Embed Size (px)
DESCRIPTION
Lecture 1: Nutrition in Parasite: Part 1 Protozoa
Citation preview

BVT 211 – BIOLOGY OF VECTORS
AND PARASITES
Dr. Nik Ahmad Irwan Izzauddin B. Nik Him
Room: G09/403
Email: [email protected]


60% - Final exam
40% - Coursework
- 12% - Prof Wahab
- 14% - Dr. Nik
- 14% - Dr. Hamady

Nutrition and excretion/secretion in parasites
Ecology of parasites
Treatment and control methods in parasitology
Introduction to vectors
Arthropod vector Mosquitoes
Mosquito-borne diseases:
1. Malaria
2. Dengue
3. Filariasis

NUTRITION AND
EXCRETION/SECRETION IN PARASITES
Part 1: Protozoa

HOW A PARASITE GETS NUTRIENT FROM
HOST?
2 system are employed by parasites:
a) Direct nutrients transfer through membranes
b) Through the involvement of various specialized
organs/ structures – permanent/temporary.

Based on the kinds of food/ nutrients taken in – 2 class of
protozoa
1) Phagothropic
– if protozoa takes in large particles of food
2 ) Saprozoic – Obtaining nourishment by absorption of
dissolved organic and inorganic materials
The manner of nutrition in Protozoa varies, depending on
species and where it lives.

PHAGOTHROPIC
Feeding by engulfing a food cell or particle and ingesting
it in a phagocytic vacuole
In Entamoebidae family, there is a structure called
pseudopodia (plural)/ pseudopodium (singular) involved
in feeding.
In ciliates there is a specialized groove or chamber in the
cell where phagocytosis takes place, called the cytostome
or mouth.

SPOROZOA
The Sporozoa are parasitic protozoans that lack
locomotor organs.
They have no cilia, no flagella, no pseudopods.
At some state in their life histories, they are usually
intracellular parasites.
The sporozoan can’t move or obtain food by itself and it
needs to depend on it’s host for transportation and to
obtain food.
Feeding on the cells and body fluids of a main organism
during their life cycle.

PART 1: PROTOZOA
Need to reiterate the following points:
- Protozoa consist of a single cell, although many species
contain more than one nucleus during all or portions of
their life cycles
- Cytoplasm contained with a membrane
- can be divided into ecto and endoplasm
- many organelles/ structures
- mitochondria, Golgi apparatus, lysosomes,
endoplasmic reticulum, ribosome, contractile
vacuoles, flagellum, cilium etc

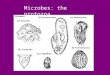
a) Pentatrichomonas hominis, a
harmless commensal of the human
digestive tract.
b) A species of Trypanosoma, 15–30
μm, from the bloodstream of
vertebrates (both a and b have
undulating membranes).
c) Free-living Amoeba sp., 100–150 μm,
showing lobopodia.
d) Actinosphaerium sp., 200 μm (many
species are much smaller), with
actinopodia.
e) Arcella vulgaris, a freshwater shelled
ameba, about 100 μm with lobopodia.
f) Globigerina sp., a marine
foraminiferan up to 800 μm, with
filopodia.
g) Oocyst of Levineia canis, (35–42)
(27–33) μm, a coccidian parasite of
dogs.
h) Zoothamnium sp. colony, individuals
50–60 μm, colony up to 2 mm tall, an
obligate ectocommensal ciliate of
aquatic invertebrates.
i) Euplotes sp., 100–170 μm, a free-
living ciliate with ventral cirri and
prominent oral membranes.
j) Tetrahymena sp., ∼60 μm, a free-
living ciliate showing ciliary rows
(kineties)

ORGANELLES – WITH DIFFERENT FUNCTIONS
Mitochondria – respiration, exception – anaerobic
- Mitochondria, the organelles that bear enzymes of oxidative phosphorylation and the tricarboxylic acid cycle, often have tubular rather than lamellar cristae.
- In addition, some amebas have branched tubular cristae, but in other protozoan groups the cristae may be absent altogether.
- Mitochondria may be present as a single, large body, as in some flagellates, or arranged as elongated, sausage-shaped structures, as occur in pellicular ridges of some ciliates.
Ribosomes, endoplasmic reticulum
– Secretory functions

Contractile vacuoles
– excretion of excess water
Flagellum and cilium
– locomotion and food gathering
Pseudopodia - are temporary extensions of the cell membrane and
- are found in amebas as well as in a variety of cell types in other organisms.
- Pseudopodia function in locomotion and feeding.
All this organelles directly involved with nutrition process

PHYLUM RHIZOPODA
.

Take up dissolved organics directly across the cell membranes,
The most common mechanisms of ingestion are pinocytosisand phagocytosis.
The size of the food vacuoles varies greatly, depending on the primarily on the size of the food materials ingested.
Ingestion can occur anywhere on the surface of the body, there being no distinct cytostome.
Most rhizopodans are carnivores and frequently predaceous.

A food vacuole forms from invagination in the cell surface –sometimes called a food cup – that pinches off and drops inward.
This process called endocytosis, occurs in response to stimulus at the environment.
Vacuole formation in rhizopodans may be induced either mechanical or chemical stimuli
Even nonfood items may be incorporated into food vacuoles, but they are soon ingested.
Not only the size of food item, but also the amount of water taken in during feeding determine the size of the food vacuole.

is the case when a molecule causes the cell membrane to bulge
inward, forming a vesicle
surrounding of food particles
- pushing of food particles into its cytoplasm

If live prey been ingested, they generally die within a few
minutes from the paralytic and proteolytic enzymes
present.
Undigested material that remains within the vacuole wall
reincorporated into cell membrane.
In most rhizopodans this process of cell defecation may
occur anywhere on the body, but in some active forms it
tend to take place at or near the trailing end of the moving
cell.

The streaming of protoplasm inside the
pseudopods moves the amoeba
forward.
Amoeba make contacts with a food
particle.
The pseudopods surround the particle.
After the food is corralled by the amoeba,
an opening in the membrane allows the
food particle to pass into the cell.
Inside the cell, the food is enclosed within
food vacuoles, digested by enzymes,
and assimilated by the amoeba.
The amoeba expels particles that are not
acceptable as food.

B) Uptake of dissolved
nutrients through a
pinocytotic channel in
Amoeba
C) Two ameba ingested
ciliates by phagocytosis.

PHYLUM KINETOPLASTIDA
Trypanosoma brucei, a blood
stream parasite

All kinetoplastids are heterotropic.
Free-living bodonids capture particulate food, primarilly
bacteria, with the aid of anterior flagellum and ingest
through a permanent cytostome.
The cytostome leads to a cytopharynx which is supported
by microtubules
At the base of the cytopharynx, food is enclosed in food
vacuoles by endocytosis


Unfortunately, little is known about feeding mechanisms in trypanosomes, all of which are parasitic.
Some trypanosomes have cytostome-cytopharyngeal complex through which proteins are ingested.
The proteins are taken into food vacuoles by pinocyctosis at the base of cytopharynx.
It has also reported that some trypanosomes can take in proteins by pinocyctosis from the membrane lining the flagellarpocket or by some sort of cell membrane mediated mechanism.

PHYLUM APICOMPLEXA
An apicomplexan sporozoite or merozoite
illustrating the apical complex and other
structures typical of this life-cycle stage.

The alveoli are interrupted at both the anterior and posterior ends, and at tiny invaginations of the cell membranes called microspores, which have been implicated in feeding.
Nutrient ingestion is thought to occur primarily by pinocytosisor phagocytosis at the microspores.
In the haemosporidians, ingestion of the host’s cytoplasm through the microspores has been observed.
Absorption of the nutrients in some gregarines at the point where the parasite attaches to the host’s cell.

B and C: The body of a
gregarine is commonly into
3 three recognizable
regions

PLASMODIUM
Malaria parasites transport carbohydrates, amino acids, purine nucleosides, fatty acids, complex lipids, anions and cationsand the presence of the parasite confers upon the infected red cell pathological alterations in nutrient transport that may favour the development of the parasite.
The asexual stages of the parasite within the red cell lack stored carbohydrate but require considerable quantities of glucose to fuel their active metabolism and division.
Infected erythrocytes use between 10 and 50 times more glucose than uninfected cells and the parasite appears to induce permeability changes in the red cell membrane which facilitate the passage of host glucose and amino acids into the erythrocyte.

The nutritional source of amino acids for intracellular stages of malaria is not fully understood; haemoglobin digestion undoubtedly provides significant amounts, but the infected red cell also shows increased transport of free amino acids in culture.
In P. falciparum, these changes in amino acid transport rates are first seen 15 hours after invasion and the infected erythrocyte loses energy'coupled transport systems in favourof diffusion; whether these amino acids enter the parasite itself by carrier mediated transport or by diffusion remains to be determined and awaits the development of methods permitting culture of the asexual stages of the parasite outside the red cell.

Malaria parasites transport exogenous purine nucleosides but not pyrimidines.
This may be related to their inability to synthesize the purinering de novo.
Parasites liberated from red cells may accumulate certain purines (e.g. adenosine, guanosine and hypoxanthine) and can incorporate radioactivity from labelled adenosine, AMP and ATP.
Several studies have demonstrated that lipids (i.e. free fatty acids, cholesterol and phospholipid) are readily incorporated into malaria parasites resident within the red cell but the transport processes involved are unknown.

PHYLUM DIPLOMONADIDA

Most diplomonads are phagotrophic and feed on
bacteria.
These forms have a cytostome through which
endocytosis of the bacteria occur.
Other genera susch as Giardia lack of cytostomes and
are saprozoic, feeding by pinocytosis on mucous
secretions of the host’s intestinal tissue.

All parabasilids are heterotrophic and lack of cystome.
Fluid is taken up by pinocytosis in depression of cell surface.
Some take in by phagoctosis.
In heypermastigotes, pseudopodia formed in a sensitive region
at the posterior end of the cell engulf wood particles.
Trichomonads also form pseudopodia that engulf bacteria,
cellular debris and leukocytes

(a) Trichomonas foetus; (b) Trichomonas vaginalis

PHYLUM CHILIOPHORA

The ciliates include may different feeding types.
Some are filter feeders, other capture and ingest other protista or small invertebrates, many eat algal filaments or diatoms, some graze on attached bacteria, and a few are saprophytic parasites.
In most ciliates, feeding is restricted to a specialized oral area containing the cytostome or ‘the cell mouth’.
Food vacuoles are formed at the cytostome and then are circulated through the cytoplasm a digestion occurs.

There are a variety of structures associated with, and modifications of the cytostome.
Holozoic ciliates that ingest relatively large food items usually possess a nonciliated tube – called the cytopharnx, which extend from cytosome deep into the cytoplasm.
The wall of the cytopharynx often reinforced with rods of microtubules (nematodesmata).
In Didinium, the cytopharynx is normally everted to form a projection that sticks to prey and then inverts back into the cell, thus pulling the prey into food vacuole.

In this way Didinium can engulf its relatively gigantic prey.
Other ciliates, such as hypostomes, have complex
nematodesmal baskets in which microtubules work
together to withdrawn filament of algae into cytostome,
reminiscent on the way sucks up a piece of spaghetti.
In most of these ciliates, the cilia around the mouth are
relatively simple

Other ciliates, including many of more familiar form (e.g
Stentor) are suspension feeders.
These often lack or have reduced cytopharynxes.
They have elaborate speciall oral cilia for creating water
currents, and filtering structures or scrapping devices.
Their cytostomes often sit in a depression on the cell
surface.

The size of food eaten by such ciliates depends on the nature of feeding current and when present, the size of depression.
The oral ciliates often consists of compound ciliary organeles, call the adoral zone of membranelles or simply the AZM, on the other side of cytostome and a row of closely situated paired cilia which is frequently called the paroral membrane on the other side.
E.G Euplotes, Stentor, and Vorticella.
Many hypotrichs e.g Euplotes that move about the substratum with their oral region oriented ventrally use their specialized oral ciliature to swirl settle materials into suspension and then into buccal cavity for ingestion.


Among the most specialized cialite feeding methods are
those use by suctorians, which lack cilia as adults and
instead have knobbed feeding tentacles.
A few suctorians, have two types of tentacles, one form
for food capture, and another one for digestion.
The swelling at the tips of the tentacles contains
extrusomes called haptocysts, which are discharged upon
contact with potential prey.

Portions of the haptocyst penetrate the victim and hold it
to the tentacle.
Sometime prey are actually paralysed after contact with
haptocycts, presumably by enzymes released during
discharge.
Following attachment to the prey, a temporary tube forms
within the tentacle and the contents of the prey are
sucked into tentacle and incorporated into food vacuoles

Phylum Ciliophora: Feeding in
suctorian ciliate Acinate.
A) Acinate has capitate feeding
tentacles; note the absence of
cilia
B) Contact with prey and firing of
haptocysts into prey
C) Shortening the tentacle and
formation of a temporary
feeding duct within the ring of
microtubules
D) Drawing of contents of prey into
duct and formation of food
vacuole

Several other types are extrusomes are present in the ciliates.
Some predatory ciliates have tubular extrusomes, called toxicysts, in the oral region of the cell.
During feeding, the toxicysts are extruded and release their contents, which apparently include both paralytic and digestive enzymes.
Active prey are first immobilised and then partially digested food is later taken into food vacuoles.

Some ciliates have organelles called mucocysts located
on the beneath the pellicle.
Mucocysts discharge mucus onto the surface of the cell
as a protective coating; they may also play a role in cyst
formation.
Other have trichocysts, which contain nail-shaped
structures that can be discharged through the pellicle.
This structure serve a defensive function.

Extrusomes in ciliates:
A) Toxicysts
B) Mucocysts
C) The pellicle showing of mucocysts of (rised dots) just
below the surface

EXCRETION AND OSMOREGULATION
Most protozoa appear to be ammonotelic; that is, they
excrete most of their nitrogen as ammonia, most of which
readily diffuses directly through the cell membrane into
the surrounding medium.
Other sometimes unidentified waste products are also
produced, at least by intracellular parasites.
After these substances are secreted they are
accumulated within their host cell and, on the death of the
infected cell, have toxic effects on the host.
Carbon dioxide, lactate, pyruvate, and short-chain fatty
acids are also common waste products.

Contractile vacuoles are probably more involved with
osmoregulation than excretion.
Because free living, freshwater protozoa are hypertonic to
their environment, they imbibe water continuously by
osmosis.
Contractile vacuoles effectively pump out the water.
Marine species and most parasites do not form these
vacuoles, probably because they are more isotonic to
their environment.
However, Balantidium species have contractile vacuoles.

CONTRACTILE VACUOLE
Water is collected into the central ring of the vacuole and actively
transported from the cell

RFERENCES;
Foundations of Parasitology
Parasitology – An Integrated Approach