Embed Size (px)
Citation preview

Water clarity in Lake Mendota since 1900:
responses to differing levels of nutrients
and herbivory
Richard C. Lathrop, Stephen R. Carpenter, and Lars G. Rudstam
Abstract: Water transparency, phosphorus concentrations, and herbivory in eutrophic Lake Mendota have been highlyvariable this century. To assess the effects of both nutrients and herbivory on water clarity, mean Secchi disk readings werecomputed for six seasonal periods for each year during 1900–1993 that had readings (71 years but not in all periods). Weassigned each year one of three levels of nutrients and one of two levels of herbivory on the basis of data independent ofSecchi readings to avoid circularity in our analyses. For each seasonal period, we tested for differences in Secchi readingsattributable to nutrients, herbivory, and their interaction. Our results indicate that in years with high herbivory, water claritywas significantly greater during all periods including summer when blue-green algal blooms frequently occurred. Highernutrient levels only led to decreased water clarity during the summer and destratification periods; water clarity was notaffected by nutrient levels during other periods. Our analysis of Lake Mendota’s long-term Secchi disk record indicates thatherbivory can strongly influence water clarity in large eutrophic lakes, and that the greatest improvements in summer waterclarity in those lakes may be achieved by combining phosphorus reduction programs with biomanipulation techniques toincrease herbivory.
Résumé: La limpidité de l’eau, la concentration du phosphore et la phytophagie ont considérablement varié dans lelac Mendota, un lac eutrophe, au cours de ce siècle. Afin d’évaluer les effets de la concentration des nutriants et de laphytophagie sur la limpidité de l’eau, nous avons calculé les moyennes de lectures faites au disque de Secchi en fonction desix périodes saisonnières pour toutes les années entre 1900 et 1993 où ces renseignements existent (soit pour 71 ans, mais paspour toutes les périodes). Nous avons attribué à chaque année un niveau de nutriants sur trois possibles et un niveau dephytophagie sur deux possibles en nous appuyant sur des données indépendantes des lectures au disque de Secchi afin d’éviterla création de cercles vicieux dans nos analyses. Pour chaque période saisonnière, nous avons cherché des différences dans leslectures au disque de Secchi qui soient attribuables aux nutriants, à la phytophagie et à leurs interactions. Nos résultatsrévèlent que, les années d’intense phytophagie, l’eau était significativement plus limpide à toutes les périodes, y compris l’été,alors qu’il s’est souvent produit des proliférations d’algues bleues. À des concentrations supérieures de nutriants, le seul effetobservé était une perte de limpidité durant l’été et pendant les périodes de déstratification. La limpidité de l’eau n’a pas étémodifiée par la teneur en nutriants durant les autres périodes. Notre analyse des lectures à long terme au disque de Secchi de lalimpidité de l’eau du lac Mendota indique que la phytophagie peut fortement influer sur la limpidité de l’eau des grands lacseutrophes et que les plus grandes améliorations sur ce plan au cours de la période estivale, dans ces lacs, peut être obtenue encombinant des programmes pour réduire les apports de phosphore à des techniques de biomanipulation visant à favoriser laphytophagie.[Traduit par la Rédaction]
Introduction
The role of nutrients versus zooplankton grazing in controllingphytoplankton blooms in eutrophic lakes is controversial.Many studies show that changes in fish communities cascade
through the food web to change phytoplankton biomass(Hrbáeek et al. 1961; Brooks and Dodson 1965; Shapiro andWright 1984; Gulati et al. 1990; Reinertsen et al. 1990; Perssonet al. 1992; Findlay et al. 1994; Carpenter et al. 1996). How-ever, successful biomanipulations often involve lakes only afew hectares in area for only a few years (Shapiro 1990;Benndorf 1990; Reynolds 1994). Moreover, biomanipulationmay not work in eutrophic lakes where high nutrient suppliesfavor dense growths of blue-green algae that are not readilygrazed by zooplankton (McQueen et al. 1986; Benndorf 1987,1990; Elser and Goldman 1991; Reynolds 1994). Thus, thereis uncertainty about the capacity of grazing to affect phyto-plankton biomass and water clarity in larger, more eutrophiclakes over many years.
Can grazing improve water clarity in a large eutrophic lakefor periods of years to decades, at varying levels of nutrients?The long-term limnological record of Lake Mendota (Wiscon-sin, U.S.A.) allows us to test this question. For this large strati-fied lake (3985 ha, 25.3 m maximum depth), Secchi disk
Received September 27, 1995. Accepted March 29, 1996.J13089
R.C. Lathrop.1 Bureau of Research, Wisconsin Department ofNatural Resources, 1350 Femrite Drive, Monona, WI 53716,U.S.A., and Center for Limnology, 680 North Park Street,University of Wisconsin, Madison, WI 53706, U.S.A.S.R. Carpenter and L.G. Rudstam.2 Center for Limnology,680 North Park Street, University of Wisconsin, Madison, WI53706, U.S.A.
1 Author to whom all correspondence should be addressed.e-mail: [email protected]
2 Present address: Cornell Biological Field Station, 900Shackelton Point Road, Bridgeport, NY 13030, U.S.A.
Can. J. Fish. Aquat. Sci. 53: 2250–2261 (1996).
2250
© 1996 NRC Canada

readings exist for many years since 1900. Extensive historicalinformation on lake phosphorus (P) concentrations andzooplankton and fish populations allows nutrient and her-bivory levels to be assigned for different time periods. Ongo-ing research has elucidated important food web interactionsamong the lake’s key piscivores, planktivores, and herbivores(Kitchell 1992). Lake Mendota, which has a history of algalblooms since at least the late 1800s (Brock 1985; Lathrop andCarpenter 1992a), has experienced a major increase in P load-ing rates during this century (Lathrop 1990, 1992). Becauseplanktivore populations have also varied greatly in Lake Men-dota, episodes of low and high herbivory have occurred duringperiods of both low and high P loadings.
Long-term studies like those done on Lake Mendota havebeen increasingly recognized for their contribution to the un-derstanding of ecosystem function (Likens 1983, 1989; Schin-dler 1987, 1988, Edmondson 1991). Long time series canidentify ecological processes that are masked in shorter studiesbecause of the long lags in certain processes (Magnuson 1990;Schindler 1990; Edmondson 1991; Likens 1992). Our analysisof Lake Mendota’s long-term responses to differing levels ofnutrients and herbivory provides insights into whether bioma-nipulation can be expected to produce sustained results inlarge, stratified eutrophic lakes. Our results suggest that her-bivory can affect water clarity in all seasons and that the great-est improvement in summer water clarity results from bothnutrient reductions and fish manipulations that allow her-bivory to increase.
Methods
Water clarity dataSecchi disk readings for Lake Mendota during 1900–1993 were com-piled from many published and unpublished sources. Five differentstyles of disks have been used: 10-cm white, 20-cm black–white,25-cm black–white, 20-cm white, and 30-cm white. Because readingsfrom the five disk styles are not exactly comparable, all disk readingswere standardized to a 20-cm black–white disk. Disk data sources for1900–1989, the style of disk used by each observer, and disk stand-ardization factors (range 0.956–1.129) were given in Lathrop (1992).Major data sources included readings by E.A. Birge, C. Juday, andtheir students for the early 1900s, the Madison Public Health Depart-ment for the 1920s through the 1940s, University of Wisconsin (UW)graduate students for many years from 1961 to 1981, the Wisconsin
Department of Natural Resources since the mid-1970s, and the UWsince 1987. When more than one reading was taken on a given day, adaily average was computed. Readings taken at stations other than thelake’s central deep area were ignored.
A total of 1701 daily Secchi disk readings from 70 calendar yearsduring 1900–1993 were found for Lake Mendota. However, manyyears had too few readings to document seasonal fluctuations withina given year; 9 years had only one reading. To assess pattern in thelong-term record, Secchi readings were stratified into six seasonalperiods: winter, spring turnover, early stratification, summer, destrati-fication, and fall turnover. These six periods were distinguished by thefollowing criteria: (i) the lake’s thermal structure in each of the sixperiods was relatively different; (ii) hypolimnetic anoxia was re-stricted to the summer and destratification periods; (iii) dissolved re-active phosphorus (DRP) concentrations in the surface waters weregenerally above analytical detection levels in all periods except sum-mer (Lathrop 1992); and (iv) the periods mirrored the seasonal suc-cession of phytoplankton and zooplankton in Lake Mendota (Lathropand Carpenter 1992a, 1992b), which closely followed the PlanktonEcology Group model for eutrophic lakes (Sommer et al. 1986).
We used fixed calendar dates to distinguish between all open-water seasonal periods (Table 1). However, we used the lake’s re-corded freeze-up date to separate each year’s fall turnover and winterperiods, and the recorded ice-out date to separate each year’s winterand spring turnover periods. The long-term average freeze-up andice-out dates for Lake Mendota are December 20 and April 5, butconsiderable year to year variation in the dates has occurred (Robert-son et al. 1992). As a result, Secchi disk readings taken through theice in December were considered as part of the next year’s winterrecord. This caused a total of 71 years to have Secchi records in ourdata set.
For each of the six seasonal periods in a given year, means of thestandardized Secchi disk readings were computed; their correspond-ing standard deviations were also computed if three or more readingswere available (Appendix 1). We assigned each year that had Secchireadings to one of three levels of nutrients and one of two levels ofherbivory (Appendix 2). To avoid circularity in our analysis, we ig-nored Secchi disk information in assigning each year’s nutrient andherbivory levels.
Nutrient levelsWe assigned a low nutrient level to all years prior to 1945, becauseDRP concentrations in Lake Mendota increased dramatically at theend of World War II (Fig. 1). This shift in nutrient status was relatedto an increase in DRP concentrations in the lake’s tributaries that werereceiving sewage effluents (Lathrop 1992). While sewage effluentshad been discharged in earlier years, loadings apparently increased asa result of urban growth and new commercial activity at the war’s end.The sewage was finally diverted from the lake after 1971.
We classified nutrient levels for all years between 1945 and 1951as medium. Other than the major increase in DRP concentrations thatoccurred around 1945, we did not believe that year to year variationsin DRP were reliable indicators of medium versus high nutrient lev-els. (As stated earlier, summer DRP concentrations since 1925 werealmost always low and near analytical detection levels. However,spring DRP concentrations appeared to be affected by algal bloomdensities and zooplankton grazing.) The medium classification for1945–1951 was reasonable because the application of P-based fertil-izers on Wisconsin farmland was minimal during the 1940s; its usageincreased markedly in the 1960s and peaked in the late 1970s(Lathrop 1992).
For years since 1961, we used a spring (January to March) runoffindex to determine whether Lake Mendota’s nutrient level was me-dium or high. This index was based on continuous discharge recordsobtained since 1954 by the United States Geological Survey on BlackEarth Creek, a stream draining a watershed immediately to the westof Lake Mendota’s watershed (Lathrop 1990). For the index, runoff
Period Mean n s
Winter (freeze-up to ice-out) 4.74 43 2.086Spring turnover (ice-out to 10 May
(day 130)) 2.79 52 1.133Early stratification (11 May to 29 June
(days 131–180)) 3.76 59 1.224Summer (30 June to 2 Sept.
(days 181–245)) 2.29 60 0.789Destratification (3 Sept. to 12 Oct.
(days 246–285)) 2.47 55 0.706Fall turnover (13 Oct. (day 286) to
freeze-up) 3.24 53 0.895
Note: Long-term means were computed from seasonal mean readings ofindividual years with Secchi disk readings (from Appendix 1).
Table 1.Long-term means of Secchi disk readings for differentseasonal periods in Lake Mendota during 1900–1993.
Lathrop et al. 2251
© 1996 NRC Canada

was separated from base flow in the mean daily discharge records. Wefelt that this index was a good indicator of annual P loadings to thelake, because runoff at Black Earth Creek was highly correlated withrunoff at Lake Mendota’s tributaries for years when continuous dis-charge records were available (Lathrop 1990), and because springrunoff contained much higher concentrations of biologically availableP than runoff during the late spring or summer months (Lathrop1986).
For our nutrient level criteria, we designated a year as having highnutrients if spring runoff was greater than the long-term mean runofffor 1954–1993, or if the 2-year running average spring runoff wasgreater than the long-term mean (Fig. 2). This allowed a realisticcarry-over effect from the previous year’s loading to influence theassigned nutrient level. To test this runoff index method for assigningnutrient levels, we compared the outcome with April total P (TP)
concentration data that were available for 1971–1993. All years werereadily classified except for 1978 (Fig. 2) because P loadings arestrongly correlated to stream flow (Lathrop 1986). While spring run-off in 1977–1978 was low, TP in April 1978 remained high after5 years of high runoff (1972–1976); runoff in late spring 1978 wasalso high (Lathrop 1992). Consequently, we designated 1978 as ahigh-nutrient year, but used the runoff criteria to determine nutrientlevels for all other years since 1961.
Herbivory levelsWe assigned a high herbivory level to each year if we had evidencethat the larger-bodied Daphnia pulicaria was the dominant Daphniaspecies during April to June, and a low herbivory level if the smaller-bodied Daphnia galeata mendotae (or Daphnia retrocurva) domi-nated. These spring months encompassed the clear-water phase inLake Mendota when Daphnia densities were usually the greatest(Lathrop and Carpenter 1992a, 1992b). The difference in grazingpotential between the two Daphnia species has been shown to be large(Burns 1969; Threlkeld 1979, 1980; Leibold 1990), and thus couldcause a marked effect on water clarity during different seasons de-pending on which species dominated. While we recognize that a her-bivory index based on spring Daphnia populations may not have beenalways valid for late summer or fall because herbivory could havedecreased owing to predation by an unusually large hatch of plank-tivorous fish or increased owing to a large summer mortality of adultplanktivorous fish, spring herbivory levels should be indicative ofannual herbivory for most years. Because any misrepresentation of ayear’s late summer or fall herbivory level should introduce additionalvariance in our analyses, our approach would be conservative fordetecting the effects of herbivory.
Spring Daphnia densities were available for many years duringthe early 1900s and for 1976–1993 (Fig. 3). For all sampling datesduring April to June, we computed time-weighted mean densities forboth the large and small species. Thus, the relative proportions of thesmall and large species were most heavily influenced by the samplingdates having the greatest densities. For most years in the historicalrecord, the spring Daphnia population was almost exclusively com-prised of one species. The only years that had Secchi records andsubstantial densities of both D. pulicaria and D. g. mendotae were1985 and 1986, which we assigned as having low and high herbivory,respectively. Daphnia biomass data clearly indicated that 1986 had amuch higher herbivory level than 1985, which was similar in biomassto the preceding years dominated by D. g. mendotae (Rudstam et al.1993).
Fig. 1. Mean annual (the previous October through September)DRP concentrations in the surface waters of Lake Mendota forselected years during 1925–1993. Means were computed frommonthly concentration data; interpolations for months without datawere done if 6 or more months in a given year had data. Datasources are summarized in Lathrop (1992).
Fig. 2. Spring (January–March) runoff water volumes for BlackEarth Creek, a watershed immediately west of Lake Mendota’swatershed, 1954–1993. April TP concentrations for surface watersof Lake Mendota, 1971–1993. Data sources are summarized inLathrop (1992).
Fig. 3. The relative proportion of Daphnia species in Lake Mendotaduring April–June for selected years since 1895. Data sources aresummarized in Lathrop and Carpenter (1992).
Can. J. Fish. Aquat. Sci. Vol. 53, 19962252
© 1996 NRC Canada

The herbivory level for other years was indexed to relative abun-dance records of cisco (Coregonus artedi) (Fig. 4), the principalplanktivore that has been shown to suppress D. pulicaria in LakeMendota (Luecke et al. 1992; Rudstam et al. 1993). Historical ac-counts indicate that cisco were abundant in the lake during the late1800s and the early 1900s when commercial fishing for cisco wasimportant (Lathrop et al. 1992; Magnuson and Lathrop 1992). Exceptfor occasional die-offs of older individuals during warm summers, thepopulation apparently remained large until about 1941 following amajor die-off in 1940. The population was considered low for manyyears thereafter; cisco were even thought to be extinct in Lake Men-dota after small die-offs in the early 1950s. However, as a result of anunexpectedly large recruitment in 1977, the biomass of cisco quicklyincreased and remained high until a massive summer mortality oc-curred in 1987 (Fig. 4). Calculated planktivory rates were high be-cause of this large cisco biomass between 1978 and 1987; rates weredramatically lower in years following the cisco die-off (Rudstam et al.1993). Since the 1987 die-off, cisco populations have remained lowin Lake Mendota.
While the cisco biomass data were not needed to determine her-bivory levels for 1976–1993 because Daphnia density data wereavailable, the inverse relationship between cisco densities and thebody size of the dominant Daphnia species during those years(Figs. 3–4) indicates the usefulness of the cisco index for assigningherbivory levels for other years. Other accounts of Daphnia popula-tions for selected years during the 1950s to the early 1970s (Lathropand Carpenter 1992b) also corroborate the cisco–Daphnia relation-ship. Finally, paleolimnological analysis of a core taken in 1987 indi-cated good correspondence between the percentage of daphnids thatare D. g. mendotae and cisco abundance (Kitchell and Sanford 1992;Johnson 1995).
AnalysesFor each seasonal period, we performed analysis of variance(ANOVA) to test for overall significant effects of nutrients, her-bivory, and their interaction on Secchi disk readings. Significant dif-ferences in Secchi readings between groups of years with the sixdifferent nutrient–herbivory combinations were compared via theirrespective 95% confidence intervals.
We performed ANOVA on both untransformed and log-trans-formed Secchi disk data. Normal probability plots of the untrans-
formed residuals never indicated a severe departure from normality.Also, analyses of the original data and log-transformed data gaveidentical conclusions with respect to nutrient and grazer effects andtheir interaction. Therefore, for simplicity we present analyses of un-transformed Secchi disk data.
We also analyzed the within-season variability for those years thathad enough readings to compute standard deviations. The relationshipbetween the seasonal variability estimate and its respective mean Sec-chi reading was determined for the first three open-water periods andfall turnover. Analysis was not performed for the winter period, whichhad less variability data, and the destratification period, which wasvery similar to summer. Because of the smaller number of years withvariability estimates, years with medium and high nutrient levels werecombined. Also, the low nutrient – high herbivory effect could not betested because variability data were not available.
Results
Water clarity in each of the six seasonal periods has varied inLake Mendota since 1900 (Table 1, Fig. 5). As expected, dif-ferences between seasons have also been pronounced. Waterclarity was generally greatest during the winter under the ice.During the open-water months, clarity was greatest during theearly stratification period when a clear-water phase normallyoccurred. Water clarity was lowest during the summer monthswhen blue-green algal blooms were frequently dense. Vari-ability among years was greatest during the winter period, fol-lowed by the early stratification and spring turnover periods.
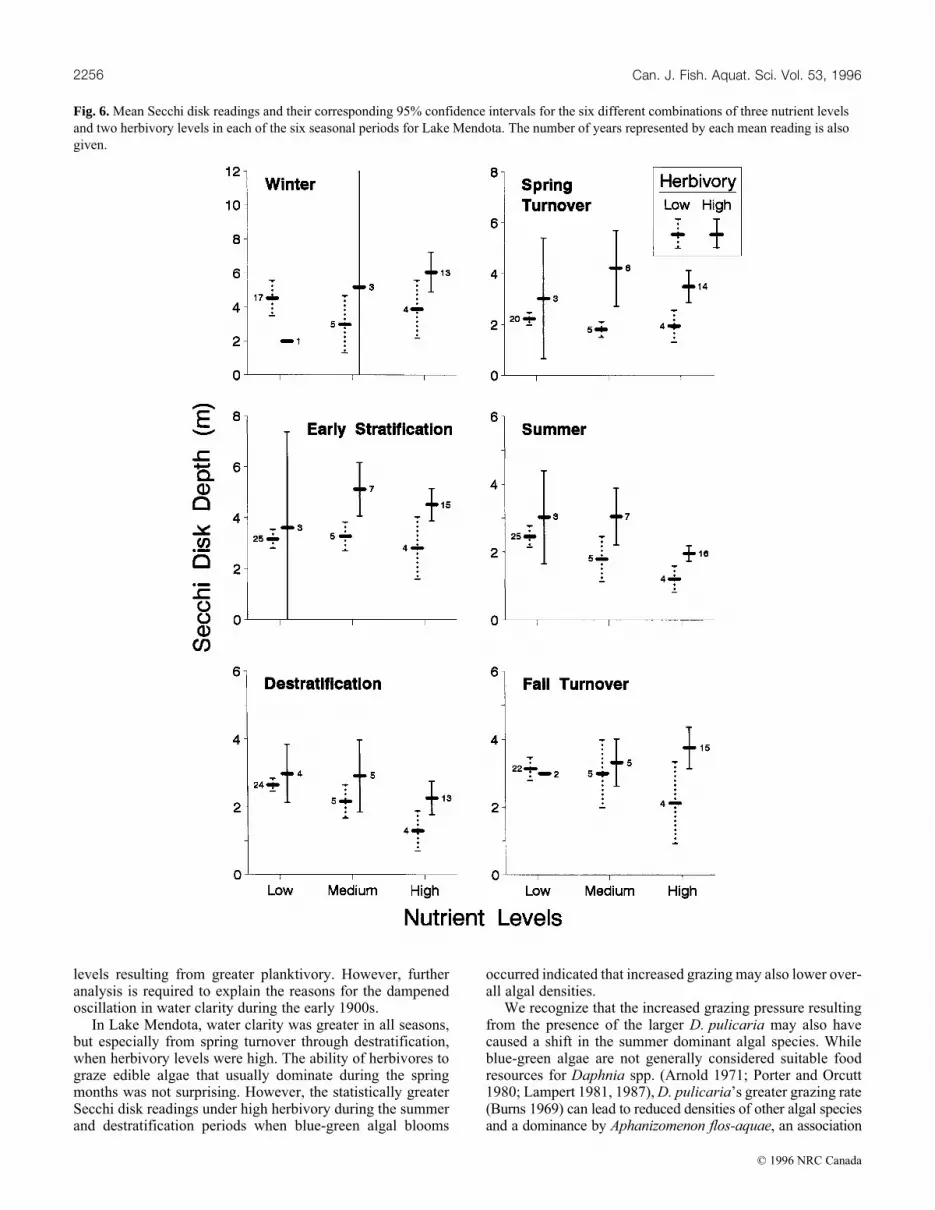
Our analysis of the effects of different nutrient and her-bivory levels on seasonal Secchi disk readings in Lake Men-dota produced statistically discernible results (Table 2, Fig. 6).In the spring turnover and early stratification periods, waterclarity was not affected by different nutrient levels, whereaswater clarity was significantly greater when herbivory washigh. In the summer and destratification periods, water claritywas affected by both nutrients and herbivory. Water claritydecreased as nutrient levels increased. Water clarity also wasgreater when herbivory was high at all three levels of nutrients.In the spring turnover through the destratification periods, theinteraction between nutrients and herbivory was not statisti-cally significant. During the fall turnover period, nutrient ef-fects were highly nonsignificant (large p values), while theherbivory and interaction terms were almost significant with ap value slightly greater than 0.05. Neither nutrients, herbivory,or their interaction were significant during the winter period.However, during the fall turnover and winter periods in yearswith high nutrients, water clarity was significantly greaterwhen herbivory was high (Fig. 6).
The within-season variability of Secchi disk readings indi-cated different responses to the three nutrient–herbivory com-binations analyzed (Fig. 7). During spring turnover, Secchivariability was small when herbivory was low at all nutrientlevels. When herbivory was high at the higher nutrient levels,Secchi variability was much greater if the mean Secchi readingwas also high. During the early stratification period, meanreadings were relatively similar for both nutrient levels at lowherbivory, but variability was greater at the higher nutrientlevels. For the higher nutrient levels, variability was similarfor both herbivory levels, but mean Secchi readings generallywere greater when herbivory was high. During the summer andfall turnover periods, the variability of Secchi disk readings
Fig. 4. The relative abundance of cisco (Coregonus artedi) in LakeMendota for selected years during 1860 to 1991. Maximal catch pernight was based on historical commercial fishing accounts; Telfordcatch per season (fall dip netting) was based on the personal diaryof J. Telford; UW sonar biomass was determined from acousticsonar and gill-net data (Rudstam et al. 1993).
Lathrop et al. 2253
© 1996 NRC Canada

Fig
.5.S
ecch
idis
kre
adin
gs(c
onve
rted
to20
-cm
blac
k–w
hite
disk
s)co
llect
eddu
ring
six
seas
onal
peri
ods
for
sele
cted
year
sdu
ring
1900
–199
3in
Lak
eM
endo
ta.T
hesh
orth
oriz
onta
lbar
isth
ese
ason
alm
ean
for
each
year
;the
vert
ical
line
isth
era
nge
ifm
ore
than
one
read
ing
was
take
n.T
hebr
oken
hori
zont
allin
eis
the
long
-ter
mm
ean
ofse
ason
alm
eans
for
indi
vidu
alye
ars.
Can. J. Fish. Aquat. Sci. Vol. 53, 19962254
© 1996 NRC Canada

was relatively small, and not very different for the three nutri-ent– herbivory combinations.
Discussion
Water clarity in Lake Mendota has been much more variablethis past century than earlier analyses had indicated. Even be-tween years when seasonal Secchi disk readings were similar,the relative effects of both nutrients and herbivory were mark-edly different. Of particular interest has been the effect of nu-trients and herbivory on water clarity during the summer whenthe lake was dominated by blue-green algae. A comparison ofSecchi readings from the early 1900s with readings from the1960s and early 1970s led to the conclusion that summer algalblooms had not increased in Lake Mendota during this pastcentury (Stewart 1976; Brock 1985). Considering Secchi read-ings obtained in more recent years plus earlier unpublishedrecords, Lathrop (1992) concluded that these periods of similarreadings were produced by different nutrient–herbivory inter-actions. The early 1900s were characterized by low nutrientsand low herbivory (owing to high planktivory), while the1960s through the mid-1970s were characterized by muchhigher nutrients and high herbivory (owing to low plank-tivory). The shallowest summer Secchi readings during thiscentury were recorded from the late 1970s through the mid-1980s when nutrients were relatively high and herbivory waslow, thus emphasizing the effect of nutrients alone on lakequality. Summer Secchi readings since the cisco die-off in
1987 generally have been similar to readings for the 1960sthrough the mid-1970s. Nutrient and herbivory levels weresimilar between those two groups of years.
Water clarity changes during the spring turnover and earlystratification periods also demonstrate the effects of herbivoryduring this century in Lake Mendota. In years when highplanktivory suppressed the larger bodied D. pulicaria, thesmaller D. g. mendotae, which has higher temperature require-ments for growth (Burns 1969; Threlkeld 1980), was not ableto reduce algal densities until late spring. In most years whenplanktivory was low, D. pulicaria populations increased ear-lier in the spring season and caused a rapid decline in algaldensities, leading to a higher variability in Secchi disk read-ings. However, in some years classified as having high her-bivory, D. pulicaria populations apparently did not increase asearly, causing Secchi disk readings in the spring turnover pe-riod to be similar to those recorded during low-herbivoryyears. Reasons for this delayed Daphnia increase are unclear.
During the early stratification period when herbivory waslow, the greater variability in Secchi readings at higher nutrientlevels in more recent years is noteworthy. This increased vari-ability could result from a decrease in ecosystem stability trig-gered by higher nutrient supply rates (Scheffer 1990;Carpenter 1992). Greater P availability fuels higher algal pro-duction, which then supports a larger Daphnia population thatovergrazes its food resources. Another factor that could havecaused the smaller variability in Secchi readings in the earlystratification period during the early 1900s is lower herbivory
Season Source SS df MS F p
Winter Nutrients 9.832 2 4.916 1.373 0.266Herbivory 1.787 1 1.787 0.499 0.484Interaction 17.588 2 8.794 2.457 0.100Error 132.443 37 3.580
Spring turnover Nutrients 0.918 2 0.459 0.663 0.520Herbivory 21.299 1 21.299 30.753 0.000Interaction 3.497 2 1.749 2.525 0.091Error 31.860 46 0.693
Early stratification Nutrients 3.802 2 1.901 1.947 0.153Herbivory 15.417 1 15.417 15.790 0.000Interaction 3.336 2 1.668 1.708 0.191Error 51.747 53 0.976
Summer Nutrients 8.568 2 4.284 10.172 0.000Herbivory 6.500 1 6.500 15.434 0.000Interaction 0.706 2 0.353 0.838 0.438Error 22.743 54 0.421
Destratification Nutrients 7.301 2 3.650 10.234 0.000Herbivory 4.163 1 4.163 11.671 0.001Interaction 0.674 2 0.337 0.945 0.396Error 17.479 49 0.357
Fall turnover Nutrients 0.262 2 0.131 0.192 0.826Herbivory 2.606 1 2.606 3.828 0.056Interaction 4.322 2 2.161 3.174 0.051Error 32.005 47 0.681
Note: Results in bold face have p values < 0.05.
Table 2.ANOVA results of the effect of nutrient and herbivory levels on seasonal Secchi disk readings recorded on Lake Mendota during1900–1993.
Lathrop et al. 2255
© 1996 NRC Canada
levels resulting from greater planktivory. However, furtheranalysis is required to explain the reasons for the dampenedoscillation in water clarity during the early 1900s.
In Lake Mendota, water clarity was greater in all seasons,but especially from spring turnover through destratification,when herbivory levels were high. The ability of herbivores tograze edible algae that usually dominate during the springmonths was not surprising. However, the statistically greaterSecchi disk readings under high herbivory during the summerand destratification periods when blue-green algal blooms
occurred indicated that increased grazing may also lower over-all algal densities.
We recognize that the increased grazing pressure resultingfrom the presence of the larger D. pulicaria may also havecaused a shift in the summer dominant algal species. Whileblue-green algae are not generally considered suitable foodresources for Daphnia spp. (Arnold 1971; Porter and Orcutt1980; Lampert 1981, 1987), D. pulicaria’s greater grazing rate(Burns 1969) can lead to reduced densities of other algal speciesand a dominance by Aphanizomenon flos-aquae, an association
Fig. 6. Mean Secchi disk readings and their corresponding 95% confidence intervals for the six different combinations of three nutrient levelsand two herbivory levels in each of the six seasonal periods for Lake Mendota. The number of years represented by each mean reading is alsogiven.
Can. J. Fish. Aquat. Sci. Vol. 53, 19962256
© 1996 NRC Canada

found in many highly eutrophic lakes (Hrbáeek 1964; Lynch1980; Ganf 1983; Andersson and Cronberg 1984; Pechar andFott 1991). Blooms of species with large colony sizes such asA. flos-aquae can result in greater Secchi disk readings thanwould occur if the blooms were dominated by a similarbiomass of blue-green algae of smaller size. This clumpingeffect may explain why Benndorf (1990), in his review ofbiomanipulation case studies of lakes with high nutrient load-ings, reported no declines in algal biomass but increases inwater clarity.
The lack of a significant nutrient effect on water clarity inLake Mendota during all seasonal periods except summer anddestratification is reasonable. DRP concentrations were wellabove analytical detection levels throughout the year except inthe summer months even during the low-nutrient years in theearly 1900s (Lathrop 1992). Because measurable DRP concen-trations indicate that P is not growth limiting to algae (Brown-lee and Murphy 1983; Prepas 1983; Fisher and Lean 1992),the fall, winter, and spring algae in Lake Mendota would nothave responded to additional nutrients. In the summer, waterclarity declines as a higher nutrient status stimulates more algal
growth even though DRP concentrations continue to be low.The same inverse relationship between nutrient levels andwater clarity occurs during the destratification period. Appar-ently, an increase in DRP and inorganic N concentrations fromentrained hypolimnetic nutrients during destratification doesnot stimulate the blue-green algal populations enough togreatly alter densities established during the summer.
Although biomanipulation efforts are mainly directed at re-ducing summer blue-green algal densities, reviews of previousstudies suggest that these efforts are ineffective in large eutro-phic lakes (Benndorf 1990; Reynolds 1994). Our analysis ofLake Mendota’s long-term Secchi disk record indicates thatgreater water clarity during the summer months can beachieved by both P reductions and increased herbivory causedby a shift to larger sized Daphnia spp. This indicates that lakesize and trophic status do not preclude lakes from achievinggreater water clarity from biomanipulation efforts that increaseherbivory. Our analysis also shows that grazer effects on waterclarity can be detected over periods of decades. In principle,there may be no time limit on the capacity of grazers to controlphytoplankton. In practice, however, sustained biomanipulation
Fig. 7. Mean Secchi disk readings and associated standard deviations for all years with at least three readings in each seasonal period. Data arestratified according to three combinations of nutrients and herbivory levels: low nutrients and low herbivory (open circles), medium–highnutrients and low herbivory (asterisks), and medium–high nutrients and high herbivory (solid circles).
Lathrop et al. 2257
© 1996 NRC Canada

may be difficult. In Lake Mendota, for example, a massivebiomanipulation effort (Kitchell 1992) could not be sustainedbecause of a tremendous increase in angling pressure on thestocked piscivores (Johnson and Staggs 1992). While sustain-ing high herbivory remains a challenge (Shapiro 1990), ouranalysis of Lake Mendota’s long-term water clarity record in-dicates that management programs incorporating both P reduc-tion and biomanipulation may produce more improvements insummer water clarity in large eutrophic lakes than if only onemanagement technique is used.
Acknowledgments
We thank T.D. Brock, R.A. Ragotzkie, D.M. Robertson,B.G. Saley, W.C. Sonzogni, and K.W. Stewart for providingor locating unpublished Secchi disk records for Lake Mendota.We also thank K.L. Cottingham, D.E. Schindler,P.A. Soranno, and C.A. Stow and anonymous reviewers fortheir many helpful comments on the draft. This project wasfunded by the Wisconsin Department of Natural Resources,the Federal Aid in Sport Fish Restoration Act under projectF-95-P, and the North Temperate Lakes Long-term EcologicalResearch program funded by National Science Foundationgrant No. DEB-9011660.
References
Andersson, G., and Cronberg, G. 1984. Aphanizomenon flos-aquaeand large Daphnia: an interesting plankton association in hyper-trophic water. In Nordisk limnologsymposium, interactions be-tween trophic levels in freshwaters. Edited by S. Bosheim andM. Nicholls. Norsk Limnologforening, Oslo, Norway. pp. 63–76.
Arnold, D.E. 1971. Ingestion, assimilation, survival, and reproduc-tion by Daphnia pulex fed seven species of blue-green algae. Lim-nol. Oceanogr. 16: 906–920.
Benndorf, J. 1987. Food web manipulation without nutrient control: auseful strategy in lake restoration? Schweiz. Z. Hydrol. 49: 237– 248.
Benndorf, J. 1990. Conditions for effective biomanipulation; conclu-sions derived from whole-lake experiments in Europe. Hydrobi-ologia, 200–201: 187–203.
Brock, T.D. 1985. A eutrophic lake: Lake Mendota, Wisconsin.Springer-Verlag, New York.
Brownlee, B.G., and Murphy, T.P. 1983. Nitrogen fixation and phos-phorus turnover in a hypertrophic lake. Can. J. Fish. Aquat. Sci.40: 1853–1860.
Burns, C.W. 1969. Relation between filtering rate, temperature, andbody size in four species of Daphnia. Limnol. Oceanogr.14: 696–700.
Carpenter, S.R. 1992. Destabilization of planktonic ecosystems andblooms of blue-green algae. In Food web management: a casestudy of Lake Mendota, Wisconsin. Edited by J.F. Kitchell. Sprin-ger-Verlag, New York. pp. 461–482.
Carpenter, S.R., Kitchell, J.F., Cottingham, K.L., Schindler, D.E.,Christensen, D.L., Post, D.M., and Voichick, N. 1996. Chloro-phyll variability, nutrient input and grazing: evidence from whole-lake experiments. Ecology, 77: 725–735.
Edmondson, W.T. 1991. The uses of ecology: Lake Washington andbeyond. University of Washington Press, Seattle, Wash.
Elser, J.J., and Goldman, C.R. 1992. Zooplankton effects on phyto-plankton in lakes of contrasting trophic status. Limnol. Oceanogr.36: 64–90.
Findlay, D.L., Kasian, S.E.M., Hendzel, L.L., Hegehr, G.W., Schin-dler, E.U., and Shearer, J.A. 1994. Biomanipulation of Lake 221in the Experimental Lakes Area (ELA): effects on phytoplanktonand nutrients. Can. J. Fish. Aquat. Sci. 51: 2794–2807.
Fisher, T.R., and Lean, D.R.S. 1992. Interpretation of radiophosphatedynamics in lake water. Can. J. Fish. Aquat. Sci. 49: 252–258.
Ganf, G.C. 1983. An ecological relationship between Aphanizomenonand Daphnia pulex. Aust. J. Mar. Freshwater Res. 34: 755–773.
Gulati, R.D., Lammens, E.H.R.R., Meijer, M.-L., and van Donk, E.(Editors). 1990. Biomanipulation tool for water management.Hydrobiologia, 200–201.
Hrbáeek, J. 1964. Contribution to the ecology of water-bloom-form-ing blue-green algae: Aphanizomenon flos aquae and Microcystisaeruginosa. Verh. Int. Ver. Theor. Angew. Limnol. 15: 837–846.
Johnson, B.M., and Staggs, M.D. 1992. The fishery. In Food webmanagement: a case study of Lake Mendota, Wisconsin. Edited byJ.F. Kitchell. Springer-Verlag, New York. pp. 353–375.
Johnson, T.B. 1995. Long-term dynamics of the zooplanktivorousfish community in Lake Mendota, Wisconsin. Ph.D. thesis, Uni-versity of Wisconsin, Madison, Wis.
Kitchell, J.F. (Editor). 1992. Food web management: a case study ofLake Mendota, Wisconsin. Springer-Verlag, New York.
Kitchell, J.F., and Sanford, P.R. 1992. Paleolimnological evidence offood web dynamics in Lake Mendota. In Food web management:a case study of Lake Mendota, Wisconsin. Edited by J.F. Kitchell.Springer-Verlag, New York. pp. 31–47.
Lampert, W. 1981. Inhibitory and toxic effects of blue-green algae onDaphnia. Int. Rev. Gesamten Hydrobiol. 66: 285–298.
Lampert, W. 1987. Laboratory studies on zooplankton-cyanobacteriainteractions. N.Z. J. Mar. Freshwater Res. 21: 483–490.
Lathrop, R.C. 1986. A simplified method for obtaining monitoredphosphorus loadings. Lake Reservoir Manage. 2: 20–26.
Lathrop, R.C. 1990. Response of Lake Mendota (Wisconsin, U.S.A.)to decreased phosphorus loadings and the effect on downstreamlakes. Verh. Int. Ver. Theor. Angew. Limnol. 24: 457–463.
Lathrop, R.C. 1992. Nutrient loadings, lake nutrients, and water clar-ity. In Food web management: a case study of Lake Mendota,Wisconsin. Edited by J.F. Kitchell. Springer-Verlag, New York.pp. 69–96.
Lathrop, R.C., and Carpenter, S.R. 1992a. Phytoplankton and theirrelationship to nutrients. In Food web management: a case studyof Lake Mendota, Wisconsin. Edited by J.F. Kitchell. Springer-Verlag, New York. pp. 97–126.
Lathrop, R.C., and Carpenter, S.R. 1992b. Zooplankton and their re-lationship to phytoplankton. In Food web management: a casestudy of Lake Mendota, Wisconsin. Edited by J.F. Kitchell. Springer-Verlag, New York. pp. 127–150.
Lathrop, R.C., Nehls, S.B., Brynildson, C.L., and Plass, K.R. 1992.The fishery of the Yahara lakes. Wis. Dep. Nat. Resour. Tech.Bull. No. 181.
Leibold, M.A. 1990. Resource edibility and the effects of predatorsand productivity on the outcome of trophic interactions. Am. Nat.134: 922–949.
Likens, G.E. 1983. A priority for ecological research. Bull. Ecol. Soc.Am. 64: 234–243.
Likens, G.E. (Editor). 1989. Long-term studies in ecology. Springer-Verlag, New York.
Likens, G.E. 1992. The ecosystem approach: its use and abuse. Ecol-ogy Institute, Olendorf–Luhe, Germany.
Luecke, C., Rudstam, L.G., and Allen, Y. 1992. Interannual patternsof planktivory 1987–89: an analysis of vertebrate and invertebrateplanktivores. In Food web management: a case study of LakeMendota, Wisconsin. Edited by J.F. Kitchell. Springer-Verlag,New York. pp. 275–301.
Lynch, M. 1980. Aphanizomenon blooms: alternate control and culti-vation by Daphnia pulex. In Evolution and ecology of zooplank-ton communities. Edited by W.C. Kerfoot. University Press ofNew England, Hanover, N.H. pp. 299–304.
Magnuson, J.J. 1990. Long-term ecological research and the invisiblepresent. BioScience, 40: 495–501.
Magnuson, J.J., and Lathrop, R.C. 1992. Historical changes in the fish
Can. J. Fish. Aquat. Sci. Vol. 53, 19962258
© 1996 NRC Canada

community. In Food web management: a case study of Lake Men-dota, Wisconsin. Edited by J.F. Kitchell. Springer-Verlag, NewYork. pp. 191–231.
McQueen, D.J., Post, J.R., and Mills, E.L. 1986. Trophic relation-ships in freshwater ecosystems. Can. J. Fish. Aquat. Sci.43: 1571–1581.
Pechar, L., and Fott, J. 1991. On the occurrence of Aphanizomenonflos-aquae var. flos-aquae in fish ponds. Int. Rev. Gesamten Hy-drobiol. 76: 57–66.
Persson, L., Johansson, L., Andersson, G., and Hamrin, S.F. 1992.Trophic interactions in temperate lake ecosystems: a test of foodchain theory. Am. Nat. 140: 59–84.
Porter, K.G., and Orcutt, J.D., Jr. 1980. Nutritional adequacy, man-ageability, and toxicity as factors that determine the food qualityof green and blue-green algae for Daphnia. In Evolution and ecol-ogy of zooplankton communities. Edited by W.C. Kerfoot. Uni-versity Press of New England, Hanover, N.H. pp. 268–281.
Prepas, E.E. 1983. Orthophosphate turnover time in a shallow pro-ductive lake. Can. J. Fish. Aquat. Sci. 40: 1412–1418.
Reinertsen, H.A., Jensen, A., Koksvik, J.I., Langelaand, A., andOlsen, Y. 1990. Effects of fish removal on the limnetic ecosystemof a eutrophic lake. Can. J. Fish. Aquat. Sci. 47: 166–173.
Reynolds, C.S. 1994. The ecological basis for the successful bioma-nipulation of aquatic communities. Arch. Hydrobiol. 130: 1–33.
Robertson, D.M., Ragotzkie, R.A., and Magnuson, J.J. 1992. Lakeice records used to detect historical and future climate changes.Clim. Change, 21: 407–427.
Rudstam, L.G., Lathrop, R.C., and Carpenter, S.R. 1993. The rise and
fall of a dominant planktivore: direct and indirect effects onzooplankton. Ecology, 74: 303–319.
Scheffer, M. 1990. Multiplicity of stable states in freshwater systems.Hydrobiologia, 200–201: 475–486.
Schindler, D.W. 1987. Detecting ecosystem responses to anthropo-genic stress. Can. J. Fish. Aquat. Sci. 44(Suppl. 1): 6–25.
Schindler, D.W. 1988. Experimental studies of chemical stressors onwhole lake ecosystems. Verh. Int. Ver. Theor. Angew. Limnol.23: 11–41.
Schindler, D.W. 1990. Experimental perturbations of whole lakes astests of hypotheses concerning ecosystem structure and function.Oikos, 57: 25–41.
Shapiro, J. 1990. Biomanipulation: the next phase — making it stable.Hydrobiologia, 200–201: 13–27.
Shapiro, J., and Wright, D.I. 1984. Lake restoration by biomanipula-tion: Round Lake, Minnesota the first two years. Freshwater Biol.14: 371–383.
Sommer, U., Gliwicz, Z.M., Lampert, W., and Duncan, A. 1986. ThePEG*-model of seasonal succession of planktonic events in freshwaters. Arch. Hydrobiol. 106: 433–471.
Stewart, K.W. 1976. Oxygen deficits, clarity, and eutrophication insome Madison lakes. Int. Rev. Gesamten Hydrobiol. 61: 563–579.
Threlkeld, S.T. 1979. The midsummer dynamics of two Daphniaspecies in Wintergreen Lake. Ecology, 60: 165–179.
Threlkeld, S.T. 1980. Habitat selection and population growth of twocladocerans in seasonal environments. In Evolution and ecologyof zooplankton communities. Edited by W.C. Kerfoot. UniversityPress of New England, Hanover, N.H. pp. 346–357.
Winter Spring turnover Early stratificationSummer
stratification
Year n Mean s n Mean s n Mean s n Mean s
19001901 3 9.40 2.816 5 1.92 0.807 4 3.75 0.379190219051906 1 0.9 9 4.04 1.473 10 2.50 0.5141907 3 3.60 0.557 4 2.68 0.435 7 3.04 0.716 8 1.83 0.1391908 1 2.11909 3 5.33 0.416 3 3.37 0.058 3 2.27 0.2311911 1 2.9 2 2.851912 1 4.2 2 2.101913 1 2.0 4 2.70 0.766 2 1.851914 3 1.87 0.493 6 2.47 0.774 8 2.29 0.3141915 8 4.38 1.478 3 2.27 0.569 7 2.46 0.412 8 2.73 0.7521916 2 4.00 3 1.43 0.513 7 2.29 0.639 8 2.64 0.6781917 3 2.27 0.252 7 2.69 0.694 8 2.69 0.8031918 4 5.33 0.789 4 2.33 0.287 14 3.32 1.303 16 2.14 0.4101919 4 1.85 0.100 5 3.90 0.752 4 2.45 0.4511920 1 1.7 5 2.78 1.666 7 2.39 0.7271921 3 4.10 0.872 4 2.35 0.436 1 2.2 3 3.77 0.5861922 4 2.78 1.014 5 2.28 0.1921923 7 3.60 2.041 4 1.55 0.252 6 3.08 0.360 2 1.751924 6 2.93 1.277 2 2.25 6 3.13 0.543 6 2.13 0.1971925 9 3.78 1.360 7 2.21 0.107 7 2.56 0.627 8 2.34 0.6971926 7 3.54 0.852 1 2.1 5 2.40 0.930 8 1.71 0.3141927 4 5.83 1.953 1 2.9 3 6.27 1.3651928 3 3.00 1.480 2 3.45 1 2.8
Destratification Fall turnover
n Mean s n Mean s
1 4.7
1 3.31 2.47 2.64 0.310 4 3.03 0.3772 2.203 1.90 0.000 5 2.58 0.303
1 1.35 2.32 0.259 5 2.40 0.3944 2.23 0.222 12 3.93 0.6046 2.28 0.293 25 3.04 0.4816 2.62 0.371 40 2.86 0.3876 2.68 0.412 44 3.09 0.4056 3.10 0.395 8 3.90 1.0184 2.15 0.173 1 2.6
5 2.78 0.390 7 3.69 0.7564 2.65 0.714 8 3.14 0.7567 3.20 0.245 4 2.85 0.3706 2.90 0.738 11 3.00 0.4712 2.25 6 3.90 0.6033 3.30 0.529 4 3.00 0.4553 2.90 0.300 4 3.48 0.3861 2.8 2 3.601 3.0 4 3.20 0.702
Appendix 1. Annual means of Secchi disk readings recorded in each seasonal period on Lake Mendota during 1900–1993.
Lathrop et al. 2259
© 1996 NRC Canada

Winter Spring turnover Early stratificationSummer
stratification
Year n Mean s n Mean s n Mean s n Mean s
1929 4 3.33 0.922 3 2.40 0.200 2 4.35 3 2.43 0.2891930 3 7.07 4.539 2 2.15 3 3.20 1.153 5 1.94 0.2701931 1 6.81933 1 2.01934 1 2.51936 1 2.8 1 3.51939 1 5.219411942 1 4.0 1 5.0 3 3.60 0.2651943 1 2.0 1 3.0 2 3.85 1 3.01944 1 2.1 1 2.0 2 2.501945 1 4.0 2 3.85 2 2.301946 1 4.0 2 5.80 2 3.351947 1 4.61951 1 1.8 4 1.85 0.420 2 3.80 1 3.31961 2 2.301962 7 7.23 3.086 3 3.57 0.252 10 4.78 2.132 10 2.35 0.7071963 3 5.83 1.4191965 1 5.3 2 1.901966 10 3.84 1.203 11 3.67 2.127 14 2.09 0.3701967 5 3.30 1.546 2 4.45 10 3.57 1.714 16 2.43 1.1971968 1 6.01971 4 4.33 2.740 14 6.54 2.736 5 2.46 0.3361972 3 8.80 4.677 4 2.70 0.283 13 4.45 1.988 17 1.66 0.3991973 5 6.20 1.020 7 2.87 0.745 5 2.96 1.756 2 1.301974 2 5.15 4 2.33 0.330 6 5.32 1.677 2 2.351975 3 2.60 0.600 3 5.37 1.914 3 2.07 0.6661976 2 6.85 7 3.89 1.119 10 4.71 2.090 13 1.45 0.4591977 3 6.20 1.664 7 3.51 1.805 8 3.81 1.720 11 1.13 0.3551978 3 3.60 3.005 3 1.57 0.058 12 3.49 1.516 15 1.41 0.5431979 9 3.89 1.587 6 2.35 0.464 10 1.76 0.734 12 1.42 0.3351980 7 2.80 0.666 3 1.67 0.321 10 3.22 1.676 14 0.97 0.2581981 5 2.52 0.801 8 1.69 0.336 11 3.37 1.855 14 1.26 0.2101982 2 4.90 2 1.65 3 2.70 1.803 4 1.48 0.3591983 1 3.4 3 2.03 0.451 3 3.67 2.779 4 2.43 0.6181984 1 1.2 3 1.60 0.173 3 2.93 2.444 5 1.54 0.3211985 1 5.3 3 2.20 0.755 3 2.83 1.210 5 1.04 0.2701986 2 3.00 3 1.77 0.321 3 4.23 2.409 5 2.16 0.4341987 2 2.90 7 2.10 0.451 8 3.73 2.561 9 2.30 0.5341988 1 7.0 7 5.21 3.501 11 5.63 2.932 12 3.45 0.6691989 1 6.9 7 4.52 1.909 10 6.82 2.654 13 2.37 0.9221990 2 3.60 11 2.63 0.413 25 2.34 1.256 18 1.83 0.3451991 5 8.91 2.920 8 4.76 2.204 8 5.79 2.987 7 2.39 0.6981992 2 6.75 6 6.02 2.774 15 4.28 2.639 22 1.93 0.4311993 2 7.00 5 5.55 1.610 13 4.79 1.223 21 1.74 0.445
Appendix 1 (concluded).
Destratification Fall turnover
n Mean s n Mean s
1 3.7 1 2.42 2.20 2 3.401 2.3
1 2.33 3.33 0.5772 2.85 1 3.01 3.50 1 3.01 2.8 1 3.01 4.3 1 3.0
4 2.38 0.320 6 3.27 0.8668 3.38 1.940 8 4.13 1.243
1 1.0 2 4.304 1.88 0.411 4 3.93 1.1624 1.73 0.538 2 2.15
5 2.00 0.292 7 2.80 0.73713 3.05 0.747 8 3.21 0.458
2 2.95
1 4.68 1.48 0.442 5 2.70 1.1515 1.26 0.089 7 1.81 0.2047 1.60 0.520 7 2.50 0.5074 1.25 0.058 5 2.52 0.8148 1.56 0.141 9 2.51 0.8807 1.56 0.276 5 2.12 0.6302 2.05 1 3.72 2.35 1 2.22 2.30 1 3.81 0.8 1 1.01 2.5 2 3.503 2.60 0.400 5 3.16 0.4107 2.97 0.818 7 3.80 1.3287 2.34 0.391 8 3.58 0.575
13 3.14 1.117 10 6.16 0.9323 3.48 0.597 4 4.96 1.0968 2.55 0.604 5 4.02 1.2708 1.93 0.596 4 4.23 1.066
Can. J. Fish. Aquat. Sci. Vol. 53, 19962260
© 1996 NRC Canada

Year n Nutrients Herbivory Year n Nutrients Herbivory
1900 1 L L 1944 6 L H1901 12 L L 1945 7 M H1902 1 L L 1946 7 M H1905 1 L L 1947 1 M H1906 31 L L 1951 8 M H1907 24 L L 1961 12 H H1908 9 L L 1962 46 H H1909 9 L L 1963 3 H H1911 4 L L 1965 6 H H1912 13 L L 1966 43 H H1913 23 L L 1967 39 H H1914 48 L L 1968 1 M H1915 72 L L 1971 35 M H1916 70 L L 1972 58 H H1917 32 L L 1973 21 H H1918 43 L L 1974 14 H H1919 13 L L 1975 10 H H1920 25 L L 1976 45 H H1921 23 L L 1977 41 H H1922 20 L L 1978 47 H L1923 36 L L 1979 46 H L1924 28 L L 1980 51 H L1925 38 L L 1981 50 M L1926 28 L L 1982 14 M L1927 11 L L 1983 14 M L1928 11 L L 1984 15 M L1929 14 L L 1985 14 H L1930 17 L L 1986 16 H H1931 2 L L 1987 34 M L1933 1 L L 1988 45 M H1934 1 L L 1989 46 H H1936 2 L L 1990 79 H H1939 1 L L 1991 35 H H1941 1 L H 1992 58 M H1942 8 L H 1993 53 H H1943 8 L H
Note: L, low; M, medium; H, high.
Appendix 2. Number of Secchi disk readings recorded on Lake Mendota during 1900–1993 and theassigned levels of nutrients and herbivory for each year with readings.
Lathrop et al. 2261
© 1996 NRC Canada





























