-
PREPOZNAVANJE ANTIGENA USPECIFINOM (STEENOM) IMUNSKOM
ODGOVORU:
Receptori za antigen(molekuli koji prepoznaju Ag na
limfocitima)
- Kakva je struktura receptora za Ag ?- Kako receptori
prepoznaju Ag ?- Kako antigenski receptori limfocita uspevaju da
prepoznaju ekstremno razliite antigenem, a da pri tome isporue iste
aktivacione signale u elije?
Razvoj limfocitnog repertoara (Skup specifinosti svih limfocita)
- Kako nastaje ogromna raznolikost limfocitnih receptora?
-
Specifino prepoznavanje antigena vre receptori za antigen na B i
T limfocitima: BCR (B elijski receptor, membranski Ig)TCR (T
elijski receptor)
BCR i TCR su klonalno rasporeeni: Jedan klon/ jedna specifinost
receptora/prepoznavanje jednog Ag 109-1011 klonova limfocita Iako
klonovi limfocita prepoznaju razliite antigene, antigenski
receptori sprovode biohemijske signale koji su u osnovi isti u svim
limfocitima
-
BCR i TCR su nekovalentno vezani za druge, nepromenjive
strukture ija je funkcija da u unutranjost elije isporue
aktivacione signale pokrenute prepoznavanjem antigena
Kompleks BCR i njemu pridruenih signalnih molekula:receptorski
kompleks B elija (BCR kompleks)
Kompleks TCR i njemu pridruenih signalnih molekula:receptorski
kompleks T elija (TCR kompleks)
-
BCR KOMPLEKS: antitelo + Iga i Igb (imunoglobulin)
prepoznavanje Ag prenos signala aktivacije
-
TCR KOMPLEKS:TCR molekul+ CD3 molekul + z lanci
prepoznavanje antigenaprenos signala aktivacije
-
Heterodimer (a i b lanac)Oba lanca su usidrena u elijsku
membranuOba lanca imaju:varijabilni (V) i konstantni (C) regionV i
C regioni TCR su homologni V i C regionima IgVa i Vb sadre po tri
hipervarijabilna (CDR) regionaTCR se ne stvara u sekretornoj formi
i ne podlee afinitetnom sazrevanju.STRUKTURA TCR
-
Preko 90% T limfocita ima TCRabOko 5 do 10% T limfocita ima
TCRgd T limfociti koji ispoljavaju gd TCR su u najveem broju
prisutni u epitelu, imaju ogranienu raznolikost gd TCR su po
strukturi slini ab TCR, ali T limfociti koji imaju gd TCR
prepoznaju proteinske i neproteinske Ag, koje uglavnom NE
prezentuju klasini MHC I molekuliNKT elije: imaju ab TCR koji
prepoznaje lipide vezane za CD1 molekule (nepolimorfni, slini MHC
I)
-
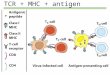
Vezivanje TCR za antigen
TCR vezuje samo peptid/MHC kompleks
-
U prepoznavanju peptid/MHC kompleksa uestvuju Va i Vb domeni TCR
TCR prepoznaje samo 1-3 aminokiselinska ostatka peptida vezanog za
MHC molekul.TCR prepoznaje samo nekoliko peptida sloenog
makromolekula: imunodominantne epitopeTCR prepoznaje istovremeno i
polimorfne delove sopstvenog MHC molekula koji nosi peptid
Kako TCR prepoznaje kompleks peptid/MHC ?
-
CD3 i z proteini TCR kompleksa Nekovalentno vezani za ab TCR
heterodimer
Kada TCR prepozna antigen,ovi proteini sprovode signale koji
dovode do aktivacije T limfocita
CD3 molekul se sastoji iz tri proteina: g, d i e
Sva tri lanca CD3 molekula imaju po jedan Ig-slian domen na
N-terminalnom kraju molekula (pripadaju imunoglobulinskoj
superfamiliji)
Svaki TCR kompleks sadri:jedan TCR ab dimer, jedan CD3 ge
heterodimer, jedan CD3 de heterodimer, i jedan disulfidno vezan zz
homodimer
-
CD3 i z proteini TCR kompleksaCitoplazmatski domeni CD3 g, d i e
lanaca sadre po jedan IMUNORECEPTORSKI TIROZINSKI AKTIVACIONI MOTIV
(ITAM)Citoplazmatski deo z lanca ima tri ITAM
ITAM igra centralnu ulogu u transdukciji signala
Ostaci tirozina u ITAM sekvenci bivaju fosforilisani kada se TCR
vee za peptid/MHC kompleks. Fosforilisani ITAM vezuju i aktiviraju
protein tirozin-kinaze u citoplazmi
ITAM-i su naeni i u: Iga i Igb proteinima (u BCR kompleksu)
nekim Fc receptorimanekim aktivacionim receptorima NK elija
-
Komponente TCR kompleksa
-
TCR i akcesorski molekuliAkcesorski molekuli- Povrinski molekuli
T limfocita koji ne prepoznaju antigen, ali uestvuju u odgovoru T
elija na antigen- Isti su na svim T elijamaKoreceptori Receptori za
kostimulatore Adhezivni molekuliFunkcija:signalizacija adhezija
-
Koreceptori: CD4 i CD8 Koreceptori, jer zajedno sa TCR uestvuju
u vezivanju za MHC molekule Vezuju se za nepolimorfne regione MHC
molekula; uestvuju u sprovoenju signala Pripadaju Imunoglobulinskoj
superfamiliji
CD4 je monomerIma 4 Ig-slina domena2 Ig-slina domena CD4
proteina se vezuju za b2 domen klase II MHC
Veina CD8 molekula postoji kao dimer: CD8a i CD8b lanacSvaki
lanac ima po 1 Ig-slini domenIg domen CD8 lanca se vezuje za a3
domen klase I MHC molekula
-
CD4 i CD8 uestvuju u ranim dogaajima transdukcije signala nakon
to T limfociti prepoznaju peptid-MHC kompleks na APCSignalna
funkcija CD4 i CD8 molekula se ostvaruje tirozin kinazama (lck
tirozin kinaza, lan lan Src familije) koje su nekovalentno vezane
za citoplazmatski rep ovih molekula.
Lck se aktivira grupisanjem CD4 ili CD8 koreceptora
Lck potom fosforilie ostatke tirozina u ITAM-ima CD3 i z lanaca,
zapoinjui aktivacionu kaskadu u T limfocitima
-
Receptori za kostimulatore
Takoe uestvuju u prenosu signala, ali za razliku od koreceptora,
prepoznaju molekule na APC koji nisu deo peptid/MHC kompleksa
R za kostimulatore na TLy: CD28 Kada se CD28 vee za
kostimulatore na APC, sprovodi signal koji funkcionie zajedno sa
signalima koji se pokreu vezivanjem TCR i koreceptora za kompleks
peptid-MHC na APC Signal koji se sprovodi posredstvom CD28 je
neophodan za aktivaciju naivnih T Ly
Kostimulatori na AP (D, makrofagama i B Ly): B7-1 (CD80) i B7-2
(CD86), vezuju se za CD28kostimulatori, jer zajedno sa antigenom
stimuliu limfocite
CTLA-4 je strukturni homolog CD28ispoljava se na aktiviranim T
limfocitima, funkcija mu je da inhibira aktivaciju T limfocita
ponitavajui signale koji se sprovode putem TCR kompleksa i CD28
-
Adhezivni molekuliInterakcija adhezivnih molekula na T
limfocitima sa odgovarajuim ligandima na APC ima za cilj da
stabilizuje vezivanje Telija za APC i time omogui uspostavljanje
signala aktivacije
Najznaajniji adhezivni molekuli pripadaju grupi heterodimernih
proteina integrina
LFA-1 (integrin) na T limfocitima se vezuje za ICAM-1 na APC
LFA-1 (antigen-1 povezan sa funkcijom leukocita, eng. leukocyte
function-associated antigen-1) ICAM-1 (intercelularni adhezivni
molekul-1)
Integrini su znaajni i za usmereno kretanje efektorskih T elija
iz cirkulacije ka mestu infekcije.
-
Imunoglobulinska superfamilijaVelika familija proteina koji
sadre globularni strukturni motiv, nazvan imunoglobulinski (Ig)
domen, najpre opisan kod antitela
Mnogi proteini od znaaja za imunski sistem pripadaju ovoj
superfamiliji:Ig, Iga/Igb, TCR, MHC I i II, CD4, CD8, CD28, g, d i
e lanci CD3, ICAM-1
-
NASTANAK REPERTOARA SPECIFINOSTIiliKako nastaje tako veliki broj
(>109) receptora za antigen razliite specifinosti ?
Repertoar se formira u toku sazrevanja limfocita !
-
Sazrevanje limfocita obuhvata tri vrste procesa:PROLIFERACIJU
nezrelih elijaIL-7 pospeuje proliferaciju progenitora B i T Ly pre
nego to ispolje antigenske receptore
EKSPRESIJU RECEPTORA za antigen TCR i BCR su proizvodi nekoliko
genskih segmenata koji su odvojeni u genomu, ali se tokom
sazrevanja Ly rekombinuju (spajaju). Rekombinacija je SLUAJNI
PROCES RAZNOLIKOST REPERTOARA Repertoar se formira somatskim
rekombinacijama genskih segmenata koji kodiraju varijabilne regione
receptora
SELEKCIJU
Zasniva se na ekspresiji kompletnog antigenskog receptora, kao i
na tome ta taj receptor prepoznaje
Tzv. pre-limfociti koji ne ispolje receptor umiru.
Nezreli T limfociti se selektuju na osnovu sposobnosti da
prepoznaju sopstvene MHC molekule (pozitivna selekcija)
Nezreli T i B limfociti se eliminiu ukoliko visokim afinitetom
prepoznaju sopstvene antigene (negativna selekcija) ime se
uklanjaju potencijalno tetni autoreaktivni limfociti.
-
U genomu limfocitnih progenitora: Ig i TCR geni se nalaze u
konfiguraciji nasleenoj od roditelja.
U toj konfiguraciji, lokusi za H i L lance Ig i a i b lance TCR
sadre:vei broj gena za varijabilni (V) i jedan ili nekoliko gena za
konstantni (C) region. Izmeu V i C gena nalaze se spojni genski
segmenti (engl. joining, J ) Samo na lokusu za teki (H) lanac Ig i
lokusu za b lanac TCR prisutni su genski segmenti raznolikosti
(engl. diversity, D)Nasleena konfiguracija genskih lokusa za
antigenske receptore
-
V- D- J rekombinacijakodira V region ! Opredeljuje limfocitni
progenitor da postane B limfocit!
Somatsku rekombinacijuobavljaju V(D)J rekombinaze
VDJ rekombinaza specifina za limfocite: RAG-1 i RAG-2 (engl.
recombinase-activating gene)
Rekombinacija i ekspresija Ig gena
-
Raznolikost antigenskih receptora (>109) rezultat
je:Kombinatorne raznolikosti Nastaje sluajnim kombinacijama V, D i
J genskih segmenata (ograniena je brojem raspoloivih V, D i J
genskih segmenata)
2. Spojne raznolikostiUklanjanje nukleotida iz V, D i J
segmenata tokom rekombinacije (egzonukleaze) ili Dodavanje novih
nukleotida na mesta V(D) J rekombinacija, pri emu nastaju tzv.
N-regioni i P-nukleotidiV(D)J spoj kodira redosled AK u CDR3!
3. Povezivanja razliitih H i L lanaca Ig, odnosno razliitih a i
b lanaca TCR
-
omoguava preivljavanje i proliferaciju pre-B elija prva
kontrolna taka u razvoju B elija(elije koje ne ispolje pre-BCR
umiru apoptozom) zaustavlja rekombinaciju gena za H lanac na drugom
hromozomu (iskljuivanje alela) svaka elija ima BCR jedne
specifinostipodstie rekombinaciju gena za L lance FAZE U SAZREVANJU
I SELEKCIJI B LIMFOCITApre-BCR (m lanac + surogat L lanac) +Iga i
IgbU pre-B elijama, Ig geni u lokusu za H lance jednog hromozoma
podleu rekombinaciji koja dovodi do nastanka m lanca
-
omoguava preivljavanje i proliferaciju nezrelih B elija (koje
ispoljavaju kompletne receptore druga kontrolna taka u razvoju B
elija zaustavlja sintezu rekombinaza i dalju rekombinaciju na
ostalim genskim lokusima za L lance svaka B elija ima samo jedan,
ili k ili l lanacAko BCR (IgM) vee antigen visokim afinitetom:
negativna selekcija (apoptoza) ili reaktivacija rekombinaza i
sinteza drugog L lanca (tzv. preureenje receptora)FAZE U SAZREVANJU
I SELEKCIJI B LIMFOCITABCR (m lanac + k ili l lanac) +Iga i Igb
-
FAZE U SAZREVANJU I SELEKCIJI B LIMFOCITAmembranski IgM i IgD +
Iga i Igb
(ispoljeni su zajedno jer iRNK rekombinovanog VDJ H lanca moe da
se spoji ili sa iRNK za Cm ili sa iRNK za Cd )
-
FAZE U SAZREVANJU I SELEKCIJI T LIMFOCITAU pro-T elijama
(CD4-CD8-, tj. DN):zapoinje V-D-J rekombinacija gena za b lanac
TCR
-
omoguava preivljavanje i proliferaciju pre-T elija prva
kontrolna taka u razvoju T Ly (elije koje ne ispolje pre-TCR umiru)
zaustavlja VDJ rekombinaciju gena za b lanac u drugom hromozomu
(iskljuivanje alela) podstie rekombinaciju gena za a lanac
TCRstimulie ekspresiju CD4 i CD8 FAZE U SAZREVANJU I SELEKCIJI T
LIMFOCITApre -TCR (b lanac + pre-Ta)+CD3 i z lanci
-
omoguava preivljavanje i proliferaciju nezrelih T elija druga
kontrolna taka u razvoju T elija ( elije koje ne ispolje a lanac
tj. TCR umiru) prepoznaje sopstveni peptid/MHC kompleks u timusu
pozitivna ili negativna selekcija odsustvo pozitivne selekcije smrt
zanemarivanjemFAZE U SAZREVANJU I SELEKCIJI T LIMFOCITA TCR (b + a
lanac)+CD3 i z lanci
-
DN (CD4-CD8-) elije: ispoljavaju pre-TCR kompleksDP (CD4+CD8+)
elije: ispoljavaju TCR kompleksJP elije (zrele, CD4+CD8- i
CD8+CD4-): ispoljavaju TCR kompleks
Pozitivna i negativna selekcija odvijaju se na nivou DP
elijaUkoliko DP elije ne prepoznaju Ag/MHC kompleks u timusu, one
umiru u timusu (smrt usled zanemarivanja)