Embed Size (px)
Citation preview

1
Astrocyte derived CXCL10 drives Accumulation of Antibody Secreting Cells in the 1 Central Nervous System during Viral Encephalomyelitis 2
3 Timothy W. Phares1, Stephen A. Stohlman1, David R. Hinton2, and Cornelia C. 4 Bergmann1# 5 6 Department of Neurosciences, Lerner Research Institute, The Cleveland Clinic, 7 Cleveland, OH 44195, USA1. Department of Pathology, Keck School of Medicine, 8 University of Southern California, Los Angeles, CA 90033, USA2. 9 10 Running title: CXCL10 dependent ASC migration 11 Number of words in abstract: 245 12 Number of words in text: 5083 13 14 #Correspondence address: 15 Dr. Cornelia C. Bergmann 16 Department of Neuroscience 17 Lerner Research Institute 18 The Cleveland Clinic 19 9500 Euclid Avenue, NC30 20 Cleveland, OH 44195 21 Phone: 216-444-5922 22 Fax: 216-444-7927 23 E-mail: [email protected] 24
Copyright © 2013, American Society for Microbiology. All Rights Reserved.J. Virol. doi:10.1128/JVI.03307-12 JVI Accepts, published online ahead of print on 9 January 2013
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

2
ABSTRACT (245) 25 Microbial infections of the central nervous system (CNS) are often associated with local 26 accumulation of antibody secreting cells (ASC). By providing a source of antibody (Ab) 27 at the site of infection, CNS localized ASC play a critical role in acute viral control and 28 preventing viral recrudescence. Following coronavirus induced encephalomyelitis, CNS 29 accumulation of ASC is chemokine (C-X-C motif) receptor 3 (CXCR3) dependent. This 30 study demonstrates that CNS expression of the CXCR3 ligand CXCL10 is the critical 31 chemokine regulating ASC accumulation. Impaired ASC recruitment in CXCL10-/- but 32 not CXCL9-/- mice was consistent with reduced CNS IgG and κ-light chain mRNA as 33 well as virus specific Ab. Moreover, the few ASC recruited to the CNS in CXCL10-/- 34 mice were confined to the vasculature, distinct from parenchymal localization in wild-35 type and CXCL9-/- mice. However, neither CXCL9 nor CXCL10 deficiency diminished 36 neutralizing serum Ab, supporting a direct role for CXCL10 in ASC migration. T cell 37 accumulation, localization and effector functions were also not affected in either CXCL9-38 /- or CXCL10-/- mice, consistent with similar control of infectious virus. There was also 39 no evidence for dysregulation of chemokines or cytokines involved in ASC regulation. 40 The distinct roles of CXCL9 and CXCL10 in ASC accumulation rather coincided with 41 their differential localization. While CXCL10 was predominantly expressed by 42 astrocytes, CXCL9 expression was confined to the vasculature/perivascular spaces. 43 These results suggest that CXCL10 is critical for two phases: recruitment of ASC to the 44 CNS vasculature, as well as ASC entry into the CNS parenchyma. 45 46 47
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

3
INTRODUCTION 48 The site of infection and access of neutralizing antibody (Ab) contribute to viral 49 control during primary as well as secondary infections. This is particularly important in 50 the central nervous system (CNS), which is a target for numerous acute encephalitic viral 51 infections as well as a reservoir of latent/persistent infections (27). Sequestration from 52 the circulation by the specialized structure of the blood brain barrier makes Ab passage 53 from the serum to the CNS inefficient. While control of acute neurotropic infections of 54 the CNS is largely T cell mediated, local Ab, produced by Ab-secreting cells (ASC), is 55 required for ultimate clearance and/or prevention of viral recrudescence (23, 36, 41, 75). 56 Continuous intrathecal Ab production thus provides an effective nonlytic mechanism of 57 sustained viral control within the CNS. This is supported by complement independent 58 Ab-mediated control of mouse hepatitis virus, Sindbis and rabies virus (35, 40, 50). The 59 CNS as a survival niche for ASC is well established in experimental animal models as 60 well as humans. However, the mechanisms of ASC accumulation and replenishment may 61 vary in distinct inflammatory conditions and involve both direct ASC migration from the 62 periphery as well as ectopic follicle formation (46, 58, 69, 74, 75). 63 As B cells differentiate into ASC and emigrate from lymphoid follicles they 64 upregulate surface expression of CXCR4 and CXCR3. While CXCR4 interactions with 65 its ligand, CXCL12, mediates homing to and survival of ASC in the bone marrow (18), 66 CXCR3 signaling is associated with ASC migration to inflamed tissue, including the 67 CNS (52). The CXCR3 ligands CXCL9, CXCL10 and CXCL11 are strongly induced in 68 the CNS following viral infection as well as during non microbial inflammation (12, 16, 69 19, 34, 37, 39, 53, 78). An essential role for CXCR3 in ASC accumulation in the CNS 70
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

4
was recently demonstrated using CXCR3 deficient (CXCR3-/-) mice infected with 71 gliatropic as well as dual hepato and neurotropic strains of coronavirus (25, 49). Both 72 virus strains cause acute encephalomyelitis which resolves into a persistent infection 73 associated with chronic, ongoing immune mediated myelin loss (9, 25). Although control 74 of infectious virus in the CNS is T cell mediated (67, 71, 72), continuously produced 75 antiviral IgG within the CNS is required to prevent viral recrudescence (41, 49, 51). 76 CXCR3 deficiency did not impair T cell mediated antiviral activity or serum neutralizing 77 Ab, but severely abrogated virus specific ASC accumulation in the CNS (49). 78 The present study sought to define the relative contribution of CXCR3 ligand(s) 79 CXCL9 and CXCL10 to ASC accumulation and regional localization. During infection 80 with the gliatropic JHM variant V2.2-1 of mouse hepatitis virus (JHMV) CXCL10 and to 81 a lesser extent CXCL9 mRNA expression is sustained after initial viral control (49, 59), 82 although both peak concomitant with peak T cell infiltration and gamma interferon (IFN-83 γ) secretion (60, 68). Neither JHMV infected CXCL9-/- nor CXCL10-/- mice exhibited 84 deficits in virus specific serum Ab or CNS T cell activity, similar to CXCR3-/- mice (49). 85 However, both ASC, as well as virus specific IgG, within the CNS were significantly 86 impaired in CXCL10-/-, but not CXCL9-/- mice. Furthermore, the few ASC recruited to 87 the CNS in CXCL10-/- mice, were confined to the vasculature, distinct from their 88 parenchymal distribution in wild-type (WT) and CXCL9-/- mice. The parenchymal 89 accumulation of ASC in WT mice was directly linked to predominant CXCL10 90 expression by astrocytes, while CXCL9 expression was confined to the microvasculature. 91 These results demonstrate a prominent, nonredundant role of CXCL10 in both ASC 92
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

5
recruitment as well as entry into the inflamed CNS parenchyma during viral 93 encephalomyelitis. 94 MATERIALS AND METHODS 95
Mice, Virus Infection and Virus Titers. C57BL/6 mice were purchased from 96 the National Cancer Institute (Frederick, MD). CXCL9-/-, CXCL10-/- and CXCR3-/- mice 97 on the C57BL/6 background were previously described (49, 66). All mice were housed 98 under pathogen free conditions at an accredited facility at the Cleveland Clinic Lerner 99 Research Institute. Mice were infected at 6-7 wks of age by intracranial injection with 100 1000 PFU of the gliatropic JHMV variant (21). Animals were scored daily for clinical 101 signs of disease with: 0, healthy; 1, ruffled fur and hunched back; 2, hind limb paralysis 102 or inability to turn to upright position; 3, complete hind limb paralysis and wasting; 4, 103 moribund or dead. All animal experiments were performed as per guidelines approved by 104 the Cleveland Clinic Lerner Research Institute Institutional Animal Care and Use 105 Committee. Virus titers within the CNS were determined in clarified supernatants by 106 plaque assay using the murine delayed brain tumor (DBT) astrocytoma as described (21). 107 Plaques were counted after 48 hour (hr) incubation at 37°C. 108 109
Gene Expression and Cytokine Analysis. Snap frozen spinal cords from 110 individual mice (n = 3-9) were placed into 1 ml Trizol (Invitrogen, Grand Island, NY) 111 and homogenized using a TissueLyser and stainless steel beads (Qiagen, Valencia, CA). 112 RNA was extracted according to the manufacturer's instructions. DNA contamination 113 was removed by treatment with DNase I for 30 minutes (min) at 37°C (DNA-free kit, 114 Ambion, Austin, TX) and cDNA was synthesized from RNA using M-MLV Reverse 115
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

6
Transcriptase (Invitrogen), oligo-dT primers and random primers (Promega, Madison, 116 WI). Quantitative real-time PCR was performed using 4 μl of cDNA and SYBR Green 117 Master Mix (Applied Biosystems, Foster City, CA) in duplicate on a 7500 Fast Real-118 Time PCR System (Applied Biosystems). PCR conditions were 10 min at 95°C followed 119 by 40 cycles at 95°C for 15 seconds (s), 60°C for 30 s and 72°C for 30 s. Real-time 120 primers for transcripts encoding APRIL, BCMA, GAPDH, JHMV nucleocapsid, IL-10, 121 IL-21, TACI, CXCL9, CXCL10, CXCL11 and CXCL12 were previously described (59, 122 62). GAPDH, IFN-γ, CXCL13, BAFF, CCL19, CCL21 and IgG mRNA levels were 123 determined using Applied Biosystems Gene Expression Arrays with Universal Taqman 124 Fast Master Mix (Applied Biosystems) in duplicate. Primer and probe sequences for κ-125 light chain mRNA detection were as described (58). PCR conditions were 20 s at 95°C 126 followed by 40 cycles at 95°C for 3 s and 60°C for 30 s. Transcript levels were 127 calculated relative to the housekeeping gene GAPDH using the following formula: 128 2[CT(GAPDH) – CT(Target Gene)] x 1000, where CT is determined as the threshold cycle at which 129 the fluorescent signal becomes significantly higher than background. 130
CXCL9 and CXCL10 protein levels in clarified CNS supernatants were assayed 131 by ELISA kits (MCX900 and SMCX100) according to the procedures supplied by the 132 manufacturer (R&D Systems, Minneapolis, MN). Optical densities were read at 450nm 133 in a Bio-Rad Model 680 microplate reader and analyzed using Microplate Manager 5.2 134 software (Bio-Rad Laboratories, Hercules, CA). 135
136 Serum and CNS Ab. Virus specific IgG within the CNS was determined in 137
clarified spinal cord supernatants by ELISA as described (49). Briefly, 96 well plates 138
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

7
were coated with 100 µl of a serum free supernatant derived from JMHV infected DBT 139 cells and incubated overnight at 4°C. Plates were washed with phosphate buffered saline 140 (PBS)-Tween 20 and non-specific binding was blocked with 10% fetal calf serum (FCS) 141 in PBS overnight at 4°C. Samples were added and incubated overnight at 4°C. After 142 washes bound IgM and IgG were detected using biotinylated goat anti-mouse IgM 143 (Jackson ImmunoResearch, West Grove, PA) or goat anti-mouse IgG2a (Southern 144 Biotech, Birmingham, AL). Secondary Ab was detected using Streptavidin HRP (BD 145 Bioscience) followed by 3,3’,5,5’ Tetramethylbenzidine (TMB Reagent Set; BD 146 Bioscience). Optical densities were read at 450nm in a Bio-Rad Model 680 microplate 147 reader and analyzed using Microplate Manager 5.2 software (Bio-Rad Laboratories). 148 Data are expressed as Arbitrary Units / Tissue where 1 Arbitrary Unit = 0.1 Absorbance 149 @ 450. Levels were calculated using the following formula: (Absorbance @ 450 / 0.1) x 150 dilution factor x volume of PBS used to homogenize spinal cord. Background levels 151 from naïve mice were subtracted. 152
Neutralizing Ab was measured as described previously (73). Briefly, triplicates 153 of serial 2-fold dilutions of heat inactivated serum from individual mice (n = 5) were 154 incubated with 50 PFU of JHMV in 96-well plates for 90 min at 37°C. DBT cells (8 × 155 104 cells/well) were then added, and plates were incubated at 37°C for 48 hr. 156 Neutralization titers represent the log10 of the highest average serum dilution that 157 inhibited cytopathic effect. 158 159
Histology. Spinal cords from PBS perfused mice were snap-frozen in Tissue-Tek 160 optimum cutting temperature compound (Sakura Finetek, Torrance, CA). Distribution of 161
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

8
CD138+ ASC, CD4+ and CD8+ T cells were determined by staining acetone-fixed, 10-μm 162 longitudinal cryosections with Ab specific for either CD138 (Abcam, Cambridge, MA), 163 CD4 or CD8 (BD, Bioscience), followed by anti-mouse or anti-rat IgG, respectively. 164 Immunoreactivity was detected using either the Vector Mouse on Mouse (M.O.M.) 165 peroxidase immunodetection kit (Vector Laboratories, Burlingame, CA) or Vectastain 166 ABC kit. Viral nucleocapsid protein was detected in paraffin embedded sections by 167 immunoperoxidase staining using anti-JHMV mouse mAb J.3.3 as described (61). 168 Digital images were captured using a Leica DMLB light microscope (Leica 169 Microsystems, Bannockburn, IL) equipped with a SPOT Insight camera (Columbia, IL). 170 Sections were scored in a blinded manner and representative fields identified based on 171 the average score of all sections in each experimental group. 172
For detection of CXCL9 and CXCL10 longitudinal sections were fixed with 4% 173 paraformaldehyde for 20 min, treated with 1% Triton X-100 in PBS at room temperature, 174 blocked for 30 min, and then stained with either rabbit anti-mouse laminin (Cedarlane 175 Laboratories, Ontario, Canada), rat anti-mouse CD138 (BD Bioscience), goat anti-mouse 176 CXCL9 (R&D Systems; AF-492-NA), goat anti-mouse CXCL10 (R&D Systems; AF-177 466-NA), rat anti-mouse glial fibrillary acidic protein (GFAP) (2.2B10, Invitogen), rabbit 178 anti-mouse ionized calcium binding adaptor molecule-1 (Iba-1) (Wako, Richmond, VA), 179 anti-mouse CD31 (BD Bioscience; 550274) or anti-JHMV monoclonal Ab overnight at 180 4°C. Secondary fluorescently labeled Ab were added, and the slides incubated for 1 hr at 181 room temperature. Sections were mounted with ProLong Gold antifade reagent with 182 4’,6-diamidino-2-phenylindole (DAPI) (Invitrogen) and analyzed using a Leica DM2500 183 microscope. 184
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

9
185 Flow cytometric analysis. For phenotypic analysis CNS derived mononuclear 186
cells were isolated from pooled brains (3-4 mice per timepoint) as described (61). 187 Expression of cell surface markers was determined by staining with Ab specific for CD4 188 (L3T4), CD8 (53-6.7) and CD45 (30-F11) (BD Bioscience). Virus specific CD8 T cells 189 were identified using H2-Db/S510 MHC class I tetramers (8). Cells were analyzed on a 190 FACSCalibur flow cytometer (BD, Mountain View, CA) using FlowJo 7.1 software 191 (Tree Star, Ashland, OR). Cell numbers were calculated based on cell yields and 192 percentages of gated live cells. 193 194
Statistical Analysis. Results are expressed as the mean + SEM for each group of 195 mice. In all cases, p < 0.05 was considered significant determined by unpaired t test. 196 Graphs were plotted and statistics assessed using GraphPad Prism 3.0 software. 197 198 on A
pril 3, 2019 by guesthttp://jvi.asm
.org/D
ownloaded from

10
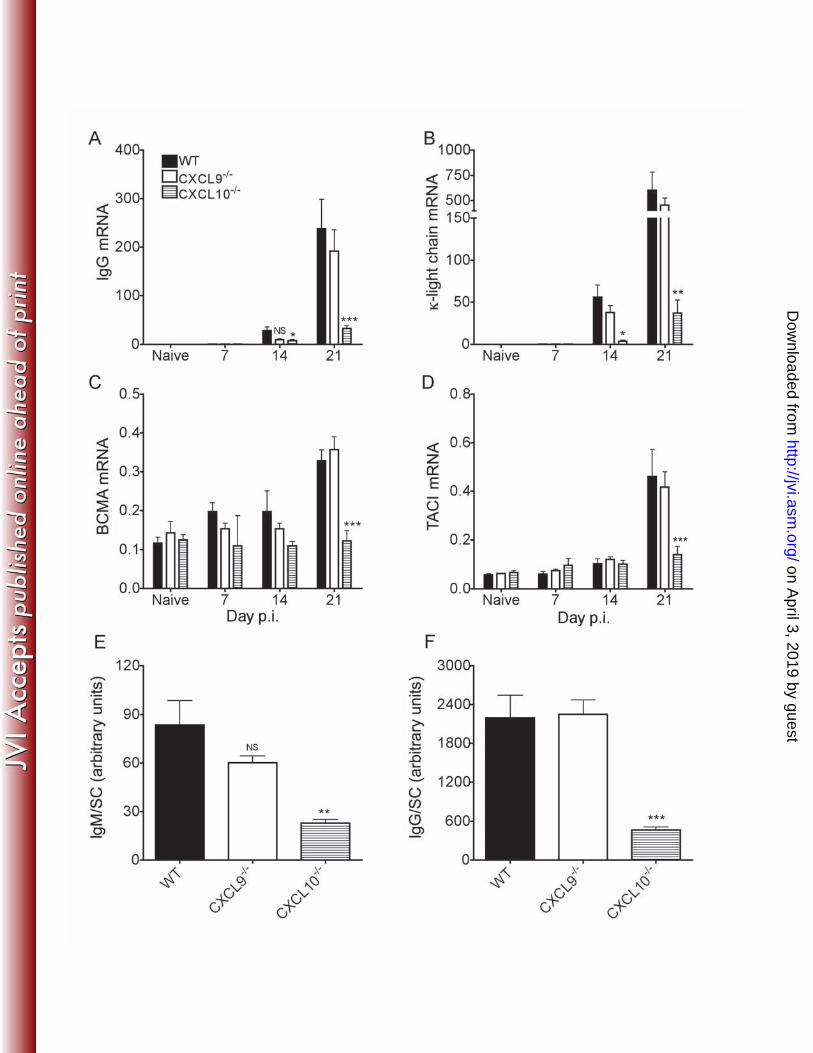
RESULTS 199 Impaired Ab production within the CNS of CXCL10-/- mice. ASC recruitment 200 into the CNS is CXCR3 dependent in C57BL/6 mice following infection with a gliatropic 201 as well as a dual hepato and neurotropic viral strain of coronavirus (25, 49). To 202 determine if the CXCR3 ligands CXCL9 and CXCL10 play interchangeable roles in 203 mediating ASC migration, gliatropic JHMV infected CXCL9-/- and CXCL10-/- mice were 204 analyzed for humoral responses in the periphery and CNS. Neutralizing Ab titers in 205 serum were similar between infected WT, CXCL9-/- and CXCL10-/- mice (Table I) 206 ensuring that neither CXCL9 nor CXCL10 deficiency affected peripheral ASC activation 207 following JHMV infection, similar to CXCR3-/- mice (49). Based on the high Ig heavy 208 and light chain expression in ASC compared to non Ab secreting B cells, IgG and κ-light 209 chain transcripts were assessed in the CNS. Total IgG and κ-light chain mRNA were 210 barely detectable and similarly low at day 7 post-infection (p.i.) in all groups (Fig. 1A 211 and B; Fig. 1 Legend). Both transcripts increased in all groups by day 14 p.i., although 212 the increase was more modest in CXCL10-/- mice. By day 21 p.i. mRNA further 213 increased by ~10-fold in WT and CXCL9-/- mice, but only reached 25% of WT values in 214 CXCL10-/- mice. Nevertheless, progressive low level ASC accumulation was evident by 215 similar transcript expression to those in WT mice at day 14 p.i. Moreover expression of 216 receptors that support ASC survival and differentiation, namely B cell maturation antigen 217 (BCMA) and transmembrane activator and calcium modulator ligand interactor (TACI) 218 (7, 14, 48, 55), were also significantly upregulated at day 21 p.i. in infected WT and 219 CXCL9-/- mice (Fig. 1C and D). By contrast, they remained near naïve levels throughout 220 infection in CXCL10-/- mice (Fig. 1C and D). The kinetics of mRNA expression thus 221
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

11
indicated similar ASC recruitment in WT and CXCL9-/- mice, but lagging levels in the 222 absence of CXCL10. To verify that Ig mRNA reflects reduced virus specific Ab 223 production, CNS supernatants from day 21 p.i. were assessed for virus specific IgM and 224 IgG. Virus specific IgM and IgG were similar between WT and CXCL9-/- mice (Fig. 1E 225 and F), with IgG exceeding IgM levels (49). By contrast, Ig levels were significantly 226 lower in CXCL10-/- mice. These data implied ASC migration and virus specific Ab 227 production within the CNS is CXCL10 dependent. 228 229 CXCL9 and CXCL10 do not alter CNS chemokine or cytokine expression. 230 To exclude the possibility of chemokine or cytokine dysregulation in CXCL10-/- and 231 CXCL9-/- mice, transcript levels of CXCR3 ligands, as well as other chemokines and 232 survival factors associated with B cell migration and survival were assessed. CXCL9 233 mRNA was ~50% lower in infected CXCL10-/- mice compared to WT mice at day 7 p.i. 234 (Fig. 2). However, the relative difference in CXCL9 expression subsequently 235 diminished, with indistinguishable CXCL9 mRNA in CXCL10-/- and WT mice by day 21 236 p.i., the peak of ASC CNS accumulation (49, 59, 73, 74) (Fig. 2). By contrast, CXCL10 237 transcripts in the CNS of infected CXCL9-/- mice showed no differences when compared 238 with infected WT mice (Fig. 2). Expression of the CXCR3 ligand CXCL11, although not 239 functional in C57BL/6 mice (52), was also similar in all groups of infected mice (Fig. 2). 240 Expression of CXCL12 mRNA, which regulates ASC recruitment and survival in bone 241 marrow (18) did not exceed basal levels throughout infection in any group (Fig. 2). 242 Potential alterations in the chemokines CXCL13, CCL19 and CCL21, all involved in 243 regulating B cell movement within lymphoid tissue and ectopic follicles (38), were also 244
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

12
assessed within the CNS. While infection increased CXCL19 mRNA to a similar extent 245 in all groups (Fig. 2), CCL21 mRNA appeared to drop transiently at 7 days p.i., but was 246 restored to basal levels by 14 days p.i. in all groups (Fig. 2). Unexpectedly however, 247 mRNA expression of CXCL13, which guides B cells into B cell zones within lymphoid 248 tissue (2), were 10- to 20-fold higher in both CXCL9-/- and CXCL10-/- compared to WT 249 mice (Fig. 2). Lastly, to determine whether CXCL10 deficiency negatively impacts 250 factors associated with ASC survival, transcripts encoding a proliferating-inducing ligand 251 (APRIL) and B cell-activating factor of the tumor necrosis factor family (BAFF) (6, 7, 252 14, 17, 65) were assessed. APRIL and BAFF mRNA increased in the CNS of infected 253 CXCL9-/- and CXCL10-/- mice with similar kinetics as in WT mice, with most extensive 254 upregulation of APRIL at day 14 p.i., and sustained elevated BAFF expression 255 throughout days 7-21 p.i. (Fig. 2). With the exception of elevated CXCL13, these results 256 indicate that expression of chemokines and survival factors involved in B cell and ASC 257 regulation are not altered in the absence of CXCL9 or CXCL10; moreover, CXCL9 258 cannot compensate for the absence of CXCL10 in ASC accumulation. 259 260
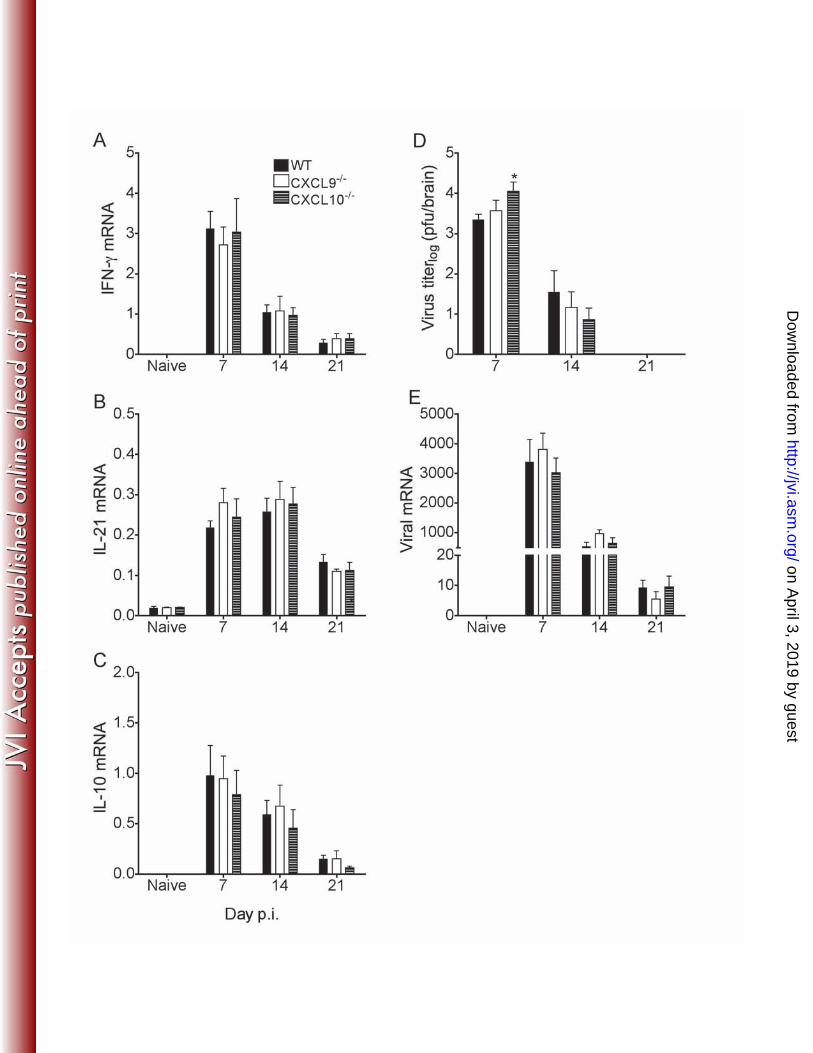
T cell function in the CNS does not require CXCL9 or CXCL10. A role for T 261 cells in promoting ASC recruitment to, and survival within the CNS is poorly explored. 262 Anti-CXCL10 Ab treatment reduced T cell recruitment following a heterologous 263 coronavirus infection, resulting in uncontrolled virus replication and death prior to ASC 264 recruitment (42). By contrast, CXCR3 deficiency did not impair T cell accumulation or 265 initial viral control in the CNS following the gliatropic virus infection used herein (49). 266 Nevertheless, CD4 T cell derived interleukin-21 (IL-21), a cytokine that sustains CD8 T 267
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

13
cell function and promotes ASC differentiation (4, 20, 24) was reduced by 50% in 268 CXCR3-/- mice relative to WT levels (49). To ascertain if defective ASC accumulation in 269 the absence of CXCL10 is due to impaired T cell function in the CNS, T cell infiltration 270 as well as antiviral and ASC promoting cytokines were analyzed. Flow cytometry 271 revealed similar CNS recruitment of CD4, total CD8 and virus specific CD8 T cells (data 272 not shown). Comparable transcript levels of antiviral IFN-γ, IL-21, and the anti-273 inflammatory and humoral promoting factor IL-10 (Fig. 3) further indicated that neither 274 the absence of CXCL9 or CXCL10 altered T cell functions in the CNS. Notably, during 275 JHMV infection IFN-γ is secreted by both CD4 and CD8 T cells (11, 57, 60), while IL-21 276 and IL-10 are produced by CD4 T cells (59, 62). Despite ~5-fold increased viral titers in 277 CXCL10-/- mice at day 7 p.i., infectious virus was controlled with similar kinetics and 278 efficiency in all groups by day 21 p.i. (Fig. 3D). Analysis of spinal cords, the primary 279 site of virus persistence, further revealed reductions of viral RNA in WT, CXCL9-/- and 280 CXCL10-/- mice by day 21 p.i. (Fig. 3E). Onset of encephalomyelitis and clinical disease 281 was similar in all groups (data not shown). These data demonstrate that CXCL10 282 promotes early virus control, but that neither the absence of CXCL9 or CXCL10 283 significantly impairs T cell migration or antiviral function in the CNS prior to ASC 284 accumulation. 285
286 CXCL9 and CXCL10 have regionally distinct localization in the CNS. The 287 apparently distinct functions of CXCL9 and CXCL10 in regulating ASC accumulation 288 were unanticipated based on both their abilities to promote ASC transmigration in vitro 289 (32). Differential in vivo recruitment may therefore be attributed to distinct quantities or 290
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

14
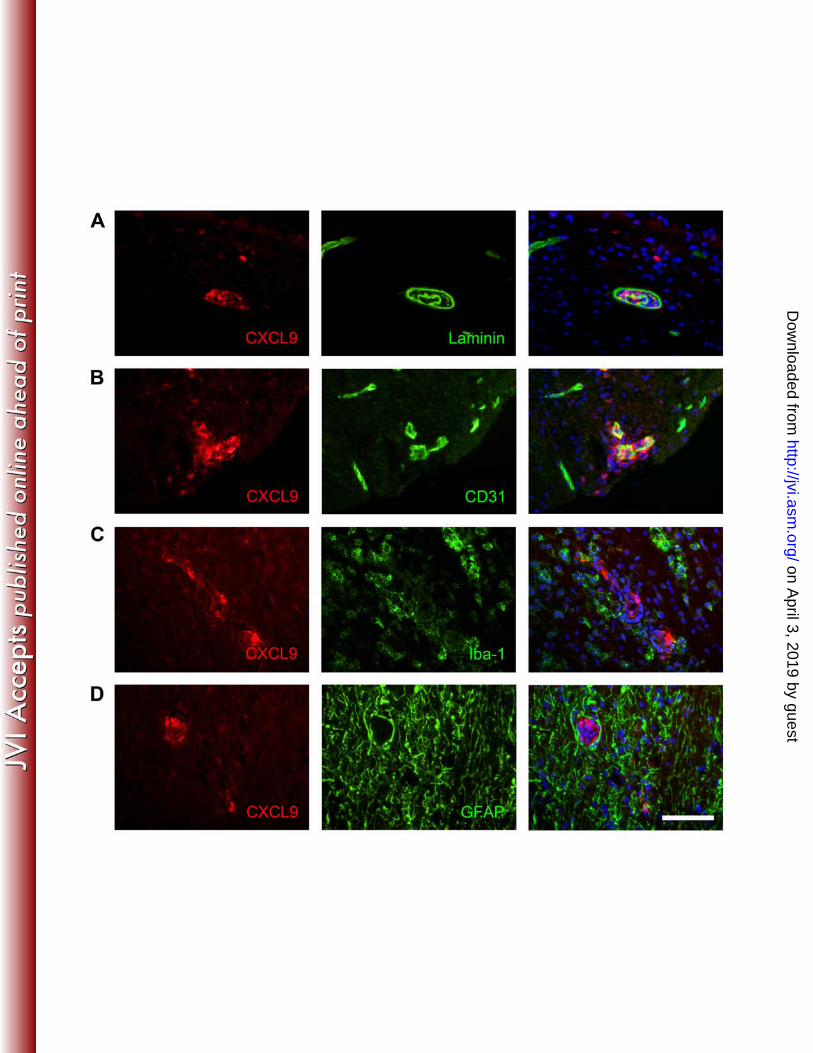
localization. CXCL10 mRNA expression was ~3-fold higher compared to CXCL9 291 mRNA in the CNS of WT mice at day 7 p.i.; however mRNA levels were similar at days 292 14 and 21 days p.i. (Fig. 2). Enhanced expression of CXCL10 at earlier timepoints may 293 therefore result in more regionally concentrated and prolonged presentation of CXCL10. 294 Measurement of both chemokines in clarified CNS supernatants by ELISA indicated 295 peak values at day 7 p.i., with overall levels of CXCL10 exceeding those of CXCL9 296 (Table II). However, both CXCL9 and CXCL10 protein declined rapidly by day 10 p.i., 297 distinct from sustained mRNA expression (Fig. 2). These data suggest chemokine 298 secretion is either rapidly reduced and/or extracellular chemokine is bound by 299 surrounding matrix or consumed and thus not available to detection by ELISA. CXCL9 300 and CXCL10 production in the CNS has been associated with different cellular sources 301 (12, 16, 19, 34, 37, 39, 53, 78), suggesting distinct anatomical presentation. CXCL10 302 mRNA is prominently associated with astrocytes during viral as well as non microbial 303 neuroinflammation (12, 16, 34), whereas CXCL9 mRNA is associated with microglia (4, 304 16, 19, 34). Thus, astrocyte mediated CXCL10 expression may provide a stronger signal 305 at the glia limitans to attract ASC into the parenchyma. To test this hypothesis, spinal 306 cords from JHMV infected mice were analyzed for chemokine expression, prior to 307 emergence of ASC at day 14 p.i. (59, 73, 74). CXCL10 was indeed most prominently 308 expressed in GFAP+ astrocytes at day 7 p.i., with limited expression by Iba-1+ 309 microglia/macrophages (Fig. 4). CXCL10 expression remained prominent in astrocytes 310 at day 10 p.i., coinciding with initial ASC recruitment, and was still detectable at day 21 311 p.i. Notably, during viral persistence at day 21 p.i., CD138+ ASC located proximal to 312 CXCL10+ astrocytes (data not shown). Lastly, the absence of CXCL10 expression by 313
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

15
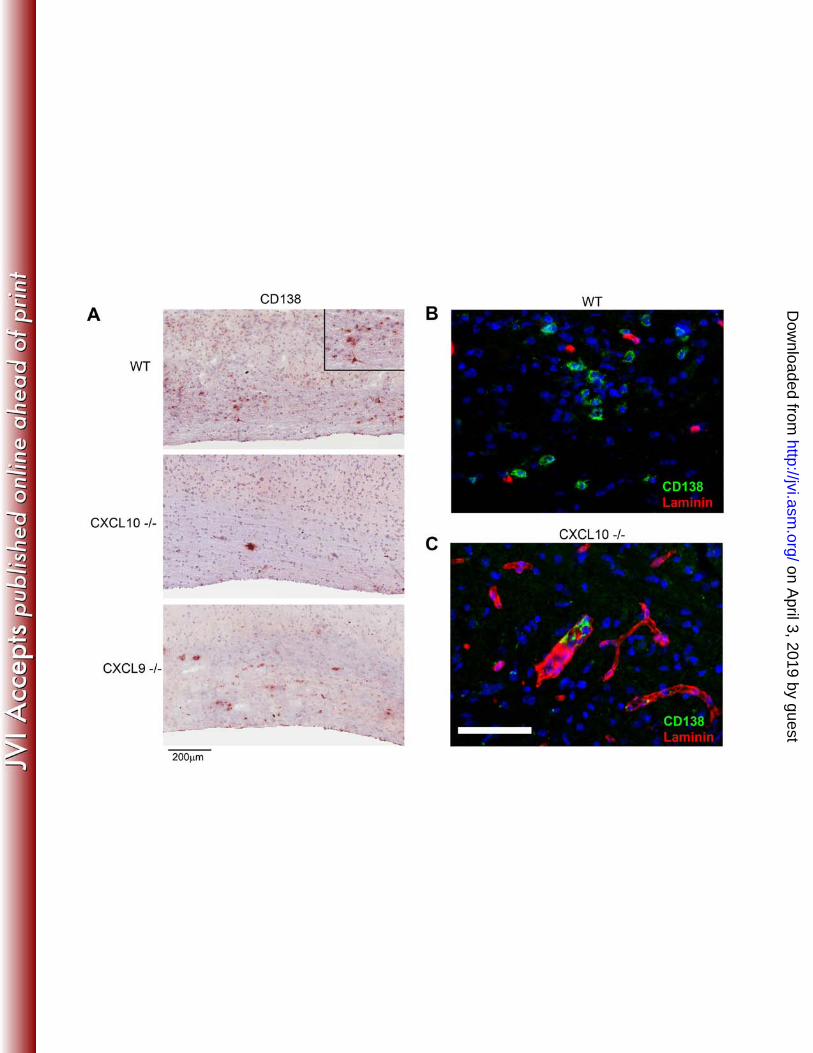
virus infected cells supported sparse astrocyte infection (33) and IFN mediated CXCL10 314 expression (44, 45, 56). In contrast to CXCL10, CXCL9 was exclusively associated with 315 blood vessels (Fig 5) and appeared contiguous with the endothelial matrix as shown by 316 dual labeling of CXCL9 and laminin (Fig. 5). The endothelial marker CD31 further 317 revealed that some, but not all CXCL9 colocalized with endothelial cells (Fig. 5). 318 CXCL9 was also detected in few perivascular Iba-1+ microglia /macrophages, but not in 319 parenchymal microglia/macrophages and did not colocalize with GFAP+ astrocytes (Fig. 320 5). These data clearly uncover highly restricted, nonoverlapping expression patterns of 321 CXCL9 and CXCL10 in vivo. 322 323 CXCL10 controls ASC trafficking into the CNS parenchyma. To determine 324 whether CXCL9 and CXCL10 deficiency affected the anatomical distribution of ASC, 325 spinal cords of persistently infected mice were analyzed for the frequency and 326 distribution of ASC. The number and distribution of CD138+ ASC were similar 327 comparing WT and CXCL9-/- mice (Fig. 6), with CD138+ ASC predominantly in 328 demyelinated lesions and adjacent white matter. By contrast, almost no CD138+ ASC 329 were detected in the white matter of CXCL10-/- mice (Fig. 6). Moreover, the few CD138+ 330 ASC were confined to the vasculature, often in small clusters (Fig. 6). Dual labeling for 331 CD138 and laminin confirmed that the few ASC present in CXCL10-/- mice were 332 perivascular (Fig. 6). Reduced numbers of ASC as well as vascular retention in the CNS 333 of CXCL10-/- mice suggests that CXCL10 is not only instrumental in recruiting ASC, but 334 also in mediating their migration into the parenchyma. 335
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

16
CXCR3 ligands also act as cues for T cell migration and prolonged interaction 336 with target cells within inflamed tissue (28, 29). However, neither the absence of 337 CXCL9 or CXCL10 affected the anatomical localization of CD4+ or CD8+ T cells within 338 the CNS during viral induced encephalomyelitis (Fig. 7). Similar to WT mice, both 339 populations were detected in the white matter (Fig. 7). Numbers of virally infected cells 340 and their distribution were also comparable in WT and CXCL10-/- mice (data not shown), 341 consistent with similar viral mRNA (Fig. 3). Essentially all positive cells were detected 342 in the white matter and exhibited small round morphology consistent in appearance with 343 oligodendrocytes (data not shown). These data suggest that differences in ASC 344 localization were not attributed to either distinct distribution of viral antigen or T cells. 345 346
Impaired, yet sustained Ig expression in CXCL10-/- mice coincides with 347 ongoing virus control. Ongoing viral control in the CNS of infected CXCL10-/- mice, 348 despite significantly impaired ASC accumulation, differs from the lack of control in 349 CXCR3-/- mice. Virus specific IgG levels in the CNS were therefore directly compared to 350 determine if this discrepancy reflected higher local Ab production in CXCL10-/- relative 351 to CXCR3-/- mice. Indeed, IgG was ~6-fold higher in CXCL10-/- compared to CXCR3-/- 352 mice at 21 days p.i. (Fig. 8A). Similarly, CNS expression of IgG and κ-light chain 353 mRNA was increased 5- and 8-fold, respectively, in infected CXCL10-/- versus CXCR3-/- 354 mice (data not shown). These data indicate that virus specific IgG coincides with total Ig 355 mRNA expression. To assess whether the low, yet elevated levels of virus specific IgG 356 in CXCL10-/- versus CXCR3-/- mice are sufficient to sustain viral control WT and 357 CXCL10-/- mice were monitored during persistence. IgG and κ-light chain transcripts 358
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

17
dropped less than 2-fold in WT mice and were sustained at similarly low levels in 359 CXCL10-/- mice between days 21 and 63 p.i. (Fig. 8B and C). Persisting viral mRNA in 360 the spinal cord was comparable between WT and CXCL10-/- mice at day 21 p.i. and 361 progressively declined throughout day 63 p.i. in both groups (Fig. 8D). However after 35 362 days p.i., viral RNA transcripts declined more slowly in the absence of CXCL10 as 363 indicated by 6- and 8-fold higher viral mRNA at days 49 and 63 p.i. (Fig. 8D). Notably 364 however, infectious virus remained below detection in CXCL10-/- mice (data not shown). 365 These results suggest that few ASC can be highly effective at sustaining protective ASC 366 responses in the CNS. 367 368
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

18
DISCUSSION 369 Infections of the CNS in humans as well as experimental models are commonly 370 associated with the presence of effector T cells and ASC or elevated Ig in cerebrospinal 371 fluid. Although numerous studies have characterized chemokines and their receptors 372 involved in T cell accumulation in the CNS, the chemokines driving ASC recruitment to 373 the CNS are less well understood. Local ASC provide a protective role in reducing viral 374 replication, when T cells alone fail to eliminate virus. A better understanding of events 375 underlying ASC CNS accumulation may thus lead to more effective vaccination 376 strategies. CXCR3 was recently identified as the chemokine receptor mediating ASC 377 trafficking into the CNS in two different models of neurotropic coronavirus infection (25, 378 49). Herein, we demonstrate a dominant role of the CXCR3 ligand CXCL10 in 379 regulating CXCR3 mediated ASC recruitment to the CNS following infection with a 380 gliatropic coronavirus. The absence of CXCL10 not only diminished overall ASC 381 recruitment, but trapped the few migrating ASC within the CNS vasculature. These data 382 suggested CXCL10 acts both to recruit ASC to the vasculature and mediates entry into 383 the CNS parenchyma. 384
The preferential CNS recruitment of ASC by CXCL10 rather than CXCL9 385 correlated with their distinct localization patterns. CXCL10 colocalized primarily with 386 astrocytes and expression was more abundant and dispersed than CXCL9. CXCL10 387 production by astrocytes is consistent with in situ CXCL10 mRNA detection in astrocyte 388 during MHV infection as well as experimental allergic encephalomyelitis (16, 39). 389 Moreover, the inability to colocalize CXCL10 with JHMV infected cells, suggests 390 CXCL10 is induced in astrocytes in response to proximally secreted IFN-α/β, IFN-γ 391
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

19
and/or TNF (56). By contrast, during West Nile virus encephalitis, neurons are 392 predominant sources of CXCL10 and also the major cell type supporting virus replication 393 (37). CXCL10 is also in neurons during murine cerebral malaria, with only rare 394 expression by endothelial cells and no expression by astrocytes (15). However, the 395 prominent affect of CXCL10 on overall ASC recruitment, not only parenchymal entry, 396 suggests CXCL10 is also presented at vascular sites to promote local recruitment. The 397 mechanisms underlying distinct CXCL10 production in these models may reside in the 398 relative magnitudes of direct microbe-induced signals versus lymphocyte-derived 399 cytokines, such as IFN-γ. Thus, under conditions of high IFN-γ levels, astrocytes may be 400 the main CXCL10 producers, irrespective of virus tropism. By contrast, if local IFN-γ 401 production is less prominent the response of individually infected cell populations may 402 dominate. 403
In contrast to CXCL10, CXCL9 was only detected in the microvasculature. This 404 observation is consistent with CXCL9 detection on endothelial cells during cerebral 405 malaria (15), as well as vascular localization in R. rickettsii infection and primary CNS 406 lymphoma (76, 78). Vascular CXCL9 presentation also suggests local production of 407 IFN-γ, a major inducer of CXCL9, by early infiltrating perivascular CD4 T cells (68) 408 responding to perivascular antigen presenting cells (5). The association of CXCL9 with 409 perivascular cuffing further supports the concept that infiltrating cells may produce 410 CXCL9 for presentation by nearby endothelial cells (30). However, the inability to 411 detect CXCL9 in microglia/macrophages during JHMV infection was surprising as these 412 myeloid cells are capable of producing CXCL9 during CNS inflammation (16, 19, 34, 413 39). Furthermore, IFN-γ is abundantly expressed in the CNS during JHMV infection 414
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

20
(60) and sufficient to induce major histocompatibility complex class II on microglia and 415 major histocompatibility complex class I on oligodendrocytes (10, 31, 47). These data 416 suggest CXCL9 detection in microglia may be hampered by limited chemokine 417 production or rapid release. Indeed, preliminary analysis of purified microglia from 418 infected spinal cords revealed both CXCL9 and CXCL10 are upregulated at the 419 transcriptional level (data not shown), supporting a contribution of microglia to CXCR3 420 ligand production. The absence of CXCL9 expression by astrocytes is consistent with 421 their lack of PU.1, a key determinant for CXCL9 expression by myeloid cells (19). 422
Impaired ASC recruitment in the absence of CXCL10 could not be attributed to 423 reduced expression of other chemokines or cytokines known to affect ASC development 424 or survival. However, elevated CXCL13 in both CXCL9-/- and CXCL10-/-, but not 425 CXCR3-/- infected mice (49) were surprising, as CXCL13 upregulation is associated with 426 ectopic follicle formation and ASC accumulation during neuroborreliosis (54). While 427 CXCL13 is expressed by stromal cells in lymphoid tissue, both dendritic cells and 428 macrophages may be additional sources during inflammation (38). Although increased 429 CXCL13 suggested increased B cell recruitment, we did not detect ectopic follicle like 430 structures (data not shown). Moreover, similar and significantly decreased Ig production 431 in CXCL9-/- and CXCL10-/- mice, respectively, relative to WT mice, argues that CXCL13 432 does not promote ASC accumulation in this model, similar to results from Sindbis virus 433 infection and autoantigen induced CNS inflammation (63). 434
No adverse effects on virus control in CXCL9-/- and CXCL10-/- mice were 435 consistent with unaltered T cell function in the CNS. The distinct findings using 436 CXCL10 Ab mediated disruption may reside in the use of a different MHV variant and 437
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

21
dose, as well as experimental approaches (42, 43, 70). However, no evidence for either 438 loss of viral control during persistence or increased morbidity, despite significantly 439 reduced Ig production in infected CXCL10-/- relative to WT and CXCL9-/- mice was 440 unanticipated, given that virus was increased in CXCR3-/- mice (49). These contrasting 441 results may be partially explained by elevated virus specific Ab in the CNS of CXCL10-/- 442 relative to CXCR3-/- mice. Furthermore, the confined localization of T cells to white 443 matter, distinct from the widespread T cell distribution in CXCR3-/- mice may limit 444 disease severity (49, 53). Lastly, CXCR3 expression on neurons as well as microglia 445 may have physiological roles in modulating neuronal function, neuron-glia interactions 446 and neuroprotection (1, 3, 13, 22, 64, 77, 79, 81). Specifically, CXCR3 expression on 447 neurons mediates increased susceptibility to neuronal death in vitro following West Nile 448 virus infection (81). While this would suggest a detrimental rather than protective 449 function of CXCR3 signaling on neurons, our data suggest CXCL9 and CXCL10 do not 450 play a major role or compensate for each other in influencing CNS resident glia or 451 neurons during JHMV infection. 452 In summary, the results demonstrate a crucial role for CXCL10 in promoting ASC 453 accumulation within the CNS during a viral infection. Astrocytes as prominent sources 454 of CXCL10, as well as ASC survival factors BAFF and APRIL may further be 455 strategically relevant for focal ASC retention. Whether activated astrocytes, play 456 additional roles in directly promoting B cell differentiation, as recently described for 457 CXCL10 expressing macrophages (80), remains to be investigated. Overall the data 458 highlight distinct regulation of T cell and ASC migration to the CNS and provide novel 459
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

22
insights into intervention strategies to increase protective humoral immunity during CNS 460 infections or impede detrimental B cells implicated in autoimmunity (26). 461 462 ACKNOWLEDGMENTS 463 This work was supported by U.S. National Institutes of Health grants NS064932 and AI 464 47249. We sincerely thank Dr Shabbir Hussain for his contributions in evaluating 465 pathogenesis, gene expression analysis, and flow cytometry. Wenqiang Wei, Eric Barron 466 and Ernesto Barron provided excellent technical assistance with immunohistochemistry. 467
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

23
FIGURE LEGENDS 468 Figure 1. Ab production within the CNS is impaired in the absence of CXCL10. 469 Relative transcript levels of IgG (A), κ-light chain (B), BCMA (C) and TACI (D) in 470 spinal cords of naïve and infected WT, CXCL9-/- and CXCL10-/- mice assessed by real-471 time PCR. Data depict the mean ± SEM relative to GAPDH mRNA of two to three 472 independent experiments with at least 3 mice per timepoint per experiment. Significant 473 differences between WT and CXCL9-/- or CXCL10-/- mice were denoted by *, P < 0.05 474 **, P < 0.005 and ***, P < 0.001. NS = not significant, P > 0.05. Day 7 p.i. expression 475 levels were as follows: IgG, WT 0.26 ± 0.09, CXCL9-/- 0.17 ± 0.04, CXCL10-/- 0.17 ± 476 0.03 (A); κ-light chain, WT 0.45 ± 0.07, CXCL9-/- 0.48 ± 0.10, CXCL10-/- 0.63 ± 0.03 477 (B). (E and F) Virus specific IgM and IgG2a levels in clarified supernatants from spinal 478 cord homogenates at day 21 p.i. assessed by ELISA. Arbitrary units reflect total Ab 479 levels per spinal cord from 4-6 mice. Significant differences between WT and CXCL9-/- 480 or CXCL10-/- mice denoted by **, P < 0.005 and ***, P < 0.001. NS = not significant, P 481 > 0.05. 482 483 Figure 2. Chemokine and cytokine expresssion in spinal cords of infected WT, 484 CXCL9-/- and CXCL10-/- mice. Relative transcript levels in spinal cords of naïve and 485 infected WT, CXCL9-/- and CXCL10-/- mice assessed by real-time PCR. Data depict the 486 mean ± SEM relative to GAPDH mRNA of two to three independent experiments with at 487 least 3 mice per timepoint per experiment. Significant differences between WT and 488 CXCL9-/- or CXCL10-/- mice denoted by *, P < 0.05 and ***, P < 0.001. NS = not 489 significant, P > 0.05. 490
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

24
491 Figure 3. CXCL9 and CXCL10 independent antiviral T function in the CNS. 492 Relative transcript levels of IFN-γ (A), IL-21 (B) and IL-10 (C) in spinal cords of naïve 493 and infected WT, CXCL9-/- and CXCL10-/- mice assessed by real-time PCR. Data depict 494 the mean ± SEM relative to GAPDH mRNA of two to three independent experiments 495 with at least 3 mice per timepoint per experiment. (D) Viral titers determined by plaque 496 assay expressed as the mean ± SEM of two independent experiments with at least 3 mice 497 per timepoint per experiment. Data are on a log10
scale. Significant differences in viral 498 titers between WT and CXCL10-/- mice denoted by *, P < 0.05. (E) Viral nucleocapsid 499 mRNA in spinal cords determined by real-time PCR. Data are the mean + SEM relative 500 to GAPDH mRNA of two to three independent experiments with at least 3 mice per 501 timepoint per experiment. 502 503 Figure 4. Astrocytes express CXCL10 during JHMV infection. Spinal cord sections 504 from infected WT mice at day 7 p.i. analyzed for CXCL10 expression (A-C left panels). 505 CXCL10+ staining identified with anti-CXCL10 Ab (red). Astrocytes (A middle panel), 506 microglia/macrophages (B middle panel) and viral nucleocapsid protein (C middle panel) 507 were visualized with anti-GFAP, anti-Iba-1 or anti-JHMV J.3.3 Ab, respectively (green). 508 DAPI staining (blue) was used to identify cell nuclei. Merge images are shown in far 509 right panels. Bar, 50 μm. 510 511 Figure 5. CXCL9 is primarily detected at blood vessels. Spinal cord sections from 512 infected WT mice at day 7 p.i. analyzed for CXCL9 expression (A-D left panels). 513
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

25
CXCL9+ staining identified with anti-CXCL9 Ab (red). Laminin (A middle panel), 514 endothelium (B middle panel), microglia/macrophages (C middle panel) and astrocytes 515 (D middle panel) were visualized with anti-laminin, anti-CD31 anti-Iba-1 Ab or anti-516 GFAP respectively (green). DAPI staining (blue) was used to identify cell nuclei. Merge 517 images are shown in far right panels. Bar, 50 μm. 518 519 Figure 6. Parenchymal trafficking of CD138+ ASC is specifically impaired in 520 CXCL10-/- mice. (A) Longitudinal sections of spinal cords from infected WT (day 28 521 p.i.), CXCL9-/- (day 28 p.i.) and CXCL10-/- (day 25 p.i.) mice analyzed for CD138+ cells. 522 CD138+ cells identified with anti-CD138 Ab (brown) with hematoxylin counterstain. 523 Panels represent areas reflecting the average blinded score value of each group. (B and 524 C) Longitudinal sections of spinal cords from infected WT and CXCL10-/- mice at day 21 525 p.i. analyzed for CD138+ cells and laminin. CD138+ cells identified with anti-CD138 Ab 526 (green). Laminin visualized with anti-laminin Ab (red). DAPI staining (blue) was used to 527 identify cell nuclei. Merge images are shown. Bar, 50 μm. 528 529 Figure 7. T cell trafficking within the CNS is CXCL9 and CXCL10 independent. 530 Longitudinal sections of spinal cords from infected WT (day 28 p.i.), CXCL9-/- (day 28 531 p.i.) and CXCL10-/- (day 25 p.i.) mice analyzed for CD4+ and CD8+ T cell distribution 532 using anti-CD4 or anti-CD8 Ab (brown) with hematoxylin counterstain. Bar, 200 μm. 533 534 Figure 8. Local CNS Ab production is less impaired in CXCL10-/- compared to 535 CXCR3-/- mice. (A) Virus specific IgG2a in clarified supernatants from spinal cord 536
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

26
homogenates of infected WT, CXCL10-/- and CXCR3-/- mice at day 21 p.i. assessed by 537 ELISA. Arbitrary units reflect total Ab levels per spinal cord from three individual mice 538 per timepoint. Significant differences between WT and CXCL10-/- mice denoted by ***, 539 P < 0.001. Significant differences between CXCL10-/- and CXCR3-/- mice denoted by 540 ###, P < 0.001. (B-D) Relative transcript levels of IgG (B), κ-light chain (C) and viral 541 nucleocapsid mRNA (D) in spinal cords of infected WT and CXCL10-/- mice assessed by 542 real-time PCR at the indicated timepoint. Data depict the mean ± SEM relative to 543 GAPDH mRNA with at least 3 mice per timepoint. Significant differences between viral 544 nucleocapsid expression between WT and CXCL10-/- mice denoted by *, P < 0.05. 545 546
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

27
REFERENCES 547 1. Ambrosini, E., and F. Aloisi. 2004. Chemokines and glial cells: a complex 548
network in the central nervous system. Neurochemical research 29:1017-1038. 549 2. Ansel, K. M., V. N. Ngo, P. L. Hyman, S. A. Luther, R. Forster, J. D. 550
Sedgwick, J. L. Browning, M. Lipp, and J. G. Cyster. 2000. A chemokine-551 driven positive feedback loop organizes lymphoid follicles. Nature 406:309-314. 552
3. Bajetto, A., R. Bonavia, S. Barbero, and G. Schettini. 2002. Characterization 553 of chemokines and their receptors in the central nervous system: 554 physiopathological implications. J Neurochem 82:1311-1329. 555
4. Barker, B. R., M. N. Gladstone, G. O. Gillard, M. W. Panas, and N. L. 556 Letvin. 2010. Critical role for IL-21 in both primary and memory anti-viral CD8+ 557 T-cell responses. Eur J Immunol 40:3085-3096. 558
5. Bartholomaus, I., N. Kawakami, F. Odoardi, C. Schlager, D. Miljkovic, J. W. 559 Ellwart, W. E. Klinkert, C. Flugel-Koch, T. B. Issekutz, H. Wekerle, and A. 560 Flugel. 2009. Effector T cell interactions with meningeal vascular structures in 561 nascent autoimmune CNS lesions. Nature 462:94-98. 562
6. Belnoue, E., M. Pihlgren, T. L. McGaha, C. Tougne, A. F. Rochat, C. Bossen, 563 P. Schneider, B. Huard, P. H. Lambert, and C. A. Siegrist. 2008. APRIL is 564 critical for plasmablast survival in the bone marrow and poorly expressed by 565 early-life bone marrow stromal cells. Blood 111:2755-2764. 566
7. Benson, M. J., S. R. Dillon, E. Castigli, R. S. Geha, S. Xu, K. P. Lam, and R. 567 J. Noelle. 2008. Cutting edge: the dependence of plasma cells and independence 568 of memory B cells on BAFF and APRIL. J Immunol 180:3655-3659. 569
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

28
8. Bergmann, C. C., J. D. Altman, D. Hinton, and S. A. Stohlman. 1999. Inverted 570 immunodominance and impaired cytolytic function of CD8+ T cells during viral 571 persistence in the central nervous system. J Immunol 163:3379-3387. 572
9. Bergmann, C. C., T. E. Lane, and S. A. Stohlman. 2006. Coronavirus infection 573 of the central nervous system: host-virus stand-off. Nature reviews. Microbiology 574 4:121-132. 575
10. Bergmann, C. C., B. Parra, D. R. Hinton, R. Chandran, M. Morrison, and S. 576 A. Stohlman. 2003. Perforin-mediated effector function within the central 577 nervous system requires IFN-gamma-mediated MHC up-regulation. J Immunol 578 170:3204-3213. 579
11. Bergmann, C. C., B. Parra, D. R. Hinton, C. Ramakrishna, K. C. Dowdell, 580 and S. A. Stohlman. 2004. Perforin and gamma interferon-mediated control of 581 coronavirus central nervous system infection by CD8 T cells in the absence of 582 CD4 T cells. J Virol 78:1739-1750. 583
12. Bhowmick, S., R. Duseja, S. Das, M. B. Appaiahgiri, S. Vrati, and A. Basu. 584 2007. Induction of IP-10 (CXCL10) in astrocytes following Japanese encephalitis. 585 Neuroscience letters 414:45-50. 586
13. Biber, K., I. Dijkstra, C. Trebst, C. J. De Groot, R. M. Ransohoff, and H. W. 587 Boddeke. 2002. Functional expression of CXCR3 in cultured mouse and human 588 astrocytes and microglia. Neuroscience 112:487-497. 589
14. Bossen, C., T. G. Cachero, A. Tardivel, K. Ingold, L. Willen, M. Dobles, M. 590 L. Scott, A. Maquelin, E. Belnoue, C. A. Siegrist, S. Chevrier, H. Acha-591 Orbea, H. Leung, F. Mackay, J. Tschopp, and P. Schneider. 2008. TACI, 592
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

29
unlike BAFF-R, is solely activated by oligomeric BAFF and APRIL to support 593 survival of activated B cells and plasmablasts. Blood 111:1004-1012. 594
15. Campanella, G. S., A. M. Tager, J. K. El Khoury, S. Y. Thomas, T. A. 595 Abrazinski, L. A. Manice, R. A. Colvin, and A. D. Luster. 2008. Chemokine 596 receptor CXCR3 and its ligands CXCL9 and CXCL10 are required for the 597 development of murine cerebral malaria. Proc Natl Acad Sci U S A 105:4814-598 4819. 599
16. Carter, S. L., M. Muller, P. M. Manders, and I. L. Campbell. 2007. Induction 600 of the genes for Cxcl9 and Cxcl10 is dependent on IFN-gamma but shows 601 differential cellular expression in experimental autoimmune encephalomyelitis 602 and by astrocytes and microglia in vitro. Glia 55:1728-1739. 603
17. Cassese, G., S. Arce, A. E. Hauser, K. Lehnert, B. Moewes, M. Mostarac, G. 604 Muehlinghaus, M. Szyska, A. Radbruch, and R. A. Manz. 2003. Plasma cell 605 survival is mediated by synergistic effects of cytokines and adhesion-dependent 606 signals. J Immunol 171:1684-1690. 607
18. Cyster, J. G. 2003. Homing of antibody secreting cells. Immunological reviews 608 194:48-60. 609
19. Ellis, S. L., V. Gysbers, P. M. Manders, W. Li, M. J. Hofer, M. Muller, and I. 610 L. Campbell. 2010. The cell-specific induction of CXC chemokine ligand 9 611 mediated by IFN-gamma in microglia of the central nervous system is determined 612 by the myeloid transcription factor PU.1. J Immunol 185:1864-1877. 613
20. Elsaesser, H., K. Sauer, and D. G. Brooks. 2009. IL-21 is required to control 614 chronic viral infection. Science 324:1569-1572. 615
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

30
21. Fleming, J. O., M. D. Trousdale, F. A. el-Zaatari, S. A. Stohlman, and L. P. 616 Weiner. 1986. Pathogenicity of antigenic variants of murine coronavirus JHM 617 selected with monoclonal antibodies. J Virol 58:869-875. 618
22. Flynn, G., S. Maru, J. Loughlin, I. A. Romero, and D. Male. 2003. Regulation 619 of chemokine receptor expression in human microglia and astrocytes. J 620 Neuroimmunol 136:84-93. 621
23. Fragkoudis, R., C. M. Ballany, A. Boyd, and J. K. Fazakerley. 2008. In 622 Semliki Forest virus encephalitis, antibody rapidly clears infectious virus and is 623 required to eliminate viral material from the brain, but is not required to generate 624 lesions of demyelination. J Gen Virol 89:2565-2568. 625
24. Frohlich, A., J. Kisielow, I. Schmitz, S. Freigang, A. T. Shamshiev, J. Weber, 626 B. J. Marsland, A. Oxenius, and M. Kopf. 2009. IL-21R on T cells is critical 627 for sustained functionality and control of chronic viral infection. Science 628 324:1576-1580. 629
25. Gil-Cruz, C., C. Perez-Shibayama, S. Firner, A. Waisman, I. Bechmann, V. 630 Thiel, L. Cervantes-Barragan, and B. Ludewig. 2012. T helper cell- and CD40-631 dependent germline IgM prevents chronic virus-induced demyelinating disease. 632 Proc Natl Acad Sci U S A 109:1233-1238. 633
26. Gold, M., R. Pul, J. P. Bach, M. Stangel, and R. Dodel. 2012. Pathogenic and 634 physiological autoantibodies in the central nervous system. Immunological 635 reviews 248:68-86. 636
27. Griffin, D. E. 2003. Immune responses to RNA-virus infections of the CNS. Nat 637 Rev Immunol 3:493-502. 638
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

31
28. Groom, J. R., and A. D. Luster. 2011. CXCR3 in T cell function. Experimental 639 cell research 317:620-631. 640
29. Groom, J. R., and A. D. Luster. 2011. CXCR3 ligands: redundant, collaborative 641 and antagonistic functions. Immunology and cell biology 89:207-215. 642
30. Guarda, G., M. Hons, S. F. Soriano, A. Y. Huang, R. Polley, A. Martin-643 Fontecha, J. V. Stein, R. N. Germain, A. Lanzavecchia, and F. Sallusto. 2007. 644 L-selectin-negative CCR7- effector and memory CD8+ T cells enter reactive 645 lymph nodes and kill dendritic cells. Nat Immunol 8:743-752. 646
31. Hamo, L., S. A. Stohlman, M. Otto-Duessel, and C. C. Bergmann. 2007. 647 Distinct regulation of MHC molecule expression on astrocytes and microglia 648 during viral encephalomyelitis. Glia 55:1169-1177. 649
32. Hauser, A. E., G. F. Debes, S. Arce, G. Cassese, A. Hamann, A. Radbruch, 650 and R. A. Manz. 2002. Chemotactic responsiveness toward ligands for CXCR3 651 and CXCR4 is regulated on plasma blasts during the time course of a memory 652 immune response. J Immunol 169:1277-1282. 653
33. Hindinger, C., J. M. Gonzalez, C. C. Bergmann, B. Fuss, D. R. Hinton, R. D. 654 Atkinson, W. B. Macklin, and S. A. Stohlman. 2005. Astrocyte expression of a 655 dominant-negative interferon-gamma receptor. Journal of neuroscience research 656 82:20-31. 657
34. Hofer, M. J., S. L. Carter, M. Muller, and I. L. Campbell. 2008. Unaltered 658 neurological disease and mortality in CXCR3-deficient mice infected 659 intracranially with lymphocytic choriomeningitis virus-Armstrong. Viral 660 Immunol 21:425-433. 661
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

32
35. Hooper, D. C., K. Morimoto, M. Bette, E. Weihe, H. Koprowski, and B. 662 Dietzschold. 1998. Collaboration of antibody and inflammation in clearance of 663 rabies virus from the central nervous system. J Virol 72:3711-3719. 664
36. Hooper, D. C., T. W. Phares, M. J. Fabis, and A. Roy. 2009. The production of 665 antibody by invading B cells is required for the clearance of rabies virus from the 666 central nervous system. PLoS Negl Trop Dis 3:e535. 667
37. Klein, R. S., E. Lin, B. Zhang, A. D. Luster, J. Tollett, M. A. Samuel, M. 668 Engle, and M. S. Diamond. 2005. Neuronal CXCL10 directs CD8+ T-cell 669 recruitment and control of West Nile virus encephalitis. J Virol 79:11457-11466. 670
38. Lalor, S. J., and B. M. Segal. 2010. Lymphoid chemokines in the CNS. J 671 Neuroimmunol 224:56-61. 672
39. Lane, T. E., V. C. Asensio, N. Yu, A. D. Paoletti, I. L. Campbell, and M. J. 673 Buchmeier. 1998. Dynamic regulation of alpha- and beta-chemokine expression 674 in the central nervous system during mouse hepatitis virus-induced demyelinating 675 disease. J Immunol 160:970-978. 676
40. Levine, B., J. M. Hardwick, B. D. Trapp, T. O. Crawford, R. C. Bollinger, 677 and D. E. Griffin. 1991. Antibody-mediated clearance of alphavirus infection 678 from neurons. Science 254:856-860. 679
41. Lin, M. T., D. R. Hinton, N. W. Marten, C. C. Bergmann, and S. A. 680 Stohlman. 1999. Antibody prevents virus reactivation within the central nervous 681 system. J Immunol 162:7358-7368. 682
42. Liu, M. T., B. P. Chen, P. Oertel, M. J. Buchmeier, D. Armstrong, T. A. 683 Hamilton, and T. E. Lane. 2000. The T cell chemoattractant IFN-inducible 684
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

33
protein 10 is essential in host defense against viral-induced neurologic disease. J 685 Immunol 165:2327-2330. 686
43. Liu, M. T., H. S. Keirstead, and T. E. Lane. 2001. Neutralization of the 687 chemokine CXCL10 reduces inflammatory cell invasion and demyelination and 688 improves neurological function in a viral model of multiple sclerosis. J Immunol 689 167:4091-4097. 690
44. Luster, A. D., and J. V. Ravetch. 1987. Biochemical characterization of a 691 gamma interferon-inducible cytokine (IP-10). J Exp Med 166:1084-1097. 692
45. Luster, A. D., R. L. Weinshank, R. Feinman, and J. V. Ravetch. 1988. 693 Molecular and biochemical characterization of a novel gamma-interferon-694 inducible protein. J Biol Chem 263:12036-12043. 695
46. Magliozzi, R., S. Columba-Cabezas, B. Serafini, and F. Aloisi. 2004. 696 Intracerebral expression of CXCL13 and BAFF is accompanied by formation of 697 lymphoid follicle-like structures in the meninges of mice with relapsing 698 experimental autoimmune encephalomyelitis. J Neuroimmunol 148:11-23. 699
47. Malone, K. E., S. A. Stohlman, C. Ramakrishna, W. Macklin, and C. C. 700 Bergmann. 2008. Induction of class I antigen processing components in 701 oligodendroglia and microglia during viral encephalomyelitis. Glia 56:426-435. 702
48. Mantchev, G. T., C. S. Cortesao, M. Rebrovich, M. Cascalho, and R. J. 703 Bram. 2007. TACI is required for efficient plasma cell differentiation in response 704 to T-independent type 2 antigens. J Immunol 179:2282-2288. 705
49. Marques, C. P., P. Kapil, D. R. Hinton, C. Hindinger, S. L. Nutt, R. M. 706 Ransohoff, T. W. Phares, S. A. Stohlman, and C. C. Bergmann. 2011. 707
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

34
CXCR3-dependent plasma blast migration to the central nervous system during 708 viral encephalomyelitis. J Virol 85:6136-6147. 709
50. Matthews, A. E., E. Lavi, S. R. Weiss, and Y. Paterson. 2002. Neither B cells 710 nor T cells are required for CNS demyelination in mice persistently infected with 711 MHV-A59. J Neurovirol 8:257-264. 712
51. Matthews, A. E., S. R. Weiss, M. J. Shlomchik, L. G. Hannum, J. L. 713 Gombold, and Y. Paterson. 2001. Antibody is required for clearance of 714 infectious murine hepatitis virus A59 from the central nervous system, but not the 715 liver. J Immunol 167:5254-5263. 716
52. Muller, M., S. Carter, M. J. Hofer, and I. L. Campbell. 2010. Review: The 717 chemokine receptor CXCR3 and its ligands CXCL9, CXCL10 and CXCL11 in 718 neuroimmunity--a tale of conflict and conundrum. Neuropathol Appl Neurobiol 719 36:368-387. 720
53. Muller, M., S. L. Carter, M. J. Hofer, P. Manders, D. R. Getts, M. T. Getts, 721 A. Dreykluft, B. Lu, C. Gerard, N. J. King, and I. L. Campbell. 2007. CXCR3 722 signaling reduces the severity of experimental autoimmune encephalomyelitis by 723 controlling the parenchymal distribution of effector and regulatory T cells in the 724 central nervous system. J Immunol 179:2774-2786. 725
54. Narayan, K., D. Dail, L. Li, D. Cadavid, S. Amrute, P. Fitzgerald-Bocarsly, 726 and A. R. Pachner. 2005. The nervous system as ectopic germinal center: 727 CXCL13 and IgG in lyme neuroborreliosis. Ann Neurol 57:813-823. 728
55. O'Connor, B. P., V. S. Raman, L. D. Erickson, W. J. Cook, L. K. Weaver, C. 729 Ahonen, L. L. Lin, G. T. Mantchev, R. J. Bram, and R. J. Noelle. 2004. 730
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

35
BCMA is essential for the survival of long-lived bone marrow plasma cells. J Exp 731 Med 199:91-98. 732
56. Oh, J. W., L. M. Schwiebert, and E. N. Benveniste. 1999. Cytokine regulation 733 of CC and CXC chemokine expression by human astrocytes. J Neurovirol 5:82-734 94. 735
57. Parra, B., C. C. Bergmann, D. R. Hinton, R. Atkinson, and S. A. Stohlman. 736 2001. IFN-gamma secreted by virus-specific CD8+ T cells contribute to CNS 737 viral clearance. Adv Exp Med Biol 494:335-340. 738
58. Phares, T. W., R. B. Kean, T. Mikheeva, and D. C. Hooper. 2006. Regional 739 differences in blood-brain barrier permeability changes and inflammation in the 740 apathogenic clearance of virus from the central nervous system. J Immunol 741 176:7666-7675. 742
59. Phares, T. W., C. P. Marques, S. A. Stohlman, D. R. Hinton, and C. C. 743 Bergmann. 2011. Factors supporting intrathecal humoral responses following 744 viral encephalomyelitis. J Virol 85:2589-2598. 745
60. Phares, T. W., S. A. Stohlman, D. R. Hinton, R. Atkinson, and C. C. 746 Bergmann. 2010. Enhanced antiviral T cell function in the absence of B7-H1 is 747 insufficient to prevent persistence but exacerbates axonal bystander damage 748 during viral encephalomyelitis. J Immunol 185:5607-5618. 749
61. Phares, T. W., S. A. Stohlman, M. Hwang, B. Min, D. R. Hinton, and C. C. 750 Bergmann. 2012. CD4 T cells promote CD8 T cell immunity at the priming and 751 effector site during viral encephalitis. J Virol 86:2416-2427. 752
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

36
62. Puntambekar, S. S., C. C. Bergmann, C. Savarin, C. L. Karp, T. W. Phares, 753 G. I. Parra, D. R. Hinton, and S. A. Stohlman. 2011. Shifting hierarchies of 754 interleukin-10-producing T cell populations in the central nervous system during 755 acute and persistent viral encephalomyelitis. J Virol 85:6702-6713. 756
63. Rainey-Barger, E. K., J. M. Rumble, S. J. Lalor, N. Esen, B. M. Segal, and D. 757 N. Irani. 2011. The lymphoid chemokine, CXCL13, is dispensable for the initial 758 recruitment of B cells to the acutely inflamed central nervous system. Brain, 759 behavior, and immunity 25:922-931. 760
64. Rappert, A., K. Biber, C. Nolte, M. Lipp, A. Schubel, B. Lu, N. P. Gerard, C. 761 Gerard, H. W. Boddeke, and H. Kettenmann. 2002. Secondary lymphoid tissue 762 chemokine (CCL21) activates CXCR3 to trigger a Cl- current and chemotaxis in 763 murine microglia. J Immunol 168:3221-3226. 764
65. Rolink, A. G., J. Tschopp, P. Schneider, and F. Melchers. 2002. BAFF is a 765 survival and maturation factor for mouse B cells. Eur J Immunol 32:2004-2010. 766
66. Rosenblum, J. M., N. Shimoda, A. D. Schenk, H. Zhang, D. D. Kish, K. 767 Keslar, J. M. Farber, and R. L. Fairchild. 2010. CXC chemokine ligand 768 (CXCL) 9 and CXCL10 are antagonistic costimulation molecules during the 769 priming of alloreactive T cell effectors. J Immunol 184:3450-3460. 770
67. Savarin, C., C. C. Bergmann, D. R. Hinton, R. M. Ransohoff, and S. A. 771 Stohlman. 2008. Memory CD4+ T-cell-mediated protection from lethal 772 coronavirus encephalomyelitis. J Virol 82:12432-12440. 773
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

37
68. Savarin, C., S. A. Stohlman, R. Atkinson, R. M. Ransohoff, and C. C. 774 Bergmann. 2010. Monocytes regulate T cell migration through the glia limitans 775 during acute viral encephalitis. J Virol 84:4878-4888. 776
69. Serafini, B., B. Rosicarelli, R. Magliozzi, E. Stigliano, and F. Aloisi. 2004. 777 Detection of ectopic B-cell follicles with germinal centers in the meninges of 778 patients with secondary progressive multiple sclerosis. Brain Pathol 14:164-174. 779
70. Stiles, L. N., M. P. Hosking, R. A. Edwards, R. M. Strieter, and T. E. Lane. 780 2006. Differential roles for CXCR3 in CD4+ and CD8+ T cell trafficking 781 following viral infection of the CNS. Eur J Immunol 36:613-622. 782
71. Stohlman, S. A., D. R. Hinton, B. Parra, R. Atkinson, and C. C. Bergmann. 783 2008. CD4 T cells contribute to virus control and pathology following central 784 nervous system infection with neurotropic mouse hepatitis virus. J Virol 82:2130-785 2139. 786
72. Sussman, M. A., R. A. Shubin, S. Kyuwa, and S. A. Stohlman. 1989. T-cell-787 mediated clearance of mouse hepatitis virus strain JHM from the central nervous 788 system. J Virol 63:3051-3056. 789
73. Tschen, S. I., C. C. Bergmann, C. Ramakrishna, S. Morales, R. Atkinson, 790 and S. A. Stohlman. 2002. Recruitment kinetics and composition of antibody-791 secreting cells within the central nervous system following viral 792 encephalomyelitis. J Immunol 168:2922-2929. 793
74. Tschen, S. I., S. A. Stohlman, C. Ramakrishna, D. R. Hinton, R. D. Atkinson, 794 and C. C. Bergmann. 2006. CNS viral infection diverts homing of antibody-795 secreting cells from lymphoid organs to the CNS. Eur J Immunol 36:603-612. 796
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

38
75. Tyor, W. R., and D. E. Griffin. 1993. Virus specificity and isotype expression of 797 intraparenchymal antibody-secreting cells during Sindbis virus encephalitis in 798 mice. J Neuroimmunol 48:37-44. 799
76. Valbuena, G., W. Bradford, and D. H. Walker. 2003. Expression analysis of 800 the T-cell-targeting chemokines CXCL9 and CXCL10 in mice and humans with 801 endothelial infections caused by rickettsiae of the spotted fever group. Am J 802 Pathol 163:1357-1369. 803
77. van Weering, H. R., H. W. Boddeke, J. Vinet, N. Brouwer, A. H. de Haas, N. 804 van Rooijen, A. R. Thomsen, and K. P. Biber. 2011. CXCL10/CXCR3 805 signaling in glia cells differentially affects NMDA-induced cell death in CA and 806 DG neurons of the mouse hippocampus. Hippocampus 21:220-232. 807
78. Venetz, D., M. Ponzoni, M. Schiraldi, A. J. Ferreri, F. Bertoni, C. Doglioni, 808 and M. Uguccioni. 2010. Perivascular expression of CXCL9 and CXCL12 in 809 primary central nervous system lymphoma: T-cell infiltration and positioning of 810 malignant B cells. Int J Cancer 127:2300-2312. 811
79. Xia, M. Q., B. J. Bacskai, R. B. Knowles, S. X. Qin, and B. T. Hyman. 2000. 812 Expression of the chemokine receptor CXCR3 on neurons and the elevated 813 expression of its ligand IP-10 in reactive astrocytes: in vitro ERK1/2 activation 814 and role in Alzheimer's disease. J Neuroimmunol 108:227-235. 815
80. Xu, W., H. Joo, S. Clayton, M. Dullaers, M. C. Herve, D. Blankenship, M. T. 816 De La Morena, R. Balderas, C. Picard, J. L. Casanova, V. Pascual, S. Oh, 817 and J. Banchereau. 2012. Macrophages induce differentiation of plasma cells 818 through CXCL10/IP-10. J Exp Med 209:1813-1823. 819
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

39
81. Zhang, B., J. Patel, M. Croyle, M. S. Diamond, and R. S. Klein. 2010. TNF-820 alpha-dependent regulation of CXCR3 expression modulates neuronal survival 821 during West Nile virus encephalitis. J Neuroimmunol 224:28-38. 822
823 824
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

Table I. Normal serum neutralizing Ab titers in CXCR3 ligand deficient mice.
Day p.i. WT CXCL9-/- CXCL10-/-
7 1.2 + 0.04a 1.2 + 0.08 1.3 + 0.07
14 2.0 + 0.08 2.0 + 0.09 1.9 + 0.05
21 2.3 + 0.07 2.2 + 0.04 2.7 + 0.07 a Neutralization titers represent the log10 of the highest average serum dilution that inhibited viral cytopathic effect. Data are presented as mean + SEM of 5 individual mice per timepoint.
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from

Table II. CNS levels of CXCL9 and CXCL10.
Day p.i. CXCL9a CXCL10a
Naive BDc BD
5 1.8 ± 0.6b 79.5 ± 7.6
7 9.5 ± 1.6 143.9 ± 11.8
10 2.1 ± 0.6 8.2 ± 1.7 a CXCL9 and CXCL10 (ng/spinal cord) assessed by ELISA. b Data are presented as mean + SEM of 3 individual mice per timepoint. c BD, Below Detection
on April 3, 2019 by guest
http://jvi.asm.org/
Dow
nloaded from