Embed Size (px)
DESCRIPTION
Биокинетика. Чернышев Андрей Витальевич ( лекции ) [email protected] Некрасов Вячеслав Михайлович (семинары) [email protected] р.т.: 333-32-40. Гл. 1. Введение в биокинетику. Установление количественных закономерностей биологических процессов на молекулярном уровне. Цель:. - PowerPoint PPT Presentation
Citation preview

БиокинетикаБиокинетика
Чернышев Андрей ВитальевичЧернышев Андрей Витальевич((лекциилекции))
Некрасов Вячеслав МихайловичНекрасов Вячеслав Михайлович(семинары)(семинары)
р.т.: р.т.: 333-32-40333-32-40
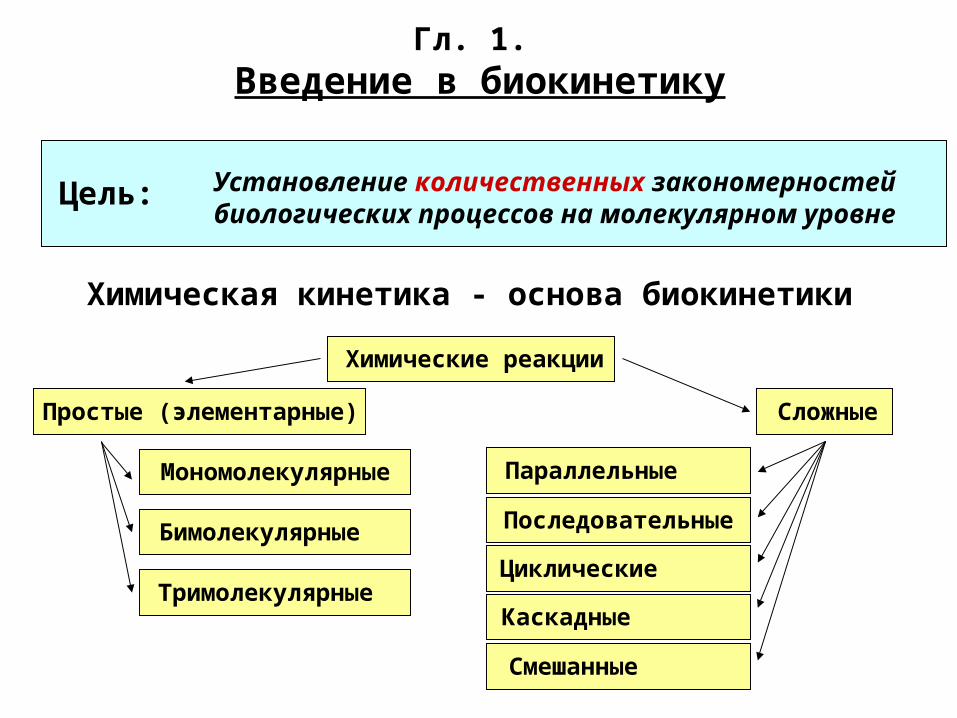
Гл. 1. Введение в биокинетику
Установление количественных закономерностей биологических процессов на молекулярном уровне
Химическая кинетика - основа биокинетики
Цель:
Химические реакции
Простые (элементарные) Сложные
Мономолекулярные
Бимолекулярные
Тримолекулярные
Параллельные
Последовательные
Циклические
Смешанные
Каскадные
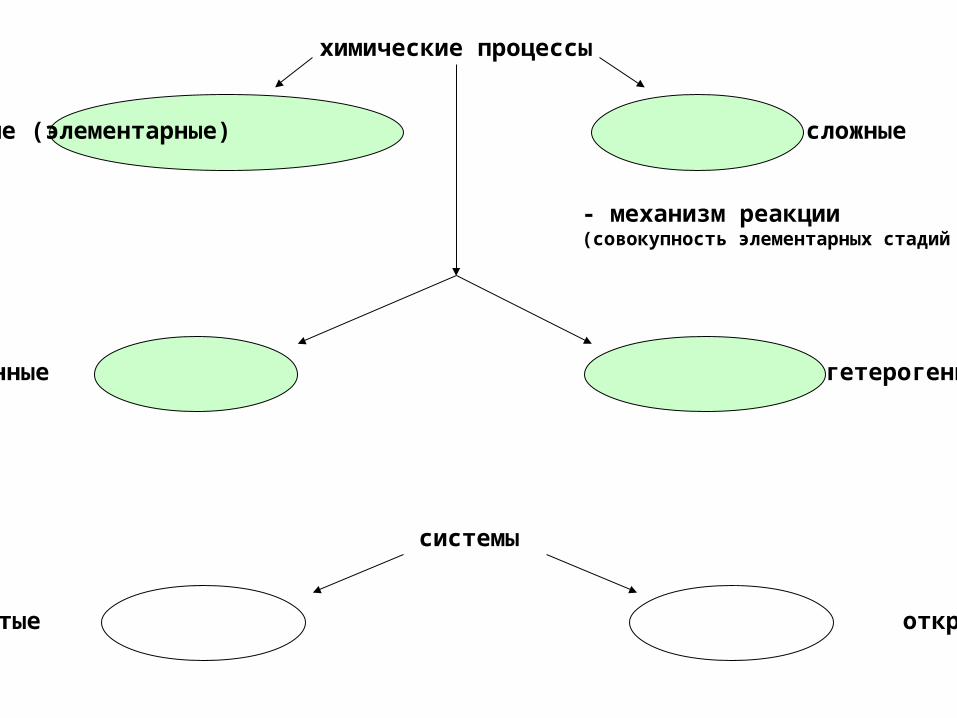
простые (элементарные) сложные
химические процессы
- механизм реакции(совокупность элементарных стадий )
гомогенные гетерогенные
замкнутые открытые
системы
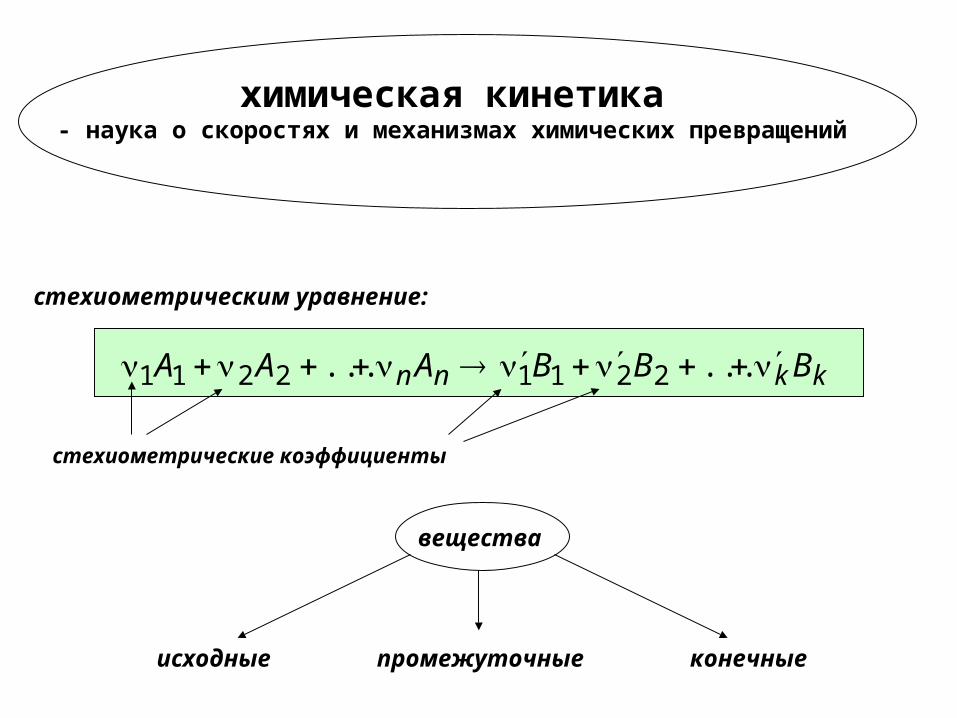
химическая кинетика - наука о скоростях и механизмах химических превращений
kknn BBBAAA ...... 22112211
стехиометрическим уравнение:
стехиометрические коэффициенты
исходные промежуточные конечные
вещества

Скорость химической реакции
dt
dN
Vdt
dN
Vw ji
B
j
A
i
1111
dt
Bd
dt
Adw
j
j
i
i
11V = const
Элементарная реакция: w = число элементарных актов реакции в единице объема в единицу времени.
Сложная реакция: может быть введено понятие единой скорости процесса, когда концентрации промежуточных веществ пренебрежимо малы
по сравнению с концентрациями реагентов и продуктов реакции (иначе - скорость реакции по конкретному веществу).
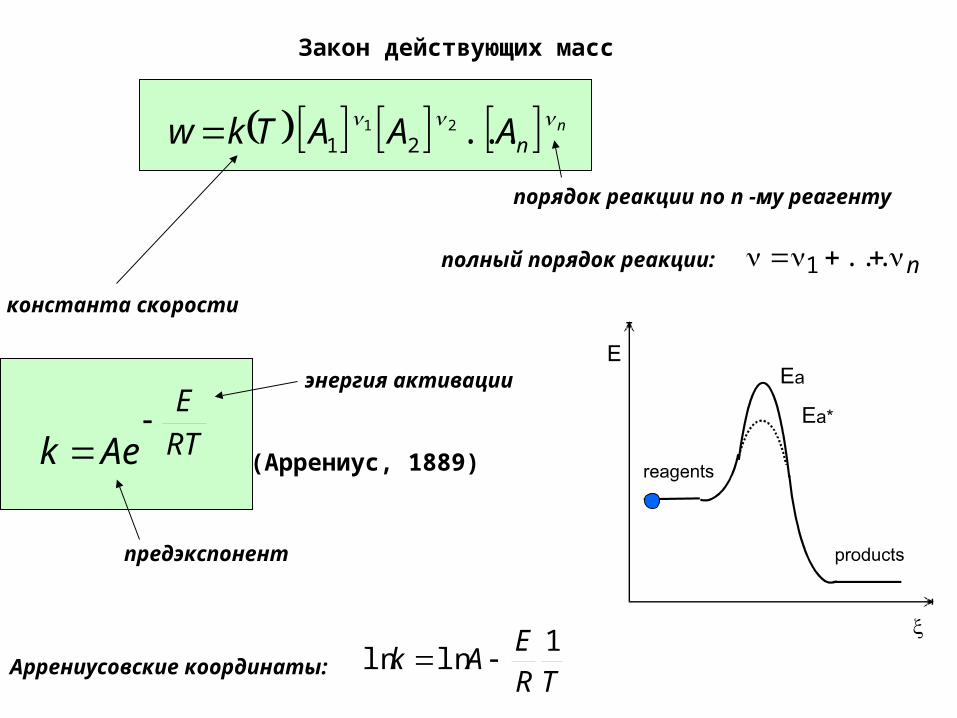
Закон действующих масс
n
nAAATkw ...21
21
константа скорости
порядок реакции по n -му реагенту
n ...1полный порядок реакции:
RT
E
Aek
(Аррениус, 1889)
энергия активации
предэкспонент
TR
EAk
1lnln Аррениусовские координаты:

Закон действующих масс с учетом микроскопических состояний
)(...)()()(...)()( 222111222111 kkknnn jBjBjBiAiAiA
nkn
llnnjjjiii
iA
liAiAiAk
dt
dN
Vw
)(...)()(
1121
2121 2211......)(
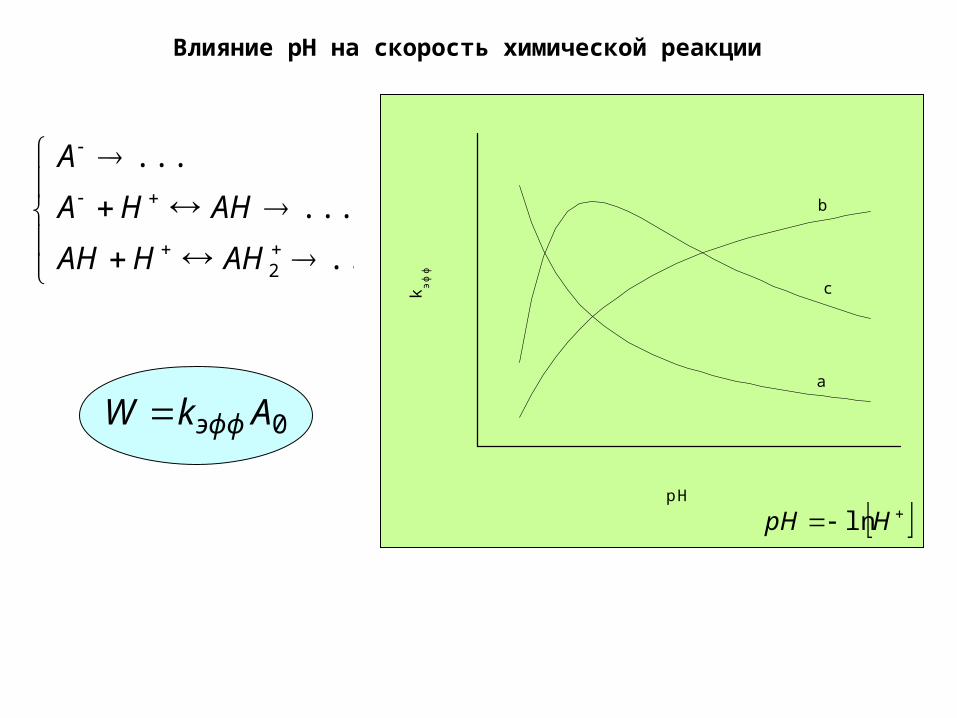
Влияние pH на скорость химической реакции
...
...
...
2AHHAH
AHHA
A
0AkW эфф
c
b
a
k эф
ф
pH
HpH ln
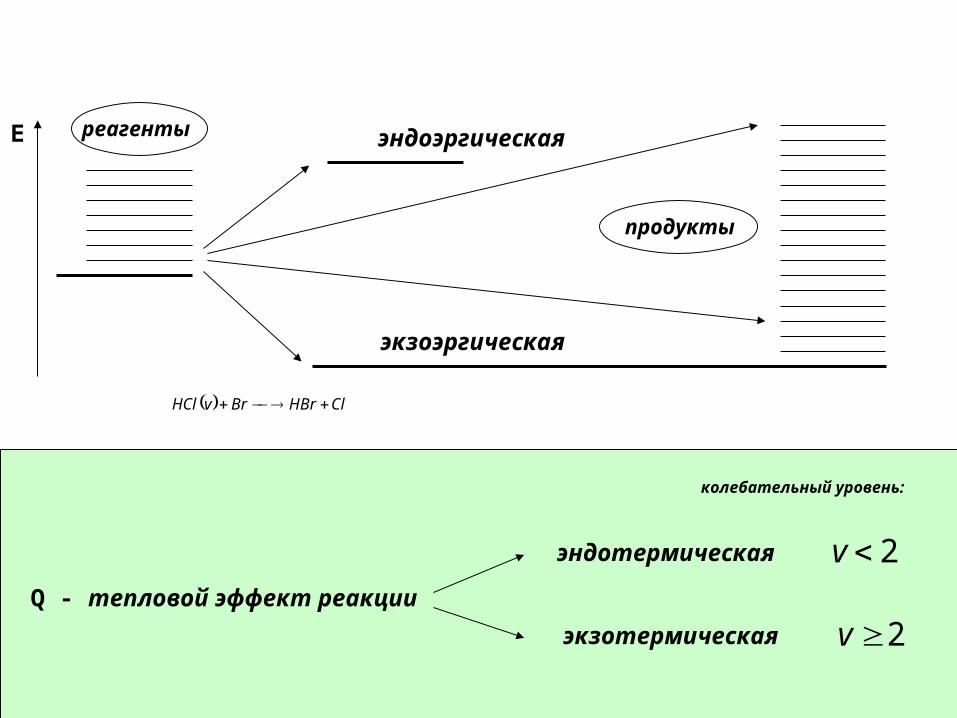
Q - тепловой эффект реакции
E реагенты
продукты
экзоэргическая
эндоэргическая
эндотермическая
экзотермическая
ClHBrBrvHCl
2v
колебательный уровень:
2v
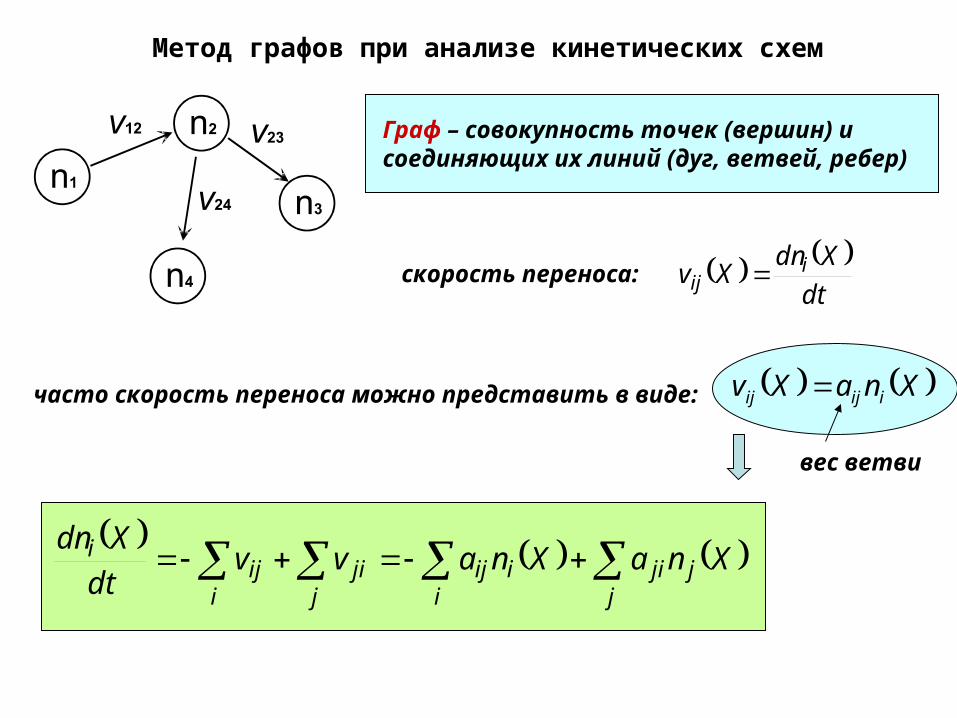
Метод графов при анализе кинетических схем
Граф – совокупность точек (вершин) и соединяющих их линий (дуг, ветвей, ребер)
dt
XdnXv i
ij
XnaXv iijij часто скорость переноса можно представить в виде:
скорость переноса:
вес ветви
j
jjii
iijj
jii
iji XnaXnavvdt
Xdn
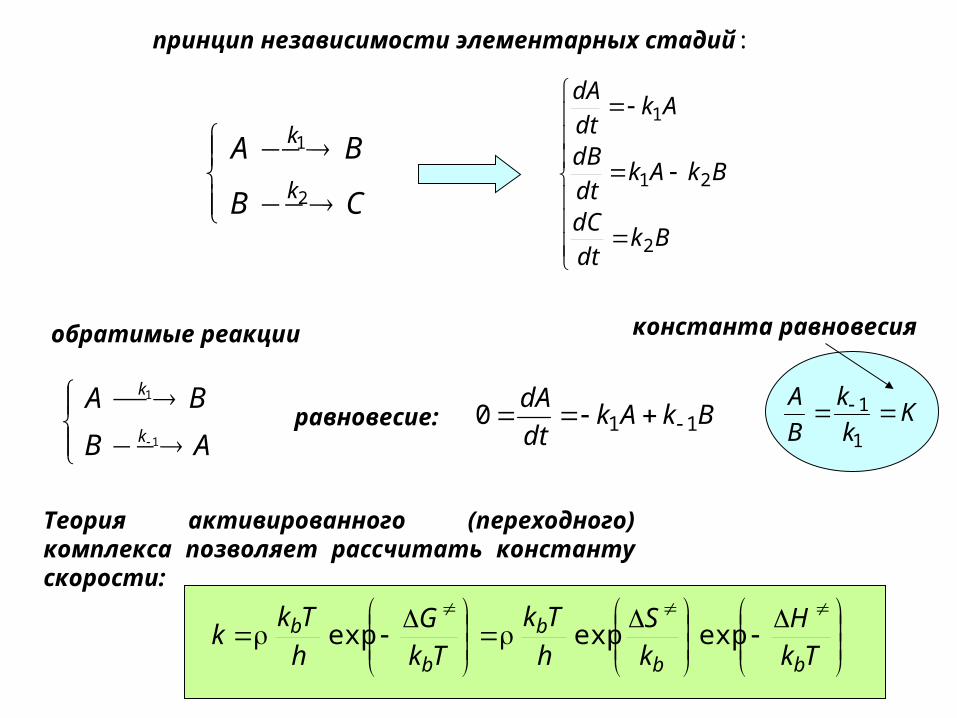
принцип независимости элементарных стадий:
CB
BAk
k
2
1
Bkdt
dC
BkAkdt
dB
Akdt
dA
2
21
1
обратимые реакции
AB
BAk
k
1
1
BkAkdt
dA110 K
k
k
B
A
1
1равновесие:
константа равновесия
Tk
H
k
S
h
Tk
Tk
G
h
Tkk
bb
b
b
b expexpexp
Теория активированного (переходного) комплекса позволяет рассчитать константу скорости:

m
jjj
kn
iii AA
11
Теория активированного (переходного) комплекса
kT
E
n
iA
e
Z
Z
h
kTk
i
0
1
~
~
Статистические суммы:
ядэлколврп ZZZZZZ
2/3
3)3( 2 mkT
hV
Zп
2)1( 21
IkTZвр
2)2( 21
IkTZвр
3
2/32/1)3( 21
kTIIIZ CBA
вр
1
1
1
s
i
kT
h
кол
i
eZ
.. состоснэл gZ
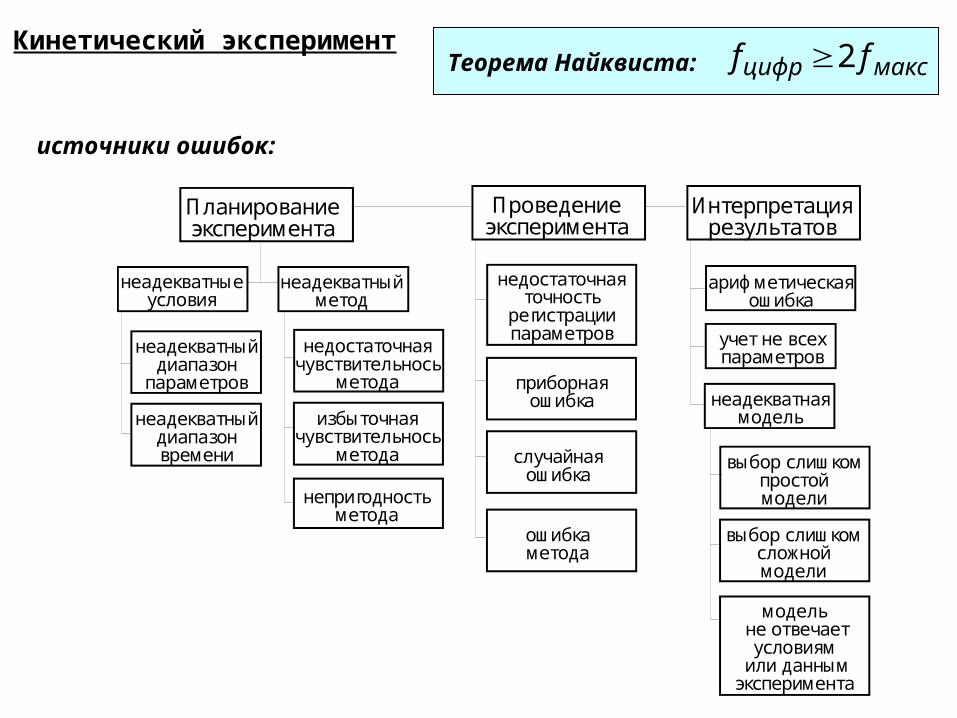
Кинетический эксперимент Теорема Найквиста: максцифр ff 2
Планированиеэксперимента
Проведениеэксперимента
Интерпретациярезультатов
неадекватныеусловия
неадекватныйдиапазон
параметров
неадекватныйдиапазонвремени
неадекватныйметод
недостаточнаячувствительнось
метода
избыточнаячувствительнось
метода
непригодностьметода
недостаточнаяточность
регистрациипараметров
приборнаяошибка
случайнаяошибка
ошибкаметода
неадекватнаямодель
выбор слишкомпростоймодели
модель не отвечает
условиям или данным
эксперимента
выбор слишкомсложноймодели
учет не всехпараметров
арифметическаяошибка
источники ошибок:
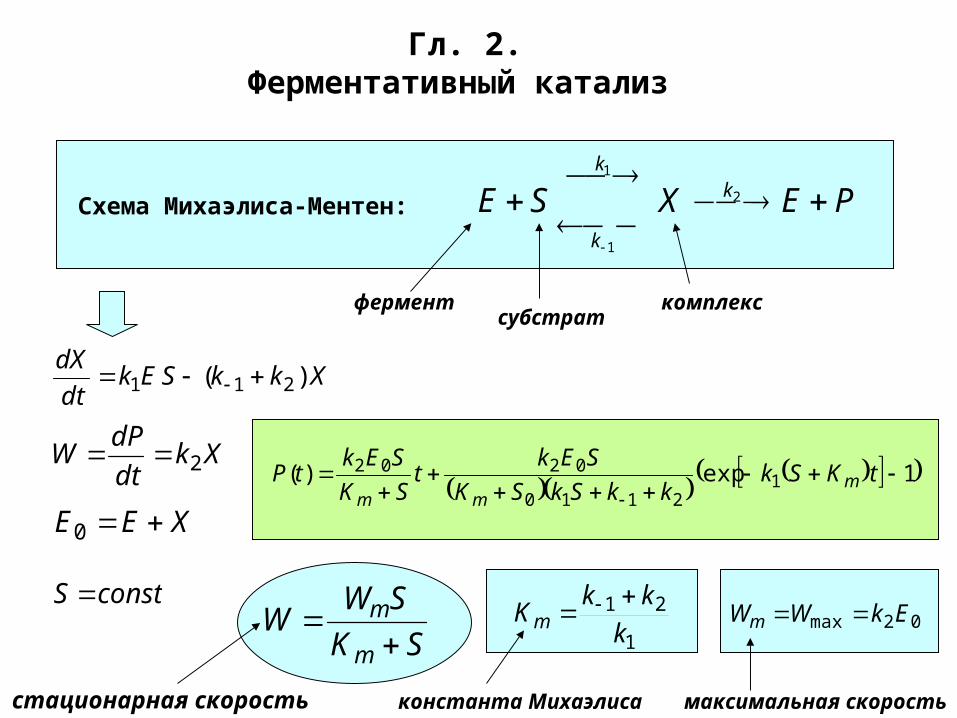
Гл. 2.Ферментативный катализ
Схема Михаэлиса-Ментен: PEXSE k
k
k
2
1
1
ферментсубстрат
комплекс
XkkSEkdt
dX)( 211
Xkdt
dPW 2
XEE 0
constS
1exp)( 12110
0202
tKSkkkSkSK
SEkt
SK
SEktP m
mm
SK
SWW
m
m
1
21
k
kkKm
02max EkWWm
константа Михаэлиса максимальная скоростьстационарная скорость

Схема Wm Km
Некоторые кинетические схемы, приводящие к уравнению Михаэлиса
PESX
XSEk
k
2
1
2
1
PEXXSE kk
k
k
32
1
1
21
PEXXSE nkn
k
k
k
12
1
1
...1
011 ESk
032
32 Ekk
kk
1
2
0
1n
i ik
E
2
11
k
Sk
3121
31
kkkk
kk
1
221
1
1n
i ikkk
k
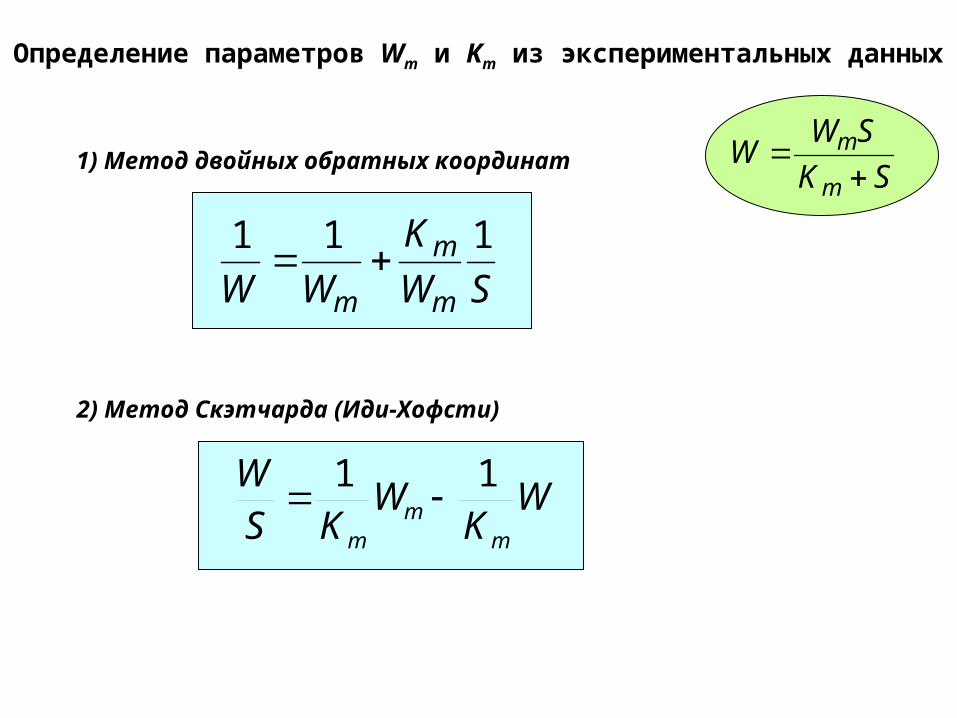
Определение параметров Wm и Km из экспериментальных данных
1) Метод двойных обратных координат
2) Метод Скэтчарда (Иди-Хофсти)
SW
K
WW m
m
m
111
WK
WKS
W
mm
m
11
SK
SWW
m
m

Определение концентрации активных центров:
По содержанию белка(если один активный центр)
Путем необратимого ингибирования
m
PE 0
)( 00 IEkW
1.
2.
Определение имитирующей стадии:
Измерение нестационарной кинетики
Использование субстратов с различной структурой и различной реакционной способностью
1.
2.
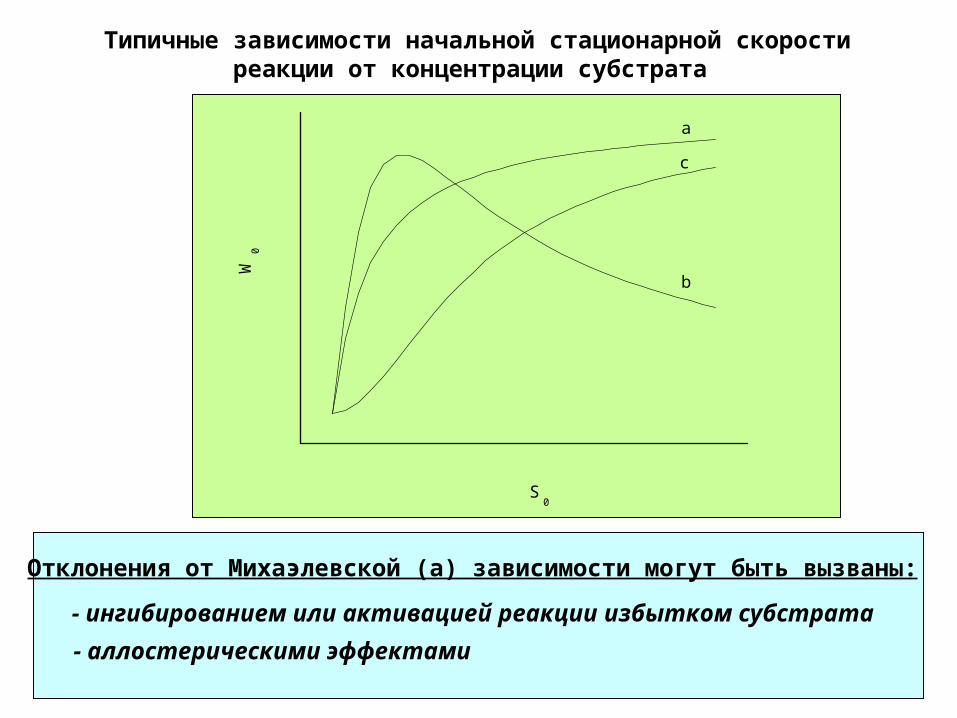
Типичные зависимости начальной стационарной скорости реакции от концентрации субстрата
c
b
a
W0
S0
- аллостерическими эффектами
- ингибированием или активацией реакции избытком субстрата
Отклонения от Михаэлевской (а) зависимости могут быть вызваны:
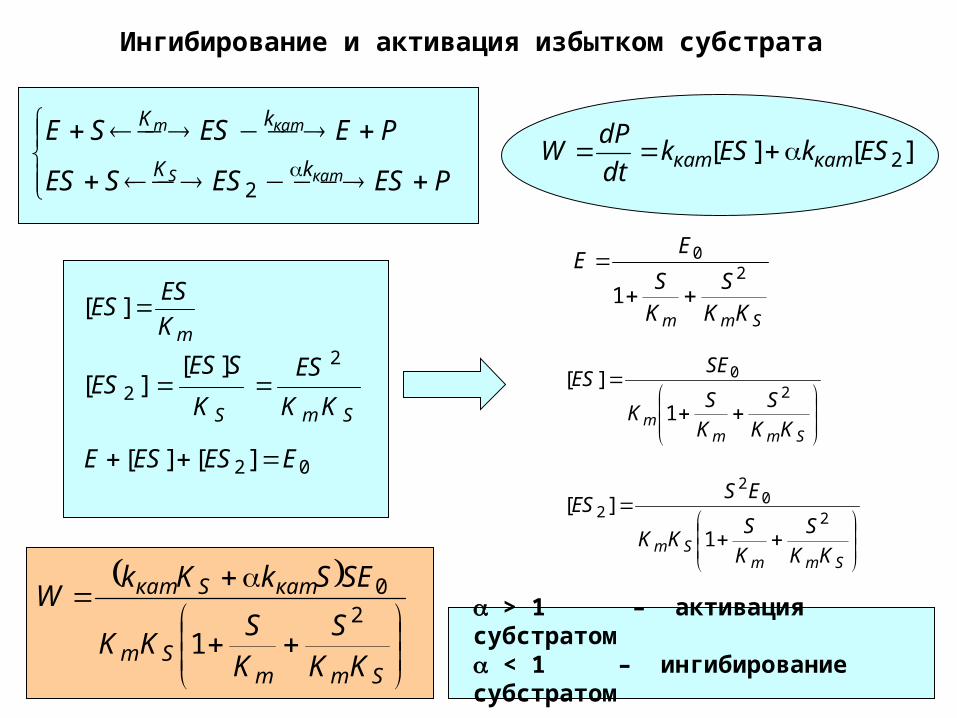
Ингибирование и активация избытком субстрата
PESESSES
PEESSE
катS
катm
kK
kK
2
][][ 2ESkESkdt
dPW каткат
mK
ESES ][
SmS KK
ES
K
SESES
2
2][
][
02 ][][ EESESE
Smm KK
S
K
S
EE
20
1
Smmm KK
S
K
SK
SEES
20
1
][
SmmSm KK
S
K
SKK
ESES
20
2
2
1
][
SmmSm
катSкат
KK
S
K
SKK
SESkKkW
20
1 > 1 – активация субстратом < 1 – ингибирование субстратом
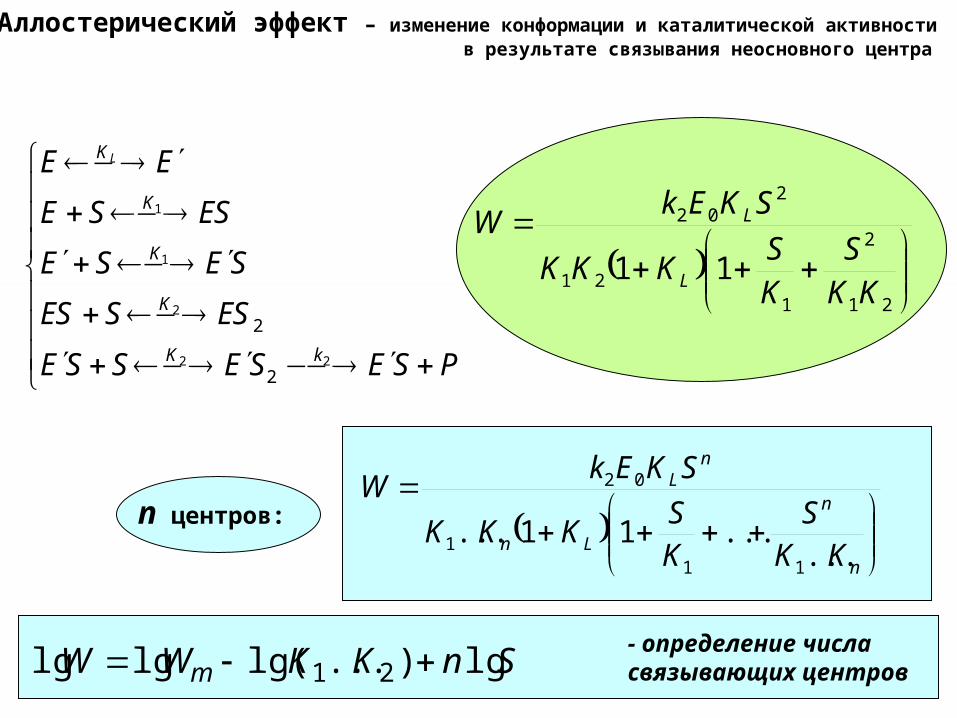
Аллостерический эффект – изменение конформации и каталитической активности в результате связывания неосновного центра
PSESESSE
ESSES
SESE
ESSE
EE
kK
K
K
K
KL
22
2
1
1
2
2
21
2
121
202
11KK
SKS
KKK
SKEkW
L
L
n
n
Ln
nL
KKS
KS
KKK
SKEkW
......11...
111
02
n центров:
SnKKWW m lg)...lg(lglg 21 - определение числасвязывающих центров

Многосубстратные реакции
Механизм тройного комплекса “Пинг-понг” механизм
PESES
SESSES
ESSE
k
K
K
21
2121
11
2
1
21
21
1
121
021
1KKSS
KS
KK
ESkSW
21
21
1
12110 ][][
KK
SES
K
ESESESESEE
21
21
1
1
0
1KK
SS
K
SE
E
PEXSSX
XESSEkK
kK
22
11
22
11
2
22
1
112211 ][][0
K
XSk
K
ESkXSkESk
dt
dX
2
2
212
121
1
1210 1][][
K
S
SKk
ESKk
K
ESEXSXESEE
2
2
212
121
1
1
0
11K
S
SKk
SKk
K
S
EE
2
2
2
1
1
1
01
11SK
kk
SK
EkW

Нестационарная кинетика ферментативных реакций
Предстационарная кинетика многостадийной реакции
PEXXSE nkn
k
k
k
k
1
2
2
1
1
...1
j
n
iiijj CXBX
dt
d
1
nn
n
ii
nnnnnn
iiiiiiii
XkdtdP
XEE
XkkXkdt
dX
XkkXkXkdt
dX
XkkXkSEkdt
dX
1
10
11
1111
1212211
...
...
01
)( AeAtXn
i
tjij
i
tAkeA
ktP n
n
i
t
i
nin
i01
11 1)(
i – собственные значения матрицы Bij Aij и A0 – константы

Релаксационная кинетика
При быстром внешнем воздействии на систему (изменение температуры, давления, пр.) время, которое нужно системе для достижения нового равновесия (или стационарного состояния), зависит от скорости химической реакции (и иногда от скорости диффузии реагентов).
EAAEk
k
1
1 резко меняется температура: T →T0 + T
меняется константа равновесия: K → K0 + K
TRT
HK
KK
20
ln
][][
11 EAkEAkdt
EAd
Кинетика перехода в новоеравновесное состояние:
)(
)(
)]([][][
tAAA
tEEE
tEAAEEA
Если отклонение от равновесия невелико:
][][][
11111 EAkAkEkAEkAEkdt
EAd
tkAkEkkAkEk
AEkAEkEA 111
111
11 exp1][
][
1111
kAkEk - время релаксации
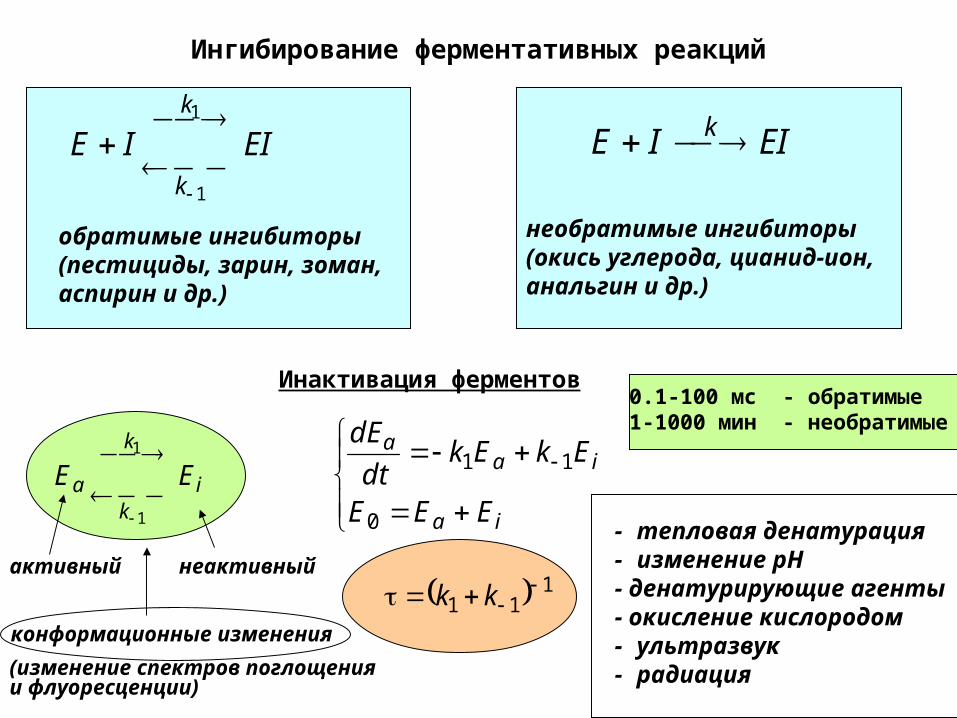
Ингибирование ферментативных реакций
EIIEk
k
1
1
обратимые ингибиторы(пестициды, зарин, зоман, аспирин и др.)
EIIE k
необратимые ингибиторы (окись углерода, цианид-ион, анальгин и др.)
Инактивация ферментов
ik
k
a EE
1
1
активный неактивный
ia
iaa
EEE
EkEkdt
dE
0
11
111
kk
- тепловая денатурация- изменение pH - денатурирующие агенты- окисление кислородом- ультразвук- радиация
конформационные изменения
(изменение спектров поглощения и флуоресценции)
0.1-100 мс - обратимые 1-1000 мин - необратимые

1. Простейшая кинетическая схема инактивации с равновесием конформеров:
ikK EEE 21
k
K
1
2. Инактивации подвержены оба конформера:
ik
ik
K
EE
EE
EE
22
11
21
2
1
Kkk
K
21
1
3. Более общий случай для системы с участием n конформеров:
k
KKKK nn 1221 1...111
ik
nKKK EEEEE n 121 ...321

4. Диссоциативный механизм инактивации:
5. Инактивация в процессе реакции:
ikK EEE in 222
ik
ik
ik
ik
kK
EPE
ESE
ESES
EE
PEESSE
4
3
2
1

Методы дискриминации механизмов инактивации и определения кинетических характеристик реакции:
- анализ зависимости выхода продукта от концентрации фермента;
- зависимость степени конверсии субстрата от степени инактивированности фермента;
- проведение реакции при низких степенях конверсии субстрата и низких концентрациях фермента;
- проведение реакции при больших концентрациях фермента;
- предынкубация фермента с компонентами реакции;
- использование интегральных уравнений реакции.

Полиферментные системы. Сопряженные ферментные реакции
PSS EE 2121
22
22
22
22
11
112
SK
SV
dt
dP
SK
SV
SK
SV
dt
dS
m
m
m
m
m
m
избыток S1 :t
SK
SV
SK
SVdS
S
m
m
m
m
2
022
22
11
11
2
tVWW
V
K
S
WV
KVS m
m
mm
mm21
1
2
2
2
12
222 11ln
11
111 SK
SVW
m
m
0111
2
2
2
W
V
K
S m
m 11
2
22
W
VK
Sm
m
V2m>>W1 : tK
V
W
V
K
S
K
S
m
mm
mm 2
2
1
2
2
2
2
2 1ln
S2/K2m<<1 :
m
m
m K
S
W
V
K
S
2
2
1
2
2
21ln
tK
V
m
m m
m
eWV
KtS 2
2
1)( 12
22
1)( 2
2
12
21
tK
V
m
m m
m
eWV
KtWtP
m
m
V
K
2
2
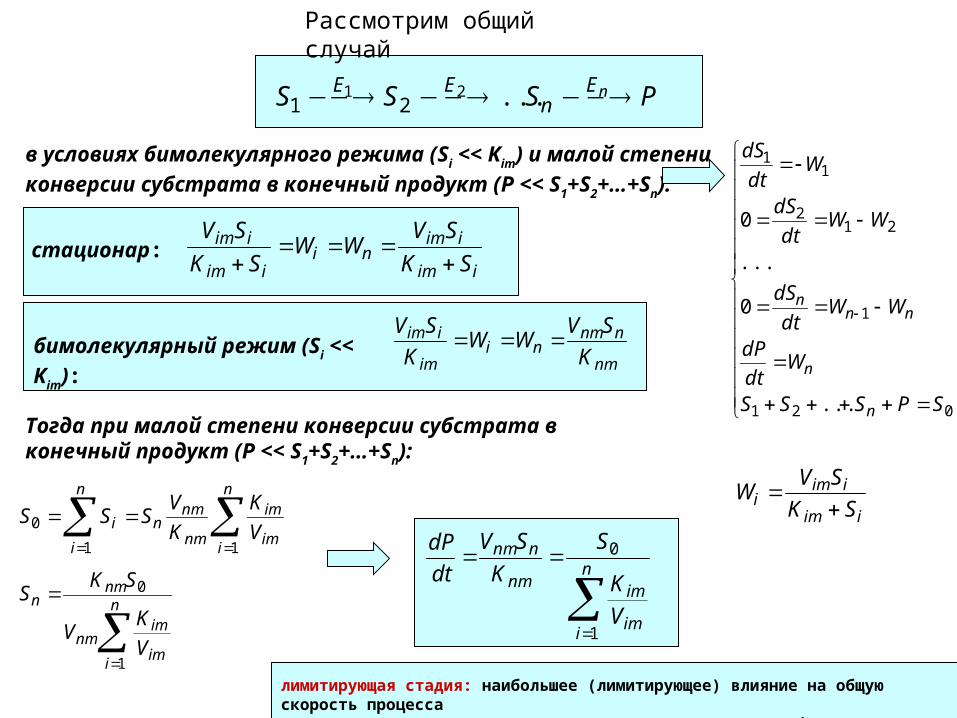
PSSS nEn
EE ...2121
Рассмотрим общий случай
в условиях бимолекулярного режима (Si << Kim) и малой
степени конверсии субстрата в конечный продукт (P << S1+S2+…+Sn):
021
1
212
11
...
0
...
0
SPSSS
Wdt
dP
WWdt
dS
WWdt
dS
Wdt
dS
n
n
nnn
iim
iimi SK
SVW
iim
iimni
iim
iim
SK
SVWW
SK
SV
стационар:
nm
nnmni
im
iim
K
SVWW
K
SVбимолекулярный режим (Si << Kim):
Тогда при малой степени конверсии субстрата в конечный продукт (P << S1+S2+…+Sn):
n
iim
im
nm
nmn
n
i
i V
K
K
VSSS
11
0
n
iim
imnm
nmn
V
KV
SKS
1
0
n
iim
imnm
nnm
V
K
S
K
SV
dt
dP
1
0
лимитирующая стадия: наибольшее (лимитирующее) влияние на общую скорость процесса оказывают ферменты, имеющие наименьший параметр Vim/Kim.
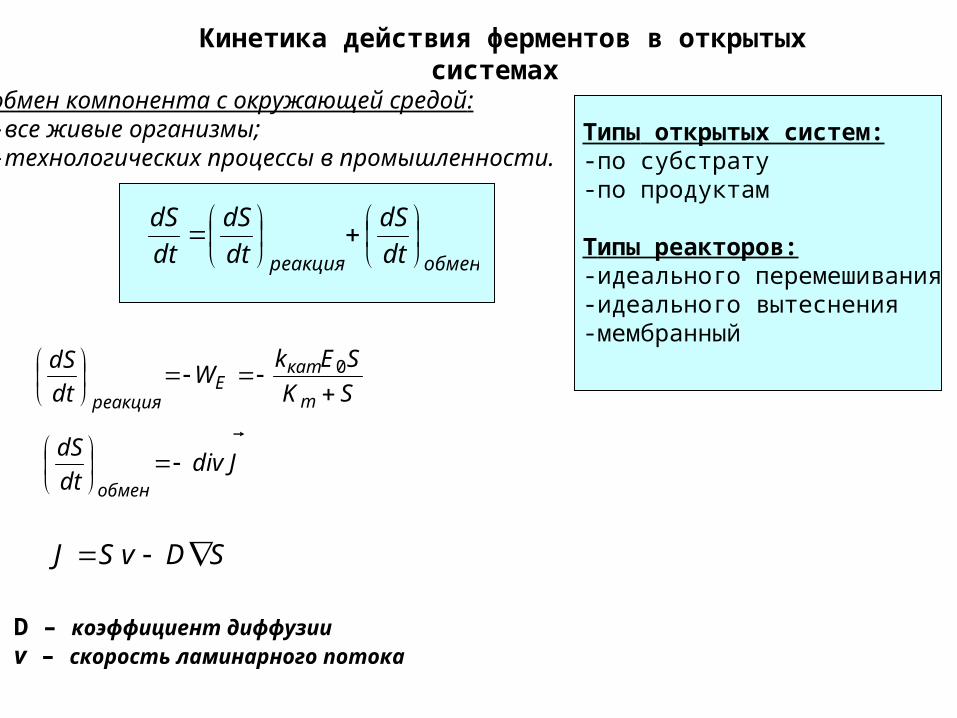
Кинетика действия ферментов в открытых системах
обмен компонента с окружающей средой:-все живые организмы; -технологических процессы в промышленности.
Типы открытых систем: -по субстрату-по продуктам
Типы реакторов:-идеального перемешивания-идеального вытеснения-мембранный
обменреакция dt
dS
dt
dS
dt
dS
SK
SEkW
dt
dS
m
катE
реакция
0
Jdivdt
dS
обмен
SDvSJ
D – коэффициент диффузииv – скорость ламинарного потока
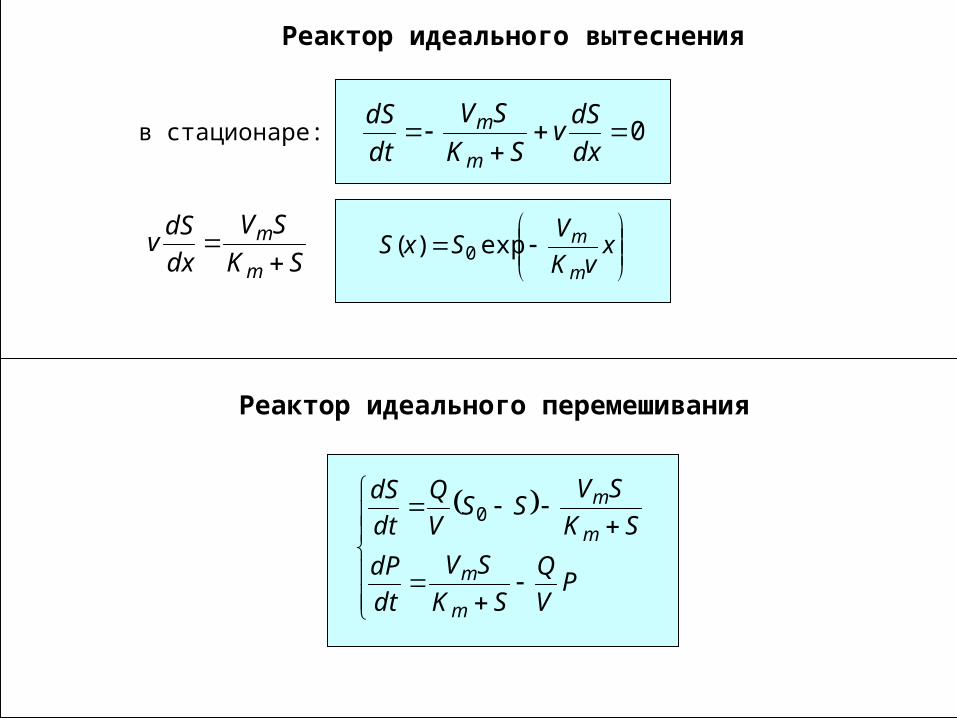
Реактор идеального вытеснения
0
dx
dSv
SK
SV
dt
dS
m
mв стационаре:
SK
SV
dx
dSv
m
m
x
vK
VSxS
m
mexp)( 0
Реактор идеального перемешивания
PV
Q
SK
SV
dt
dP
SK
SVSS
V
Q
dt
dS
m
m
m
m0

Гл. 3. Молекулярная рецепция
Рецепторы – химические соединения на поверхности или внутри клеток, посредством которых происходит распознавание веществ (лигандов) и формирование клеточного ответа.
Лиганды – химические вещества, способные реагировать с рецепторами.
LRRLk
k
1
1][
][11 LRkLRk
dt
LRd
Если L >> R: tkLk
k
kL
RLLR 101
1
10
00 exp1][
- изотерма Ленгмюра (сорбции)
][
][][
0
1
1
LRLRRL
LRRL
kk
K
L
LRLR
KR
K
][][
110 - уравнение Скэтчарда

рецепторы
- белки, различающиеся разными участками или третичной структурой;
- трансмембранные белки;
- образуют четвертичную структуру с углеводами, гликопротеидами, фосфолипидами мембран;
- в процессе функционирования может меняться третичная и четвертичная структуры (конформация рецептора, структура и состав связанных с ним молекул) и сам рецептор (фосфорилирование и дефосфорилировать и др.);
- центры связывания лигандов: COOH-группы дикарбоновых кислот, NH2-группы диаминовых кослот, OH-группы гидроксиаминокислот, SH-группы цистеина, гидрофобные участки аминокислот и др.; - участвует несколько активных участков связывающего центра;
- большая степень сродства к лигандам, чем у ферментов к субстратам.
лиганды
- различное химическое строение (белковые, пептидные и др.)

Лиганды:
Агонисты – связываясь с рецепторами, активно вызывают биологический ответ клетки (стимулируют клеточные функции).
Антагонисты – не вызывают активного клеточного ответа, препятствуют связыванию агонистов с рецепторами (угнетают кл. функции).
Принцип структурной комплиментарности (“ключ-замок”)
Проведение и усиление рецепторного сигнала:
- максимальный биологический ответ клетки наблюдался даже тогда, когда лишь незначительная часть рецепторов связана с лигандами;
- кривая зависимости биологического ответа клетки от концентрации добавленного лиганда во многих случаях имеет сложный колоколообразный вид;
- различие механизма действия агонистов и антагонистов рецепторов одного типа.

Вторичные мессенджеры – внутриклеточные молекулы, осуществляющие сопряжение рецепторов с внутриклеточными эфферентными системами (ферментами, ионными каналами, геномом и т.д.).
Первичными мессенджерами раньше называли лиганды, так как многие из лигандов осуществляют сопряжение эндокринных желез с эфферентными клетками.
Классификация рецепторов по их сопряжению со вторичными мессенджерами:
- рецепторы-ионные каналы (несколько глобул интегральных белков на мембране, в неактивном состоянии рецептора ионный канал закрыт);
- рецепторы, сопряженные с ферментами (одна из глобул белка-рецептора обладает каталитической активностью, образование лиганд-рецепторного комплекса приводит к изменению каталитической активности);
- рецепторы сопряженные с G-белками (рецепторная молекула сопряжена в с внутриклеточной стороны с особым ГТФ-связывающим белком, - G-белком, - через G-белок рецепторы могут менять активность внутриклеточных ферментативных систем и вызывать открытие ионных каналов).

G-белок сопряженные рецепторы
- семь раз пронизывает цитоплазматическую мембрану;- NH2-конец располагается внеклеточно;- три внутриклеточных и три внеклеточных “петли”;- лиганды обычно связывает вторая внеклеточная петля;- оканчивается внутри клетки гидрофильным COOH-концом;- третья внутриклеточная петля и COOH-конец осуществляют сопряжение с G-белком.
- состоит из двух субъединиц: и ;- -субъединица связывает ГТФ;- - субъединица осуществляет крепление G-белка к цитолазматической мембране;- -субъединица связывается с COOH концом и третьей петлей рецептора;- - субъединица связывается только с COOH-концом.
G-белок
1) Gs-белки (s-субъединица, стимулируют АТФ→цАМФ, ингибируют Ca2+ каналы);
2) Gi-белки (ингибируют АТФ→цАМФ);
3) Go-белки (угнетают потенциал-зависимые Ca2+ каналы, стимулируют К+ каналы);
4) др.

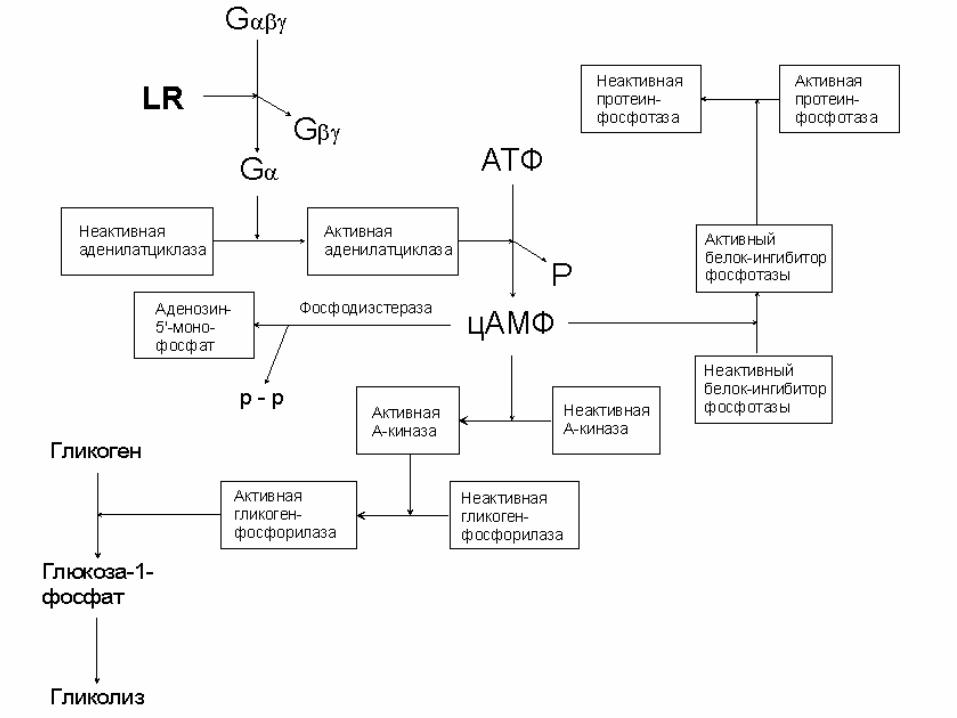
Схема функционирования G-сопряженного рецептора:
ГДФGГДФG
ГДФGGГДФG
EГДФGEГДФG
PEГДФGEГТФG
цАМФАТФ
EГТФGEГТФG
GГТФGRLГТФGRL
ГТФGRLГТФGRL
GRLGRL
RLRL
RLLR
EГТФG
клазааденилатци
aa
aa
aa
ai
ii
][
][][
][][
][][
][][
][
10-100 G-белков
10-100 молекул цАМФ
усиление в 100-10000 раз!
х
цАМФ
в 104-108 раз!!
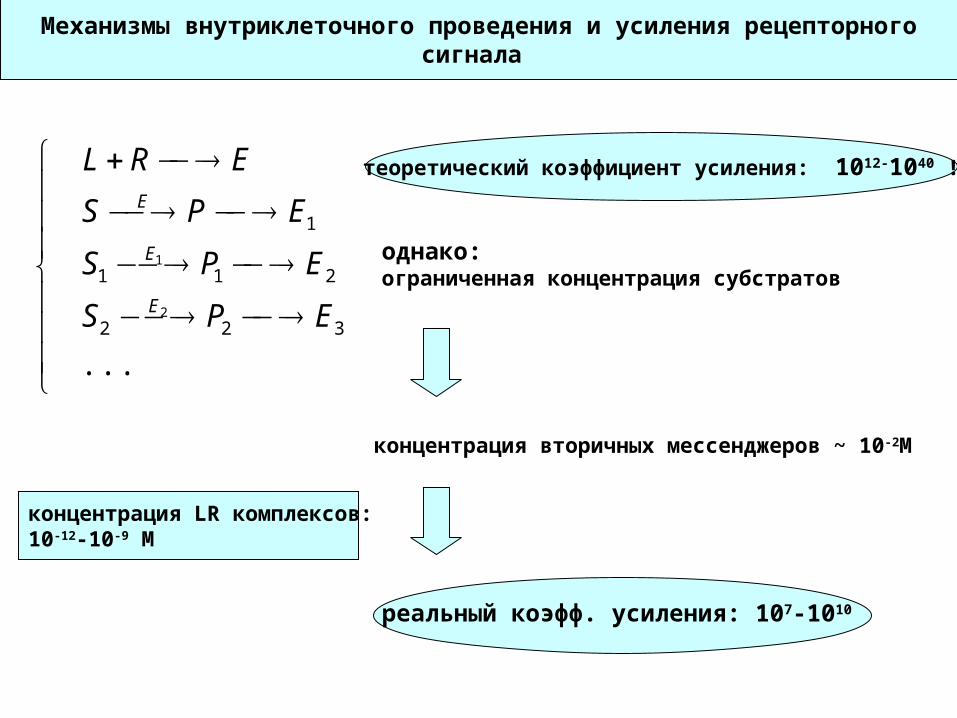
Механизмы внутриклеточного проведения и усиления рецепторного сигнала
...322
211
1
2
1
EPS
EPS
EPS
ERL
E
E
E
теоретический коэффициент усиления: 1012-1040 !
однако:ограниченная концентрация субстратов
концентрация вторичных мессенджеров ~ 10-2M
концентрация LR комплексов: 10-12-10-9 М
реальный коэфф. усиления: 107-1010

Инактивация рецепторного сигнала
I. Уменьшение концентрации LR комплексов:
а) Диссоциация LR (напр. из-за уменьшения концентрации лигандов в растворе вследствии разрушения лиганда ферментами).
б) Повышение константы скорости диссоциации лиганд-рецепторного комплекса (активация рецептора).
в) Изменение концентрации свободных или связанных рецепторов на мембране (эндоцитоз и экзоцитоз).
II. Инактивация сопряжения LR комплеска с ферментами:
а) Самоинактивация -субъединицы G-белка (ГТФ-азная активность -субъединицы).
б) Десенситизация ферментной системы (процессы фосфорилирования-дефосфорилирования, метилирования-деметилирования рецептора, G-белков, ферментов и т.д.).
III. Инактивация фермента, синтезирующего ключевые регуляторы клеточного ответа (напр. инактивация вторичным мессенджером – арахидоновой кислотой – ферментов синтеза эйкозаноидов (простагландинов, тромбоксанов, липоксинов)
IV. Взаимодействие лиганда с различными типами рецепторов: 1) стимулирует 2) ингибирует 3) не влияет

Дискриминация моделей
1) Ингибиторование рецепторов (использование конкурентных антагонистов).
2) Ингибирование мембранного транспорта (разрушение клеток).
3) Уменьшение мембранного транспорта пептидных фрагментов.
4) Уменьшение эндоцитоза (мембранные стабилизаторы).
5) Увеличение проницаемости мембран (пермиолизаторы).
6) Использование димеров лигандов (димеры труднее диффундируют).
7) Использование ингибиторов макроэргов (уменьшение скорости энергозависимых процессов транспорта и деградации ферментов).
8) Ингибирование ферментов (уменьшение скорости деградации лигандов).
9) Добавление ионов металлов (меняют аффинность рецепторов).
10) Изменение pH, ионной силы, температуры (транспорт, деградация, связывание и пр.).
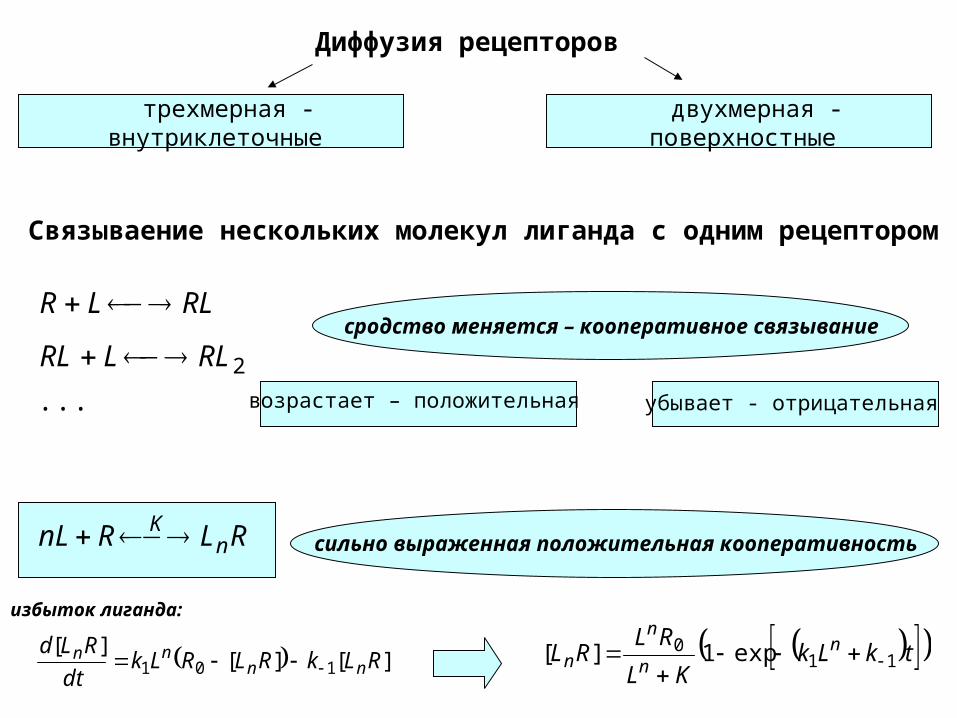
Диффузия рецепторов
трехмерная - внутриклеточные
двухмерная -поверхностные
Связываение нескольких молекул лиганда с одним рецептором
...2RLLRL
RLLR
сродство меняется – кооперативное связывание
возрастает – положительная убывает - отрицательная
RLRnL nK сильно выраженная положительная кооперативность
][][][
101 RLkRLRLkdt
RLdnn
nn tkLk
KL
RLRL n
n
n
n 110 exp1][
избыток лиганда:

Координаты Хилла
KLnRLR
RL
n
n logloglog0
LRLR
RL
n
n log;log0
Тангенс угла наклона () связан со степенью коопретивности: > 1 положительная кооперативность, < 1 отрицательная коперативность, = 1 отсутствие кооперативности.
Координаты Бьеррума
y
yKL
1logloglog
Ly
Ly
log;log
log;
LK
L
R
LRy
0
][
Число перегибов равно числу типов мест связывания.Абцисса точки перегиба равна логарифму константы диссоциации.
степень насыщения

RLLRLL
RLLRLL
RLLRLL
RLLRLL
RLRL
RLRL
2222
2121
1212
1111
22
11
1) Конкуренция за места связывания: а) неконкурентное (разные места связывания); б) конкурентное (одно место связывания); в) бесконкурентное (лиганд L2 связывается с L1R комплексом).
2) Изменение аффинности рецепторов: а) без изменения аффинности (L1R и R имеют одинаковую аффинность к
L2); б) с изменением аффинности.
3) Наличие взаимодействия между лигандами: а) без взаимодействия L1 и L2; б) с взаимодействием.

Феномен колебаний рецепторного связывания 1991 г.
RXXR
RLLR
XK
K1
- периодические (период порядка 10 минут)- апериодические
wtXBX sin0модулятор
[RL] (t)
Возможным источником колебаний может быть соотношение ГТФ / ГДФ, возникающее при гликолитических колебаниях.

Учет функции распределения клеток по количеству рецепторов на мембране
,NC,NCA
,yxC,yxCA
,yxCx,yCA
Nk
Nk
yxk
yxk
yxk
yxk
011
...
2211
11
1,11
,02
1,11
2,22
,1
1,12
x y
tyxСyxktAyxkdt
tdA
tyxСyxktyxСyxk
tyxСtAyxktyxСtAyxkdt
tyxdС
),,(),()(),()(
...
),1,1()1,1(),,(),(
),,()(),(),1,1()()1,1(),,(
21
22
11
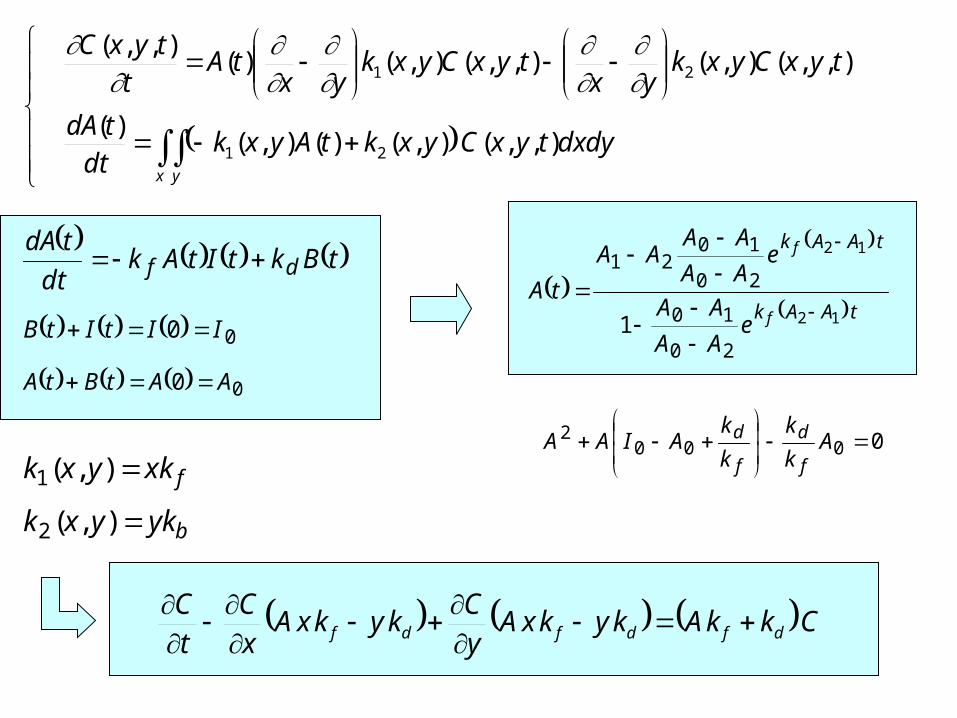
fxkyxk ),(1
x y
dxdytyxCyxktAyxkdt
tdA
tyxCyxkyx
tyxCyxkyx
tAt
tyxC
),,(),()(),()(
),,(),(),,(),()(),,(
21
21
bykyxk ),(2
00 IItItB
00 AAtBtA
tBktItAkdt
tdAdf
tAAk
tAAk
f
f
eAA
AA
eAA
AAAA
tA12
12
20
10
20
1021
1
00002
A
k
k
k
kAIAA
f
d
f
d
CkkAkykxAyC
kykxAxC
tC
dfdfdf
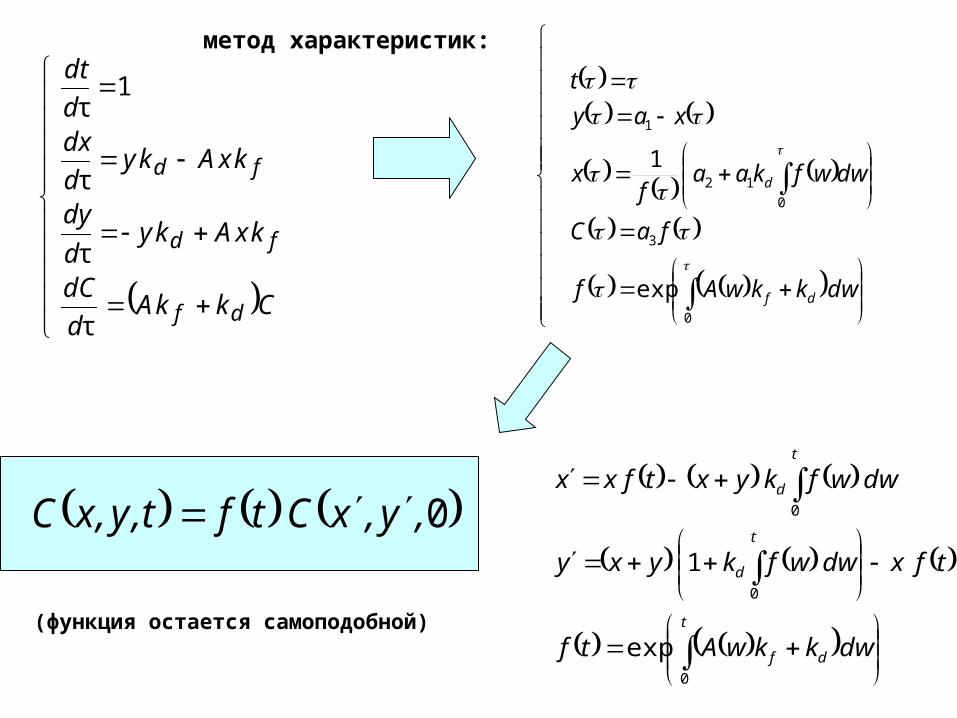
CkkAd
dC
kxAkyd
dy
kxAkyd
dxd
dt
df
fd
fd
τ
τ
τ
1τ
метод характеристик:
0
3
0
12
1
exp
1
dwkkwAf
faC
dwwfkaaf
x
xay
t
df
d
0,y,xCtft,y,xC
t
df
t
d
t
d
dwkkwAtf
tfxdwwfkyxy
dwwfkyxtfxx
0
0
0
exp
1
(функция остается самоподобной)
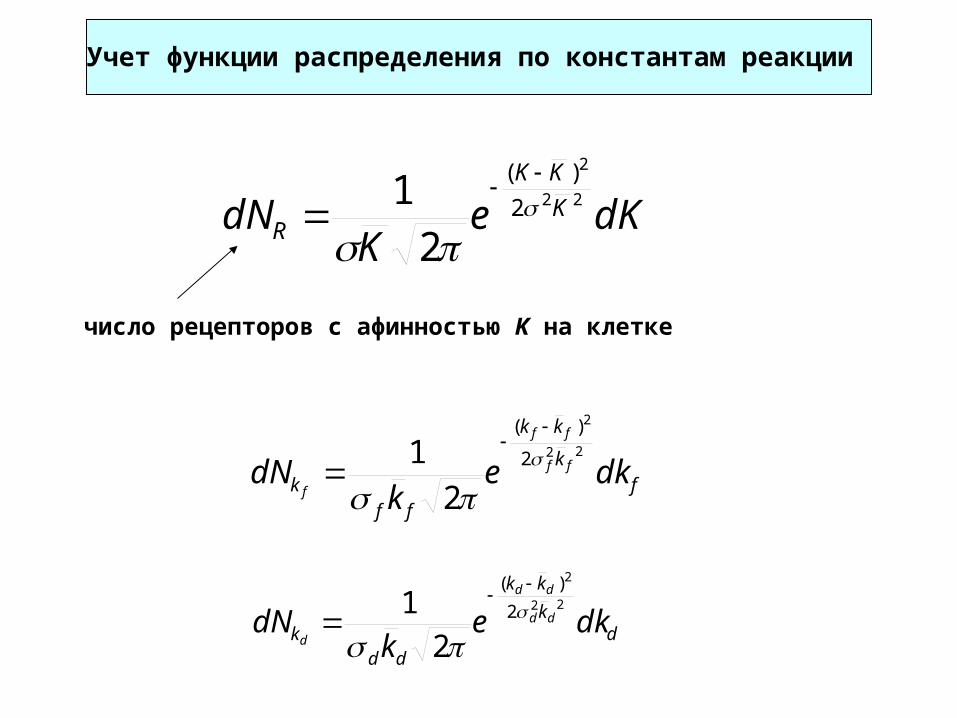
Учет функции распределения по константам реакции
dKeK
dN K
KK
R
22
2
2
)(
2
1
число рецепторов с афинностью K на клетке
fk
kk
ffk dke
kdN ff
ff
f
22
2
2
)(
2
1
dk
kk
ddk dke
kdN dd
dd
d
22
2
2
)(
2
1
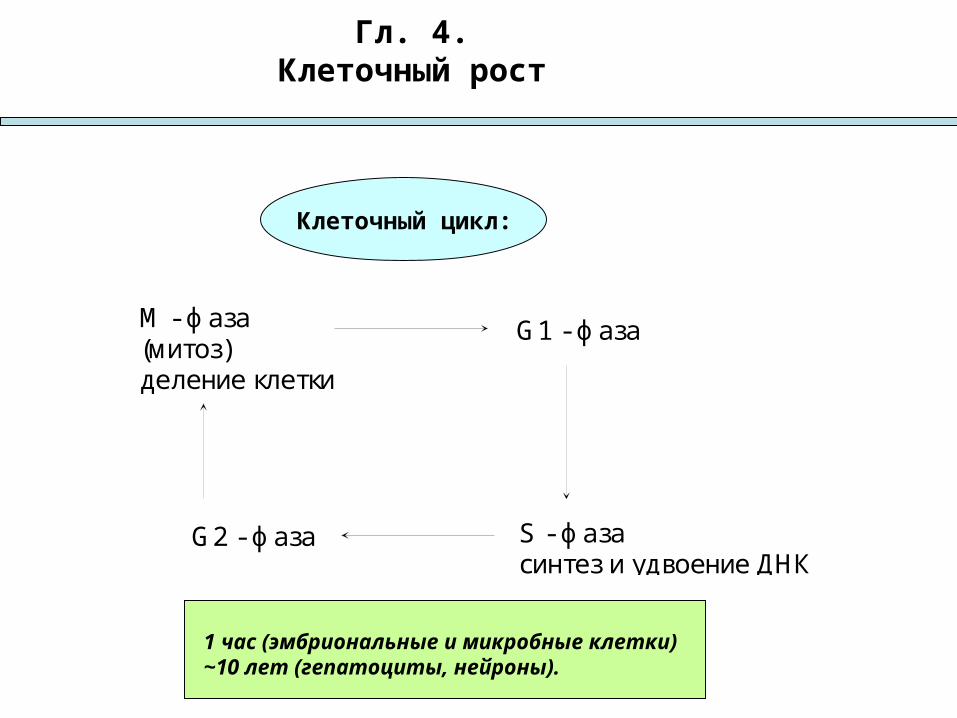
Гл. 4.Клеточный рост
М - фаза(митоз)деление клетки
G1 - фаза
S - фазасинтез и удвоение ДНК
G2 - фаза
Клеточный цикл:
1 час (эмбриональные и микробные клетки)~10 лет (гепатоциты, нейроны).

Культивирование клеток -> изучение клеточного цикла in vitro(культуры, линии клеток).
Фазы роста: 1) индукционный (лаг-фаза) – адаптация клеток к среде, перестройка их метаболизма;2) экспоненциальный рост (много митозов);3) линейный рост (мало митозов);4) замедление роста;5) стационарная фаза;6) отмирание культуры.
Причины замедления роста – истощение субстрата, накопление токсических продуктов и др.
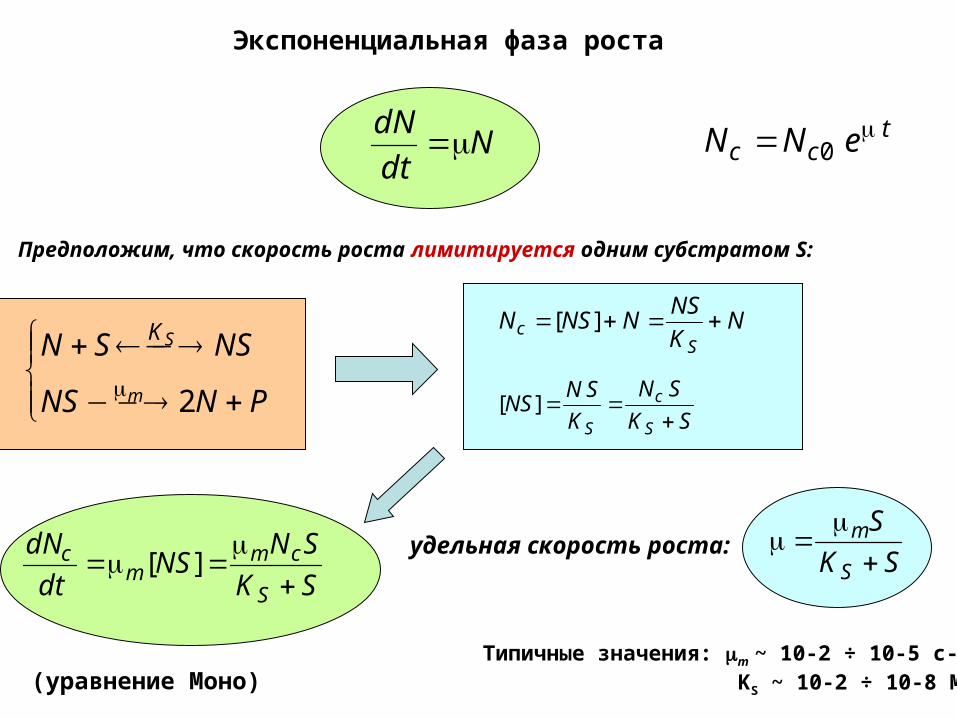
Экспоненциальная фаза роста
Ndt
dN
PNNS
NSSN
m
SK
2
Предположим, что скорость роста лимитируется одним субстратом S:
NK
NSNNSN
Sc ][
SK
SN
K
SNNS
S
c
S ][
SK
SNNS
dt
dN
S
cmm
с
][ SK
S
S
m
удельная скорость роста:
(уравнение Моно) Типичные значения: m ~ 10-2 ÷ 10-5 c-1,
KS ~ 10-2 ÷ 10-8 M.
tсс eNN 0

Многосубстратные процессы
пример (M. thermoautotrophicum): OHCHCOH 2422 24
возможные механизмы:
PNNSS
NSSSNS
NSNS
митоз
K
K
221
2121
11
2
1
21
21
2
21SS
KK
S
Km
тройной комплекс
PNXS
XSXS
PXNS
NSNS
митоз
K
K
22
22
1
11
2
1
2
2
1
11S
K
S
Km
пинг-понг
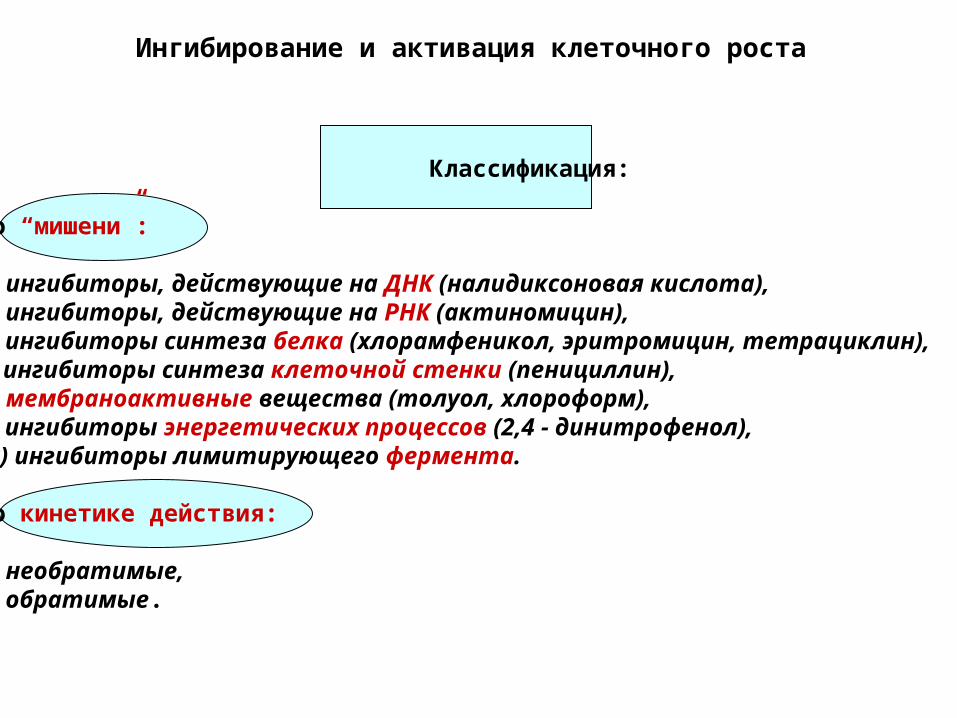
Ингибирование и активация клеточного роста
Классификация:
По “мишени”:
а) ингибиторы, действующие на ДНК (налидиксоновая кислота),б) ингибиторы, действующие на РНК (актиномицин),в) ингибиторы синтеза белка (хлорамфеникол, эритромицин, тетрациклин),г) ингибиторы синтеза клеточной стенки (пенициллин),д) мембраноактивные вещества (толуол, хлороформ),е) ингибиторы энергетических процессов (2,4 - динитрофенол),ж) ингибиторы лимитирующего фермента.
По кинетике действия:
а) необратимые,б) обратимые.
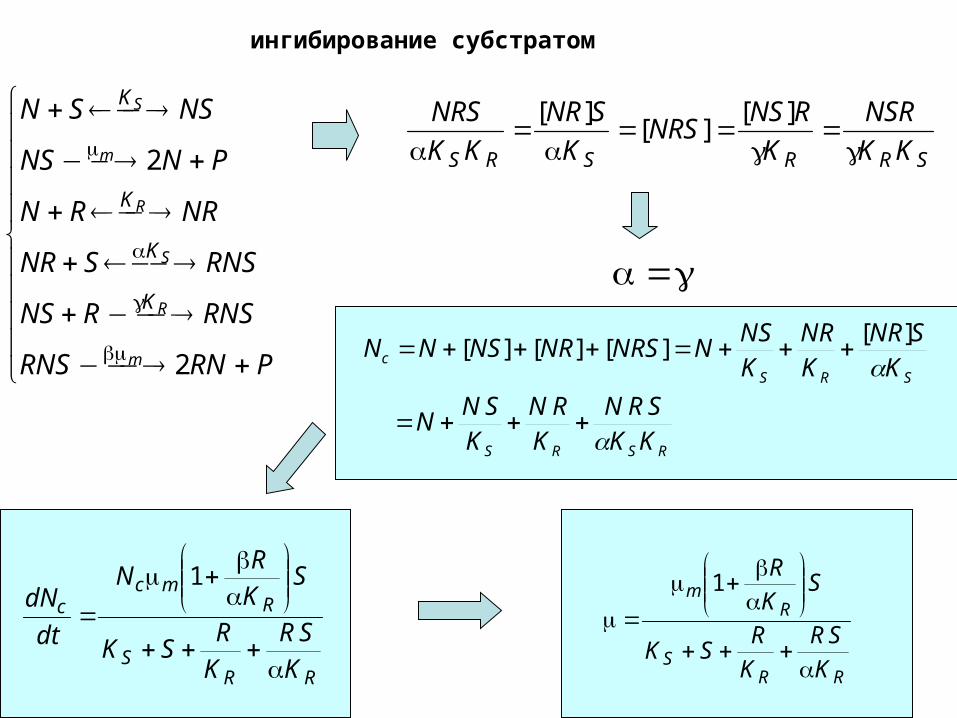
ингибирование субстратом
PRNRNS
RNSRNS
RNSSNR
NRRN
PNNS
NSSN
m
R
S
R
m
S
K
K
K
K
2
2 SRRSRS KK
NSR
K
RNSNRS
K
SNR
KK
NRS
][
][][
RSRS
SRSc
KKSRN
KRN
KSN
N
KSNR
KNR
KNS
NNRSNRNSNN
][][][][
RRS
Rmc
c
K
SR
K
RSK
SK
RN
dt
dN
1
RRS
Rm
K
SR
K
RSK
SK
R
1

Влияние pH
- на субстрат (аминокислоты);
- на ферменты клетки.
inexin HH
dt
dH
Кинетика переноса ионов водорода через мембрану клетки:
inex HHВ условиях равновесия:
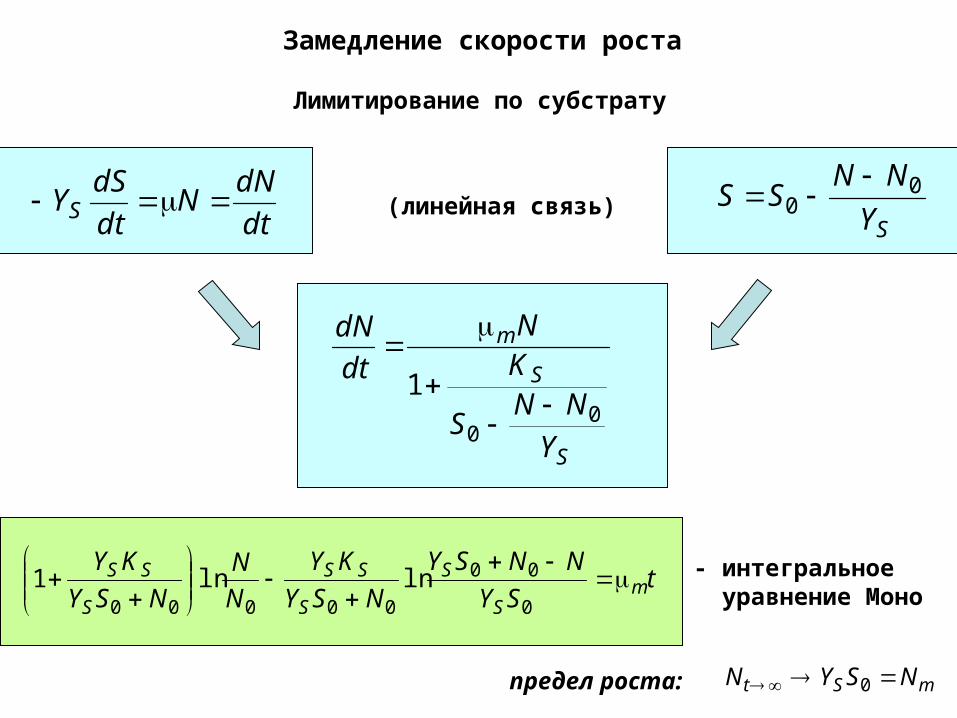
Замедление скорости роста
Лимитирование по субстрату
dt
dNN
dt
dSYS (линейная связь)
SY
NNSS 0
0
S
S
m
Y
NNS
KN
dt
dN
00
1
tSY
NNSY
NSY
KY
N
N
NSY
KYm
S
S
S
SS
S
SS
0
00
00000lnln1
mSt NSYN 0предел роста:
- интегральное уравнение Моно

Ингибирование избытком субстрата
NX
XSSX
XNS
m
i
S
K
K
2
SiSiSc KK
SNS
K
SNN
K
XS
K
SNNXSXNN
SiS
c
KK
S
K
S
NN
2
1
iS
cm
S
mm
c
K
SSK
SN
K
SNX
dt
dN2

Ингибирование субстратом в условиях истощения по субстрату
S
iS
m
Y
NNSS
K
SSK
SN
dt
dN
00
2
tNNYKN
NN
S
K
N
N
K
S
S
Km
Sim
S
i
S
0
0
0
0
0
11lnln1

Ингибирование продуктами
NPPN
PNX
XNS
i
m
S
K
K
2iS
c K
NP
K
SNNNPXNN ][
dt
dN
dt
dPY c
P 00 ccP NNPPY
P
cc
i
SS
cm
Smm
c
Y
NNP
K
KSK
SN
K
SNX
dt
dN
00
t
SK
SNN
SKKY
K
N
N
SKKY
KNPY
S
mcc
SiP
S
c
c
SiP
ScP
0
0
00 ln1
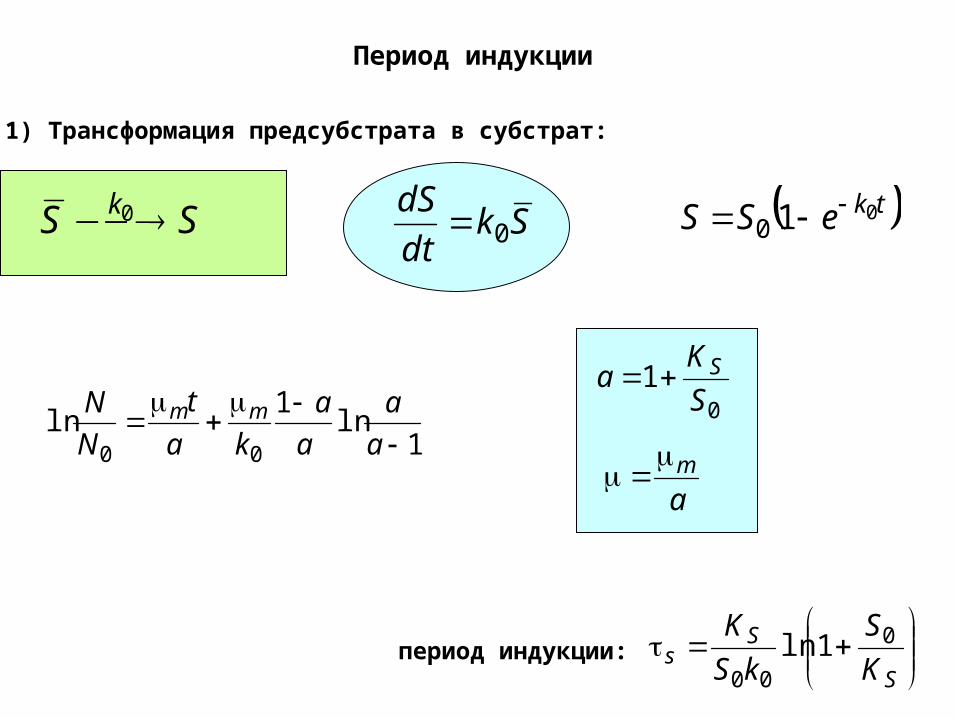
Период индукции
1) Трансформация предсубстрата в субстрат:
SS k 0 Skdt
dS0 tkeSS 010
1ln
1ln
00
a
a
a
a
ka
t
N
N mm 01
S
Ka S
am
S
Ss K
S
kS
K 0
001lnпериод индукции:
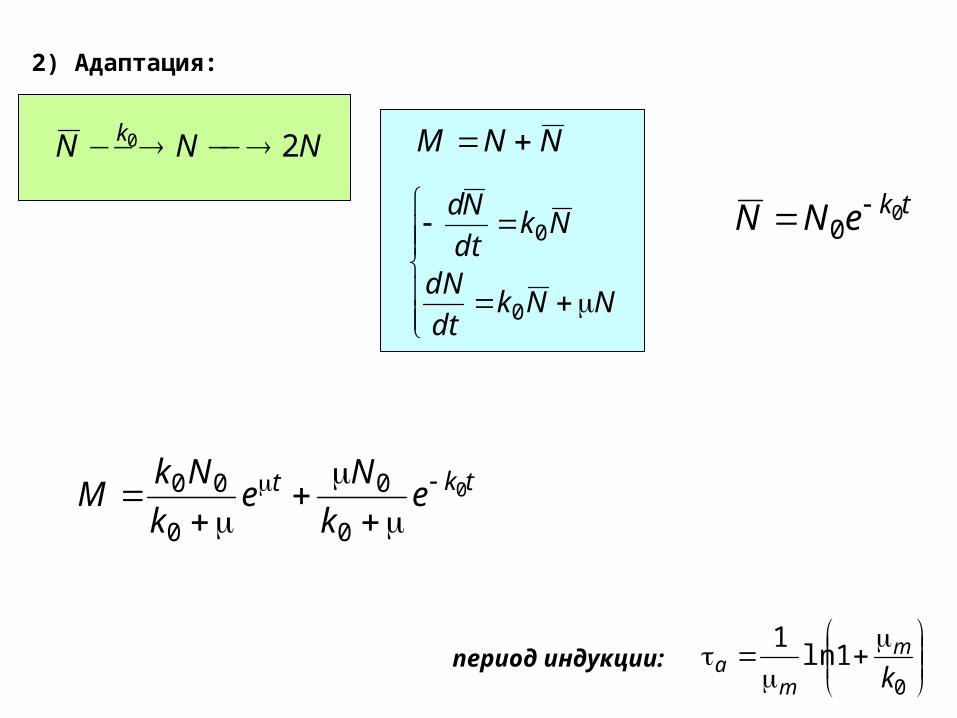
2) Адаптация:
NNN k 20 NNM
NNkdt
dN
Nkdt
Nd
0
0tkeNN 0
0
tkt ek
Ne
k
NkM 0
0
0
0
00
0
1ln1
km
maпериод индукции:

3) Расход ингибитора роста:
NINI k 0 Ikdt
dI0 tkeII 0
0
tk
ie
K
IN
dt
dN001
0
0
kK
I
ii период индукции:
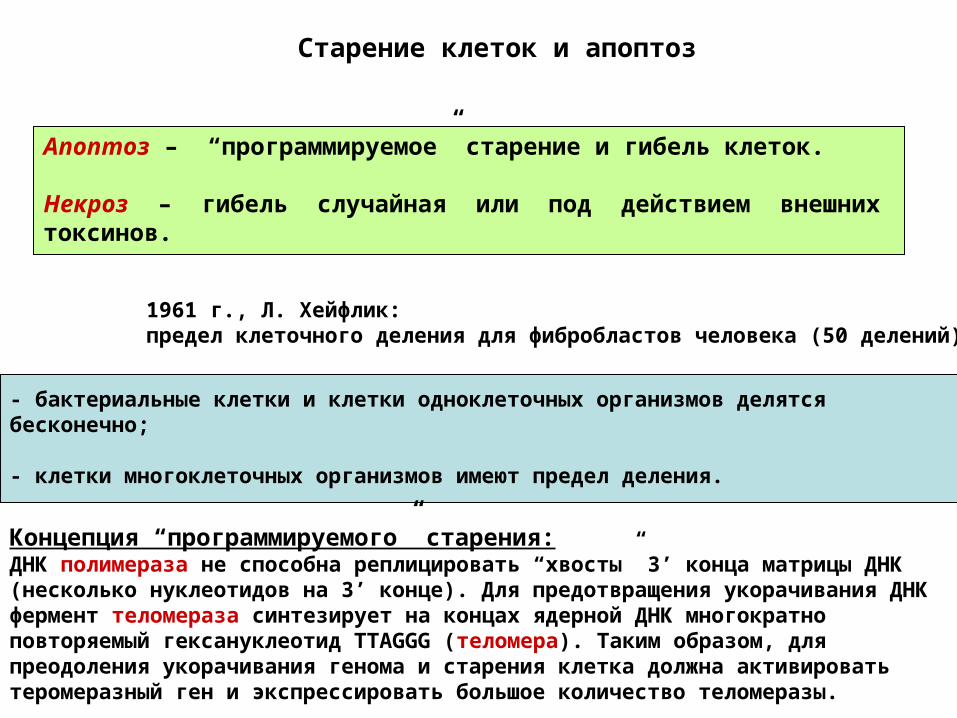
Старение клеток и апоптоз
Апоптоз – “программируемое” старение и гибель клеток.
Некроз – гибель случайная или под действием внешних токсинов.
1961 г., Л. Хейфлик: предел клеточного деления для фибробластов человека (50 делений).
- бактериальные клетки и клетки одноклеточных организмов делятся бесконечно;
- клетки многоклеточных организмов имеют предел деления.
Концепция “программируемого” старения:ДНК полимераза не способна реплицировать “хвосты” 3’ конца матрицы ДНК (несколько нуклеотидов на 3’ конце). Для предотвращения укорачивания ДНК фермент теломераза синтезирует на концах ядерной ДНК многократно повторяемый гексануклеотид TTAGGG (теломера). Таким образом, для преодоления укорачивания генома и старения клетка должна активировать теромеразный ген и экспрессировать большое количество теломеразы.

Удельная скорость клеточного роста в экспоненциальной фазе
PS
ДНКSS RE
Предположения:
- скорость синтеза ДНК пропорциональна концентрации S’ и характеризуется константой скорости R;
- скорость биосинтеза лимитирующего фермента и белков репликационного комплекса намного выше скорости синтеза ДНК, так что можно считать E=const.
SSK
ESk
dt
dS
SДНКdt
d
RS
кат
R
0
dt
SdSK
ESkS
S
кат
R
1
В квазистационаре:
tSK
ESktДНК
S
кат
R
R
)]([
RS
кат
R
R
SK
ESkB
ESk
BSK
катR
SRR
R
2ln
время репликации
базовое количество ДНК
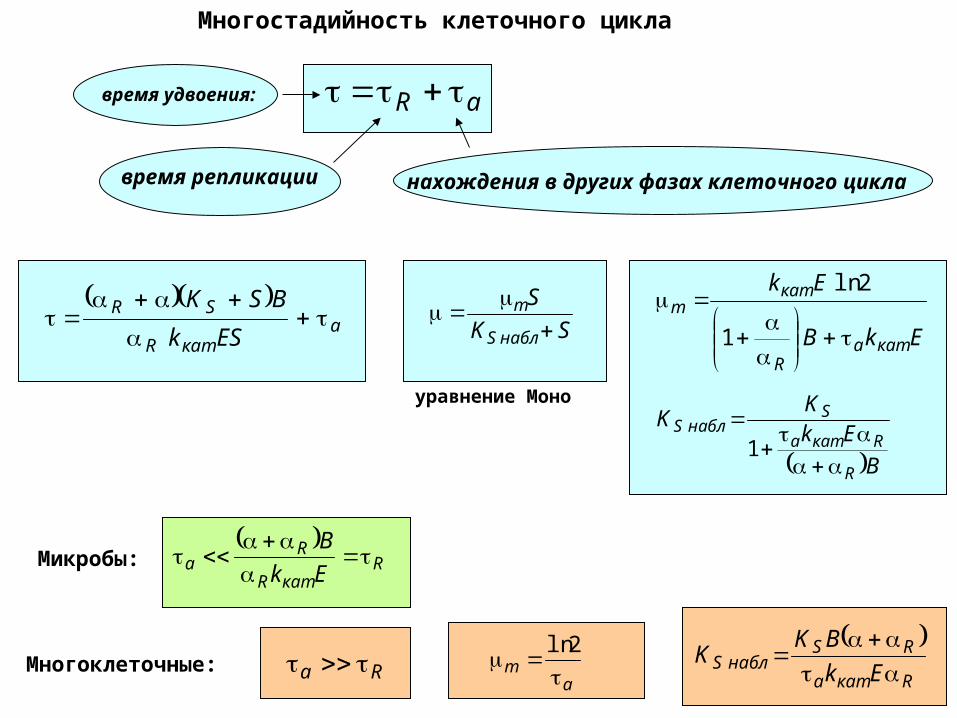
Многостадийность клеточного цикла
aR
время репликации нахождения в других фазах клеточного цикла
время удвоения:
a
катR
SR
ESk
BSK
SK
S
наблS
m
EkB
Ek
катaR
катm
1
2ln
BEk
KK
R
Rкатa
SнаблS
1
уравнение Моно
R
катR
Ra Ek
B
Микробы:
Многоклеточные: Ra a
m
2ln Rкатa
RSнаблS Ek
BKK
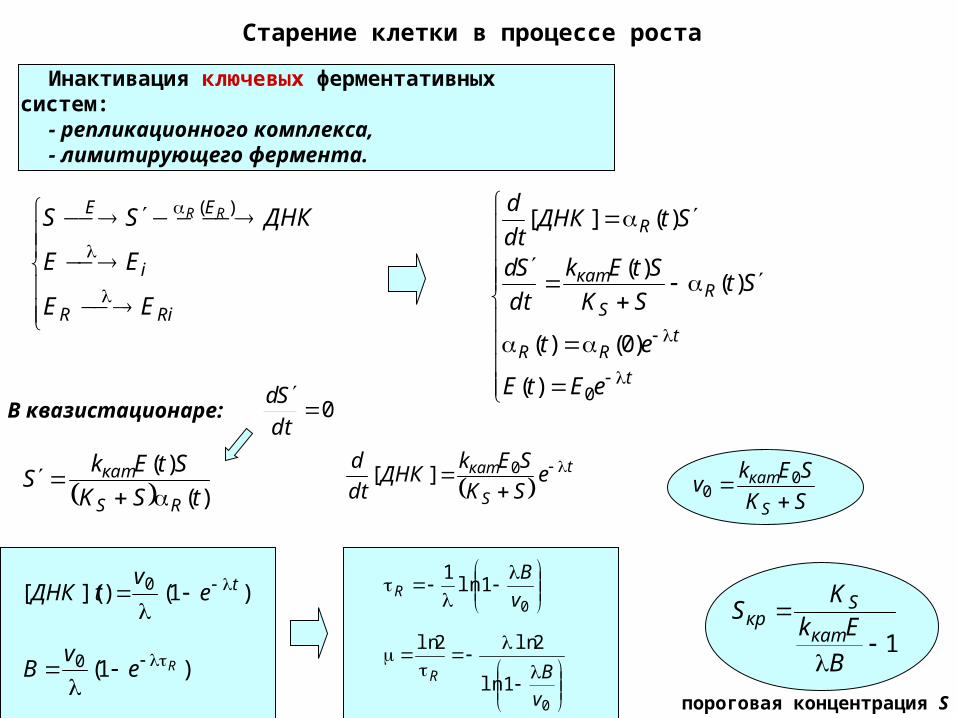
Старение клетки в процессе роста
Инактивация ключевых ферментативных систем:
- репликационного комплекса,- лимитирующего фермента.
RiR
i
EE
EE
EE
ДНКSS RR )(
t
tRR
RS
кат
R
eEtE
et
StSK
StEk
dt
Sd
StДНКdt
d
0)(
)0()(
)()(
)(][
)(
)(
tSK
StEkS
RS
кат
В квазистационаре: 0
dt
Sd
t
S
кат eSK
SEkДНК
dt
d
0][
)1()]([ 0 tev
tДНК
SK
SEkv
S
кат
0
0
)1(0 Rev
B
01ln
1
v
BR
01ln
2ln2ln
v
BR
1
B
EkK
Sкат
Sкр
пороговая концентрация S

Кинетические модели апоптоза
1) Прогрессирующая некомпетентность:
ai
aaaa
Ndt
dN
NNtdt
dN)(
способные к делению клетки
ia NNN
aaNdt
dN
0dt
dNaВозможна ситуация: (максимальное количество некомпетентных клеток)
S
mS
S
mm
Y
NtNSK
Y
NtNS
00
00
)(
)(
Если a дается уравнением Моно, то:
m
SSm
KSYNtN 00)(
tm - время достижения максимума по Na
m
Sm
KtS )(

1) начальный период:
ama ,
tNN amaa ,0 exp
tNN am
am
ai
,,
0 exp
tNN am
am
aam
,
,
0, exp
tNN maa exp,
2) режим истощения субстрата (на больших временах):
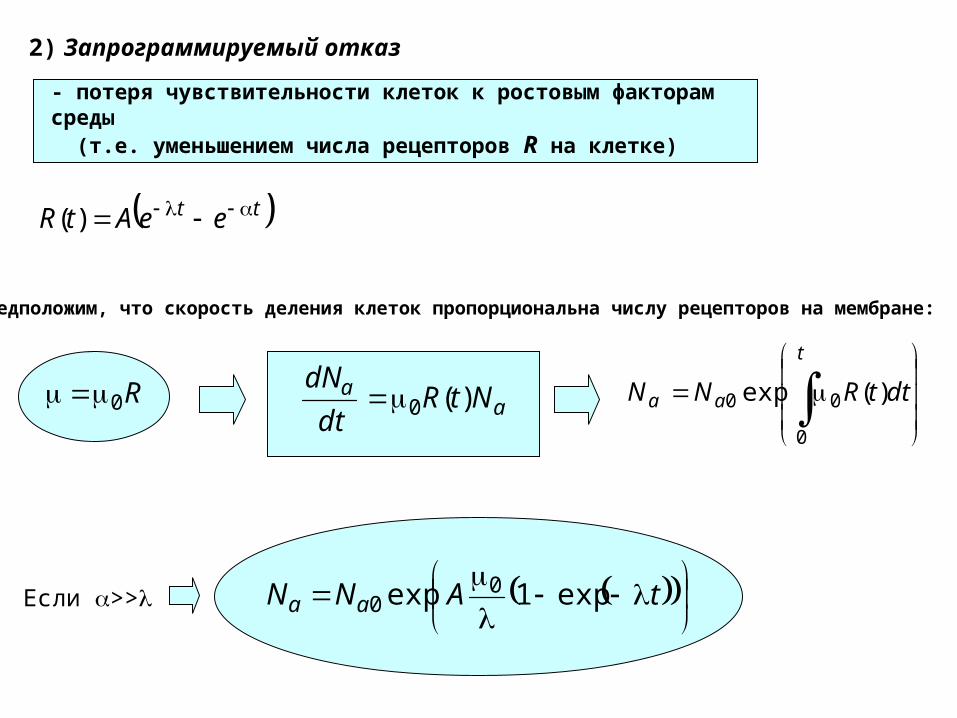
2) Запрограммируемый отказ
- потеря чувствительности клеток к ростовым факторам среды (т.е. уменьшением числа рецепторов R на клетке)
tt eeAtR )(
R0
Предположим, что скорость деления клеток пропорциональна числу рецепторов на мембране:
aa NtR
dt
dN)(0
t
aa dttRNN
0
00 )(exp
tANN aa exp1exp 00Если >>

Модель “хищник-жертва”
2212
2111
NNNdt
dN
NNaNdt
dN
жертва хищник
Существует стационарное состояние:
0
0
2
1
dt
dNdt
dN
aN
N
ст
ст
2
1
Фазовый портрет (связь между N1 и N2):
1
11
стN
Nn
2
22
стN
Nn
212
121
1
1
nndt
dn
nnadt
dn 12
21
1
2
1
1
nna
nn
dn
dn
1
1
12
2
2 11dn
an
ndn
n
n
consten
en
a
nn
21
21
фазовый портретколебательной траектории
Средняя численность популяции равна стационарной численности
T
dtnan
dn
0
21
1 1 201 )(ln0 naTaTtnT
12 n
11 n

Ассоциации микроорганизмов
симбиоза до антагонизма
Пример: ассоциация двух микроорганизмов M1 и M2
PSS MM 2121
dt
dM
Ydt
dP
dt
dM
Ydt
dM
Ydt
dS
dt
dM
Ydt
dS
SK
MS
dt
dM
SK
MS
dt
dM
P
PS
S
2
122
11
22
2222
11
1111
2
12
1
1
11
1
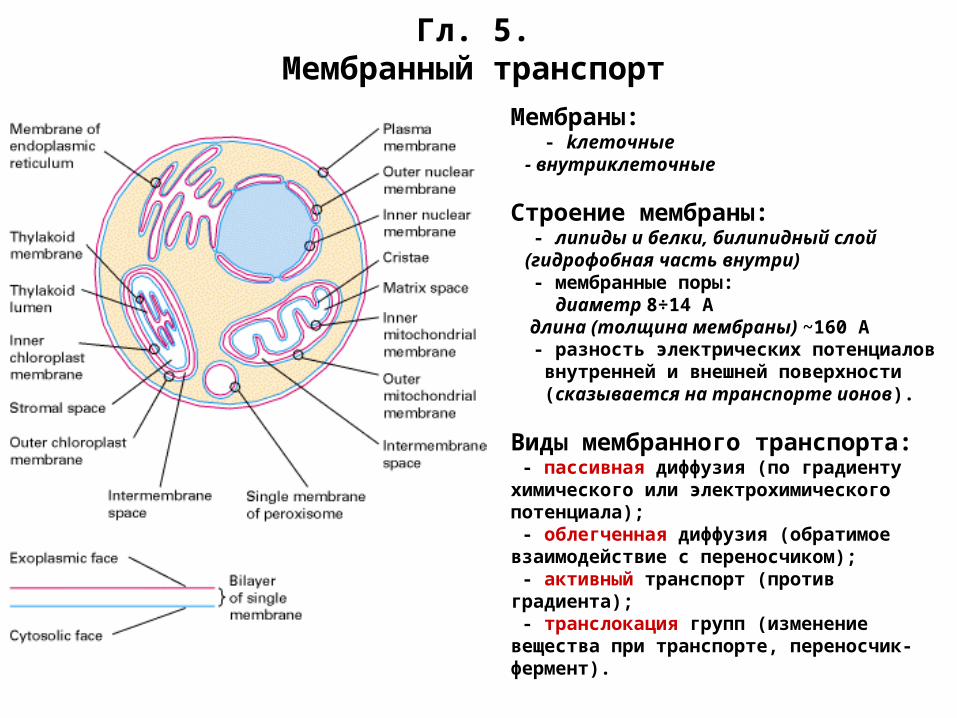
Гл. 5.Мембранный транспорт
Мембраны: - kлеточные - внутриклеточные
Строение мембраны: - липиды и белки, билипидный слой (гидрофобная часть внутри) - мембранные поры: диаметр 8÷14 А длина (толщина мембраны) ~160 А - разность электрических потенциалов внутренней и внешней поверхности (сказывается на транспорте ионов).
Виды мембранного транспорта: - пассивная диффузия (по градиенту химического или электрохимического потенциала); - облегченная диффузия (обратимое взаимодействие с переносчиком); - активный транспорт (против градиента); - транслокация групп (изменение вещества при транспорте, переносчик-фермент).
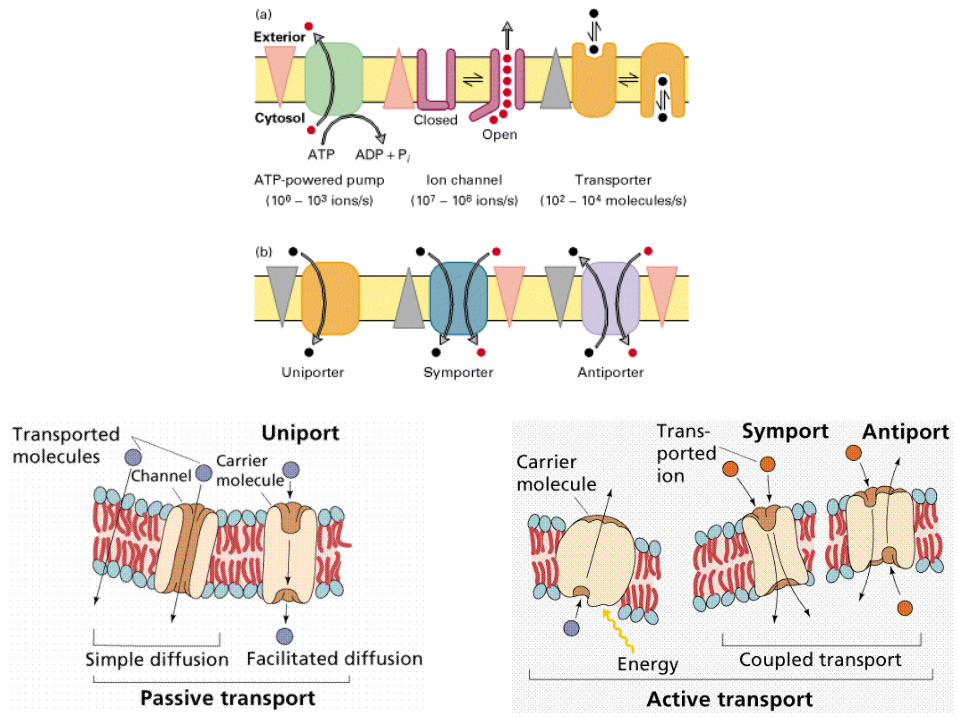

Пассивный транспорт
0
ii Jdiv
t
С
iii
iii CDRT
FzCDJ
число Фарадея
заряд (целое число, в единицах электронов) i-го иона
потенциал электрического поля в мембране
i
iiCzF
dx
dDC
RT
Fz
dx
dCDJ
Одномерный случай:
xdx
d
в линейном
приближении:
xDC
RT
Fz
dx
dCDJ
e
i
C
C
x
dC
xDC
RT
FzJ
Ddxx
0
RT
zF
RT
zF
ie
exRT
eCCFzD
J
1
x
uRT
x
Dp
Коэффициент проницаемости:
подвижность
RT
zF
ie eCC
ie CCpJ
i
e
e
i
Cl
Cl
H
H
Если молекула не заряжена:
В равновесии (J = 0): (уравнение Нерста)
Доннаново равновесие
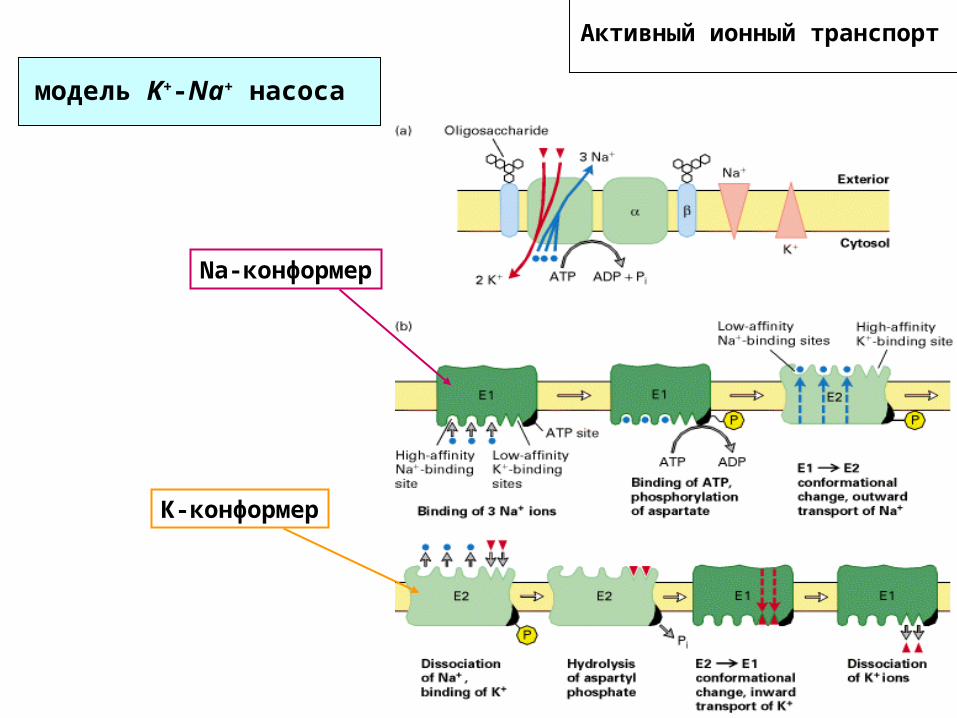
Активный ионный транспорт
модель K+-Na+ насоса
Na-конформер
К-конформер
)(
)(
)(
)(
.)..(
.)..(
3
3
3
3
2
2
1
1
KтранспортпассивныйKK
NaтранспортпассивныйNaNa
белкамолекулыпереворотФNaEФNaE
белкамолекулыпереворотKEKE
мембрповвнешннаNaФKEФNaEK
мембрповвнутрнаKАДФФNaEАТФKENa
eJ
i
eJ
i
ek
k
i
ek
k
i
eek
k
ee
iik
k
ii
K
Na
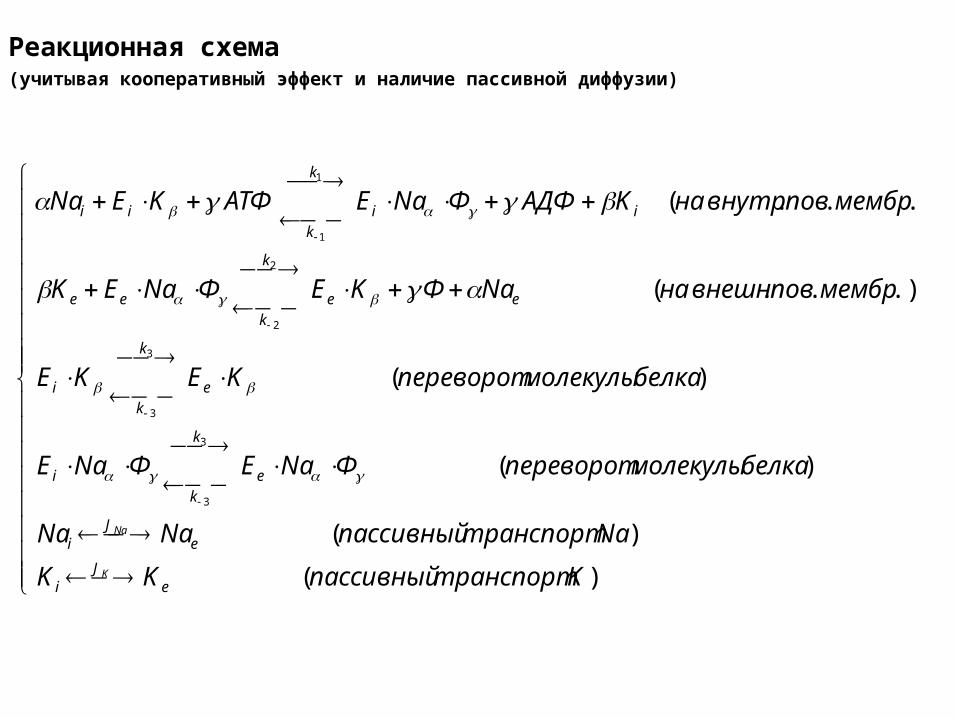
Реакционная схема(учитывая кооперативный эффект и наличие пассивной диффузии)
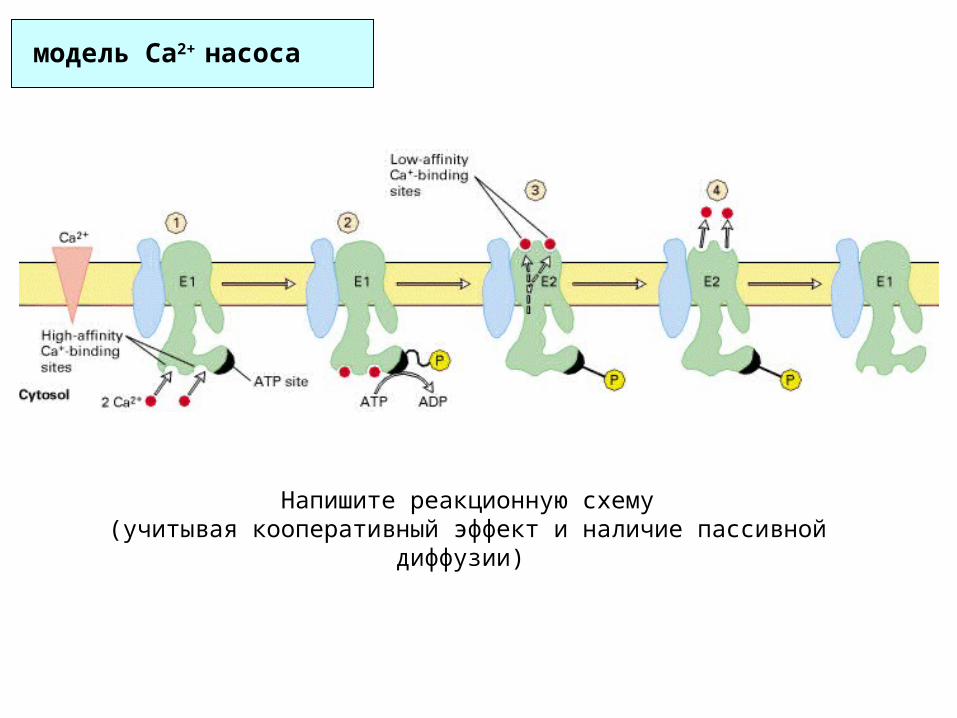
модель Ca2+ насоса
Напишите реакционную схему(учитывая кооперативный эффект и наличие пассивной диффузии)

Высокая селективность ионных каналов к определенному типу ионов
93.0
Na
Li
p
p16.04
Na
NH
p
p09.0
Na
K
p
p
3.2
K
Tl
p
p13.04
K
NH
p
p01.0
K
Na
p
p
натриевый канал:
калиевый канал:
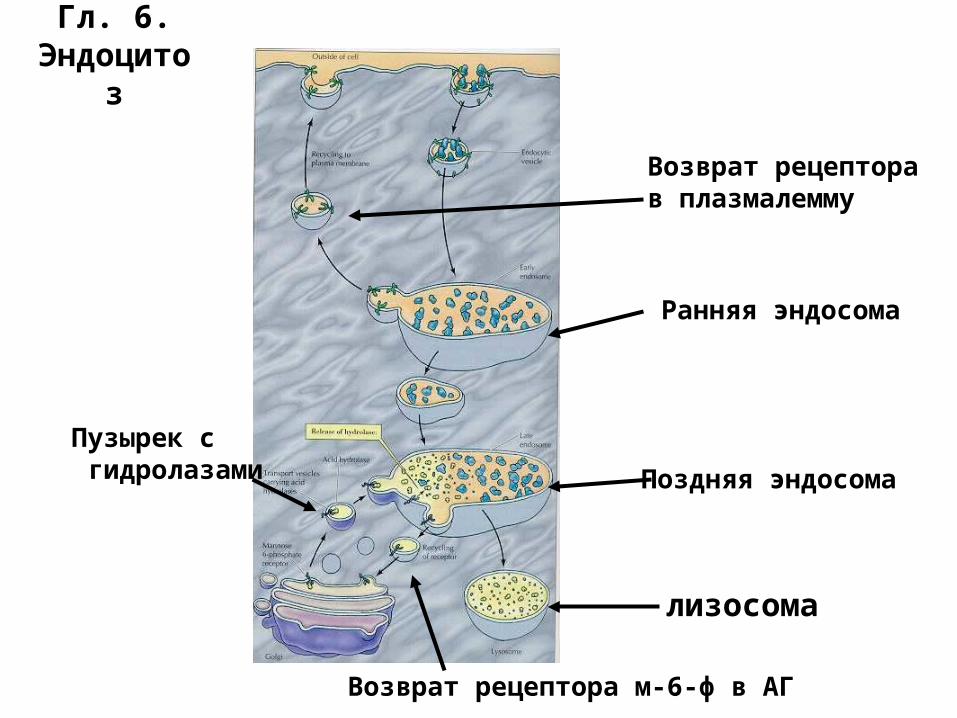
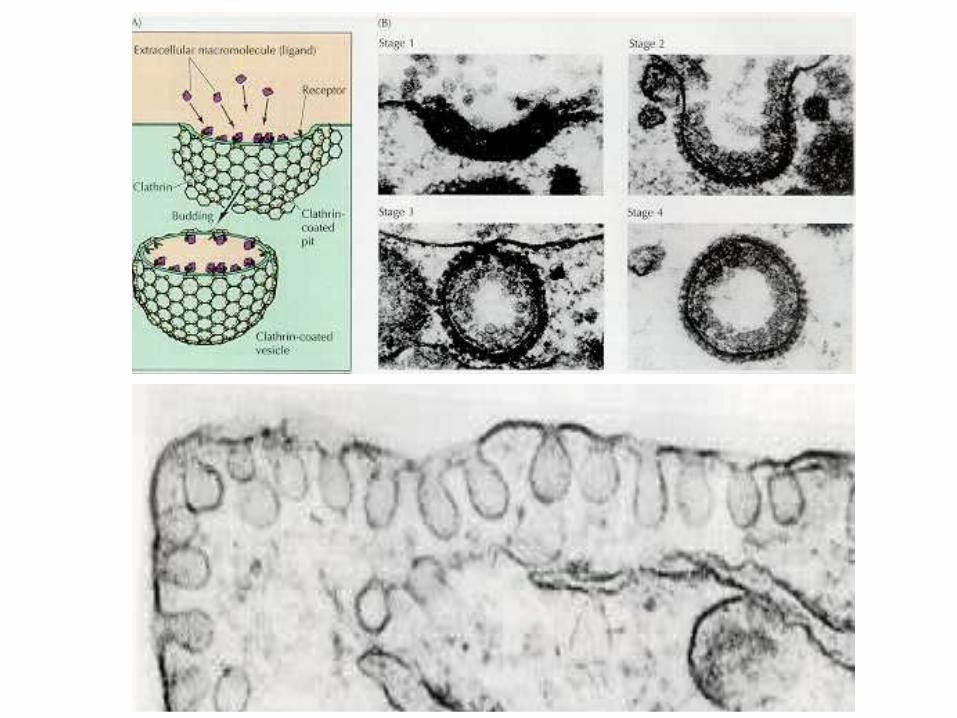
Гл. 6.Эндоцитоз
Ранняя эндосома
Пузырек с гидролазами Поздняя эндосома
лизосома
Возврат рецепторав плазмалемму
Возврат рецептора м-6-ф в АГ
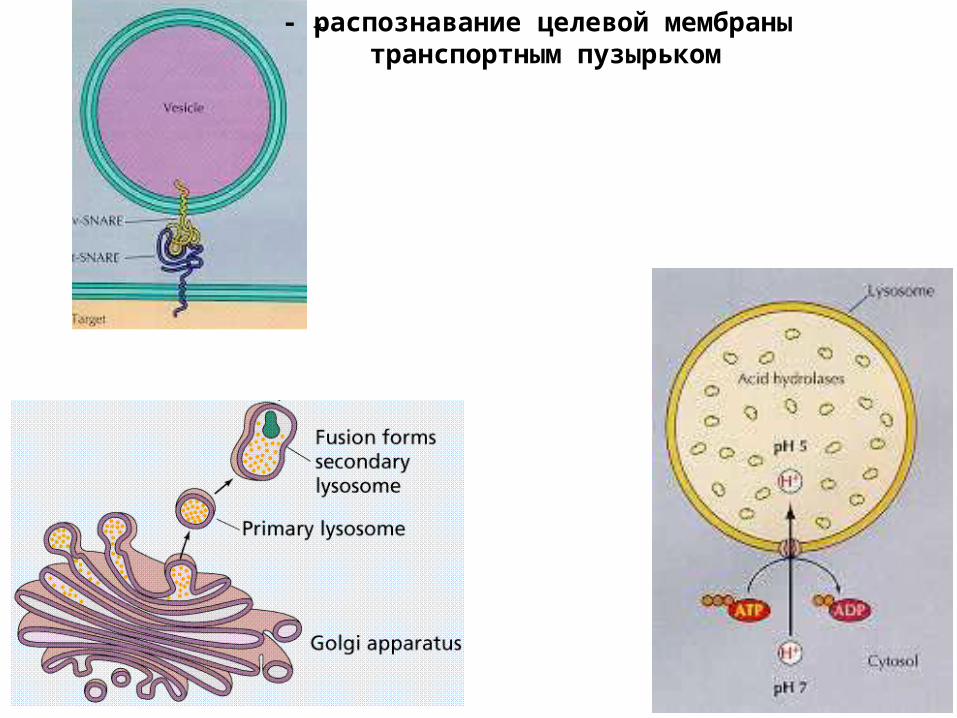
- распознавание целевой мембраны транспортным пузырьком
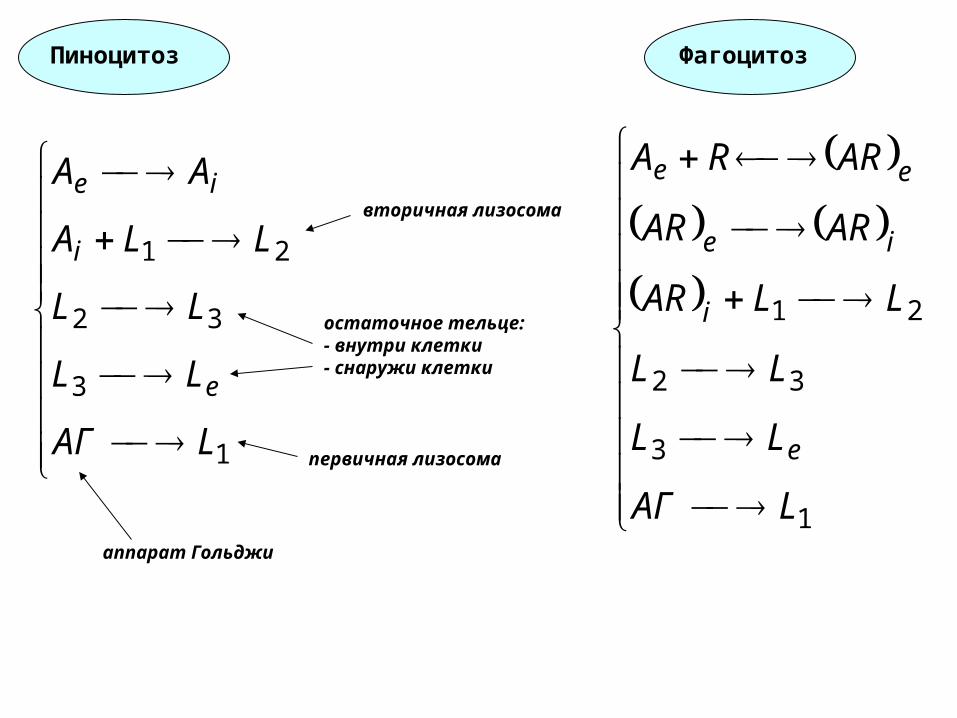
1
3
32
21
LАГ
LL
LL
LLA
AA
e
i
ie
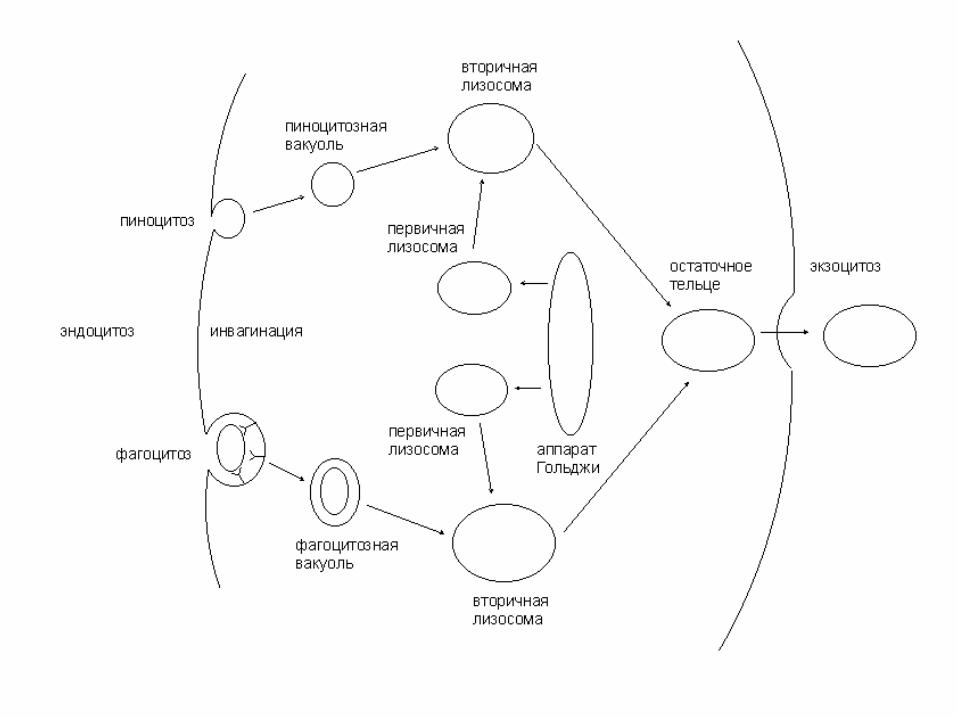
Пиноцитоз Фагоцитоз
1
3
32
21
LАГ
LL
LL
LLAR
ARAR
ARRA
e
i
ie
ee
первичная лизосома
вторичная лизосома
остаточное тельце:- внутри клетки- снаружи клетки
аппарат Гольджи

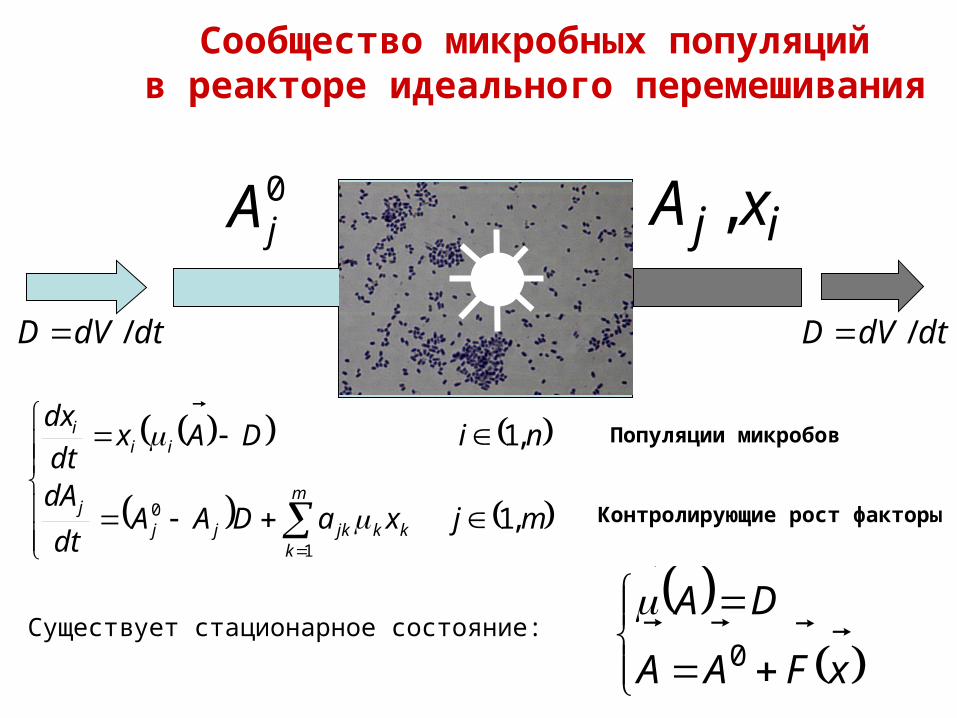
Сообщество микробных популяцийв реакторе идеального перемешивания
m
kkkjkjj
j
iii
mjxaDAAdt
dA
niDAxdt
dx
1
0 ,1
,1
Популяции микробов
Контролирующие рост факторы
0jA ij xA ,
Существует стационарное состояние:
xFAA
DA
0
dtdVD / dtdVD /
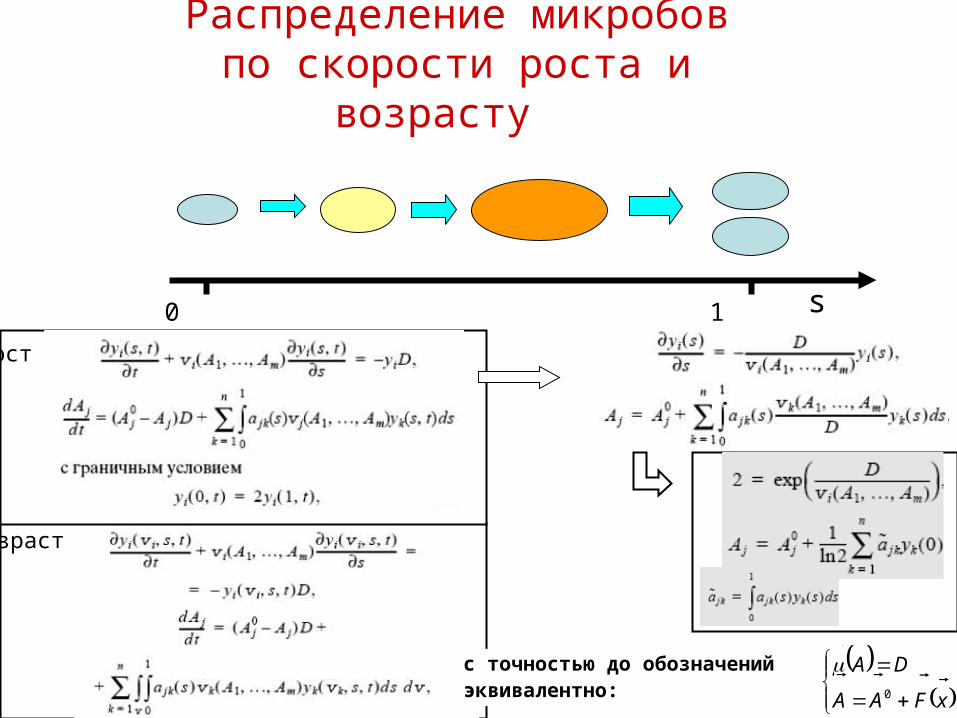
Распределение микробовпо скорости роста и возрасту
s0 1
с точностью до обозначенийэквивалентно:
рост
возраст
xFAA
DA
0
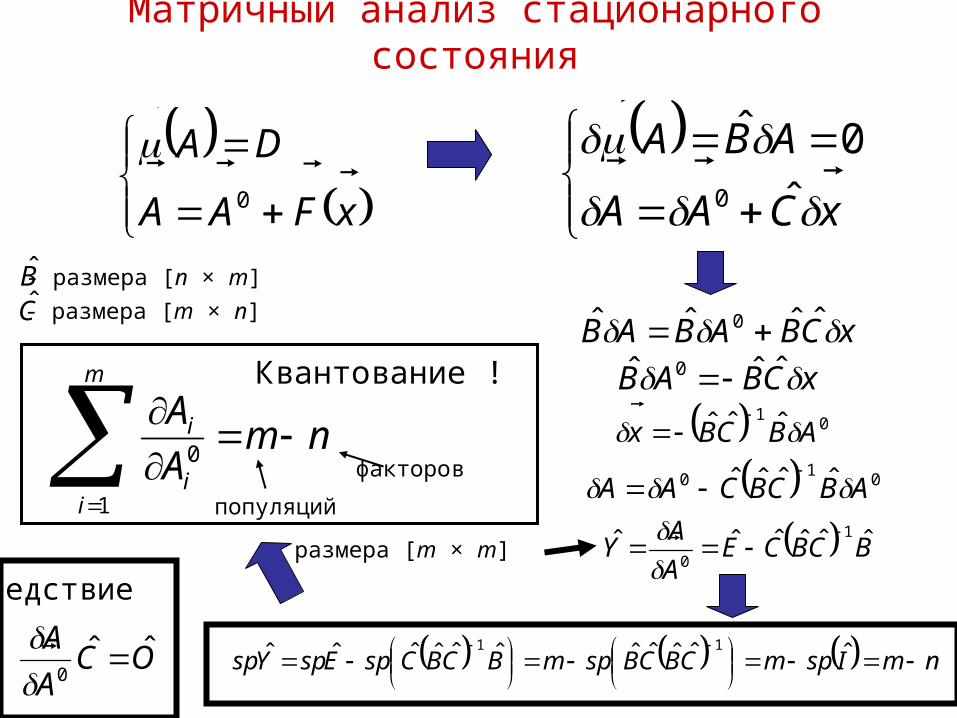
Матричный анализ стационарного состояния
nmA
Am
ii
i
1
0
OCA
A ˆˆ0
xFAA
DA
0
xCAA
ABA
ˆ
0ˆ
0
xCBABAB ˆˆˆˆ 0
BC
- размера [n × m]
- размера [m × n]
xCBAB ˆˆˆ 0
01 ˆˆˆ ABCBx
01
0 ˆˆˆˆ ABCBCAA
BCBCEA
AY ˆˆˆˆˆˆ 1
0
размера [m × m]
nmIspmCBCBspmBCBCspEspYsp
ˆˆˆˆˆˆˆˆˆˆˆ 11
Квантование !
следствие
популяций
факторов
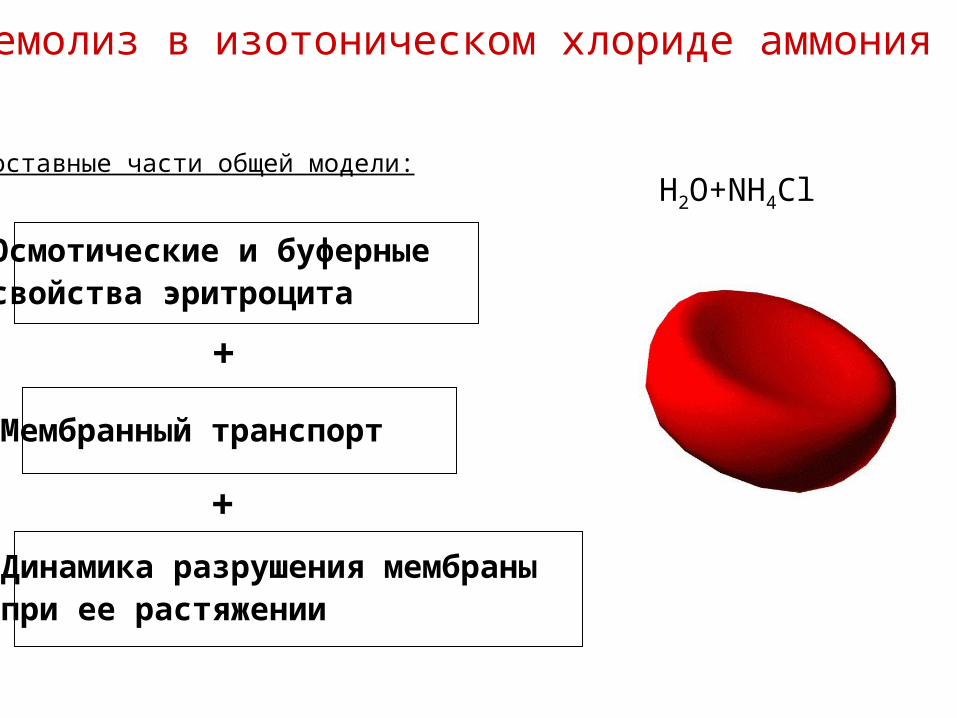
Гемолиз в изотоническом хлориде аммония
H2O+NH4Cl
Осмотические и буферныесвойства эритроцита
Мембранный транспорт
Динамика разрушения мембраныпри ее растяжении
Составные части общей модели:
+
+
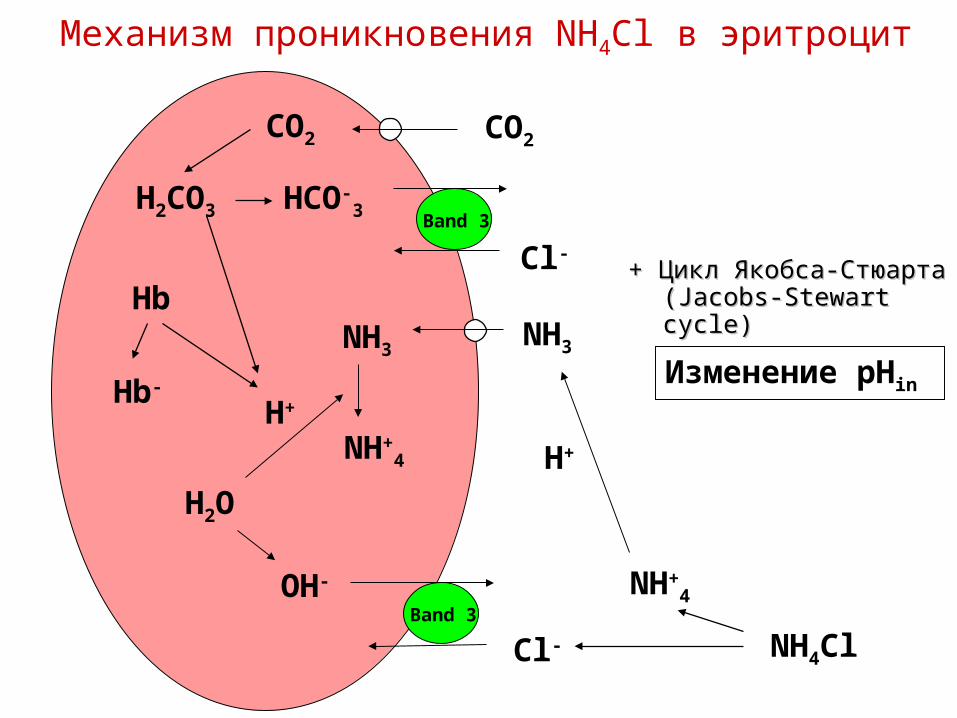
Механизм проникновения NH4Cl в эритроцит
NH3
H+
NH+4
Cl-
NH3
NH+4
H+
OH-
H2O
Band 3
NH4Cl
Hb
Hb-
Band 3HCO-
3
Cl-
CO2CO2
H2CO3
Изменение pHin
+ Цикл Якобса-Стюарта + Цикл Якобса-Стюарта ((Jacobs-Stewart cycleJacobs-Stewart cycle))
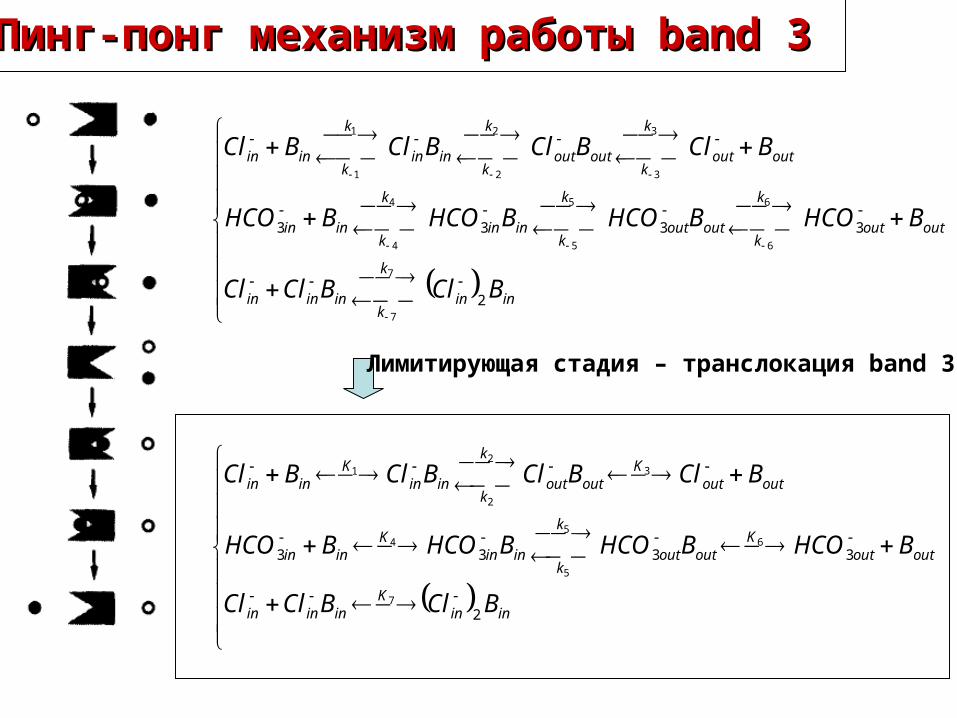
inink
k
ininin
outoutk
k
outoutk
k
inink
k
inin
outoutk
k
outoutk
k
inink
k
inin
BClBClCl
BHCOBHCOBHCOBHCO
BClBClBClBCl
2
3333
7
7
6
6
5
5
4
4
3
3
2
2
1
1
Пинг-понг механизм работы Пинг-понг механизм работы band 3band 3
ininK
ininin
outoutK
outoutk
k
ininK
inin
outoutK
outoutk
k
ininK
inin
BClBClCl
BHCOBHCOBHCOBHCO
BClBClBClBCl
2
3333
7
6
5
54
3
2
21
Лимитирующая стадия – транслокация band 3

Три стадии гемолиза
0303
0
1 0.00110.00719 0.0238084.1 wwwww
w HbNaHCOHbNaHCOV
V
1. Быстрое увеличение (на ~20%) начального объема (< 0.2 сек) - релаксация клетки в новой среде:
2. Медленное дальнейшее увеличение объема до сферы (> 10 сек):
31
2141 /
/GVV
GGGVV
dt
d
wwww
aww NVHbNaHCO
BNaHCOG
103
183
4 ][549.0][693.0162
10][
0031 )]([0.00261][0.09077][0.063471883.5 ww inClHbNaHCOG
0032 )]([0.0032][0.01643][0.0041155.64 ww inClHbNaHCOG
0033 )]([0.00097][0.00805][0.013249641.0 ww inClHbNaHCOG
3. Растяжение сферической клетки со спонтанным разрывом (> 10 сек):
tn
tndt
d /exp/ 2/1
0 0
0
S
SSKS
напряженность мембранывремя жизни мембраныконцентрация клеток
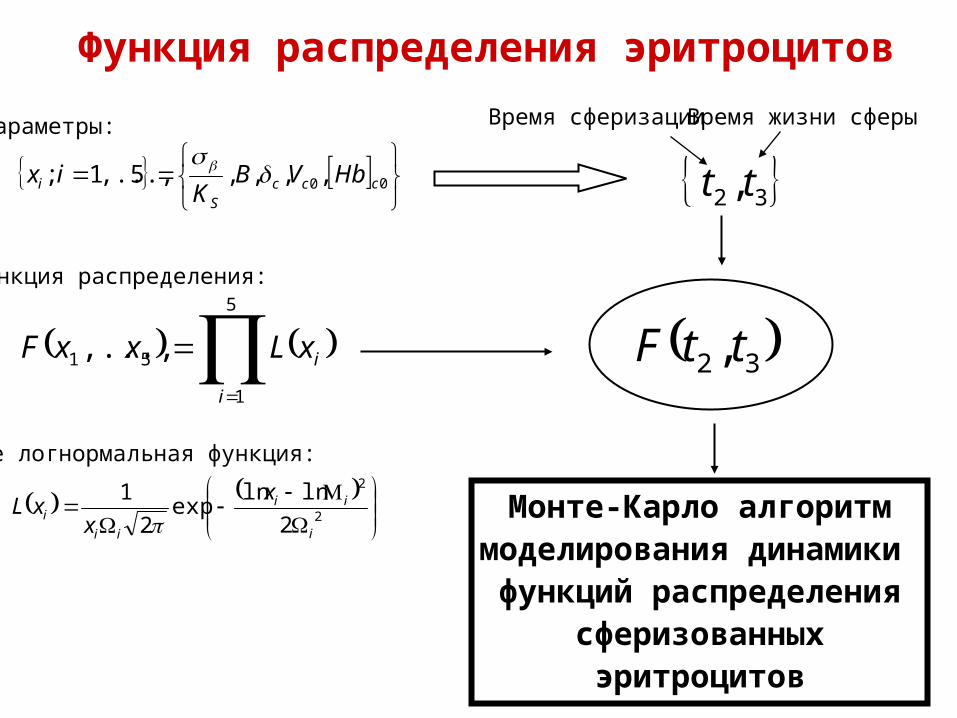
Функция распределения эритроцитов
5
1
51 ,...,
i
ixLxxF
2
2
2
lnlnexp
2
1
i
ii
ii
i
x
xxL
00 ,,,,5,...,1; cccS
i HbVBK
ix
Параметры:
Функция распределения:
где логнормальная функция:
32 , tt
Время сферизацииВремя жизни сферы
Монте-Карло алгоритммоделирования динамики функций распределения
сферизованныхэритроцитов
32 , ttF
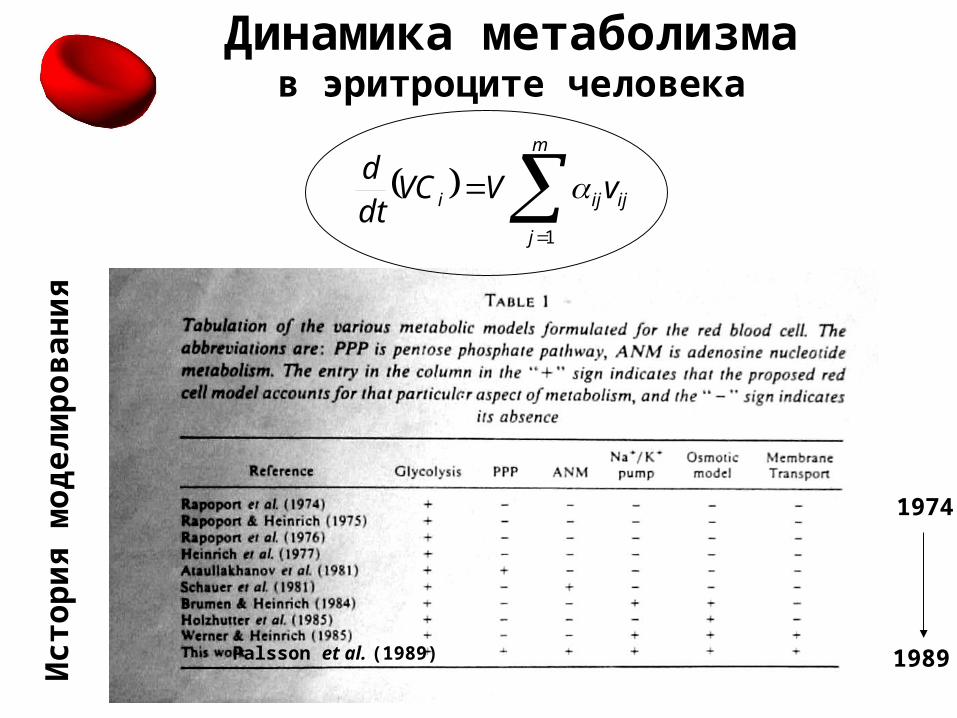
Динамика метаболизмав эритроците человека
Ист
ор
ия
мо
дел
ир
ов
ани
я
1989
1974
Palsson et al. (1989)
m
j
ijiji vVVCdtd
1

Аб
бр
еви
атур
а м
етаб
ол
ито
в э
ри
тро
ци
та

Аб
бр
еви
атур
а м
етаб
ол
ито
в э
ри
тро
ци
та
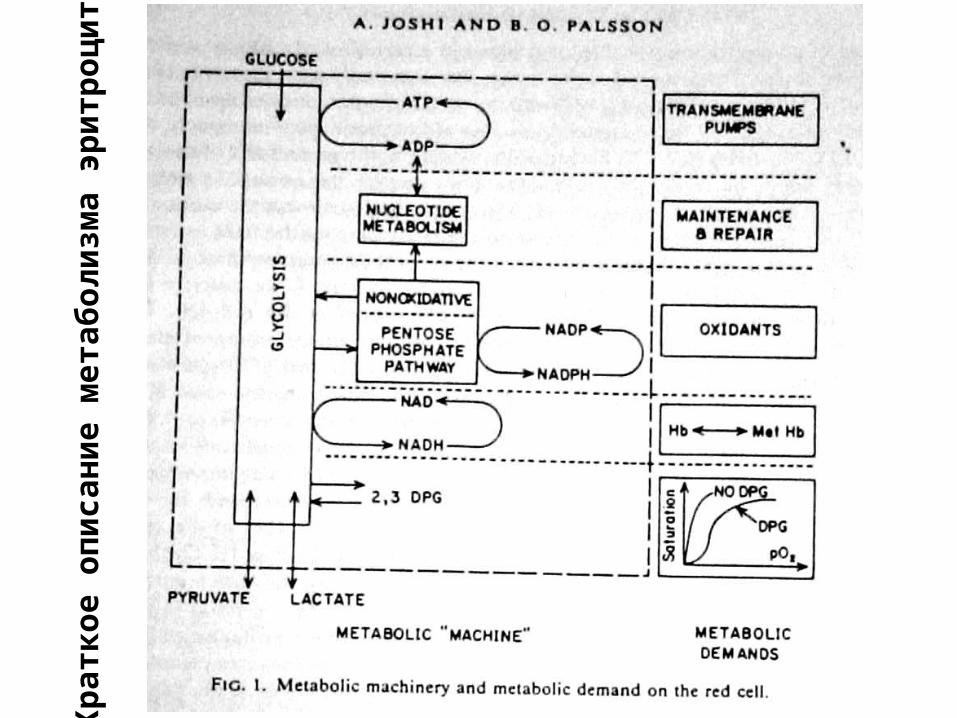
Кр
атко
е о
пи
сан
ие
мет
або
ли
зма
эри
тро
ци
та
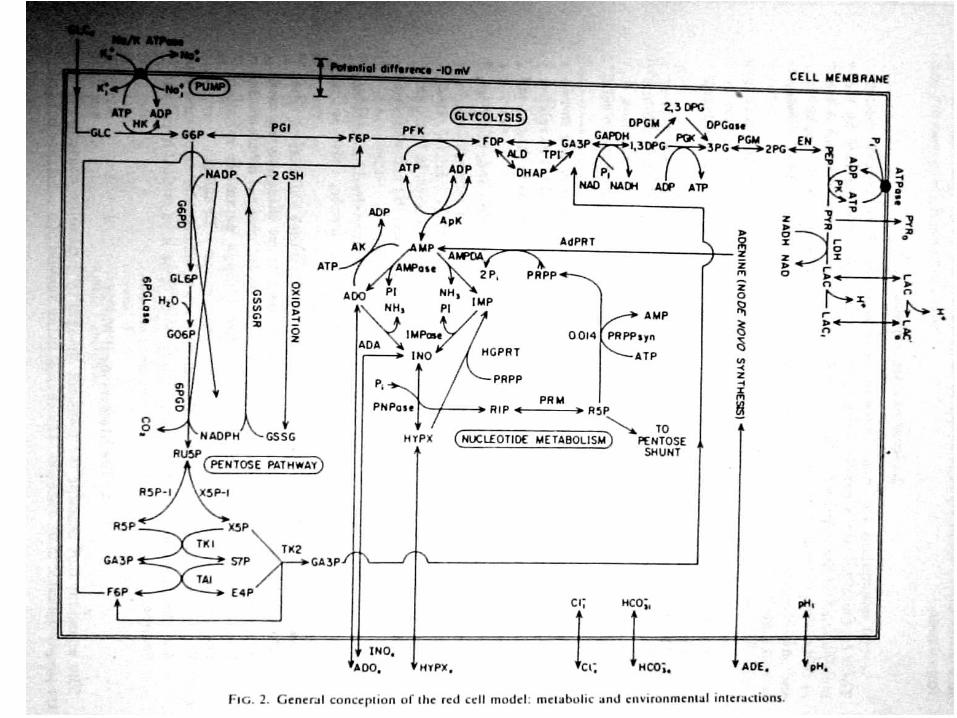


Основные ограничения:
1) Электронейтральность
2) Баланс осмотического давления
n
i
iiCz
1
0
n
i
osmiiCRT
1
mMinHbHbHbHb20258.00645.01
7.892
6.691
2211
10
10
1/41/236.15
pH
pH
Hb
b
b
bbbbz
(буферные свойства)
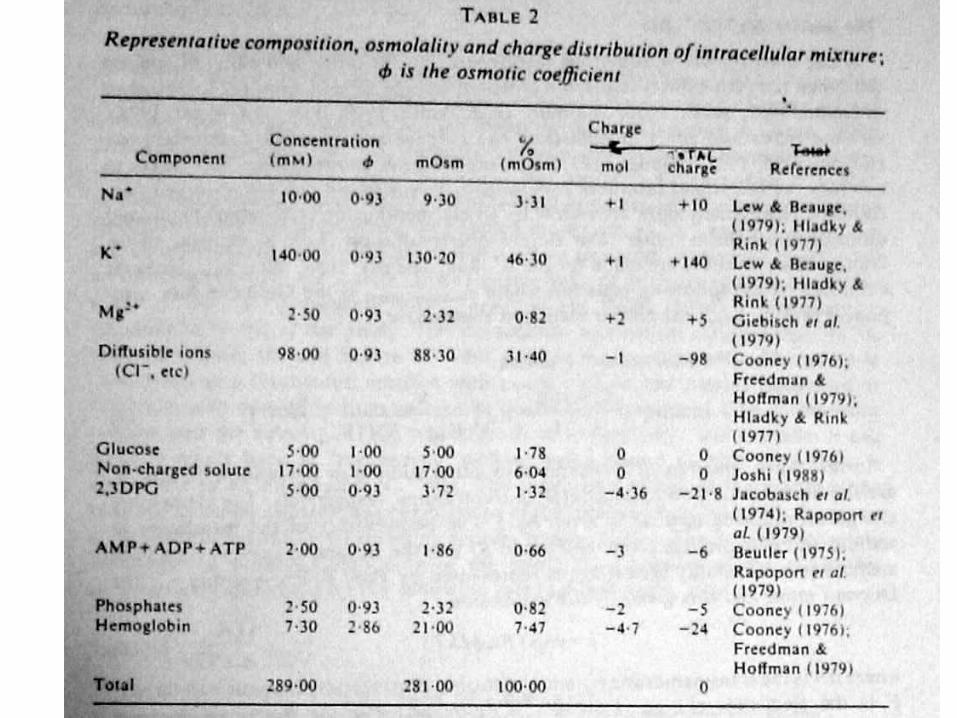

Трансмебранный K+/Na+ обмен
pumpKleakin
pumpNaleakin
vvKdtd
vvNadtd
2
3
,
,
exexinexex
exexm
ATPpumppump
inXm
in
exXm
exXm
inexXdfacilitatefusionelectrodifXleak
KBKkkkkBBNaBKKBBB
KBKV
KATP
ATPv
RTFr
rXK
rX
XK
Xv
r
XrXrzkvvv
2
2
131221
3
3
2
221
2
2
,
,,,2
2
,
//1/2
2/2/
/exp
1ln
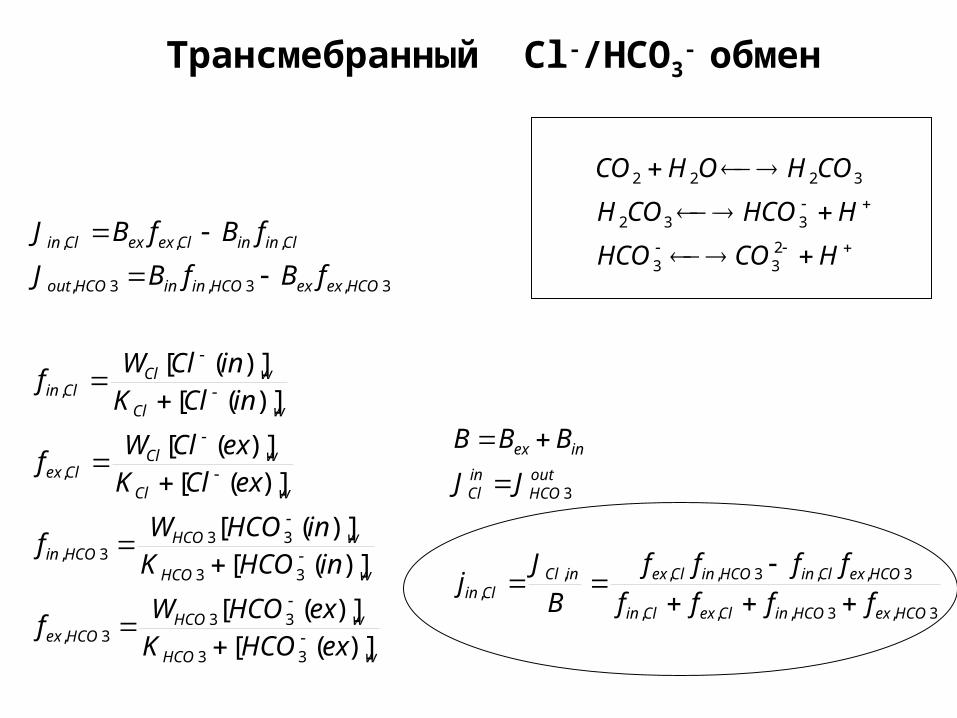
Трансмебранный Cl-/HCO3- обмен
wHCO
wHCOHCOex
wHCO
wHCOHCOin
wCl
wClClex
wCl
wClClin
HCOexexHCOininHCOout
ClininClexexClin
exHCOKexHCOW
f
inHCOKinHCOW
f
exClKexClW
f
inClKinClW
f
fBfBJ
fBfBJ
)]([)]([
)]([)]([
)]([)]([
)]([)]([
33
333,
33
333,
,
,
3,3,3,
,,,
3,3,,,
3,,3,,,,
3
HCOexHCOinClexClin
HCOexClinHCOinClexinClClin
outHCO
inCl
inex
ffff
ffff
B
Jj
JJ
BBB
HCOHCO
HHCOCOH
COHOHCO
233
332
3222
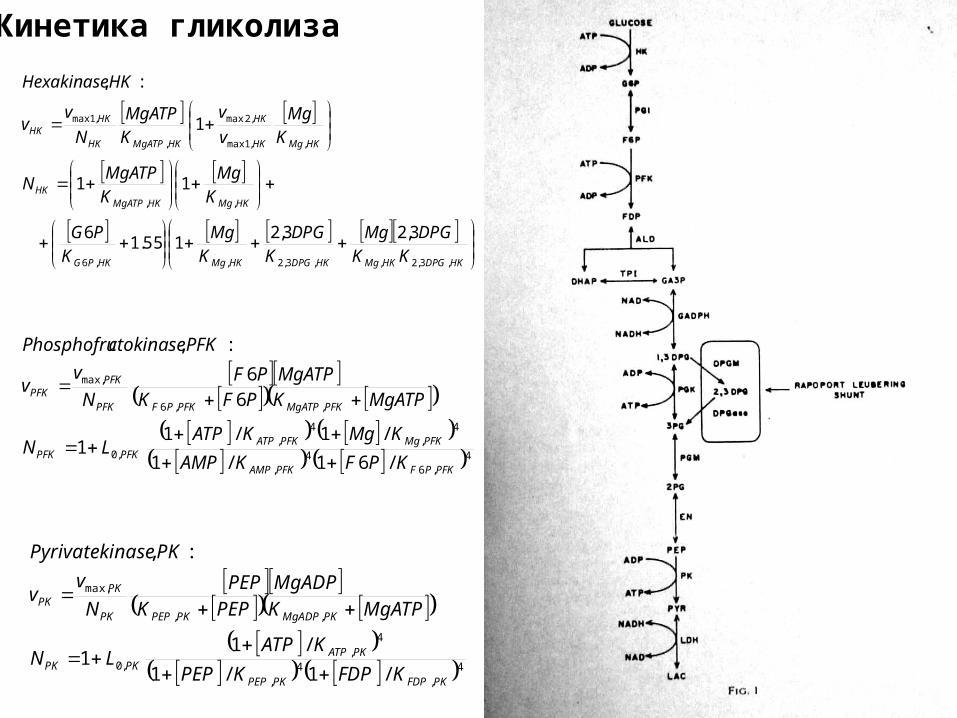
Кинетика гликолиза
HKDPGHKMgHKDPGHKMgHKPG
HKMgHKMgATPHK
HKMgHK
HK
HKMgATPHK
HKHK
KK
DPGMg
K
DPG
K
Mg
K
PG
K
Mg
K
MgATPN
K
Mg
v
v
K
MgATP
N
vv
HKHexakinase
,3,2,,3,2,,6
,,
,,1max
,2max
,
,1max
3,23,2155.1
6
11
1
:,
4,6
4,
4,
4,
,0
,,6
max,
/61/1
/1/11
66
:,
PFKPFPFKAMP
PFKMgPFKATPPFKPFK
PFKMgATPPFKPFPFK
PFKPFK
KPFKAMP
KMgKATPLN
MgATPKPFKMgATPPF
N
vv
PFKctokinasePhosphofru
4,
4,
4,
,0
,,
max,
/1/1
/11
:,
PKFDPPKPEP
PKATPPKPK
PKMgADPPKPEPPK
PKPK
KFDPKPEP
KATPLN
MgATPKPEPKMgADPPEP
N
vv
PKkinasePyrivate
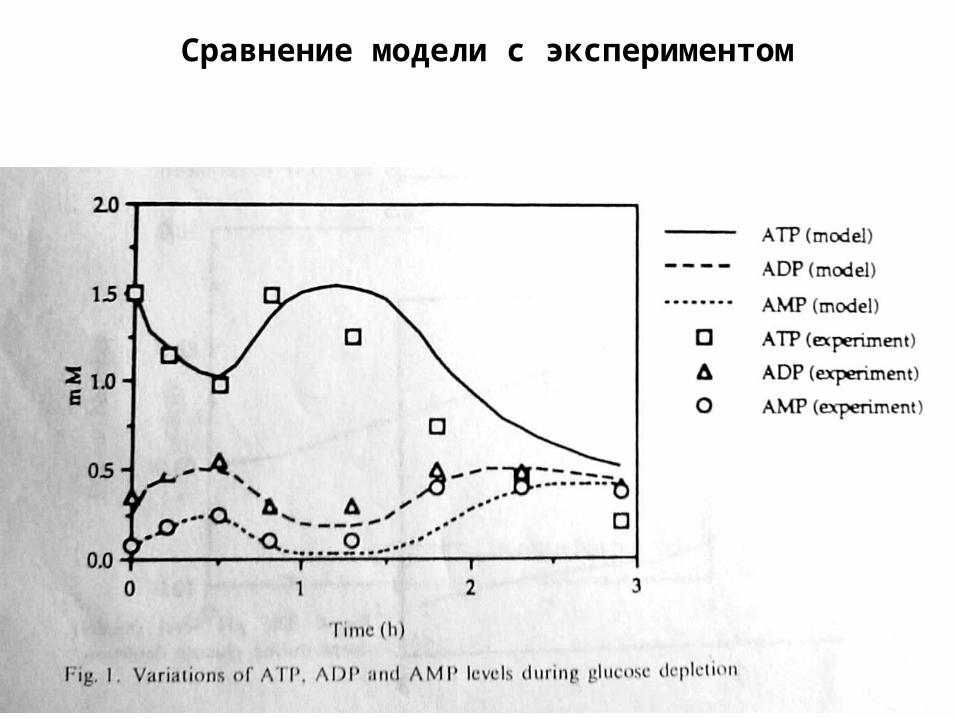
Сравнение модели с экспериментом
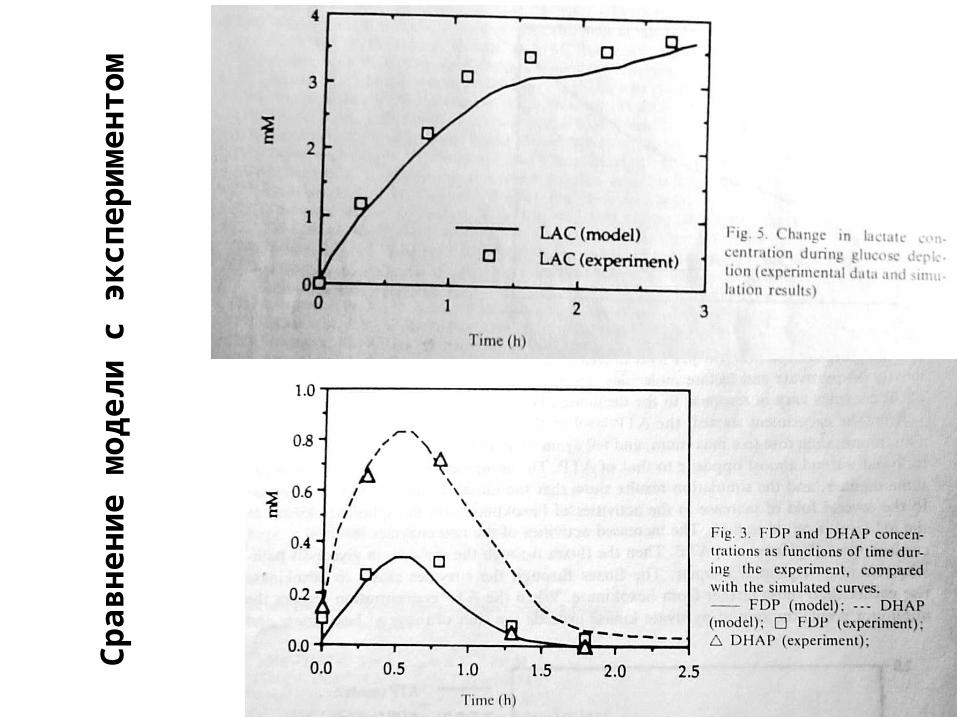
Ср
авн
ени
е м
од
ели
с э
ксп
ери
мен
том
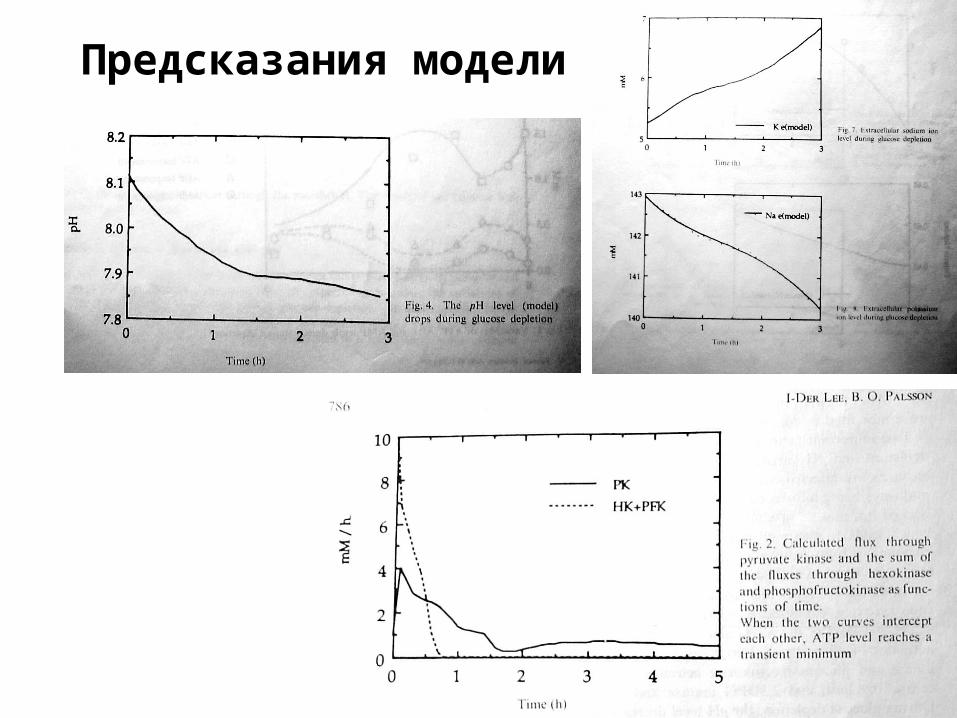
Предсказания модели






























