Embed Size (px)
DESCRIPTION
第二章 蛋白质的结构与功能 Chapter 2 Structure and Function of Protein. 主讲人:段江燕. 蛋白质 (protein) 是由氨基酸为单位组成的一类重要的生物大分子,是生命的物质基础。. 第一节 蛋白质通论. 蛋白质的组成与分类 蛋白质的大小与分子量 蛋白质的构象 蛋白质的功能. 第二节 蛋白质的组成-氨基酸. 组成蛋白质的基本单位是氨基酸。 天然的蛋白质完全水解,最后都可得到约二十种不同的氨基酸。除 脯氨酸 和 甘氨酸 外,其余均属于 L-α- 氨基酸 。 氨基酸的功能 ( 1 )、组成蛋白质 - PowerPoint PPT Presentation
Citation preview

第二章 蛋白质的结构与功能 第二章 蛋白质的结构与功能Chapter 2 Structure Chapter 2 Structure
and Function of Proteinand Function of Protein
主讲人:段江燕

蛋白质 (protein) 是由氨基酸为单位组成的一类重要的生物大分子,是生命的物质基础。

第一节 蛋白质通论 第一节 蛋白质通论
蛋白质的组成与分类蛋白质的大小与分子量蛋白质的构象蛋白质的功能

第二节 蛋白质的组成-氨基酸 第二节 蛋白质的组成-氨基酸组成蛋白质的基本单位是氨基酸。天然的蛋白质完全水解,最后都可得到约二十种不同的氨基酸。除脯氨酸和甘氨酸外,其余均属于 L-α-氨基酸。
氨基酸的功能( 1)、组成蛋白质( 2)、一些 aa及其衍生物充当化学信号分子 -氨基丁酸,色氨酸衍生物: 5-羟色胺、褪黑激素,都是神经递质,甲状腺素( Tyr 衍生物) ,吲哚乙酸( Trp衍生物)都是激素
( 3)、氨基酸是许多含 N分子的前体物 核酸的含氮碱基、血红素、叶绿素的合成都需要 aa( 4)、一些基本氨基酸和非基本 aa是代谢中间物 精氨酸、瓜氨酸、鸟氨酸 )尿素循环的中间物,

COOH │
H2N — C — H │ R
一、氨基酸一、氨基酸 (amino acid)(amino acid)的结构与通式的结构与通式

基本氨基酸(编码的蛋白质氨基酸):组成蛋白质, 20种,
稀有氨基酸:是多肽合成后由基本 aa经酶促修饰而来。
非蛋白质氨基酸:存在于生物体内但不组 成蛋白质 (约 150种 )。

二、基本氨基酸的分类:二、基本氨基酸的分类:
根据 R侧链基团性质的不同,可将氨基酸进行分类
杂环氨基酸、芳香族氨基酸与脂肪族氨基酸
极性氨基酸与非极性氨基酸

11、、按照按照 RR基的化学结构分类基的化学结构分类 ( ( 11)、)、 RR为脂肪烃基的氨基酸(为脂肪烃基的氨基酸( 55
种)种)

(( 22)、)、 RR中含有羟基和硫的氨基酸(共中含有羟基和硫的氨基酸(共 44种)种)

(( 33)、)、 RR中含有酰胺基团(中含有酰胺基团( 22种)种)

(( 44)、)、 RR中含有酸性基团(中含有酸性基团( 22种)种)

(( 55)、)、 RR中含碱性基团(中含碱性基团( 33种)种)

( 6 )、 芳香族氨基酸
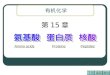
(( 77)、杂环族氨基酸()、杂环族氨基酸( 11种)种)
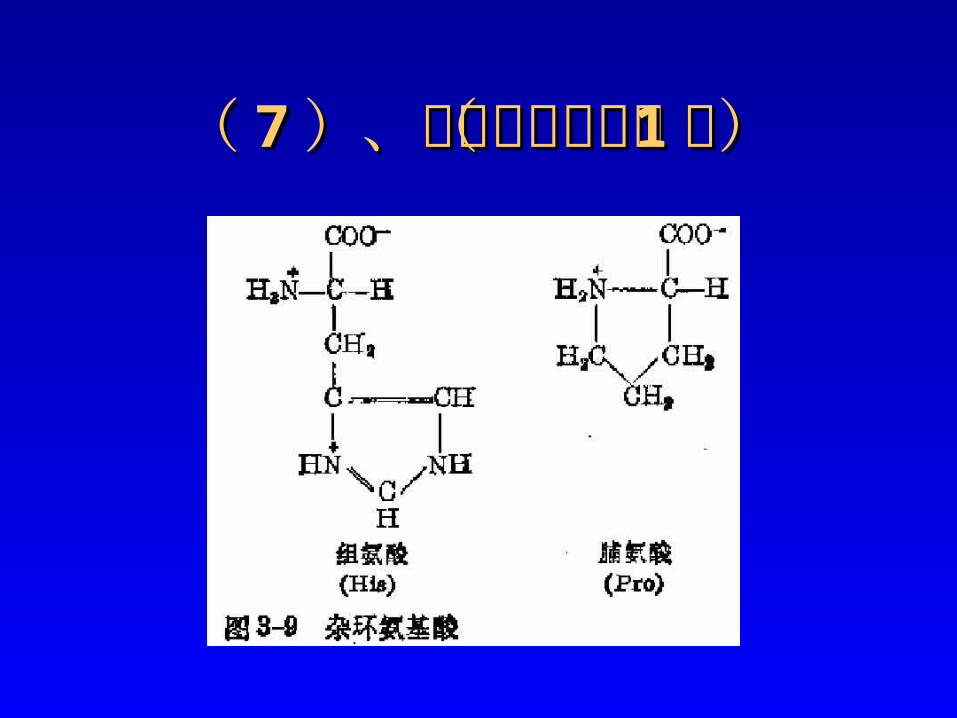
2、极性分类可分为:
a)非 极 性 氨 基 酸 :Ala, Val, Leu, Ile, Pro, Phe,Try,Met。
b)极 性 氨 基 酸 :Gly,Tyr, Cys, Ser, Thr, Asn,Gln; Lys,Arg,His;Asp,Glu.

三、氨基酸的理化性质:三、氨基酸的理化性质:1.两性解离及等电点:(1) 氨基酸在晶体和水溶液中主要以兼性离子形式存在① → 晶体溶点高 离子晶格,不是分子晶格。② →不溶于非极性溶剂 极性分子③ → 介电常数高 水溶液中的氨基酸是极性分子。(2) 氨基酸的两性解离和酸碱滴定曲线 HA → A- + H+ pH=pKa/ +Log[A-]/ [HA] pKa/ 就是 HA50%解离( [碱 ] = [酸 ])时溶液的
pH值, Ka/就是此时溶液的 [H+]当 HA50%解离时,溶液的 pH值等于 pKa/pH = pKa/ 时, [碱 ] = [酸 ]( 50%解离) pH > pKa/ 时, [碱 ] > [酸 ](> 50%解离) pH < pKa/ 时, [酸 ] > [碱 ](< 50%解离)


氨基酸分子是一种两性电解质。通过改变溶液的 pH可使氨基酸分子的
解离状态发生改变。氨基酸分子带有相等正、负电荷时,溶液的 pH值称为该氨基酸的等电点( pI )。

OH- OH-
R—CH—COOH R—CH—COO- R—CH—COO- │ H+ │ H+ │ NH3
+ NH3+ NH2
pH < pI pH = pI pH > pI
2.
2.
2.
3
3
R
R
pKNHpKapIa
pKCOOHpKapIa
NHpKCOOHpKapIa
碱性
酸性
中性

2.紫外吸收性质:
组成天然蛋白质分子的 20 种氨基酸中,只有色氨酸、酪氨酸和苯丙氨酸对紫外光有光吸收。其吸收峰在 280nm左右,以色氨酸吸收最强。
可利用此性质采用紫外分分光度法测定 蛋白质的含量。

氨基酸的光吸收性 Tyr、 Phe、 Trp的 R基含有共轭双键,在 220-300nm近紫外区有吸收。
λ( nm ) εTyr 275 1.4×103Phe 257 2.0×102Trp 280 5.6×103

3.氨基酸的化学反应( 1)氨基参与的反应与亚硝酸反应( Van Slyke 法测定氨基酸)Sanger 反应 : 2.4一二硝基氟苯( DNFB)Edman反应:苯异硫氰酸酯, PITC 生成西佛碱的反应( Schiff):西佛碱是某些酶促反应的中间产物(如转氨基反应的中间产物)。
甲醛反应

(( 22)羧基参与的反应)羧基参与的反应
1、成盐、成酯反应2、成酰氯的反应 氨基被保护后,羧基可与二氯亚砜或
五氯化磷反应,生成酰氯, 使羧基活化,在多肽的人工合成中常用。
3 、叠氮反应 :活化羧基,可用于肽的人工合成
4、脱羧基反应: Glu脱羧形成 γ-氨基丁酸

(( 33)) α-NHα-NH22和和 α-COOHα-COOH共同参加的反共同参加的反
应 应
与茚三酮反应 茚三酮在弱酸中与 α-氨基酸共热,引起氨基酸的氧化脱氨,脱羧反应,最后,
——茚三酮与反应产物 氨和还原茚三酮反应,生成紫色物质( λmax=570nm)。Pro、 Hy-Pro与直接形成黄色化合物
( λmax=440nm)成肽反应

(( 44)侧链)侧链 RR基参加的反应基参加的反应——用于蛋白质的化学修饰——用于蛋白质的化学修饰
TrpArgHisTyrCysPro

三、氨基酸的分离和分析
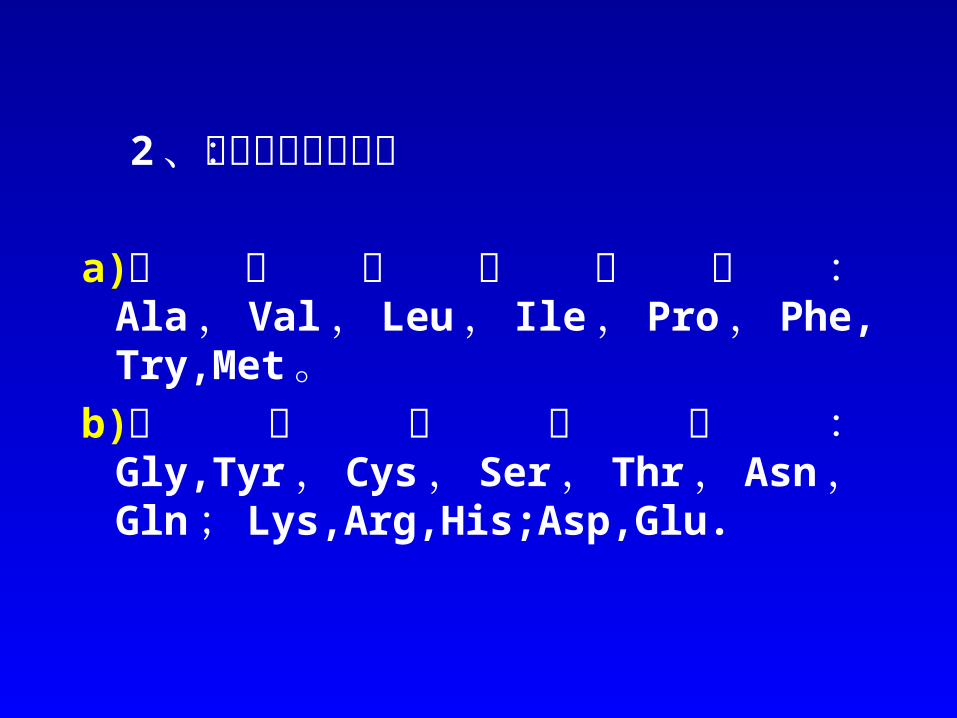
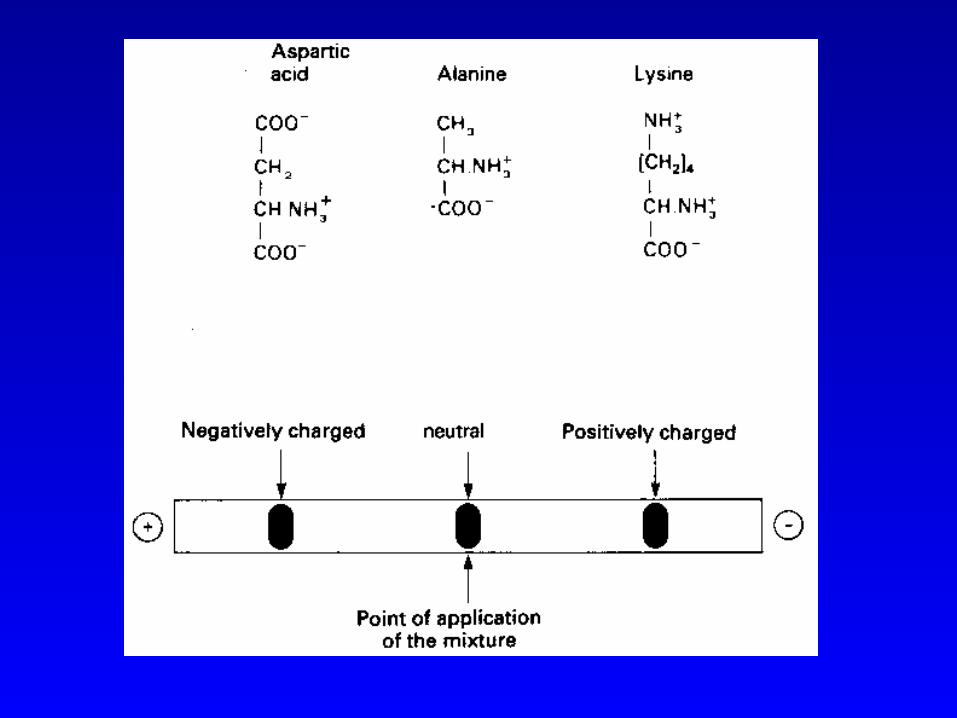
(一)基于电荷性质差异的分离方法1、电泳分离 电泳的基本原理 Glu、 Leu 、 His 、 Lys, 4种混合样,在 pH6.0时,泳动方向及相对速度:
Glu Leu His LyspI 3.22 5.98 7.59 9.74
2、离子交换柱层析

(二)基于分配系数(极性)差异的分离——方法 分配层析
1 、 纸层析2、薄层层析3、分配柱层析

第三节 蛋白质的分子结构 第三节 蛋白质的分子结构
蛋白质是由许多氨基酸单位通过肽键连接起来的,具有特定分子结构的高分子化合物。
蛋白质的分子结构可人为划分为一、二、三、四级结构。除一级结构外,蛋白质的二、三、四级结构均属于空间结构,即构象。
构象是由于有机分子中单键的旋转所形成的。蛋白质的构象通常由非共价键(次级键)来维系。

一、一级结构一、一级结构
(一)氨基酸的连接方式1、肽键2、二硫键3、铰链4、异肽键蛋白质是由若干氨基酸的氨基与羧基经脱水缩合而连接起来形成的长链化合物。一个氨基酸分子的 α-羧基与另一个氨基酸分子的 α-氨基在适当的条件下经脱水缩合即生成肽。

两氨基酸单位之间的酰胺键,称为肽键。多肽链中的氨基酸单位称为氨基酸残基。
多肽链具有方向性,头端为氨基端(N端),尾端为羧基端( C端)。
凡氨基酸残基数目在 50个以上,且具有特定空间结构的肽称蛋白质;凡氨基酸残基数目在 50个以下,且无特定空间结构者称多肽。


谷胱甘肽(GSH):全称为 γ-谷氨酰半胱氨酰甘氨酸。其巯基可氧化、还原,故有还原型(GSH)与氧化型(GSSG )两种存在形式。
Glu-Cys-Gly Glu-Cys-Gly SH S S Glu-Cys-Gly

谷胱甘肽的生理功用:解毒作用:与毒物或药物结合,消除其毒性作用;
参与氧化还原反应:作为重要的还原剂,参与体内多种氧化还原反应;
保护巯基酶的活性:使巯基酶的活性基团 -SH维持还原状态;
维持红细胞膜结构的稳定:消除氧化剂 对红细胞膜结构的破坏作用。

(二)一级结构的测序(二)一级结构的测序

1、 测序的一般步骤( 1) 测定蛋白质分子中多肽链的数目( 2) 拆分蛋白质分子中的多肽链。( 3) 断裂链内二硫键。( 4 ) 分析多肽链的 N末端和 C末端。
( 5) 测定多肽链的氨基酸组成。( 6) 多肽链部分裂解成肽段。( 7) 测定各个肽段的氨基酸顺序( 8) 确定肽段在多肽链中的顺序。( 9) 确定多肽链中二硫键的位置。

2 、 端基分析①N端分析★DNS-cl法:最常用,黄色荧光,灵敏度极高, DNS-多肽水解后的 DNS-氨基酸不需要提取。
★DNFB法: Sanger试剂, DNP-多肽,酸水解,黄色DNP-氨基酸,有机溶剂(乙酸乙酯)抽提分离,纸层析、薄层层析、液相等
★ PITC法: Edman法,逐步切下。无色 PTH-氨基 酸,有机溶剂抽提,层析。

②C端分析★肼解法无水肼 NH2NH2 100 5-10h℃ 。
苯甲醛沉淀氨基酸的酰肼, C端游离氨基酸留在上清中。
Gln、 Asn、 Cys、 Arg不能用此法。★羧肽酶法( Pro不能测)羧肽酶 A:除 Pro、 Arg、 Lys外的所有
C端 a.a
羧肽酶 B:只水解 Arg、 Lys
N H2N… … … … …Val—Ser—Gly C

3、 肽链的部分裂解和肽段的分离纯化
( 1) 、 化学裂解法
① —溴化氰 Met—X— 产率 85%
② —亚碘酰基苯甲酸 Trp—X— 产率 70-100%
③NTCB( 2-硝基 -5- —硫氰苯甲酸) X—Cys—
④羟胺 NH2OH —Asn—Gly—
约 150个氨基酸出现一次

( 2) 、 酶法裂解① 胰蛋白酶 Lys X (X ≠ Pro) Arg—— X ② 胰凝乳蛋白酶 Tyr——X (X ≠ Pro) Trp——X
Phe——X胃蛋白酶 Phe(Trp 、 Try 、 Leu)——Phe(Trp 、 Try 、
Leu)③Glu 蛋白酶 Glu——X
( Vs蛋白酶)④Arg 蛋白酶 Arg——X⑤Lys 蛋白酶 X——Lys⑥Pro 蛋白酶 Pro——X

( 4)二硫键的确定(对角线电泳法)
碘乙酰胺封闭 -SH
胃蛋白酶酶解蛋白质第一向电泳
—过甲酸氧化 S—S—生成 -SO3H
第二向电泳分离出含二硫键的两条短肽,测序与拼接出的肽链比较,定出二硫键的位置。

(三)蛋白质的一级结构与生物功能(三)蛋白质的一级结构与生物功能
★蛋白质的一级结构决定高级结构和功能

★★ 同源蛋白质一级结构的种属差异与生物进化同源蛋白质一级结构的种属差异与生物进化
通过比较同源蛋白质的氨基酸序列的差异可以研究不同物种间的亲源关系和进化。
亲源关系越远,同源蛋白的氨基酸顺序差异就越大。
同源蛋白质:在不同的生物体内行使相同或相似功能的蛋白质。如:血红蛋白在不同的脊椎动物中都具有输送氧气的功能,细胞色素在所有的生物中都是电子传递链的组分。

★★ —蛋白质一级结构的突变 分子病—蛋白质一级结构的突变 分子病
分子病:基因突变引起的某个功能蛋白的某一个或几个氨基酸残基发生了遗传性替代从而导致整个分子的三维结构发生改变,功能部分或全部丧失。
镰刀形红细胞贫血现是由于血红蛋白发生了遗传突变引起的链第 6位的 aa线基由正常的 Glu变成了疏水性的 Val
★一级结构的部分切除与蛋白质的激活

二、二级结构二、二级结构(一)、蛋白质分子中的非共价键(一)、蛋白质分子中的非共价键 ((次级次级
键键 ))1. 氢键:氢键 (hydrogen bond)的形成常见于连接在一电负性很强的原子上的氢原子,与另一电负性很强的原子之间。
氢键在维系蛋白质的空间结构稳定上起着重要的作用。
氢键的键能较低 (~12kJ/mol),因而易被破坏。

蛋白质分子中氢键的形成

2. 疏水键:
非极性物质在含水的极性环境中存在时,会产生一种相互聚集的力,这种力称为疏水键或疏水作用力。
蛋白质分子中的许多氨基酸残基侧链也是非极性的,这些非极性的基团在水中也可相 互 聚 集 , 形 成 疏 水 键 , 如Leu, Ile, Val, Phe, Ala等的侧链基团。

3. 离子键(盐键):
离子键 (salt bond)是由带正电荷基团与带负电荷基团之间相互吸引而形成的化学键。
在近中性环境中,蛋白质分子中的酸性氨基酸残基侧链电离后带负电荷,而碱性氨基酸残基侧链电离后带正电荷,二者之间可形成离子键。

蛋白质分子中离子键的形成

4.范德华氏( van der Waals)引力:原子之间存在的相互作用力。

蛋白质的二级结构是指蛋白质多肽链主链
原子局部的空间结构,但不包括与其他肽
段的相互关系及侧链构象的内容。维系蛋
白质二级结构的主要化学键是氢键。

酰胺平面(肽平面)酰胺平面(肽平面)
1. 由于 C=O双键中的 π电子云与 N“原子上的未共用电子对发生 电子共
”振 ,使肽键具有部分双键的性质,不能自由旋转。
2.与肽键相连的六个原子构成刚性平面结构,称为肽单元或肽键平面。但由于α-碳原子与其他原子之间均形成单键,因此两相邻的肽键平面可以作相对旋转。


——肽键平面 由于肽键具有部分双
键的性质,使参与肽键构成的六个
原子被束缚在同一平面上,这一平
面称为肽键平面或肽单元。


★肽键的结构特点:( 1)酰胺氮上的孤对电子与相邻羰基之间形成共振杂化体。( 2)肽键具有部分双键性质,不能自由旋转。( 3)肽键具有平面性,组成肽键的 4个原子和 2个 Cα几乎处在同一平面内(酰氨平面)。( 4)肽键亚氨基在 pH0---14内不解离。( 5)肽链中的肽键一般是反式构型,而 Pro的肽键可能出现顺、反两种构型.

★多肽链可以看成由 Cα串联起来的无数个酰胺平面组成

★当 Φ(Ψ)旋转键两侧的主链呈顺式时,规定
Φ(Ψ) =0°
★从 Cα沿键轴方向看,顺时针旋转的 Φ
(Ψ)角为正值,反之为负值。

(二)蛋白质二级结构的类型:(二)蛋白质二级结构的类型:
蛋白质的二级结构主要包括 α- 螺
旋, β-折迭, β-转角及无规卷曲
等几种类型。

1. 1. α-α-螺旋螺旋::

α-螺旋是多肽链的主链原子沿一中心轴盘绕所形
成的有规律的螺旋构象,其结构特征为:
① 二面角:Φ= -57°, Ψ= - 48°,是一种右手螺旋② 每圈螺旋: 3.6个 a.a 残基, 高度: 0.54nm
③ 每个残基绕轴旋转 100°,沿轴上升 0.15nm
④ 氨基酸残基侧链向外⑤ 相邻螺圈之间形成链内氢链,氢键的取向几乎与中心轴平行。
⑥ 肽键上 C=O氧与它后面( C端)第四个残基上的N-H氢间形成氢键。

典型的 α螺旋用 3.613表示,3.6表示每圈螺旋包括 3.6个残基13表示氢键封闭的环包括 13个原子。 封闭环原子数 =3n+4( n=1、 2、 .....)( n:后边第 n个 aa)
2.27螺旋( n=1)310 螺旋( n=2)613螺旋( n=3)4.316螺旋( n=4)
2.27 310 3.613 4.316
n=1 n=2 n=3 n=4

影响 α-螺旋稳定的因素有:
⑴ 极大的侧链基团(存在空间位阻);
⑵ 连续存在的侧链带有相同电荷的氨基酸残基(同种电荷的互斥效应);
⑶ 有 Pro 等亚氨基酸存在(不能形 成氢键)。

2. β-2. β-折迭:折迭:

β-折迭是由若干肽段或肽链排列起来所形成的扇面状片层构象,其结构特征为:
⑴ 由若干条肽段或肽链平行或反平行排列组成片状结构;
⑵ 主链骨架伸展呈锯齿状;
⑶ 借相邻主链之间的氢键维系。

β-折迭包括平行式和反平行式两种类型

3. β-3. β-转角:转角:

β-转角是多肽链 180°回折部分所形成的一种二级结构,其结构特征为:
⑴ 主链骨架本身以大约 180°回折 ;
⑵ 回折部分通常由四个氨基酸残基构成 ;
⑶构象依靠第一残基的 -CO基与第四残基的 -NH 基之间形成氢键来维系。

4. 4. 无规卷曲:无规卷曲:
无规卷曲是指多肽链主链部分形成的
无规律的卷曲构象。

RNase的分子结构
α-螺旋
β-折迭
β-转角
无规卷曲

三、超二级结构 由若干个相邻的二级结构单元( α-
螺旋、 β-折叠、 β-转角及无规卷曲)组
合在一起,彼此相互作用,形成有规则
的、在空间上能够辨认的二级结构组合
体。

1、复绕 α-螺旋( αα 结构 )
22 、 、 ββxxββ结构结构33、、 ββ曲折(曲折( ββ--
meandermeander ) )44 、 、 回形拓扑结回形拓扑结
构(希腊钥匙) 构(希腊钥匙)55 、 、 ββ-- 折叠桶 折叠桶66 、 、 αα--螺旋螺旋 --ββ转角转角 --αα--螺旋螺旋

锌指结构 锌指结构 ((螺旋螺旋 --折迭折迭 --折迭模序折迭模序 ))

转录因子转录因子MyoDMyoD的螺旋的螺旋 --环环 --螺旋螺旋模序模序

四、四、 结构域 结构域domaindomain,, motif(motif(模块模块 ))
在二级结构及超二级结构的基础上,多肽链进一步卷曲折叠,组装成几个相对独立的、近似球形的三维实体。

结构域是球状蛋白的折叠单位
较小的蛋白质分子或亚基往往是单结构域的。
结构域一般有100-200氨基酸残基。
结构域之间常常有一段柔性的肽段相连,形成所谓的铰链区,使结构域之间可以发生相对移动。
结构域承担一定的生物学功能,几个结构域协同作用,可体现出蛋白质的总体功能。例如,脱氢酶类的多肽主链有两个结构域,一个为 NAD+结合结构域,一个是起催化作用的结构域,两者组合成脱氢酶的脱氢功能区。
结构域间的裂缝,常是酶的活性部位,也是反应物的出入口


★EF手:钙结合蛋白中,含有 Helix-Loop-Lelix结构
★锌指: DNA结合蛋白中, 2个His、 2个 Cys结合一个 Zn
★亮氨酸拉链: DNA结合蛋白中,由亮氨酸倒链形成的拉链式结构

五、蛋白质的三级结构五、蛋白质的三级结构
1、三级结构: 整个多肽链在二级结构、超二级结构和结构域的基础上盘旋、折叠,形成的特定的整个空间结构。
维系三级结构的化学键主要是非共价键(次级键),如疏水键、氢键、盐键、范氏引力
等,但也有共价键,如二硫键等。

22、三级结构有以下特点:、三级结构有以下特点:
许多在一级结构上相差很远的 aa碱基在三级结构上相距很近。
球形蛋白的三级结构很密实,大部分的水分子从球形蛋白的核心中被排出,这使得极性基团间以及非极性基团间的相互用成为可能。
大的球形蛋白 (200aa以上 ),常常含有几个结构域,

33、球状蛋白质三维结构特征、球状蛋白质三维结构特征
1、含有丰富的二级结构元件2、具有明显的折叠层次3、分子呈现球状或椭球状4、疏水侧链埋藏在分子内部,亲水侧链暴露在外表
5、表面常常有空穴,配体结合部位(活性中心)

44、肌红蛋白的结构与功能:、肌红蛋白的结构与功能:
( 1 ) 肌红蛋白的三级结构哺乳动物肌肉中储氧的蛋白质。由一条多肽链(珠蛋白)和一个血红素辅基组成。球状分子,单结构域。 8段直的 α-螺旋组成,分别命名为 A、 B、 C…H,拐弯处是由 1~8个氨基酸组成的松散肽段(无规卷曲)。4 个 Pro 残 基 各 自 处 在 一 个 拐 弯 处 , 另 外 4 个 是
Ser、 Thr、 Asn、 Ile。
血红素辅基 ,扁平状,结合在肌红蛋白表面的一个洞穴内。


(( 22 )、 )、 肌红蛋白的氧合曲线肌红蛋白的氧合曲线

胰岛素分子的三级结构胰岛素分子的三级结构

溶菌酶分子的三级结构溶菌酶分子的三级结构

磷酸丙糖异构酶和丙酮酸激酶的三级结构磷酸丙糖异构酶和丙酮酸激酶的三级结构


六、蛋白质的四级结构六、蛋白质的四级结构
蛋白质的四级结构( quaternary structure) 就是指蛋白质分子中亚基的立体排布,亚基间的相互作用与接触部位的布局。
亚基 (subunit) 就是指参与构成蛋白质四级结构的、每条具有三级结构的多肽链。
维系蛋白质四级结构的是氢键、盐键、范氏引力、疏水键等非共价键。

四级缔合在结构和功能上的优越性四级缔合在结构和功能上的优越性
1、降低比表面积,增加蛋白质的稳定性2、丰富蛋白质的结构,以行使更复杂的功能。
3、提高基因编码的效率和经济性。4、使酶的催化基团汇集,提高催化效率。5、形成一定的几何形状,如细菌鞭毛。6、适当降低溶液渗透压。7、具有协同效应和别构效应,实现对酶活性的调节

亚基的立体排布亚基的立体排布

血红蛋白的结构与功能血红蛋白的结构与功能
1 、 血红蛋白的结构: 成人: HbA : α2β2 98% ,
HbA2: α2δ2 2% 胎儿: HbF α2γ2
早期胚胎: α2ε2
▲接近于球体, 4个亚基分别在四面体的四个角上,每个亚基上有一个血红素辅基。
▲α、 β链的三级结构与肌红蛋白的很相似,

血红蛋白血红蛋白 (hemoglobin)(hemoglobin)的四级结构的四级结构


当血红蛋白的一个 α亚基与氧分子结合以后,可引起其他亚基的构象发生改变,对氧的亲和力增加,从而导致整个分子的氧结合力迅速增高,使血红蛋白的氧饱和曲线
“呈 S”形。这种由于蛋白质分子构象改变而导致蛋白质分子功能发生改变的现象称为变构效应。

血红蛋白的变构血红蛋白的变构

★ 别构蛋白:除了有活性部位(结合底物)外,还有别构部位(结合调节物)。有时活性部位和别构部位分属不同的亚基(活性亚基和调节亚基),活性部位之间以及活性部位和调节部位之间通过蛋白质构象的变化而相互作用。
★ 别构效应:别构蛋白的别构部位与效应物的结合改变了蛋白质的构象,从而对活性部位产生的影响。
★同促效应(同位效应):一种配体的的结合对于其它部位结合同种配体的能力的影响,包括(正)协同效应和负协同效应。
★异促效应(异位效应):就是别构效应,别构部位与效应物的结合对活性部位的影响。
★ 正协同效性:一种配基的结合促进后续配基的结合, S型配体结合曲线。
★负协同效性:一种配基的结合抑制促进后续配基的结合。★正效应物:促进活性部位与配基结合的别构效应物。★负效应物:抑制活性部位与配基结合的别构效应物。

22、血红蛋白的氧饱和曲线、血红蛋白的氧饱和曲线

33 、 、 波耳效应及其生理意义波耳效应及其生理意义
血红蛋白上有 CO2和 BPG 结合部位,因此,血红蛋白还能运输 CO2 。
波耳效应:增加 CO2的浓度、降低 pH 能显著提高血红蛋白亚基间的协同效应,降低血红蛋白对 O2的亲和力,促进 O2的释放,反之,高浓度的 O2也能促使血红蛋白释放 H+ 和 CO2 。

44 、 、 BPGBPG的别构效应的别构效应
BPG是血红蛋白的负效应物。BPG( 2,3-二磷酸甘油酸)通过与它的两个 β亚基形成盐键稳定了血红蛋白的脱氧态的构象,因而降低脱氧血红蛋白的氧亲和力。
无 BPG时, P50=1 torr,BPG=4.5mM时, P50=26 torr
BPG进一步提高了血红蛋白的输氧效率。在肺部, PO2超过 100torr,因此,即使没有 BPG,血红蛋白也能被饱和,在组织中, PO2低, BPG降低血红蛋白的氧亲和力,加大血红蛋白的卸氧量。
( 1)高山适应和肺气肿的生理补偿变化; BPG升高。( 2)血库储血时加入肌苷可 BPG。

第四节 蛋白质的理化性质 第四节 蛋白质的理化性质
(一)蛋白质的两性解离与等电点:由于蛋白质分子中氨基酸残基的侧链上存在游离的氨基和游离的羧基,因此蛋白质与氨基酸一样具有两性解离的性质,因而也具有特定的等电点。

(二)蛋白质的胶体性质:(二)蛋白质的胶体性质:
蛋白质分子的颗粒直径已达1~100nm,处于胶体颗粒的范围。因此,蛋白质具有亲水溶胶的性质。
蛋白质分子表面的水化膜和表面电荷是稳定蛋白质亲水溶胶的两个重要因
素。

(三)蛋白质的变性、沉淀和凝固:(三)蛋白质的变性、沉淀和凝固:
在某些物理或化学因素的作用下,蛋白质严格的空间结构被破坏(不包括肽键的断裂),从而引起蛋白质若干理化性质和生物学性质的改变,称为 蛋 白 质 的 变 性(denaturation)。

引起蛋白质变性的因素有:
① 物理因素:高温、高压、紫外线、电离辐射、超声波等;
② 化学因素:强酸、强碱、有机溶剂、重金属盐等。

变性蛋白质的性质改变:① 物理性质:旋光性改变,溶解度下降,沉降率升高,粘度升高,光吸收度增加等;
② 化学性质:官能团反应性增加,易被蛋白酶水解。
③ 生物学性质:原有生物学活性丧失,抗原性改变。

蛋白质变性的可逆性:a)蛋白质在体外变性后,绝大多数情况下是不能复性的;
b)如变性程度浅,蛋白质分子的构象未被严重破坏;或者蛋白质具有特殊的分子结构,并经特殊处理则可以复性。

核糖核酸酶的变性与复性核糖核酸酶的变性与复性

蛋白质分子相互聚集而从溶液中析出的现象称为沉淀。
变性后的蛋白质由于疏水基团的暴露而易于沉淀,但沉淀的蛋白质不一定都是变性后的蛋白质。
加热使蛋白质变性并凝聚成块状称为凝固。因此,凡凝固的蛋白质一定
发生变性。

第五节、蛋白质的分离与纯化第五节、蛋白质的分离与纯化
(一)盐析与有机溶剂沉淀:1. 盐析:在蛋白质溶液中加入大量中性盐,以破坏蛋白质的胶体性质,使蛋白质从溶液中沉淀析出,称为盐析。

常用的中性盐有:硫酸铵、氯化钠、硫酸钠等。
盐析时,溶液的 pH在蛋白质的等电点处效果最好。
盐析沉淀蛋白质时,通常不会引起蛋白质的变性。

分段盐析: 半饱和硫酸铵溶液可沉淀血浆球蛋白,而饱和硫酸铵溶液可沉淀血浆清蛋白。

2. 有机溶剂沉淀蛋白质:
凡能与水以任意比例混合的有机溶剂,如乙醇、甲醇、丙酮等,均可用于沉淀蛋白质。
① ② 沉淀原理是: 脱水作用; 使水的介电常数降低,蛋白质溶解度降低。

(二)电泳:(二)电泳:
带电粒子在电场中移动的现象称为电泳( electrophoresis) 。
蛋白质分子在溶液中可带净的负电荷或带净的正电荷,故可在电场中发生移动。不同的蛋白质分子所带电荷量不同,且分子大小也不同,故在电场中的移动速度也不同,据此可互相分离。

(三)层析:(三)层析:
层析( chromatography) 是一种利用混合物中各组分理化性质的差异,在相互接触的两相(固定相与流动相)之间的分布不同而进行分离分析的技术方法。
主要的层析技术有离子交换层析,凝胶层
析,吸附层析及亲和层析等。

凝胶过滤分离蛋白质凝胶过滤分离蛋白质

(四)超速离心:(四)超速离心:
利用物质密度的不同,经超速离心后,分布于不同的液层而分离。
超速离心也可用来测定蛋白质的分子量,蛋白质的分子量与其沉降系数S 成正比。

三、氨基酸顺序分析三、氨基酸顺序分析
蛋白质多肽链的氨基酸顺序分析,即蛋白质一级结构的测定,主要包括以下几个步骤:
1. 分离纯化蛋白质,得到一定量的蛋白质纯品。
2. 取一定量的样品进行完全水解,再用氨基酸自动分析仪,测定蛋白质的氨基酸
组成。

蛋白质的氨基酸组成分析蛋白质的氨基酸组成分析

3. 分析蛋白质的 N-端和 C-端氨基酸。
通 常 采 用 2,4- 二 硝 基 氟 苯( DNFB)法和肼解法分别确定蛋白质的N-端和 C-端氨基酸残基。

DNFBDNFB法分析法分析 N-N-端氨基酸残基端氨基酸残基

肼解法分析肼解法分析 C-C-端氨基酸残基端氨基酸残基

4. 采用特异性的酶(如胰蛋白酶)或化学试剂(如溴化氰)将蛋白质裂解成为长短不一的若干条肽段(必须作两套)。

胰蛋白酶(胰蛋白酶( trypsin)trypsin)的裂解作用的裂解作用

BrCNBrCN的裂解作用的裂解作用

5. 5. 分离纯化单一肽段。分离纯化单一肽段。
6. 6. 使用氨基酸顺序测定仪,测定各使用氨基酸顺序测定仪,测定各
条肽段的氨基酸顺序。一般采用条肽段的氨基酸顺序。一般采用
EdmanEdman降解法降解法,用异硫氰酸苯酯进,用异硫氰酸苯酯进
行反应,将氨基酸降解后,逐一进行行反应,将氨基酸降解后,逐一进行
测定。测定。

EdmanEdman降解法测定氨基酸序列降解法测定氨基酸序列
异硫氰酸
苯酯

氨基酸自动测序仪氨基酸自动测序仪

7. 将两套不同肽段的氨基酸顺序进行比较,以获得完整的蛋白质分子的氨基酸顺序。