Embed Size (px)
Citation preview

Vocal and LocomotorResponses of Piglets to SocialIsolation and Reunion
Valentina Colonnello1,2
Paolo Iacobucci1,2
Ruth C. Newberry1
1Department of Animal Sciencesand Department of Veterinary
and Comparative AnatomyPharmacology and Physiology
Center for the Study ofAnimal Well-being
Washington State UniversityPullman, WA 99164-6351
E-mail: [email protected]
2Department of PsychologyUniversity of Rome ‘‘Sapienza’’
Via dei Marsi 78, 00185 Rome, Italy
ABSTRACT: Potentiation of infant isolation calls following a brief reunion withthe mother is considered an index of filial bonding in altricial rodents. Weinvestigated potentiation of isolation and reunion responses in 15-day-oldunweaned domestic piglets (Sus scrofa domesticus). When piglets were re-isolatedfollowing a brief, comforting reunion with their mother and littermates in theirhome pen, they displayed a persistence (relative potentiation) of calling andjumping. In contrast, when re-isolated following a brief interaction with theirmother or an unfamiliar sow in a familiar or unfamiliar location, or with littermatesalone, calling rate and locomotion dropped. Subsequently, piglets spent more timenear their mother’s face if they had previously interacted with an unfamiliar sowrather than their mother, consistent with maternal recognition. Although we did notdetect maternal potentiation as described in altricial rodents, filial attachmentbonding was clearly evident in piglet responses, especially during reunions. � 2009Wiley Periodicals, Inc. Dev Psychobiol 52: 1–12, 2010.
Keywords: maternal potentiation; filial attachment; social bonds; vocalizations;domestic pig; swine
INTRODUCTION
In humans, a filial attachment bond can be distinguished
from other kinds of social relationships by the selective
seeking of proximity to a caregiver to obtain security and
comfort, in conjunction with emotional distress during
periods of involuntary separation from the caregiver,
emotionally significant reunions, and a persistent as
opposed to transitory bond (Ainsworth, 1989; Bowlby,
1973; Cassidy, 1999). In other mammals, some degree of
filial attachment of an infant to its mother is needed for
nurturance of the infant. However, there is variation across
species in the strength and selectivity of social attachment
depending on life history variables (Newberry &
Swanson, 2008), and not all filial relationships meet the
criteria for an attachment bond as described in humans.
Filial attachment in humans has been investigated
by means of separation-reunion procedures, such as the
‘‘strange situation’’ test (Ainsworth, 1989; Polan & Hofer,
1999). Infant responses to separation have also been
investigated extensively in rodent models (Herman &
Panksepp, 1978; Hofer, Brunelli, & Shair, 1993; Hofer &
Shair, 1978; Moles, Kieffer, & D’Amato, 2004; Muller,
Brunelli, Moore, Myers, & Shair, 2005; Panksepp,
Nelson, & Bekkedal, 1997; Pettijohn, 1979). Analysis of
vocalizations following separation or isolation has been of
particular interest for gaining insights into rodent filial
attachment (Brunelli & Hofer, 2007; Iacobucci, Colon-
nello, Pavan, & Ricceri, 2007; Shair, 2007).
Studies exploiting a repeated isolations test, which is
a three-session procedure involving a first isolation
followed by a brief reunion with the caregiver and
then a second isolation, have revealed a ‘‘potentiation’’ of
calls by infant laboratory rats during the second isolation
(Hofer, Brunelli, Masmela, & Shair, 1996; Hofer,
Brunelli, & Shair, 1994; Hofer, Masmela, Brunelli, &
Developmental Psychobiology
Received 4 February 2009; Accepted 10 August 2009Valentina Colonnello and Paolo Iacobucci contributed equally to this
work.Correspondence to: R. C. NewberryContract grant sponsor: National Council of the University of Rome
‘‘Sapienza,’’ ItalyPublished online 22 September 2009 in Wiley InterScience
(www.interscience.wiley.com). DOI 10.1002/dev.20406
� 2009 Wiley Periodicals, Inc.

Shair, 1998; Shair, 2007; Shair, Masmela, Brunelli, &
Hofer, 1997). Specifically, whereas control pups that
remain isolated in the second session of the test show a
reduction in calling during the third session, pups that
are re-isolated after experiencing a brief reunion with
their mother call at over two times the rate observed in the
first isolation, an effect attributed to a reward expectancy
violation (Hofer et al., 1996; Polan & Hofer, 1999).
This phenomenon, dubbed ‘‘maternal potentiation,’’ is
selectively induced by reunion with the mother and not
by littermates or home cage shavings. Nevertheless,
interaction with an unfamiliar adult female during session
2 of the test provokes a similar increase in calling rate
indicating that, despite the terminology, the potentiation is
not elicited solely by the rat’s mother. Furthermore,
when rat pups are reared in the presence of an adult
male, potentiation of calling also emerges after a
brief interaction with an unfamiliar adult male (reviewed
by Shair, 2007). A ‘‘contact quieting’’ effect, whereby
the rate of calling decreases dramatically during reunions
with the mother, has also been reported in rats (Hofer &
Shair, 1978; Shair, 2007). This effect is less selective
than maternal potentiation, being evoked by many
other factors that could be perceived as comforting
including exposure to littermates, warmth, a furry sub-
strate and home cage bedding (Hofer & Shair, 1978; Shair,
2007).
The human filial attachment bond is a unique
psychological relationship of the infant to the caregiver
that is highly emotionally significant during both
separations and reunions (Ainsworth, 1989). Data do not
robustly support the existence of this kind of bond in infant
laboratory rats. Among several critical aspects (for a
review, see Panksepp, 2003), we consider the lack of
specific greeting responses upon reunion with the dam or
littermates as a main issue. Even so, the rat studies suggest
that the phenomenon of maternal potentiation may be
useful for characterizing the emotional content of social
bonds in species showing more selective filial attachment.
Research using the repeated isolations procedure in
the guinea pig provides some support for this idea. In
the guinea pig, exposure to the mother does not increase
the rate of calling by infants in the second isolation, as
found in rats, but it does prevent the decrease in the
rate of calling observed in control pups (Hennessy, Miller,
& Shair, 2006). This relative, although not absolute,
potentiation is selective for the mother and is not observed
following interaction with littermates or other adult
females. Although a promising finding, the external
validity of maternal potentiation as an index of infant
emotional attachment to the mother (and other caregivers)
would be strengthened by evidence for this phenomenon
in other mammalian species exhibiting filial attachment
bonds.
In the present study, we investigated maternal poten-
tiation in the domestic pig, Sus scrofa domesticus, a
widely available precocial species that is endowed with a
sophisticated social life commencing with the develop-
ment of maternal recognition soon after birth (Morrow-
Tesch & McGlone, 1990; Newberry & Wood-Gush, 1985,
1986; Parfet & Gonyou, 1991; Schon, Puppe, Gromyko, &
Manteuffel, 1999). Piglets and sows are highly vocal
during typical social interactions, such as suckling bouts
and reunion episodes, and during brief periods of
involuntary isolation (Algers, 1993; Illmann, Schrader,
Spinka & Sustr, 2002; Puppe, Schon, & Wendland, 1999;
Spinka, Stehulova, Zacharova, Maletinska, & Illmann,
2002; Weary, Appleby, & Fraser, 1999; Weary, Ross, &
Fraser, 1997). When isolated, pigs usually begin
emitting relatively quiet, low-pitched grunts interspersed
with silence (listening). If a reunion is not immediately
forthcoming, isolation-induced calls escalate in rate,
pitch, volume, and duration (Fraser, 1975), correlated
with activation of a physiological stress response
(Schrader & Todt, 1998). As in reunions between infants
and caretakers in humans (Ainsworth, Blehar, Waters, &
Wall, 1978), reunions between piglets and sows are
characterized by an active ‘‘greeting’’ interaction. Imme-
diately upon reunion with their mother, piglets typically
emit a rapid series of ‘‘quack’’ calls near the sow’s face
(Illmann et al., 2002). Young piglets direct these greeting
calls specifically towards their mother, although occa-
sionally older piglets may also direct these calls towards
other familiar adults in the social group (Newberry,
personal observation). Piglets show signs of individual
recognition of their mother’s voice, responding to her
nursing calls by running directly towards her rather than to
other sows emitting nursing calls during synchronized
nursing bouts (Newberry & Wood-Gush, 1985). There is
also evidence that sows are able to recognize their own
piglets by voice (Appleby, Weary, Taylor, & Illmann,
1999; Illmann et al., 2002; Maletinska, Spinka, Vichova,
& Stehulova, 2002; Weary, Lawson, & Thompson, 1996).
From these findings, it can be inferred that domestic
piglets form a selective attachment bond with their
mother.
The aim of our study was twofold: (1) to investigate the
face validity of the domestic pig for studies of filial
attachment bonding, and (2) to investigate whether the
level of maternal potentiation detected in laboratory rats
can be found in a nonrodent species exhibiting filial
attachment bonding. We focused on two criteria charac-
terizing infant attachment bonds: responses to involuntary
separation and responses to reunion. Since reunion calls
are evident in piglet–sow interactions (in contrast to rat
infant–mother interactions), we decided to observe the
reunion subsequent to the second as well as the first
isolation session, introducing a fourth session into the
Developmental Psychobiology2 Colonnello, Iacobucci, and Newberry

classic repeated isolations procedure. To establish
whether young piglets exhibit an emotional response to
involuntary displacement from, and reunion with, their
mother in the absence of an immediate need for warmth
or nourishment, we conducted the research in a warm
environment, excluded relatively small piglets from our
sample, and tested piglets after completion of nursing
episodes. We took these steps to rule out potential
confounding effects as it is known that small, cold, and
hungry piglets call more when isolated, and elicit a
stronger response from their mother, than larger, warmer
and more satiated piglets (Weary & Fraser, 1995; Weary
et al., 1996, 1997).
The study comprised four experiments. In Experiment
1, we evaluated both male and female piglets for the
presence of maternal potentiation. We predicted that a
brief reunion with the mother and littermates in the home
pen following a temporary separation would increase (i.e.,
potentiate) the rate of calling during a second isolation
whereas control piglets not experiencing the reunion
would show a decrease in calling rate during the second
isolation period. If a brief reunion increases the aversive-
ness of a subsequent isolation, it is possible that it would
also increase the rewarding nature of the following
reunion. We predicted, therefore, that the reunited piglets
would show a stronger vocal response during a second
reunion than the control piglets.
The results of Experiment 1 indicated a persistence
of isolation calls after reunion with the mother and
littermates. To understand whether this effect was induced
specifically by interaction with the mother, in Experiment
2, we focused on the vocal and locomotor responses of
piglets when re-isolated following a brief interaction
with their mother or an unfamiliar adult female alone in
an unfamiliar location. We hypothesized that piglet
responses to re-isolation are specifically influenced by
the brief interaction with their mother. Our prediction was
that piglets briefly reunited with their mother, but not an
unfamiliar sow, would call at a greater rate in the second
than the first isolation. By contrast, we predicted that
interaction with an unfamiliar female would produce a
decline in call rate during the second isolation. We also
expected a difference in the rate of calls emitted during the
reunions. In particular, we predicted a stronger response in
piglets exposed to the mother during both the reunion
sessions. There was an absence of maternal potentiation in
Experiment 2, possibly because the sows were agitated in
the unfamiliar location used for reunions. Therefore, in
Experiment 3, we exposed piglets to their mother or an
unfamiliar sow alone in the sow’s home pen. Predictions
were as for Experiment 2. In Experiment 4, we
investigated whether the persistence of calling detected
in Experiment 1 could be induced by exposure to siblings,
or the familiar home pen, alone. We hypothesized that
neither of these conditions would be sufficient to elicit a
potentiation effect.
Because we found no sex differences in Experiment 1,
we conducted Experiments 2, 3, and 4 with females only.
We were especially interested in female responses
because, in a natural context, pigs live in a matrilinear
society comprising several sows and their young, and
female offspring tend to remain in the natal group as adults
and maintain long-term relationships with their mother
whereas males disperse (Gonyou, 2001; Graves, 1984;
Newberry & Wood-Gush, 1986).
All testing was carried out on postnatal day 15. At this
age, the mother-young relationship is well established and
nursing episodes occur approximately once per hour. In a
natural context, the piglets would be actively exploring
their environment at this age, resulting in brief voluntary
separations from the mother and the use of vocalizations
and locomotion to relocate the mother in different
locations within the home range (Newberry & Wood-
Gush, 1985, 1986).
METHODS
Subjects, Housing, and Husbandry
Subjects were 15-day-old crossbred domestic piglets (genetic
background: sows predominantly Large White and boars from
Landrace, Yorkshire, Duroc and Hampshire lineages) housed
with their mother (1st to 6th parity) and littermates in 2.1 m
long� 1.8 m wide farrowing pens with porous floors. The pens
were located in farrowing rooms containing 12 pens within the
Washington State University Swine Center. Average litter size
was 11 piglets (range 8–14 piglets). In accordance with standard
practices in the United States, piglets were weighed at birth,
tooth clipped, tail docked, ear notched, and vaccinated against
atrophic rhinitis on postnatal day (PND) 2, and males were
castrated on PND 7. Room temperature was 17� 1�C, and a heat
lamp in each pen provided supplementary heat for the piglets.
Fluorescent lighting was provided for 11 hr daily, with lights on
at 0530 hr. Sows were fed a standard lactation diet twice daily
and provided with water ad libitum.
Treatments
Experiment 1—With Mother and Littermates in Home Pen
Versus Empty Pen. Four piglets (2 males and 2 females) of
similar body weight at birth, from each of eight litters born
within a 12-day period, were selected for testing. Small piglets
(runts) were excluded from the study. Of the four selected
subjects per litter, one male and one female were randomly
assigned to a Home Pen (HP) treatment group and the other male
and female to an Empty Pen (EP) treatment group (within litter
design). They were marked on the back with a wax livestock
marker 48 hr prior to testing to facilitate identification. Each
subject (n¼ 32) was subjected to a four-session test on PND 15:
(1) a 7-min isolation session (Iso1), (2) a 1-min reunion session
Developmental Psychobiology Isolation and Reunion Responses of Piglets 3

(R1), (3) a second 7-min isolation session (Iso2), and
(4) a second 1-min reunion session (R2). During the two
isolation sessions, piglets were individually isolated in an empty
1.7 m� 1.5 m observation arena with a concrete floor, located in
an empty observation room. During R1, piglets in the HP group
were placed in their home pen containing their mother and
littermates and piglets whereas piglets in the EP group were
placed alone in an empty farrowing pen of the same size and
features as their home pen, located in an empty farrowing room.
During R2, piglets from both groups were reunited with their
mother and littermates in their home pen. The same procedures
were used in all subsequent experiments with the following
exceptions.
Experiment 2—With Mother Versus Stranger in Unfamiliar
Pen. One female from each of eight litters was assigned to a
Mother in Unfamiliar Pen (MUP) treatment group and a second
female per litter was assigned to a Stranger in Unfamiliar Pen
(SUP) treatment (within litter design). For the test, each piglet
was: (1) isolated for 7 min (Iso1), (2) placed in a farrowing pen
alone with her biological mother (MUP group) or an unfamiliar
lactating sow (SUP group) in an empty farrowing room for 1 min
(R1), (3) re-isolated for 7 min (Iso2), and (4) reunited with her
mother and littermates in the home pen for 1 min (R2). The
mother or stranger was moved temporarily into the empty
farrowing room and placed alone in a farrowing pen with
food and water 2–9 min before R1. The unfamiliar sow was
genetically unrelated to the piglet being tested (i.e., no aunts or
adult sisters were used) and, to ensure that the sow’s appearance,
odor and vocalizations were unfamiliar to the piglet, was from a
different farrowing room to that in which the piglet resided. A
different unfamiliar sow was used for testing the SUP piglet of
each litter. After the piglet was removed for Iso2, the sow was
returned to her home pen for R2.
Experiment 3—With Mother Versus Stranger in Sow’s Home
Pen. In this experiment, one female from each of eight litters
was assigned to a Mother in Home Pen (MHP) treatment group
while a second female was assigned to a Stranger in her
Home Pen (SHP) treatment (within litter design). Each piglet
was: (1) isolated for 7 min (Iso1), (2) placed in her own farrowing
pen alone with her biological mother (MHP) or alone with an
unfamiliar lactating sow in the stranger’s home pen (SHP) for
1 min (R1), (3) re-isolated for 7 min (Iso2), and (4) reunited with
her mother and littermates in the home pen for 1 min (R2). All
piglets from the home pen of the mother or stranger were
removed prior to R1 (placed in an empty farrowing room as a
littermate group), and returned prior to R2. The stranger sow was
genetically unrelated to the piglet being tested, but was from the
same farrowing room to that in which the piglet resided. A
different stranger sow was used for testing the SHP piglet of each
litter.
Experiment 4—With Littermates in Home Pen Versus Empty
Home Pen. Two female piglets from each of eight litters were
randomly assigned to a Littermates in Home Pen (LHP) or
Empty Home Pen (EHP) treatment group, respectively (within
litter design). Each subject was tested in: (1) a 7-min isolation
session (Iso1), (2) a 1-min reunion session (R1) in the home
pen with her littermates (LHP) or in the home pen alone (EHP),
(3) a second 7-min isolation session (Iso2), and (4) a second
1-min reunion session (R2) with her mother and littermates in the
home pen. Mothers in LHP, and mothers and littermates in EHP,
were removed to a farrowing pen in an empty farrowing room
prior to R1, and returned prior to R2.
General Procedures
Subjects from each litter were tested in counterbalanced order
for sex (Experiment 1) and experimental treatment. All testing
was carried out between 0900 and 1500 hr. Because different
litters within a farrowing group were born on different days, tests
at PND 15 were conducted over multiple days. The durations of
the isolation and reunion sessions were selected taking into
consideration the frequency of nursing episodes (about one per
hour) and our intention to observe separation responses without
inducing excessive stress or fatigue in piglets. The reunion
sessions were particularly brief given that ‘‘greeting’’ calls are
emitted only briefly and immediately upon reunion with the
mother (Illmann et al., 2002).
It took approximately 40 s to transport piglets between
farrowing rooms and the observation room where Iso1 and Iso2
occurred. Piglets were carried in a clean opaque gray box
(61 cm� 40 cm� 42 cm), used to minimize stress related to
handling. Preliminary observations indicated that piglets emitted
few calls when transported in the box (data not reported). During
reunion sessions, piglets were introduced from the back of the
pen to minimize any disturbance to the sow resulting from
human presence. The temperature in the farrowing rooms and
observation room was set at an equivalent level and adjusted
prior to each test if needed. After each isolation session, the floor
of the observation arena was cleaned with paper towels and a
diluted alcohol solution, and two fans were activated to eliminate
odor. The empty farrowing room used for reunion sessions in
Experiment 2 was cleaned with a pressure hose at the end of each
reunion session.
To avoid testing when piglets were hungry, piglets were not
disturbed during nursing bouts. Since nursing occurred at
intervals of approximately 1 hr, we were able to test two piglets
per inter-nursing interval, starting after the litter completed
the post-letdown teat massage phase of the nursing bout
(Algers, 1993; Jensen, Gustafsson, & Augustsson, 1998). During
Experiment 1, the sow started to nurse her piglets during R1 on
three occasions. In these cases, the test was terminated and a
substitute piglet from that litter, of the same sex and approximate
birth weight as the original subject, was tested after the nursing
bout had finished.
Recording Equipment
During isolation sessions, data were collected using a video
camera (Sony DCR-HC21 Handycam MiniDV Camcorder,
Sony Electronics Inc., San Diego, CA) and a uni-directional
(cardiod) microphone (Sennheiser/Microphone Madness
MM-HLSC-Lapel; working range 30–20,000 Hz; Microphone
Madness, Inc., Palm Coast, FL) suspended from the ceiling. The
Developmental Psychobiology4 Colonnello, Iacobucci, and Newberry

camera and microphone were connected to a video monitor and
computer, respectively, located in an adjacent room. The
computer was equipped with a Creative Sound Blaster 16 PCI
sound card (recording sampling rate 48 kHz, 16 bit; Creative
Labs, Inc., Milpitas, CA) and ‘‘SeaPro’’ sound recording
software (CIBRA, http://www.unipv.it/webcib/res_software_
uk.html, University of Pavia, Pavia IT). To avoid the recording
of calls associated with handling during introduction to the
arena, recording started 10 s following placement of the piglet
into the arena. A Sony DCR-HC21 Handycam MiniDV
Camcorder was used to collect audiovisual recordings during
the reunion sessions.
Data Collected
We recorded both vocalizations and locomotor activity to
elucidate whether the hypothesized maternal potentiation in pigs
was restricted solely to calling as in rats (Hofer et al., 1996;
Myers et al., 2004; Shair et al., 1997), or extended to activity as
reported in guinea pigs (Hennessy et al., 2006). In addition to
determining the number of calls uttered by the piglet during the
7-min isolation sessions, we assessed two measures of locomotor
activity: (1) the number of crossings of virtual lines dividing the
observation arena into five equal squares as viewed on a monitor
during videotape analysis, where a crossing was counted when a
line was crossed with all four trotters, and (2) the number of
jumps, defined as jumping up against the wall with all four
trotters leaving the floor. During the 1-min reunion sessions, we
determined the rate of calling and the time spent in proximity to
the sow’s face, defined as the total number of seconds spent with
the piglet’s head within 10 cm of the sow’s mouth. All behavioral
data were collected by trained observers blind to the treatments,
and exhibiting high inter-observer concordance (>95%).
In Experiment 1, we recorded piglet body temperature just
before Iso1 and just after Iso2 using an infrared thermometer to
confirm whether body temperature of piglets in the two treatment
groups remained unaffected by exposure to different rooms
during R1. We did not collect body temperatures in the other
experiments because R1 sessions were conducted in the same
room for all piglets.
Statistical Analysis
Statistical analyses were conducted using Statview II software
(Abacus Concepts, Inc., Berkeley, CA). Because the isolation
and reunion sessions differed in duration and context, the data on
responses to isolation and reunion were analyzed separately for
each experiment. Except where noted, data were analyzed using
ANOVA, with litter as a blocking factor. Jumping frequencies
during isolations were not normally distributed and, therefore, an
SQRT transformation was performed prior to analysis. Post-hoc
means comparisons were performed using Tukey’s Honestly
Significant Differences test.
For Experiment 1 isolation sessions, we analyzed treatment
and sex effects on vocalizations, line crossing, jumping, and
body temperature, with test session (Iso1 vs. Iso2) as a repeated
measure. In R1, piglets in the EP treatment were isolated and,
therefore, their behavior was not comparable with that of HP
piglets during R1, or behavior in R2. Therefore, we compared
vocalizations and time spent near the sow’s face during R1
versus R2 sessions by piglets in the HP treatment only, with sex
as a main effect and test session (R1 vs. R2) as a repeated
measure. For R2, we analyzed treatment and sex effects on these
variables. There were no main effects or interactions involving
the factor ‘sex of piglet’ and so sex differences were not
considered further.
In Experiments 2 and 3 we determined treatment effects on
vocalizations, line crossing, and jumping during isolations, and
vocalizations and time spent near the sow’s face during reunions,
with test session (Iso1 vs. Iso2 or R1 vs. R2) as the repeated
measure.
In Experiment 4, we determined treatment effects on
vocalizations, line crossings, and jumping during isolations,
with test session (Iso1 vs. Iso2) as the repeated measure. Because
EHP piglets were isolated during R1, their behavior was not
compared with that of LHP piglets in R1, or their own behavior in
R2. We compared piglet vocalizations by LHP piglets during R1
(sow absent) versus R2, and vocalizations and time spent near the
sow’s face by EHP versus LHP piglets in R2, using paired t-tests.
RESULTS
Experiment 1—With Mother and Littermates inHome Pen (HP) Versus Empty Pen (EP)
Isolation Sessions. A brief interaction with the mother
and littermates during R1 affected the number of calls
emitted during Iso2 (Treatment: F(1, 7)¼ 6.1, p¼ .04).
The number of calls was significantly lower in Iso2 than
Iso1 in the EP group; by contrast, the HP group showed no
change in rate of calling (Session: F(1, 7)¼ 11.7,
p¼ .011; Treatment� Session: F(1, 7)¼ 19.5, p¼ .003;
post-hoc comparison EP-Iso1 versus EP-Iso2, p< .01;
Fig. 1A).
There was a treatment� session effect on jumping
(F(1, 7)¼ 5.8, p¼ .047). Piglets in the HP group jumped
more than piglets in the EP group during Iso2 (p< .05),
whereas there was no difference between treatments in
Iso1 (Fig. 1B). For the HP group, the post-hoc comparison
between Iso1 and Iso2 was p¼ .051. There was no
treatment effect on line crossings in the isolation sessions
(p> .05). Fewer line crossings (n/7 min) occurred in Iso2
(40.9� 6.2) than Iso1 (86.2� 6.3; Session: F(1,
7)¼ 102.9, p< .001) but this decrease was not treatment
specific (Treatment� Session: p> .05).
Piglet body temperature was stable from Iso1
(32.7� .15�C) to Iso2 (33.5� .26�C) in both treatment
groups (main effects and interaction, p> .05).
Reunion Sessions. In the HP group, the rate of calling
(R1: 7.9� 1.0; R2: 6.5� 1.4 calls/min) and time spent
in proximity to the sow’s face (R1: 13.1� 1.1 s; R2:
10.6� .9 s) did not differ across reunions sessions
Developmental Psychobiology Isolation and Reunion Responses of Piglets 5
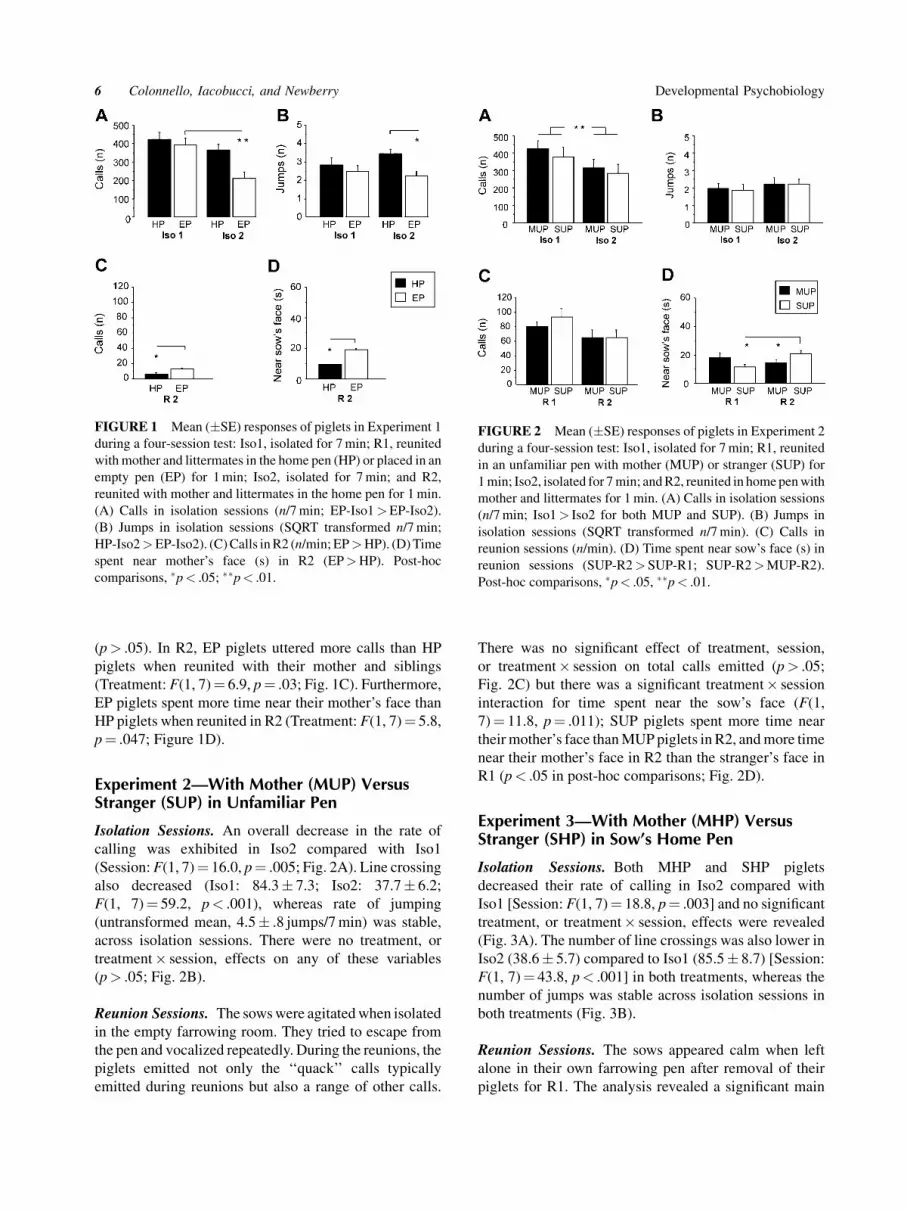
(p> .05). In R2, EP piglets uttered more calls than HP
piglets when reunited with their mother and siblings
(Treatment: F(1, 7)¼ 6.9, p¼ .03; Fig. 1C). Furthermore,
EP piglets spent more time near their mother’s face than
HP piglets when reunited in R2 (Treatment: F(1, 7)¼ 5.8,
p¼ .047; Figure 1D).
Experiment 2—With Mother (MUP) VersusStranger (SUP) in Unfamiliar Pen
Isolation Sessions. An overall decrease in the rate of
calling was exhibited in Iso2 compared with Iso1
(Session: F(1, 7)¼ 16.0, p¼ .005; Fig. 2A). Line crossing
also decreased (Iso1: 84.3� 7.3; Iso2: 37.7� 6.2;
F(1, 7)¼ 59.2, p< .001), whereas rate of jumping
(untransformed mean, 4.5� .8 jumps/7 min) was stable,
across isolation sessions. There were no treatment, or
treatment� session, effects on any of these variables
(p> .05; Fig. 2B).
Reunion Sessions. The sows were agitated when isolated
in the empty farrowing room. They tried to escape from
the pen and vocalized repeatedly. During the reunions, the
piglets emitted not only the ‘‘quack’’ calls typically
emitted during reunions but also a range of other calls.
There was no significant effect of treatment, session,
or treatment� session on total calls emitted (p> .05;
Fig. 2C) but there was a significant treatment� session
interaction for time spent near the sow’s face (F(1,
7)¼ 11.8, p¼ .011); SUP piglets spent more time near
their mother’s face than MUP piglets in R2, and more time
near their mother’s face in R2 than the stranger’s face in
R1 (p< .05 in post-hoc comparisons; Fig. 2D).
Experiment 3—With Mother (MHP) VersusStranger (SHP) in Sow’s Home Pen
Isolation Sessions. Both MHP and SHP piglets
decreased their rate of calling in Iso2 compared with
Iso1 [Session: F(1, 7)¼ 18.8, p¼ .003] and no significant
treatment, or treatment� session, effects were revealed
(Fig. 3A). The number of line crossings was also lower in
Iso2 (38.6� 5.7) compared to Iso1 (85.5� 8.7) [Session:
F(1, 7)¼ 43.8, p< .001] in both treatments, whereas the
number of jumps was stable across isolation sessions in
both treatments (Fig. 3B).
Reunion Sessions. The sows appeared calm when left
alone in their own farrowing pen after removal of their
piglets for R1. The analysis revealed a significant main
Developmental Psychobiology
FIGURE 1 Mean (�SE) responses of piglets in Experiment 1
during a four-session test: Iso1, isolated for 7 min; R1, reunited
with mother and littermates in the home pen (HP) or placed in an
empty pen (EP) for 1 min; Iso2, isolated for 7 min; and R2,
reunited with mother and littermates in the home pen for 1 min.
(A) Calls in isolation sessions (n/7 min; EP-Iso1>EP-Iso2).
(B) Jumps in isolation sessions (SQRT transformed n/7 min;
HP-Iso2>EP-Iso2). (C) Calls in R2 (n/min; EP>HP). (D) Time
spent near mother’s face (s) in R2 (EP>HP). Post-hoc
comparisons, �p< .05; ��p< .01.
FIGURE 2 Mean (�SE) responses of piglets in Experiment 2
during a four-session test: Iso1, isolated for 7 min; R1, reunited
in an unfamiliar pen with mother (MUP) or stranger (SUP) for
1 min; Iso2, isolated for 7 min; and R2, reunited in home pen with
mother and littermates for 1 min. (A) Calls in isolation sessions
(n/7 min; Iso1> Iso2 for both MUP and SUP). (B) Jumps in
isolation sessions (SQRT transformed n/7 min). (C) Calls in
reunion sessions (n/min). (D) Time spent near sow’s face (s) in
reunion sessions (SUP-R2> SUP-R1; SUP-R2>MUP-R2).
Post-hoc comparisons, �p< .05, ��p< .01.
6 Colonnello, Iacobucci, and Newberry

effect of session on calling [F(1, 7)¼ 14.9, p¼ .006], with
both MHP and SHP piglets calling less in R2 than R1.
Treatment, and treatment� session, effects on calling
were nonsignificant (Fig. 3C). However, SHP piglets
spent more time near their mother’s face than MHP piglets
in R2, and more time near their mother’s face in R2
than the stranger’s face in R1 [Treatment� Session:
[F(1, 7)¼ 8.2, p¼ .02; p< .01 in post-hoc comparisons;
Fig. 3D].
Experiment 4—With Littermates in Home Pen(LHP) Versus Empty Home Pen (EHP)
Isolation Sessions. Piglets in both LHP and EHP groups
called less in Iso2 compared with Iso1 [Session: F(1,
7)¼ 29.2, p¼ .001; Fig. 4A], and crossed fewer lines
[Iso1: 85.1� 6.2; Iso2: 34.9� 5.1; Session: F(1,
7)¼ 55.2, p< .001], but no significant treatment, or
treatment� session, effects were revealed. The rate of
jumping was stable across isolation sessions in both
treatments (p> .05; Fig. 4B).
Reunion Sessions. LHP piglets emitted more calls
during reunion with their mother and littermates in
R2 (10.1� 2.9 calls/min) than during reunion with
littermates alone in R1 (3.9� 1.2; Session: T(7)¼�2.81, p¼ .026). They also called less in R1 than piglets
in an empty home pen in R1 (50.1� 11.4). No differences
in rate of calling (Fig. 4C), or time spent near the mother’s
face (Fig. 4D), were detected between the LHP and EHP
piglets during the second reunion (p> .05).
DISCUSSION
Responses to Isolation
Based on previous findings in rats (Shair, 2007), we
predicted that a brief reunion with the mother
and littermates in the home pen during R1 would result
in a higher rate of calling during Iso2 than Iso1 whereas,
in the absence of a reunion during R1, calling rate
would be lower in Iso2 than Iso1. We also hypothesized
that potentiation of calling would be specific to
the mother rather than being elicited by other social
stimuli such as littermates and unfamiliar sows. Thus, we
predicted that reunion with the mother would potentiate
calling whereas interaction with an unfamiliar sow
would produce a decline in call rate during the second
isolation.
Developmental Psychobiology
FIGURE 3 Mean (�SE) responses of piglets in Experiment 3
during a four-session test: Iso1, isolated for 7 min; R1, reunited
in sow’s home pen with mother (MHP) or stranger (SHP) for
1 min; Iso2, isolated for 7 min; and R2, reunited in home pen with
mother and littermates for 1 min. (A) Calls in isolation sessions
(n/7 min; Iso1> Iso2 for both MHP and SHP). (B) Jumps in
isolation sessions (SQRT transformed n/7 min). (C) Calls in
reunion sessions (n/min; R1>R2 for both MHP and SHP).
(D) Time spent near sow’s face (s) in reunion sessions
(SHP-R2> SHP-R1; SHP-R2>MHP-R2). Post-hoc compari-
sons, �p< .05, ��p< .01.
FIGURE 4 Mean (�SE) responses of piglets in Experiment 4
during a four-session test: Iso1, isolated for 7 min; R1, reunited
in home pen with littermates (LHP) or alone (EHP) for 1 min;
Iso2, isolated for 7 min; and R2, reunited in home pen with
mother and littermates for 1 min. (A) Calls in isolation sessions
(n/7 min; Iso1> Iso2 for both LHP and EHP). (B) Jumps in
isolation sessions (SQRT transformed n/7 min). (C) Calls in R2
(n/min). (D) Time spent near mother’s face (s) in R2. Post-hoc
comparisons, �p< .05, ��p< .01.
Isolation and Reunion Responses of Piglets 7

Contrary to our predictions, piglets did not show the
classic form of potentiation described in rats (elevated
calling in Iso2 relative to Iso1) in any of the four
experiments. We expected, and found, less calling and line
crossing in Iso2 than Iso1 in the absence of a reunion with
the mother in R1, including when placed in an empty
unfamiliar pen, alone in the home pen, with a stranger sow,
or with littermates in the absence of the mother. However,
unexpectedly, we found that less calling and line crossing
also occurred in Iso2 than Iso1 when piglets were reunited
specifically with their mother in R1. In Experiment 2, we
thought that this weakened response in Iso2, antithetical to
maternal potentiation, could have been due to R1 taking
place in an unfamiliar location. The sows appeared
agitated and the piglets displayed a high rate of calling
under this condition, suggesting that the R1 reunion was
not comforting. However, the results of Experiment 3
show that, even when the R1 reunion occurred with an
apparently calm mother in the home pen, fewer calls and
line crossings occurred in Iso2 than Iso1. Therefore, we
were not able to detect a maternal potentiation effect
specific to reunion with the mother, contrary to our
prediction based on the rat literature.
Despite the absence of maternal potentiation, when
piglets were reunited with their whole family (mother and
littermates) in their familiar home pen in R1, they
exhibited a persistence, rather than a drop, in calling from
Iso1 to Iso2, unlike their response in any other condition
that we tested. This finding suggests that reunion with
a calm mother and littermates in the home pen was
maximally reassuring, thereby stimulating, or perhaps
physically enabling (due to relative preservation of
energetic reserves in R1), the relatively strong reinstate-
ment behavior in Iso2. This outcome makes sense from an
evolutionary perspective because it would presumably be
adaptive to call in an attempt to reinstate contact when
conditions favor a safe reunion, and to be quieter when
sows have recently been signaling distress, as in the MUP
and SUP conditions, or when a potentially dangerous
change in the environment has recently been detected
(e.g., pen or room is different, stranger is present, or
mother and/or littermates are missing, in R1).
Hennessy et al. (2006) observed a similar persistence
rather than potentiation of calling in Iso2 in guinea pigs,
which they referred to as a relative potentiation, but the
response was specific to the mother. Although the piglets
exhibited maternal recognition (see below), it appears that
they did not discriminate the mother alone as the primary
source of comfort under the conditions of this study,
possibly because the piglets were confined in farrowing
pens and littermates were always present along with the
mother. If so, one might expect a similar lack of maternal
specificity in laboratory rodents, which are confined in
cages. At least under confined housing conditions, the
difference between piglet and rodent responses may relate
to the fact that rodent maternal care is more individu-
alized; unlike rodent mothers, sows do not lick individual
piglets or retrieve them in their mouth. Nevertheless,
housing sows and litters in get-away pens that allow sows
to leave their piglets voluntarily for brief periods (Pajor,
Kramer, & Fraser, 2000), or under free-range conditions,
would allow piglets to develop a more individualized
response to isolation and reunion from their mother.
Observations of pigs under free-range conditions suggest
that it is the mother who is the most important determinant
of the movements and social groupings of unweaned
piglets (Newberry & Wood-Gush, 1986). Although 15-
day-old piglets also direct affiliative behavior towards
their littermates and spend more time in close proximity
with their littermates than with their mother, they do not
exhibit strong preferences for specific littermates at this
age whereas they clearly have a selective attachment to
their mother (Newberry & Wood-Gush, 1986).
In contrast to the strong potentiation of calling in rats,
the lesser persistence response exhibited by piglets and
guinea pigs during a second isolation may be related to
their precociality, making them better equipped to cope
with short separations from their mother and littermates in
an unfamiliar location than relatively helpless rat pups. By
15 days of age, piglets are capable of navigating among
different nest sites as well as exploring new areas and
playing out of visual, and sometimes auditory, contact
with their mother (Donaldson, Newberry, Spinka, &
Cloutier, 2002; Newberry & Wood-Gush, 1985, 1986;
Newberry, Wood-Gush, & Hall, 1988). Under commercial
swine production conditions, they are also able to survive
weaning at this age.
In altricial laboratory mice subjected to the repeated
isolation paradigm (Iacobucci et al., 2005; Scattoni,
Crawley, & Ricceri, 2009), the extent of calling given
by infants in a second isolation can vary according to
genetic strain, age, and methodological details. Therefore,
we cannot rule out the possibility that pigs might exhibit a
potentiation of calling, equivalent to that observed in rats,
under some circumstances.
We took multiple steps to ensure that our piglets were
not hungry or cold during testing, thereby minimizing
the piglets’ immediate need for resources to maintain
physiological homeostasis, a factor known to influence
calling rate in piglets (Weary & Fraser, 1995; Weary et al.,
1996, 1997). The persistence of calling in Experiment 1,
in the absence of pain or urgent physiological needs,
points to the importance of psychological comfort derived
from contact with the mother and littermates in the safety
of the nest (home pen) in eliciting a relatively strong
response in Iso2. Although we found no treatment-
specific potentiation of line crossing in the observation
arena from Iso1 to Iso2 in any experiment, in agreement
Developmental Psychobiology8 Colonnello, Iacobucci, and Newberry

with findings in rats (Shair, 2007), we did observe a
persistence (and almost a potentiation, p¼ .051) of
jumping in Iso2 of Experiment 1 following reunion with
the mother and littermates in the home pen. The brief
reunion may have stimulated renewed attempts to escape
from the observation arena in anticipation of reinstating
contact with the family. An active response by piglets can
be explained by their precocial nature, unlike that of
altricial, relatively immobile rat pups whose calls elicit a
maternal retrieval response. Likewise, exposure to the
mother prevented a drop in motor activity during a second
isolation in another precocial species, the guinea pig
(Hennessy et al., 2006). However, in piglets, jumping
responses were persistent across both isolation sessions
regardless of treatment. In contrast, line crossings
probably dropped in Iso2 because the relatively small
arena had already been explored in Iso1 and, now, activity
was focused on escape from the arena.
It remains unclear whether the degree of potentiation
following re-isolation accurately depicts the strength of
filial attachment, given that both piglets and guinea pigs
show a relative rather than absolute potentiation of calling
despite having a more selective filial attachment bond than
rats.
Responses to Reunion
Although piglets called and moved around the arena less
in Iso2 than Iso1 regardless of whether they had just met
their mother alone, or another adult female, in R1, these
results were not due to failure to recognize their mother.
Piglets spent more time near their mother’s face during R2
than a stranger’s face during R1, and more time near their
mother’s face in R2 if they had just met another sow rather
than their own mother in R1, regardless of whether R1
occurred in a relatively familiar or unfamiliar location.
These results confirm that the piglets were able to
distinguish between their mother and the unfamiliar
sow, as reported in other studies of maternal recognition
by piglets (Illmann et al., 2002; Morrow-Tesch &
McGlone, 1990; Parfet & Gonyou, 1991; Puppe et al.,
1999).
We predicted that piglets reunited with their mother in
the presence or absence of their littermates during R1
would show a stronger vocal response during R2 than
piglets placed in an empty pen or exposed to an unfamiliar
sow during R1. In contrast to our prediction, the rate of
calls emitted by piglets during R2 was greater for EP than
HP piglets in Experiment 1 and comparable across the
pairs of treatments evaluated in the other experiments.
Looking at all treatments together, it appears that there
was a numerical tendency for piglets to call more in R2 if
they had encountered an unfamiliar pen or sow in R1 (EP,
MUP, SUP, SHP) than if they had been placed in their own
home pen in R1 (HP, MHP, LHP, EHP). Modulation of
reunion calling as a function of exposure to novel
conditions indicates that these calls were not emitted in
a fixed, predetermined pattern but varied according to the
context of the social reunion. Time spent close to the
mother’s face in R2 was significantly higher for piglets
that had been placed in a novel pen or with another sow
(EP, SUP, SHP) rather than with their own mother (HP,
MUP, MHP) in R1, suggesting that piglets that experi-
enced a longer separation from their mother, especially in
a novel location, were more motivated to seek proximity
to her face.
The rate of calling during the reunions was strongly
affected by the quality of the interaction with the sow. In
Experiment 1, the mean call rate during the 1-min
reunions (for R1 of the HP group and R2 of both the HP
and EP groups) was 14.3� .8 calls/min whereas the mean
rate of calling during the 7-min isolations (for both the HP
and EP groups across both isolation sessions) was
49.6� 2.9 calls/min. Although these numbers are not
directly comparable because the reunion sessions were
shorter than the isolation sessions, piglets usually vocalize
only briefly during reunions and so prolonging the reunion
sessions would have resulted in a lower, rather than higher,
mean calling rate/min. Our data are, thus, indicative of a
dramatic ‘‘contact quieting’’ effect during the reunions in
Experiment 1, as previously described in rats (Hofer &
Shair, 1978; Shair, 2007). ‘‘Contact quieting’’ was also
evident in Experiment 4, when the presence of littermates
in R1 lowered calling compared to Iso1 or exposure to the
empty home pen. By contrast, in Experiment 2, the mean
rate of calling during reunions (for both MUP and SUP
groups in R1 and R2) was 75� 5.4 calls/min, comprising
a variety of vocalizations and not just the ‘‘quack’’ calls
typical of face-to-face interaction with the sow. This value
was even higher than the mean rate of calling during
isolations in Experiment 2 (50.1� 3.7 calls/min, for both
MUP and SUP groups across Iso1 and Iso2), supporting
the interpretation that the reunions were less comforting in
the presence of a sow alone. A high rate of calling in R1
also occurred when piglets met a relatively calm sow
alone in her home pen in Experiment 3, suggesting that it
was the absence of littermates that prompted the elevated
levels of calling. Calling was reduced following reunion
with the whole family in R2 in Experiment 3 but not in
Experiment 2, possibly due to greater disturbance from
the R1 reunion with an agitated sow in an unfamiliar room.
The absence of sex differences in responses to reunion
in Experiment 1 is interesting given that, theoretically, one
would expect that, under good conditions, mothers of
polygynous species should show greater maternal invest-
ment in sons than daughters (Trivers & Willard, 1973).
The high prolificacy of sows and rapid growth of piglets
under intensive farm management (as in the present study)
Developmental Psychobiology Isolation and Reunion Responses of Piglets 9

could be interpreted as conditions favoring male-biased
investment. One mechanism by which male offspring
could command greater maternal investment than females
would be to perform more calling (a form of ‘‘begging’’
behavior) than females when reunited with their mother.
However, this was not observed, probably because piglets
suckle as a litter rather than individually and differences in
milk provisioning by the sow to individual piglets within a
litter are primarily determined by direct competition
among piglets for high-yielding teats and, following
establishment of the teat order, by the amount of teat
massage performed by each piglet towards ‘‘its’’ teat
(Dostaikova, Spinka & Kindlmann, 2002).
CONCLUSIONS
The combined gestalt of the mother and littermates in their
familiar location induced a optomistic persistence of
reinstatement responses (protest) following re-isolation,
whereas any unfamiliar conditions encountered during the
first reunion provoked despair-like responses when
piglets were re-isolated. We refer to despair rather than
habituation to the isolation because, especially for EP and
SUP piglets, responses in R2 were suggestive of having
missed the mother and siblings. Findings on proximity-
seeking towards the mother, expressed by calls and
physical proximity, show the value of adding the second
reunion to the classic isolation–reunion–isolation proce-
dure.
Our results, in combination with those of Hennessy
et al. (2006) in guinea pigs, suggest that the potentiation
paradigm is extendable to precocial species but these
species may be more likely to exhibit persistence rather
than the potentiation observed in rats. The observation
that piglets exhibited emotional responses to separation
from, and reunion with, both their mother and littermates
indicates the importance of careful evaluation of
responses in relation to the biology of individual species.
We have demonstrated that the domestic pig can provide
useful insights into aspects of attachment, such as
production of ‘‘greeting’’ calls and seeking of maternal
proximity upon reunion, that cannot be addressed in rat
models. Our data extend understanding of filial attach-
ment bonding in the domestic pig, and reveal similarities
between piglet and human infant responses to both
isolation and reunion.
NOTES
V.C. was supported by a scholarship from the National Council
of the University of Rome ‘‘Sapienza,’’ Italy. The authors are
grateful to Professor Gianni Pavan (University of Pavia, IT) for
providing access to the SeaPro Sound Emission Analyzer
software for recording and analysis of audible and ultrasonic
vocalizations, Winnie Chan, Nicole Dossey, Joe Gallegos,
Chesica Jones, Satomi Kaneko, Jacob Larson, Brooke Reed,
Rachel Reynolds, and Kaori Tokuhisa for help with data
collection, Asa Timm and the staff of the WSU Swine Center
for animal care, and two anonymous reviewers for helpful
comments on the manuscript.
REFERENCES
Ainsworth, M. D. S. (1989). Attachment beyond infancy.
American Psychologist, 44, 709–716.
Ainsworth, M. D. S., Blehar, M. C., Waters, E., & Wall, S.
(1978). Patterns of attachment: A psychological study of the
strange situation. Hillsdale, NJ: Lawrence Erlbaum Asso-
ciates.
Algers, B. (1993). Nursing in pigs: Communicating needs and
distributing resources. Journal of Animal Science, 71, 2826–
2831.
Appleby, M. C., Weary, D. M., Taylor, A. A., & Illmann, G.
(1999). Vocal communication in pigs: Who are nursing
piglets screaming at? Ethology, 105, 881–892.
Bowlby, J. (1973). Separation: Anxiety & anger. London:
Hogarth Press.
Brunelli, S. A., & Hofer, M. A. (2007). Selective breeding for
infant rat separation-induced ultrasonic vocalizations: Devel-
opmental precursors of passive and active coping styles.
Behavioural Brain Research, 182, 193–207.
Cassidy, J. (1999). The nature of the child’s ties. In J. Cassidy,
& P. R. Shaver (Eds.), Handbook of attachment: Theory,
research, and clinical applications (pp. 3–20). New York:
Guilford Press.
Donaldson, T. M., Newberry, R. C., Spinka, M., & Cloutier, S.
(2002). Effects of early play experience on play behaviour of
piglets after weaning. Applied Animal Behaviour Science,
79, 221–231.
Dostaikova, I., Spinka, M., & Kindlmann, P. (2002). Begging
for milk: Evolution of teat massaging in suckling pigs.
Journal of Theoretical Biology, 215, 321–332.
Fraser, D. (1975). Vocalizations of isolated piglets. I. Sources of
variation and relationships among measures. Applied Animal
Ethology, 1, 387–394.
Gonyou, H. W. (2001). The social behaviour of pigs. In L. J.
Keeling, & H. W. Gonyou (Eds.), Social behaviour in farm
animals (pp. 147–176). Wallingford, UK: CAB Interna-
tional.
Graves, H. B. (1984). Behavior and ecology of wild and feral
swine (Sus scrofa). Journal of Animal Science, 58, 482–492.
Hennessy, M. B., Miller, E. E., & Shair, H. N. (2006). Brief
exposure to the biological mother ‘‘potentiates’’ the isolation
behavior of precocial Guinea pig pups. Developmental
Psychobiology, 48, 653–659.
Herman, B. H., & Panksepp, J. (1978). Effects of morphine and
naloxone on separation distress and approach attachment:
Evidence for opiate mediation of social affect. Pharmaco-
logy, Biochemistry, and Behavior, 9, 213–220.
Developmental Psychobiology10 Colonnello, Iacobucci, and Newberry

Hofer, M. A., Brunelli, S. A., Masmela, J., & Shair, H. N.
(1996). Maternal interactions prior to separation potentiate
isolation-induced calling in rat pups. Behavioral Neuro-
science, 110, 1158–1167.
Hofer, M. A., Brunelli, S. A., & Shair, H. N. (1993). Ultrasonic
vocalization responses of rat pups to acute separation and
contact comfort do not depend on maternal thermal cues.
Developmental Psychobiology, 26, 81–95.
Hofer, M. A., Brunelli, S. A., & Shair, H. N. (1994).
Potentiation of isolation-induced vocalization by brief
exposure of rat pups to maternal cues. Developmental
Psychobiology, 27, 503–517.
Hofer, M. A., Masmela, J., Brunelli, S. A., & Shair, H. N.
(1998). The ontogeny of maternal potentiation of the infant
rats’ isolation call. Developmental Psychobiology, 33, 189–
201.
Hofer, M. A., & Shair, H. (1978). Ultrasonic vocalization
during social interaction and isolation in 2-weeek-old rats.
Developmental Psychobiology, 11, 495–504.
Iacobucci, P., Branchi, I., D’Adamo, P., Toniolo, D., Alleva, E.,
& Ricceri, L. (2005). Early cognitive impairment in
Gdi1 deficient mice, a mouse model of X-linked non-
specific mental retardation. Department of Cell Biology and
Neurosciences meeting, June 14–16, 2005, Italian National
Institute of Health, Rome IT.
Iacobucci, P., Colonnello, V., Pavan, G., & Ricceri, L. (2007). A
qualitative analysis of ultrasound vocalizations emitted by
12-day-old rats during homing test. In: Proc. 40th Annual
Meeting of the International Society for Developmental
Psychobiology, October 31–November 3, 2007, San Diego,
CA.
Illmann, G., Schrader, L., Spinka, M., & Sustr, P. (2002).
Acoustical mother-offspring recognition in pigs (Sus scrofa
domesticus). Behaviour, 139, 487–505.
Jensen, P., Gustafsson, M., & Augustsson, H. (1998). Teat
massage after milk ingestion in domestic piglets: An
example of honest begging? Animal Behaviour, 55, 779–
786.
Maletinska, M., Spinka, M., Vichova, J., & Stehulova, I. (2002).
Individual recognition of piglets by sows in the early post-
partum period. Behaviour, 139, 975–991.
Moles, A., Kieffer, B. L., & D’Amato, F. R. (2004). Deficit in
attachment behavior in mice lacking the mu-opioid receptor
gene. Science, 304, 1983–1986.
Morrow-Tesch, J., & McGlone, J. J. (1990). Sources of
maternal odors and the development of odor preferences
in baby pigs. Journal of Animal Science, 68, 3563–3571.
Muller, J. M., Brunelli, S. A., Moore, H., Myers, M. M., &
Shair, H. N. (2005). Maternally modulated infant separation
responses are regulated by D2-family dopamine receptors.
Behavioral Neuroscience, 119, 1384–1388.Myers, M. M., Ali, N., Weller, A., Brunelli, S. A., Tu, A. Y.,
Hofer, M. A., et al. (2004). Brief maternal interaction
increases number, amplitude, and bout size of isolation-induced
ultrasonic vocalizations in infant rats (Rattus norvegicus).
Journal of Comparative Psychology, 118, 95–102.Newberry, R. C., & Swanson, J. C. (2008). Implications
of breaking mother-young social bonds. Applied Animal
Behaviour Science, 110, 3–23.
Newberry, R. C., & Wood-Gush, D. G. M. (1985). The suckling
behaviour of domestic pigs in a semi-natural environment.
Behaviour, 95, 11–25.
Newberry, R. C., & Wood-Gush, D. G. M. (1986). Social
relationships of piglets in a seminatural environment. Animal
Behaviour, 34, 1311–1318.Newberry, R. C., Wood-Gush, D. G. M., & Hall, J. W. (1988).
Playful behaviour of piglets. Behavioural Processes, 17,
205–216.Pajor, E. A., Kramer, D. L., & Fraser, D. (2000). Regulation
of contact with offspring by domestic sows: Temporal
patterns and individual variation. Ethology, 106, 37–51.
Panksepp, J. (2003). Can anthropomorphic analyses of
separation cries in other animals inform us about the
emotional nature of social loss in humans? Comment on
Blumberg and Sokoloff (2001). Psychological Review, 110,
376–388.
Panksepp, J., Nelson, E., & Bekkedal, M. (1997). Brain
systems for the mediation of social separation-distress and
social-reward. Evolutionary antecedents and neuropeptide
intermediaries. Annals of the New York Academy of
Sciences, 807, 78–100.
Parfet, K. A., & Gonyou, H. W. (1991). Attraction of newborn
piglets to auditory, visual, olfactory and tactile stimuli.
Journal of Animal Science, 69, 125–133.
Pettijohn, T. F. (1979). Attachment and separation distress in
the infant guinea pig. Developmental Psychobiology, 12,
73–81.Polan, H. J., & Hofer, M. A. (1999). Psychobiological origins of
infant attachment and separation responses. In J. Cassidy, &
P. R. Shaver (Eds.), Handbook of attachment: Theory,
research, and clinical applications (pp. 162–180). New York:
Guilford Press.
Puppe, B., Schon, P. C., & Wendland, K. (1999). Monitoring of
piglets’ open field activity and choice behaviour during the
replay of maternal vocalization: A comparison between
Observer and PID technique. Laboratory Animals, 33, 215–220.
Scattoni, M. L., Crawley, J., & Ricceri, L. (2009). Ultrasonic
vocalizations: A tool for behavioural phenotyping of mouse
models of neurodevelopmental disorders. Neuroscience and
Biobehavioral Reviews, 33, 508–515.
Schrader, L., & Todt, D. (1998). Vocal quality is correlated with
levels of stress hormones in domestic pigs. Ethology, 104,
859–876.
Schon, P. C., Puppe, B., Gromyko, T., & Manteuffel, G. (1999).
Common features and individual differences in nurse
grunting of domestic pigs (Sus scrofa): A multi-parametric
analysis. Behaviour, 136, 49–66.
Shair, H. N. (2007). Acquisition and expression of a socially
mediated separation response. Behavioural Brain Research,
182, 180–192.
Shair, H. N., Masmela, J. R., Brunelli, S. A., & Hofer,
M. A. (1997). Potentiation and inhibition of ultrasonic
vocalization of rat pups: Regulation by social cues.
Developmental Psychobiology, 30, 195–200.
Spinka, M., Stehulova, I., Zacharova, J., Maletinska, J., & Illmann,
G. (2002). Nursing behaviour and nursing vocalisations in
domestic sows: Repeatability and relationship with maternal
investment. Behaviour, 139, 1077–1097.
Developmental Psychobiology Isolation and Reunion Responses of Piglets 11

Trivers, R. L., & Willard, D. E. (1973). Natural selection of
parental ability to vary the sex ratio of offspring. Science,
179, 90–92.
Weary, D. M., Appleby, M. C., & Fraser, D. (1999).
Responses of piglets to early separation from the
sow. Applied Animal Behaviour Science, 63, 289–300.
Weary, D. M., & Fraser, D. (1995). Calling by domestic piglets:
Reliable signals of need? Animal Behaviour, 50, 1047–1055.
Weary, D. M., Lawson, L. G., & Thompson, B. K. (1996). Sows
show stronger responses to isolation calls of piglets
associated with greater levels of piglet need. Animal
Behaviour, 52, 1247–1253.
Weary, D. M., Ross, S. K., & Fraser, D. (1997). Vocalizations by
isolated piglets: A reliable indicator of piglet need directed
towards the sow. Applied Animal Behaviour Science, 53,
249–257.
Developmental Psychobiology12 Colonnello, Iacobucci, and Newberry





























