Embed Size (px)
DESCRIPTION
hy
Citation preview

TATAP MUKA 6
STRATEGI KLONING (1)

CLONE-STRATEGY
requires
DNA fragments
Genomics DNA mRNA PCR
DNA Fragment
for cloning
DNA Synthesis
• Mechanically• Restriction
enzyme
cDNA
Synthesised tothrough
Obtained from
Aligning DNA fragment to Vector
Method required
Biomolecular eecombinant
Inserted DNA /Vector concatameric
Blunt-end ligation Linker application Adapter application Homopolymer tailing Ligation of cohesive
end
Aligned to vector using
Resulted in
• cDNA lybrary• Genomics library
Resulted in
through
Can be applied for
Gene isolation
Produces
Resulted in

Learning Outcome (LO)
LO 54: menjelaskan tahapan kloning DNA
LO 55: menjelaskan materi awal kloning DNA
LO 56: menjelaskan sintesis cDNA
LO 57: menjelaskan kloning cDNA ke dalam plasmid
LO 58: menjelaskan penggunaan linker, adaptor, dan
homopolymer tailing
LO 59: menjelaskan kloning cDNA ke dalam bakteriofaga

Cloning strategies
LO 54: menjelaskan tahapan kloning DNA
Four steps of cloning
Cutting DNA strands into fragments
Alignment of DNA fragments to vector to allow recombinance
Propagation of DNA recombinant in the host cell
Identification and selection of targetted DNA
recombinant

LO 54: menjelaskan tahapan kloning DNA
cDNA synthesis
Cutting of Dna fragment using
restriction enzymeMechanicall
cuttingChemical synthesis
Transformation Transfection In vitro packing of DNA
Blunt end ligation
Linker application
Homopolymer tailing
Cohesive end ligation
Preparation of DNA fragment
Ligation
Inserting DNA to the host’s cell
Path ways

Initial matterial for cloning
LO 55: menjelaskan materi awal kloning
Genomic DNA mRNARepresenting whole genome
of an organismIf the genome is too big, it
might cause big problem especially if the main purpose is to isolate single gene
Has intron, controlling area and repeating sekuens.
Representing genetic information expressed by a particular cell
Not all genomic DNA is represented in an mRNA population .
Can only represent abundance of particular mRNA
Contains sequence of gene codes, due to elimination of intron along processing of mRNA

Note: The diversity of mRNAs is indicated by the number of different mRNA molecules. There is one mRNA that is present in chick oviduct cells at a very high level (100000 molecules per cell). This mRNA encodes ovalbumin, the major egg white protein. Source: After Old & Primrose (1989), Principles of Gene Manipulation, 4th edition, Blackwell. Mouse data from Young et al. (1976), Biochemistry 15, 2823–2828, copyright (1976) American Chemical Society. Chick data from Axel et al. (1976), Cell 11, 247–254, copyright (1976) Cell Press. Reproduced with permission.
LO 55: menjelaskan materi awal kloning

Sintesis cDNA
LO 56: menjelaskan sintesis cDNA
Fig. 6.2. Synthesis of cDNA. Poly(A) RNA (mRNA) is used as the starting material
short oligo(dT) primer is annealed to the poly(A) tail on the mRNA
reverse transcriptase to begin copying the mRNA
mRNA is removed
short double-stranded hairpin loop
second-strand synthesis by a DNA polymerase
double-stranded cDNA is trimmed with S1 nuclease

Producing cDNA from mRNA

The above statements might cause some problems as follows:1) Sinthesis of cDNA might probably not efficientlly
run especially when mRNA is relatively long.2) Application of S1 nuclease could relieve some
important bases at 5’ end.
The latest method to synthesis cDNA is by using an Oligo(dG)-primed second-strand cDNA synthesis (Figure 6.3).
LO 56: menjelaskan sintesis cDNA

Fig. 6.3. Oligo(dG)-primed second-strand cDNA synthesis
LO 56: menjelaskan sintesis cDNA
mRNA·cDNA hybrid
tailed with C residues using terminal transferase
An oligo(dG) primer is annealed to the C tails
hydrolyses the mRNA
double-stranded full-length cDNA molecule

Plasmid is still, being used as vector especially when isolation of preffered cDNA including small numbers of screened-clones.
Ligation of cDNA fragment to the vector usually run in either of the following methods : Ligation of blunt-
end linker homopolimer
tailing
Cloning of cDNA to the plasmid vector
LO 57: menjelaskan kloning cDNA ke dalam plasmid
Sinthesys of cDNA
Blunt-end ligation
linker Homopolymer tailing

Blunt-end ligation is a ligation process of DNA molecule to the blunt-end, done by which is done by DNA ligase
The blunt-end might be obtained from : Applicatiion of S1 nuclease Filling the cohesive-end with the DNA polimerase.
Blunt-ent ligation is inefficent ligation process since there is no particular relation between molecules to hold the DNA strand before the DNA ligase forms fosfodiester bonds to produce DNA recombinant.
Probability for joining two blunt-ends, the ligation might be done using high concentration DNA

Theoritically, when DNA vector is mixed with cDNA, it might produced: Recombinant with one insert. Self-ligate to make a circular molecule or insert / vector DNA could form concatemers
Practically, to avoid self ligation vector is treated by fosfatase (either BAP or CIP).
To solve problems in joining blunt-ends the following methods are applied : Linker Adaptor Homopolymer tailing

Fig. 6.4. Use of linkers. (a) The 10-mer 5’-CCGAATTCGG-3’ contains the recognition site for EcoRI. (b) The linker is added to blunt-ended DNA using DNA ligase. (c) The construct is then digested with EcoRI, which cleaves the linker to generate protruding 5’ termini.
LO 58: menjelaskan penggunaan linker, adaptor, dan homopolymer tailing

Fig. 6.5. Use of adaptors. In this example a BamHI adaptor (5’-GATCCCCGGG-3’) is annealed with a single-stranded HpaII linker (3’-GGGCCC-5’) to generate a double stranded cohesive-ended molecule, as shown in (a). This is added to blunt-ended DNA using DNA ligase. The DNA therefore gains protruding 5’ termini without the need for digestion with a restriction enzyme, as shown in (b). The 5’ terminus of the adaptor can be dephosphorylated to prevent self ligation.
LO 58: menjelaskan penggunaan linker, adaptor, dan homopolymer tailing
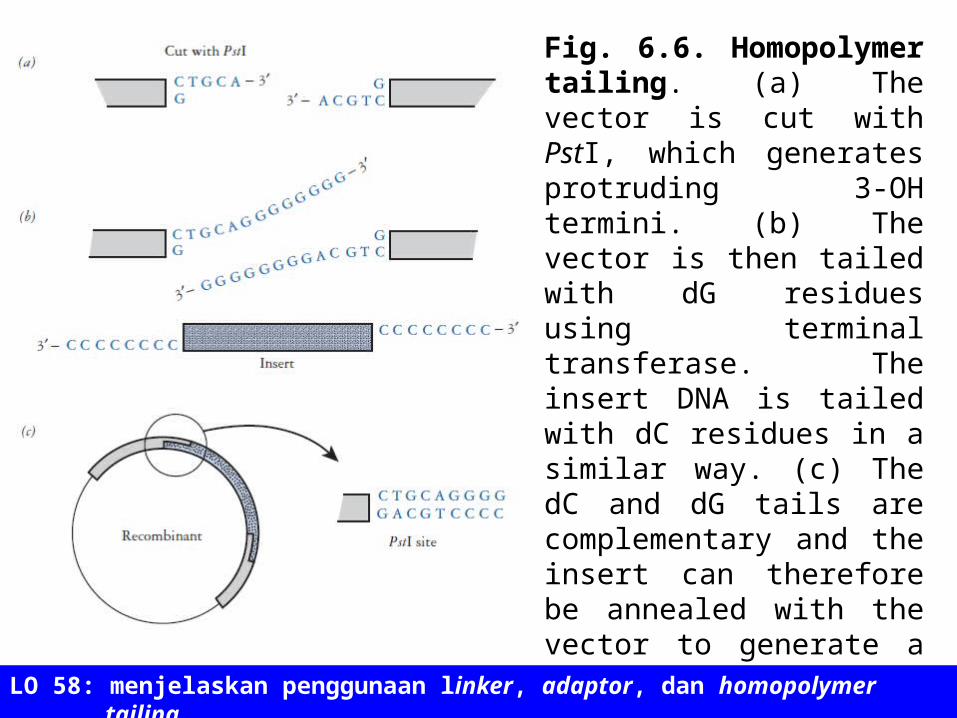
Fig. 6.6. Homopolymer tailing. (a) The vector is cut with PstI, which generates protruding 3-OH termini. (b) The vector is then tailed with dG residues using terminal transferase. The insert DNA is tailed with dC residues in a similar way. (c) The dC and dG tails are complementary and the insert can therefore be annealed with the vector to generate a recombinant. The PstI sites are regenerated at the ends of the insert DNA, as shown.
LO 58: menjelaskan penggunaan linker, adaptor, dan homopolymer tailing

(a) The ds cDNA is treated with EcoRI methylase,
(b) which methylates any internal EcoRI recognition sequences.
(c) EcoRI linkers are then added to the ends of the methylated cDNA, and the linkers digested with EcoRI.
(d) The methylation prevents digestion at internal sites, and the result is a cDNA with EcoRI cohesive ends.
(e) This can be ligated into the EcoRI site of a λ vector such as λgt10.
LO 59: menjelaskan kloning cDNA ke dalam bakteriofaga
Cloning cDNA into bacteriophage
Fig. 6.7 Cloning cDNA in λ vectors using linkers

Cloning cDNA in phage vectors using linkers
LO 59: menjelaskan kloning cDNA ke dalam bakteriofaga

MINGGU DEPAN TATAP MUKA 7












![KLONING ekosari[Compatibility Mode]](https://img.pdfslide.net/doc/110x75/587b2fb61a28aba4788b4d3b/kloning-ekosaricompatibility-mode.jpg)
















