Embed Size (px)
Citation preview

E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
Ava i l ab l e on l i ne a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /yexc r
Research Article
α-Syntrophin is required for the hepatocyte growthfactor-induced migration of cultured myoblasts
Min Jeong Kima, Stanley C. Froehnerb, Marvin E. Adamsb,⁎, Hye Sun Kima,⁎⁎aDepartment of Biological Science, Ajou University, Suwon 443-749, Republic of KoreabDepartment of Physiology & Biophysics, University of Washington, Seattle, Washington 98195, USA
A R T I C L E I N F O R M A T I O N
⁎ Corresponding author. Fax: +1 206 685 061⁎⁎ Corresponding author. Fax: +82 31 219 161
E-mail addresses: [email protected] (M.E. AdaAbbreviations: PH, pleckstrin homology; SU,
AB, α; β2, syntrophin double knockout; FLA, traified Eagle's medium; DAPI, 4′,6-diamidino-2-p
0014-4827/$ – see front matter © 2011 Elseviedoi:10.1016/j.yexcr.2011.09.017
A B S T R A C T
Article Chronology:
Received 17 May 2011Revised version received28 September 2011Accepted 29 September 2011Available online 6 October 2011
Syntrophins are adaptor proteins that link intracellular signaling molecules to the dystrophin basedscaffold. In this study, we investigated the function of syntrophins in cell migration, one of the earlysteps in myogenic differentiation and in regeneration of adult muscle. Hepatocyte growth factor(HGF) stimulates migration and lamellipodia formation in cultured C2 myoblasts. In the migratingcells, syntrophin concentrated in the rear-lateral region of the cell, opposite of the lamellipodia,instead of being diffusely present throughout the cytoplasm of non-migrating cells. When theexpression of α-syntrophin, the major syntrophin isoform of skeletal muscle, was reduced by trans-fection with the α-syntrophin-specific siRNA, HGF stimulation of lamellipodia formation was pre-vented. Likewise, migration of myoblasts from α-syntrophin knockout mice could not be
stimulated by HGF. However, HGF-induced migration was restored in myoblasts isolated from atransgenic mouse expressing α-syntrophin only in muscle cells. Treatment of C2 myoblasts withinhibitors of PI3-kinase not only reduced the rate of cell migration, but also impaired the accumula-tion of syntrophins in the rear-lateral region of the migrating cells. Phosphorylation of Akt wasreduced in the α-syntrophin siRNA-treated C2 cells. These results suggest that α-syntrophin is re-quired for HGF-induced migration of myoblasts and for proper PI3-kinase/Akt signaling.
© 2011 Elsevier Inc. All rights reserved.
Keywords:
α-SyntrophinMyoblast migrationHepatocyte growth factor
α-Syntrophin-specific siRNAα-Syntrophin knock-out mousePI3-kinase/Akt signaling
Introduction
Cell migration is one of the essential steps in muscle regenera-tion in adult myofibers as well as in embryonic development.The satellite cells, myogenic stem cells in adult skeletal muscle,can be activated to proliferate in response to various stimulisuch as injury and exercise [1,2]. The activated cells migrate tothe injured sites by chemotaxis and start to proliferate. Hepato-cyte growth factor (HGF) is one of the chemo-attractants
9.5.ms), [email protected] (syntrophin unique; HGF, hnsgenic mouse expressinghenylindole; PI, propidium
r Inc. All rights reserved.
released in response to muscle injury [1,3]. In addition, HGF fa-cilitates both non-directional and directional migration of cul-tured C2C12 myoblasts [4]. In response to the stimulus tomigrate, the actin-cytoskeleton is reorganized and lamellipodiaform in the direction of the leading edge of migrating cells.Other proteins, including PTEN, accumulate at the rear of themigrating cell or trailing edge, opposite of the lamellipodia [5].Inhibitors of PI3-kinase, LY294002 and Wortmannin, preventHGF-induced lamellipodial formation and cell migration [4,6].
H.S. Kim).epatocyte growth factor; C57, C57bl6/J; αKO, α-syntrophin knockout;full length α-syntrophin only in muscle cells; DMEM, Dulbecco's mod-iodide.

2915E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
The syntrophins are a family of scaffolding proteins thatlink signaling proteins to the dystrophin protein complex [7].They interact with multiple signaling proteins via two pleck-strin homology (PH1) domains, a PDZ domain, and a conservedsyntrophin unique (SU) domain [8–10]. Both the SU domainand second PH domain are required for binding to dystrophinand other members of the dystrophin protein family (utrophin,dystrobrevins, and DRP2) [10]. The PDZ domain of syntrophinscan bind to a variety of signaling proteins, including kinases,various channels, and nitric oxide synthase [11–17]. Syntro-phins also have been reported to associate with actin, calmod-ulin, and phosphatidylinositols [18]. There are five differentsyntrophin isoforms (α, β1, β2, γ1, and γ2). Four isoforms(all except γ1) are expressed in skeletal muscle, but α-syntrophin is the predominant isoform in this tissue [8].
Studies on syntrophin function have largely focused on ma-ture myofibers and the neuromuscular junction. In the culturedmyoblasts, however, α-syntrophin is expressed from the earlystages of differentiation when dystrophin, a major binding pro-tein of syntrophin, has not yet been expressed [19]. Little isknown about the function of syntrophin in the early stages ofmuscle development. Recently, we have shown that α-syntrophin is involved in the expression of myogenin, a key myo-genic regulatory factor, by interaction with MLL5 in the earlystages of myogenic differentiation [20]. In this study, we have fo-cused on the intracellular localization of syntrophin, particularlyin the migrating muscle cells. We investigated the changes in in-tracellular localization of syntrophin following HGF stimulation.By using cultured C2 myoblasts and primary myoblast culturesfrom α-syntrophin knockout mice, we investigated the functionof syntrophin in promoting cell migration in response to HGF.Our results demonstrate that α-syntrophin is required for theHGF-induced migration of cultured myoblasts.
Materials and methods
Materials
HGF was purchased from Peprotech (Rocky Hill, NJ). Anti-PTENand anti-actin antibodies, control siRNA, α-syntrophin-specificsiRNA, and protein A/G were from Santa Cruz Biotechnology(Santa Cruz, CA). Anti-p85, anti-Met, anti-pMet (Y1234), andanti-pMet (Y1349) antibodies were from Cell Signaling (Dan-vers, PA). LY294002, Wortmannin and collagen type I wereobtained from Sigma (St. Louis, MO). Lipofectamine 2000,rhodamine-phalloidin, propidium iodide (PI), 4′,6-diamidino-2-phenylindole (DAPI), fluorescein isothiocyanate (FITC)-conjugated, and tetramethyl rhodamine isothiocyanate (TRITC)-conjugated antibodies were from Invitrogen (Carlsbad, CA).pGFPN3 was from Clontech (Mountain View, CA). HindIII andBamHI were from Fermentas (Hanover, MD). Anti-pan-syntrophinand anti-α-syntrophin antibodieswere used as previously described[21]. All of the reagents for primary cell culture were also fromInvitrogen. Strainers, 24 well plates, and cell culture inserts forthree dimensional chamber assay were from BD Falcon (SanJose, CA). The polyvinylidenedifluoride (PVDF) membrane wasfrom Millipore (Billerica, MA). Bradford assay kit was from Bio-Rad(Hercules, CA).
Primary culture of skeletal muscle cells
Primary culture of the skeletal muscle was performed as previous-ly described [20]. In brief, skeletal muscle cells were isolated fromthe forelimbs and hindlimbs of neonatal mice (0–1 day old) fromC57BL6/J wild-type (C57), α-syntrophin-knockout (αKO) [22],α-syntrophin transgenic (FLA) [23], and α/β2-syntrophin-doubleknockout mice (AB) [24]. Skeletal muscles were dissected free ofother tissues including skin, lipid, and bone then minced with for-ceps. Single muscle cells were isolated with treatment of 0.05%trypsin EDTA in phosphate buffered saline (PBS) for 45 min andpassed through a 70 μm strainer. Cells were pre-plated on a100 mm culture dish to remove fibroblasts and to enrich for mus-cle cells twice for 1 h each. Experimental procedures performed onmice were approved by the Institutional Animal Care and UseCommittee of the University of Washington.
Three-dimensional migration chamber assay
A three-dimensional migration chamber assaywas performed using atissue culture insert (8.0 μm pore size) in 24-well plates. The uppersurface of the membrane was coated with collagen type I and300 μl of cell suspension (5.0×104 cells/ml) was applied to the coat-ed membrane. The lower chamber of the insert contained serum freemedium (200 μl) with or without HGF (50 ng/ml). After incubation at37 °C for 1 h, the cell suspension was removed from the chamber andthe surface of the lower chamber was fixed with ice-cold methanol.To recognize moving cells, the lower surface was stained with a he-matoxylin solution and cells in the upper chamber were removedwith a cotton tip. The hematoxylin-stained cells in the lower surfaceof the membrane were counted using an inverted microscope. Eachexperiment was independently carried out four times.
HGF-induced migration assay
C2 myoblasts were seeded on 6-well tissue culture plates with1.0×104 cells/ml. Cells were grown in 10% fetal bovine serum(FBS) containing Dulbecco's modified Eagle's medium (DMEM)for 6 h. Cells were serum-starved by incubating in 0.5% FBS con-taining DMEM for 3 h. Cells were incubated with HGF (50 ng/ml)for 1 h and then fixed with 4% paraformaldehyde. To visualizethe cytoskeleton, F-actin was stained with rhodamine-phalloidin.Migrating cells with lamellipodia were quantified by visual analy-sis on at least 10 randomly selected fields, such that >300 cellswere counted per field.
Immunofluorescence assay
Cells were seeded on a cover-slip in 6-well plates (1.0×104cells/ml) and grown in a 5% CO2 humidified incubator for 6 h. Cells trea-ted with or without HGF were fixed with 4% paraformaldehyde inPBS for 10 min and permeabilized with 0.5% Triton X-100 in PBS.Cells were blocked with 10% normal goat serum in PBS for 10 min.All of the primary antibodies were incubated for 1 h and TRITC- orFITC-conjugated secondary antibodies were incubated for 40 minat room temperature. All of the primary antibodies were diluted1:50 and secondary antibodies were diluted 1:100. Both DAPI andPI were used to stain nuclei. Images were taken with a ZeissLSM510 confocal laser scanning microscope or Zeiss Axioskop 40FL microscope.

2916 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
siRNA transfection
Transfection of α-syntrophin-specific siRNA was performed as de-scribed previously [20]. In brief, C2 myoblasts were grown in 24-well plates at a density of 5.0×104 cells/ml for 24 h. Cells weretransfected with control siRNA or α-syntrophin-specific siRNAby incubation with lipofectamine 2000 for 6 h. The cells wereallowed to recover from the transfection by transferring intofresh 10% FBS-containing medium and incubated for 18 h. Subse-quently, cells were treated with HGF (50 ng/ml) and incubatedfor 1 h.
Transfection of GFP fusion α-syntrophin
To determine the localization of exogenous α-syntrophin, the full-length α-syntrophin cDNA [8] was cloned into the HindIII and BamHIsite of pGFPN3. For transfection of GFP fusion α-syntrophin, C2 cellswere grown in 6-well plates (5.0×104 cells/ml) for 24 h in anantibiotics-free DMEM and transfected with GFP fusion α-syntrophinby incubationwith lipofectamine 2000 for 6 h. Cellswere allowed to re-cover from the transfection by transferring in 10%FBS-containingmedi-um for 18 h. Subsequently, HGF (50 ng/ml) was added into 0.5% FBS-containing medium and incubated for 1 h.
Immunoprecipitation assay
At the indicated culture time, cellswere rinsedwith cold PBS, harvestedwith a scraper, and disrupted by ultrasonication in RIPA buffer, pH 7.4,containing 9.1 mMNa2HPO4, 1.7 mMNaH2PO4, 150mMNaCl, 0.5% so-dium deoxycholate, 0.1% SDS, 1 mM PMSF, and 0.1% protease inhibitorcocktail. Cell lysates were centrifuged at 15,000×g for 3min to removecell debris and then protein concentrationwas determined by Bradfordassay. For pre-clearance, cell extracts (500 μg/500 μl) were incubatedwith 20 μl of proteinA/G for 30minon ice. The proteinswere incubatedwith anti-PTEN or anti-syntrophin antibodies overnight at 4 °C. ProteinA/G (20 μl) was added and incubated for 1 h at 4 °C. The immune-complexes were collected by centrifugation and washed with coldPBS for 3 times, then the proteins were separated with SDS-polyacrylamide gel electrophoresis (SDS-PAGE).
Immunoblot assay
To determine protein expression, cells were rinsed twice with coldPBS and mixed with SDS-sample buffer (1.0 M Tris/HCl, pH 6.8,containing 10% glycerol, 2% SDS, 0.025% bromo-phenol blue, and5% β-mercaptoethanol) and boiled in 100 °C for 5 min. Equalamounts of protein were separated by SDS-PAGE and transferredonto PVDF membrane. The membranes were pre-blocked with5% bovine serum albumin (BSA) and incubated with the indicatedprimary antibodies. After incubation with peroxidase-conjugatedsecondary antibodies, the immunoreactive protein bands were vi-sualized by enhanced chemiluminescence detection with DigitalLuminescent Image Analyzer LAS-1000 (Fuji film, Japan). Band in-tensity was determined by Scion image (Fredrick, MD).
Statistical analysis
Results are presented as mean±S.E.M. For the statistical analysisof cell migration, two tailed Student's unpaired t test was per-formed. A value of p<0.05 was considered to be significant.
Results
Syntrophin specifically concentrates at the trailing edge ofmigrating C2 myoblasts
HGF has been reported to regulate cell migration in a variety ofcell types [4,6,25,26]. We examined whether HGF can induce mi-gration of C2 cells under our culture conditions. Myoblasts cul-tured on a cover-slip with HGF were stained with rhodamine-phalloidin which detects actin stress fibers. The cytoskeleton hasto rearrange for morphological change in migrating cells [27]and the lamellipodia are formed in the direction of the movingcells [28,29]. As shown in Fig. 1A, typical morphology of migratingcells with lamellipodia (arrows) were found following HGF incu-bation. To examine how many cells respond to HGF under thesame conditions, cells were exposed to HGF for the indicated pe-riods in a low serum media (Fig. 1B). The cells with lamellipodiawere counted in randomly chosen fields. Just before HGF treat-ment (0 min), approximately 23% of cells in the fields showedlamellipodia. After incubation with HGF for 30 min, however, thepercentage of lamellipodia-forming cells increased to 39%, andremained relatively constant at 60 min. In contrast, without HGF,the percentage of cells with lamellipodia did not increase with60 min incubation.
Syntrophins can bind to filamentous actin (F-actin) via its in-ternal domains such as the second PH and the SU domains in car-diac and skeletal muscle [18]. We therefore examined theintracellular localization of syntrophins during actin reorganiza-tion in the HGF-induced migrating cells. Without HGF, syntro-phins were distributed throughout the cytoplasm of non-migrating cells (Fig. 2A). However, when the cells were incubatedwith HGF, syntrophins concentrated to the rear and lateral part ofthe cells, distinctly separate from the lamellipodia (arrowheads inFig. 2A). Because PTEN is known to accumulate in the rear and lat-eral part of cells stimulated with chemo-attractant [30, 31], it iswidely used as a marker for the rear-lateral part of the migratingcells. In contrast, PI3-kinase localizes at the leading-edge of cellstreated with chemo-attractant and stimulates cell migration invarious cell types [6, 32–34]. We also found that PTEN is localizedin the rear-lateral region of the HGF-induced C2 cells (asterisks inFig. 2A), while it dispersed in the cytoplasmwithout HGF. To showthe localization of PI3-kinase, cells were stained with anti-p85 an-tibody, the PI3-kinase regulatory subunit. As expected, p85 wasfound in the region of lamellipodia in the HGF-induced cells (ar-rows in Fig. 2A). In co-immunolabeling experiments, PTEN andsyntrophins co-localized in the rear-lateral part of the migratingcells (arrows in Fig. 2B). However, syntrophins and p85 separatedin the HGF-induced migrating cells (asterisks in Fig. 2B). To con-firm the localization of syntrophin in the HGF-induced migratingcells, C2 cells were transfected with GFP fusion α-syntrophinand visualized under confocal laser scanning microscope. TheGFP protein can be seen in the rear part of the migrating cells(arrow in Fig. 2C). Then we examined whether syntrophins asso-ciate with PTEN by performing an immunoprecipitation assay.α-Syntrophin was precipitated by the anti-PTEN antibody, andconversely, PTEN was precipitated by the anti-pan-syntrophinantibody (Fig. 2D). These results suggest that syntrophins con-centrate at the rear-lateral part of the HGF-induced migratingcells.
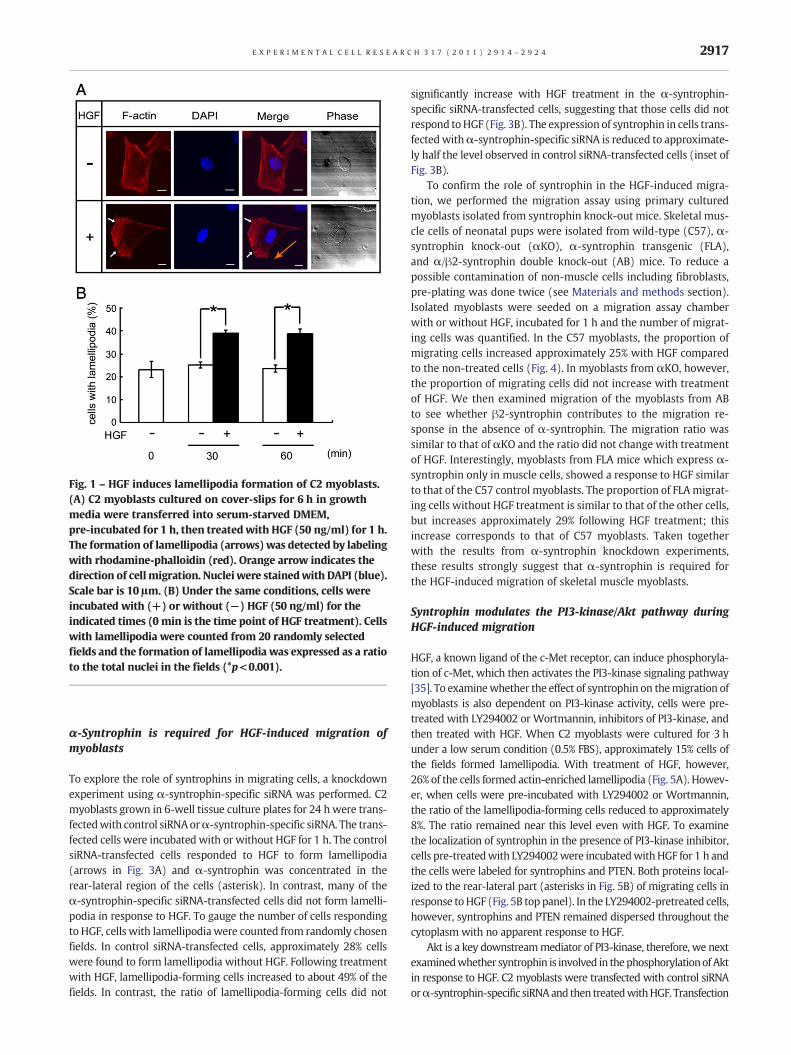
Fig. 1 – HGF induces lamellipodia formation of C2 myoblasts.(A) C2 myoblasts cultured on cover-slips for 6 h in growthmedia were transferred into serum-starved DMEM,pre-incubated for 1 h, then treated with HGF (50 ng/ml) for 1 h.The formation of lamellipodia (arrows) was detected by labelingwith rhodamine-phalloidin (red). Orange arrow indicates thedirection of cellmigration. Nuclei were stainedwith DAPI (blue).Scale bar is 10 μm. (B) Under the same conditions, cells wereincubated with (+) or without (−) HGF (50 ng/ml) for theindicated times (0 min is the time point of HGF treatment). Cellswith lamellipodia were counted from 20 randomly selectedfields and the formation of lamellipodia was expressed as a ratioto the total nuclei in the fields (*p<0.001).
2917E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
α-Syntrophin is required for HGF-induced migration ofmyoblasts
To explore the role of syntrophins in migrating cells, a knockdownexperiment using α-syntrophin-specific siRNA was performed. C2myoblasts grown in 6-well tissue culture plates for 24 h were trans-fectedwith control siRNAorα-syntrophin-specific siRNA. The trans-fected cells were incubated with or without HGF for 1 h. The controlsiRNA-transfected cells responded to HGF to form lamellipodia(arrows in Fig. 3A) and α-syntrophin was concentrated in therear-lateral region of the cells (asterisk). In contrast, many of theα-syntrophin-specific siRNA-transfected cells did not form lamelli-podia in response to HGF. To gauge the number of cells respondingto HGF, cells with lamellipodia were counted from randomly chosenfields. In control siRNA-transfected cells, approximately 28% cellswere found to form lamellipodia without HGF. Following treatmentwith HGF, lamellipodia-forming cells increased to about 49% of thefields. In contrast, the ratio of lamellipodia-forming cells did not
significantly increase with HGF treatment in the α-syntrophin-specific siRNA-transfected cells, suggesting that those cells did notrespond toHGF (Fig. 3B). The expression of syntrophin in cells trans-fectedwithα-syntrophin-specific siRNA is reduced to approximate-ly half the level observed in control siRNA-transfected cells (inset ofFig. 3B).
To confirm the role of syntrophin in the HGF-induced migra-tion, we performed the migration assay using primary culturedmyoblasts isolated from syntrophin knock-out mice. Skeletal mus-cle cells of neonatal pups were isolated from wild-type (C57), α-syntrophin knock-out (αKO), α-syntrophin transgenic (FLA),and α/β2-syntrophin double knock-out (AB) mice. To reduce apossible contamination of non-muscle cells including fibroblasts,pre-plating was done twice (see Materials and methods section).Isolated myoblasts were seeded on a migration assay chamberwith or without HGF, incubated for 1 h and the number of migrat-ing cells was quantified. In the C57 myoblasts, the proportion ofmigrating cells increased approximately 25% with HGF comparedto the non-treated cells (Fig. 4). In myoblasts from αKO, however,the proportion of migrating cells did not increase with treatmentof HGF. We then examined migration of the myoblasts from ABto see whether β2-syntrophin contributes to the migration re-sponse in the absence of α-syntrophin. The migration ratio wassimilar to that of αKO and the ratio did not change with treatmentof HGF. Interestingly, myoblasts from FLA mice which express α-syntrophin only in muscle cells, showed a response to HGF similarto that of the C57 control myoblasts. The proportion of FLAmigrat-ing cells without HGF treatment is similar to that of the other cells,but increases approximately 29% following HGF treatment; thisincrease corresponds to that of C57 myoblasts. Taken togetherwith the results from α-syntrophin knockdown experiments,these results strongly suggest that α-syntrophin is required forthe HGF-induced migration of skeletal muscle myoblasts.
Syntrophin modulates the PI3-kinase/Akt pathway duringHGF-induced migration
HGF, a known ligand of the c-Met receptor, can induce phosphoryla-tion of c-Met, which then activates the PI3-kinase signaling pathway[35]. To examinewhether the effect of syntrophin on themigration ofmyoblasts is also dependent on PI3-kinase activity, cells were pre-treated with LY294002 or Wortmannin, inhibitors of PI3-kinase, andthen treated with HGF. When C2 myoblasts were cultured for 3 hunder a low serum condition (0.5% FBS), approximately 15% cells ofthe fields formed lamellipodia. With treatment of HGF, however,26% of the cells formed actin-enriched lamellipodia (Fig. 5A). Howev-er, when cells were pre-incubated with LY294002 or Wortmannin,the ratio of the lamellipodia-forming cells reduced to approximately8%. The ratio remained near this level even with HGF. To examinethe localization of syntrophin in the presence of PI3-kinase inhibitor,cells pre-treatedwith LY294002were incubatedwithHGF for 1 h andthe cells were labeled for syntrophins and PTEN. Both proteins local-ized to the rear-lateral part (asterisks in Fig. 5B) of migrating cells inresponse toHGF (Fig. 5B top panel). In the LY294002-pretreated cells,however, syntrophins and PTEN remained dispersed throughout thecytoplasm with no apparent response to HGF.
Akt is a key downstreammediator of PI3-kinase, therefore, we nextexaminedwhether syntrophin is involved in thephosphorylationofAktin response to HGF. C2 myoblasts were transfected with control siRNAorα-syntrophin-specific siRNAand then treatedwithHGF. Transfection
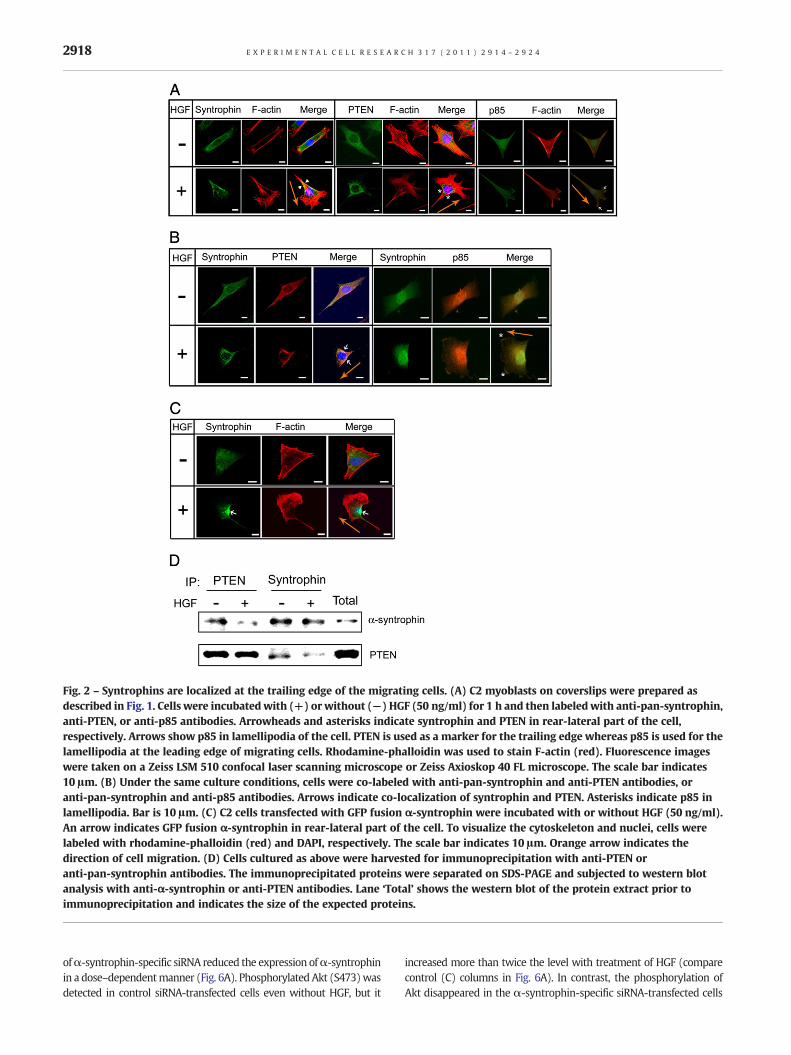
Fig. 2 – Syntrophins are localized at the trailing edge of the migrating cells. (A) C2 myoblasts on coverslips were prepared asdescribed in Fig. 1. Cells were incubated with (+) or without (−) HGF (50 ng/ml) for 1 h and then labeled with anti-pan-syntrophin,anti-PTEN, or anti-p85 antibodies. Arrowheads and asterisks indicate syntrophin and PTEN in rear-lateral part of the cell,respectively. Arrows show p85 in lamellipodia of the cell. PTEN is used as a marker for the trailing edge whereas p85 is used for thelamellipodia at the leading edge of migrating cells. Rhodamine-phalloidin was used to stain F-actin (red). Fluorescence imageswere taken on a Zeiss LSM 510 confocal laser scanning microscope or Zeiss Axioskop 40 FL microscope. The scale bar indicates10 μm. (B) Under the same culture conditions, cells were co-labeled with anti-pan-syntrophin and anti-PTEN antibodies, oranti-pan-syntrophin and anti-p85 antibodies. Arrows indicate co-localization of syntrophin and PTEN. Asterisks indicate p85 inlamellipodia. Bar is 10 μm. (C) C2 cells transfected with GFP fusion α-syntrophin were incubated with or without HGF (50 ng/ml).An arrow indicates GFP fusion α-syntrophin in rear-lateral part of the cell. To visualize the cytoskeleton and nuclei, cells werelabeled with rhodamine-phalloidin (red) and DAPI, respectively. The scale bar indicates 10 μm. Orange arrow indicates thedirection of cell migration. (D) Cells cultured as above were harvested for immunoprecipitation with anti-PTEN oranti-pan-syntrophin antibodies. The immunoprecipitated proteins were separated on SDS-PAGE and subjected to western blotanalysis with anti-α-syntrophin or anti-PTEN antibodies. Lane ‘Total’ shows the western blot of the protein extract prior toimmunoprecipitation and indicates the size of the expected proteins.
2918 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
ofα-syntrophin-specific siRNA reduced the expression ofα-syntrophinin a dose–dependentmanner (Fig. 6A). Phosphorylated Akt (S473)wasdetected in control siRNA-transfected cells even without HGF, but it
increased more than twice the level with treatment of HGF (comparecontrol (C) columns in Fig. 6A). In contrast, the phosphorylation ofAkt disappeared in the α-syntrophin-specific siRNA-transfected cells
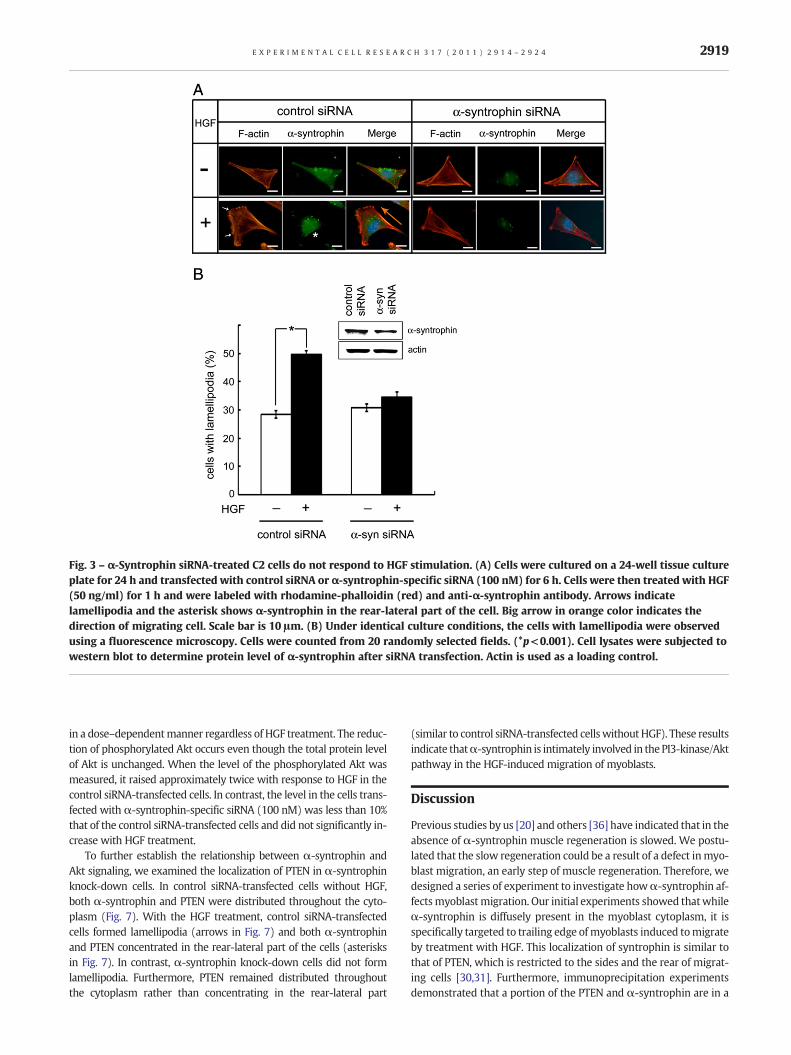
Fig. 3 – α-Syntrophin siRNA-treated C2 cells do not respond to HGF stimulation. (A) Cells were cultured on a 24-well tissue cultureplate for 24 h and transfected with control siRNA or α-syntrophin-specific siRNA (100 nM) for 6 h. Cells were then treated with HGF(50 ng/ml) for 1 h and were labeled with rhodamine-phalloidin (red) and anti-α-syntrophin antibody. Arrows indicatelamellipodia and the asterisk shows α-syntrophin in the rear-lateral part of the cell. Big arrow in orange color indicates thedirection of migrating cell. Scale bar is 10 μm. (B) Under identical culture conditions, the cells with lamellipodia were observedusing a fluorescence microscopy. Cells were counted from 20 randomly selected fields. (*p<0.001). Cell lysates were subjected towestern blot to determine protein level of α-syntrophin after siRNA transfection. Actin is used as a loading control.
2919E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
in a dose–dependentmanner regardless of HGF treatment. The reduc-tion of phosphorylated Akt occurs even though the total protein levelof Akt is unchanged. When the level of the phosphorylated Akt wasmeasured, it raised approximately twice with response to HGF in thecontrol siRNA-transfected cells. In contrast, the level in the cells trans-fected with α-syntrophin-specific siRNA (100 nM) was less than 10%that of the control siRNA-transfected cells and did not significantly in-crease with HGF treatment.
To further establish the relationship between α-syntrophin andAkt signaling, we examined the localization of PTEN in α-syntrophinknock-down cells. In control siRNA-transfected cells without HGF,both α-syntrophin and PTEN were distributed throughout the cyto-plasm (Fig. 7). With the HGF treatment, control siRNA-transfectedcells formed lamellipodia (arrows in Fig. 7) and both α-syntrophinand PTEN concentrated in the rear-lateral part of the cells (asterisksin Fig. 7). In contrast, α-syntrophin knock-down cells did not formlamellipodia. Furthermore, PTEN remained distributed throughoutthe cytoplasm rather than concentrating in the rear-lateral part
(similar to control siRNA-transfected cellswithoutHGF). These resultsindicate thatα-syntrophin is intimately involved in the PI3-kinase/Aktpathway in the HGF-induced migration of myoblasts.
Discussion
Previous studies by us [20] and others [36] have indicated that in theabsence of α-syntrophin muscle regeneration is slowed. We postu-lated that the slow regeneration could be a result of a defect inmyo-blast migration, an early step of muscle regeneration. Therefore, wedesigned a series of experiment to investigate howα-syntrophin af-fectsmyoblastmigration. Our initial experiments showed thatwhileα-syntrophin is diffusely present in the myoblast cytoplasm, it isspecifically targeted to trailing edge ofmyoblasts induced tomigrateby treatment with HGF. This localization of syntrophin is similar tothat of PTEN, which is restricted to the sides and the rear of migrat-ing cells [30,31]. Furthermore, immunoprecipitation experimentsdemonstrated that a portion of the PTEN and α-syntrophin are in a
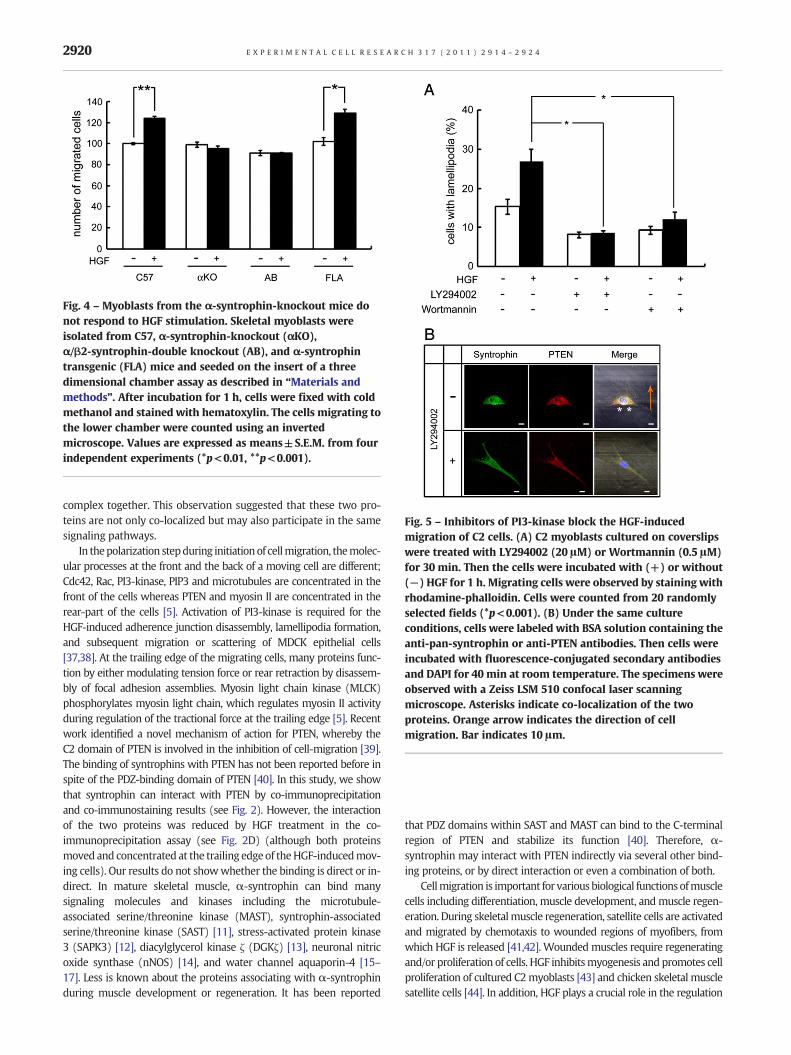
Fig. 4 – Myoblasts from the α-syntrophin-knockout mice donot respond to HGF stimulation. Skeletal myoblasts wereisolated from C57, α-syntrophin-knockout (αKO),α/β2-syntrophin-double knockout (AB), and α-syntrophintransgenic (FLA) mice and seeded on the insert of a threedimensional chamber assay as described in “Materials andmethods”. After incubation for 1 h, cells were fixed with coldmethanol and stained with hematoxylin. The cells migrating tothe lower chamber were counted using an invertedmicroscope. Values are expressed as means±S.E.M. from fourindependent experiments (*p<0.01, **p<0.001).
Fig. 5 – Inhibitors of PI3-kinase block the HGF-inducedmigration of C2 cells. (A) C2 myoblasts cultured on coverslipswere treated with LY294002 (20 μM) or Wortmannin (0.5 μM)for 30 min. Then the cells were incubated with (+) or without(−) HGF for 1 h. Migrating cells were observed by staining withrhodamine-phalloidin. Cells were counted from 20 randomlyselected fields (*p<0.001). (B) Under the same cultureconditions, cells were labeled with BSA solution containing theanti-pan-syntrophin or anti-PTEN antibodies. Then cells wereincubated with fluorescence-conjugated secondary antibodiesand DAPI for 40 min at room temperature. The specimens wereobserved with a Zeiss LSM 510 confocal laser scanningmicroscope. Asterisks indicate co-localization of the twoproteins. Orange arrow indicates the direction of cellmigration. Bar indicates 10 μm.
2920 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
complex together. This observation suggested that these two pro-teins are not only co-localized but may also participate in the samesignaling pathways.
In thepolarization stepduring initiationof cellmigration, themolec-ular processes at the front and the back of a moving cell are different;Cdc42, Rac, PI3-kinase, PIP3 and microtubules are concentrated in thefront of the cells whereas PTEN and myosin II are concentrated in therear-part of the cells [5]. Activation of PI3-kinase is required for theHGF-induced adherence junction disassembly, lamellipodia formation,and subsequent migration or scattering of MDCK epithelial cells[37,38]. At the trailing edge of the migrating cells, many proteins func-tion by either modulating tension force or rear retraction by disassem-bly of focal adhesion assemblies. Myosin light chain kinase (MLCK)phosphorylates myosin light chain, which regulates myosin II activityduring regulation of the tractional force at the trailing edge [5]. Recentwork identified a novel mechanism of action for PTEN, whereby theC2 domain of PTEN is involved in the inhibition of cell-migration [39].The binding of syntrophins with PTEN has not been reported before inspite of the PDZ-binding domain of PTEN [40]. In this study, we showthat syntrophin can interact with PTEN by co-immunoprecipitationand co-immunostaining results (see Fig. 2). However, the interactionof the two proteins was reduced by HGF treatment in the co-immunoprecipitation assay (see Fig. 2D) (although both proteinsmoved and concentrated at the trailing edge of theHGF-inducedmov-ing cells). Our results do not showwhether the binding is direct or in-direct. In mature skeletal muscle, α-syntrophin can bind manysignaling molecules and kinases including the microtubule-associated serine/threonine kinase (MAST), syntrophin-associatedserine/threonine kinase (SAST) [11], stress-activated protein kinase3 (SAPK3) [12], diacylglycerol kinase ζ (DGKζ) [13], neuronal nitricoxide synthase (nNOS) [14], and water channel aquaporin-4 [15–17]. Less is known about the proteins associating with α-syntrophinduring muscle development or regeneration. It has been reported
that PDZ domains within SAST and MAST can bind to the C-terminalregion of PTEN and stabilize its function [40]. Therefore, α-syntrophin may interact with PTEN indirectly via several other bind-ing proteins, or by direct interaction or even a combination of both.
Cellmigration is important for various biological functions ofmusclecells including differentiation, muscle development, andmuscle regen-eration. During skeletalmuscle regeneration, satellite cells are activatedand migrated by chemotaxis to wounded regions of myofibers, fromwhich HGF is released [41,42]. Wounded muscles require regeneratingand/or proliferation of cells. HGF inhibitsmyogenesis and promotes cellproliferation of cultured C2myoblasts [43] and chicken skeletal musclesatellite cells [44]. In addition, HGF plays a crucial role in the regulation
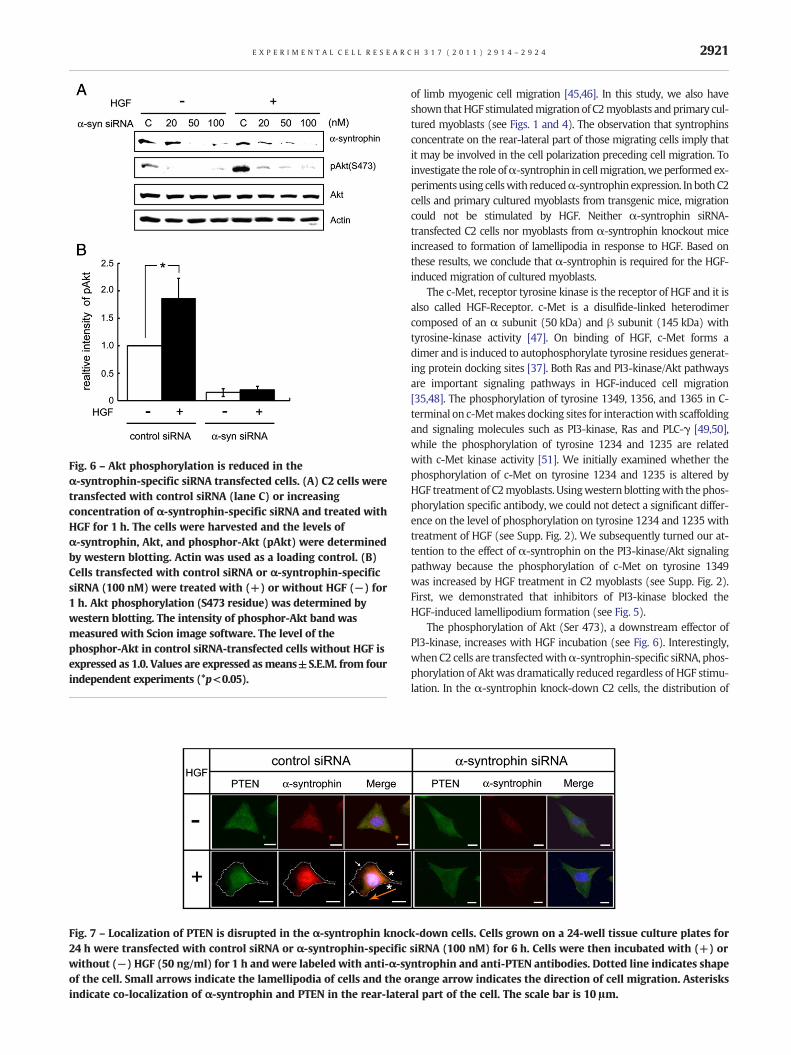
Fig. 6 – Akt phosphorylation is reduced in theα-syntrophin-specific siRNA transfected cells. (A) C2 cells weretransfected with control siRNA (lane C) or increasingconcentration of α-syntrophin-specific siRNA and treated withHGF for 1 h. The cells were harvested and the levels ofα-syntrophin, Akt, and phosphor-Akt (pAkt) were determinedby western blotting. Actin was used as a loading control. (B)Cells transfected with control siRNA or α-syntrophin-specificsiRNA (100 nM) were treated with (+) or without HGF (−) for1 h. Akt phosphorylation (S473 residue) was determined bywestern blotting. The intensity of phosphor-Akt band wasmeasured with Scion image software. The level of thephosphor-Akt in control siRNA-transfected cells without HGF isexpressed as 1.0. Values are expressed asmeans±S.E.M. from fourindependent experiments (*p<0.05).
Fig. 7 – Localization of PTEN is disrupted in the α-syntrophin knoc24 h were transfected with control siRNA or α-syntrophin-specificwithout (−) HGF (50 ng/ml) for 1 h and were labeled with anti-α-syof the cell. Small arrows indicate the lamellipodia of cells and the oindicate co-localization of α-syntrophin and PTEN in the rear-later
2921E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
of limb myogenic cell migration [45,46]. In this study, we also haveshown thatHGF stimulatedmigration of C2myoblasts andprimary cul-tured myoblasts (see Figs. 1 and 4). The observation that syntrophinsconcentrate on the rear-lateral part of those migrating cells imply thatit may be involved in the cell polarization preceding cell migration. Toinvestigate the role ofα-syntrophin in cellmigration,weperformed ex-periments using cellswith reducedα-syntrophin expression. In bothC2cells and primary cultured myoblasts from transgenic mice, migrationcould not be stimulated by HGF. Neither α-syntrophin siRNA-transfected C2 cells nor myoblasts from α-syntrophin knockout miceincreased to formation of lamellipodia in response to HGF. Based onthese results, we conclude that α-syntrophin is required for the HGF-induced migration of cultured myoblasts.
The c-Met, receptor tyrosine kinase is the receptor of HGF and it isalso called HGF-Receptor. c-Met is a disulfide-linked heterodimercomposed of an α subunit (50 kDa) and β subunit (145 kDa) withtyrosine-kinase activity [47]. On binding of HGF, c-Met forms adimer and is induced to autophosphorylate tyrosine residues generat-ing protein docking sites [37]. Both Ras and PI3-kinase/Akt pathwaysare important signaling pathways in HGF-induced cell migration[35,48]. The phosphorylation of tyrosine 1349, 1356, and 1365 in C-terminal on c-Metmakes docking sites for interactionwith scaffoldingand signaling molecules such as PI3-kinase, Ras and PLC-γ [49,50],while the phosphorylation of tyrosine 1234 and 1235 are relatedwith c-Met kinase activity [51]. We initially examined whether thephosphorylation of c-Met on tyrosine 1234 and 1235 is altered byHGF treatment of C2myoblasts. Usingwestern blottingwith the phos-phorylation specific antibody, we could not detect a significant differ-ence on the level of phosphorylation on tyrosine 1234 and 1235 withtreatment of HGF (see Supp. Fig. 2). We subsequently turned our at-tention to the effect of α-syntrophin on the PI3-kinase/Akt signalingpathway because the phosphorylation of c-Met on tyrosine 1349was increased by HGF treatment in C2 myoblasts (see Supp. Fig. 2).First, we demonstrated that inhibitors of PI3-kinase blocked theHGF-induced lamellipodium formation (see Fig. 5).
The phosphorylation of Akt (Ser 473), a downstream effector ofPI3-kinase, increases with HGF incubation (see Fig. 6). Interestingly,whenC2 cells are transfectedwithα-syntrophin-specific siRNA, phos-phorylation of Akt was dramatically reduced regardless of HGF stimu-lation. In the α-syntrophin knock-down C2 cells, the distribution of
k-down cells. Cells grown on a 24-well tissue culture plates forsiRNA (100 nM) for 6 h. Cells were then incubated with (+) orntrophin and anti-PTEN antibodies. Dotted line indicates shaperange arrow indicates the direction of cell migration. Asterisksal part of the cell. The scale bar is 10 μm.

2922 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
PTEN in the rear-lateral part of the cells was disrupted even with HGFtreatment (see Fig. 7). In this case, it could be that PTEN dephosphor-ylates PIP3, which inhibits phosphorylation of Akt. These results sug-gest that syntrophin is involved in the PI3-kinase/Akt pathway inthe HGF-induced lamellipodium formation.
α-Syntrophin knockout mice do not have specific symptomsfor muscular dystrophy although they do have aberrant neuro-muscular junctions [22]. Previously, we have shown that bothdifferentiation of α-syntrophin siRNA C2 cells and the regenera-tion of muscle in the α-syntrophin knock-out mouse is delayedcompared to controls [20]. In this study, we show that both α-syntrophin siRNA C2 cells and the myoblasts from the α-syntrophin knockout mice do not respond to HGF stimulationto initiate cell migration (see Figs. 3 and 4). These results suggestthat deficiency of α-syntrophin results in insensitivity to HGF,which may cause delayed differentiation and regeneration.
Based on our observation that syntrophin is concentrated atthe trailing edge of migrating myoblasts, we can propose mecha-nisms for syntrophin function. Intracellular calcium levels are im-plicated in the disassembly of adhesions at the trailing edge [52].Important targets of calcium are the calcium-regulated phospha-tase, calcineurin [53] and calcium-activated protease, calpain[54], which is involved in adhesion disassembly. Syntrophin canbind to calmodulin via C-terminal 24 amino acids in the SU do-main in a Ca2+-dependent manner and α-syntrophin also canbind to plasma membrane Ca2+/calmodulin-dependent ATPase(PMCA) [55,56]. Both syntrophin and dystrophin are phosphory-lated in vivo and in vitro by Ca2+/calmodulin dependent kinase II(CamKII) [57] which is also required for the migration of PDGF-stimulated vascular smooth muscle cell [58]. Together thesereports suggest that syntrophins have a role in trailing edge re-traction in a calcium-dependent manner. The PH1 domain of α-syntrophin also binds phosphoinositol 4, 5 bisphosphate (PtdIns(4,5)P2) [59,60] which is formed by phosphatidylinositol phos-phate 5 kinase or PTEN. PtdIns(4,5) P2 is involved in actin organi-zation and focal adhesion formation [61,62]. In addition, theheterotrimeric Gαβγ is bound by syntrophin in a laminin-dependent manner [63]. It is likely that syntrophins function bylinking these diverse signaling molecules to form a functionalcomplex at the trailing edge that can modulate cell migration.
Supplementary materials related to this article can be foundonline at doi:10.1016/j.yexcr.2011.09.017.
Acknowledgments
This work was supported by the Korea Research Foundation grantfunded by the Korean Government (MOEHRD, Basic Research Pro-motion Fund) (KRF-2008-531-E00004) to HSK and by NIH grantNS33145 to SCF.
R E F E R E N C E S
[1] P. Seale, M.A. Rudnicki, A new look at the origin, function, and“stem-cell” status of muscle satellite cells, Dev. Biol. 218 (2000)115–124.
[2] E. Schultz, D.L. Jaryszak, C.R. Valliere, Response of satellite cells tofocal skeletal muscle injury, Muscle Nerve 8 (1985) 217–222.
[3] S.M. Sheehan, R. Tatsumi, C.J. Temm-Grove, R.E. Allen, HGF is anautocrine growth factor for skeletal muscle satellite cells in vitro,Muscle Nerve 23 (2000) 239–245.
[4] K. Kawamura, K. Takano, S. Suetsugu, S. Kurisu, D. Yamazaki, H.Miki, T. Takenawa, T. Endo, N-WASP and WAVE2 actingdownstream of phosphatidylinositol 3-kinase are required formyogenic cell migration induced by hepatocyte growth factor, J.Biol. Chem. 279 (2004) 54862–54871.
[5] A.J. Ridley, M.A. Schwartz, K. Burridge, R.A. Firtel, M.H. Ginsberg,G. Borisy, J.T. Parsons, A.R. Horwitz, Cell migration: integratingsignals from front to back, Science 302 (2003) 1704–1709.
[6] T.E. Taher, P.W. Derksen, O.J. de Boer, M. Spaargaren, P. Teeling,A.C. van der Wal, S.T. Pals, Hepatocyte growth factor triggerssignaling cascades mediating vascular smooth muscle cellmigration, Biochem. Biophys. Res. Commun. 298 (2002) 80–86.
[7] D.E. Albrecht, S.C. Froehner, Syntrophins and dystrobrevins:defining the dystrophin scaffold at synapses, Neurosignals 11(2002) 123–129.
[8] M.E. Adams, T.M. Dwyer, L.L. Dowler, R.A. White, S.C. Froehner,Mouse alpha 1- and beta 2-syntrophin gene structure,chromosome localization, and homology with a discs largedomain, J. Biol. Chem. 270 (1995) 25859–25865.
[9] G. Piluso, M. Mirabella, E. Ricci, A. Belsito, C. Abbondanza, S. Servidei,A.A. Puca, P. Tonali, G.A. Puca, V. Nigro, Gamma1- andgamma2-syntrophins, two novel dystrophin-binding proteinslocalized in neuronal cells, J. Biol. Chem. 275 (2000) 15851–15860.
[10] A.H. Ahn, C.A. Freener, E. Gussoni, M. Yoshida, E. Ozawa, L.M.Kunkel, The three human syntrophin genes are expressed indiverse tissues, have distinct chromosomal locations, and eachbind to dystrophin and its relatives, J. Biol. Chem. 271 (1996)2724–2730.
[11] C. Lumeng, S. Phelps, G.E. Crawford, P.D. Walden, K. Barald, J.S.Chamberlain, Interactions between beta 2-syntrophin and afamily of microtubule-associated serine/threonine kinases, Nat.Neurosci. 2 (1999) 611–617.
[12] M. Hasegawa, A. Cuenda, M.G. Spillantini, G.M. Thomas, V.Buee-Scherrer, P. Cohen, M. Goedert, Stress-activated proteinkinase-3 interacts with the PDZ domain of alpha1-syntrophin. Amechanism for specific substrate recognition, J. Biol. Chem. 274(1999) 12626–12631.
[13] A. Hogan, L. Shepherd, J. Chabot, S. Quenneville, S.M. Prescott,M.K. Topham, S.H. Gee, Interaction of gamma 1-syntrophin withdiacylglycerol kinase-zeta. Regulation of nuclear localization byPDZ interactions, J. Biol. Chem. 276 (2001) 26526–26533.
[14] Y. Miyagoe-Suzuki, S.I. Takeda, Association of neuronal nitricoxide synthase (nNOS) with alpha1-syntrophin at thesarcolemma, Microsc. Res. Tech. 55 (2001) 164–170.
[15] J.D. Neely, M. Amiry-Moghaddam, O.P. Ottersen, S.C. Froehner, P.Agre, M.E. Adams, Syntrophin-dependent expression andlocalization of aquaporin-4 water channel protein, Proc. Natl.Acad. Sci. U. S. A. 98 (2001) 14108–14113.
[16] M. Inoue, Y. Wakayama, J.W. Liu, M. Murahashi, S. Shibuya, H.Oniki, Ultrastructural localization of aquaporin 4 andalpha1-syntrophin in the vascular feet of brain astrocytes,Tohoku J. Exp. Med. 197 (2002) 87–93.
[17] M.E. Adams, Y. Tesch, J.M. Percival, D.E. Albrecht, J.I. Conhaim, K.Anderson, S.C. Froehner, Differential targeting of nNOS and AQP4to dystrophin-deficient sarcolemma by membrane-directed{alpha}-dystrobrevin, J. Cell Sci. 121 (2008) 48–54.
[18] Y. Iwata, M. Sampaolesi, M. Shigekawa, S. Wakabayashi,Syntrophin is an actin-binding protein the cellular localization ofwhich is regulated through cytoskeletal reorganization inskeletal muscle cells, Eur. J. Cell Biol. 83 (2004) 555–565.
[19] S.T. Cooper, A.L. Maxwell, E. Kizana, M. Ghoddusi, E.C. Hardeman,I.E. Alexander, D.G. Allen, K.N. North, C2C12 co-culture on afibroblast substratum enables sustained survival of contractile,highly differentiated myotubes with peripheral nuclei and adultfast myosin expression, Cell Motil. Cytoskeleton 58 (2004)200–211.

2923E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
[20] M.J. Kim, S.H. Hwang, J.A. Lim, S.C. Froehner, M.E. Adams, H.S.Kim, Alpha-syntrophin modulates myogenin expression indifferentiating myoblasts, PLoS One 5 (2010) e15355.
[21] M.F. Peters, M.E. Adams, S.C. Froehner, Differential association ofsyntrophin pairs with the dystrophin complex, J. Cell Biol. 138(1997) 81–93.
[22] M.E. Adams, N. Kramarcy, S.P. Krall, S.G. Rossi, R.L. Rotundo, R.Sealock, S.C. Froehner, Absence of alpha-syntrophin leads tostructurally aberrant neuromuscular synapses deficient inutrophin, J. Cell Biol. 150 (2000) 1385–1398.
[23] M.E. Adams, H.A. Mueller, S.C. Froehner, In vivo requirement of thealpha-syntrophin PDZ domain for the sarcolemmal localization ofnNOS and aquaporin-4, J. Cell Biol. 155 (2001) 113–122.
[24] M.E. Adams, N. Kramarcy, T. Fukuda, A.G. Engel, R. Sealock, S.C.Froehner, Structural abnormalities at neuromuscular synapseslacking multiple syntrophin isoforms, J. Neurosci. 24 (2004)10302–10309.
[25] K. Bandow, T. Ohnishi, M. Tamura, I. Semba, Y. Daikuhara,Hepatocyte growth factor/scatter factor stimulates migration ofmuscle precursors in developing mouse tongue, J. Cell. Physiol.201 (2004) 236–243.
[26] I. Royal, N. Lamarche-Vane, L. Lamorte, K. Kaibuchi, M. Park,Activation of cdc42, rac, PAK, and rho-kinase in response tohepatocyte growth factor differentially regulates epithelial cellcolony spreading and dissociation, Mol. Biol. Cell 11 (2000)1709–1725.
[27] T.D. Pollard, G.G. Borisy, Cellular motility driven by assembly anddisassembly of actin filaments, Cell 112 (2003) 453–465.
[28] C. Ballestrem, B. Wehrle-Haller, B.A. Imhof, Actin dynamics inliving mammalian cells, J. Cell Sci. 111 (Pt 12) (1998) 1649–1658.
[29] M. Vicente-Manzanares, M. Rey, M. Perez-Martinez, M. Yanez-Mo,D. Sancho, J.R. Cabrero, O. Barreiro, H. de la Fuente, K. Itoh, F.Sanchez-Madrid, The RhoA effector mDia is induced during T cellactivation and regulates actin polymerization and cell migration in Tlymphocytes, J. Immunol. 171 (2003) 1023–1034.
[30] S. Funamoto, R. Meili, S. Lee, L. Parry, R.A. Firtel, Spatial andtemporal regulation of 3-phosphoinositides by PI 3-kinase andPTEN mediates chemotaxis, Cell 109 (2002) 611–623.
[31] F.I. Comer, C.A. Parent, PI 3-kinases and PTEN: how oppositeschemoattract, Cell 109 (2002) 541–544.
[32] P. Devreotes, C. Janetopoulos, Eukaryotic chemotaxis:distinctions between directional sensing and polarization, J. Biol.Chem. 278 (2003) 20445–20448.
[33] S. Merlot, R.A. Firtel, Leading the way: directional sensingthrough phosphatidylinositol 3-kinase and other signalingpathways, J. Cell Sci. 116 (2003) 3471–3478.
[34] E.A. Goncharova, A.J. Ammit, C. Irani, R.G. Carroll, A.J. Eszterhas, R.A.Panettieri, V.P. Krymskaya, PI3K is required for proliferation andmigration of human pulmonary vascular smooth muscle cells, Am. J.Physiol. Lung Cell. Mol. Physiol. 283 (2002) L354–L363.
[35] R. van der Voort, T.E. Taher, P.W. Derksen, M. Spaargaren, R. vander Neut, S.T. Pals, The hepatocyte growth factor/Met pathway indevelopment, tumorigenesis, and B-cell differentiation, Adv.Cancer Res. 79 (2000) 39–90.
[36] Y. Hosaka, T. Yokota, Y. Miyagoe-Suzuki, K. Yuasa, M. Imamura, R.Matsuda, T. Ikemoto, S. Kameya, S. Takeda,Alpha1-syntrophin-deficient skeletal muscle exhibitshypertrophy and aberrant formation of neuromuscular junctionsduring regeneration, J. Cell Biol. 158 (2002) 1097–1107.
[37] I. Royal, M. Park, Hepatocyte growth factor-induced scatter ofMadin–Darby canine kidney cells requires phosphatidylinositol3-kinase, J. Biol. Chem. 270 (1995) 27780–27787.
[38] A. Khwaja, K. Lehmann, B.M. Marte, J. Downward,Phosphoinositide 3-kinase induces scattering and tubulogenesisin epithelial cells through a novel pathway, J. Biol. Chem. 273(1998) 18793–18801.
[39] M. Raftopoulou, S. Etienne-Manneville, A. Self, S. Nicholls, A. Hall,Regulation of cell migration by the C2 domain of the tumorsuppressor PTEN, Science 303 (2004) 1179–1181.
[40] M. Valiente, A. Andres-Pons, B. Gomar, J. Torres, A. Gil, C.Tapparel, S.E. Antonarakis, R. Pulido, Binding of PTEN to specificPDZ domains contributes to PTEN protein stability andphosphorylation by microtubule-associated serine/threoninekinases, J. Biol. Chem. 280 (2005) 28936–28943.
[41] R. Bischoff, Chemotaxis of skeletal muscle satellite cells, Dev. Dyn.208 (1997) 505–515.
[42] K.J. Miller, D. Thaloor, S. Matteson, G.K. Pavlath, Hepatocytegrowth factor affects satellite cell activation and differentiation inregenerating skeletal muscle, Am. J. Physiol. Cell Physiol. 278(2000) C174–C181.
[43] S. Anastasi, S. Giordano, O. Sthandier, G. Gambarotta, R. Maione,P. Comoglio, P. Amati, A natural hepatocyte growth factor/scatterfactor autocrine loop in myoblast cells and the effect of theconstitutive Met kinase activation on myogenic differentiation, JCell Biol 137 (1997) 1057–1068.
[44] Y. Leshem, D.B. Spicer, R. Gal-Levi, O. Halevy, Hepatocyte growthfactor (HGF) inhibits skeletal muscle cell differentiation: a rolefor the bHLH protein twist and the cdk inhibitor p27, J. Cell.Physiol. 184 (2000) 101–109.
[45] F. Bladt, D. Riethmacher, S. Isenmann, A. Aguzzi, C. Birchmeier,Essential role for the c-met receptor in the migration of myogenicprecursor cells into the limb bud, Nature 376 (1995) 768–771.
[46] R. Gal-Levi, Y. Leshem, S. Aoki, T. Nakamura, O. Halevy,Hepatocyte growth factor plays a dual role in regulating skeletalmuscle satellite cell proliferation and differentiation, Biochim.Biophys. Acta 1402 (1998) 39–51.
[47] L. Beviglia, K.Matsumoto, C.S. Lin, B.L. Ziober, R.H. Kramer, Expressionof the c-Met/HGF receptor in human breast carcinoma: correlationwith tumor progression, Int. J. Cancer 74 (1997) 301–309.
[48] Y. Ueoka, K. Kato, Y. Kuriaki, S. Horiuchi, Y. Terao, J. Nishida, H.Ueno, N. Wake, Hepatocyte growth factor modulates motility andinvasiveness of ovarian carcinomas via Ras-mediated pathway,Br. J. Cancer 82 (2000) 891–899.
[49] C. Birchmeier, W. Birchmeier, E. Gherardi, G.F. Vande Woude,Met, metastasis, motility and more, Nat. Rev. Mol. Cell Biol. 4(2003) 915–925.
[50] S. Benvenuti, P.M. Comoglio, The MET receptor tyrosine kinase ininvasion and metastasis, J. Cell. Physiol. 213 (2007) 316–325.
[51] M. Hecht, M. Papoutsi, H.D. Tran, J. Wilting, L. Schweigerer,Hepatocyte growth factor/c-Met signaling promotes theprogression of experimental human neuroblastomas, Cancer Res.64 (2004) 6109–6118.
[52] J. Lee, A. Ishihara, G. Oxford, B. Johnson, K. Jacobson, Regulation ofcell movement is mediated by stretch-activated calciumchannels, Nature 400 (1999) 382–386.
[53] B. Hendey, C.B. Klee, F.R. Maxfield, Inhibition of neutrophilchemokinesis on vitronectin by inhibitors of calcineurin, Science258 (1992) 296–299.
[54] A. Glading, D.A. Lauffenburger, A. Wells, Cutting to the chase:calpain proteases in cell motility, Trends Cell Biol. 12 (2002) 46–54.
[55] B.J. Newbell, J.T. Anderson, H.W. Jarrett, Ca2+-calmodulinbinding to mouse alpha1 syntrophin: syntrophin is also aCa2+-binding protein, Biochemistry 36 (1997) 1295–1305.
[56] J.C. Williams, A.L. Armesilla, T.M. Mohamed, C.L. Hagarty, F.H.McIntyre, S. Schomburg, A.O. Zaki, D. Oceandy, E.J. Cartwright,M.H. Buch, M. Emerson, L. Neyses, The sarcolemmal calcium pump,alpha-1 syntrophin, and neuronal nitric-oxide synthase are partsof a macromolecular protein complex, J. Biol. Chem. 281 (2006)23341–23348.
[57] R. Madhavan, H.W. Jarrett, Calmodulin-activated phosphorylationof dystrophin, Biochemistry 33 (1994) 5797–5804.
[58] P.J. Pfleiderer, K.K. Lu, M.T. Crow, R.S. Keller, H.A. Singer,Modulation of vascular smooth muscle cell migration by calcium/calmodulin-dependent protein kinase II-delta 2, Am. J. Physiol.Cell Physiol. 286 (2004) C1238–C1245.
[59] P.S. Chockalingam, S.H. Gee, H.W. Jarrett, Pleckstrin homologydomain 1 of mouse alpha 1-syntrophin binds phosphatidylinositol4,5-bisphosphate, Biochemistry 38 (1999) 5596–5602.

2924 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 7 ( 2 0 1 1 ) 2 9 1 4 – 2 9 2 4
[60] J. Yan, W. Wen, W. Xu, J.F. Long, M.E. Adams, S.C. Froehner, M.Zhang, Structure of the split PH domain and distinct lipid-bindingproperties of the PH-PDZ supramodule of alpha-syntrophin,EMBO J. 24 (2005) 3985–3995.
[61] I. van den Bout, N. Divecha, PIP5K-driven PtdIns(4,5)P2synthesis: regulation and cellular functions, J. Cell Sci. 122 (2009)3837–3850.
[62] K. Ling, N.J. Schill, M.P. Wagoner, Y. Sun, R.A. Anderson, Movin' onup: the role of PtdIns(4,5)P(2) in cell migration, Trends Cell Biol.16 (2006) 276–284.
[63] Y.W. Zhou, S.A. Oak, S.E. Senogles, H.W. Jarrett, Laminin-alpha1globular domains 3 and 4 induce heterotrimeric G protein binding toalpha-syntrophin's PDZ domain and alter intracellular Ca2+ inmuscle, Am. J. Physiol. Cell Physiol. 288 (2005) C377–C388.