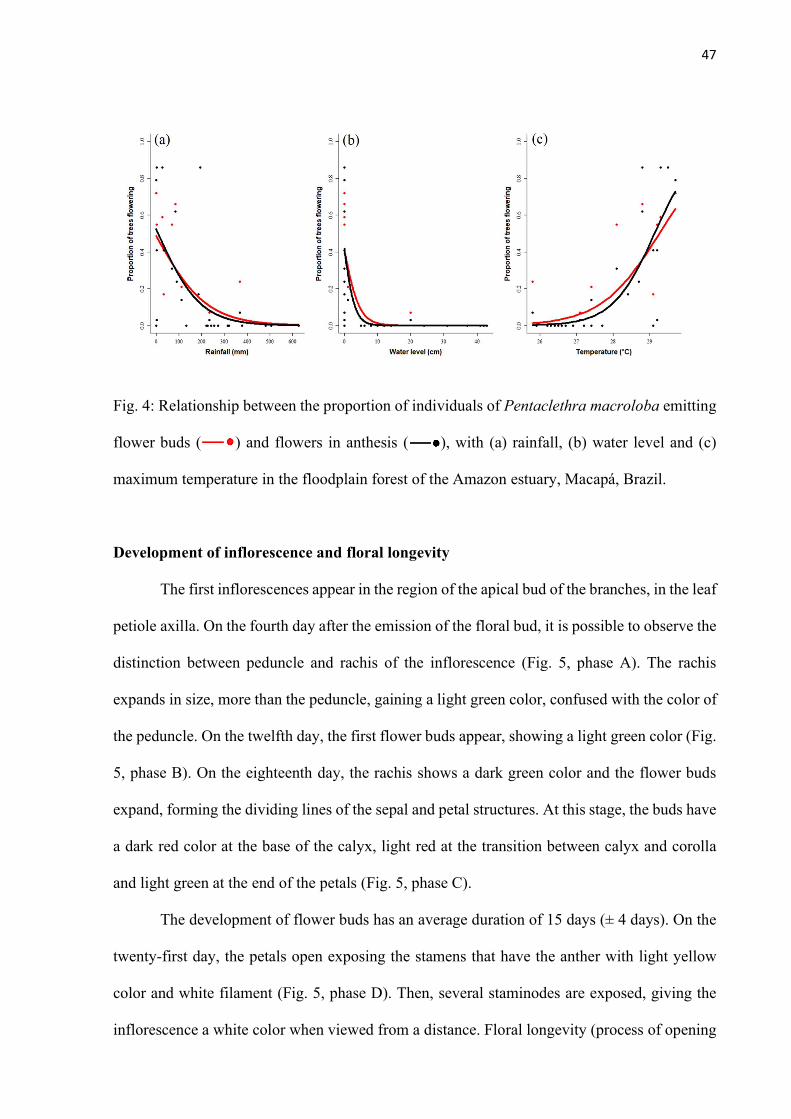
47
Fig. 4: Relationship between the proportion of individuals of Pentaclethra macroloba emitting
flower buds ( ) and flowers in anthesis ( ), with (a) rainfall, (b) water level and (c)
maximum temperature in the floodplain forest of the Amazon estuary, Macapá, Brazil.
Development of inflorescence and floral longevity
The first inflorescences appear in the region of the apical bud of the branches, in the leaf
petiole axilla. On the fourth day after the emission of the floral bud, it is possible to observe the
distinction between peduncle and rachis of the inflorescence (Fig. 5, phase A). The rachis
expands in size, more than the peduncle, gaining a light green color, confused with the color of
the peduncle. On the twelfth day, the first flower buds appear, showing a light green color (Fig.
5, phase B). On the eighteenth day, the rachis shows a dark green color and the flower buds
expand, forming the dividing lines of the sepal and petal structures. At this stage, the buds have
a dark red color at the base of the calyx, light red at the transition between calyx and corolla
and light green at the end of the petals (Fig. 5, phase C).
The development of flower buds has an average duration of 15 days (± 4 days). On the
twenty-first day, the petals open exposing the stamens that have the anther with light yellow
color and white filament (Fig. 5, phase D). Then, several staminodes are exposed, giving the
inflorescence a white color when viewed from a distance. Floral longevity (process of opening
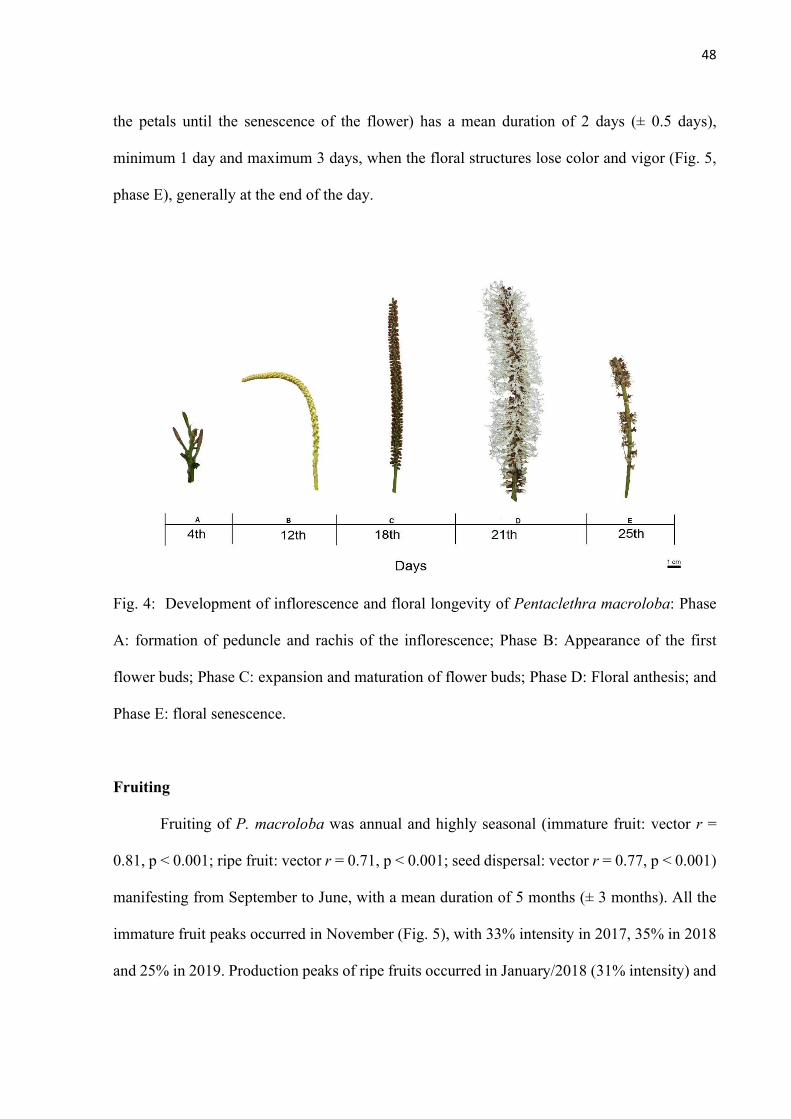
48
the petals until the senescence of the flower) has a mean duration of 2 days (± 0.5 days),
minimum 1 day and maximum 3 days, when the floral structures lose color and vigor (Fig. 5,
phase E), generally at the end of the day.
Fig. 4: Development of inflorescence and floral longevity of Pentaclethra macroloba: Phase
A: formation of peduncle and rachis of the inflorescence; Phase B: Appearance of the first
flower buds; Phase C: expansion and maturation of flower buds; Phase D: Floral anthesis; and
Phase E: floral senescence.
Fruiting
Fruiting of P. macroloba was annual and highly seasonal (immature fruit: vector r =
0.81, p < 0.001; ripe fruit: vector r = 0.71, p < 0.001; seed dispersal: vector r = 0.77, p < 0.001)
manifesting from September to June, with a mean duration of 5 months (± 3 months). All the
immature fruit peaks occurred in November (Fig. 5), with 33% intensity in 2017, 35% in 2018
and 25% in 2019. Production peaks of ripe fruits occurred in January/2018 (31% intensity) and
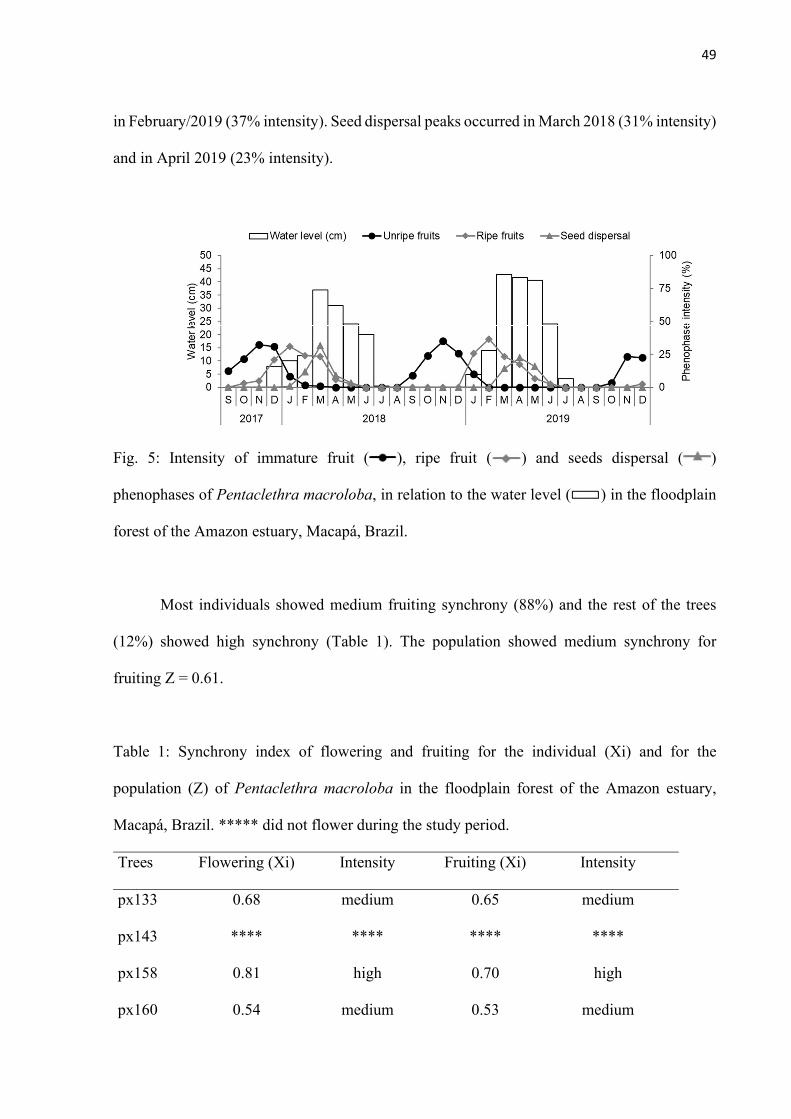
49
in February/2019 (37% intensity). Seed dispersal peaks occurred in March 2018 (31% intensity)
and in April 2019 (23% intensity).
Fig. 5: Intensity of immature fruit ( ), ripe fruit ( ) and seeds dispersal ( )
phenophases of Pentaclethra macroloba, in relation to the water level ( ) in the floodplain
forest of the Amazon estuary, Macapá, Brazil.
Most individuals showed medium fruiting synchrony (88%) and the rest of the trees
(12%) showed high synchrony (Table 1). The population showed medium synchrony for
fruiting Z = 0.61.
Table 1: Synchrony index of flowering and fruiting for the individual (Xi) and for the
population (Z) of Pentaclethra macroloba in the floodplain forest of the Amazon estuary,
Macapá, Brazil. ***** did not flower during the study period.
Trees Flowering (Xi) Intensity Fruiting (Xi) Intensity
px133 0.68 medium 0.65 medium
px143 **** **** **** ****
px158 0.81 high 0.70 high
px160 0.54 medium 0.53 medium
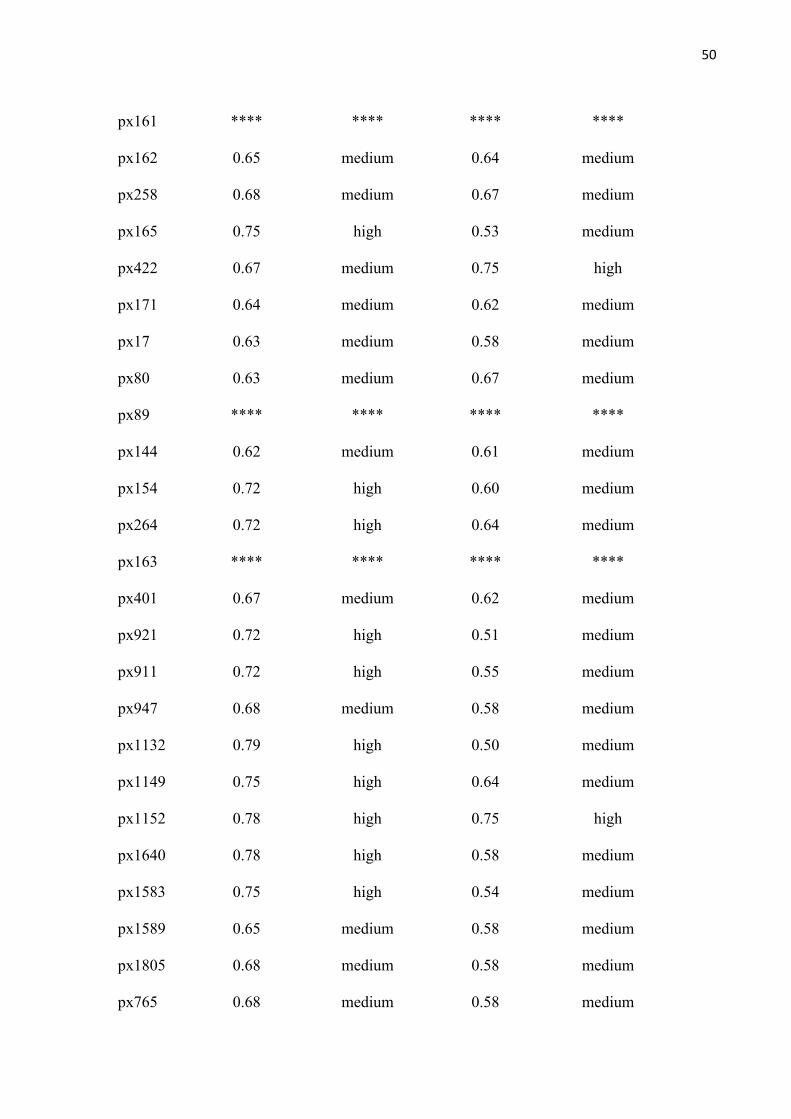
50
px161 **** **** **** ****
px162 0.65 medium 0.64 medium
px258 0.68 medium 0.67 medium
px165 0.75 high 0.53 medium
px422 0.67 medium 0.75 high
px171 0.64 medium 0.62 medium
px17 0.63 medium 0.58 medium
px80 0.63 medium 0.67 medium
px89 **** **** **** ****
px144 0.62 medium 0.61 medium
px154 0.72 high 0.60 medium
px264 0.72 high 0.64 medium
px163 **** **** **** ****
px401 0.67 medium 0.62 medium
px921 0.72 high 0.51 medium
px911 0.72 high 0.55 medium
px947 0.68 medium 0.58 medium
px1132 0.79 high 0.50 medium
px1149 0.75 high 0.64 medium
px1152 0.78 high 0.75 high
px1640 0.78 high 0.58 medium
px1583 0.75 high 0.54 medium
px1589 0.65 medium 0.58 medium
px1805 0.68 medium 0.58 medium
px765 0.68 medium 0.58 medium
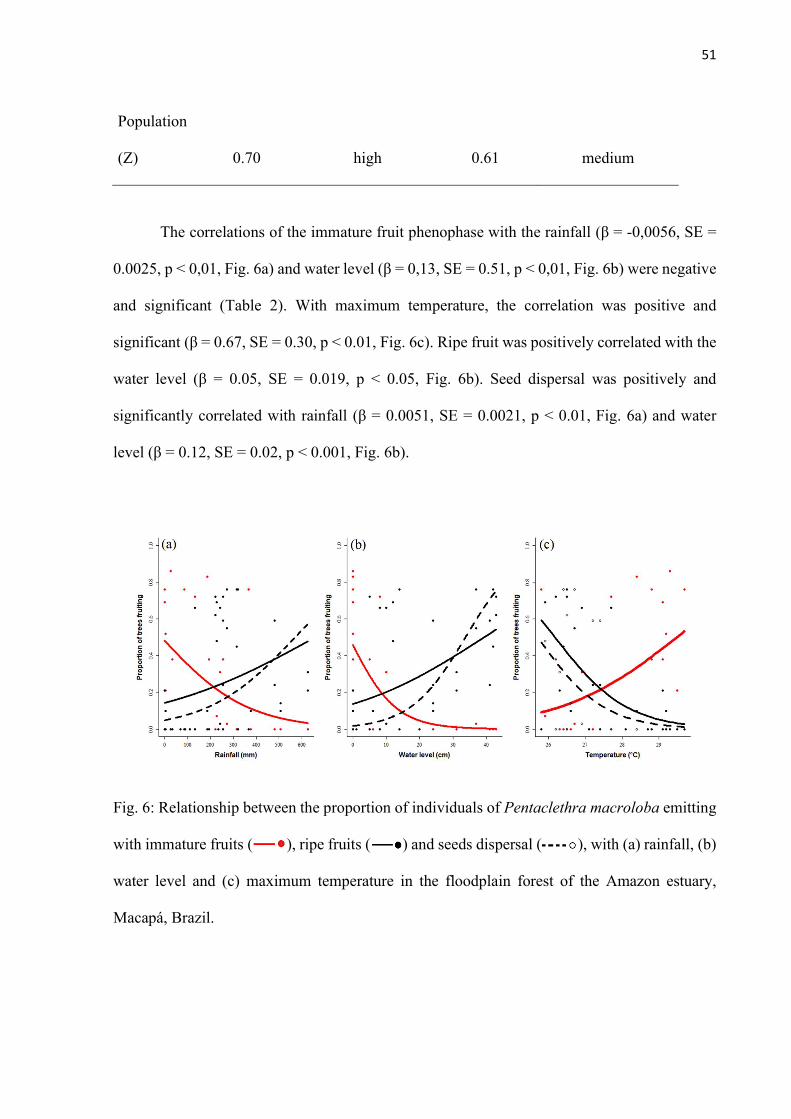
51
Population
(Z) 0.70 high 0.61 medium
The correlations of the immature fruit phenophase with the rainfall (β = -0,0056, SE =
0.0025, p < 0,01, Fig. 6a) and water level (β = 0,13, SE = 0.51, p < 0,01, Fig. 6b) were negative
and significant (Table 2). With maximum temperature, the correlation was positive and
significant (β = 0.67, SE = 0.30, p < 0.01, Fig. 6c). Ripe fruit was positively correlated with the
water level (β = 0.05, SE = 0.019, p < 0.05, Fig. 6b). Seed dispersal was positively and
significantly correlated with rainfall (β = 0.0051, SE = 0.0021, p < 0.01, Fig. 6a) and water
level (β = 0.12, SE = 0.02, p < 0.001, Fig. 6b).
Fig. 6: Relationship between the proportion of individuals of Pentaclethra macroloba emitting
with immature fruits ( ), ripe fruits ( ) and seeds dispersal ( ), with (a) rainfall, (b)
water level and (c) maximum temperature in the floodplain forest of the Amazon estuary,
Macapá, Brazil.
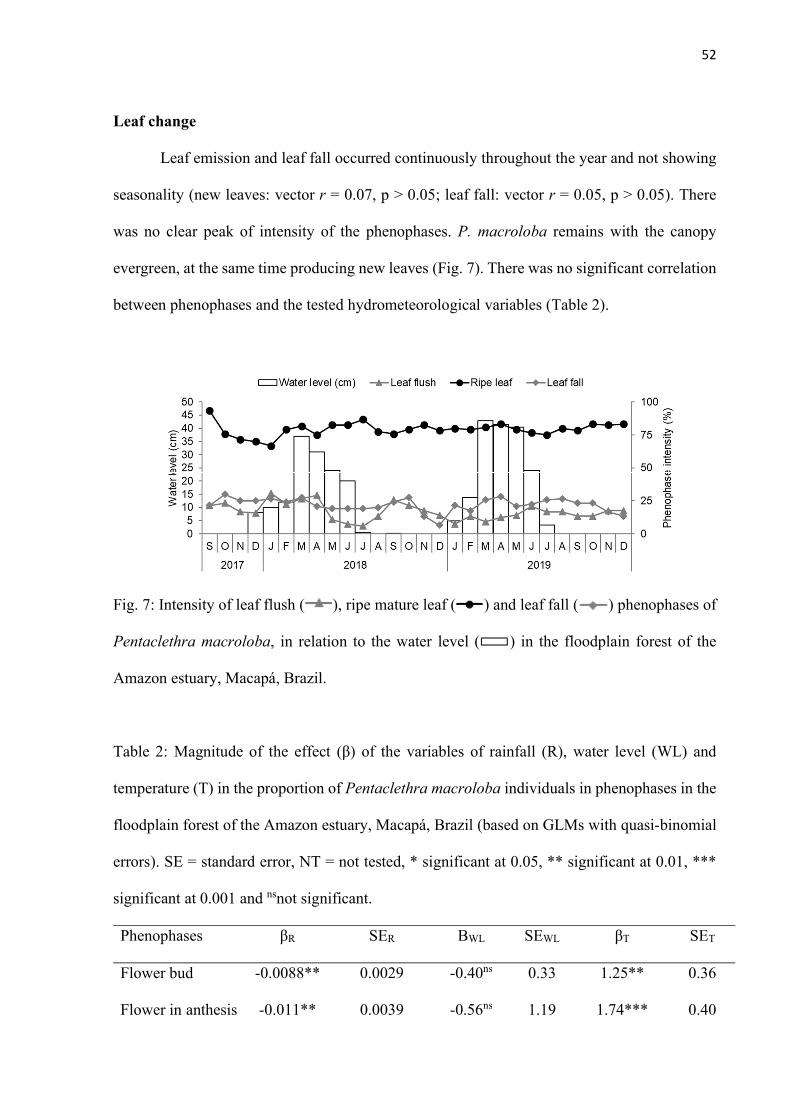
52
Leaf change
Leaf emission and leaf fall occurred continuously throughout the year and not showing
seasonality (new leaves: vector r = 0.07, p > 0.05; leaf fall: vector r = 0.05, p > 0.05). There
was no clear peak of intensity of the phenophases. P. macroloba remains with the canopy
evergreen, at the same time producing new leaves (Fig. 7). There was no significant correlation
between phenophases and the tested hydrometeorological variables (Table 2).
Fig. 7: Intensity of leaf flush ( ), ripe mature leaf ( ) and leaf fall ( ) phenophases of
Pentaclethra macroloba, in relation to the water level ( ) in the floodplain forest of the
Amazon estuary, Macapá, Brazil.
Table 2: Magnitude of the effect (β) of the variables of rainfall (R), water level (WL) and
temperature (T) in the proportion of Pentaclethra macroloba individuals in phenophases in the
floodplain forest of the Amazon estuary, Macapá, Brazil (based on GLMs with quasi-binomial
errors). SE = standard error, NT = not tested, * significant at 0.05, ** significant at 0.01, ***
significant at 0.001 and nsnot significant.
Phenophases βR SER ΒWL SEWL βT SET
Flower bud -0.0088** 0.0029 -0.40ns 0.33 1.25** 0.36
Flower in anthesis -0.011** 0.0039 -0.56ns 1.19 1.74*** 0.40
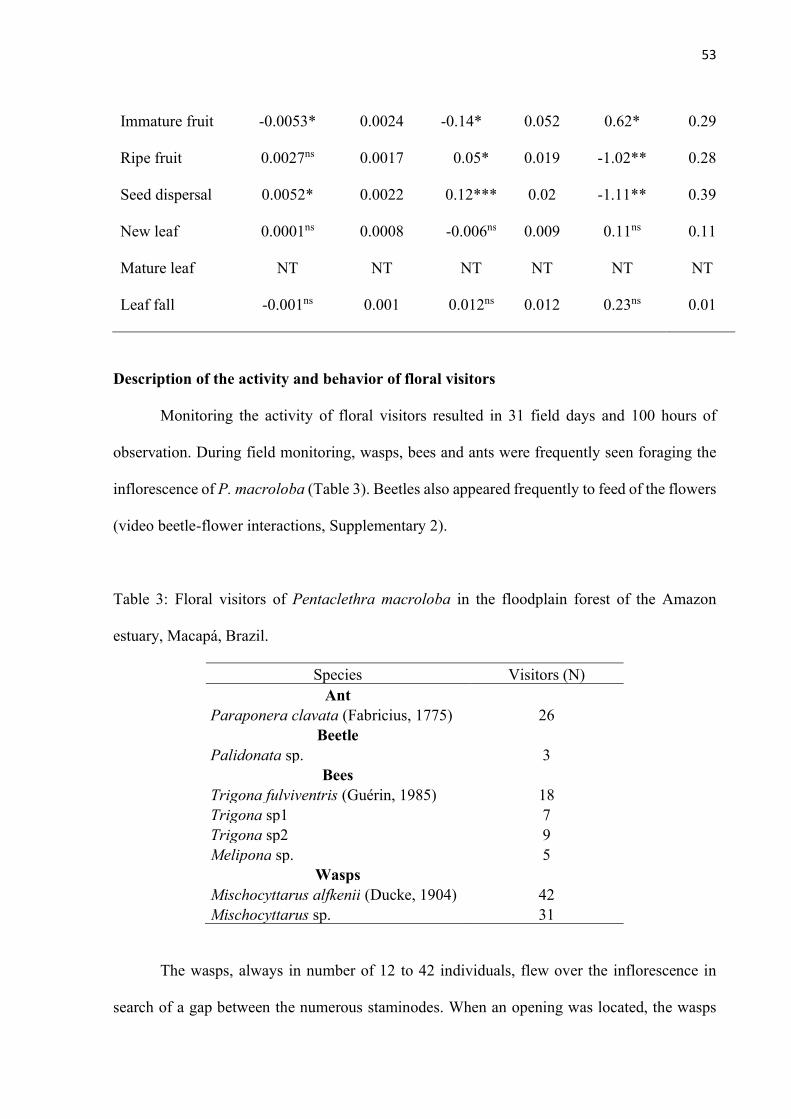
53
Immature fruit -0.0053* 0.0024 -0.14* 0.052 0.62* 0.29
Ripe fruit 0.0027ns 0.0017 0.05* 0.019 -1.02** 0.28
Seed dispersal 0.0052* 0.0022 0.12*** 0.02 -1.11** 0.39
New leaf 0.0001ns 0.0008 -0.006ns 0.009 0.11ns 0.11
Mature leaf NT NT NT NT NT NT
Leaf fall -0.001ns 0.001 0.012ns 0.012 0.23ns 0.01
Description of the activity and behavior of floral visitors
Monitoring the activity of floral visitors resulted in 31 field days and 100 hours of
observation. During field monitoring, wasps, bees and ants were frequently seen foraging the
inflorescence of P. macroloba (Table 3). Beetles also appeared frequently to feed of the flowers
(video beetle-flower interactions, Supplementary 2).
Table 3: Floral visitors of Pentaclethra macroloba in the floodplain forest of the Amazon
estuary, Macapá, Brazil.
Species Visitors (N) Ant
Paraponera clavata (Fabricius, 1775) 26 Beetle
Palidonata sp. 3 Bees
Trigona fulviventris (Guérin, 1985) 18 Trigona sp1 7 Trigona sp2 9 Melipona sp. 5
Wasps Mischocyttarus alfkenii (Ducke, 1904) 42 Mischocyttarus sp. 31
The wasps, always in number of 12 to 42 individuals, flew over the inflorescence in
search of a gap between the numerous staminodes. When an opening was located, the wasps

54
landed on the staminodes and went towards the corolla of the flower (where the stamens and
ovary are located) and remained there for 5 to 7 minutes foraging and vibrating the hind legs
(video wasp-flower interactions, Supplementary 3). Bees explored the flowers in small groups
of individuals, from three to five insects, and always flew in circles around the inflorescence to
reach the interior of the flower. The bees remained inside the flower for 8 to 11 minutes and
they alternated between several flowers at the same time. Bees were seen foraging flower buds,
in addition to flowers, together with the ants (video bee-ant-flower interactions, supplementary
4).
Wasps frequently visited the flowers between the hours of 12:00 to 14:00 pm, with peak
visitation at 12:00 pm (Fig. 8a). The preferred time of the bees occurred between 08:00 to 12:30
pm, with peak visitation at 10:00 am (Fig. 8b). Ants had a wider visitation time, but the peak
visitation occurred at 11:00 am (Fig. 8c). When the wasps identified other intruder insects trying
to exploit an inflorescence dominated by the group, they exhibited aggressive behavior and
went to attack the intruders with their stingers. This behavior may justify the low number of
species that visited the inflorescence of P. macroloba (Table 3).
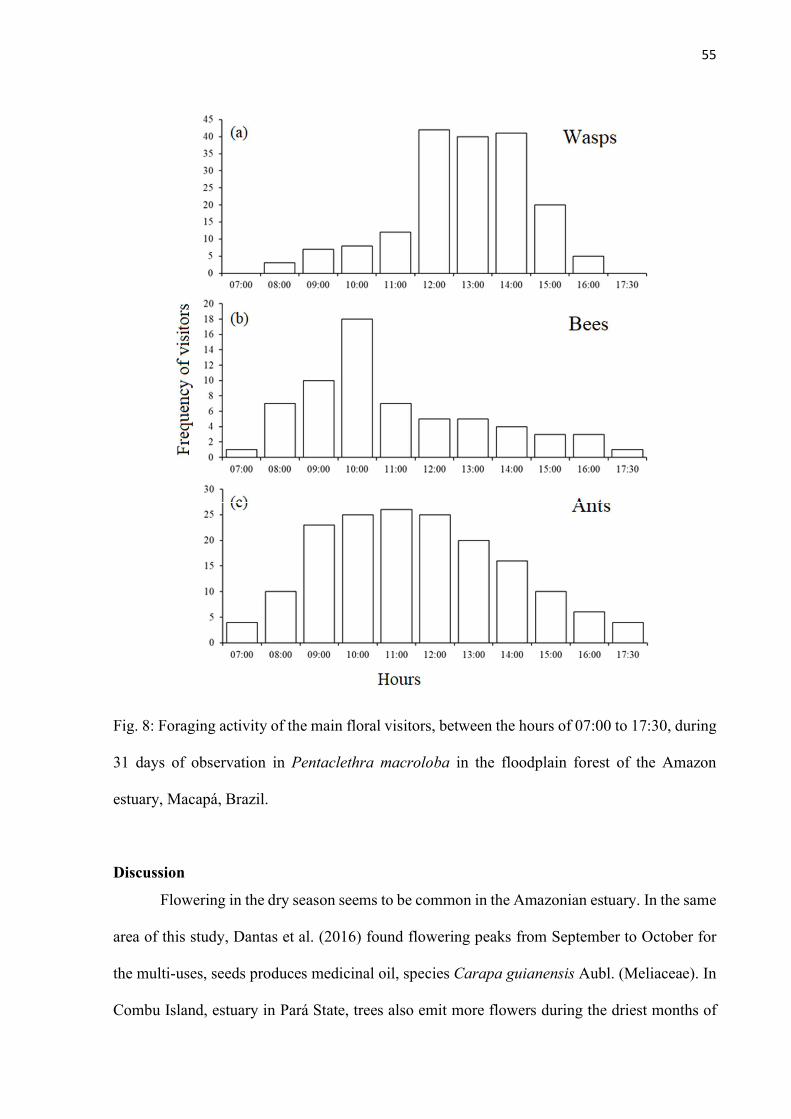
55
Fig. 8: Foraging activity of the main floral visitors, between the hours of 07:00 to 17:30, during
31 days of observation in Pentaclethra macroloba in the floodplain forest of the Amazon
estuary, Macapá, Brazil.
Discussion
Flowering in the dry season seems to be common in the Amazonian estuary. In the same
area of this study, Dantas et al. (2016) found flowering peaks from September to October for
the multi-uses, seeds produces medicinal oil, species Carapa guianensis Aubl. (Meliaceae). In
Combu Island, estuary in Pará State, trees also emit more flowers during the driest months of

56
the year, from July to December (Cattanio et al. 2004). The flowering period of P. macroloba
differs from the trees of flooded forests in the Central Amazon. Some trees produce flowers at
the beginning of the flood period (Worbes 1997) and others at the peak of the river flood
(Schöngart et al. 2002). However, Haugaasen and Peres (2005) studied the phenology of
arboreal communities in the várzea and igapó forests of the lower Purus River, west of the
Central Amazon, and found that species from both environments showed marked flowering
peaks during the dry season, from July to October.
Flower production in the dry season, where temperatures are high and the rainfall is low,
becomes a reproductive trigger for Pentaclethra macroloba. The intense rains of the rainy
season in the Amazon can damage the floral structure, affecting pollen viability and stigma
receptivity (Dantas et al. 2016). For floral visitors of P. macroloba, such as wasps and bees, the
dry season is ideal for foraging flowers, because rain interferes with the pollination activity of
the insects. In a broad review of the effects of rainfall on pollinator-plant interactions, Lawson
and Rands (2019) found that rainfall causes mechanical restrictions on the insect wings and
interferes with their sensory signals. In general, rainfall can cause various effects on insects and
other arthropods, which can seriously damage trophic interactions (Chen et al. 2019).
P. macroloba population showed highly synchronized flowering between individuals.
Synchronized flowering favors cross-pollination (Nilsson and Wästljung 1987; Stevenson et al.
2008; Narbona et al. 2011), as pollinators can forage several individuals during the day, while
mixing and transferring pollen from neighboring trees (Augspurger 1981; Braga and Collevatti
2011). The low flowering intensity of P. macroloba, between 23% to 25%, indicates that the
species does not show mass-flowering, in which the tree produces a large number of flowers at
once (Augspurger 1980). Although the population of P. macroloba shows synchronism in
flowering, the branches of the tree making flowering are not synchronized. When one branches

57
is in floral senescence, another is already bearing new inflorescences, ensuring continuous
production and of low intensity during the flowering period.
Synchronized flowering, however, can also be a disadvantage for trees, due to attract
the attention of predators (Rathcke and Lacey 1985). During the flowering period of P.
macroloba, it was common to observe undeveloped inflorescences on the forest floor. This is
probably due to the presence of predators, such as beetles found in inflorescences. For Kevan
and Baker (1983), beetles are common in trees with synchronized flowering, as they feed of the
plant sap and floral structures. However, several other factors can contribute to flower abortion
(Stephenson 1981; Bawa and Webb 1984), such as water deficiency (Rapoport et al. 2012).
This factor could affect more directly the development of P. macroloba inflorescence, since its
flowering is concentrated in the dry season.
The period that trees remain with its flowers open is an important aspect to understand
the plant-pollinator interactions (Primack 1985). The floral development of P. macroloba can
last up to 20 days and anthesis three days. This cost can be rewarded, in trees with low floral
longevity such as P. macroloba, for a high frequency of pollinators (Spigler 2017), no being
necessary an extended anthesis for several days, which avoids geitonogamy and the
displacement of nutritional energy for the flower.
The behavior shown by wasps and bees when foraging the flower, lead to the assumption
that they are legitimate pollinators of P. macroloba. At the La Selva Biological Station in Costa
Rica, Bawa et al. (1985) found small bees pollinating P. macroloba. These little Hymenoptera
are attracted by the sweet smell that the flowers release (personal observation). The nectar
guides present on the petals (Dantas et al. 2021) attract insects towards the nectar gland at the
base of the ovary (Barros et al. 2017). This increases the chances of the reproductive organs of
the flower being touched by the movement of the insect when they enter the corolla.

58
Ants were also frequently observed foraging the flower buds of P. macroloba. Perhaps
the ants are attracted by the extrafloral gland present at the base of the P. macroloba leaf (Dantas
et al. 2021). Extrafloral nectar is rich in carbohydrates and amino acids, essential energy source
for canopy ants (Byk and Del-Claro 2011). Ants of the species Paraponera clavata (Fabricius,
1775) have mutualistic interactions with individuals of P. macroloba from Central America.
The tree offers extrafloral nectary and shelter for the ant and the ant rewards protecting the plant
of the herbivores (Bennett and Breed 1985).
Other studies also show the predominance of species fruiting and dispersing propagules
at the peak of flooding in the forest, both in Central Amazonia (Worbes 1997; Schöngart et al.
2002; Haugaasen and Peres 2005; Ferreira and Parolin 2007; Hawes and Peres 2016) and in the
Amazon estuary (Cattanio et al. 2004; Dantas et al. 2016). This shows that the flood pulse is an
important driver of the phenological patterns of the Amazon flooded forests.
Fruiting of P. macroloba was influenced by the increased of the rainfall and water level
inside the forest. In floodplain forest, the flood pulse and rainfall act as synchronizers of the
phenological rhythms of many species, mainly in the stage of ripening of the fruits (Parolin et
al. 2010). This period is ideal for species to disperse their propagules, because many trees
depend of the ichthyofauna (Kubitzki and Ziburski 1994) and river water to transport their seeds
(Moegenburg 2002; Cunha et al. 2017).
The ripening time of the fruits reflects on the successful dispersion of the seeds (Rathcke
and Lacey 1985). The high temperatures that the fruits receive during the development in the
dry season, favor the maturation and desiccation of the fruits, which is essential for the release
of the seeds when the fruits are ripe in the rainy season. According to van Rheede van
Oudtshoorn and van Rooyen (1999), Fabaceae trees which release their seeds by the explosive
dehiscence of the fruits, need water for the cells of the dry tissue of the fruits to absorb the
liquid and cause tension in the valves. As a result, the valves twist in the abscission zone of the

59
fruit and expel the seeds away from the tree. In the case of P. macroloba, the seeds can be
expelled up to 10 m (Hartshorn 1983).
The synchrony of the seed dispersal with the rainy season and flood peak of the Amazon
River, allows P. macroloba to disperse its seeds over a long distance by hydrocoria (Williamson
and Costa 2000). The flattened shape (deltoid) and the accumulation of air between the
cotyledons, allow the seed to glide when leaving the fruit, and float when reaching the water of
the Amazon estuary obtaining greater efficiency in dispersion (Dantas et al. 2021). Correa et
al. (2018) suggest that the synchronization of fruiting time with the flooding peak contributed
to the evolution of the patterns of hydrocoric and ichthyochoric dispersion found in the flooded
forests of the Amazon. The authors emphasize that these two types of dispersion also directed
the phenological behavior of the trees, so that they respond presenting more attractive and
fleshy fruits for the fish and mechanisms adapted to buoyancy in water.
P. macroloba remains evergreen while unfolding and losing leaves, and there is no clear
relationship with the hydrometeorological variables. The characteristic of presenting always
ripe leaves and continuous production of new leaves, can decrease the herbivory rate of flowers
and fruits. Many leaves in the canopy can block and hide the reproductive elements from the
view of herbivores. In addition, Klimas et al. (2012) suggest that the continuous production of
leaves increases the photosynthetic area of the canopy, allowing the species to stock higher
amounts of carbohydrates in the leaves to use in the reproductive phase.
Conclusions
The high degree of synchrony of the reproductive phenophases with the seasons of the
year, suggests that P. macroloba has high adaptation and reproductive strategies in the tidal
floodplains of the Amazon estuary, this may be one of the factors that explain its dominance in
this environment.

60
The seasonality of the rainy season has a major influence on the reproductive phenology
of P. macroloba. The decrease in rainfall from September to December causes an increase in
air temperature, which is essential to stimulate the flowering and the production of the first
fruits. The increase in rainfall favors fruit ripening and triggers their explosive dehiscence
mechanism. The flood pulse of the Amazon River may be a synchronizing agent in the P.
macroloba seed dispersal phenophase, this environmental drive is essential for the transport of
seeds and colonization of other areas.
Our results will be fundamental to assist in the good practices of P. macroloba seed
management and in the valuation of non-wood forest products in estuarine floodplain forests.
Acknowledgements
We thank Conselho Nacional de Desenvolvimento Científico e Tecnológico, Programa
de Pós-graduação em Ecologia (PPGEco/INPA), Instituto Nacional de Pesquisas da Amazônia,
Grupo Ecologia, Monitoramento e Uso Sustentável de Áreas Úmidas
(MAUA/PELD/CNPq/FAPEAM) and students of the Forest Engineering course/ Instituto
Macapaense de Ensino Superior (Gabriel, Pedro and Tailon).
Conflicts of interest
The authors declare that there is no any conflict of interest.
References
Agostinelli C, Lund U (2017) Circular Statistics, from ``Topics in circular Statistics’’
Alvares CA, Stape JL, Sentelhas PC, et al (2014) Köppen’s climate classification map for
Brazil. Meteorol Zeitschrift 22:711–728. https://doi.org/10.1127/0941-2948/2013/0507
Augspurger CK (1983) Phenology, flowering synchrony, and fruit set of six Neotropical shrubs.

61
Biotropica 15:257. https://doi.org/10.2307/2387650
Augspurger CK (1981) Reproductive synchrony of a tropical shrub: experimental studies on
effects of pollinators and seed predators in Hybanthus prunifolius (Violaceae). Ecology
62:775–788. https://doi.org/10.2307/1937745
Augspurger CK (1980) Mass-flowering of a tropical shrub (Hybanthus prunifolius): influence
on pollinator attraction and movement. Evolution (N Y) 34:475–488.
https://doi.org/10.1111/j.1558-5646.1980.tb04837.x
Banov D, Banov F, Bassani AS (2014) Case series: the effectiveness of fatty acids from pracaxi
oil in a topical silicone base for scar and wound therapy. Dermatol Ther (Heidelb) 4:259–
269. https://doi.org/10.1007/s13555-014-0065-y
Barros TC, Pedersoli GD, Paulino J V, Teixeira SP (2017) In the interface of caesalpinioids and
mimosoids: comparative floral development elucidates shared characters in
Dimorphandra mollis and Pentaclethra macroloba (Leguminosae). Am J Bot 104:218–
232. https://doi.org/10.3732/ajb.1600308
Bawa KS, Bullock SH, Perry DR, et al (1985) Reproductive biology of tropical lowland rain
forest trees. II. pollination systems. Am J Bot 72:346–356.
https://doi.org/10.2307/2443527
Bawa KS, Webb CJ (1984) Flower, fruit and seed abortion in tropical forest trees: implications
for the evolution of paternal and maternal reproductive patterns. Am J Bot 71:736.
https://doi.org/10.2307/2443371
Bennett B, Breed MD (1985) On the association between Pentaclethra macroloba
(Mimosaceae) and Paraponera clavata (Hymenoptera: Formicidae) Colonies. Biotropica
17:253. https://doi.org/10.2307/2388226
Braga AC, Collevatti RG (2011) Temporal variation in pollen dispersal and breeding structure
in a bee-pollinated Neotropical tree. Heredity (Edinb) 106:911–919.

62
https://doi.org/10.1038/hdy.2010.134
Byk J, Del-Claro K (2011) Ant-plant interaction in the Neotropical savanna: direct beneficial
effects of extrafloral nectar on ant colony fitness. Popul Ecol 53:327–332.
https://doi.org/10.1007/s10144-010-0240-7
Carim M de JV, Wittmann FK, Piedade MTF, et al (2016) Composition, diversity, and structure
of tidal “Várzea” and “Igapó” floodplain forests in eastern Amazonia, Brazil. Brazilian J
Bot. https://doi.org/10.1007/s40415-016-0315-6
Cattanio JH, Anderson AB, Rombold JS, Nepstad DC (2004) Phenology, litterfall, growth, and
root biomass in a tidal floodplain forest in the Amazon estuary. Rev Bras Botânica 27:703–
712. https://doi.org/10.1590/S0100-84042004000400010
Chen C, Harvey JA, Biere A, Gols R (2019) Rain downpours affect survival and development
of insect herbivores: the specter of climate change? Ecology 100:1–10.
https://doi.org/10.1002/ecy.2819
CHM (2019) Tábuas de Maré. https://www.marinha.mil.br/chm/tabuas-de-mare. Accessed 12
Dec 2019
Correa SB, de Oliveira PC, Nunes da Cunha C, et al (2018) Water and fish select for fleshy
fruits in tropical wetland forests. Biotropica 50:312–318.
https://doi.org/10.1111/btp.12524
Cunha AC, Mustin K, dos Santos ES, et al (2017) Hydrodynamics and seed dispersal in the
lower Amazon. Freshw Biol 1–9. https://doi.org/10.1111/fwb.12982
Dantas AR, Guedes MC, Vasconcelos CC, et al (2021) Morphology, germination, and
geographic distribution of Pentaclethra macroloba (Fabaceae ): a hyperdominant
Amazonian tree. Rev Biol Trop 69:1–16
Dantas AR, Lira-Guedes AC, Mustin K, et al (2016) Phenology of the multi-use tree species
Carapa guianensis in a floodplain forest of the Amazon Estuary. Acta Bot Brasilica

63
30:618–627. https://doi.org/10.1590/0102-33062016abb0282
Dantas AR, Marangon LC, Guedes MC, et al (2017) Spatial distribution of a population of
Pentaclethra macroloba (Willd.) Kuntze in a floodplain forest of the Amazon estuary. Rev
Árvore 41:1–11. https://doi.org/10.1590/1806-90882017000400006
Ferreira LV, Parolin P (2007) Tree phenology in central amazonian floodplain forests: effects
of water level fluctuation and precipitation at community and population level. Botânica
Pesqui 58:139–155. https://doi.org/10.2307/2261447
Fouks B, Wagoner KM (2019) Pollinator parasites and the evolution of floral traits. Ecol Evol
9:6722–6737. https://doi.org/10.1002/ece3.4989
Fournier L (1974) Un método cuantitativo para la medición de características fenológicas en
árboles. Turrialba 24:422–423
Freitas J da L, Malheiros MA de B, Vasconcelos PCS (2003) Processsos fenológicos de
Taperebá (Spondias mombin L.) e Pracaxi (Pentaclethra macroloba (Wild) O.Kuntz) em
ecossistema florestal de várzea na Ilha do Pará, Afuá, Pará. Rev Ciências Agrárias 163–
172
Hartshorn GS (1983) Pentaclethra macroloba (Gavilan). In: Janzen DH (ed) Costa Rican
Natural History. Universityof Chicago Press, pp 301–303
Haugaasen T, Peres CA (2005) Tree phenology in adjacent amazonian flooded and unflooded
forests. Biotropica 37:620–630. https://doi.org/10.1111/j.1744-7429.2005.00079.x
Hawes JE, Peres CA (2016) Patterns of plant phenology in Amazonian seasonally flooded and
unflooded forests. Biotropica 48:465–475. https://doi.org/10.1111/btp.12315
IBGE - Instituto Brasileiro de Geografia e Estatística (2012) Manual técnico da vegetação
brasileira : sistema fitogeográfico : inventário das formações florestais e campestres :
técnicas e manejo de coleções botânicas : procedimentos para mapeamentos, 2nd edn.
IBGE, Rio de Janeiro

64
INMET (2019) Banco de Dados Meteorológicos para Ensino e Pesquisa.
nmet.gov.br/portal/index.php?r=bdmep/bdmep. Accessed 20 Dec 2019
Junk WJ (2001) The flood pulse concept of large rivers: learning from the tropics. SIL
Proceedings, 1922-2010 27:3950–3953.
https://doi.org/10.1080/03680770.1998.11901733
Junk WJ, Piedade MTF (2010) An introduction to South American wetland forests: distribution,
definitions and general characterization. In: Amazonian floodplain forests: ecophysiology,
biodiversity and sustainable management. pp 3–25
Junk WJ, Piedade MTF, Lourival R, et al (2014) Brazilian wetlands: their definition,
delineation, and classification for research, sustainable management, and protection.
Aquat Conserv Mar Freshw Ecosyst 24:5–22. https://doi.org/10.1002/aqc.2386
Kevan PG, Baker HG (1983) Insects as flower visitors and pollinators. Annu Rev Entomol
28:407–453. https://doi.org/10.1146/annurev.en.28.010183.002203
Klimas CA, Kainer KA, Wadt LH, et al (2012) Control of Carapa guianensis phenology and
seed production at multiple scales: a five-year study exploring the influences of tree
attributes, habitat heterogeneity and climate cues. J Trop Ecol 28:105–118.
https://doi.org/10.1017/S0266467411000630
Kubitzki K, Ziburski A (1994) Seed dispersal in flood plain forests of Amazonia. Biotropica
26:30. https://doi.org/10.2307/2389108
Lawson DA, Rands SA (2019) The effects of rainfall on plant–pollinator interactions.
Arthropod Plant Interact 13:561–569. https://doi.org/10.1007/s11829-019-09686-z
Lieth H (1974) Phenology and seasonality modeling. Springer Berlin Heidelberg, Berlin,
Heidelberg
Luize BG, Magalhães JLL, Queiroz H, et al (2018) The tree species pool of Amazonian wetland
forests: which species can assemble in periodically waterlogged habitats? PLoS One 13:1–

65
13. https://doi.org/10.1371/journal.pone.0198130
Malucelli TS, Maia FR, Varassin IG (2018) Breeding system and pollination of Pleroma
trichopodum DC. (Melastomataceae): A potential species for the restoration of atlantic
forest in Southern Brazil. Acta Bot Brasilica 32:402–409. https://doi.org/10.1590/0102-
33062018abb0103
Mendoza I, Peres CA, Morellato LPC (2017) Continental-scale patterns and climatic drivers of
fruiting phenology: a quantitative Neotropical review. Glob Planet Change 148:227–241.
https://doi.org/10.1016/j.gloplacha.2016.12.001
Moegenburg SM (2002) Spatial and temporal variation in hydrochory in Amazonian floodplain
forest. Biotropica 34:606. https://doi.org/10.1646/0006-
3606(2002)034[0606:satvih]2.0.co;2
Morellato LPC, Alberti LF, Hudson IL (2010a) Applications of circular statistics in plant
phenology: a case studies approach. In: Hudson IL, Keatley MR (eds) Phenological
Research. Springer Netherlands, Dordrecht, pp 339–359
Morellato LPC, Camargo MGG, D’Eça Neves FF, et al (2010b) The influence of sampling
method, sample size, and frequency of observations on plant phenological patterns and
interpretation in tropical forest trees. In: Hudson IL, Keatley MR (eds) Phenological
research: methods for environmental and climate change analysis. Springer Netherlands,
Dordrecht, pp 99–121
Morellato LPC, Camargo MGG, Gressler E (2013) A review of plant phenology in South and
Central America. In: Schwartz MD (ed) Phenology: an integrative environmental science.
Springer Netherlands, Dordrecht, pp 91–113
Morellato LPC, Talora DC, Takahasi A, et al (2000) Phenology of Atlantic rain forest trees: a
comparative study. Biotropica 32:811–823. https://doi.org/10.1111/j.1744-
7429.2000.tb00620.x

66
Narbona E, Ortiz PL, Arista M (2011) Linking self-incompatibility, dichogamy, and flowering
synchrony in two Euphorbia species: alternative mechanisms for avoiding self-
fertilization? PLoS One 6:e20668. https://doi.org/10.1371/journal.pone.0020668
Newbery DM, Chuyong GB, Zimmermann L (2006) Mast fruiting of large ectomycorrhizal
African rain forest trees: importance of dry season intensity, and the resource-limitation
hypothesis. New Phytol 170:561–579. https://doi.org/10.1111/j.1469-8137.2006.01691.x
Newstrom LE, Frankie GW, Baker HG (1994) A new classification for plant phenology based
on flowering patterns in lowland tropical rain forest trees at La Selva, Costa Rica.
Biotropica 26:141. https://doi.org/10.2307/2388804
Nilsson SG, Wästljung U (1987) Seed predation and cross-pollination in mast-seeding beech
(Fagus sylvatica) Patches. Ecology 68:260–265. https://doi.org/10.2307/1939256
Oliveira PE, Rech AR (2018) Floral biology and pollination in Brazil: History and possibilities.
Acta Bot Brasilica 32:321–328. https://doi.org/10.1590/0102-33062018abb0255
Parolin P, Wittmann F, Schöngart J (2010) Tree phenology in Amazonian floodplain forests.
In: Junk WJ, Piedade MTF, Wittmann F, et al. (eds) Amazonian floodplain forests:
ecophysiology, biodiversity and sustainable management, 1st edn. Springer Netherlands,
pp 105–126
Pinto ER (2014) Solos, hidrologia e estrutura populacional de pracuubeiras em florestas de
várzea do estuário amazônico. Dissertation, Universidade Federal do Amapá
PPBio (2019) Protocolo 1 - Insetos capturados com armadilhas atrativas. http://ppbio.museu-
goeldi.br/?q=pt-br/protocolo-1-insetos-capturados-com-armadilhas-atrativas. Accessed 6
Jan 2019
Primack RB (1985) Longevity of individual flowers. Annu Rev Ecol Syst 16:15–37.
https://doi.org/10.1146/annurev.es.16.110185.000311
R core Team (2019) R: A Language and Environment for Statistical Computing

67
Rapoport HF, Hammami SBM, Martins P, et al (2012) Influence of water deficits at different
times during olive tree inflorescence and flower development. Environ Exp Bot 77:227–
233. https://doi.org/10.1016/j.envexpbot.2011.11.021
Rathcke B, Lacey EP (1985) Phenological patterns of terrestrial plants. Annu Rev Ecol Syst
16:179–214. https://doi.org/10.1146/annurev.es.16.110185.001143
Sakai S (2001) Phenological diversity in tropical forests. Popul Ecol 43:77–86.
https://doi.org/10.1007/PL00012018
Schöngart J, Piedade MTF, Ludwigshausen S, et al (2002) Phenology and stem-growth
periodicity of tree species in Amazonian floodplain forests. J Trop Ecol 18:581–597.
https://doi.org/10.1017/S0266467402002389
Silva JO da., Fernandes RS, Ticli FK, et al (2007) Triterpenoid saponins, new metalloprotease
snake venom inhibitors isolated from Pentaclethra macroloba. Toxicon 50:283–291.
https://doi.org/10.1016/j.toxicon.2007.03.024
Spigler RB (2017) Plasticity of floral longevity and floral display in the self-compatible biennial
Sabatia angularis (Gentianaceae): untangling the role of multiple components of
pollination. Ann Bot 119:167–176. https://doi.org/10.1093/aob/mcw195
Stephenson AG (1981) Flower and fruit abortion: proximate causes and ultimate functions.
Annu Rev Ecol Syst 12:253–279. https://doi.org/10.1146/annurev.es.12.110181.001345
Stevenson PR, Castellanos MC, Cortés AI, Link A (2008) Flowering patterns in a seasonal
tropical lowland forest in Western Amazonia. Biotropica 40:559–567.
https://doi.org/10.1111/j.1744-7429.2008.00417.x
ter Steege H, Pitman NCA, Sabatier D, et al (2013) Hyperdominance in the Amazonian tree
flora. Science (80- ) 342:. https://doi.org/10.1126/science.1243092
van Rheede van Oudtshoorn K, van Rooyen MW (1999) Autochory. In: van Rheede van
Oudtshoorn, Karen van Rooyen MW (ed) Dispersal biology of desert plants: adaptations

68
of desert organisms, 1st edn. Springer, Berlin, Heidelberg, pp 81–91
van Schaik CP, Terborgh JW, Wright SJ (1993) The phenology of tropical forests: adaptive
significance and consequences for primary consumers. Annu Rev Ecol Syst 24:353–377.
https://doi.org/10.1146/annurev.es.24.110193.002033
Vieira MF, Fonseca RS, Araújo LM (2012) Floração, polinização e sistemas reprodutivos em
florestas tropicais. In: Martins SV (ed) Ecologia de florestas tropicais do Brasil, 2nd edn.
Editora UFV, Viçosa, pp 53–84
Vilchez B, Chazdon R, Alvarado W (2007) Fenología reproductiva de las especies del dosel
superior en seis sitios de la Región Huetar Norte de Costa Rica. Kurú Rev For 4:1–16
Vilhena JE de S, Lima e Silva RB, Freitas J da L (2018) Climatologia do Amapá: quase um
século de história. Gramma, Rio de Janeiro
Williamson GB, Costa F (2000) Dispersal of Amazonian tress: hydrocory in Pentaclethra
macroloba. Biotropica 32(3):548–552. https://doi.org/doi.org/10.1646/0006-
3606(2000)032[0548:DOATHI]2.0.CO;2
Wittmann F, Schöngart J, Junk WJ (2010) Phytogeography, species diversity, community
structure and dynamics of Central Amazonian floodplain forests. In: Junk WJ, Piedade
MTF, Wittmann FK, et al. (eds) Amazonian floodplain forests: ecophysiology,
biodiversity and sustainable management, Ecological Studies. Springer Dordrecht
Heidelberg London, London, pp 61–102
Worbes M (1997) The forest ecosystem of the floodplains. In: Junk WJ (ed) The central
Amazon floodplain: ecology of a pulsing system, 1st edn. Springer-Verlag Berlin
Heidelberg, pp 223–265

69
Capítulo 3
________________________________________ Dantas, A.R., Guedes, M.C., Lira-Guedes, A.C & Piedade, M.T.F. Produção e predação de frutos e sementes de Pentaclethra macroloba (Fabaceae) em floresta de várzea do estuário amazônico. Manuscrito em preparação para Biotropica.

70
Produção e predação de frutos e sementes de Pentaclethra macroloba (Fabaceae) em floresta
de várzea do estuário do Rio Amazonas
1*Adelson Rocha Dantas, 2Marcelino Carneiro Guedes, 2Ana Cláudia Lira-Guedes, 1Maria
Teresa Fernandez Piedade
1Instituto Nacional de Pesquisas da Amazônia, Programa de Pós-graduação em Ecologia,
69067-375 Manaus – AM, Brasil; 2Empresa Brasileira de Pesquisa Agropecuária, 68903-419
Macapá – AP, Brasil
*Correspondence author:
Email: [email protected]; Phone: +(55) 9699128-8033
Keywords: pracaxi, dormência cambial, inundação polimodal, óleo de pracaxi, árvore
hiperdominante, hidrocoria.

71
Resumo
Pouco se conhece sobre os fatores que afetam a taxa de produção das árvores ao longo
das várzeas do Rio Amazonas. Pentaclethra macroloba (pracaxizeiro) é uma árvore
hiperdominante na Amazônia, socioeconomicamente importante na região e altamente
explorada, devido ao óleo de suas sementes ser um poderoso fitoterápico. Nós quantificamos a
produção de frutos e sementes e avaliamos a taxa de predação de 33 pracaxizeiros no nordeste
da Amazônia, durante o período de fevereiro a dezembro de 2019. Foram utilizados modelos
lineares para relacionar a produção de sementes com o diâmetro das árvores, densidade de
pracaxizeiros, crescimento cambial, precipitação e altura de inundação. Foi aplicado teste de
Kruskal-Wallis para avaliar as variações morfométricas das sementes produzidas por três
populações de pracaxizeiros ao longo do estuário. No total, foram produzidos 261 frutos (9 ± 8
frutos) e 380 sementes (54 ± 50 sementes). A taxa de predação foi de 5% (18 sementes), com a
maioria das sementes predadas por larvas de mariposa 50%. No período de produção de
sementes, o crescimento diamétrico de P. macroloba desacelera (r = -0,76; F = 12,7; p =
0,006**) e o número de sementes aumenta no período de inundação da floresta (r = 0,90; F =
40,3; p = 0,0001***). Para as três populações, as medidas de comprimento (H = 49,2; p =
0,0001***), largura (H = 62,5; p = 0,0001***), espessura (H = 137,2; p = 0,0001***) e peso
fresco (H = 70,4; p = 0,0001***) das sementes são altamente variáveis ao longo do estuário. A
alta interação da produção de P. macroloba com a dinâmica de inundação do estuário
amazônico e a baixa taxa de predação, podem resultar em maiores taxas de produção e múltiplas
estratégias de colonização, explicando a sua dominância nesse ambiente.

72
1. INTRODUÇÃO
Os produtos florestais não madeireiros (PFNMs) exercem grande função ecológica e
econômica. Todas as partes do vegetal e compostos secundários que a árvore produz são
considerados PFNMs, excluíndo a madeira (Ticktin 2004). As múltiplas alternativas dos
PFNMs são fundamentais na valorização da floresta, pois o extrativismo desses produtos
permite uma renda extra para a comunidade local e a conservação da floresta (Ticktin 2015),
preservando também as árvores.
As florestas de várzea da Amazônia são uma importante fonte de PFNMs. Wittmann et
al. (2010) investigaram o uso de 186 espécies arbóreas de áreas alagáveis da Amazônia Central,
e constataram que do total de espécies amostradas, 135 espécies (72,6%) são utilizadas pela
população ribeirinha, sendo que 49 dessas espécies (36,3%) são usadas para uma única
finalidade e a grande maioria é utilizada para múltiplos propósitos. Dentre as espécies
estudadas, 98 espécies produzem produtos não madeireiros, uma grande parte dos quais é
utilizada na medicina popular. No estuário amazônico, os PFNMs são fundamentais para a
subsistência das populações ribeirinhas, sendo neste ambiente encontradas várias espécies de
múltiplos usos e produtoras de PFNMs, como: Euterpe oleracea Mart. (a polpa do fruto é muito
consumida nos estados do Amapá e do Pará), Carapa guianensis Aubl. (a semente possui óleo
medicinal), Virola surinamensis (Rol. ex Rottb.) Warb. (manteiga extraída do fruto é muito
utilizada na indústria de cosmético), entre outras.
Pentaclethra macroloba (Willd.) Kuntze é uma espécie da família Fabaceae, conhecida
popularmente na Amazônia pelo nome “pracaxi” (Dantas et al. 2017) ou “pracaxizeiro”. A
espécie é hiperdominante na bacia Amazônia (ter Steege et al. 2013), especialmente nas
florestas de várzea do estuário amazônico (Carim et al. 2016). O pracaxizeiro apresenta
múltiplas utilidades. A casca do tronco possui saponinas terpenóides com ação anti-
hemorragica contra o veneno de víboras do gênero Bothrops (Silva et al. 2007), porém, o
principal PFNM desta espécie é a semente oleaginosa, muito explorada pelos ribeirinhos do

73
estuário amazônico. Das semente é extraído um óleo com propriedades medicinais, indicado
para tratar dores musculares, inflamações (Crespi & Guerra 2013) e cicatrização de ferimentos
(Banov et al. 2014). As comunidades tradicionais da Ilha de Cotijuba, estado do Pará, utilizam
o óleo para tratar pessoas com câncer (Crespi & Guerra 2013), mas ainda não existem estudos
científicos comprovando a sua eficácia.
Atualmente, o óleo das sementes da espécie é bastante visado pelas indústrias de
cosméticos, por apresentar alta concentração de ácidos graxos que hidratam e proporcionam
maciez à pele (Costa et al. 2014). No mercado internacional, o litro do óleo pode custar de
US$50 a US$75 e as indústrias de cosméticos o utilizam no preparo de sabonetes, hidratantes,
esfoliantes e xampus (Teixeira et al. 2020).
Para alguma espécies que produzem mais de um PFNM, o efeito combinado de várias
colheitas pode ter impacto danoso nos processos ecofisiológicos dessas árvores de múltiplas
utilidades (Avocèvou-Ayisso et al. 2009). As taxas de sobrevivência, crescimento e de
reprodução do vegetal podem ser seriamente afetadas por extrações desordenadas (Ticktin
2004) e subsequentes.
A falta de conhecimento sobre aspectos básicos da ecologia de P. macroloba é
preocupante, pois as sementes dessa espécie já vêm sendo extensivamente exploradas na região
do estuário amazônico, o que pode seriamente afetar a estrutura demográfica da população.
Determinar a capacidade produtiva de uma árvore é um fator chave para auxiliar em estratégias
de manejo e conservação das espécies com potencial econômico, como P. macroloba. O
monitoramento e quantificação da produção de frutos e sementes é fundamental para determinar
o quanto a espécie produz e quais fatores bióticos e abióticos influenciam na produção. Os
fatores que afetam a taxa de produção de frutos e sementes em espécies arbóreas podem ser
melhor entendidos quando associados com as características intrínsecas das espécies (Pinto et
al. 2019), e com as variáveis ambientais do habitat.

74
A variabilidade no tamanho da semente dentro e fora de uma população arbórea é um
traço funcional que revela as múltiplas estratégias de colonização do vegetal, um aspecto chave
da biologia evolutiva de plantas. Variações no tamanho da semente podem ser atribuídas à
herança genética dos pais e à posição do fruto na copa (Simons & Johnston 2000). As variadas
amplitudes de inundação, a heterogeneidade topográfica e o status nutricional das áreas úmidas
amazônica, são fatores que modelam o tamanho da semente de várias espécies vegetais
adaptadas à esse tipo de ambiente (Parolin, 2000). No estuário amazônico, a inundação diária
do Rio Amazonas é um agente dispersor efetivo de sementes (Cunha et al. 2017) de muitas
plantas, pois o vai e vem diário do pulso de inundação polimodal maximiza a dispersão à longa
distância e aumenta a probabilidade da semente colonizar novas áreas. Nesse sentido, pode-se
esperar alta variabilidade de tamanhos de sementes produzidas pelas as árvores, principalmente
por P. macroloba devido a sua dominância na região do estuário.
A identificação dos principais predadores de propágulos das árvores tropicais é
importante para entender a demografia da espécie, além de auxiliar em planos de conservação
e de manejo dos recursos naturais. Isto se deve ao fato de que os predadores de sementes
exercem um impacto significativo na produção das espécies economicamente importantes,
podendo influenciar na demografia das árvores hiperdominantes (Silman et al. 2003), como P.
macroloba.
Entretanto, é sabido que P. macroloba produz compostos secundários em suas sementes,
que são tóxicos para a maioria dos animais (Janzen 1970, McKenna & McKenna 2006) e este
pode ser um dos fatores que favoreça sua dominância nas florestas da América Central e da
Amazônia. Nesse caso, são esperadas baixas taxas de predação para a espécie nas áreas de
estudo. Diante disso, nós determinamos neste estudo a taxa de predação de sementes e
avaliamos os fatores ambientais da várzea estuarina e intrínsecos de P. macroloba relacionados
à sua produção.

75
2. MÉTODOS
2.1. Área de estudo
O monitoramento da produção de sementes ocorreu no Campo Experimental do
Mazagão (CEM), pertencente à Empresa Brasileira de Pesquisa Agropecuária do Amapá. O
CEM está localizado no sudeste do estado do Amapá (00° 06' S, 51° 16' W), cidade de Mazagão,
Brasil (Dantas, Lira-Guedes, Guedes, Piedade, & Batista, 2020). Além da população de do P.
macroloba do CEM, foram obtidos dados morfométricos de sementes de populações de P.
macroloba em área ao longo do estuário, sendo: na Área de proteção ambiental da Fazendinha
(APA da Fazendinha) (Dantas et al. 2021) e na Ilha de Santana (IS) (dados cedidos por Soares,
Santos, & Silva, 2019) que ficam a 17 km (00º4’33’’S e 51º10’18’’W) e 12 km (00º3’10’’S e
51º7’38’’W), respectivamente, da área deste estudo.
O sistema climático da região é Am tropical chuvoso (Alvares et al. 2014), sendo que
no estado do Amapá o período chuvoso tem início em dezembro com pico de chuvas acima de
60 mm no mês de março e finaliza no mês de agosto. O período de estiagem inicia no mês de
setembro, quando o regime de precipitação fica abaixo de 60 mm mensais (Vilhena et al. 2018).
O solo do estuário amazônico é classificado como Gleissolo Melânico, de textura siltosa
e com predomínio de esmectita, ilita, caulinita, goetita, anatasio e quartzo (Pinto 2014). A
vegetação é classificada como Floresta Ombrófila Densa Aluvial (IBGE, 2012), com
dominância das espécies Mora paraensis (Ducke) Ducke, Astrocaryum murumuru Mart.,
Pentaclethra macroloba (Willd.) Kuntze, Carapa guianensis Aubl. e Virola surinamensis
(Rol.) Warb. (Carim et al. 2016).
2.2. Amostragem
Foram selecionadas 33 árvores com diâmetro na altura do peito DAP > 5 cm (DAP entre
17 a 50 cm) para o monitoramento da produção de frutos e sementes no CEM. Os critérios de
seleção das árvores foram: bom aspecto fitossanitário, dominante e codominante no estrato
arbóreo e distanciamento entre árvores de 50 m. Dentre essas 33 árvores, sete foram

76
selecionadas para instalação de cercas coletoras de sementes. As sete árvores foram cercadas
com tela de polietileno de 1 m de altura, capturando toda a área de projeção da copa no solo
(Wadt et al. 2017) e adicionando mais 2 m para fora da área, por causa da deiscência explosiva
dos frutos que lançam as sementes para fora da copa (Hartshorn 1983). O cercamento das
árvores é importante em floresta de várzea estuarina, pois a inundação pode levar as sementes
de um dado indivíduo e trazer sementes de outras áreas, prejudicando as estimativas de
produção da árvore monitorada (Wadt et al. 2017).
A produção de frutos e sementes foi monitorada durante o período de fevereiro a
dezembro de 2019 em intervalos de 15 dias. A quantidade de frutos foi estimada pelo método
de visualização e contagem diretamente na copa (Minor & Kobe 2019), com auxílio de binoculo
de 60x. As sementes coletadas dentro da cerca foram acondicionadas em sacos plásticos
etiquetadas e transportadas para o laboratório de sementes da Embrapa Amapá, onde foi
realizada a triagem, separando as sementes sadias das predadas. Das sementes sadias, foram
obtidos dados morfométricos de comprimento (mm), largura (mm), espessura (mm) e peso
fresco (g) (Figura 1) com auxílio de paquímetro digital (Carbografite® modelo 150, precisão
de 0,01 mm) e balança analítica (Shimadzu® modelo AUW22OD, precisão de 0,001 g),
respectivamente.

77
Figura 1: Sementes de Pentaclethra macroloba coletada na floresta de várzea do estuário
amazônico. Comprimento (L), largura (W) e espessura (T).
As variáveis ambientais coletadas foram altura de inundação no tronco das árvores e
precipitação mensal. A altura da inundação foi medida em barbantes brancos instalados
paralelamente ao tronco de cada árvore monitorada. A água do rio, rica em sedimento, mancha
o barbante branco e deixa gravada a altura da inundação, facilitando a medição com fita métrica.
A cada avaliação, os barbantes foram trocados para a medição subsequente. Dados de
precipitação pluviométrica mensal foram obtidos da estação climatológica de Macapá
(00°02’07’’ S e 51°05’55’’ W), armazenados no banco de dados do Instituto Nacional de
Meteorologia (INMET 2019).
As variáveis intrínsecas de P. macroloba coletadas foram: crescimento diamétrico,
densidade populacional e diâmetro do tronco. O crescimento diamétrico das árvores cercadas
foi aferida por meio de bandas dendrométricas instaladas, em julho de 2018, na altura de 1,30
m no tronco da árvore. Após um mês de ajustamento das bandas dendrométricas, o crescimento
diamétrico foi medido por meio de paquímetro digital. O efeito da densidade populacional na
produção de sementes foi testado por meio dos pracaxizeiros presentes em um raio de 100 m

78
do entorno das árvores monitoradas. A medida de diâmetro foi obtida na altura de 1,30 no tronco
de cada árvore com auxílio de fita métrica.
Para avaliar a variabilidade no tamanho de sementes produzidas ao longo do estuário
amazônico, foram obtidos dados morfométricos de 379 sementes de sete árvores no CEM, 100
sementes de 15 árvores na Ilha de Santana e 134 sementes de 30 árvores na APA da Fazendinha.
2.3 Análise dos dados
Foram realizadas estatísticas descritivas das amostras morfométricas para obtenção dos
valores da média, desvio padrão e coeficiente de variação de todas as características ligadas a
produção e à biometria das sementes (Dantas, Ribeiro, Guedes, & Guedes, 2014).
A produção mensal de sementes de P. macroloba (representada pelo número de
sementes coletadas mensalmente nos coletores) foi relacionada com o diâmetro da árvore (cm),
crescimento diamétrico (mm), densidade populacional (número de árvores encontradas em um
raio de 100 m, dividido pela área da circunferência m² ha-1), altura da inundação (cm) e
precipitação pluviométrica mensal acumulada (mm). A magnitude das relações foi analisada
pelo coeficiente de Pearson (r) e por modelos lineares. A significância das relações foi avaliada
pelo teste da Análise da Variância (***significativo a 0,001; **significativo a 0,01;
*significativo a 0,5). Foi aplicado teste de Kruskal-Wallis para avaliar as variações
morfométricas das sementes produzidas pelas três populações de pracaxizeiros ao longo do
estuário amazônico.
A taxa de predação foi determinada pelo quociente entre o número de sementes predadas
e o número total de sementes coletadas, com o resultado multiplicado por 100 (Lira-Guedes &
Jesus-Barros 2017). Todas as análises foram realizadas no programa R (R core Team 2019).
3. RESULTADOS
Foram visualizados 261 frutos na copa das 33 árvores monitoradas, média de 9 frutos
por árvore (± 8 frutos) e coeficiente de variação de 89%. Os coletores instalados em sete
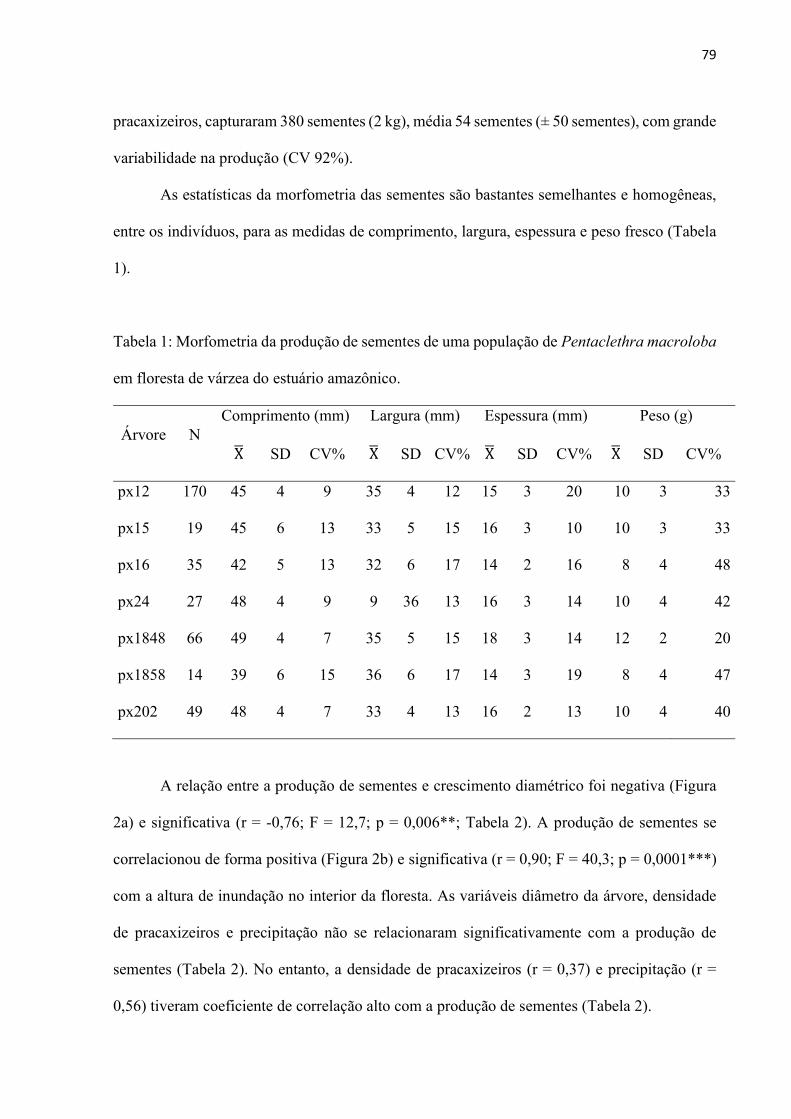
79
pracaxizeiros, capturaram 380 sementes (2 kg), média 54 sementes (± 50 sementes), com grande
variabilidade na produção (CV 92%).
As estatísticas da morfometria das sementes são bastantes semelhantes e homogêneas,
entre os indivíduos, para as medidas de comprimento, largura, espessura e peso fresco (Tabela
1).
Tabela 1: Morfometria da produção de sementes de uma população de Pentaclethra macroloba
em floresta de várzea do estuário amazônico.
Árvore N Comprimento (mm) Largura (mm) Espessura (mm) Peso (g)
X SD CV% X SD CV% X SD CV% X SD CV%
px12 170 45 4 9 35 4 12 15 3 20 10 3 33
px15 19 45 6 13 33 5 15 16 3 10 10 3 33
px16 35 42 5 13 32 6 17 14 2 16 8 4 48
px24 27 48 4 9 9 36 13 16 3 14 10 4 42
px1848 66 49 4 7 35 5 15 18 3 14 12 2 20
px1858 14 39 6 15 36 6 17 14 3 19 8 4 47
px202 49 48 4 7 33 4 13 16 2 13 10 4 40
A relação entre a produção de sementes e crescimento diamétrico foi negativa (Figura
2a) e significativa (r = -0,76; F = 12,7; p = 0,006**; Tabela 2). A produção de sementes se
correlacionou de forma positiva (Figura 2b) e significativa (r = 0,90; F = 40,3; p = 0,0001***)
com a altura de inundação no interior da floresta. As variáveis diâmetro da árvore, densidade
de pracaxizeiros e precipitação não se relacionaram significativamente com a produção de
sementes (Tabela 2). No entanto, a densidade de pracaxizeiros (r = 0,37) e precipitação (r =
0,56) tiveram coeficiente de correlação alto com a produção de sementes (Tabela 2).
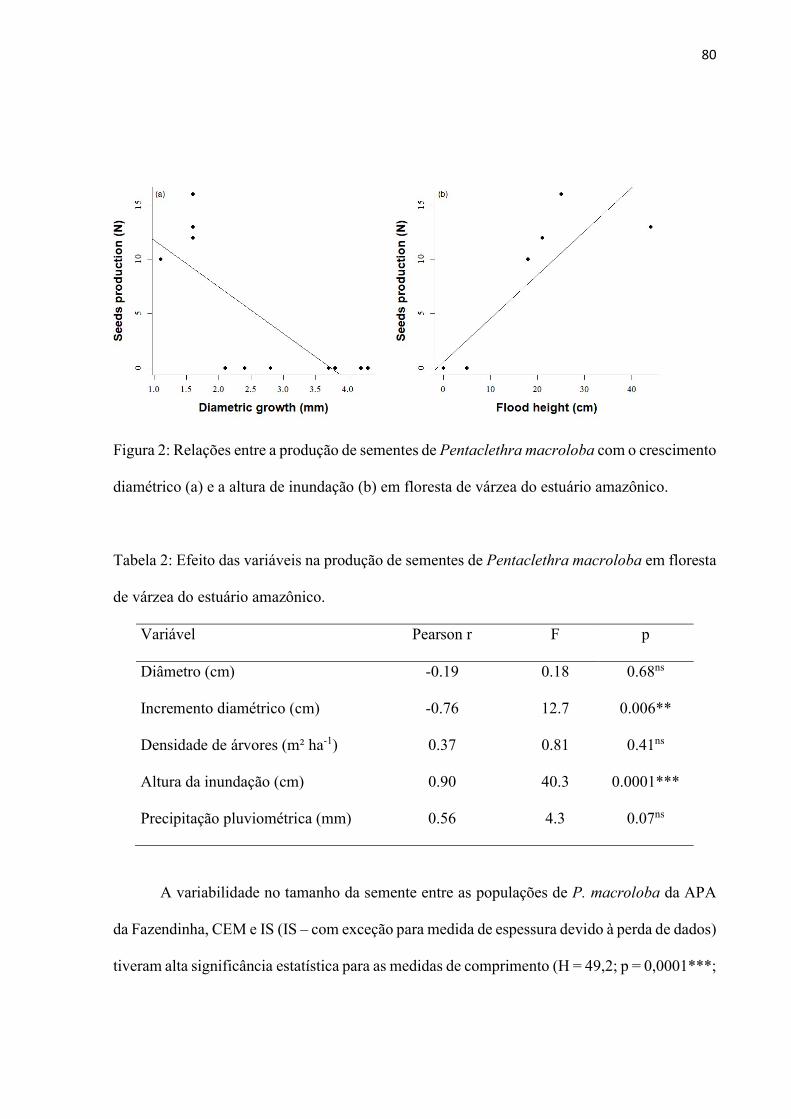
80
Figura 2: Relações entre a produção de sementes de Pentaclethra macroloba com o crescimento
diamétrico (a) e a altura de inundação (b) em floresta de várzea do estuário amazônico.
Tabela 2: Efeito das variáveis na produção de sementes de Pentaclethra macroloba em floresta
de várzea do estuário amazônico.
Variável Pearson r F p
Diâmetro (cm) -0.19 0.18 0.68ns
Incremento diamétrico (cm) -0.76 12.7 0.006**
Densidade de árvores (m² ha-1) 0.37 0.81 0.41ns
Altura da inundação (cm) 0.90 40.3 0.0001***
Precipitação pluviométrica (mm) 0.56 4.3 0.07ns
A variabilidade no tamanho da semente entre as populações de P. macroloba da APA
da Fazendinha, CEM e IS (IS – com exceção para medida de espessura devido à perda de dados)
tiveram alta significância estatística para as medidas de comprimento (H = 49,2; p = 0,0001***;
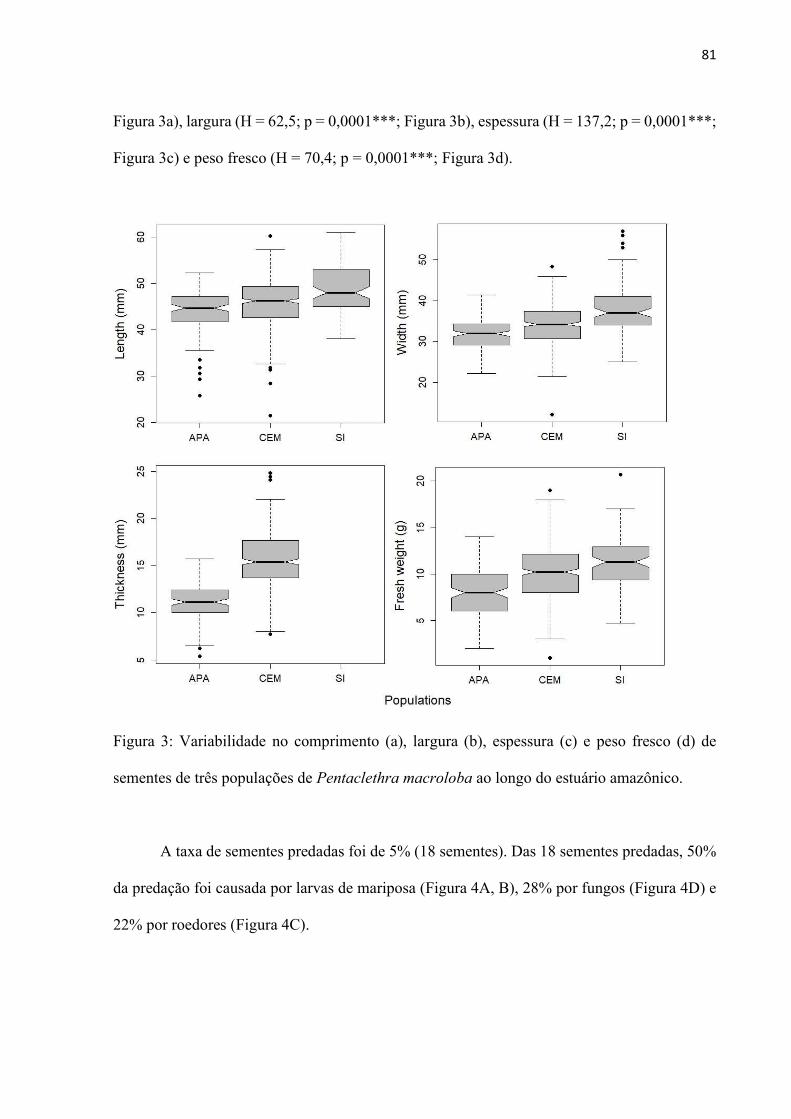
81
Figura 3a), largura (H = 62,5; p = 0,0001***; Figura 3b), espessura (H = 137,2; p = 0,0001***;
Figura 3c) e peso fresco (H = 70,4; p = 0,0001***; Figura 3d).
Figura 3: Variabilidade no comprimento (a), largura (b), espessura (c) e peso fresco (d) de
sementes de três populações de Pentaclethra macroloba ao longo do estuário amazônico.
A taxa de sementes predadas foi de 5% (18 sementes). Das 18 sementes predadas, 50%
da predação foi causada por larvas de mariposa (Figura 4A, B), 28% por fungos (Figura 4D) e
22% por roedores (Figura 4C).

82
Figura 4: Predação por larvas de mariposa (A e B), por roedores (C) e por fungos (D) em
sementes de Pentaclethra macroloba em floresta de várzea do estuário amazônico.
4. DISCUSSÃO
A alta adaptabilidade de P. macroloba ao ambiente diariamente inundado do estuário
amazônico se mostra no ajuste de seus processos reprodutivos ao ciclo de inundação desse
ambiente. O resultado é uma maior interação das suas fases crucias de vida, a produção e a
dispersão de sementes, com o ambiente inundado. Isso se reflete em maiores taxas de produção
e variabilidade de formas e tamanhos de propágulos produzidas por P. macroloba.
A população de P. macroloba do estuário amazônico apresenta maior produção de
frutos, em comparação com a população da espécie da América Central. Minor & Kobe (2019)
estimaram a produção de frutos de P. macroloba na Estação Biológica La Selva, Costa Rica,
pelo mesmo método de visualização e contagem dos frutos na copa, e constataram que 367
árvores monitoradas, com diâmetro entre 18 cm a 119 cm, produziram juntas 188 frutos. Já em
nosso estudo, as 33 árvores monitoradas, com diâmetro entre 17 cm a 55 cm, produziram 261
frutos. A variabilidade na produção de frutos entre as duas populações pode estar relacionada
ao grau nutricional do solo. O solo da Estação Biológica La Selva é de origem antiga e de baixa
fertilidade, localizado em regiões vulcânicas da Costa Rica (McDade & Hartshorn 1994). Os
solos das florestas de várzea do estuário amazônico apresentam alta concentração dos nutrientes
Ca, Mg, K e P (Montagnini & Muñiz-Miret 1999), por serem banhados pelo Rio Amazonas que

83
transporta alta carga de material sedimentar, rica em nutrientes, de origem andina e pré-andina
(Meade 2008).
Pentaclethra macroloba apresentou alta variabilidade na quantidade de frutos e
sementes produzidas entre árvores (de 1 a 26 frutos), ou seja, alguns indivíduos produzem mais
que outros. A variabilidade intrapopulacional na produção de frutos e sementes é uma
característica intrínseca das árvores tropicais, aliado à heterogeneidade ambiental da floresta.
Esse comportamento pode evidenciar uma exaustão reprodutiva e alternância na máxima
produção. Segundo Obeso (2002), a atividade reprodutiva para uma planta requer bastante
dispêndio de energia e alocação de recursos nutricionais. Assim, existe um ano de alta
produção, exigindo da árvore grande investimento nutricional e energético, seguido de um ou
dois anos com baixa ou média produção, em que a árvore está se recuperando fisiologicamente.
Segundo Lucas-Borja & Vacchiano (2018) esse padrão de produção que varia de ano para ano
e entre indivíduos é um fator chave do recrutamento natural das populações arbóreas. A
quantidade de sementes produzidas e a variabilidade da produção entre árvores, mostra que P.
macroloba apresenta aptidão para integrar sistemas de manejo de coleta de sementes.
Segundo Minor & Kobe (2019), a produção de frutos e de semente em árvores tropicais
pode ser altamente desigual entre plantas da mesma espécie, devido à fatores ambientais.
Klimas et al. (2012), estudando a produção de sementes de Carapa guianensis em floresta de
várzea e de terra firme por quatro anos no estado do Acre, Brasil, constataram que a população
da várzea produz mais sementes do que a população da terra firme. Os autores atribuem esses
resultados à maior fertilidade dos solos de várzea, o que contribui para o aumento da produção.
Outro fator que afeta a atividade reprodutiva do vegetal é a mudança climática. Wright &
Calderon (2005) verificaram, em uma comunidade arbórea da Ilha de Barro Colorado no
Panamá, que eventos moderados de El Niño, contribuíram para o aumento da taxa de produção
de sementes da comunidade arbórea. Segundo os autores, o aumento moderado da temperatura

84
e da radiação solar provavelmente contribuiu para o aumento da produtividade das árvores, pois
as suas copas estariam mais expostas a luz, aumentando a taxa fotossintética da planta.
O tamanho do diâmetro não foi um bom preditor para explicar a produção de sementes
de P. macroloba. Esse resultado também foi encontrado no estudo de Minor & Kobe (2019) no
qual os autores constataram que grandes indivíduos de P. macroloba produzem relativamente
poucos frutos. As árvores pioneiras, como P. macroloba, têm como características produzir
frutos e sementes logo nos estágios iniciais da juventude, devido ao curto tempo de vida,
independente do diâmetro. Quando se trata de espécies de longa vida e de estágio sucessional
clímax, o diâmetro pode ser um bom preditor para explicar variabilidades na produção de frutos
e sementes. No estuário amazônico, Londres et al. (2017) modelaram a produção de sementes
da espécie multi-uso andirobeira (Carapa guianensis) relacionando os tipos de floresta
(restinga, baixio e terra preta) onde a espécie ocorre, com parâmetro de iluminação da copa,
forma da copa e tamanho do diâmetro do tronco. Os autores constataram que o diâmetro foi o
melhor preditor para explicar a produção de sementes. O diâmetro da castanheira (Bertholletia
excelsa) explicou 50% da produção de frutos em um modelo misto que incluía os parâmetros
forma da copa, infestação por lianas na copa e fertilidade do solo na floresta de terra firme da
Reserva Extrativista Chico Mendes, Acre (Kainer et al. 2007).
A correlação negativa e significativa entre a produção de sementes com o crescimento
diamétrico mostrou uma estagnação da atividade cambial durante o período de frutificação de
P. macroloba. A reprodução é crucial para a árvore gerar descendentes e perpetuar os seus
genes e isso justifica alocar recursos fotossintéticos para formação de estruturas reprodutivas
(Bazzaz et al. 1987) e deixar outros processos fisiológicos, como a atividade cambial, em
segundo plano. Outro fator que contribui para a diminuição do ritmo de crescimento diamétrico,
é a dispersão de sementes de P. macroloba coincidir com o período de inundação da floresta.

85
Segundo Schöngart et al. (2002), a inundação das áreas úmidas funciona como estressor hídrico
para a planta (assim como a seca em floresta de terra firme), pois a água limita a oxigenação
para as raízes, dificulta a condutividade hídrica do tronco para copa e diminui a taxa de
crescimento cambial.
O aumento do nível do Rio Amazonas favorece o pracaxizeiro a dispersar maiores
quantidades de sementes. As sementes de P. macroloba são adaptadas à dispersão hidrocórica
(Williamson & Costa 2000) e a maré diária do Rio Amazonas facilita o transporte das sementes
por longas distâncias, permitindo a colonização de novas áreas (Moegenburg 2002). As árvores
do estuário Amazônico sincronizam a fase de frutificação e de dispersão com o período de
maior cheia do rio (Cattanio et al., 2004; Dantas et al., 2016), pois além da água, muitas
dependem da ictiofauna para dispersar suas sementes (Kubitzki & Ziburski 1994).
Foi encontrada uma correlação alta entre a produção de sementes e a densidade
populacional (r = 0,37), evidenciando que indivíduos isolados produzem menos do que
indivíduos mais agregados. Vizinhos coespecíficos podem ter efeitos positivos indiretos na taxa
individual de reprodução (Jones & Comita 2010). O padrão agregado de distribuição especial
de P. macroloba (Dantas et al., 2017) deve facilitar a polinização cruzada, pois curtos espaços
entre árvores possibilitam ao polinizador percorrer várias árvores em pequeno espaço de tempo.
Já árvores isoladas não teriam essa vantagem e a probabilidade de autopolinização seria maior
para as espécies monoicas, como P. macroloba (Barros et al. 2017). Segundo Hirayama et al.
(2005), a polinização cruzada proporciona altas taxas de formação de frutos e sementes e na
autopolinização ocorrem altas taxas de mortalidade de óvulos, devido a autoincompatibilidade
polínica.
A variabilidade no tamanho da semente de P. macroloba ao longo do estuário, mostra
as suas múltiplas estratégias de colonização. Segundo Williamson & Costa (2000), sementes
maiores têm maior probabilidade de flutuar na água, por terem mais bolsões de ar entre os

86
cotilédones, o que favorece a flutuação na correnteza do rio e a colonização de áreas distantes.
Já sementes menores submergem logo quando são expelidas da vargem e permanecem perto da
árvore parental. Para a árvore, a produção de pequenas sementes se torna uma vantagem, pois
necessita de pouca energia para formação da mesma (Parolin 2000). Por outro lado, as taxas de
desenvolvimento do embrião e de sobrevivência da plântula aumentam quando as sementes são
maiores, por apresentarem maior massa endospermática e reservas nutritivas (Michaels et al.
1988, Moegenburg 1996).
A baixa taxa de predação de sementes (5%) de P. macroloba sugere a existência de
barreiras defensivas contra a herbivoria. Possivelmente, isso se deve ao fato de que as sementes
da espécie apresentam substancias tóxicas, como alcaloides e aminoácidos livres (Hartshorn
1972), o que as torna impalatáveis para a maioria dos herbívoros. Para a espécie Carapa
guianensis, que também ocorre no estuário amazônico, mas que não dispõe de defensivos
químicos, a taxa de predação por larvas de mariposa do gênero Hypsipyla pode chegar a 62%
(Pinto et al. 2013).
Na área de deste estudo, as sementes de P. macroloba foram frequentemente predadas
por larvas de mariposa. Na Estação Biológica La Selva na Costa Rica, McKenna & McKenna
(2006) constataram que o principal predador das sementes de P. macroloba é a larva da
mariposa Carmenta surinamensis (Möschler) (Lepidoptera: Sesiidae), que tem efeito redutor
na taxa de germinação e no crescimento da plântula da espécie. As estimativas de perdas de
sementes provenientes da predação natural e o tempo de coleta das mesmas devem ser levados
em consideração para o extrativismo do óleo de pracaxi, pois a alta umidade da floresta de
várzea favorece o rápido acometimento por fungos e ataques por larvas de insetos considerando
intervalos de coletas de quinze em quinze dias.
Somente alguns vertebrados se atrevem à digerir sementes de P. macroloba, pois devido
a sua toxidade a maioria dos animais as evita (Guariguata et al. 2000). Em La Selva, papagaios

87
(Pionus senilis (Spix, 1824)) e esquilos (Sciurus variegatoides Ogilby, 1839) são predadores
conhecidos que consomem pequenas quantidades de sementes de P. macroloba. Neste estudo,
as sementes roídas podem ter sido atacadas por cotias (Dasyprocta azarae Lichtenstein, 1823),
que frequentemente aparecem em busca de sementes de andiroba. Esses roedores podem, até
certo ponto, tolerar os compostos alcaloides tóxicos que estão presentes nas sementes de muitas
espécies vegetais (Guimarães et al. 2003).
5. CONCLUSÕES
A baixa taxa de predação de sementes de P. macroloba e a variabilidade no tamanho
das sementes produzida por essa espécie, são fatores que contribuem para formação de grandes
adensamentos populacionais no estuário do Rio Amazonas e para sua hiperdominância na
Amazônia brasileira. Nossos resultados mostram um aumento do número de sementes
produzidas quando a inundação no interior da floresta aumenta, mostrando que o pulso de
inundação diário do estuário amazônico exerce papel chave nos ciclos reprodutivos e nos
processos ecofisiológicos desta espécie. No entanto, monitoramentos de duração mais longa
são fundamentais para entender a variabilidade que ocorre dentro da população e entre anos na
produção de frutos e sementes, e para determinar taxas de extração sustentável de sementes.
AGRADECIMENTOS
Ao Programa de Pós-graduação em Ecologia (INPA/PPGEco), Conselho Nacional de
Desenvolvimento Científico e Tecnológico (bolsa de doutorado, número 142316/2016-4),
Projeto Kamukaia 3 - Valorização dos produtos florestais não madeireiros na Amazônia/
Embrapa Amapá (número 02.13.07.007.00.00), Grupo Ecologia, Monitoramento e Uso
Sustentável de Áreas Úmidas (MAUA/PELD/CNPq/FAPEAM), e aos técnicos da Embrapa
Amapá: Paulo, Adijalma, Enoque e Jonas.

88
REFERÊNCIAS
ALVARES, C. A., J. L. STAPE, P. C. SENTELHAS, J. L. DE M. GONÇALVES, and G. SPAROVEK.
2014. Köppen’s climate classification map for Brazil. Meteorol. Zeitschrift 22: 711–728.
AVOCÈVOU-AYISSO, C., B. SINSIN, A. ADÉGBIDI, G. DOSSOU, and P. VAN DAMME. 2009.
Sustainable use of non-timber forest products: Impact of fruit harvesting on Pentadesma
butyracea regeneration and financial analysis of its products trade in Benin. For. Ecol.
Manage. 257: 1930–1938.
BANOV, D., F. BANOV, and A. S. BASSANI. 2014. Case Series: The effectiveness of fatty acids
from pracaxi oil in a topical silicone base for scar and wound therapy. Dermatol. Ther.
(Heidelb). 4: 259–269.
BARROS, T. C., G. D. PEDERSOLI, J. V PAULINO, and S. P. TEIXEIRA. 2017. In the interface of
caesalpinioids and mimosoids: comparative floral development elucidates shared
characters in Dimorphandra mollis and Pentaclethra macroloba (Leguminosae). Am. J.
Bot. 104: 218–232.
BAZZAZ, F. A., N. R. CHIARIELLO, P. D. COLEY, and L. F. PITELKA. 1987. Allocating resources
to reproduction and defense. Bioscience 37: 58–67.
CARIM, M. DE J. V., F. K. WITTMANN, M. T. F. PIEDADE, J. R. DA SILVA GUIMARÃES, and L. DE
CÁSSIA LEÔNCIO TOSTES. 2016. Composition, diversity, and structure of tidal “Várzea”
and “Igapó” floodplain forests in eastern Amazonia, Brazil. Brazilian J. Bot.
CATTANIO, J. H., A. B. ANDERSON, J. S. ROMBOLD, and D. C. NEPSTAD. 2004. Phenology,
litterfall, growth, and root biomass in a tidal floodplain forest in the Amazon estuary. Rev.
Bras. Botânica 27: 703–712.
COSTA, M. N. F. DOS S., M. A. P. MUNIZ, C. A. B. NEGRÃO, C. E. F. DA COSTA, M. L. N.
LAMARÃO, L. MORAIS, J. O. C. SILVA JÚNIOR, and R. M. RIBEIRO COSTA. 2014.
Characterization of Pentaclethra macroloba oil. J. Therm. Anal. Calorim. 115: 2269–
2275.

89
CRESPI, B., and G. A. D. GUERRA. 2013. Ocorrência, coleta, processamento primário e usos do
pracaxi (Pentaclethra macroloba (Willd.) Kuntze) na Ilha de Cotijuba, Belém- PA. Rev.
Bras. Agroecol. 8: 176–189.
CUNHA, A. C., K. MUSTIN, E. S. DOS SANTOS, É. W. G. DOS SANTOS, M. C. GUEDES, H. F. A.
CUNHA, P. C. C. ROSMAN, and L. DA S. L. STERNBERG. 2017. Hydrodynamics and seed
dispersal in the lower Amazon. Freshw. Biol. 1–9.
DANTAS, A., A. LIRA-GUEDES, M. GUEDES, M. PIEDADE, and A. BATISTA. 2020. Population
dynamics of Attalea excelsa (Arecaceae) in floodplain forest of the amazonian estuary. J.
Trop. For. Sci. 32: 105–113.
DANTAS, A. R., A. C. LIRA-GUEDES, K. MUSTIN, W. C. S. APARÍCIO, and M. C. GUEDES. 2016.
Phenology of the multi-use tree species Carapa guianensis in a floodplain forest of the
Amazon Estuary. Acta Bot. Brasilica 30: 618–627.
DANTAS, A. R., L. C. MARANGON, M. C. GUEDES, A. L. P. FELICIANO, and A. C. LIRA-GUEDES.
2017. Spatial distribution of a population of Pentaclethra macroloba (Willd.) Kuntze in a
floodplain forest of the Amazon estuary. Rev. Árvore 41: 1–11.
DANTAS, A. R., G. G. RIBEIRO, A. C. L. GUEDES, and M. C. GUEDES. 2014. Produção de frutos
de urucurizeiros, Attalea excelsa Mart. (Arecaceae), em floresta de várzea no estuário do
Rio Amazonas. Biota Amaz. 4: 108–114.
GUARIGUATA, M. R., J. J. R. ADAME, and B. FINEGAN. 2000. Seed Removal and Fate in Two
Selectively Logged Lowland Forests with Constrasting Protection Levels\rRemoción y
Destino de Semillas en Dos Bosques de Tierras Bajas Selectivamente Talados con Niveles
Contrastantes de Protección. Conserv. Biol. 14: 1046–1054.
GUIMARÃES, P. R., J. JOSÉ, M. GALETTI, and J. R. TRIGO. 2003. Quinolizidine alkaloids in
Ormosia arborea seeds inhibit predation but not hoarding by agoutis (Dasyprocta
leporina). J. Chem. Ecol. 29: 1065–1072.

90
HARTSHORN, G. 1972. Ecological lifeHistory and population dynamics of Pentaclethra
macroloba and Stryphnodendron excelsum. University of Washington.
HARTSHORN, G. S. 1983. Pentaclethra macroloba (Gavilan). In D. H. Janzen (Ed.) Costa Rican
Natural History. pp. 301–303, Universityof Chicago Press.
HIRAYAMA, K., K. ISHIDA, and N. TOMARU. 2005. Effects of pollen shortage and self-
pollination on seed production of an endangered tree, Magnolia stellata. Ann. Bot. 95:
1009–1015.
IBGE - INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. 2012. Manual técnico da
vegetação brasileira : sistema fitogeográfico : inventário das formações florestais e
campestres : técnicas e manejo de coleções botânicas : procedimentos para mapeamentos
2nd ed. IBGE, Rio de Janeiro.
INMET. 2019. Banco de Dados Meteorológicos para Ensino e Pesquisa. Available at:
nmet.gov.br/portal/index.php?r=bdmep/bdmep [Accessed December 20, 2019].
JANZEN, D. H. 1970. Herbivores and the Number of Tree Species in Tropical Forests. Am. Nat.
104: 501–528.
JONES, F. A., and L. S. COMITA. 2010. Density-dependent pre-dispersal seed predation and fruit
set in a tropical tree. Oikos 119: 1841–1847.
KAINER, K. A., L. H. O. WADT, and C. L. STAUDHAMMER. 2007. Explaining variation in Brazil
nut fruit production. For. Ecol. Manage. 250: 244–255.
KLIMAS, C. A., K. A. KAINER, L. H. WADT, C. L. STAUDHAMMER, V. RIGAMONTE-AZEVEDO, M.
F. CORREIA, and L. M. DA SILVA LIMA. 2012. Control of Carapa guianensis phenology
and seed production at multiple scales: a five-year study exploring the influences of tree
attributes, habitat heterogeneity and climate cues. J. Trop. Ecol. 28: 105–118.
KUBITZKI, K., and A. ZIBURSKI. 1994. Seed Dispersal in Flood Plain Forests of Amazonia.
Biotropica 26: 30.

91
LIRA-GUEDES, A. C., and C. R. JESUS-BARROS. 2017. Avaliação da predação de sementes de
andirobeiras e registro de Hypsipyla spp. In L. H. Wadt, L. M. H. Santos, M. P. de M.
Bentes, and V. B. V. Oliveira (Eds.) Produtos florestais não madeireiros: Guia
Metodológico da Rede Kamukaia. pp. 108–116, Embrapa, Brasilia.
LONDRES, M., M. SCHULZE, C. L. STAUDHAMMER, and K. A. KAINER. 2017. Population
Structure and Fruit Production of Carapa guianensis (Andiroba) in Amazonian Floodplain
Forests: Implications for Community-Based Management. Trop. Conserv. Sci.
LUCAS-BORJA, M. E., and G. VACCHIANO. 2018. Interactions between climate, growth and seed
production in Spanish black pine (Pinus nigra Arn. ssp. salzmannii) forests in Cuenca
Mountains (Spain). New For. 49: 399–414.
MCDADE, L. A., and G. S. HARTSHORN. 1994. La Selva Biological Station. In L. A. McDade,
K. S. Bawa, H. A. Hespenheide, and G. S. Hartshorn (Eds.) La Selva: ecology and natural
history of a Neotropical rain forest. pp. 6–14, The University of Chicago Press, Chicago.
MCKENNA, D. D., and K. M. MCKENNA. 2006. Sesiid moths reduce germination, seedling
growth, and survivorship in Pentaclethra macroloba (Mimosoideae), a locally dominant
lowland neotropical tree. Biotropica 38: 508–513.
MEADE, R. H. 2008. Transcontinental Moving and Storage: The Orinoco and Amazon Rivers
Transfer the Andes to the Atlantic. Large Rivers Geomorphol. Manag. 45–63.
MICHAELS, H. J., B. BENNER, A. P. HARTGERINK, T. D. LEE, S. RICE, M. F. WILLSON, and R. I.
BERTIN. 1988. Seed size variation: magnitude, distribution, and ecological correlates.
Evol. Ecol. 2: 157–166.
MINOR, D. M., and R. K. KOBE. 2019. Fruit production is influenced by tree size and size-
asymmetric crowding in a wet tropical forest. Ecol. Evol. 9: 1458–1472.
MOEGENBURG, S. M. 1996. International association for ecology Sabal palmetto seed size:
causes of variation, choices of predators, and consequences for sedlings. Oecologia 106:

92
539–543.
MOEGENBURG, S. M. 2002. Spatial and temporal variation in hydrochory in Amazonian
floodplain forest. Biotropica 34: 606.
MONTAGNINI, F., and N. MUÑIZ-MIRET. 1999. Vegetation and soils of tidal floodplains of the
Amazon estuary: a comparison of várzea and terra firme forests in Pará, Brazil. J. Trop.
For. Sci. 11: 420–437.
OBESO, J. R. 2002. The costs of reproduction in plants. New Phytol. 155: 321–348.
PAROLIN, P. 2000. Seed mass in Amazonian floodplain forests with contrasting nutrient
supplies. J. Trop. Ecol. 16: 417–428.
PINTO, A. A., B. R. TELES, N. DOS ANJOS, and S. R. M. COUCEIRO. 2013. Predação de sementes
de andiroba [Carapa guianensis Aubl. e Carapa procera DC. (Meliaceae)] por insetos na
Amazônias. Rev. Arvore 37: 1115–1123.
PINTO, E. R. 2014. Solos, hidrologia e estrutura populacional de pracuubeiras em florestas de
várzea do estuário amazônico. Universidade Federal do Amapá.
PINTO, L. C. L., I. P. S. RODRIGUES, and M. A. DRUMOND. 2019. Population structure and fruit
productivity analyses in support of the use of Caryocar brasiliense. Floresta e Ambient.
26: 1–11.
R CORE TEAM. 2019. R: A Language and Environment for Statistical Computing.
SCHÖNGART, J., M. T. F. PIEDADE, S. LUDWIGSHAUSEN, V. HORNA, and M. WORBES. 2002.
Phenology and stem-growth periodicity of tree species in Amazonian floodplain forests.
J. Trop. Ecol. 18: 581–597.
SILMAN, M. R., J. W. TERBORGH, R. A. KILTIE, S. ECOLOGY, and N. FEB. 2003. Population
regulation of a dominant rain forest tree by a major seed predator. Ecology 84: 431–438.
SILVA, J. O. DA., R. S. FERNANDES, F. K. TICLI, C. Z. OLIVEIRA, M. V. MAZZI, J. J. FRANCO, S.
GIULIATTI, P. S. PEREIRA, A. M. SOARES, and S. V SAMPAIO. 2007. Triterpenoid saponins,

93
new metalloprotease snake venom inhibitors isolated from Pentaclethra macroloba.
Toxicon 50: 283–291.
SIMONS, A. M., and M. O. JOHNSTON. 2000. Variation in seed traits of Lobelia inflata
(Campanulaceae): Sources and fitness consequences. Am. J. Bot. 87: 124–132.
SOARES, R. N., R. O. DOS SANTOS, and B. M. DA S. E SILVA. 2019. Morphological aspects and
anatomy of the fruit, seeds and seedlings of Pentaclethra macroloba (willd.) Kuntze
(Fabaceae). J. Seed Sci. 41: 452–460.
TER STEEGE, H. ET AL. 2013. Hyperdominance in the Amazonian tree flora. Science (80-. ). 342.
TEIXEIRA, G. L., L. G. MACIEL, S. MAZZUTTI, C. B. GONÇALVES, S. R. S. FERREIRA, and J. M.
BLOCK. 2020. Composition, thermal behavior and antioxidant activity of pracaxi
(Pentaclethra macroloba) seed oil obtained by supercritical CO2. Biocatal. Agric.
Biotechnol. 24.
TICKTIN, T. 2004. The ecological implications of harvesting non-timber forest products. J. Appl.
Ecol. 41: 11–21.
TICKTIN, T. 2015. The ecological sustainability of non-timber forest product harvest: Principles
and methods. In C. M. Shackleton, A. K. Pandey, and T. Ticktin (Eds.) Ecological
Sustainability for Non-timber Forest Products: Dynamics and Case Studies of Harvesting.
pp. 31–52, Taylor & Francis Group, London.
VILHENA, J. E. DE S., R. B. LIMA E SILVA, and J. DA L. FREITAS. 2018. Climatologia do Amapá:
quase um século de história. Gramma, Rio de Janeiro.
WADT, L. H., H. TONINI, and M. C. GUEDES. 2017. Monitoramento da produção de frutos e
sementes de produtos florestais não madereiros. In L. H. Wadt, L. M. H. Santos, M. P. de
M. Bentes, and V. B. V. Oliveira (Eds.) Produtos florestais não madereiros: guia
metodológico da rede Kamukaia. pp. 41–48, Brasilia.
WILLIAMSON, G. B., and F. COSTA. 2000. Dispersal of Amazonian Tress: Hydrocory in

94
Pentaclethra macroloba. Biotropica 32(3): 548–552.
WITTMANN, F., J. SCHONGART, J. M. BRITO, A. D. O. WITTMANN, M. T. F. PIEDADE, P. PAROLIN,
W. J. JUNK, and J. GUILLALMET. 2010. Manual of trees from Central Amazonian várzea
floodplains. INPA, Manaus.
WRIGHT, S. J., and O. CALDERON. 2005. Seasonal, El Niño and longer term changes in flower
and seed production in a moist tropical forest. Ecol. Lett. 0: 051128082709002.

95
Capítulo 4
________________________________________ Dantas, A.R., Guedes, M.C., Lira-Guedes, A.C., Schöngart, J & Piedade, M.T.F. Demographic structure and growth pattern of Pentaclethra macroloba (Willd.) Kuntze in the floodplain forest of the Amazon River estuary. Under review in Annals of Forest Science.

96
Demographic structure and growth pattern of Pentaclethra macroloba (Willd.) Kuntze in the floodplain forest of the Amazon River estuary Adelson Rocha DANTAS1*, Marcelino Carneiro GUEDES2, Ana Cláudia LIRA-GUEDES2, Jochen SCHÖNGART1, Maria Teresa Fernandez PIEDADE1 * Corresponding author 1 Instituto Nacional de Pesquisas da Amazônia, Programa de Pós-graduação em Ecologia, Manaus, Brasil 2 Empresa Brasileira de Pesquisa Agropecuária, Macapá, Brasil Email address: [email protected], [email protected], [email protected], [email protected], [email protected] Short title: Natural history of a population of Pentaclethra macroloba in the Amazon estuary Keywords: pracaxi oil, growth rings, management, growth strategies, polymodal flood, natural history Contributions of the co-authors Adelson Rocha Dantas: Investigation, Conceptualization, Methodology, Formal analysis, Visualization, Writing - original draft; Marcelino Carneiro Guedes: Writing - review & editing, Conceptualization, Supervision; Ana Cláudia Lira-Guedes: Writing - review & editing, Conceptualization, Project administration, Funding acquisition; Jochen Schöngart: Conceptualization, Methodology, Formal analysis, Writing - review & editing; Maria Teresa Fernandez Piedade: Writing - review & editing, Conceptualization, Supervision. Acknowledgements We thank the Programa de Pós-graduação em Ecologia (PPGEco/INPA), Instituto Nacional de Pesquisas da Amazônia, Conselho Nacional de Desenvolvimento Científico e Tecnológico/ Ph.D. scholarship [grant number 142316/2016-4], Embrapa Amapá, Grupo Ecologia, Monitoramento e Uso Sustentável de Áreas Úmidas (MAUA/PELD/CNPq/FAPEAM), environmental agents: Nerivan and Ermerson and students of the Forest Engineering course: Gabriel, Isabelly, Marcos (UEAP), Cinthia (UNB), Tailon, Gabriel and Pedro (IMMES). Funding This work was supported by Empresa Brasileira de Pesquisa Agropecuári do Amapá/ Kamukaia Project [grant numbers 02.13.07.007.00.00]. Data availability: Mandatory The datasets generated and/or analyzed during the current study are available in the Zenodo repository. Declaration on conflicts of interest: Mandatory

97
The authors declare that they have no conflict of interest. Total number of characters: including spaces, references and captions 55,625 Number of tables: 0 Number of figures: 8

98
Demographic structure and growth pattern of Pentaclethra macroloba (Willd.) Kuntze in
the floodplain forest of the Amazon River estuary
Key message Climatic seasonality and daily tide of the Amazon River are important
environmental drives that influence the growth patterns of the trees in the Amazon estuary.
Abstract
Context Pentaclethra macroloba is a Neotropical species, hyperdominant in the Amazon and
of multiple-uses. Medicinal properties of its seeds oil make this species vulnerable to
overexploitation. Little is known about the natural history of P. macroloba populations in the
Amazon Basin and the effect of the daily tide in the life cycle of trees.
Aim Was to determine the demographic and growth patterns and the environmental factors that
affect the growth of P. macroloba.
Methods In a 136.56 ha forest in the Northeast of the Brazilian Amazon, we quantified all adult
trees (diameter ≥ 5 cm) and we installed two plots of 1 ha each for inventory of the regenerants
(diameter > 5 cm). We model the growth pattern based on the width of the growth rings. We
installed dendrometric bands to relate the diametric growth with temperature, precipitation and
flood level.
Results We recorded 2,072 adult trees and 240 regenerants. P. macroloba presented cambial
dormancy in the rainy season (t = -2.62; p <0.01) and of river flooding (t = -3.01; p <0.01). The
existence of rings in P. macroloba is an important discovery, as only Mora paraensis had its
growth rings recorded in the Amazon estuary.
Conclusion Demographic and growth patterns of P. macroloba respond of the estuarine
floodplain forest environmental heterogeneity and also reflect its life history over time.
Keywords: pracaxi oil, growth rings, management, growth strategies, polymodal flood, natural
history

99
1. Introduction
Flooded areas of the Amazon are constituted by a complex interconnected system of
habitats formed by variations in topography and flood regimes. In the estuarine region of the
Amazon River and its white water tributaries, the floodplain forests occupy approximately
199,281 km² (Junk and Piedade 2010). The Amazon estuary has two daily flood cycles or a
predictable polymodal flood pulse (Junk et al. 2011) since, the oceanic tide influences this
region.
Flood dynamics of floodplain forests of the Amazon form an environmental filter that
selects the most well adapted species to the constant adversities of the environment (Luize et
al. 2018). Pentaclethra macroloba (Willd.) Kuntze is well adapted to the daily flood of the
Amazon estuary, as it has a higher population density among the trees cataloged in the estuarine
environment (Carim et al. 2016), being listed among the 10 hyperdominant species in the
Amazon (ter Steege et al. 2020). Hartshorn (1983) considers the existence of three large
populations of P. macroloba in Latin America. The first population, widely studied, is
concentrated in Costa Rica, mainly in the La Selva Forest Reserve, where the species present
monodominance. The second population is concentrated in the extension from Panama to
Colombia. The third and last population is located in the northeast of the Brazilian Amazon and
for this, the natural history is little known.
P. macroloba, known by the popular name “pracaxi” or “pracaxizeiro”, belongs to the
Fabaceae family and is used by local people for various purposes (Dantas et al. 2017). The main
product is the oil extracted from the seeds, considered a potent herbal medicine for healing
diabetic ulcers (Simmons et al. 2015), wounds, and burns (Banov et al. 2014). Also, the trunk
bark of P. macroloba has terpenoid saponins that have anti-hemorrhagic action against the
venom of the vipers of genus Bothrops (Silva et al. 2007).
The pracaxi oil can be considered an essential product of socio-biodiversity, as the oil
extraction is a source of extra income for riverine populations. People from traditional
communities collect and sell the oil to the cosmetic industries, that uses as raw material, or they
also sell on the local fair for R$100,00 the liter of oil. Riverine residents and the Amazonian
population, in general, use the oil as a natural medicine to relieve muscle pain. There are also
reports of traditional use to treat snake bites, wounds in general and those caused by
leishmaniasis (Sarquis et al. 2019).

100
Cosmetic industries have shown great interest in pracaxi oil due to its high concentration
of fatty acids with emollient action on skin hydration (Costa et al. 2014). The number of
patented pharmaceutical and cosmetic products based on pracaxi oil is currently growing, both
in the national and international market (Oliveira et al. 2020). With this growing demand, the
sustainability of the seed collection becomes worrying, as a massive extraction may change the
dynamics and demographics of the population.
The knowledge of the demographic pattern and the ecological factors that influence the
population structure allows establishing management actions aimed at the sustainability of the
production of the target species of exploitation. In floodplain forests, the amplitude and duration
of flooding are preponderant factors that influence the structure, physiology (Parolin et al.
2010), richness, diversity and spatial distribution pattern of forest communities (Ferreira and
Stohlgren 1999). In Central Amazonia, floods above 10 m in height and lasting up to 270 days
result in drastic changes in the plant metabolism (Parolin et al. 2004) and tree growth (Schöngart
et al. 2002). However, in the Amazon estuary, the flood regime is daily and of short duration.
The knowledge of the adaptive responses of trees to this environment’s conditions is still
incipient.
The plant colonization process can be influenced by the dispersion mechanism and
abiotic characteristics of the habitat. Many species from flooded areas of the Amazon
synchronize the fruiting phase with the flooding period of the rivers (Kubitzki and Ziburski
1994; Schöngart et al. 2002), favoring seeds dispersion by water and fishes (Parolin et al. 2013).
The alternation of aquatic and terrestrial phases has notable effects on the growth rate of the
floodplain trees (Schöngart et al. 2002), which can be observed in the differentiation of the
anatomical wood structures that appear as growth rings (Worbes and Fichtler 2010).
The process of a plant establishing in the habitat and its life history can be analyzed
through dendrochronological studies. Dendrochronology is a crucial tool to understand the past
of a species by analyzing the growth rings that form annually on the tree trunk. Much was
questioned about the existence of annual rings of the trees in the flood environment of the
Amazon, but studies show that several species form rings (Worbes and Junk 1989; Schöngart
et al. 2005, 2017) in response to floods that can last up to 243 days and reach a height of up to
6 m in the trunk of the Central Amazon trees (Schöngart et al. 2002). In the Amazon estuary,
Mora paraensis is the only species known that forms growth rings (Miranda et al. 2018).
However, it is not clear whether the daily flood and of low amplitude has influence on the
formation of the annual tree rings.

101
In this study, we used dendrocrology to analyze the species P. macroloba, looking for
the presence of growth rings, and evaluate the influence of the daily tide of the Amazon River
in the formation of these structures. Additionally, we map the past life trajectory of P.
macroloba to explain the current demographic and growth patterns exhibited by this species
and to assist in public policies aimed at the management and conservation of this important
natural resource.
2. Materials and methods
2. 1 Site description
The study was carried out in the floodplain forests of the Fazendinha Environmental
Protection Area (APA), a reserve of 136.59 ha, and in the Mazagão Experimental Field (CEM),
a 55.95 ha area (Fig.1). Both areas are located in the Amapá State, Brazil. The APA is a
conservation unit for sustainable use located in the Macapá city 00º03’04,24” S e 51°07’42,72”
W. The CEM belongs to the Brazilian Agricultural Research Corporation, located in the
Mazagão city 00º02’33" S e 51º15’24" W.
Fig.1 Two populations of Pentaclethra macroloba located in the Fazendinha Environmental
Protection Area (APA da Fazendinha) and in the Mazagão Experimental Field (CEM), Amazon
estuary region.

102
The estuarine forests in this study are influenced by the tide of the Amazon River, having
as characteristic the occurrence of two daily flood pulses (Junk et al. 2011), where the flood
height can reach up to 4 m in the rainy season at the mouth of the Amazon River (Cunha et al.
2017). These areas are formed by a rich system of channels and streams that regulate the entry
and exit of water from the Amazon River.
In CEM, the various slopes of the soil result in different flooding levels, forming high
and low floodplain environments (Wittmann et al. 2002). In the low floodplain, the flooding of
the river reaches the soil daily, reaching up to 60 cm in the trunk of the trees during the rainy
season and high tides of the Amazon River. In the high floodplain, the soil has a slope from 3
m to 3.5 m high, hindering the entry of the daily tide in this environment. However, the high
floodplain is partially flooded in the rainy season (Dantas et al. 2020).
The climate system in the region is Am type, rainy tropical (Alvares et al. 2014). In the
Amapá State, the rainy season start in December, and the rains are concentrated in March, when
the peak of rainfall remains above 60 mm until August. The dry season starts in September
when the rainfall is below 60 mm monthly (Vilhena et al. 2018).
The soils are Melanic Gleisols, characteristic of the floodplain region of the Amazon
estuary. These soils are silty texture and with predominance of smectite, illite, kaolinite,
goethite, anatase, and quartz (Pinto 2014).
The vegetation is classified as Dense Alluvial Ambrophilous Forest(IBGE 2012). The
five most dominant species, among the 98 registered species, are: Mora paraensis (Ducke)
Ducke (Fabaceae), Astrocaryum murumuru Mart. (Arecaceae), Pentaclethra macroloba
(Willd.) Kuntze (Fabaceae), Carapa guianensis Aubl. (Meliaceae) and Matisia paraensis
Huber (Malvaceae) (Carim et al. 2016).
2.2 Inventory of adult trees
The population structure of adult trees of P. macroloba was evaluated based on an
inventory performed in 100% of the area, in the period from September to November 2017 in
the APA. All trees with diameter at breast height 1,30 m DBH ≥ 5 cm present in the study area
were inventoried. The diameters of the trees were measured, at 1.30 m from the ground, with a
tape measure and identified with numerical tags of zinc. The coordinates of all individuals were
obtained with a GPS device (Garmin 60CSx) with ± 3 m accuracy.

103
The total height of each tree was measured with a laser tape measure (Bosch - GLM 40),
with a precision of 1.5 mm and a range of 40 m. In obtaining height, the observer positioned
himself below the tree's crown, close to the trunk, and visualized the last branch to launch the
laser towards it. The final height of the tree was estimated from the sum of the observer height
(with the arm extended) and the measurement captured on tape measure.
Qualitative data evaluated were: a) reproductive status: presence or absence of fruits or
vestiges of reproductive structures in the crown - the inventory was carried during the flowering
period and the appearance of the first fruits, and b) sociological position, classified as: dominant
(the tree occupies the upper layer of the canopy, where the crown is completely exposed to
light), codominant (the tree occupies the intermediate layer of the canopy, where the crown is
not entirely exposed to light, due to partial shading of neighboring dominant trees) and
suppressed (the tree occupies the understory of the forest, where there is no incidence of direct
light in the crown) (Dawkins 1958).
2.3 Regeneration sampling
The structure of natural regeneration was evaluated in two square plots of 100 m x 100
m (1 ha) each, installed at the same topographic level and separated by 177 m. The inventory
was carried in April 2019 at APA, and all seedlings with height < 1.60 m and all saplings with
height ≥ 1.60 m and diameter < 5 cm were measured (Klimas et al. 2007), tagged, and
georeferenced. Seedlings have their diameters measured at soil height (DSH) and saplings at
breast height (DBH) with the aid of a digital caliper (Carbografite® model 150, accuracy of
0.01 mm) and the height of the individuals was measured with a standard measuring tape.
2.4 Sampling of radial wood
We selected 38 trees without evident phytosanitary problems, with straight trunk and
DBH > 5 cm, of different diametric classes to determine age and growth patterns. Trees with
large buttresses, eccentric trunk and with anomalies in the trunk were avoided. Two wood
samples were obtained per tree, with the aid of increment bore (5.15 mm in diameter and 400
mm in length), at the DBH. The samples were transported to the Embrapa Amapá
Dendrochronology Laboratory, where they were fixed on a wooden support and polished with

104
sandpaper of different grits (100, 200, 400 and 1500 grains/mm2) for better visualization of the
growth rings.
Growth rings were visualized with the aid of a stereomicroscope (Leica EZ4D) with an
attached camera, the images of each sample were obtained at 20x magnification. The images
were obtained in the ".jpg" format with a resolution of 1200 dpi and calibrated in the 1 mm
scale. The growth ring width was measured using the ImageProPus program (version 4.5.0.29).
2.5 Monitoring of diametric growth and hydrometeorological variables
In the CEM, the diametric growth of P. macroloba was evaluated through dendrometric
bands installed in July 2018 on the trunk of 30 trees at the DBH. Trees of different diametric
classes, dominant and codominant in the canopy and at different topographic levels (10 trees in
the high floodplain and 20 in the low floodplain) were selected. After a month of adjusting of
the dendrometer bands, the readings of the diametric growth were measured with a digital
caliper.
The height of the flood was measured in white strings installed parallel to the trunk of
each monitored tree. The water, full of river sediment, stains the white string leaving the flood
height recorded, facilitating the measurement with a tape. The monthly data of accumulated
rainfall and maximum mean temperature were obtained from the Climatological Station of
Macapá (INMET 2019). The monitoring of the diametric growth and the height of the flood
lasted one year and six months, from July 2018 to December 2019, and the data were always
obtained at the end of each month.
2.6 Data analysis
2.6.1 Demographic structure
The parameters of the demographic structure analyzed were: population density
(Equation 1), diametric classes (Equation 2, suggested by Sturges 1926), height classes, basal
area (Equation 3), spatial distribution, age group and growth rates.
PD = (1)
C = 1 + 3.333 LN (N) (2)

105
g = .
; G = ∑ g (3)
PD = population density (ha); N = total number of sample individuals; A = total area sampled;
C = number of diametric classes; LN = natural log; gi = basal area of individual i; 𝜋 =
circumference perimeter (approximate value of 3.141592); dbh = diameter at breast height
measured at 1.30 m from the soil (40000 = conversion value for meters when the dbh is in
centimeters); and G = basal area of the population.
Geographical coordinates of each individual were used to describe the pattern of the
spatial distribution of the population through the univariate Ripley’s K Function (Ripley 1981)
(Equation 4). In the univariate method, the null hypothesis is of Complete Spatial Randomness
(CSR). The 𝐾(𝑠) function must be transformed to the 𝐿(𝑠) function (Equation 5) for better
visualization of the results graphically. The CSR hypothesis was tested by the Monte Carlo test,
with 1000 confidence envelopes of the Poisson pattern surrounding the distance vector s, down
5% and up 5%, of the transformed function 𝐿(𝑠) (Goreaud et al. 1997).
𝐾(𝑠) = ∑ ∑ 𝑊 𝑥 , 𝑥 𝐼(||𝑥 − 𝑥 || < 𝑠), 𝑒 𝑠 > 0 (4)
𝐿(𝑠) = − 𝑠 (5)
𝐾(𝑠) = expected number of events; n = number of trees; (||𝑥 − 𝑥 ||) = Euclidean distance
between locations 𝑥 and 𝑥 ; s = arbitrary distance vector; 𝑊 𝑥 , 𝑥 = edge effect correction
function that shows the proportion of the circumference with center 𝑥 and radius ||𝑥 − 𝑥 ||
that is outside the study region; 𝜆 = number of trees divided by the area of the study region,
being a non-vicious estimator of the process intensity; 𝐿(𝑠) = is the result of the transformed
function 𝐾(𝑠); and 𝐾 = is the Ripley function estimator, with the distance vector s.
To verify the degree of detachment of the population of the expected random
distribution, the aggregation index R (Equation 6) was calculated by the mean distance of the
nearest neighbor tree (Clark and Evans 1954):
𝑅 =
(6)
�� = average distance from the nearest neighbor = ∑
; 𝑟 = distance of the nearest neighbor to
individual i; n = total number of individuals in the population; �� = expected distance to the
nearest neighbor = ; and 𝜌 = tree density.

106
The random pattern occurs when R = 1, if the R index approaches zero the pattern is
grouped and if the R index has a maximum limit around 2.15 the pattern is regular. The z value
was calculated to determine whether the observed distribution was significantly different from
the expected random pattern (Klimas et al. 2007). The K function was determined using the
Splancs package (Rowlingson and Diggle 2017) and the aggregation index R implemented in
the Spatstat package (Baddeley and Turner 2005), both from the R program (R core Team
2019).
2.6.2 Age and growth patterns
Age was determined by quantifying of the growth ring structures formed by marginal
parenchyma (Figure 2) (Fichtler et al. 2003). For hollow trees, the missing rings were estimated
by the mean number of rings of the other samples (Brienen and Zuidema 2006). The structures
were visualized and quantified using a stereomicroscope with an attached camera.
Fig.2 Anatomical structure of the growth rings, formed by bands of marginal parenchyma, of
Pentaclethra macroloba in the Amazon estuary.
The current radial increment (based on the width of the growth rings) was adjusted to
the field diameter to construct the cumulative diametric growth curve for each individual

107
(Brienen and Zuidema 2006). The cumulative curve of each individual was used to obtain the
mean diametric growth curve, which was adjusted to the sigmoid regression model (Equation
7) proposed by Schöngart et al. (2007). The relationship between the height and diameter of
each individual was assessed and adapted using the nonlinear regression model (Equation 8,
Schöngart 2008).
DBH = [ ]
(7)
H = DBH × d/DBH + e (8)
a, b, c, d, and e are estimated parameters of the model.
The rates of Current Annual Increment (CAI; Equation 9) and Mean Annual Increment
(MAI; Equation 10) of diameter, height and biomass were derived for the corresponding age of
each individual (Schöngart et al. 2007). To estimate aboveground biomass (Equation 11) we
used the pantropical allometric equation proposed by Chave et al. (2014).
CAI = CG( ) − CG( ) (9)
MAI = CG( )/ (10)
𝐵𝑖𝑜𝑚𝑎𝑠𝑠 = 0.0673 × (𝐻 × 𝐷𝐵𝐻 × 𝜌) . (11)
CG = accumulated growth in different years t throughout the species lifespan; and ρ = wood
density (for P. macroloba the wood density is 0.65 g cm-3 Reyes et al., 1992).
The successional trajectory of P. macroloba in the forest canopy was evaluated using
the patterns of release and suppression that occurred throughout its history of growth over time.
These patterns were detected from the series of widths of the growth rings of each individual,
and interpreted by the expression proposed by (Nowacki and Abrams 1997):
%𝐺𝐶 = × 100 (12)
%𝐺𝐶 = percentage of growth change in the preceding 10 years and subsequent 10 years; M1 =
mean diametric growth of preceding 10 years; and M2 = mean diametric growth of subsequent
10 years.
Release events occur when relative growth is above 100% for five consecutive years
and suppression events occur when relative growth is below -50% for five consecutive years
(Brienen et al. 2010). The growth trajectory in diameter of the species was classified into four
patterns: (1) direct growth: occurs when the tree has no release or suppression events; (2) one
release event: when the tree shows an increase in relative growth for five consecutive years; (3)

108
one suppression event: when the tree shows a decrease in relative growth for five consecutive
years; and (4) multiple release and suppression events: when two or more release and
suppression events occur (Nowacki and Abrams 1997; Schöngart et al. 2015).
The relationship between the diametric growth of P. macroloba and the hydroclimatic
variables (temperature, precipitation and flood level in high and low floodplain environments)
was evaluated using multiple regressions (Schöngart et al. 2002) and the statistical differences
between the two environments were evaluated by the ANOVA test (Dantas et al. 2020).
3. Results
3.1 Population structure of adults
2,072 individuals were quantified, the mean population density was 15 individuals per
ha-1. The population is distributed in 12 classes of diameter, with 4 cm of interclass amplitude
and mean DBH of 23 cm (± 9 cm). The central classes, which have diameters between 18.1 cm
to 32 cm, concentrate greater population density (9 individuals ha-1) than the first (DBH from
5 cm to 18 cm with 4 individuals ha-1) and the last classes (DBH > 32.1 cm have 2 individuals
ha-1), tending to a log-normal distribution pattern (Fig.3a).
Total basal area of the population was 98.13 m², equivalent to 0.72 m² ha-1, with a mean
of 0.048 m² (± 0.035 m²) per tree. The diameter classes from 18.1 cm to 41 cm are the that most
concentrate basal area (85.80 m²), representing 86% of the total basal area.
The majority of the population consists of trees with reproductive capacity. In total,
1,235 trees (59% of the population) had some reproductive characteristics, such as flowers and
fruits or traces of both. Minimum and maximum reproductive DBH were 7 cm and 70 cm,
respectively, with a mean of 26 cm (± 7 cm). Most reproductive trees (79%) are concentrated
between the DBH from 18.1 cm to 36 cm (Fig.3a, red bars).
In the forest canopy, most individuals were classified as codominant (61%), followed
by dominant (25%) and suppressed (14%). The population has a mean height of 12.7 m (± 5.4
m) and the distribution pattern is similar to the diameter (Fig.3b).
The population of P. macroloba presented aggregated pattern in all distance scales of
the vector s(m) (Figure 4a), being evidenced by the aggregation index < 1 (R = 0.63; p < 0.002)
and by the distribution map (Fig.4b). The mean distance between neighboring trees was 8.5 m
(± 6.4 m), with minimum and maximum of 0.26 m and 54.5 m, respectively. Reproductive
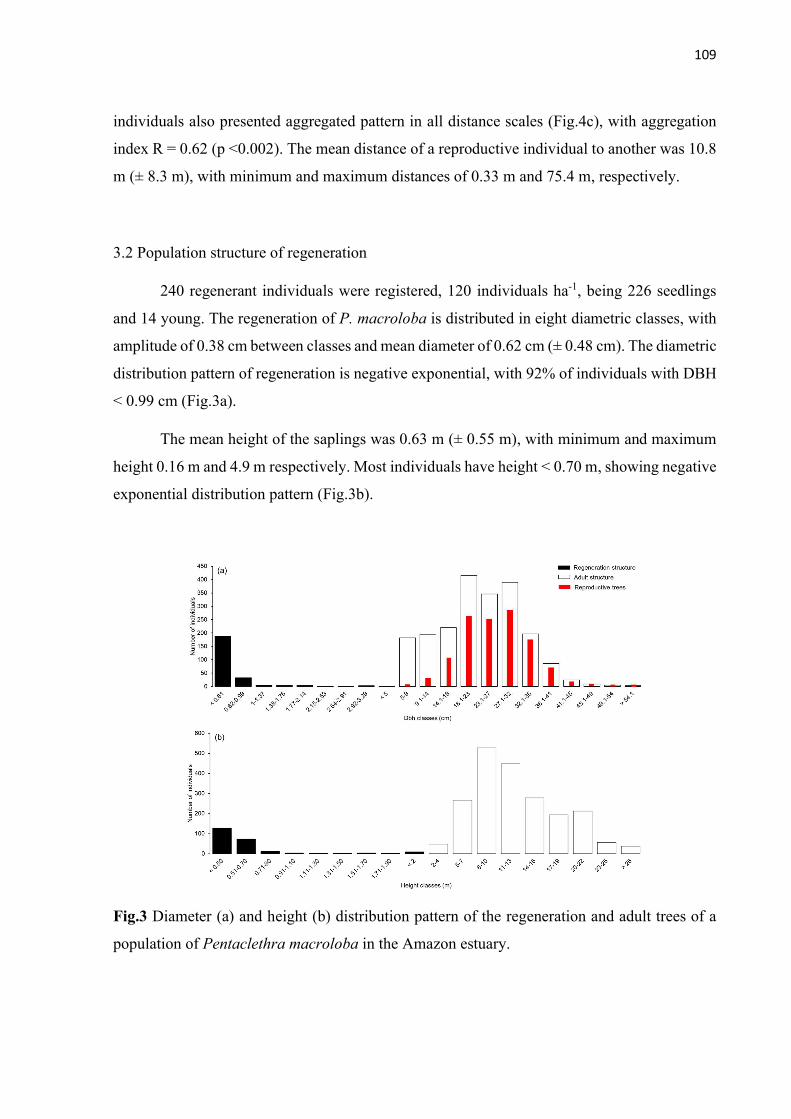
109
individuals also presented aggregated pattern in all distance scales (Fig.4c), with aggregation
index R = 0.62 (p <0.002). The mean distance of a reproductive individual to another was 10.8
m (± 8.3 m), with minimum and maximum distances of 0.33 m and 75.4 m, respectively.
3.2 Population structure of regeneration
240 regenerant individuals were registered, 120 individuals ha-1, being 226 seedlings
and 14 young. The regeneration of P. macroloba is distributed in eight diametric classes, with
amplitude of 0.38 cm between classes and mean diameter of 0.62 cm (± 0.48 cm). The diametric
distribution pattern of regeneration is negative exponential, with 92% of individuals with DBH
< 0.99 cm (Fig.3a).
The mean height of the saplings was 0.63 m (± 0.55 m), with minimum and maximum
height 0.16 m and 4.9 m respectively. Most individuals have height < 0.70 m, showing negative
exponential distribution pattern (Fig.3b).
Fig.3 Diameter (a) and height (b) distribution pattern of the regeneration and adult trees of a
population of Pentaclethra macroloba in the Amazon estuary.
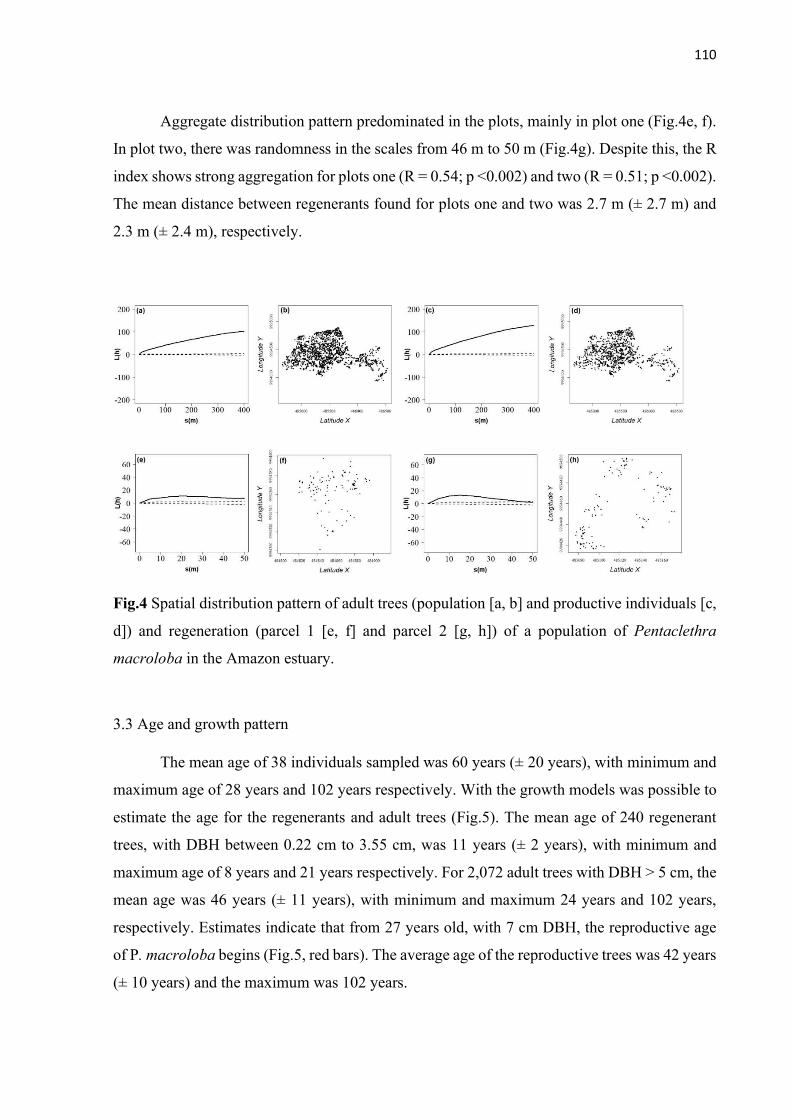
110
Aggregate distribution pattern predominated in the plots, mainly in plot one (Fig.4e, f).
In plot two, there was randomness in the scales from 46 m to 50 m (Fig.4g). Despite this, the R
index shows strong aggregation for plots one (R = 0.54; p <0.002) and two (R = 0.51; p <0.002).
The mean distance between regenerants found for plots one and two was 2.7 m (± 2.7 m) and
2.3 m (± 2.4 m), respectively.
Fig.4 Spatial distribution pattern of adult trees (population [a, b] and productive individuals [c,
d]) and regeneration (parcel 1 [e, f] and parcel 2 [g, h]) of a population of Pentaclethra
macroloba in the Amazon estuary.
3.3 Age and growth pattern
The mean age of 38 individuals sampled was 60 years (± 20 years), with minimum and
maximum age of 28 years and 102 years respectively. With the growth models was possible to
estimate the age for the regenerants and adult trees (Fig.5). The mean age of 240 regenerant
trees, with DBH between 0.22 cm to 3.55 cm, was 11 years (± 2 years), with minimum and
maximum age of 8 years and 21 years respectively. For 2,072 adult trees with DBH > 5 cm, the
mean age was 46 years (± 11 years), with minimum and maximum 24 years and 102 years,
respectively. Estimates indicate that from 27 years old, with 7 cm DBH, the reproductive age
of P. macroloba begins (Fig.5, red bars). The average age of the reproductive trees was 42 years
(± 10 years) and the maximum was 102 years.
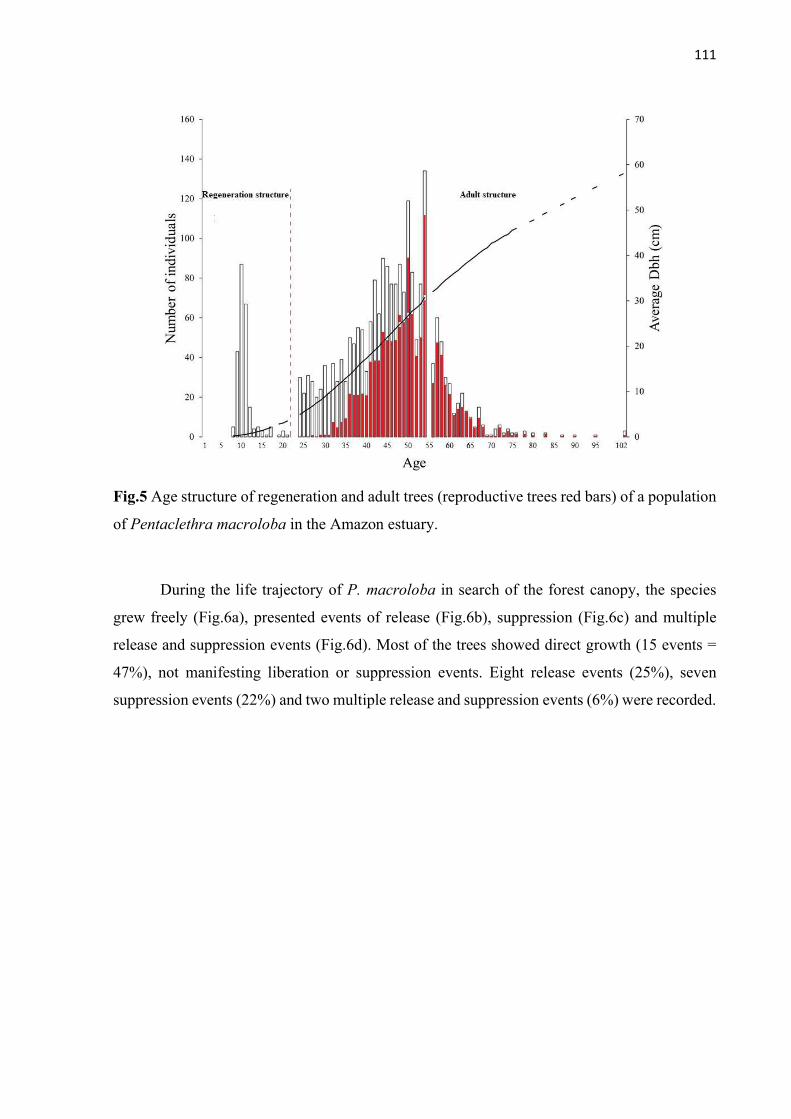
111
Fig.5 Age structure of regeneration and adult trees (reproductive trees red bars) of a population
of Pentaclethra macroloba in the Amazon estuary.
During the life trajectory of P. macroloba in search of the forest canopy, the species
grew freely (Fig.6a), presented events of release (Fig.6b), suppression (Fig.6c) and multiple
release and suppression events (Fig.6d). Most of the trees showed direct growth (15 events =
47%), not manifesting liberation or suppression events. Eight release events (25%), seven
suppression events (22%) and two multiple release and suppression events (6%) were recorded.
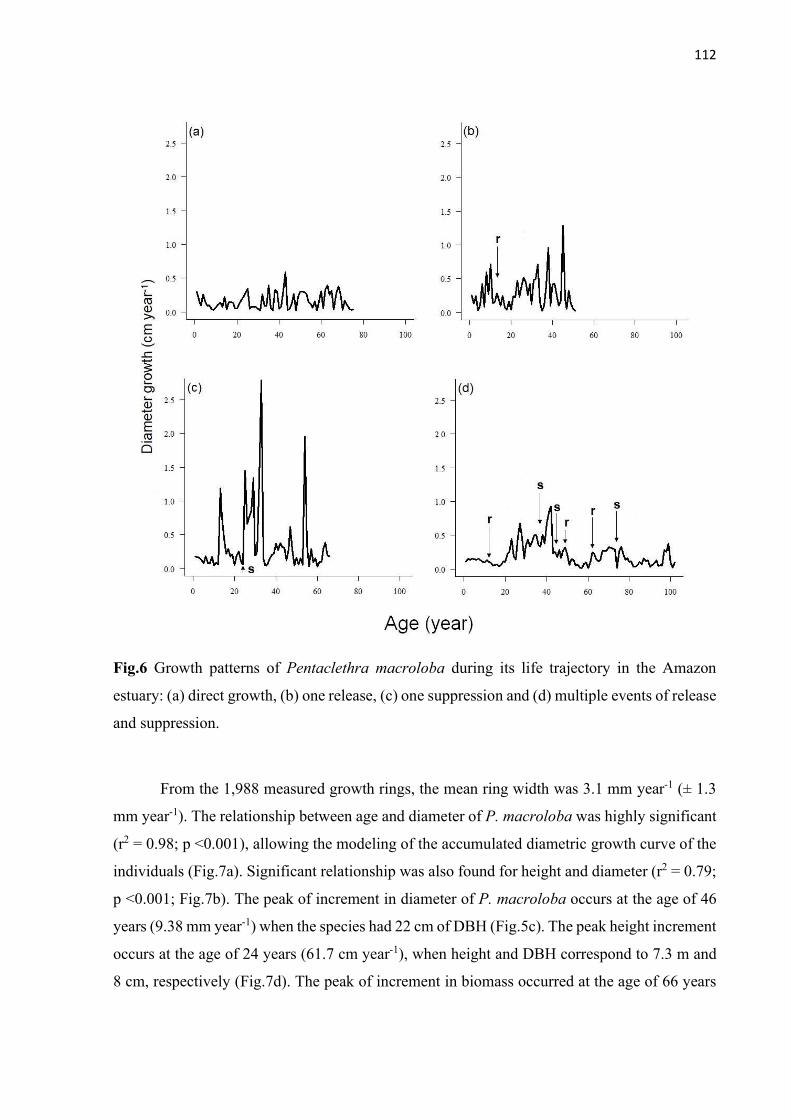
112
Fig.6 Growth patterns of Pentaclethra macroloba during its life trajectory in the Amazon
estuary: (a) direct growth, (b) one release, (c) one suppression and (d) multiple events of release
and suppression.
From the 1,988 measured growth rings, the mean ring width was 3.1 mm year-1 (± 1.3
mm year-1). The relationship between age and diameter of P. macroloba was highly significant
(r2 = 0.98; p <0.001), allowing the modeling of the accumulated diametric growth curve of the
individuals (Fig.7a). Significant relationship was also found for height and diameter (r2 = 0.79;
p <0.001; Fig.7b). The peak of increment in diameter of P. macroloba occurs at the age of 46
years (9.38 mm year-1) when the species had 22 cm of DBH (Fig.5c). The peak height increment
occurs at the age of 24 years (61.7 cm year-1), when height and DBH correspond to 7.3 m and
8 cm, respectively (Fig.7d). The peak of increment in biomass occurred at the age of 66 years
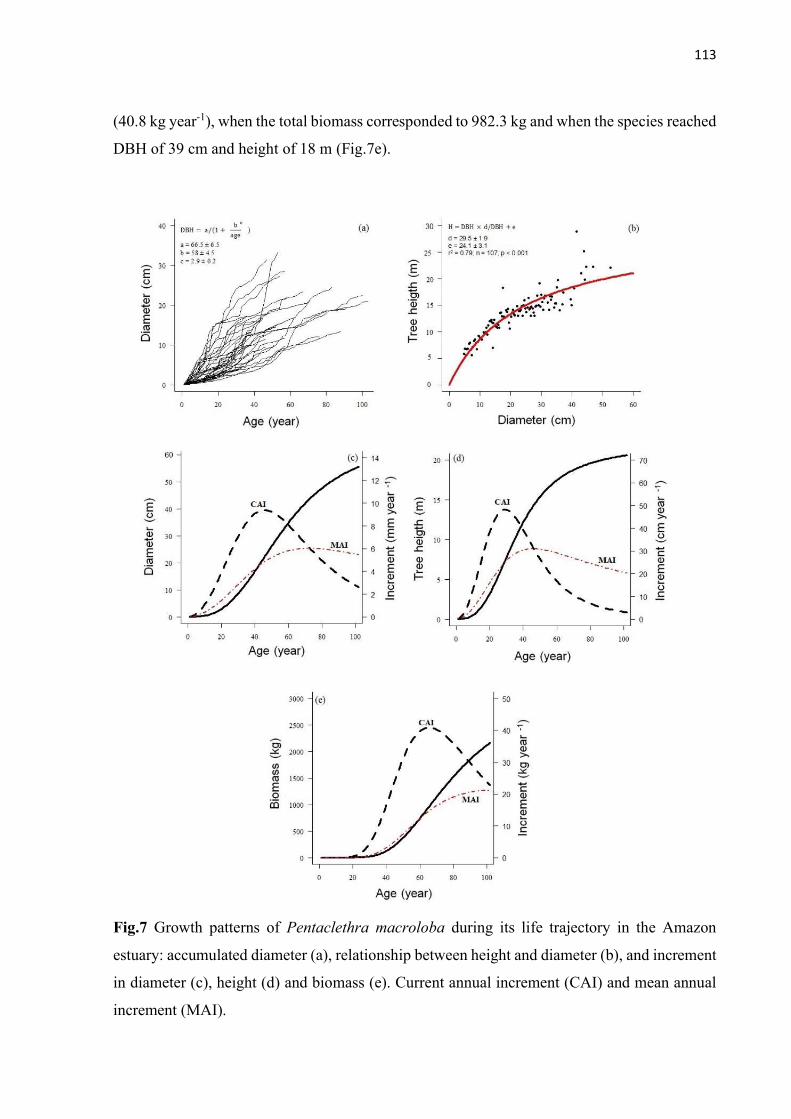
113
(40.8 kg year-1), when the total biomass corresponded to 982.3 kg and when the species reached
DBH of 39 cm and height of 18 m (Fig.7e).
Fig.7 Growth patterns of Pentaclethra macroloba during its life trajectory in the Amazon
estuary: accumulated diameter (a), relationship between height and diameter (b), and increment
in diameter (c), height (d) and biomass (e). Current annual increment (CAI) and mean annual
increment (MAI).

114
3.4 Influence of the hydrometeorological variables in the diametric growth
The mean increment of the high floodplain trees was greater (0.25 mm ± 0.56 mm) than
the pracaxi trees found in the low floodplain (0.24 mm ± 0.61 mm), but without statistical
significance (F = 0.001; p = 0.97). The mean level of flooding between high and low floodplain
differs statistically (F = 6; p <0.01), with low floodplain trees experiencing greater flooding
amplitudes (7.81 cm ± 13.08 cm), compared with the trees of the high floodplain (0.25 cm ±
0.54 cm).
The trees in both environments show greater cambium activity during the dry season
(from September to December), where the tides of the Amazon River do not reach the forest
soil (Fig.8a) due to a decrease in rainfall (Fig.8b) and the elevation of the temperature (Fig.8c).
In the rainy season (Fig.8b) due to the highest level of forest flood (this period corresponds
from January to July, with greater intensity from March to April), the diametric increment of
the species decreases. The peaks of diametric increments were recorded in the high and low
floodplain in September (1.2 mm) and October (0.92 mm) of 2018, respectively. The cambium
activity of the pracaxi trees of low floodplain has significant and negative correlation with
flooding (R2 = 0.47; t = -3.01; p <0.01) and with precipitation (R2 = 0.41; t = -2.62; p <0.01).
With the maximum temperature, the correlation was significant and positive (R2 = 0.44; t =
2.81; p = <0.05). In the high floodplain, the cambium activity of the trees also correlates
negatively with flood and precipitation and positively with the maximum temperature, but
without statistical significance.
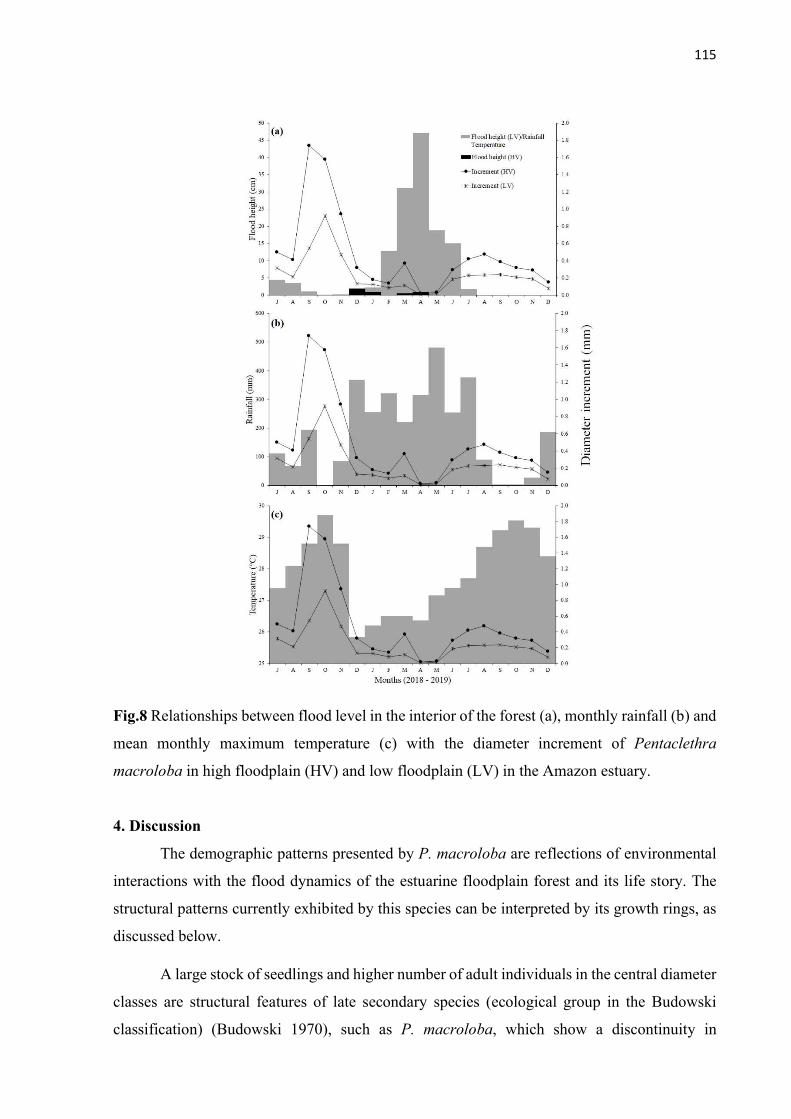
115
Fig.8 Relationships between flood level in the interior of the forest (a), monthly rainfall (b) and
mean monthly maximum temperature (c) with the diameter increment of Pentaclethra
macroloba in high floodplain (HV) and low floodplain (LV) in the Amazon estuary.
4. Discussion
The demographic patterns presented by P. macroloba are reflections of environmental
interactions with the flood dynamics of the estuarine floodplain forest and its life story. The
structural patterns currently exhibited by this species can be interpreted by its growth rings, as
discussed below.
A large stock of seedlings and higher number of adult individuals in the central diameter
classes are structural features of late secondary species (ecological group in the Budowski
classification) (Budowski 1970), such as P. macroloba, which show a discontinuity in

116
recruiting individuals over time (Peters 1996) and depend of disturbances on greater magnitude
to establish in the canopy forest. P. macroloba seedlings are adapted to low light of the
understory and persist for a long time suppressed by the tree crowns waiting for openings in the
canopy (Oberbauer and Strain 1985), what was evidenced in our study by the longevity of the
regenerants, that persist a mean of 11 years in the understory. This is possible because
regenerants can have their growth and survival stimulated even with limited light entering in
the understory, with only 40 minutes in average of direct sun light (Bazzaz and Pickett 1980).
Flood dynamics of the estuarine floodplain forest is another factor that influences the
population structure of P. macroloba. The estuarine floodplain soils are quite fragile, due to
daily tides that deposit and remove sediments from the forest, causing erosion. According to
Metzger et al. (1997), this dynamic leaves the rhizosphere unstable, directly affecting seedling
development and the stability of adult trees. The dry season of the estuary (from September to
December) is also a factor that affects the recruitment of saplings. During this period, the
seedlings experience a prolonged water stress, because the flood does not reach the interior of
the forest due to decreased rainfall. In a seasonally flooded forest in Darien (Panama), Lopez
and Kursar (2007) found that the seedling mortality rate of P. macroloba was higher in the dry
season (43%) than in the rainy season (9.4%). These environmental conditions regulate the
dominance of P. macroloba and maintain the population growth rate stable. Seedlings that can
better adapt to environmental conditions are the few that will pass to the last size classes of the
regeneration structure.
The sigmoid growth pattern over time for P. macroloba (Fig.5) shows different periods
of its life trajectory. According to Köhl et al. (2017), the sigmoid growth curve reveals three
phases: young, maturity and senescence phases. Possibly the log-normal pattern of the diameter
distribution of P. macroloba is reflecting in these three phases (comparison of Fig.3a and 7c).
Classes from 5 cm to 18 cm in diameter belong to young individuals, representing 28.8% of the
population and are on the increasing curve of the diametric increment. The classes from 18.1
cm to 32 cm belong to mature individuals, represent 55.6% of the population and are at the peak
of diametric increment. The classes > 32.1 cm belong to senescent trees, represent 15.6% of the
population, are on the decreasing curve of the diametric increment and are formed by trees with
the major biomass stocks.
Over time, older trees of P. macroloba with larger diameters are the ones that
accumulate more biomass (Fig.7e). Over the time of the plant development, the production rate
of woody material of the tree increases, by the increase of the photosynthetic rate, due to

117
expansion of the leaf and crown area (Sillett et al. 2010). This increase shows the ecological
relevance of old trees as carbon reservoirs (Köhl et al. 2017).
In the young phase, individuals experience more direct growth and release events,
therefore, they have a higher growth rate. However, when individuals reach maturity, the peak
of growth reaches the maximum limit and the individuals begin to settle in the canopy and the
growth slows down. Few individuals that reach maturity pass to the senescent phase of life, as
evidenced by the age structure of populations in this study. (Fig.5). The pattern of diametric
distribution found in this study corroborates to other studies in different parts of the Amazon
estuary, such as the Mazagão Experimental Field, in the South of Amapá, where Dantas et al.
(2017) also found a log-normal form of diametric distribution studying a population composed
of 1,015 trees. This pattern seems to be repeated in other areas of distribution of the species
because in populations of Costa Rica, Galván et al. (2003) found a similar pattern of diametric
distribution for the adult trees of P. macroloba, with higher population density in the central
diameter classes.
Many of the aggregate patterns found in tropical trees may be related to the dispersion
process of the species (Condit et al. 2000) and to the environmental factors of the habitat
(Dantas et al. 2017). When individuals of P. macroloba disperse their seeds by the explosive
dehiscence of the fruits (primary autochoric dispersion), launch their propagules at a distance
of approximately 10 m (Hartshorn 1983), allowing them to be transported by the
hydrodynamics of the river (secondary dispersion by hydrocory). The outflow and inflow of
the daily estuarine tide facilitates the exit and entry of forest seeds (Cunha et al. 2017). Groups
of seeds can be transported over long distances or remain close to the mother tree, as many are
intercepted by branches and trunks present on the forest floor (Ziburski 1991). The result is that
few individuals germinate alone, culminating in a more aggregated pattern. The trees of the
Amazon estuary synchronize the phase of seed dispersal with the period of greatest river
flooding (Cattanio et al. 2004; Dantas et al. 2016), in which it is influenced by the rainfall
regime of the rainy season amazon and tidal cycle (Cunha et al. 2017). This strategy guarantees
the reproductive success of the mother tree and the establishment of new recruitment groups in
the floodplain soil.
In the Amazon estuary, South of Amapá State, Dantas et al. (2017) also found aggregate
pattern for an adult population of P. macroloba. The authors analyzed the spatial distribution
of trees by diametric classes and found that young trees are grouped in the first diameter classes
and randomness in the last classes. This information corroborates with Janzen's hypothesis

118
(Janzen 1970) that young individuals have a better chance of survival if they establish away
from the mother tree, by the shading it provides and by predators that may be housed in the
mother plant itself.
Different growth patterns were observed during the life cycle of P. macroloba in the
conquest of the forest canopy (Fig.4). Most of the trees studied showed direct growth, followed
by release events. This pattern reinforces the idea that P. macroloba depends on clearings to
stimulate natural regeneration (Valverde-Barrantes and Rocha 2014). The seed groups that are
intercepted by the fallen trees branches in the gap, germinate and grow freely towards the
canopy because, in these conditions, no physical barriers are preventing the entry of light.
Competition for light with other species reflects a faster increment in height than in diameter
and the predominance of codominant trees in the canopy (61% of pracaxi trees). Brienen and
Zuidema (2006) emphasize that trees that do not experience events of release or suppression
during succession probably experience very stable lighting conditions, or the light oscillations
are somewhat limiting to individuals, allowing them to grow towards the canopy in less time.
Multiple events of release and suppression during the life history of the tree, evidence
that the forest canopy opened and closed several times due to formation of a clearing or damage
in the tree crown, causing increase and decrease of growth respectively (Brienen and Zuidema
2006).
In Costa Rica, P. macroloba trees that are codominant or dominant in the tree canopy,
which are more exposed to sunlight, show more significant diametric increment than trees
suppressed by the canopy of neighboring trees (Galván et al. 2003). This is evident when P.
macroloba is subjected to silvicultural release techniques followed by refinement (elimination
of neighboring trees and undesirable species). Individuals of the species subjected to such
circumstances show rapid diametric increment (6 mm year-1), compared to individuals who
receive less silvicultural interventions (3 mm year-1) (Finegan et al. 1999).
Although the general pattern of the trees is to first increment in height and then in
diameter, as observed in P. macroloba (comparison of Fig.5b and 5c), in periodically flooded
forests, this strategy is essential for the plant to escape of the flood in its early stages of life
(Parolin 2002). According to Newbery and Ridsdale (2016), understory trees have as strategy
of concentrating more resources at height, as they seek the forest canopy to increase the
photosynthetic rate and decrease competition for light, and only from then on invest in the

119
diameter increment. This type of strategy was also observed by Miranda et al. (2018) for the
estuarine species M. paraensis.
Maximum age of 102 years found in this study for P. macroloba, shows that the species
can be classified as a late secondary of long life in the floodplains of the Amazon estuary. In
Central America, populations of P. macroloba also presented long life. In a study carried out
in the La Selva forest, Costa Rica, Lieberman et al. (1985) conducted successive measurements
on trees over 13 years, one measurement in 1969 and another in 1982, and based on the DBH
measurements, the authors projected the age of P. macroloba to a lifespan of 312 years.
The existence of growth rings for P. macroloba is an important discovery for the
floodplains of the Amazonian estuary. Until the present study only the existence of annual rings
for M. paraensis was confirmed (Miranda et al. 2018). The seasonal pattern of growth of P.
macroloba shows strong indications that the intense rains of the Amazon in combination with
the arrival of the flood in the interior of the forest, may have an influence on the formation of
its growth rings. Torrential rains that fall at the mouth of the Amazon River during the rainy
season, raises the river level (Cunha et al. 2017), resulting in daily flooding inside the forest.
The flood periodicity causes stagnation of cambial activity in P. macroloba (Fig.8), especially
in the low floodplain individuals who are more exposed to flooding. According to Worbes and
Fichtler (2010), the soaked soil leaves the rhizosphere anoxic, impairing the roots breathing and
water absorption from trees. The cambial growth of P. macroloba is resumed in the dry season,
when the flood does not reach the interior of the forest, due to the decrease in the volume of
rainfall in the region.
This behavior is similar to what occurs with trees in the Central Amazon, where they
experience a long-term and high-amplitude flood regime. The trees have a greater diameter
increase in the terrestrial phase (retreat of river level) because, in the aquatic phase, the flood
causes cambial dormancy (Schöngart et al. 2002). In the humid tropical rainforest of La Selva
in Costa Rica, in a forest with flood influence, P. macroloba has the greatest increase in
diameter (1.06 cm year-1) during the dry season, from December to April (Hazlett 1987).
5. Conclusions
The formation of annual growth rings in P. macroloba shows that the estuarine trees are
sensitive to environmental changes. They have the potential for dendroecological studies.

120
The demographic and growth patterns of P. macroloba respond to the environmental
heterogeneity of the estuarine floodplain forest and reflect its life history over time. The
seasonal rainfall of the Amazon estuary is the main factor that influences the growth of the
species, and can be associated with the formation of growth rings of P. macroloba. The flooding
of the forest has a secondary effect on the growth of P. macroloba.
We elucidate some critical gaps of the scientific knowledge about the natural history of
P. macroloba of the Amazon basin, particularly in the estuarine floodplain region. In this way,
we provide subsidies for the formulation of public policies for the good management and
conservation of natural resources of this species of multiple uses.
References
Alvares CA, Stape JL, Sentelhas PC, et al (2014) Köppen’s climate classification map for
Brazil. Meteorol Zeitschrift 22:711–728. https://doi.org/10.1127/0941-2948/2013/0507
Baddeley A, Turner R (2005) spatstat : An R Package for Analyzing Spatial Point Patterns. J
Stat Softw 12:3–4. https://doi.org/10.18637/jss.v012.i06
Banov D, Banov F, Bassani AS (2014) Case Series: The Effectiveness of Fatty Acids from
Pracaxi Oil in a Topical Silicone Base for Scar and Wound Therapy. Dermatol Ther
(Heidelb) 4:259–269. https://doi.org/10.1007/s13555-014-0065-y
Bazzaz FA, Pickett STA (1980) Physiological Ecology of Tropical Succession: A Comparative
Review. Annu Rev Ecol Syst 11:287–310.
https://doi.org/10.1146/annurev.es.11.110180.001443
Brienen RJW, Zuidema PA (2006) Lifetime growth patterns and ages of Bolivian rain forest
trees obtained by tree ring analysis. J Ecol 94:481–493. https://doi.org/10.1111/j.1365-
2745.2005.01080.x
Brienen RJW, Zuidema PA, Martínez-Ramos M (2010) Attaining the canopy in dry and moist
tropical forests: Strong differences in tree growth trajectories reflect variation in growing
conditions. Oecologia 163:485–496. https://doi.org/10.1007/s00442-009-1540-5
Budowski G (1970) The distinction between old secondary and climax species in tropical
Central American lowlands. Trop Ecol 11:44–48
Carim M de JV, Wittmann FK, Piedade MTF, et al (2016) Composition, diversity, and structure
of tidal “Várzea” and “Igapó” floodplain forests in eastern Amazonia, Brazil. Brazilian J

121
Bot. https://doi.org/10.1007/s40415-016-0315-6
Cattanio JH, Anderson AB, Rombold JS, Nepstad DC (2004) Phenology, litterfall, growth, and
root biomass in a tidal floodplain forest in the Amazon estuary. Rev Bras Botânica 27:703–
712. https://doi.org/10.1590/S0100-84042004000400010
Chave J, Réjou-Méchain M, Búrquez A, et al (2014) Improved allometric models to estimate
the aboveground biomass of tropical trees. Glob Chang Biol 20:3177–3190.
https://doi.org/10.1111/gcb.12629
Clark PJ, Evans FC (1954) Distance to Nearest Neighbor as a Measure of Spatial Relationships
in Populations. Ecology 35:445–453. https://doi.org/10.2307/1931034
Condit R, Ashton P, Baker P (2000) Spatial patterns in the distribution of tropical tree species.
Science (80- ) 288:1414–1418. https://doi.org/10.1126/science.288.5470.1414
Costa MNF dos S, Muniz MAP, Negrão CAB, et al (2014) Characterization of Pentaclethra
macroloba oil. J Therm Anal Calorim 115:2269–2275. https://doi.org/10.1007/s10973-
012-2896-z
Cunha AC, Mustin K, dos Santos ES, et al (2017) Hydrodynamics and seed dispersal in the
lower Amazon. Freshw Biol 1–9. https://doi.org/10.1111/fwb.12982
Dantas A, Lira-Guedes A, Guedes M, et al (2020) Population dynamics of Attalea excelsa
(Arecaceae) In floodplain forest of the Amazonian estuary. J Trop For Sci 32:105–113.
https://doi.org/10.26525/jtfs32.2.105
Dantas AR, Lira-Guedes AC, Mustin K, et al (2016) Phenology of the multi-use tree species
Carapa guianensis in a floodplain forest of the Amazon Estuary. Acta Bot Brasilica
30:618–627. https://doi.org/10.1590/0102-33062016abb0282
Dantas AR, Marangon LC, Guedes MC, et al (2017) Spatial distribution of a population of
Pentaclethra macroloba (Willd.) Kuntze in a floodplain forest of the Amazon estuary. Rev
Árvore 41:1–11. https://doi.org/10.1590/1806-90882017000400006
Dawkins HC (1958) The management of natural tropical high-forest with special reference to
Uganda. University of Oxford. Imperial Forestry Institute
Ferreira LV, Stohlgren TJ (1999) Effects of river level fluctuation on plant species richness,
diversity, and distribution in a floodplain forest in Central Amazonia. Oecologia 120:582–

122
587. https://doi.org/10.1007/s004420050893
Fichtler E, Clark DA, Worbes M (2003) Age and Long-term Growth of Trees in an Old-growth
Tropical Rain Forest, Based on Analyses of Tree Rings and 14C1. Biotropica 35:306–317.
https://doi.org/10.1111/j.1744-7429.2003.tb00585.x
Finegan B, Camacho M, Zamora N (1999) Diameter increment patterns among 106 tree species
in a logged and silviculturally treated Costa Rican rain forest. For Ecol Manage 121:159–
176. https://doi.org/10.1016/S0378-1127(98)00551-9
Galván O, Louman B, Galloway G, Obando G (2003) Efecto de la iluminación de la copa sobre
el crecimiento de Pentaclethra macroloba y Goethalsia meiantha e implicaciones para la
silvicultura de los bosques tropicales húmedos Por Turrialba , Costa Rica. Recur. Nat. y
Ambient. 117–126
Goreaud F, Courbaud B, Collinet F (1997) Spatial structure analysis applied to modelling of
forest dynamics: a few examples. Proc IUFRO Work Empir Process based Model For tree
stand growth Simul 21:27. https://doi.org/10.1111/ecog.01579
Hartshorn GS (1983) Pentaclethra macroloba (Gavilan). In: Janzen DH (ed) Costa Rican
Natural History. Universityof Chicago Press, pp 301–303
Hazlett DL (1987) Seasonal Cambial Activity for Pentaclethra, Goelthalsia, and Carapa Trees
in a Costa Rican Lowland Forest. Biotropica 19:357. https://doi.org/10.2307/2388633
IBGE - Instituto Brasileiro de Geografia e Estatística (2012) Manual Técnico da Vegetação
Brasileira. 2a edição revista e ampliada. Sistema Fitogeográfico. Inventário das Formações
Florestais e Campestres. Técnicas e Manejo de Coleções Botânicas. Procedimentos para
Mapeamentos.
INMET (2019) Banco de Dados Meteorológicos para Ensino e Pesquisa.
nmet.gov.br/portal/index.php?r=bdmep/bdmep. Accessed 20 Dec 2019
Janzen DH (1970) Herbivores and the Number of Tree Species in Tropical Forests. Am Nat
104:501–528. https://doi.org/10.1086/282687
Junk WJ, Piedade MTF (2010) An Introduction to South American Wetland Forests:
Distribution, Definitions and General Characterization. In: Amazonian Floodplain Forests:
Ecophysiology, Biodiversity and Sustainable Management. pp 3–25

123
Junk WJ, Piedade MTF, Schöngart J, et al (2011) A classification of major naturally-occurring
amazonian lowland wetlands. Wetlands 31:623–640. https://doi.org/10.1007/s13157-011-
0190-7
Klimas CA, Kainer KA, Wadt LHO (2007) Population structure of Carapa guianensis in two
forest types in the southwestern Brazilian Amazon. For Ecol Manage 250:256–265.
https://doi.org/10.1016/j.foreco.2007.05.025
Köhl M, Neupane PR, Lotfiomran N (2017) The impact of tree age on biomass growth and
carbon accumulation capacity: A retrospective analysis using tree ring data of three
tropical tree species grown in natural forests of Suriname. PLoS One 12:e0181187.
https://doi.org/10.1371/journal.pone.0181187
Kubitzki K, Ziburski A (1994) Seed Dispersal in Flood Plain Forests of Amazonia. Biotropica
26:30. https://doi.org/10.2307/2389108
Lieberman D, Lieberman M, Hartshorn G, Peralta R (1985) Growth rates and age-size
relationships of tropical wet forest trees in Costa Rica. J Trop Ecol 1:97–109.
https://doi.org/10.1017/S026646740000016X
Lopez OR, Kursar TA (2007) Interannual variation in rainfall, drought stress and seedling
mortality may mediate monodominance in tropical flooded forests. Oecologia 154:35–43.
https://doi.org/10.1007/s00442-007-0821-0
Luize BG, Magalhães JLL, Queiroz H, et al (2018) The tree species pool of Amazonian wetland
forests: Which species can assemble in periodically waterlogged habitats? PLoS One
13:1–13. https://doi.org/10.1371/journal.pone.0198130
Metzger JP, Bernacci LC, Goldenberg R (1997) Pattern of tree species diversity in riparian
forest fragments of different widths (SE Brazil). Plant Ecol 133:135–152.
https://doi.org/10.1023/A:1009791831294
Miranda ZP, Guedes MC, Rosa SA, Schöngart J (2018) Volume increment modeling and
subsidies for the management of the tree Mora paraensis (Ducke) Ducke based on the
study of growth rings. Trees 32:277–286. https://doi.org/10.1007/s00468-017-1630-7
Newbery DM, Ridsdale CE (2016) Neighbourhood abundance and small-tree survival in a
lowland Bornean rainforest. Ecol Res 31:353–366. https://doi.org/10.1007/s11284-016-
1345-z

124
Nowacki GJ, Abrams MD (1997) Radial-growth averaging criteria for reconstructing
disturbance histories from presettlement-origin oaks. Ecol Monogr 67:225–249.
https://doi.org/10.1890/0012-9615(1997)067[0225:RGACFR]2.0.CO;2
Oberbauer SF, Strain BR (1985) Effects of light regime on the growth and physiology of
Pentaclethra macroloba (Mimosaceae) in Costa Rica. J Trop Ecol 1:303–320.
https://doi.org/10.1017/S0266467400000390
Oliveira WS, Silva JAM, Rocha CAM (2020) Prospecção Científica e Tecnológica da
Utilização do Óleo de Pracaxi. Cad Propescção 12:1560–1571.
https://doi.org/10.9771/cp.v12i5%20Especial.32629
Parolin P (2002) Submergence tolerance vs. escape from submergence: Two strategies of
seedling establishment in Amazonian floodplains. Environ Exp Bot 48:177–186.
https://doi.org/10.1016/S0098-8472(02)00036-9
Parolin P, De Simone O, Haase K, et al (2004) Central Amazonian Floodplain Forests: Tree
Adaptations in a Pulsing System. Bot Rev 70:357–380. https://doi.org/10.1663/0006-
8101(2004)070[0357:CAFFTA]2.0.CO;2
Parolin P, Lucas C, Piedade MTF, Wittmann F (2010) Drought responses of flood-tolerant trees
in Amazonian floodplains. Ann Bot 105:129–139. https://doi.org/10.1093/aob/mcp258
Parolin P, Wittmann F, Ferreira L V. (2013) Fruit and Seed Dispersal in Amazonian Floodplain
Trees–a Review. Ecotropica19 19–36
Peters CM (1996) The Ecology and Management of Non − Timber Forest Resources. 1–176
Pinto ER (2014) Solos, hidrologia e estrutura populacional de pracuubeiras em florestas de
várzea do estuário amazônico. Universidade Federal do Amapá
R core Team (2019) R: A Language and Environment for Statistical Computing
Reyes G, Brown S, Chapman J, Lugo AE (1992) Wood densities of tropical tree species. Forest
Service
Ripley BD (1981) Spatial Statistics. John Wiley & Sons, Inc., Hoboken, NJ, USA
Rowlingson B, Diggle P (2017) Splancs: Spatial and Space Time Point Pattern Analysis
Sarquis RDSFR, Sarquis ÍR, Sarquis IR, et al (2019) The Use of Medicinal Plants in the
Riverside Community of the Mazagão River in the Brazilian Amazon, Amapá, Brazil:

125
Ethnobotanical and Ethnopharmacological Studies. Evidence-Based Complement Altern
Med 2019:1–25. https://doi.org/10.1155/2019/6087509
Schöngart J (2008) Growth-Oriented Logging (GOL): A new concept towards sustainable
forest management in Central Amazonian várzea floodplains. For Ecol Manage 256:46–
58. https://doi.org/10.1016/j.foreco.2008.03.037
Schöngart J, Bräuning A, Barbosa ACMC, et al (2017) Dendroecological Studies in the
Neotropics: History, Status and Future Challenges. In: Amoroso MM, Daniels LD, Baker
PJ, Camarero JJ (eds) Dendroecology: Tree-Ring Analyses Applied to Ecological Studies.
Springer International Publishing, Gewerbestrasse, pp 35–73
Schöngart J, Gribel R, Ferreira da Fonseca-Junior S, Haugaasen T (2015) Age and Growth
Patterns of Brazil Nut Trees (Bertholletia excelsa Bonpl.) in Amazonia, Brazil. Biotropica
47:550–558. https://doi.org/10.1111/btp.12243
Schöngart J, Piedade MTF, Ludwigshausen S, et al (2002) Phenology and stem-growth
periodicity of tree species in Amazonian floodplain forests. J Trop Ecol 18:581–597.
https://doi.org/10.1017/S0266467402002389
Schöngart J, Piedade MTF, Wittmann F, et al (2005) Wood growth patterns of Macrolobium
acaciifolium (Benth.) Benth. (Fabaceae) in Amazonian black-water and white-water
floodplain forests. Oecologia 145:454–461. https://doi.org/10.1007/s00442-005-0147-8
Schöngart J, Wittmann F, Worbes M, et al (2007) Management criteria for Ficus insipida Willd.
(Moraceae) in Amazonian white-water floodplain forests defined by tree-ring analysis.
Ann For Sci 64:657–664. https://doi.org/10.1051/forest:2007044
Sillett SC, Van Pelt R, Koch GW, et al (2010) Increasing wood production through old age in
tall trees. For Ecol Manage 259:976–994. https://doi.org/10.1016/j.foreco.2009.12.003
Silva JO da., Fernandes RS, Ticli FK, et al (2007) Triterpenoid saponins, new metalloprotease
snake venom inhibitors isolated from Pentaclethra macroloba. Toxicon 50:283–291.
https://doi.org/10.1016/j.toxicon.2007.03.024
Simmons C V, Banov F, Banov D (2015) Use of a topical anhydrous silicone base containing
fatty acids from pracaxi oil in a patient with a diabetic ulcer. SAGE Open Med Case
Reports 3:1–4. https://doi.org/10.1177/2050313X15589676
Sturges HA (1926) The Choice of a Class Interval. J Am Stat Assoc 21:65–66.

126
https://doi.org/10.1080/01621459.1926.10502161
ter Steege H, Prado PI, Lima RAF d., et al (2020) Biased-corrected richness estimates for the
Amazonian tree flora. Sci Rep 10:1–13. https://doi.org/10.1038/s41598-020-66686-3
Valverde-Barrantes OJ, Rocha OJ (2014) Logging impacts on forest structure and seedling
dynamics in a Prioria copaifera (Fabaceae) dominated tropical rain forest (Talamanca,
Costa Rica). Rev Biol Trop 62:347–357. https://doi.org/10.15517/rbt.v62i1.8504
Vilhena JE de S, Lima e Silva RB, Freitas J da L (2018) Climatologia do Amapá: quase um
século de história. Gramma, Rio de Janeiro
Wittmann F, Anhuf D, Funk WJ (2002) Tree species distribution and community structure of
central Amazonian várzea forests by remote-sensing techniques. J Trop Ecol 18:805–820.
https://doi.org/10.1017/S0266467402002523
Worbes M, Fichtler E (2010) Wood Anatomy and Tree-Ring Structure and Their Importance
for Tropical Dendrochronology. In: Junk WJ, Wittmann F, Parolin P, et al. (eds)
Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable
Management. Springer Dordrecht Heidelberg London, pp 329–346
Worbes M, Junk WJ (1989) Dating Tropical Trees by Means of ^14C From Bomb Tests.
Ecology 70:503–507. https://doi.org/10.2307/1937554
Ziburski A (1991) Dissemination, Keimung und Etablierung einiger Baumarten der
Überschwemmungswálder Amazoniens. Trop. und Subtrop. Pflanzenwelt 1–96

127
Síntese
A ampla área de distribuição de P. macroloba na Região Neotropical, evidencia
seu sucesso reprodutivo e sua adaptabilidade aos diferentes tipos de ambientes. A
maré diária do estuário amazônico é um importante fator de seleção e adaptação
morfológica dos pracaxizeiros estuarinos. O período de estiagem favorece a floração,
a atividade polínica e o surgimento dos primeiros frutos. O período chuvoso aumenta
o nível do rio Amazonas que facilmente transborda para o interior da floresta. Esse
aumento hidrometeorológico é propício para o amadurecimento dos frutos e dispersão
das sementes de P. macroloba. A produção de folhas é contínua ao longo do ano, não
havendo relação significativa com a estação do ano. Vespas e abelhas são os
principais visitantes florais de P. macroloba. A taxa de predação de sementes é baixa
(5%) e larvas de mariposa são os principais predadores. Ocorre uma grande
variabilidade no tamanho das sementes produzidas por pracaxizeiros situados ao
longo do estuário amazônico, mostrando as múltiplas estratégias de colonização
dessa espécie. Variações intrapopulacionais também foram observadas na
quantidade de frutos e sementes produzidas. P. macroloba é uma espécie secundária
tardia de longa vida e que necessita de clareiras para liberação de seu crescimento.
A estruturação dos indivíduos na floresta e o padrão de crescimento apresentados por
P. macroloba, são reflexos da sua história ecológica e de suas respostas
ecofisiológicas à inundação diária do estuário amazônico.
Neste estudo, elucidamos uma grande lacuna do conhecimento sobre a história
natural de populações de P. macroloba no estuário amazônico. O resultado desta
pesquisa será fundamental para buscar políticas públicas para o bom manejo e
conservação dos pracaxizeiros amazônicos e para fortalecer cadeias produtivas dos
produtos não madeireiros como geradoras de renda e de inclusão social das
populações ribeirinhas ao longo do Rio Amazonas.

128
Referências bibliográficas
Banov, D.; Banov, F.; Bassani, A.S. 2014. Case Series: The effectiveness of fatty acids
from pracaxi oil in a topical silicone base for scar and wound therapy. Dermatology and Therapy 4: 259–269.
Bennett, B.; Breed, M.D. 1985. On the association between Pentaclethra macroloba (Mimosaceae) and Paraponera clavata (Hymenoptera: Formicidae) Colonies. Biotropica 17: 253.
Braga, A.C.M. 2018. Uso de efluente de fossa como biofertilizante nitrogenado em mudas de açaí (Euterpe oleracea Mart.) e pracaxi (Pentaclethra macroloba (Willd.) Kuntze) sob duas condições de luminosidade. Universidade Federal do Amapá, Macapá, Amapá. 88p.
Carim, M.D.J.V.; Jardim, M.A.G. 2008. Composição florística e estrutura de floresta de várzea no município de Mazagão, Estado do Amapá, Brasil. Scientia Forestalis 36: 191–201.
Costa, M.N.F.S.; Muniz, M.A.P.; Negrão, C.A.B.; Costa, C.E.F.; Lamarão, M.L.N.; Morais, L.; et al. 2014. Characterization of Pentaclethra macroloba oil. Journal of Thermal Analysis and Calorimetry 115: 2269–2275.
Crespi, B.; Guerra, G.A.D. 2013. Ocorrência, coleta, processamento primário e usos do pracaxi (Pentaclethra macroloba (Willd.) Kuntze) na Ilha de Cotijuba, Belém- PA. Revista Brasileira de Agroecologia 8: 176–189.
Cunha, A.C.; Mustin, K.; Santos, E.S.; Santos, É.W.G.; Guedes, M.C.; Cunha, H.F.A.; et al. 2017. Hydrodynamics and seed dispersal in the lower Amazon. Freshwater Biology: 1–9.
Dantas, A.R.; Marangon, L.C.; Guedes, M.C.; Feliciano, A.L.P.; Lira-Guedes, A.C. 2017. Spatial distribution of a population of Pentaclethra macroloba (Willd.) Kuntze in a floodplain forest of the Amazon estuary. Revista Árvore 41: 1–11.
Eaton, W.D.; Anderson, C.; Saunders, E.F.; Hauge, J.B.; Barry, D. 2012. The impact of Pentaclethra macroloba on soil microbial nitrogen fixing communities and nutrients within developing secondary forests in the Northern Zone of Costa Rica. Tropical Ecology 53: 207–214.
Hartshorn, G. 1972. Ecological lifeHistory and population dynamics of Pentaclethra macrolobaand Stryphnodendron excelsum. University of Washington, 76p.
Hartshorn, G.S. 1983. Pentaclethra macroloba (Gavilan). In: Janzen, D.H. (Ed.), Costa Rican Natural History, Universityof Chicago Press, p.301–303.
Junk, W.; Piedade, M. 2000. Concepts for the sustainable management of natural resources of the middle amazon floodplain: a summary. German-Brazilian Workshop on Neotropical Ecosystems-Achievements and Prospects of Cooperative Research: 697–702.
Junk, W.J. 2000. The central amazon river floodplain: conceptos for the sustainable use of its resources. In: Junk, W.J.; Ohly, J.J.; Piedade, M.T.F.; Soares, M.G.M. (Eds.), The Central Amazon Floodplain: Actual Use and Options for a Sustainable Management, Backhuys Publishers, Leiden, p.75–94.

129
Junk, W.J.; Piedade, M.T.F. 2010. An Introduction to South American Wetland Forests: Distribution, Definitions and General Characterization. In: Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable Management, p.3–25.
Junk, W.J.; Bayley, P.B.; Sparks, R.E. 1989. The flood pulse concept in river-floodplain systems. International Large River Symposium: 110–127.
Junk, W.J.; Piedade, M.T.F.; Schöngart, J.; Wittmann, F. 2012. A classification of major natural habitats of Amazonian white-water river floodplains (várzeas). Wetlands Ecology and Management 20: 461–475.
Junk, W.J.; Piedade, M.T.F.; Cunha, C.N. da; Wittmann, F.; Schöngart, J. 2018. Macrohabitat studies in large Brazilian floodplains to support sustainable development in the face of climate change. Ecohydrology and Hydrobiology 18: 334–344.
Junk, W.J.; Piedade, M.T.F.; Schöngart, J.; Cohn-Haft, M.; Adeney, J.M.; Wittmann, F. 2011. A classification of major naturally-occurring amazonian lowland wetlands. Wetlands 31: 623–640.
Junk, W.J.; Piedade, M.T.F.; Lourival, R.; Wittmann, F.; Kandus, P.; Lacerda, L.D.; et al. 2014. Brazilian wetlands: Their definition, delineation, and classification for research, sustainable management, and protection. Aquatic Conservation: Marine and Freshwater Ecosystems 24: 5–22.
Latrubesse, E.M.; Franzinelli, E. 2002. The Holocene alluvial plain of the middle Amazon River, Brazil. Geomorphology 44: 241–257.
Lima, R.P.; Luz, P.T.S; Braga, M.; Batista, P.R.S.; Costa, C.E.F.; Zamian, J.R.; et al. 2017. Murumuru (Astrocaryum murumuru Mart.) butter and oils of buriti (Mauritia flexuosa Mart.) and pracaxi (Pentaclethra macroloba (Willd.) Kuntze) can be used for biodiesel production: Physico-chemical properties and thermal and kinetic studies. Industrial Crops and Products 97: 536–544.
Maciel, M.N.M.; Watzlawick, L.F.; Schoeninger, E.R.; Yamaji, F.M. 2003. Classificação ecológica das espécies arbóreas. Revista Acadêmica: ciências agrárias e ambiente 1: 69–78.
McClain, M.E.; Naiman, R.J. 2008. Andean influences on the biogeochemistry and ecology of the Amazon River. BioScience 58: 325–338.
Meade, R.H. 2008. Transcontinental Moving and Storage: The Orinoco and Amazon Rivers Transfer the Andes to the Atlantic. Large Rivers: Geomorphology and Management: 45–63.
Melack, J.M.; Hess, L.L. 2010. Remote Sensing of the Distribution and Extent of Wetlands in the Amazon Basin. In: Junk, W.J.; Piedade, M.T.F.; Wittmann, F.; Schöngart, J.; Parolin, P. (Eds.), Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable Management, Springer Berlin Heidelberg, p.43–59.
Melack, J.M.; Novo, E.M.L.M.; Forsberg, B.R.; Piedade, M.T.F.; Maurice, L. 2009. Floodplain ecosystem processes. In: Keller, M.; Bustamante, M.; Gash, J.; Dias, P.S. (Eds.), Amazonia and Global Change, p.525–541.
Oberbauer, S.F.; Strain, B.R. 1985. Effects of light regime on the growth and physiology of Pentaclethra macroloba (Mimosaceae) in Costa Rica. Journal of

130
Tropical Ecology 1: 303–320.
Oberbauer, S.F.; Strain, B.R.; Riechers, G.H. 1987. Field water relations of a wet-tropical forest tree species. Oecologia 71: 369–374.
Oliveira, W.S.; Silva, J.A.M.; Rocha, C.A.M. 2020. Prospecção Científica e Tecnológica da Utilização do Óleo de Pracaxi. Cadernos de Propescção 12: 1560–1571.
Pesce, C. 2009. Oleaginosas da Amazônia. 2nd ed. Museu Paraense Emílio Goeldi, Bélem, 334p.
Piedade, M.T.F.; Schöngart, J.; Wittmann, F.; Bleich, M.E.; Lopes, A. 2015. Iniciando a viagem pelas Áreas Úmidas Amazônica. In: Lopes, A.; Piedade, M.T.F. (Eds.), Conhecendo as Áreas Úmidas Amazônicas: Uma Viagem Pelas Várzeas e Igapós, Editora INPA, Manaus, p.14–22.
Pinedo-Vasquez, M.A.; Sears, R.R. 2011. Várzea Forests: Multifunctionality as a Resource for Conservation and Sustainable Use of Biodiversity. In: Pinedo-Vasquez, M.; Ruffino, M.L.; Padoch, C.; Brondízio, E.S. (Eds.), The Amazon Várzea: The Decade Past and the Decade Ahead, Springer Netherlands, Dordrecht, p.187–206.
Queiroz, J.A.L. De; Machado, S.D.A.; Hosokawa, R.T.; Silva, I.C. 2007. Estrutura e dinâmica de floresta de várzea no estuário amazônico no Estado do Amapá. Floresta 37: 339–352.
Rabelo, B.V. 2000. Atlas: Zoneamento Ecológico Econômico da área sul do Estado do Amapá. IEPA, Macapá, 44p.
Rabelo, B.V. 2005. Mazagão: realidades que devem ser conhecidas. IEPA, Macapá, 68p.
Santos, G.C. dos; Jardim, M.A.G. 2006. Florística e estrutura do estrato arbóreo de uma floresta de várzea no município de Santa Bárbara do Pará, Estado do Pará, Brasil. Acta Amazonica 36: 437–446.
Sawakuchi, H.O.; Bastviken, D.; Sawakuchi, A.O.; Krusche, A. V.; Ballester, M.V.R.; Richey, J.E. 2014. Methane emissions from Amazonian Rivers and their contribution to the global methane budget. Global Change Biology 20: 2829–2840.
Schöngart, J. 2008. Growth-Oriented Logging (GOL): A new concept towards sustainable forest management in Central Amazonian várzea floodplains. Forest Ecology and Management 256: 46–58.
Schöngart, J.; Piedade, M.T.F.; Ludwigshausen, S.; Horna, V.; Worbes, M. 2002. Phenology and stem-growth periodicity of tree species in Amazonian floodplain forests. Journal of Tropical Ecology 18: 581–597.
Shebitz, D.J.; Eaton, W. 2013. Forest structure, nutrients, and Pentaclethra macroloba growth after deforestation of Costa Rican lowland forests. ISRN Ecology 2013: 27–31.
Shebitz, D.J.; Gomez, R.; Casimir, A.; Sciences, L.; Union, M.A. 2013. A preliminary study of Nicaraguan and Costa Rican medicinal plant knowledge in the Maquenque National Wildlife Refuge. Journal of Medicinal Plants Research 7:

131
790–798.
Silva, J.O.; Fernandes, R.S.; Ticli, F.K.; Oliveira, C.Z.; Mazzi, M. V.; Franco, J.J.; et al. 2007. Triterpenoid saponins, new metalloprotease snake venom inhibitors isolated from Pentaclethra macroloba. Toxicon 50: 283–291.
Silva, J.O.; Coppede, J.S.; Fernandes, V.C.; Sant’Ana, C.D.; Ticli, F.K.; Mazzi, M. V.; et al. 2005. Antihemorrhagic, antinucleolytic and other antiophidian properties of the aqueous extract from Pentaclethra macroloba. Journal of Ethnopharmacology 100: 145–152.
Sioli, H. 1968. Principais biótopos de produção primária nas águas da Amazônia. Proc. Symp. Recent Adv. Trop. Ecol: 1–10.
ter Steege, H.; Prado, P.I.; Lima, R.A.F. d.; Pos, E.; de Souza Coelho, L.; de Andrade Lima Filho, D.; et al. 2020. Biased-corrected richness estimates for the Amazonian tree flora. Scientific Reports 10: 1–13.
Teixeira, R. da S.; Rocha, P.R.; Polonini, H.C.; Brandão, M.A.F.; Chaves, M. G.A.M.; Raposo, N.R.B. 2012. Mushroom tyrosinase inhibitory activity and major fatty acid constituents of Amazonian native flora oils. Brazilian Journal of Pharmaceutical Sciences 48: 399–404.
Wittmann, F.; Junk, W.J. 2016. Amazon River Basin. In: Finlayson, C.M.; Everard, M.; Irvine, K.; McInnes, R.; Middleton, B.; van Dam, A.; et al. (Eds.), The Wetland Book, Springer Netherlands, Dordrecht, p.1–20.
Wittmann, F.; Schöngart, J.; Junk, W.J. 2010a. Phytogeography, Species Diversity, Community Structure and Dynamics of Central Amazonian Floodplain Forests. In: Junk, W.J.; Piedade, M.T.F.; Wittmann, F.K.; Schongart, J.; Parolin, P. (Eds.), Amazonian Floodplain Forests, Amazonian Floodplain Forests: Ecophysiology, Biodiversity and Sustainable Management, Ecological Studies., Springer Dordrecht Heidelberg London, London, p.61–102.
Wittmann, F.; Schongart, J.; Montero, J.C.; Motzer, T.; Junk, W.J.; Piedade, M.T.F.; et al. 2006. Tree species composition and diversity gradients in white-water forests across the Amazon Basin. Journal of Biogeography 33: 1334–1347.
Wittmann, F.; Schongart, J.; Brito, J.M.; Wittmann, A.D.O.; Piedade, M.T.F.; Parolin, P.; et al. 2010b. Manual of trees from Central Amazonian várzea floodplains. INPA, Manaus, 286p.
Wittmann, F.; Householder, E.; Piedade, M.T.F.; De Assis, R.L.; Schöngart, J.; Parolin, P.; et al. 2013. Habitat specifity, endemism and the neotropical distribution of Amazonian white-water floodplain trees. Ecography 36: 690–707.





















![E:\TCC Paulo Ricardo Versão final [6314]](https://img.pdfslide.net/doc/110x75/6321478e537c10e8380294e2/etcc-paulo-ricardo-versao-final-6314.jpg)







