Embed Size (px)
Citation preview

11. Előadás
Nukleinsavak, nukleinsav analitika. Nukleotidok, koenzimek, vitaminok.

Savas karakterű anyagok a sejtmagból
1869-71 DNS a sejtmag fő komponense – “nuclein”
Friedrich Miescher (Svájc, 1844-1895)
1970: FM Insitute for Biomedical Research, Basel
1882 Walther Flemming: „Chromatin” elnevezés
W. Waldeyer: CHROMO (szín) SZOMA
(„coloured body”, festék láthatóvá tétel)
1934 - DNS polimer és molekulatömege kb. 500 000
E. Hammersten, T. Caspersson
- tisztított növényi vírusban (tobacco mosaic virus) is van
W. M. Stanley (USA), Nobel dij, kémia, 1946
1940 első elektronmikroszkópos kép (DNS, pozitívan töltött fehérjék)
1947-50 nukleotid összetétel E. Chargaff (USA)
1953 kettős hélix J. Watson, F. Crick (UK) (N.d. 1962)
1958 DNS polimeráz I enzim (első DNS „készítő” enzim)
1960 RNS polimeráz; mRNS felfedezése
1964 tRNSAla szekvenálása R. Holley (USA)
1966 genetikai kód megfejtése 1968 védett nukleotidok összekapcsolása (A. Todd, Khorana) 1970 DNS hasító enzim (restrikciós) felfedezése
1973 DNS fragmensek beépítése plazmidba E. coli
„Androméda-törzs”
1975 RNS kromoszóma szekvenálása; MS2 fág (3 fehérje)
1977 GENENTECH
Történeti háttér
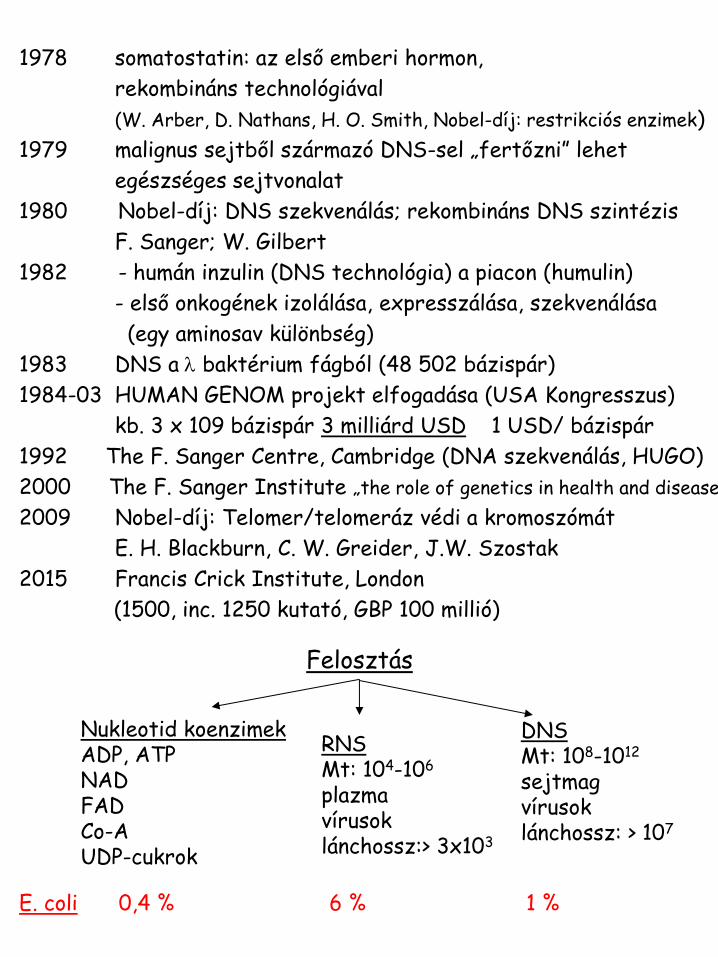
1978 somatostatin: az első emberi hormon,
rekombináns technológiával
(W. Arber, D. Nathans, H. O. Smith, Nobel-díj: restrikciós enzimek)
1979 malignus sejtből származó DNS-sel „fertőzni” lehet
egészséges sejtvonalat
1980 Nobel-díj: DNS szekvenálás; rekombináns DNS szintézis
F. Sanger; W. Gilbert
1982 - humán inzulin (DNS technológia) a piacon (humulin)
- első onkogének izolálása, expresszálása, szekvenálása
(egy aminosav különbség)
1983 DNS a l baktérium fágból (48 502 bázispár)
1984-03 HUMAN GENOM projekt elfogadása (USA Kongresszus)
kb. 3 x 109 bázispár 3 milliárd USD 1 USD/ bázispár
1992 The F. Sanger Centre, Cambridge (DNA szekvenálás, HUGO)
2000 The F. Sanger Institute „the role of genetics in health and disease”
2009 Nobel-díj: Telomer/telomeráz védi a kromoszómát
E. H. Blackburn, C. W. Greider, J.W. Szostak
2015 Francis Crick Institute, London
(1500, inc. 1250 kutató, GBP 100 millió)
Felosztás
Nukleotid koenzimek ADP, ATP NAD FAD Co-A UDP-cukrok
RNS Mt: 104-106 plazma vírusok lánchossz:> 3x103
DNS Mt: 108-1012 sejtmag vírusok lánchossz: > 107
E. coli 0,4 % 6 % 1 %
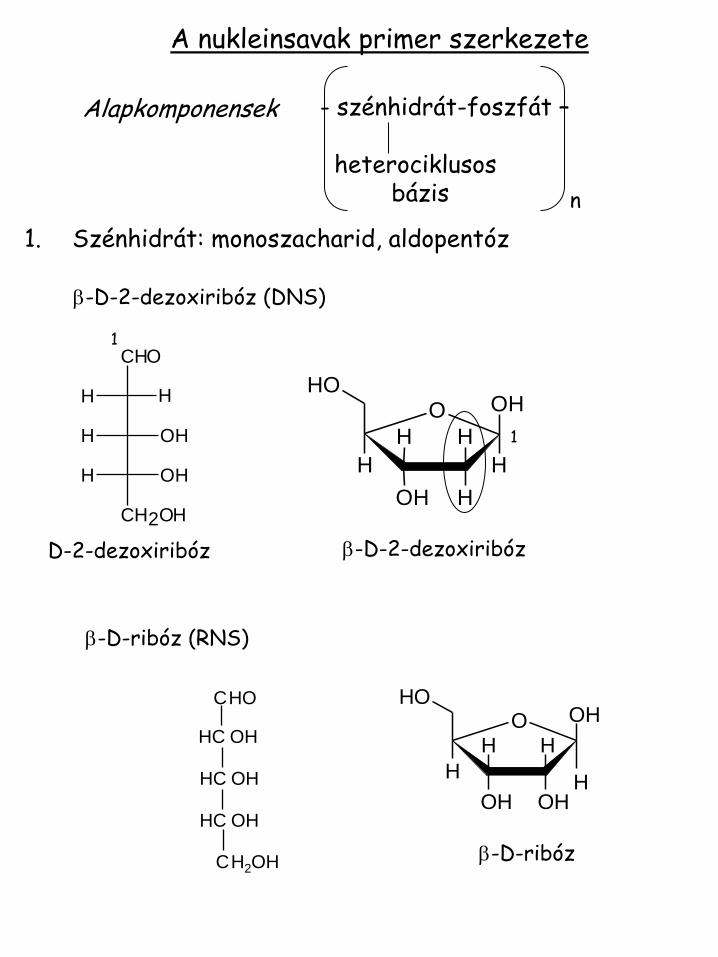
A nukleinsavak primer szerkezete
- szénhidrát-foszfát – heterociklusos bázis n
1. Szénhidrát: monoszacharid, aldopentóz b-D-2-dezoxiribóz (DNS)
CHO
CH2OH
H
H OH
H OH
H
D-2-dezoxiribóz b-D-2-dezoxiribóz
O
H
OH
H
H
H
OH
H
OH
1
1
b-D-ribóz (RNS)
C HO
HC OH
C H2OH
HC OH
HC OH
O O H
OH
H
O H
H
H
O H
H
b-D-ribóz
Alapkomponensek
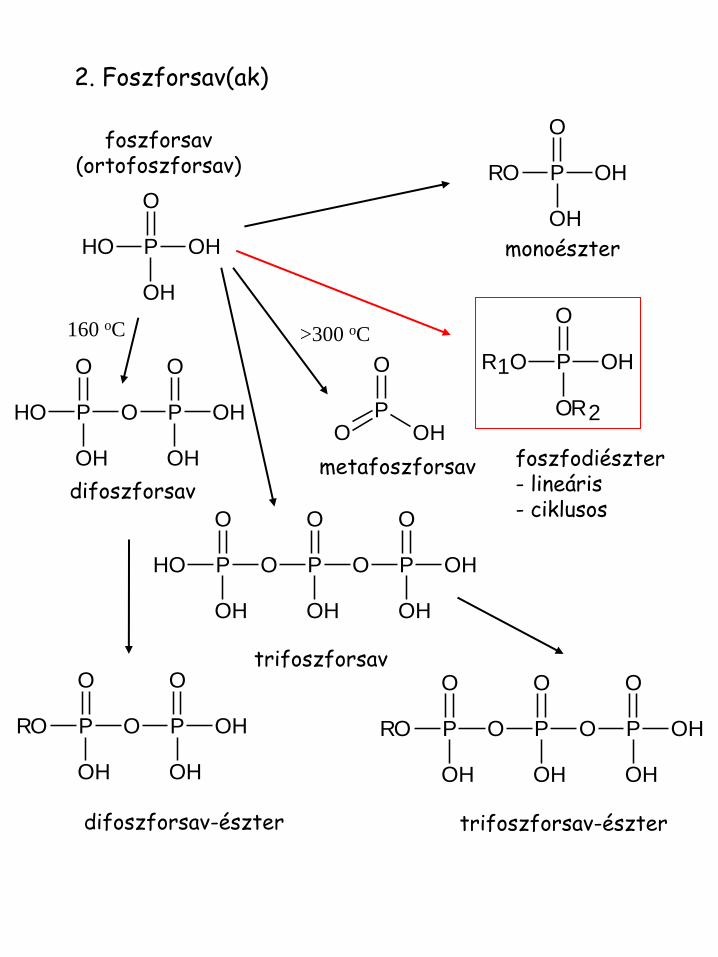
2. Foszforsav(ak)
P
O
OH
OH
OH
P
O
OH
OH
RO
P
O
OH
OR2
R1O
P
O
O
OH
OH
OH
OH
O
P
P
O
O
OH
RO
OH
OH
O
P
P
O
O
OH
OH
OH
O
O
P
OH
OH
O
P
P
O
O
OH
RO
OH
O
O
P
OH
OH
O
P
P
O
OHO
monoészter
foszfodiészter - lineáris - ciklusos
>300 oC
metafoszforsav difoszforsav
160 oC
foszforsav (ortofoszforsav)
trifoszforsav
difoszforsav-észter trifoszforsav-észter

3. Heterociklusos bázis
citozin (C)
timin R = - CH3 (T) R = - H (U) (C)
pirimidin
1 1
3
N
N H
N H 2
O N H
N H
O
O
R
adenin (A)
guanin, (G)
purin (pirimidin(d)imidazol)
1 1
3
5 7
9
a
b
c
e
N
N
N H
N
N H 2
N
N H
N H
N
N H 2
O
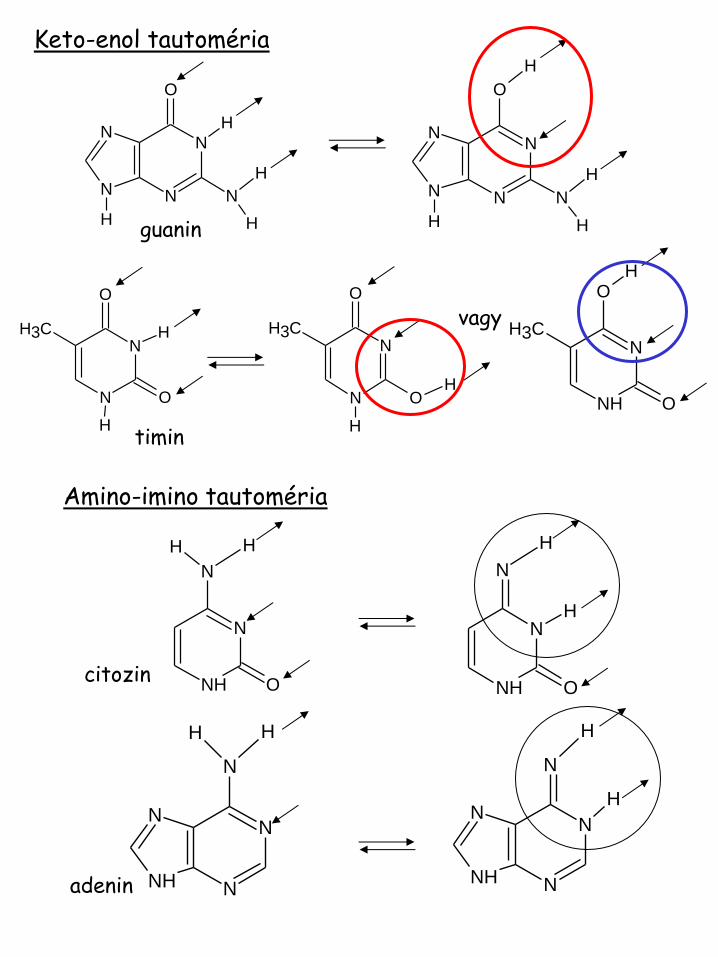
Keto-enol tautoméria
N
N
N
N
N
O
H
H
HH
N
N
N
N
N
O
H
HH
H
N
N
O
O
CH3 H
H
N
N
O
O
CH3
H
H
NH
N
O
O
CH3
H
vagy
guanin
timin
Amino-imino tautoméria
N
NH
N
O
HH
N
N
NH
N
N
HH
N
NH
N
O
H
H
N
N
NH
N
N
H
H
citozin
adenin

Nukleozidok szénhidrát és heterobázis (N – glikozid kötés)
O
H
N
H
OH
H
OH
N
NN
NH2
N
NH
O
O
CH3
O
H
H
OH
H
OH
N
N
NH2
OO
H
HH
OH
H
OH
N
NH
N
N
NH2
O
O
H
HH
OH
H
OH
dezoxicitidin dezoxitimidin
dezoxiadenozin dezoxiguanozin
b-N-glikozid
3
9
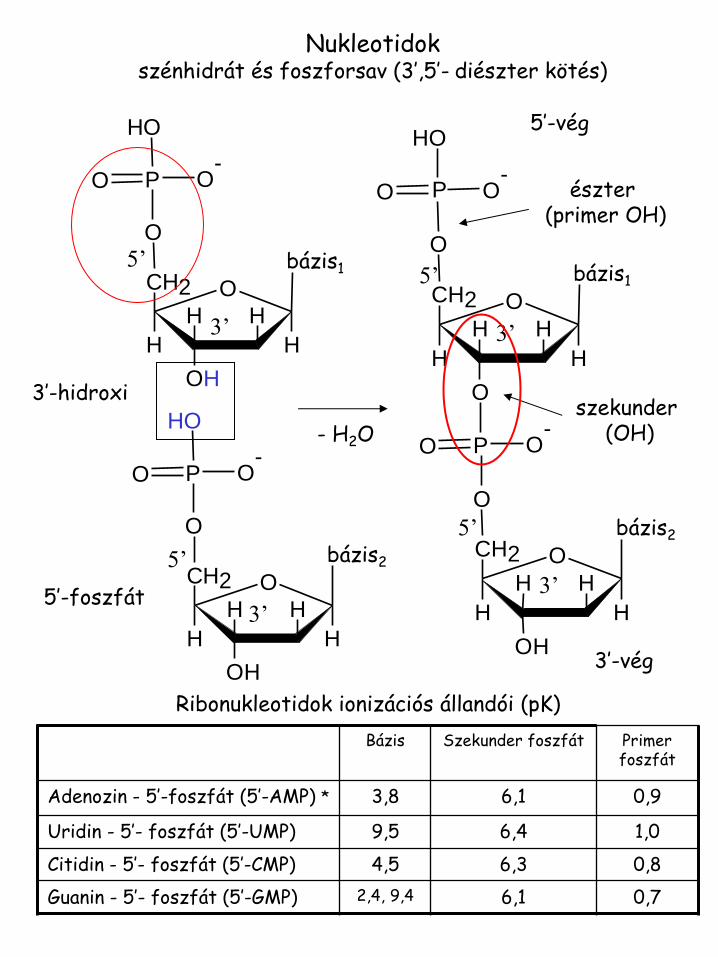
3’-hidroxi
5’-foszfát
5’-vég
3’-vég
észter (primer OH)
szekunder (OH)
Nukleotidok szénhidrát és foszforsav (3’,5’- diészter kötés)
3’
5’
3’
5’
O
H
H H
O H
C H 2
H
O
bázis1
P
O H
O O -
O
H
H H
O H
C H 2
H
O
bázis2
P
O H
O O -
3’
5’
3’
5’
O
H
H H
O
C H 2
H
O
bázis1
P
O H
O O -
O
H
H H
O H
C H 2
H
O
bázis2
P O O - - H2O
Bázis Szekunder foszfát Primer foszfát
Adenozin - 5’-foszfát (5’-AMP) * 3,8 6,1 0,9
Uridin - 5’- foszfát (5’-UMP) 9,5 6,4 1,0
Citidin - 5’- foszfát (5’-CMP) 4,5 6,3 0,8
Guanin - 5’- foszfát (5’-GMP) 2,4, 9,4 6,1 0,7
Ribonukleotidok ionizációs állandói (pK)
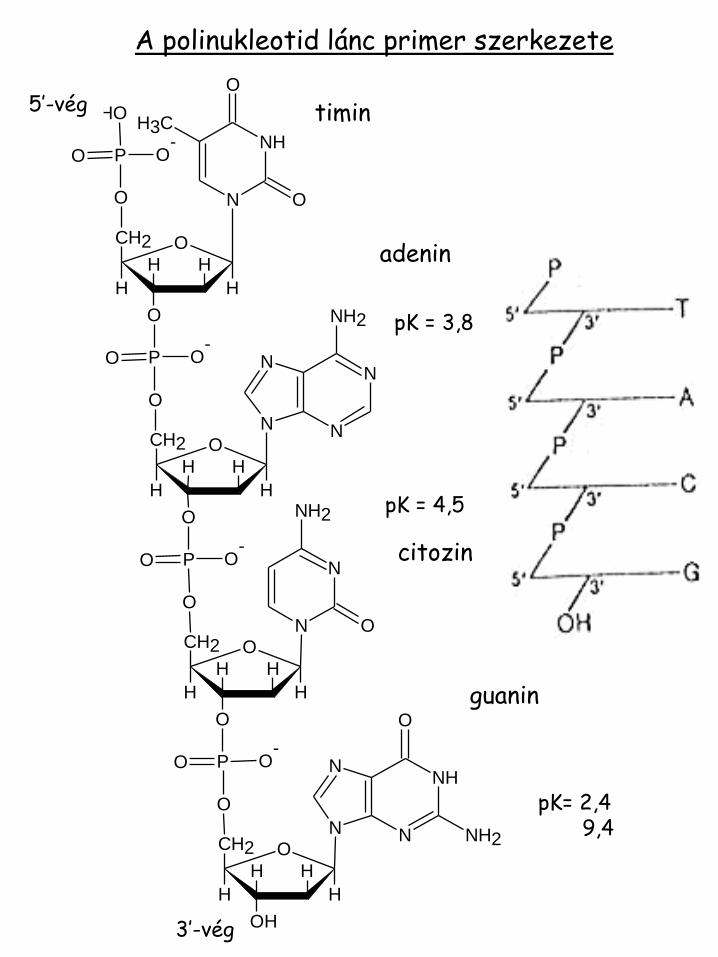
A polinukleotid lánc primer szerkezete
O
H
HH
O
CH2
H
O N
P
OH
O O-
O
H
HH
O
CH2
H
O
N
PO O-
O
H
HH
O
CH2
H
O
N
PO O-
O
H
HH
OH
CH2
H
O
N
PO O-
N
NH2
O
N
NHN
NH2
O
NH
O
O
CH3
N
NN
NH2
timin
adenin
citozin
guanin
pK = 3,8
pK = 4,5
pK= 2,4 9,4
5’-vég
3’-vég

A kettős polinukleotid lánc primer szerkezete
Rövidítések bázis
cukorrész A C
P P OH
foszfodiészter kötés
3’ 3’
5’ 5’
A C
P P
3’ 3’
5’
G
P OH
3’
5’
pApCpG [ACG]

A’
A
csó
(RNA)
Z’
Z
csó
B
C
3M NaCl
Minimális só (nem Li2+)
0.7 M MgCl2 vagy
2.5 M NaCl
72% EtOH alacsony só koncentráció
50% EtOH
Alacsony nedvességű szálakban.
Magas sókoncentráció
Nukleinsav „térszerkezetek”
Francis Crick, 1916-2004 James D. Watson, 1928-
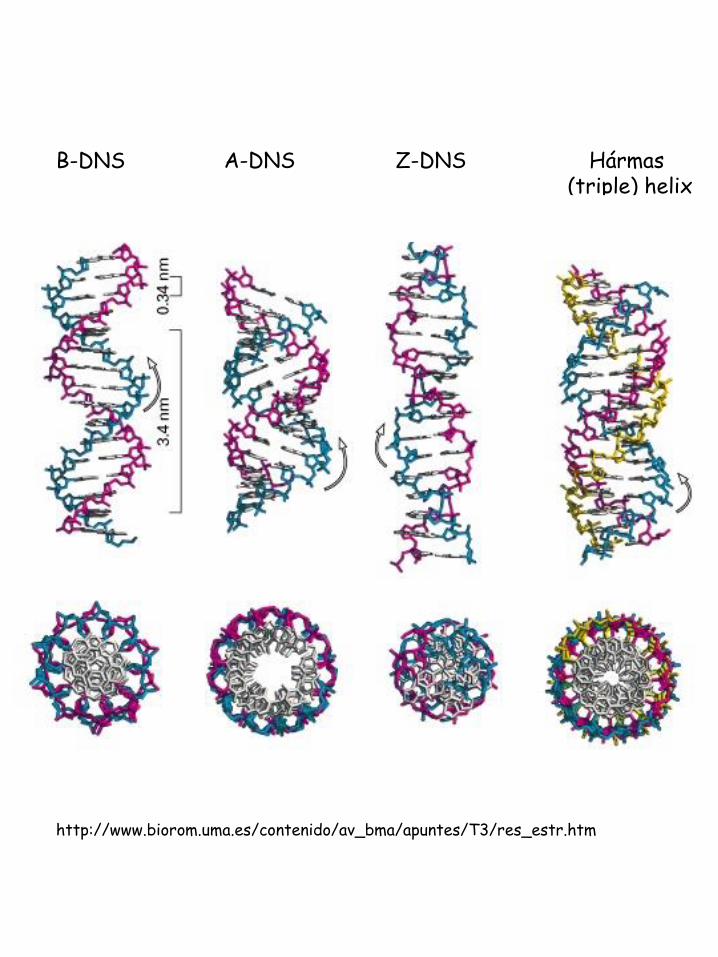
Hármas (triple) helix
http://www.biorom.uma.es/contenido/av_bma/apuntes/T3/res_estr.htm
A-DNS B-DNS Z-DNS
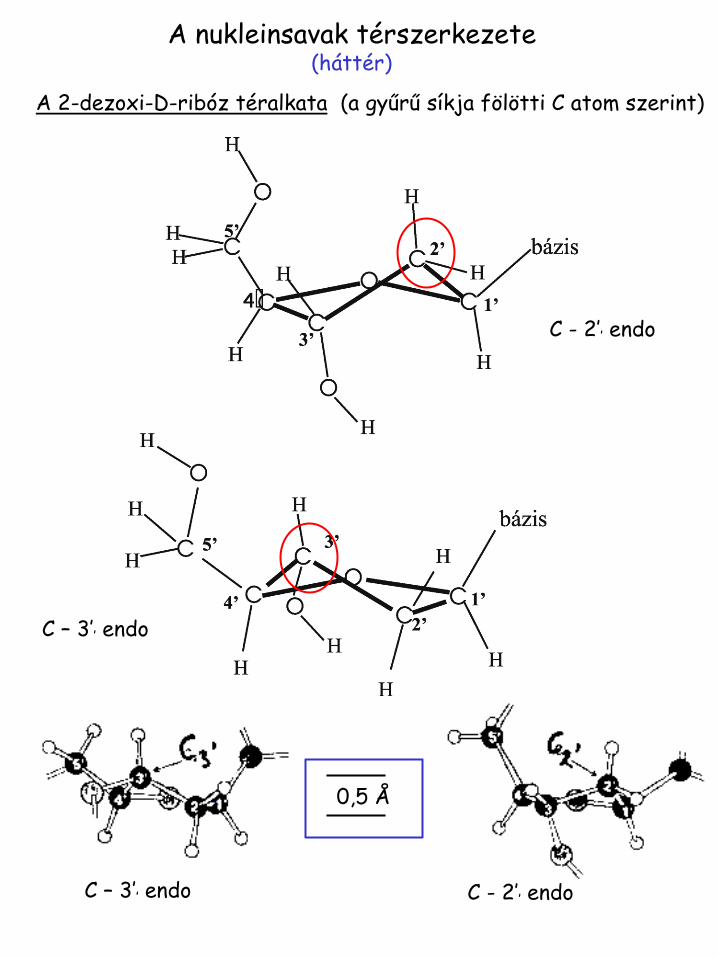
A nukleinsavak térszerkezete (háttér)
A 2-dezoxi-D-ribóz téralkata (a gyűrű síkja fölötti C atom szerint)
0,5 Å
C – 3’’ endo C - 2’’ endo
C – 3’’ endo
C - 2’’ endo

A b-glikozidkötéshez kapcsolódó konformerek
Rotáció a C - N glikozid kötés
körül
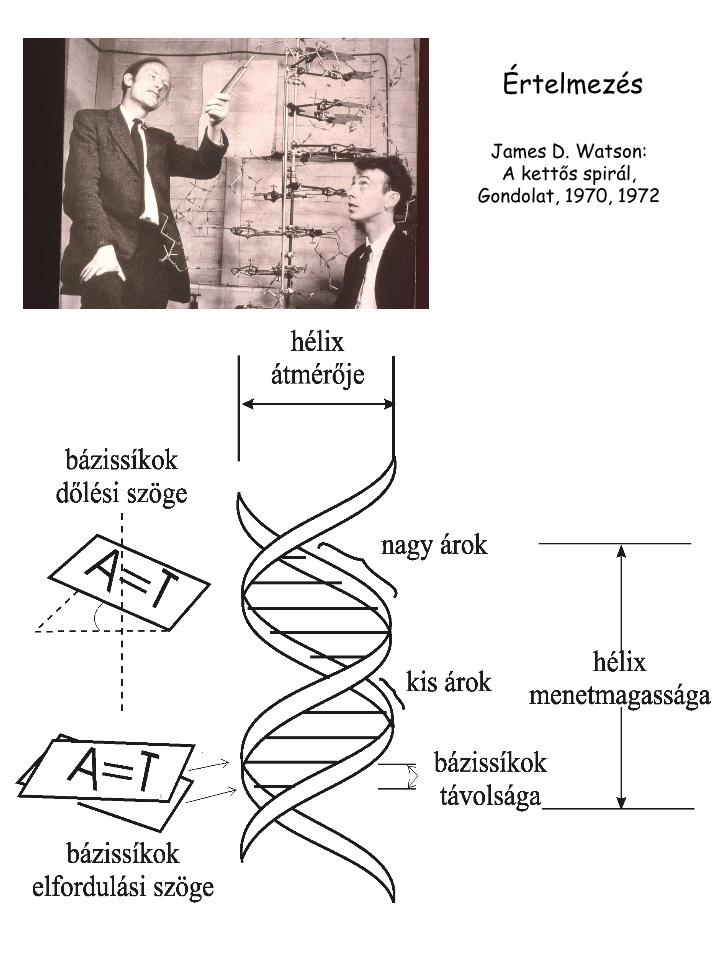
Értelmezés
James D. Watson: A kettős spirál,
Gondolat, 1970, 1972

A B Z
Hélix irány
Jobbmenetű
Jobbmenetű
Balmenetű
Bázispár per kanyar 11 10,4 12 (6 dimers)
Bázispárok távolsága 2.3 Å 3.4 Å 3.8 Å
Menetemelkedés 25,3 Å 35,4 Å 45,6 Å
Bázispár dőlésszög 19 ° 1 ° 9 °
Glikozid konformáció
Dezoxicitidin Anti Anti Anti
Dezoxiguanozin Anti Anti Syn
Szénhidrát konformáció
Dezoxicitidin C-3’-endo C-2’-endo C-2’-endo
Dezoxiguanozin C-3’-endo C-2’-endo C-3’-endo
A, B és Z típusú DNS jellemzői
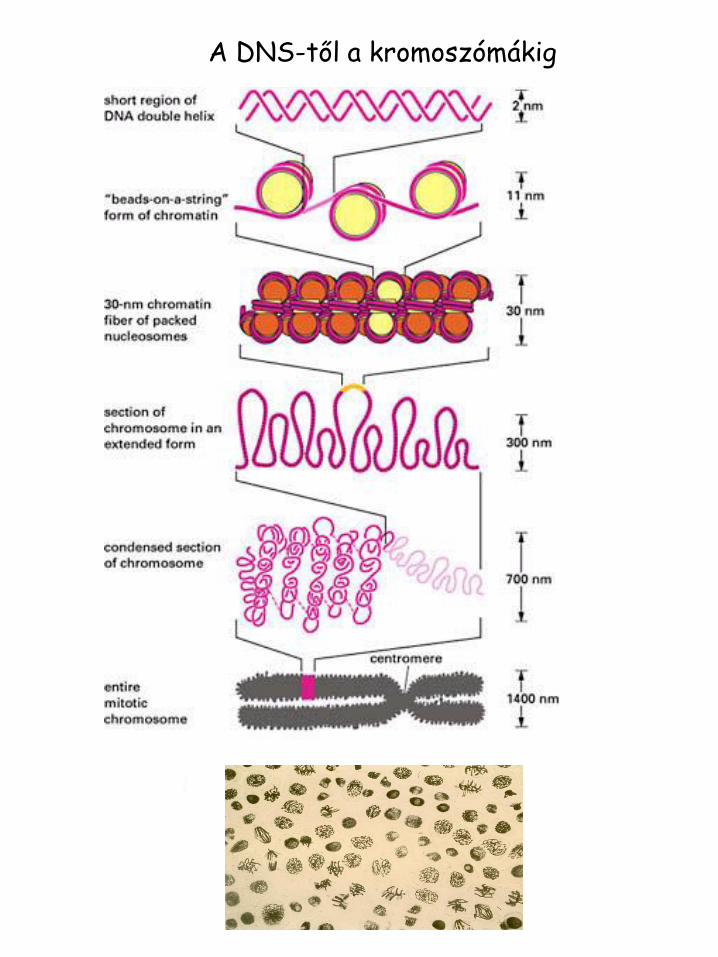
A DNS-től a kromoszómákig
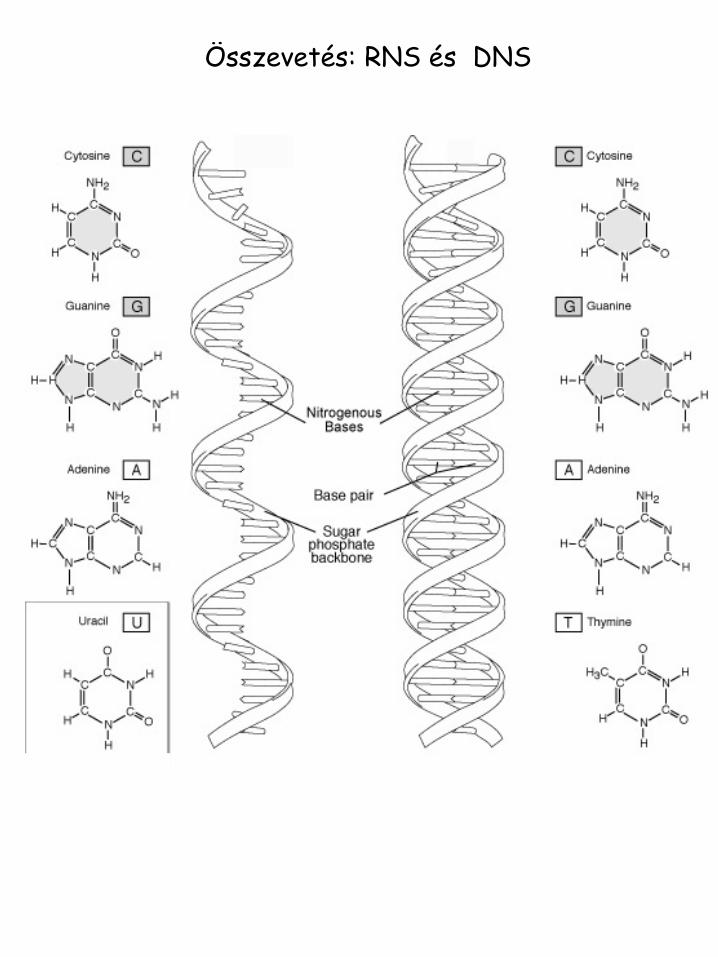
Összevetés: RNS és DNS
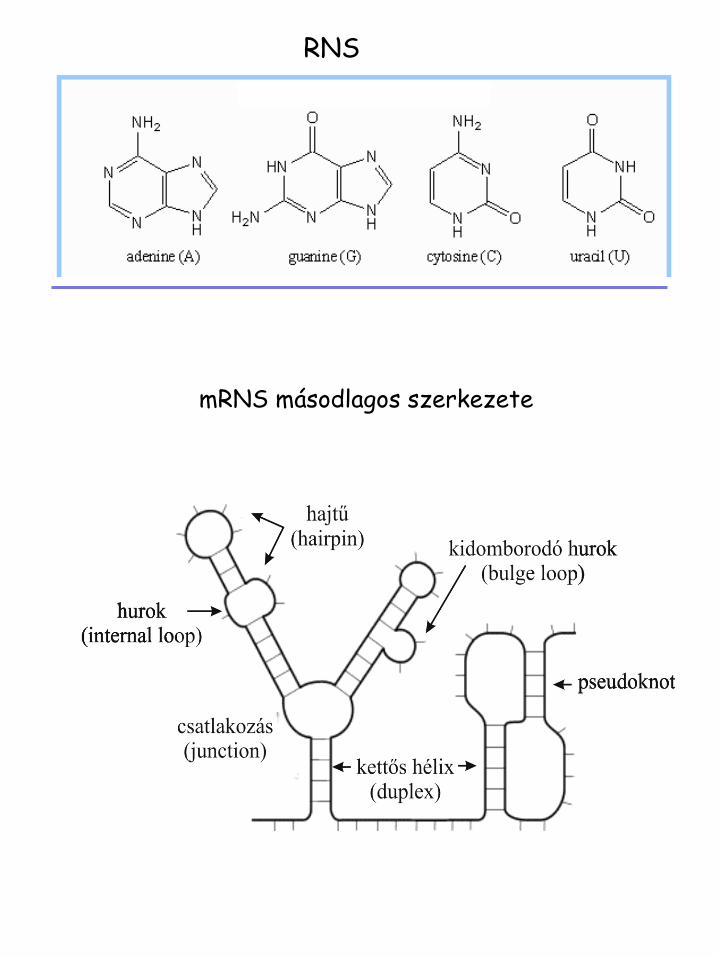
RNS
mRNS másodlagos szerkezete

tRNSAla (Holley , N.d. 1968)
(méret: 75-95 nukleotid)
Aminosav kapcsolódása
tRNS másodlagos szerkezete
Néhány módositott bázisok tRNS-ben
(75-95 nukleotid)
tRNS másodlagos szerkezete
Néhány módositott bázis a tRNS-ben
3 1 7

Aminoacil-tRNS keletkezése
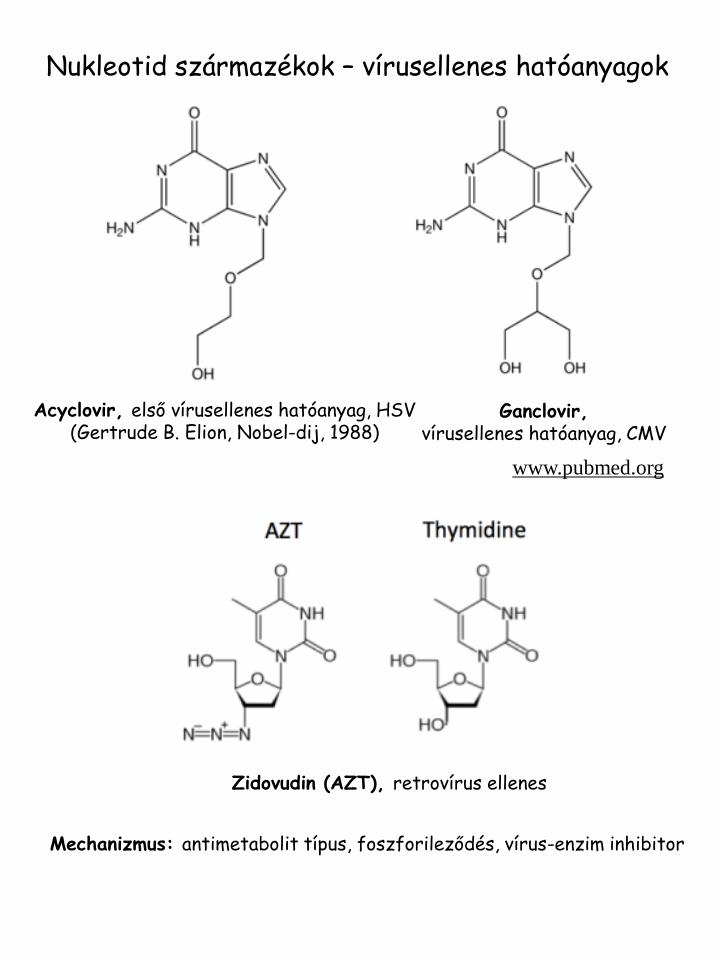
Nukleotid származékok – vírusellenes hatóanyagok
Acyclovir, első vírusellenes hatóanyag, HSV (Gertrude B. Elion, Nobel-dij, 1988)
Ganclovir, vírusellenes hatóanyag, CMV
www.pubmed.org
Zidovudin (AZT), retrovírus ellenes
Mechanizmus: antimetabolit típus, foszforileződés, vírus-enzim inhibitor
N
N
OH
OH
N
N
NH 2
OH N
N
NH 2
OH N
N
NH 2
OH F
HO
OH
HO 2 HC
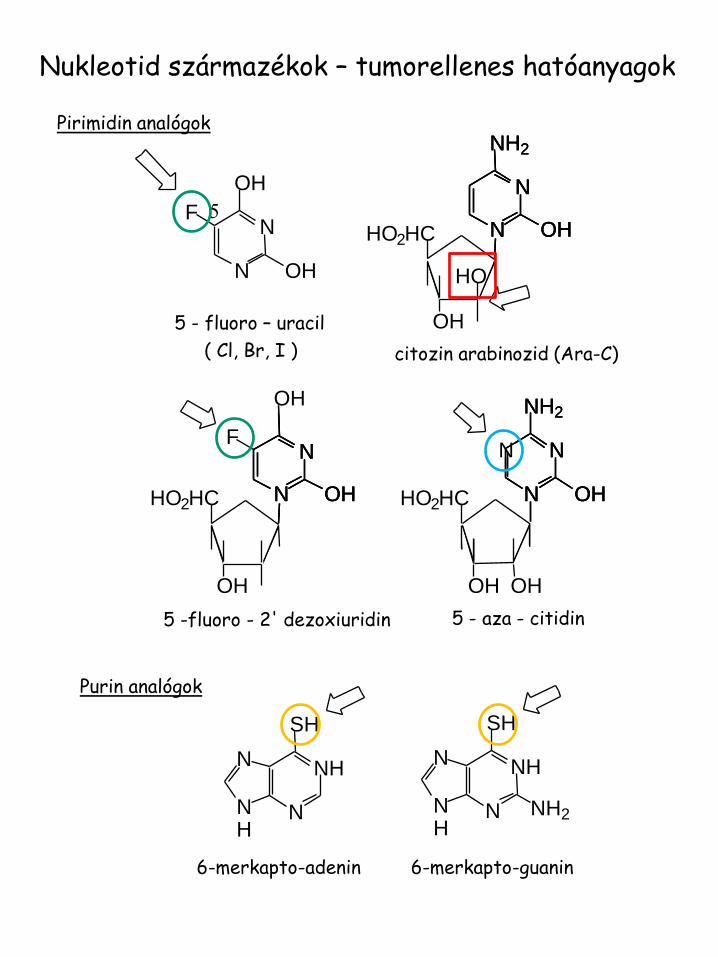
5 - fluoro – uracil
( Cl, Br, I )
5
citozin arabinozid (Ara-C)
Pirimidin analógok
N
N
NH 2
OH N
N
NH 2
OH
N
N
N
NH 2
OH
OH OH
HO 2 HC
5 - aza - citidin
N
N
OH N
N
OH N
N
OH
OH
OH
HO 2 HC
5 -fluoro - 2' dezoxiuridin
F
N
NH N
N H
SH
N
NH N
N H
SH
NH 2
6-merkapto-guanin 6-merkapto-adenin
Purin analógok
Nukleotid származékok – tumorellenes hatóanyagok

1. Elválasztás: gélelektroforézis
Nukleinsav/oligonukleotid analitika
Etidiumbromid (3,8-diamino-5-etil-6-fenil-fenantrédiumbromid) lg=254-366 nm le=590 nm
N+
C2H5
NH2 NH2
Br -
2. Kimutatás
interkaláció

Szekvencia meghatározása
1. Fragmentálás restrikciós enzimekkel (W. Arber, H. Smith, D. Nathans, 1978 Nobel-díj) baktériumokban, kékalgákban
5’ GAATTC 3’
3’ CTTAAGC 5’ EcoRI
törzs
E. coli
5’ GGCC 3’
3’ CCGG 5’ Hae III
Cél: feldarabolás
Néhány II-es típusú restrikciós (endonukleáz) enzim (>2000)
Bam HI G/GATCC Bacillus amyloliquefaciensH
Bst I G/GATCC Bacillus stearothermophilus1503-4R
Eco RI G/AATTC Escherichia coli RY 13
Fok I GGATGN9/
CCTACN13/
Flavobacterium okeanokoites
Hind II GTPy/PuAC Haemophilus influenzae Rd
Hind I A/AGCTT Haemophilus influenzae Rd
Hpa II C/CGG Haemophilus parainfluenzae
Msp I C/CGG Moraxella spezies
Not I GC/GGCCGC Nocardia otitidiscaviarum
Sac I GAGC/TC Streptomyces achromogenes
Sau 3A /GATC Staphyllococcus aureus 3A
Sma I CCC/GGG Serratia marcecescens Sb
Xma I C/CCGGG Xanthomonas malvacearum
5'- 3' irányban haladva a hasítás pontos helye: '/' jelnél ú

2a. Kémiai módosítás/hasítás (A. Maxam, W.Gilbert, 1977)
2b. Didezoxi beépítés - enzimes módszer (F. Sanger et al., 1977)
2. Nuleotid sorrend meghatározás
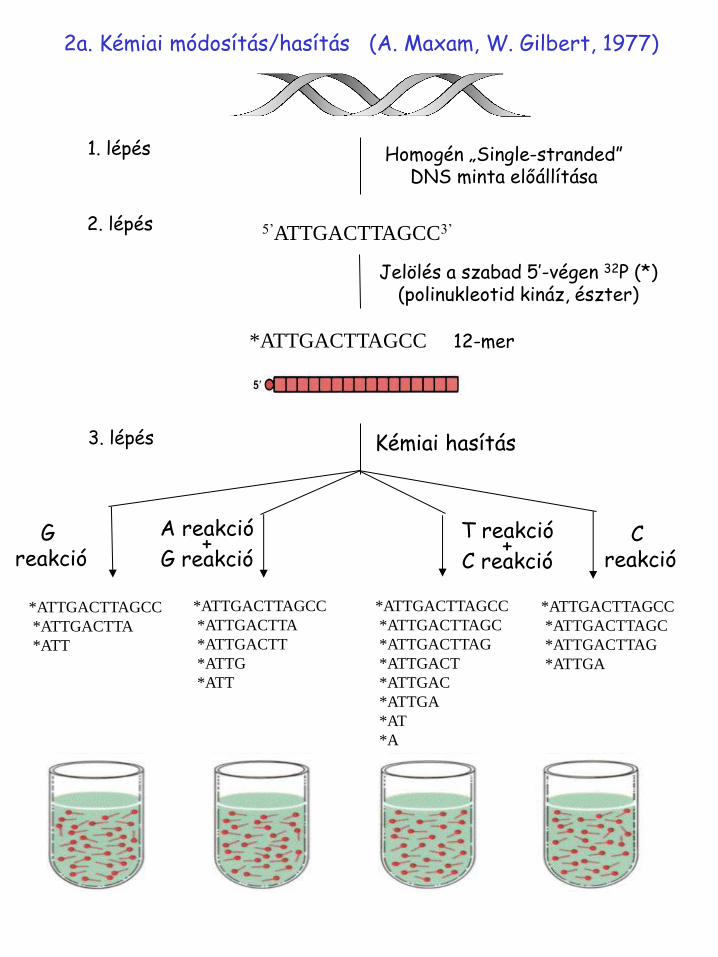
2a. Kémiai módosítás/hasítás (A. Maxam, W. Gilbert, 1977)
Homogén „Single-stranded” DNS minta előállítása
1. lépés
5’ATTGACTTAGCC3’ 2. lépés
*ATTGACTTAGCC 12-mer
Jelölés a szabad 5’-végen 32P (*) (polinukleotid kináz, észter)
3. lépés Kémiai hasítás
G reakció
A reakció +
G reakció T reakció
+ C reakció
C reakció
*ATTGACTTAGCC
*ATTGACTTA
*ATT
*ATTGACTTAGCC
*ATTGACTTA
*ATTGACTT
*ATTG
*ATT
*ATTGACTTAGCC
*ATTGACTTAGC
*ATTGACTTAG
*ATTGACT
*ATTGAC
*ATTGA
*AT
*A
*ATTGACTTAGCC
*ATTGACTTAGC
*ATTGACTTAG
*ATTGA

4. lépés: elektroforézis
http://www.slideshare.net/DurgeshSirohi/different-methods-of-gene-sequencing
Példa: G hasítás után
13
12
11
10
9
8
7
6
5
4
3
2
1
*ATTGAACTTAGCC
*ATTGAACTTA
AACTTAGCC
AACTTA
*ATT
CC
Fragmens hossz
+ +
5. lépés: autoradiográfia 6. lépés: szekvencia leolvasás
elbomlás
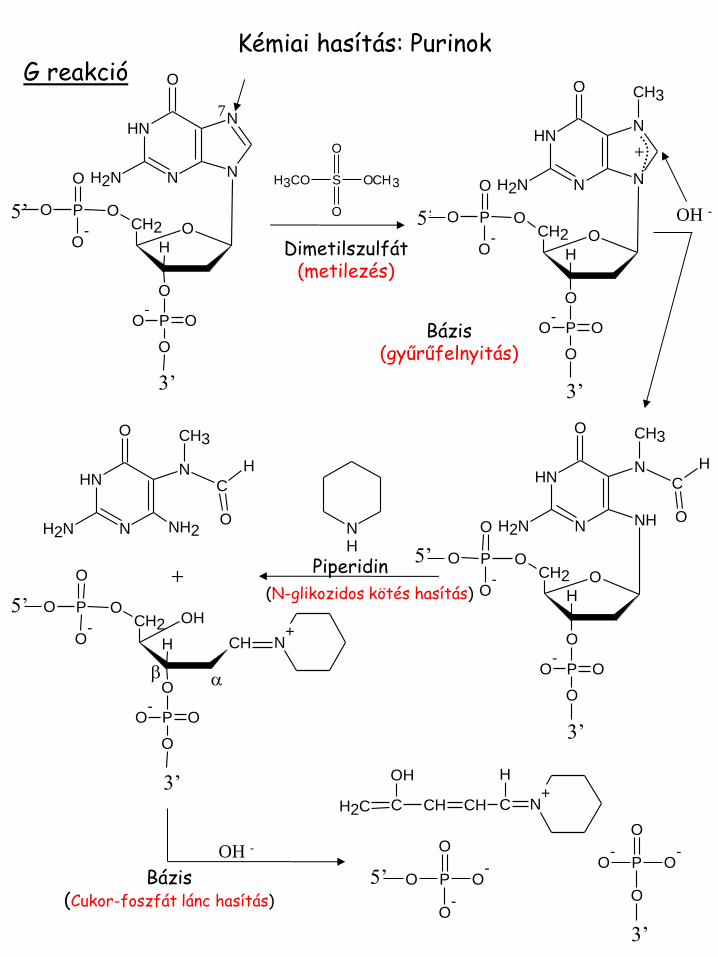
Kémiai hasítás: Purinok G reakció
S
O
O
OCH3H3CO
NH
N
NHN
NH2
O CH3
C
O
H
NH2
+
C N+
CHCHCCH2
OH H
CH
OH
H
O
CH2O
P OO-
O
P
O
O-
OH
N+
5’
O-
O-
O
O
P5’
3’
NHN
NHN
NH2
O
O
H
O
CH2O
P OO-
O
P
O
O-
OH
CH3
C
O
H
5’
3’
O-
O
O-
O
P
3’
5’
NN
NHN
NH2
O
O
H
O
CH2O
P OO-
O
P
O
O-
OH
CH3
3’
+ NN
NHN
NH2
O
O
H
O
CH2O
P OO-
O
P
O
O-
O5’
3’
7
OH -
(N-glikozidos kötés hasítás)
a b
OH -
Dimetilszulfát (metilezés)
Bázis (gyűrűfelnyitás)
Bázis (Cukor-foszfát lánc hasítás)
Piperidin
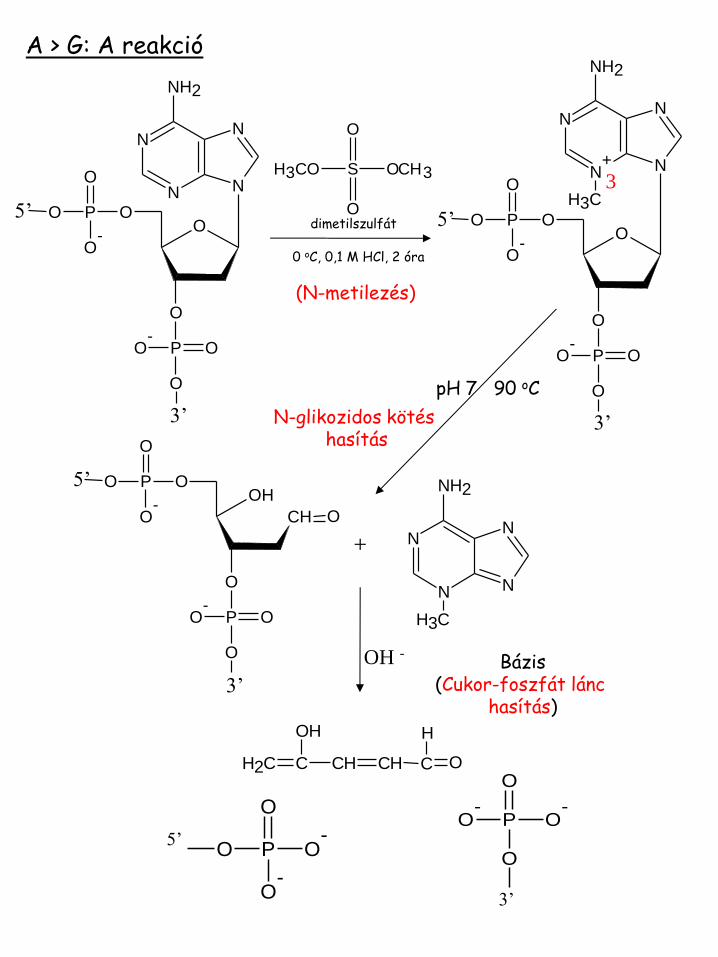
A > G: A reakció
NN
NN
NH2
CH3
CCHCHCCH2 O
OH H
+
O-
O-
O
O
P5’
O-
O
O-
O
P
3’
S
O
O
OCH3H3CO
dimetilszulfát
0 oC, 0,1 M HCl, 2 óra
CH
OHO
O
O
O-
O
P
P O
O
O-
O
5’
3’
N
OO
O
O
O-
O
PN
NN
NH2
P O
O
O-
5’
3’
N
OO
O
O
O-
O
P
N+
NN
NH2
P O
O
O-
CH35’
3’
3
pH 7 90 oC
OH -
(N-metilezés)
N-glikozidos kötés hasítás
Bázis (Cukor-foszfát lánc
hasítás)

Kémiai hasítás: Pirimidinek
N
OO
O
O
O-
O
P
NH
O
O
CH3
N
OO
O
O
O-
O
P
N
NH2
O
NHNH
OCH3
NNH
NH2
O
C
NH2 NH2
OHO
O
O
O-
O
PH
N NH2
OHO
O
O
O-
O
PH
N NH2
H
NH2 NH2hidrazin
+
piperidin
2-dezoxiribozil-hidrazon
4-Me-3-pirazolon 3-amino-pirazol
Urea
timin citozin
Hasítás a 3’ végen
3’
3 3
2 1
3’ 3’
5’ 5’
(N-metilezés)
Bázis (Cukor-foszfát lánc hasítás a 3’- végen)

C reakció
N
N
NH2
O
OO
O
O
O-
O
P
P O
O
O-
N
NHNH2
N
NH2
O
OO
O
O
O-
O
P
P O
O
O-
N
NHNH2
NO
OO
O
O
O-
O
P
P O
O
O-
NH2
NO
CH
OHO
O
O
O-
O
P
P O
O
O-
CH
OHO
O
O
O-
O
P
P O
O
O-
N NH2
+
+
+
CCHCHCCH2 O
OH H
O-
O-
O
O
P5’
O-
O
O-
O
P
3’
OHO
O
O
O-
O
PH
N+
P O
O
O-
3’
5’
OH -
piperidin
NH
NH2 NH2Hidrazin, 1 M NaCl
(gátolja a timin metileződést)
5’ 5’
5’
5’
5’
3’
3’
3’
3’ 3’

C + T: T reakció
CCHCHCCH2 O
OH H
O-
O-
O
O
P5’ O
-
O
O-
O
P
3’
N
NH2
O
O
CH3
OOO
O-
O
P
O
P O
O
O-
NH
NH
3’
5’
N
NH2
O
CH
OHOO
O-
O
P
O
P O
O
O-
3’
5’ OH
O
O
O
O-
O
PH
N+
P O
O
O-
3’
5’
OHO
O
O
O-
O
PH
N NH2
P O
O
O-
3’
5’
N
NH
O
O
CH3
OOO
O-
O
P
O
P O
O
O-
3’
5’ NH2 NH2hidrazin
piperidin
NH
OH -
N-glikozidos kötés hasítás
Bázis (Cukor-foszfát lánc
hasítás)

Maxam-Gilbert módszer - kiértékelés
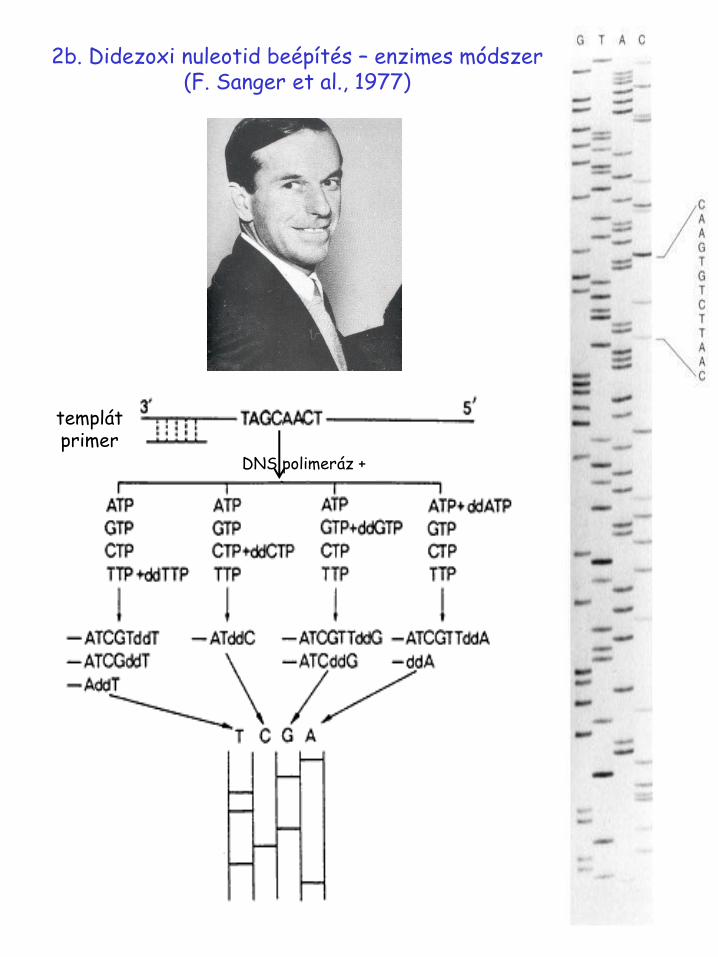
2b. Didezoxi nuleotid beépítés – enzimes módszer (F. Sanger et al., 1977)
templát primer
DNS polimeráz +

1. lépés: Didezoxi nukleotid jelölése
O
H
B
H
H
H
H
H
OPPO
H
B
H
H
H
H
H
OPPP32
b) Fluorofór (fluoreszcens jelölés, L. E. Hood, 1986)
32P
a) Radioaktív izotóp (32P)
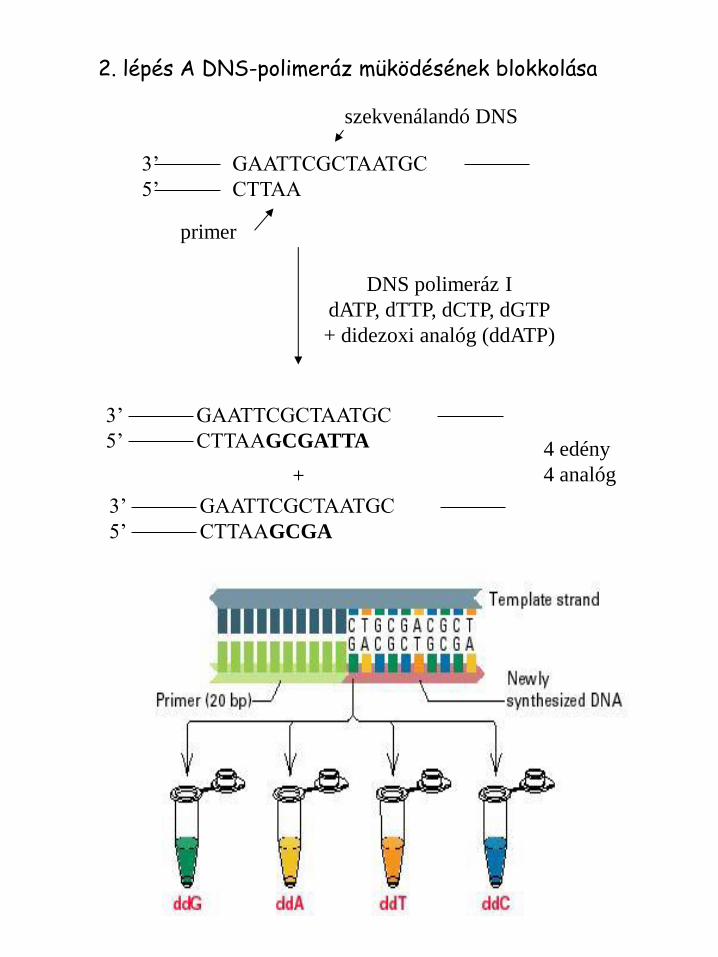
2. lépés A DNS-polimeráz müködésének blokkolása
3’ GAATTCGCTAATGC
5’ CTTAA
szekvenálandó DNS
primer
DNS polimeráz I
dATP, dTTP, dCTP, dGTP
+ didezoxi analóg (ddATP)
3’ GAATTCGCTAATGC
5’ CTTAAGCGATTA
3’ GAATTCGCTAATGC
5’ CTTAAGCGA
+
4 edény
4 analóg

5’ 3’
3’ 5’
DNS polimeráz
ddGTP ddATP ddTTP ddCTP
(32P)
Áttekintés
„primer”
DENATURÁLÁS
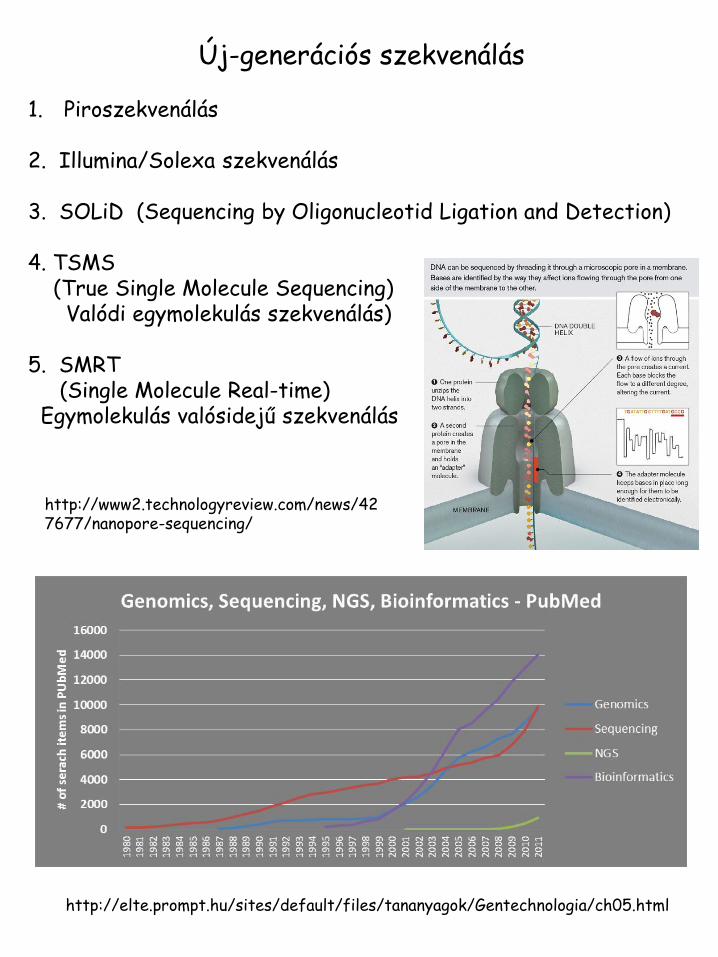
Új-generációs szekvenálás 1. Piroszekvenálás
2. Illumina/Solexa szekvenálás 3. SOLiD (Sequencing by Oligonucleotid Ligation and Detection) 4. TSMS (True Single Molecule Sequencing) Valódi egymolekulás szekvenálás) 5. SMRT (Single Molecule Real-time) Egymolekulás valósidejű szekvenálás
http://elte.prompt.hu/sites/default/files/tananyagok/Gentechnologia/ch05.html
http://www2.technologyreview.com/news/427677/nanopore-sequencing/

Oligonukleotidok szintézise
Elv:
NO
OH
O
PO OH
OH
NO
OH
O
PO OH
OH
( )
( )
1. Kondenzáció
5’-OH + H (3’)
2. Vízmentes közeg
3. Védőcsoportok
Módszerek
1. Karbodiimid módszer, oldatban (1955)
Tood (Nobel díj 1957), Khorana (Nobel díj 1975)
Kapcsoló szer: DCC/Piridin
8-12 tagszám
2. Foszfit módszer, szilárd hordozón
Beaucage, Caruthers; (1981)
(Merrifield Nobel dij, 1984)
5’ 3’

Kapcsoló szer: DCC/Piridin
Védőcsoportok: acetil, tritil
1. lépés: Védett komponensek kapcsolása
N, N – diciklohexil-karbodiimid (DCC)
Tritil (Tr)
O-acetil (Ac)
N-acetil (Ac)
N
N
N
N
NH
OO
OH
C
C
O
CH3
N
N
NH
O
OO
O
OH
OH
O
P
C
CH3
O
C
O
CH3
6
4
1. Szintézis oldatban
C
N
N
Sir Alexander R. Todd
(1907-1997)

2. lépés: A védőcsoportok eltávolítása
AO
OH
O C
OO
OH
O
P
OH
AO
Tr
O C
OO
OH
O
P
OH
OH T
OO
OH
O
P
OAc
AO
Tr
O C
OO
OH
O
P
OAc
Ac
Ac
1. Lúg
2. Sav
(80% AcOH)
Lúg
+
3. lépés: Láncnövekedés
CO
CMeO
védelem (Bz) védelem (MMTr)
Védőcsoportok
MMTr = monometoxi-tritil

2.1 Szilárd hordozó: szilika, üveggyöngy,műanyag, cellulóz, papir
szilika Si OH Si O (CH2)3 NH2szilika
1. borostyánkősavanhidrid
2. védett nukleotid
szilika Si O (CH2)3 NH C (CH2)2 C
O O
O
AO
DMTrBz2
savérzékeny
bázis érzékeny
Si O (CH2)3 NH C (CH2)2 C
O O
O
AO
OH
szilika
Bz H +
+ DMTr +
l= 498 nm
2. Szintézis szilárd fázison
(karbokation)
Foszfit módszer Beaucage, Caruthers (1981)
3-aminopropanol
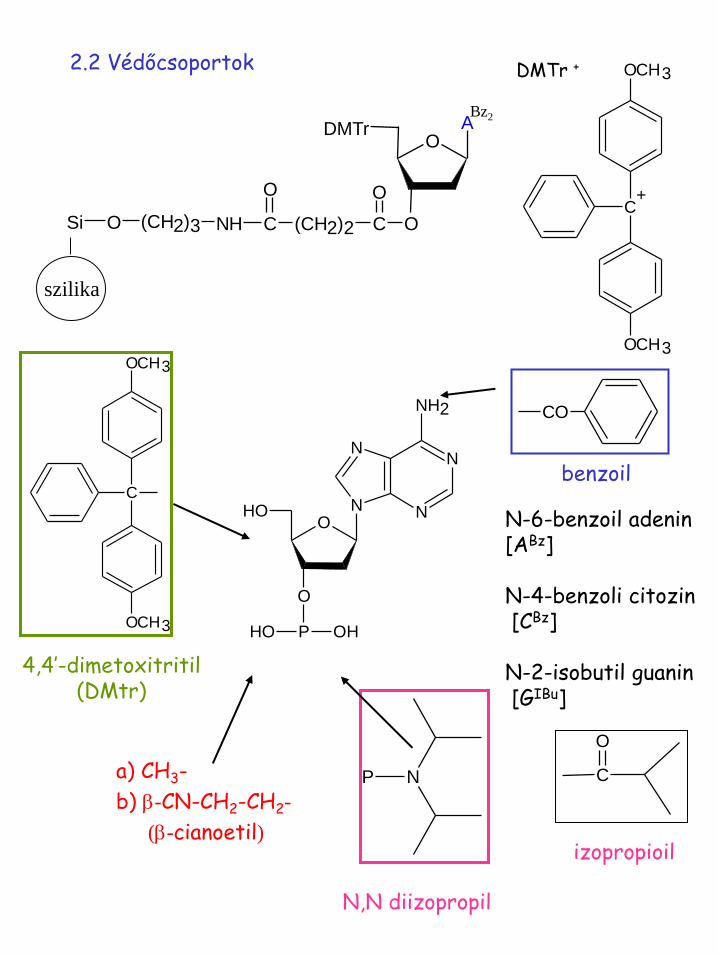
2.2 Védőcsoportok
benzoil
CO
N-6-benzoil adenin [ABz] N-4-benzoli citozin [CBz] N-2-isobutil guanin [GIBu]
C
O
C
OCH3
OCH3
4,4’-dimetoxitritil (DMtr)
a) CH3-
b) b-CN-CH2-CH2-
(b-cianoetil)
NP
N,N diizopropil
N
N
N
N
NH2
OOH
O
OH OHP
C+
OCH3
OCH3
DMTr +
szilika
Bz2
Si O (CH 2 ) 3 N H C (CH 2 ) 2 C
O O
O
A O
DMTr
izopropioil
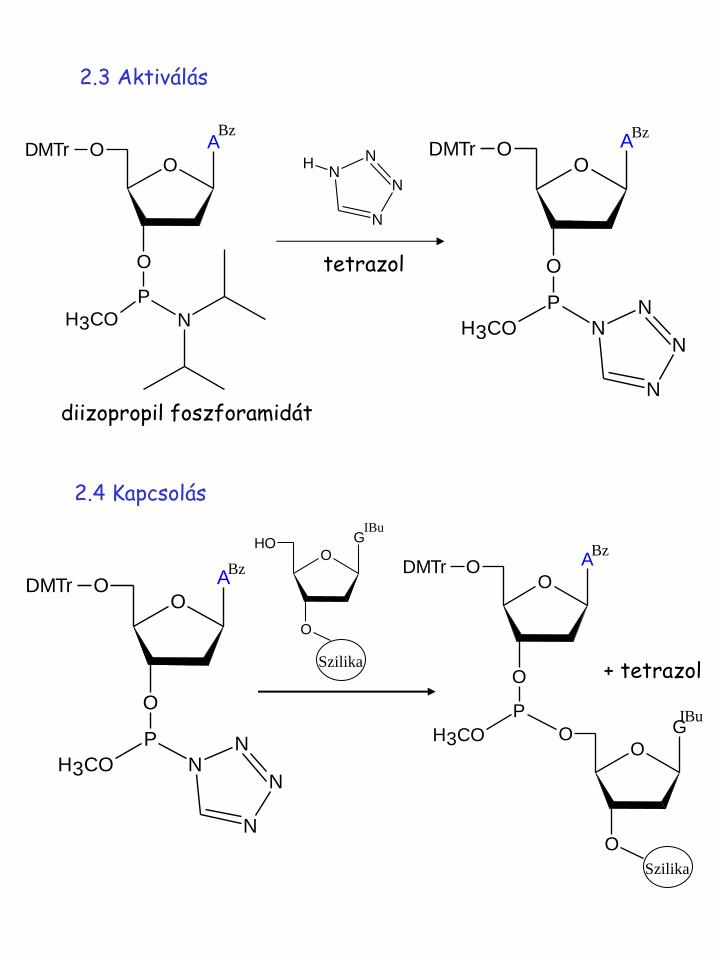
2.3 Aktiválás
A
OO
O
DMTr
P
NH3CO
N
N
NN
A
OO
O
DMTr
P
H3CO
N
N
NN
H
tetrazol
Bz Bz
diizopropil foszforamidát
2.4 Kapcsolás
N
N
NN
A
OO
O
DMTr
P
H3CO
Bz
G
OOH
O
Szilika
IBu
A
OO
O
DMTr
P
H3CO O G
O
O
Szilika
IBu
Bz
+ tetrazol
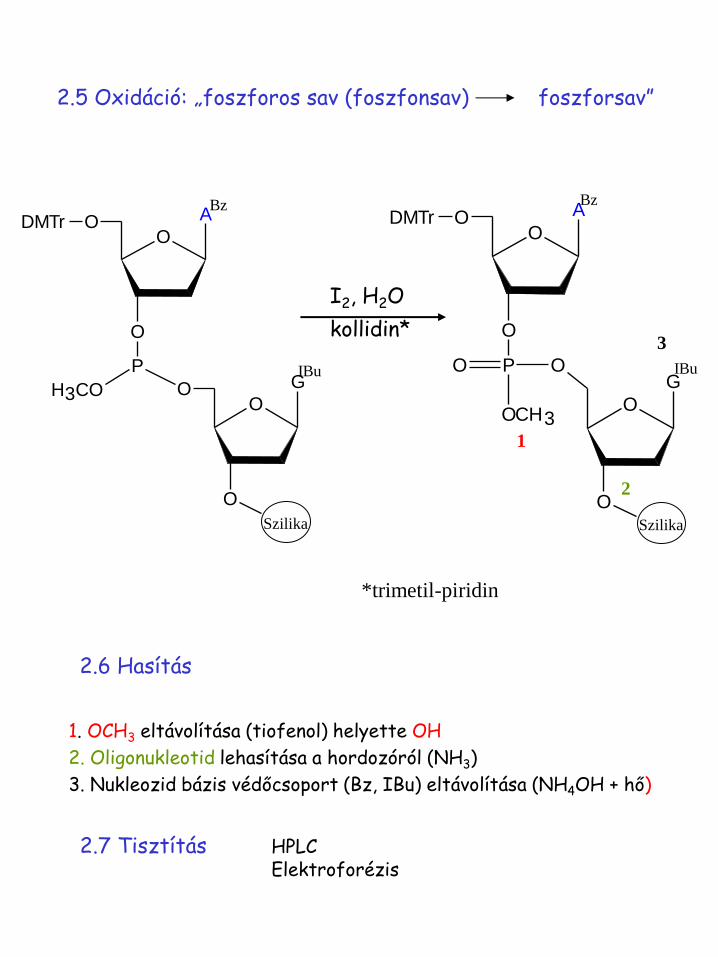
2.5 Oxidáció: „foszforos sav (foszfonsav) foszforsav”
A
OO
O
DMTr
P
H3CO O G
O
O
Szilika
IBu
Bz A
OO
O
DMTr
P
OCH3
OG
O
O
O
Szilika
IBu
Bz
3
2
1
I2, H2O
kollidin*
2.6 Hasítás
1. OCH3 eltávolítása (tiofenol) helyette OH
2. Oligonukleotid lehasítása a hordozóról (NH3)
3. Nukleozid bázis védőcsoport (Bz, IBu) eltávolítása (NH4OH + hő)
2.7 Tisztítás HPLC Elektroforézis
*trimetil-piridin
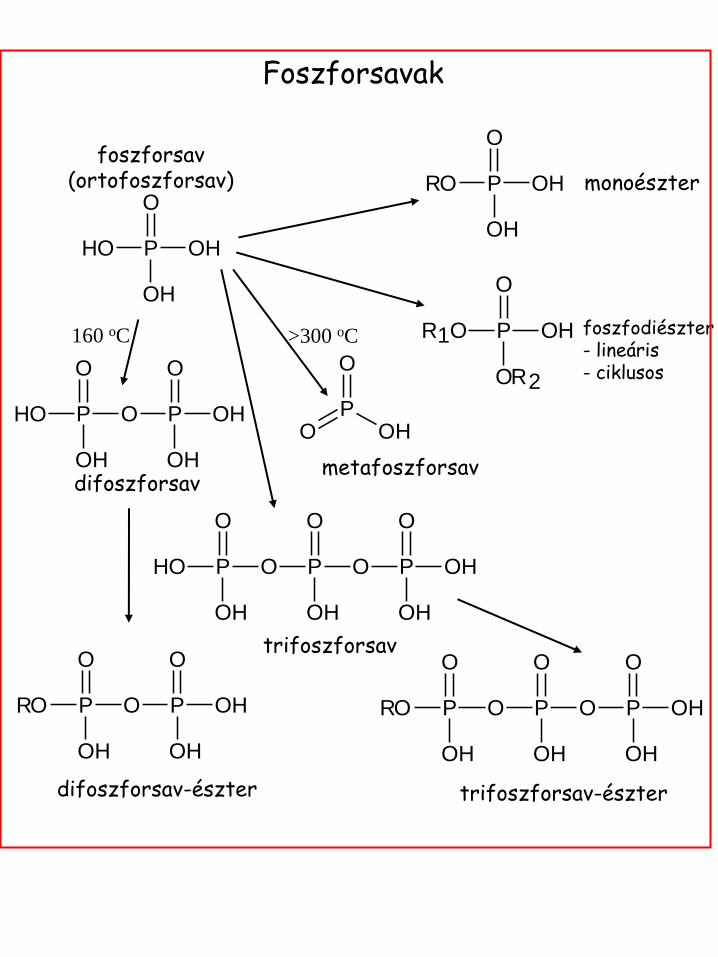
Foszforsavak
P
O
OH
OH
OH
P
O
OH
OH
RO
P
O
OH
OR2
R1O
P
O
O
OH
OH
OH
OH
O
P
P
O
O
OH
RO
OH
OH
O
P
P
O
O
OH
OH
OH
O
O
P
OH
OH
O
P
P
O
O
OH
RO
OH
O
O
P
OH
OH
O
P
P
O
OHO
monoészter
foszfodiészter - lineáris - ciklusos
>300 oC
metafoszforsav difoszforsav
160 oC
foszforsav (ortofoszforsav)
trifoszforsav
difoszforsav-észter trifoszforsav-észter

https://www.atdbio.com/content/17/Solid-phase-oligonucleotide-synthesis
Oligonukleotid szintézis ciklus (“phosphoramidite”)
1. lépés Aktiválás, kapcsolás
2. lépés „Sapkázás” (capping):
reakció ecetsavanhidriddel, a szabad OH acetilezése
3. lépés Oxidáció
4. lépés Tritil-csoport
hasítása
Következő ciklus
Védőcsoportok hasítása
Tritil-csoport hasítása
Kezdés
Oligonukleotid hasitása
a hordozóról

DNS/oligonukleotid chip (csip)

Expression levels of 50 genes most highly correlated with the acute lymphoblastic leukemia (ALL) and
acute myeloid leukemia (AML)
Expressed in ALL
Expressed in AML
Expression levels greater than the mean: red, below the mean: blue.
TR Golub et al Science 286: 531 (1999)

Koenzimek
Enzim = apoenzim (fehérje) + koenzim (nem-fehérje)
Szubsztrátspecifitás Reakciótípus
Nukleotid Fémion (példa) Egyéb
Fe (citokróm oxidáz)
NAD Cu (aszkorbinsav oxidáz) B1
FAD Zn (alkohol dehidrogenáz) B6
ATP Mn (hisztidin ammónia-liáz) B12
Koenzim A Co (glutamát mutáz) folsav
Ni (ureáz) pantoténsav
Mo (xantin oxidáz) biotin
V (nitrát reduktáz)
Se (glutation peroxidáz)
Felosztás
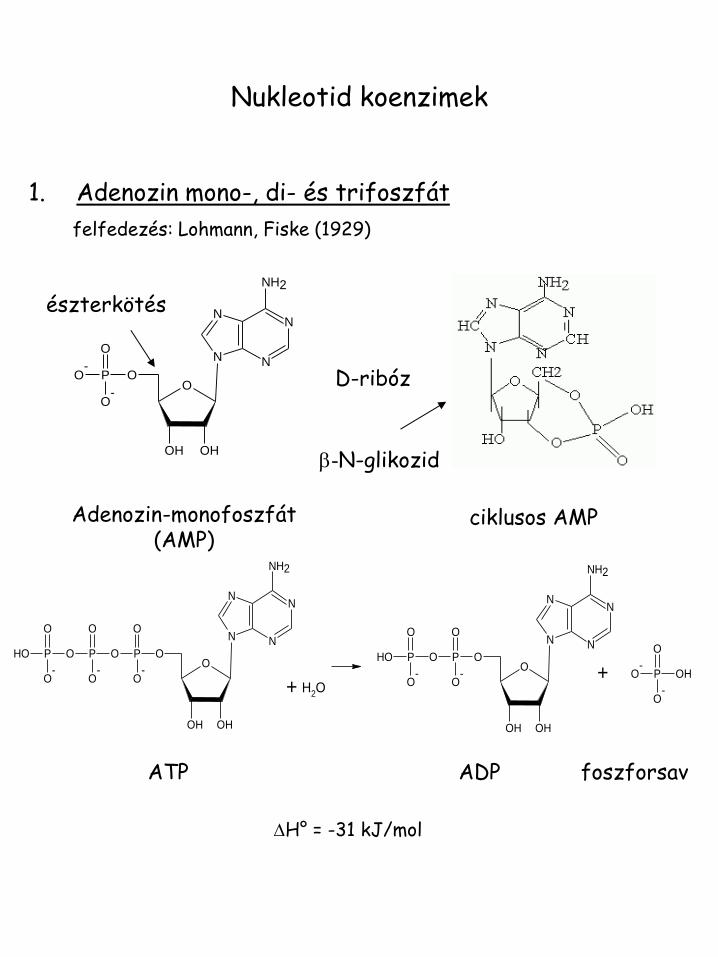
Nukleotid koenzimek
1. Adenozin mono-, di- és trifoszfát
felfedezés: Lohmann, Fiske (1929)
OO
OHOH
P
O
O-
O- N
N
N
N
NH2
OO
OHOH
P
O
O-
P
O
O
O-
OH
O-
O
O
PN
N
N
N
NH2
OO
OHOH
P
O
O-
P
O
O
O-
OHN
N
N
N
NH2
P
O
OH
O-
O-
++ H
2O
Adenozin-monofoszfát (AMP)
ATP ADP foszforsav
DH° = -31 kJ/mol
b-N-glikozid
D-ribóz
észterkötés
ciklusos AMP
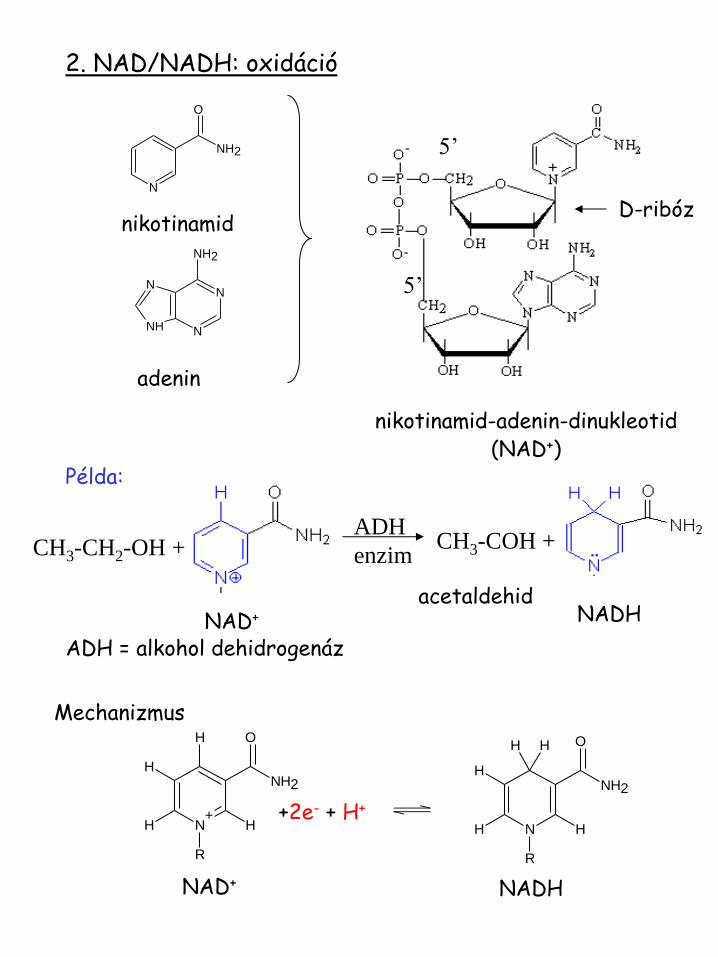
2. NAD/NADH: oxidáció
N
O
NH2
N
N
NH
N
NH2
nikotinamid
adenin
nikotinamid-adenin-dinukleotid (NAD+)
5’
5’
D-ribóz
Példa:
CH3-CH2-OH + ADH
enzim CH3-COH +
NAD+ NADH
ADH = alkohol dehidrogenáz
acetaldehid
Mechanizmus
N+
O
NH2
H
H
H H
R
N
O
NH2
H
H
H H
R
H
+2e- + H+
NAD+ NADH
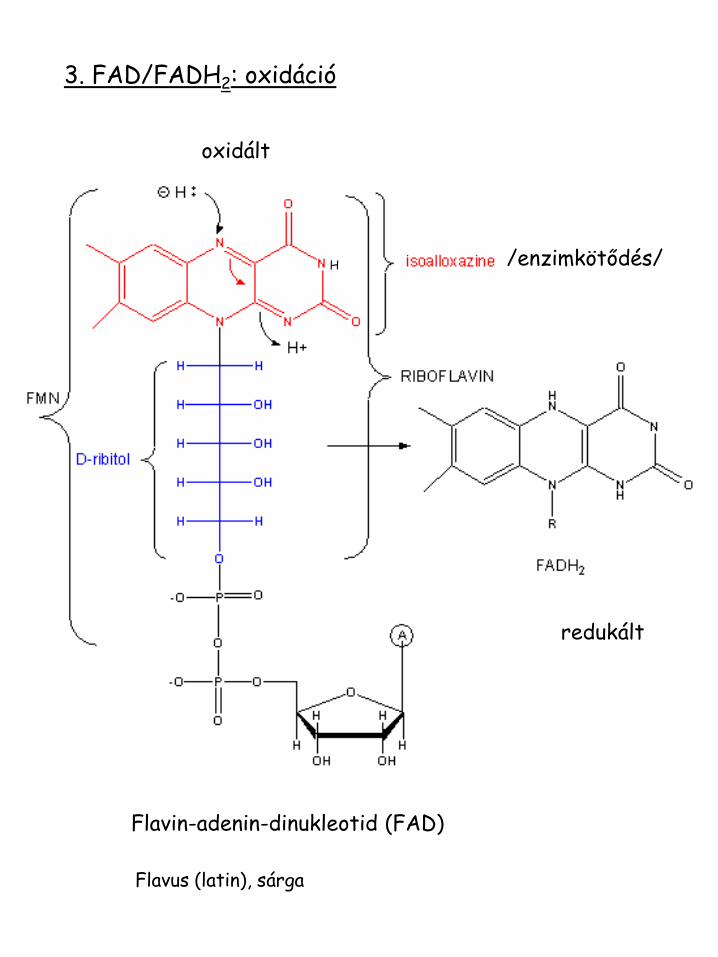
3. FAD/FADH2: oxidáció
Flavin-adenin-dinukleotid (FAD)
/enzimkötődés/ H
oxidált
redukált
Flavus (latin), sárga

4. Koenzim-A (CoA) felfedezés: Lipmann, 1946
adenozin-3’,5’-difoszfát
2,4-dihidroxi-3,3-dimetil-vajsav
b-alanin ciszteamin
pantoténsav (vitamin)
5’
3’
észter kötés
észter kötés
savamid kötés
savamid kötés
- NH –(CH2)2 - SH - NH –(CH2)2 - S – CO – CH3
S-acetil koenzim-A (Ac-CoA)

5. Koenzim-Q (ubikinon, CoQ)
(eukarióta sejtekben, mitokondriumok)
szemikinon
ubikinol
Koenzim Q4, n = 4 Koenzim Q9, n = 9 Koenzim Q10, n = 10
(Izoprén egységek)
ubikinon
Peter D. Mitchell, 1961 (Nobel díj, kémia, 1978) „for his contribution to the understanding of biological energy transfer through the formulation of the chemiosmotic theory". Később: elektrontranszport lánc koncepció – CoQ10 „protonvonzó”
1920-1992
http://www.q10facts.com/a-brief-history-of-coenzyme-q10-2/

VITAMINOK
1912: vitamin = vita amines (Casimir Funk)
„Kis mennyiségben kívülről kell felvenni.”
Nem vitaminok: esszenciális aminosavak
esszenciális zsírsavak
Provitaminok
Antropocentrikus
pl. C-vitamin: ember (tengeri malac) igen
patkány nem
p-amino-benzoesav: baktérium igen
ember nem
Kémiai szerkezet: heterogén Felosztás (oldékonyság alapján)
Vízoldékony Zsírban oldódó
B, C, H és P vitaminok A, D, E és K vitaminok
- koenzimek is - bélből szívódnak fel
- felesleg kiürül - májban raktározódnak
1958: 15-20 féle vitamin

B1 vitamin (tiamin) szerkezet 1934, szintézis 1936
Beri-beri piroszőlősav nem bomlik le
N
N
NH2
CH3S
N+
CH3
CH2CH2OHN
N
NH2
CH3S
N+
CH3
CH2CH2O
O
O-
O P
O-
O-
O P
N
N
NH2
CH3S
N+
CH3
CH2CH2O
COOH
CH3
OH
O
O-
O P
O-
O-
O P
ATP
pirimidin tiazol
tiamin-pirofoszfát (koenzim)
COOH
O
CH3
piroszőlősav:
piroszőlősav-dekarboxiláz
„aktív piruvát”
• brit tengerészek (1747) • Ch. Eijkman (1897), F. G. Hopkins [orvosi Nobel-díj (1929)] • C. Funk – vita amin (1912)
1 2
3
tiamin
észter kötés
+
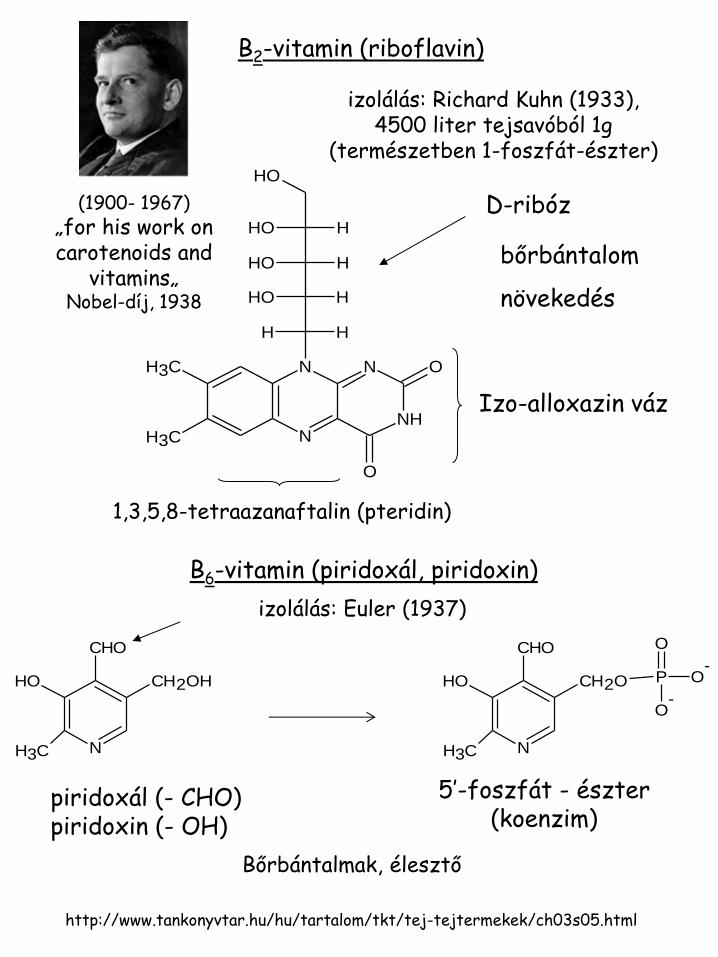
B2-vitamin (riboflavin)
izolálás: Richard Kuhn (1933), 4500 liter tejsavóból 1g
(természetben 1-foszfát-észter)
N N
NNH
O
O
OH
OH
OH
OH
CH3
CH3
H
H
H
HH
1,3,5,8-tetraazanaftalin (pteridin)
bőrbántalom
növekedés
B6-vitamin (piridoxál, piridoxin)
izolálás: Euler (1937)
NCH3
OH
CHO
CH2OH
NCH3
OH
CHO
CH2O O-
O-
O
P
piridoxál (- CHO) piridoxin (- OH)
5’-foszfát - észter (koenzim)
Bőrbántalmak, élesztő
Izo-alloxazin váz
D-ribóz
http://www.tankonyvtar.hu/hu/tartalom/tkt/tej-tejtermekek/ch03s05.html
(1900- 1967) „for his work on carotenoids and
vitamins„ Nobel-díj, 1938
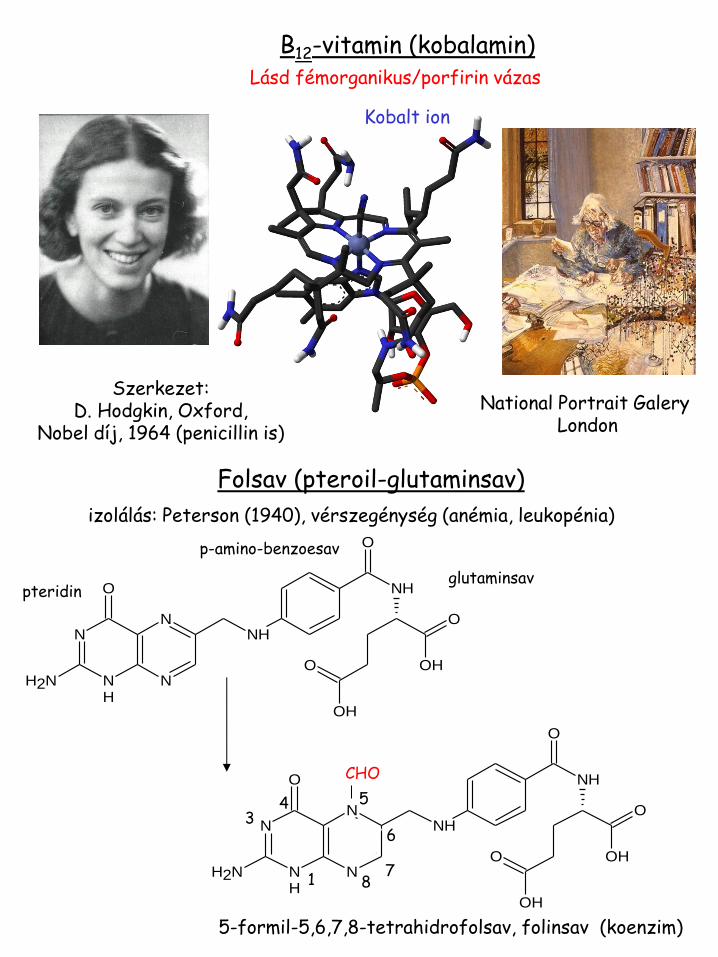
B12-vitamin (kobalamin)
Szerkezet: D. Hodgkin, Oxford,
Nobel díj, 1964 (penicillin is)
Kobalt ion
Lásd fémorganikus/porfirin vázas
National Portrait Galery London
Folsav (pteroil-glutaminsav)
izolálás: Peterson (1940), vérszegénység (anémia, leukopénia)
p-amino-benzoesav
N
NH
N
N
NHO
O
O
NH2
NH
O OH
OH
N
NH
N
N
NHO
O
O
NH2
NH
O OH
OH
glutaminsav pteridin
CHO
5-formil-5,6,7,8-tetrahidrofolsav, folinsav (koenzim)
5 4
1
3 6
7 8

C-vitamin (L-aszkorbinsav)
felfedezés: 1928 izolálás: Szentgyörgyi A. (1932)
O
OHOH
O
OH
OH
O
O
HO
HO
HO
H
H
CH2OH
=
•skorbut
1 3 4 6
H-vitamin (biotin)
imidazolidin
tetrahidrotiofén
*
* *
izolálás: Kögl (1936) 2800 kg tojássárgája 390 mg
2-keto-L-gulonsav g-laktám
1
2
6
L-aszkorbinsav)
Avidin – biotin Kd = 10-15
1 tojás 50 g, ebből 17 g sárgája 390 mg = 164 706 db tojás
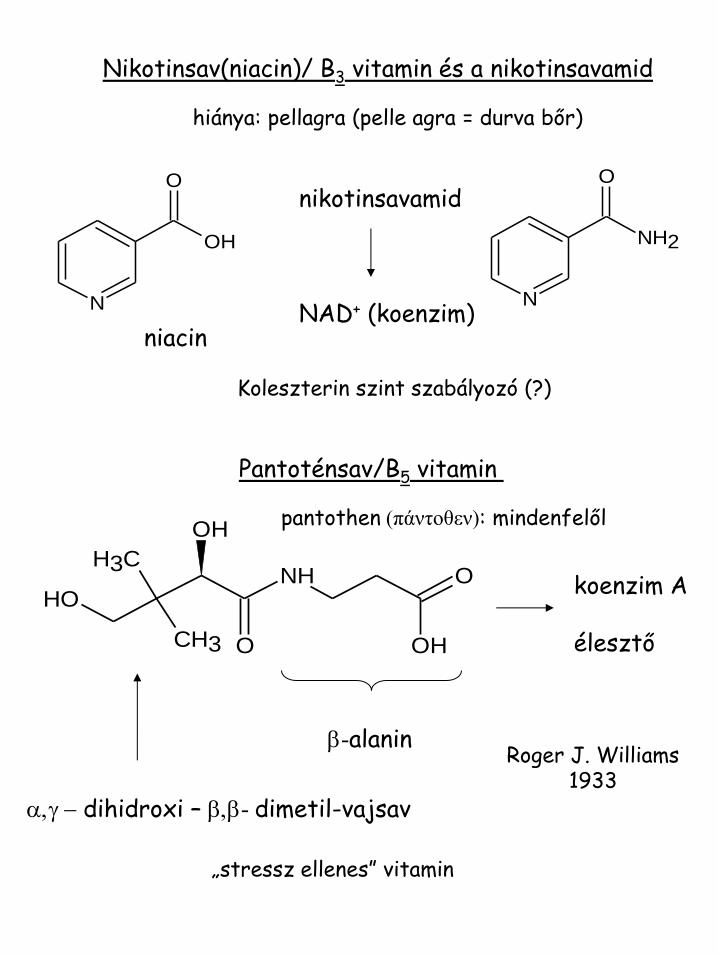
Nikotinsav(niacin)/ B3 vitamin és a nikotinsavamid
N
O
OH
N
O
NH2
hiánya: pellagra (pelle agra = durva bőr)
nikotinsavamid NAD+ (koenzim)
Pantoténsav/B5 vitamin
O
ONH
OH
OH
CH3
CH3
OH
b-alanin
koenzim A élesztő
niacin
a,g - dihidroxi – b,b- dimetil-vajsav
Koleszterin szint szabályozó (?)
„stressz ellenes” vitamin
pantothen (πάντοθεν): mindenfelől
Roger J. Williams 1933

A-vitamin
OH
O
provitamin: b-karotin
A vitamin
világos sárga olaj, fp.: 136°C A1 = retinol ,A2 = dehidroretinol
12
11
11-cisz-retinal
Kuhn, 1931, halak májából,farkasvakság, 0,5-1 mg/nap, 1 NE = 0,6 mg
CH2=C-CH=CH2
CH3
rodopszin
K1-vitamin (fillokinon) sárga olaj
O
O
CH3
CH3
CH3CH3
CH3CH3
2
4
3
1
fitil-csoport
naftokinon
2-metil-3-fitil-1,4-naftokinon
Előfordulás: káposztafélék, spenót, saláta, brokkoli, tejtermék, máj
Szerepe: antioxidáns, gyulladásgátló, Koaguláció Hiánya: vérzékenység (koaguláció hiánya miatt) Bomlás: fény, hő (100oC felett)
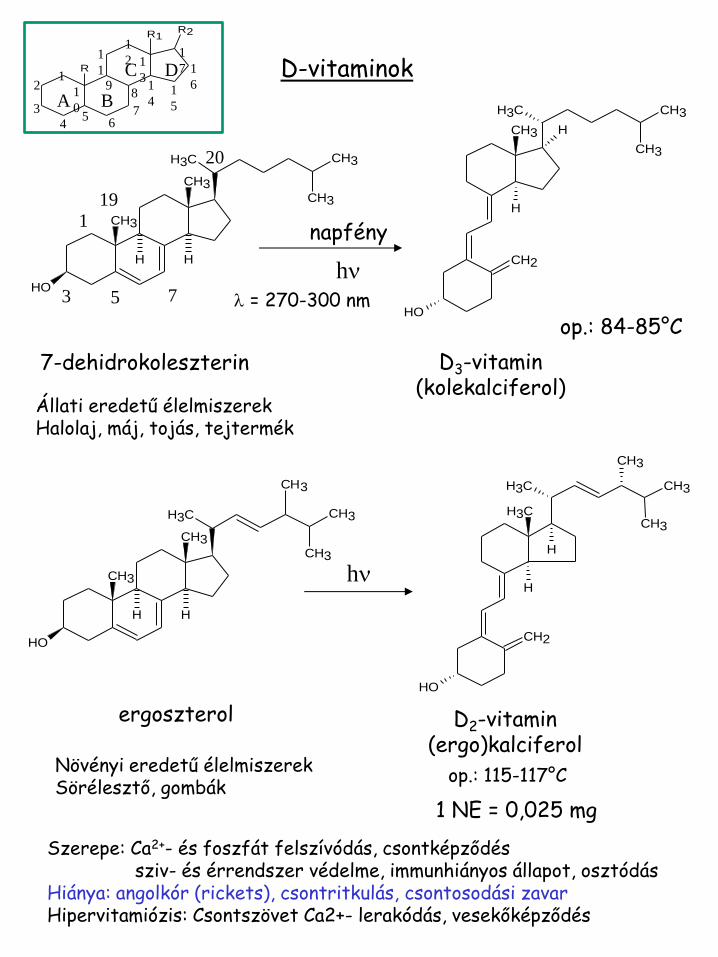
D-vitaminok
CH2
OH
CH3
CH3
CH3
CH3
H
H
CH3
CH3
HH
CH3
CH3
OH
CH3
napfény
hn
7-dehidrokoleszterin D3-vitamin (kolekalciferol)
op.: 84-85°C
Szerepe: Ca2+- és foszfát felszívódás, csontképződés sziv- és érrendszer védelme, immunhiányos állapot, osztódás Hiánya: angolkór (rickets), csontritkulás, csontosodási zavar Hipervitamiózis: Csontszövet Ca2+- lerakódás, vesekőképződés
CH2
CH3
OH
CH3
CH3
CH3
CH3
H
HCH3
CH3
HH
CH3
CH3
OH
CH3
CH3
hn
ergoszterol D2-vitamin (ergo)kalciferol
op.: 115-117°C
1 NE = 0,025 mg
l = 270-300 nm 3 7 5
19 1
20
Állati eredetű élelmiszerek Halolaj, máj, tojás, tejtermék
Növényi eredetű élelmiszerek Sörélesztő, gombák
R2
R
R1
A B
C D 1 2
3
4 5
6 7
8 9
1
0
1
1
1
2 1
3 1
4 1
5
1
6
1
7

E-vitamin (tokoferol)
O
R2
R1
R3
CH3
CH3
CH3
CH3CH3
CH3
izolálás 1936 felfedezés 1927 szintézis 1938
R1=R2=R3=CH3 a-tokoferol (5,7,8-trimetil)
R1=R3=CH3, R2= H b-tokoferol (5,8-dimetil)
R2=R3=CH3, R1= H g-tokoferol (7,8-dimetil)
halványsárga olaj
•antisterilitás (patkány)
5
7
kromán váz C16 lánc
Előfordulás: növényi olajok, napraforgó, oliva, tökmag hús, máj, tojás, tejtermék Szerepe: antioxidáns, gyulladásgátló Hiánya: vérszegénység, izomsorvadás, meddőség (embernél ritka) Hipervitamiózis: Csontszövet Ca2+- lerakódás, vesekőképződés
O
R2
R1
R3
CH3
CH3
CH3
CH3CH3
CH3
E-vitamin (tokotrienol)
τόκος [tókos], φέρειν [phérein] = születés, hordozni





























