Embed Size (px)
DESCRIPTION
第十三章 蛋白质的生物合成 Chapter 13 Biosynthesis of Protein. 一、 蛋白质的生物合成的概念. 蛋白质的生物合成过程,就是将 DNA 传递给 mRNA 的遗传信息,再具体的解译为蛋白质中氨基酸排列顺序的过程,这一过程被称为 翻译 (translation) 。. 二、 蛋白质生物合成体系. 除合成原料氨基酸外,蛋白质生物合成体系还包括 mRNA,tRNA, 核蛋白体,有关的酶,蛋白因子, ATP , GTP 等供能物质及必要的无机离子。. ㈠mRNA. mRNA 作为指导蛋白质生物合成的模板。. ⒈ 三联体密码 : - PowerPoint PPT Presentation
Citation preview

第十三章 蛋白质的生物合成 第十三章 蛋白质的生物合成 Chapter 13 Biosynthesis of ProteinChapter 13 Biosynthesis of Protein
蛋白质的生物合成过程,就是将 DNA传递给mRNA的遗传信息,再具体的解译为蛋白质中氨基酸排列顺序的过程,这一过程被称为翻译 (translation) 。
一、 蛋白质的生物合成的概念

二、蛋白质生物合成体系
除合成原料氨基酸外,蛋白质生物合成体系还包括mRNA,tRNA,核蛋白体,有关的酶,蛋白因子,ATP,GTP等供能物质及必要的无机离子。

⒈三联体密码 : mRNA从 5 ’ →3’方向,由起始密码
AUG开始到终止密码之间的区域称为一个开放读码框架,读码框架内每三个相邻的核苷酸组成三联体,代表一个氨基酸的信息,此三联体就称为密码 (coden)。共有 64 种不同的密码。
㈠mRNAmRNA作为指导蛋白质生物合成的模板。

2、三联体密码特点:连续性:密码的三联体不间断,必须三个一组连续读下去。
简并性:多个密码共同编码一个氨基酸的对象。通用性:全部生物在蛋白质生物合成中都使用同一套密码(但在线粒体或叶绿体中特殊)。摆动性:密码子与反密码子配对时,不遵守 碱基配对原则的现象。起始密码与终止密码:起始密码 -AUG;终止密码 -UAA、UAG、UGA。方向性:mRNA密码子的阅读方向是
5 ,→ 3 ,。

遗传密码的连续性遗传密码的连续性

遗传密码的简并性和通用性遗传密码的简并性和通用性

㈡ tRNA
1、反密码 (anticoden):tRNA反密码环中 部的三个 核苷酸构成三联体,可以识别
mRNA上相应的密码,此三联体就称为反密码 (anticoden)。
tRNA是转运氨基酸的工具。在氨基酸tRNA合成酶催化下,特定的 tRNA可与相应的氨基酸结合,生成氨基酰 tRNA,从而
携带氨基酸参与蛋白质的生物合成。

2、不稳定配对 :反密码的第一个核苷酸与密码第三核苷酸之间的配对,并不严格遵循碱基互补原则,称之不稳定配对。如:
5’- 反密码第一个核苷酸 3’- 密码第三核苷酸
I A、U或C U A或G G U或C

遗传密码的摆动配对遗传密码的摆动配对

起动 tRNA:能够识别mRNA中 5′端起动密码 AUG的 tRNA是一种特殊的tRNA,称为起动 tRNA。• 原核生物:起动 tRNA是tRNAi
fmet;
• 真核生物:起动 tRNA是tRNAi
met。在原核生物和真核生物中,均存在另一种携带蛋氨酸的 tRNA,识别非起动部位的蛋氨酸密码, AUG。

㈢ rRNA和核蛋白体rRNA与多种蛋白质组成核蛋白体,是肽链合成的场所。
核糖体 亚基 rRNA
蛋白质种类
原核
真核
70S30S 16S 21种
50S 5S 23S
34种
80S40S
18S 大约 33种
5S 5.8S 28S
大约 50
种
细胞
60S

核蛋白体的组装核蛋白体的组装

将 30S小亚基抱住;
大肠杆菌核蛋白体空间结构
·为一椭圆球体;·30S亚基呈哑铃状;·50S亚基带有三角,中间凹陷形成空穴,·两亚基的结合面为蛋白质生物合成的场所。

小亚基:可与mRNA、GTP和起动tRNA结合。
大亚基:具有两个不同的 tRNA结合点:• A位(右) : 受位或氨酰基位,可与新进入的氨基酰 tRNA结合 .
• P位(左) : 给位或肽酰基位,可与延伸中的肽酰基 tRNA结合。
核蛋白体大、小亚基的功能

具有转肽酶活性:将给位上的肽酰基转移给受位上的氨基酰 tRNA,形成肽键。
具有GTPase活性 :水解GTP,获得能量。
具有起动因子、延长因子及释放因子的结合部位。

多核蛋白体:由若干核蛋白体结合在一条mRNA上同时进行多肽链的翻译所形成的念球状结构称为多核蛋白体。

㈣ 起动因子( IF)
这是一些与多肽链合成起动有关的蛋
白因子。
原核生物中存在 3种起动因子( IF1-3 )。
真核生物中存在 9种起动因子( eIF )。起动因子的作用主要是促进核蛋白体小亚
基与起动 tRNA及模板mRNA结合。

EFTU(GTPase) α (GTPase)
EFT EF1 β
原核 EFTS 真核 γ
EFG(转位酶) EF2(转位酶)
㈤ 延长因子( EF)
作用:主要促使氨基酰 tRNA进入核蛋白的受 体, 并可促进移位过程。
原核生物中存在 3种延长因子( EFTU, EFTS, EFG) .真核生物中存在 2种( EF1, EF2)。

RF:主要作用是识别终止密码 ,协助多肽链的释放。
•原核生物中有 4种•真核生物中只有 1种。 RR:主要作用是促使mRNA从核蛋白体上释放。
㈥释放因子( RF)、( RR)

功能:该酶存在于胞液中,与特异氨基酸的活化以及氨基酰 tRNA的合成有关。
特点:• 每种氨基酰 tRNA合成酶对相应氨基酸以及携带氨基酸的数种 tRNA具有高度特异性 .
• 目前认为,该酶对 tRNA 的识别,是因为在tRNA的氨基酸臂上存在特定的识别密码,即第二套遗传密码。
㈦ 氨基酰 tRNA 合成酶

G3 U70
G3 U70
丙氨酸 tRNA(含密码子)
人工合成的小螺旋
(不含密码子)
Ala

㈧ 供能物质和无机离子
多肽链合成时,需 ATP、GTP作为供能物质,并需Mg2+、 K+ 参与。氨基酸活化时需消耗 2分子高能磷酸
键, 肽键形成时又消耗 2分子高能磷酸键,
故缩合一分子氨基酸残基需消耗 4分子高能磷酸键。

二、蛋白质生物合成过程二、蛋白质生物合成过程 蛋白质生物合成过程包括三大步骤 :•氨基酸的活化与搬运•活化氨基酸在核蛋白体上的缩合•多肽链合成后的加工修饰
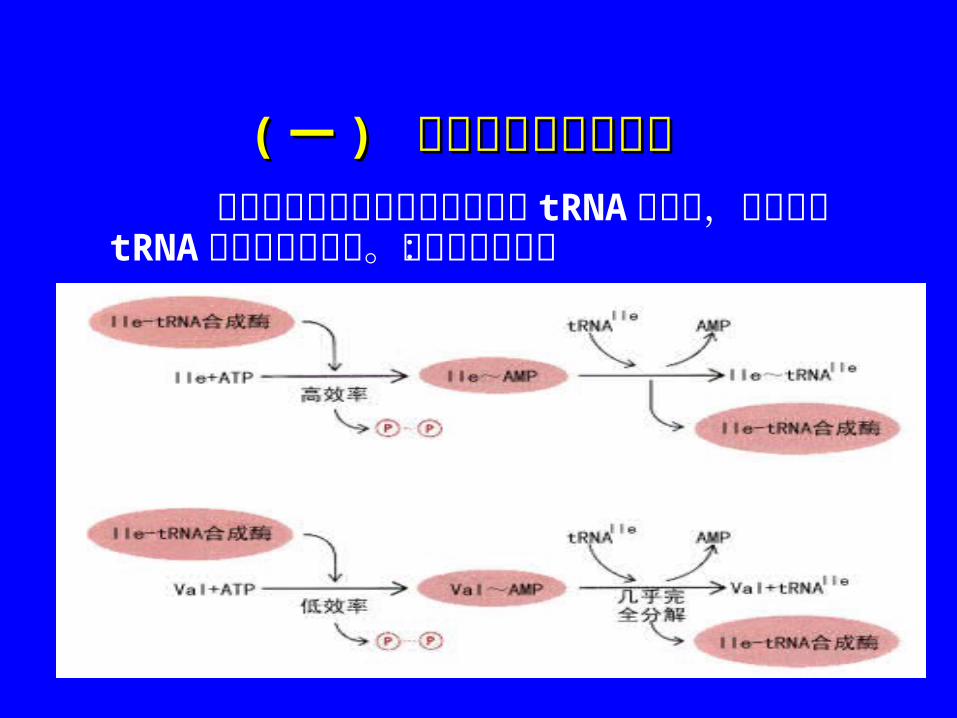
((一一 ) ) 氨基酸的活化与搬运氨基酸的活化与搬运 氨基酸的活化以及活化氨基酸与 tRNA的结合,由氨基酰 tRNA合成酶催化完成。其反应过程为:

在此反应中,特异的 tRNA3’ 端 CCA上的 2’或 3’位自由羟基与相应的活化氨基酸以酯键相连接,形成氨基酰tRNA,从而使活化氨基酸能够被搬运至核蛋白体上参与多肽链的合成。
氨基酸 tRNA 的合成,可使氨基酸活化;搬运;定位。

((二二 ))活化氨基酸的缩合活化氨基酸的缩合 --核蛋白体循环核蛋白体循环
核蛋白体循环:活化氨基酸缩合生成多肽链的过程在核蛋白体上进行。活化氨基酸在核蛋白体上反复翻译mRNA上的密码并缩合生成多肽链的循环反应过程,称为核蛋白体循环
核蛋白体循环过程:分为起始、延伸和终止三个阶段,这三个阶段在原核生物和真核生物类似,现以原核生物中的过程加以介绍。

1.起始( initiation):大致分三个阶段1)30S起始复合物的形成
2)70S起始前复合体的形成
3)70S起始复合体的形成

1)30S起始复合物的形成
﹡借助 16SrRNA近 3’端的 3’-UCCUCC-5’序列与mRNA的 SD序列互补配对 .
fMet-tRNAfmet+IF2+GTP
mRNA﹡
〔 IF1· IF3 · 30S亚基 · mRNA〕
〔 IF1· IF3 · 30S亚基 · fMet-tRNAfmet · mRNA〕
IF1、 IF3 + 70S核糖体
50S亚基
〔 IF1· IF3 · 30S亚基〕

SD序列( Shine-Dalgamo):原核mRNA的起动部位由一段富含嘌呤的特殊核苷酸顺序组成,称为 SD序列(核蛋白体结合位点, RBS),可被核蛋白体小亚基辨认结合。帽子结合蛋白( CBP):真核生物中的mRNA具有帽子结构,已知需一种特殊的帽子
结合蛋白( CBP)以识别此结构。

3) 70S起动复合体的形成
2) 70S起动前复合体的形成
•IF3从 30S起动复合体上脱落,•50S大亚基与复合体结合,形成 70S起动前复合体。•GTP被水解, IF1和 IF2从复合物上脱落。•fMet-tRNAfmet的反密码UAC与mRNA上的起动密码AUG互补结合, tRNAfmet
结合在核蛋白的给位( P 位)。

2. 延伸(elongation)EF-Tu + AA-tRNA + GTP
〔 AA-tRNA · EF-Tu · GTP 〕
进入核糖体的受位( A位)进位
EF-Tu · GDP
A位上的 AA-tRNA 中 AA和
P位的 fMet作用生成肽键成肽
EF-G · GTP
A位上的肽基 -tRNA 移向 P位
GDP
A位上 -tRNA 移向 E位
移位
E位点的 -tRNA 离开核糖体脱落

3.终止阶段 核蛋白体沿mRNA链滑
动,不断使 多肽链延长,直到 终止信号进入受位。
1) 识别: RF识别终止密码,进入核蛋白体的受
位。2) 水解: RF使转肽酶变为水解酶,多肽链与 tRNA之间的酯键被水解,多肽
链释放。3) 解离:通过水解GTP,使核蛋白体与mRNA分离, tRNA、 RF脱落,核蛋白体解离为大、小亚基。

三、多肽链合成后的加工修饰三、多肽链合成后的加工修饰 (一)一级结构的加工修饰
1.N端甲酰蛋氨酸或蛋氨酸的切除 N端甲酰蛋氨酸,必须在多肽链折迭成一定的空间结构之前被切除。
1)去甲酰化:
甲酰蛋氨酸 -肽 甲酸 + 蛋氨酸 -肽甲酰化酶
2)去蛋氨酰基:
蛋氨酰 - 肽 蛋氨酸 + 肽蛋氨酸氨基肽酶

2.氨基酸的修饰: 由专一性的酶催化进行修饰, 包括糖基化、羟基化、磷酸化、甲酰化等。
3.二硫键的形成:
由专一性的氧化酶催化,将 -SH氧化为 -S-S-。4.肽段的切除:
由专一性的蛋白酶催化,将部分肽段切除。

(二)高级结构的形成:1.构象的形成
•在分子内伴侣、辅助酶及分子伴侣的协助下,形成特定的空间构象。
2.亚基的聚合3.辅基的连接分子伴侣( chaperonins):协助肽链折
叠成蛋白质的蛋白质或酶,如HSP等。

(三)靶向输送:
蛋白质合成后,定向地被输送到其执行功能的场所称为靶向输送。大多数情况下,被输送的蛋白质分子需穿过膜性结构,才能到达特定的地点。因此,在这些蛋白质分子的氨基端,一般都带有一段疏水的肽段,称为信号肽 。

常见的信号肽由 10 ~ 40个氨基酸残基组成,N端为带正电荷的氨基酸残基,中间为疏水的核心区,而 C端由小分子氨基酸残基组成,可被信号肽酶识别并裂解。分泌型蛋白质的定向输送,就是靠信号肽与胞浆中的信号肽识别粒子( SRP)识别并特异结合,然后再通过 SRP与膜上的对接蛋白(DP)识别
并结合后,将所携带的蛋白质送出细胞。

分泌型蛋白质的靶向输送

(四)翻译抑制剂
1.四环素:抑制氨酰 -TRNA与核糖体结合。
2.嘌呤霉素:类似氨酰 -TRNA的 3’端,进入A 位, 产生未成熟肽链。
3.氯霉素:能与 50S亚基结合,阻断转肽酶活 性。
4.链霉素:能与 30S小亚基结合,引起读码错误。
5.红霉素:能与 50S亚基结合,阻断移位作用。
6.放綫菌酮:作用与氯霉素类似。





























