Embed Size (px)
Citation preview

8. Lecture SS 20005
Cell Simulations 1
V9: Characterizing the Fluxome of Biological Cells
Qualitative methods for fluxome analysis:- Graph theoretical methods
Jeong & Barabasi: scale-free metabolic networks
Arita: different view resulting from different representation
- stochiometric analyses
Extreme pathway analysis
Large-scale experimental fluxome measurement on B. subtilis wildtype
and 137 mutants.

8. Lecture SS 20005
Cell Simulations 2
First breakthrough: scale-free metabolic networks
(d) The degree distribution, P(k), of the metabolic network illustrates its scale-free topology.
(e) The scaling of the clustering coefficient C(k) with the degree k illustrates the hierarchical
architecture of metabolism (The data shown in d and e represent an average over 43
organisms).
(f) The flux distribution in the central metabolism of Escherichia coli follows a power law,
which indicates that most reactions have small metabolic flux, whereas a few reactions, with
high fluxes, carry most of the metabolic activity. It should be noted that on all three plots the
axis is logarithmic and a straight line on such log–log plots indicates a power-law scaling.
CTP, cytidine triphosphate; GLC, aldo-hexose glucose; UDP, uridine diphosphate; UMP,
uridine monophosphate; UTP, uridine triphosphate.Barabasi & Oltvai, Nature Reviews Genetics 5, 101 (2004)

8. Lecture SS 20005
Cell Simulations 3
Different opinion
Arita, PNAS 101, 1543 (2004)
According to the formal definition, in a small-world network,
(i) most nodes (metabolites in our case) have a low connection degree, and the
degree distribution follows a power law also referred to as scale-freeness;
(ii) high-degree nodes, called hubs, dominate the network, and most nodes are
clustered around hubs; and
(iii) the average path length (AL; i.e., the average of the shortest path length over
all pairs of nodes in the network) remains the theoretical minimum, that of a
random graph.
Because of its topology with few hubs, a small-world network may be resistant to
random failures: any peripheral node is likely to have a low connection degree and
is therefore expendable. In biological networks, the hubs are thought to be
functionally important and phylogenetically oldest.

8. Lecture SS 20005
Cell Simulations 4
Different opinion
Arita, PNAS 101, 1543 (2004)
Although several groups confirmed the small-world property of small-molecule
metabolisms in multiple data sources, the details of their results differ depending
on the purpose of the analysis and its data-preparation scheme.
Notable differences are attributable to the reversibility of enzymatic reactions and
to the treatment of metabolically ubiquitous compounds referred to as coenzymes
or inorganics. Table 1 summarizes differences in the major analyses and
compares the average path length (AL) and hub metabolites they identified.

8. Lecture SS 20005
Cell Simulations 5
Different opinion
Arita, PNAS 101, 1543 (2004)
All of these studies used the same
algorithmic procedure, and
discrepancies are ascribable to
the different aims of their network
analyses.
Jeong et al. computed the
proximity of metabolites by
regarding all substrates and
products in the same reaction as
adjacent (Fig. 1)

8. Lecture SS 20005
Cell Simulations 6
Basis for analysis
Arita, PNAS 101, 1543 (2004)
To reproduce biochemical pathways in the traditional metabolic map, however,
metabolites to be linked cannot be defined per se by compounds or reactions.
The biochemical link between metabolites is context-sensitive; it depends on the
conserved structural moieties in the adjacent reactions.
To accurately compute the reaction connectivity as in the traditional metabolic map,
we used digitally compiled atomic mappings, i.e., atomic position pairs between
substrates and products corresponding to the substructural moieties conserved in
each reaction.
With this information, we reassessed the global properties of metabolic networks
with special emphasis on the small-world hypothesis.

8. Lecture SS 20005
Cell Simulations 7
Basis for analysis
Arita, PNAS 101, 1543 (2004)
8. Lecture SS 20005
Cell Simulations 8
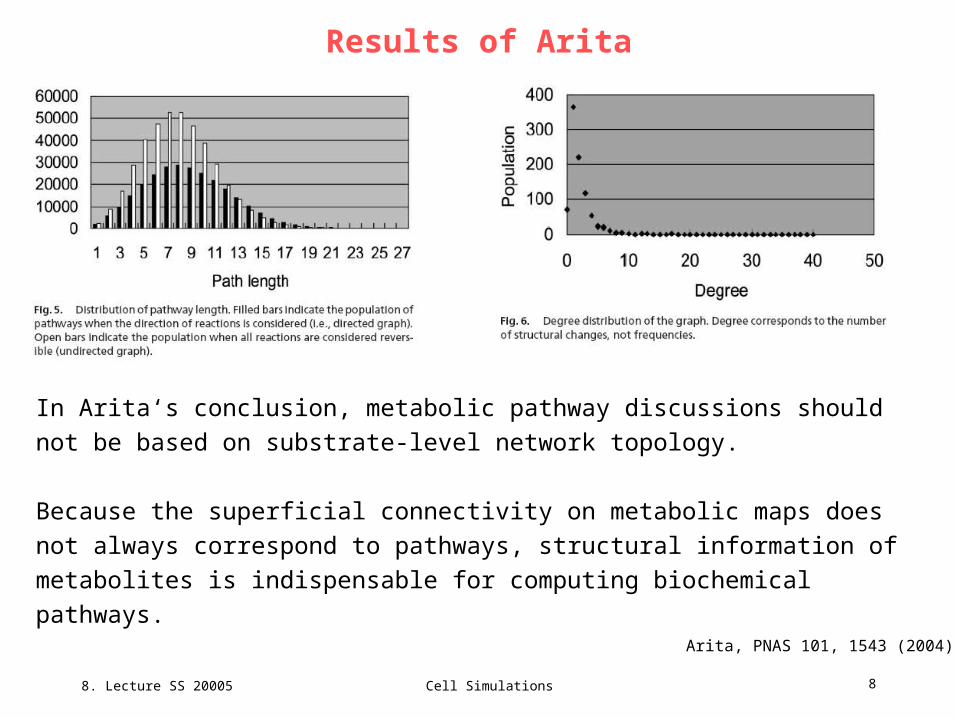
Results of Arita
Arita, PNAS 101, 1543 (2004)
In Arita‘s conclusion, metabolic pathway discussions should not be based on
substrate-level network topology.
Because the superficial connectivity on metabolic maps does not always
correspond to pathways, structural information of metabolites is indispensable for
computing biochemical pathways.

8. Lecture SS 20005
Cell Simulations 9
Review from bioinformatics III: V19 Extreme Pathwaysintroduced into metabolic analysis by the lab of Bernard Palsson
(Dept. of Bioengineering, UC San Diego). The publications of this lab
are available at http://gcrg.ucsd.edu/publications/index.html
The extreme pathway
technique is based
on the stoichiometric
matrix representation
of metabolic networks.
All external fluxes are
defined as pointing outwards.
Schilling, Letscher, Palsson,
J. theor. Biol. 203, 229 (2000)

8. Lecture SS 20005
Cell Simulations 10
Extreme Pathways – algorithm - setup
The algorithm to determine the set of extreme pathways for a reaction network
follows the pinciples of algorithms for finding the extremal rays/ generating
vectors of convex polyhedral cones.
Combine n n identity matrix (I) with the transpose of the stoichiometric
matrix ST. I serves for bookkeeping.
Schilling, Letscher, Palsson,
J. theor. Biol. 203, 229 (2000)
S
I ST

8. Lecture SS 20005
Cell Simulations 11
separate internal and external fluxes
Examine constraints on each of the exchange fluxes as given by
j bj j
If the exchange flux is constrained to be positive do nothing.
If the exchange flux is constrained to be negative multiply the
corresponding row of the initial matrix by -1.
If the exchange flux is unconstrained move the entire row to a temporary
matrix T(E). This completes the first tableau T(0).
T(0) and T(E) for the example reaction system are shown on the previous slide.
Each element of this matrices will be designated Tij.
Starting with x = 1 and T(0) = T(x-1) the next tableau is generated in the following
way:
Schilling, Letscher, Palsson,
J. theor. Biol. 203, 229 (2000)

8. Lecture SS 20005
Cell Simulations 12
idea of algorithm
(1) Identify all metabolites that do not have an unconstrained exchange flux
associated with them.
The total number of such metabolites is denoted by .
For the example, this is only the case for metabolite C ( = 1).
What is the main idea?
- We want to find balanced extreme pathways
that don‘t change the concentrations of
metabolites when flux flows through
(input fluxes are channelled to products not to
accumulation of intermediates).
- The stochiometrix matrix describes the coupling of each reaction to the
concentration of metabolites X.
- Now we need to balance combinations of reactions that leave concentrations
unchanged. Pathways applied to metabolites should not change their
concentrations the matrix entries
need to be brought to 0.Schilling, Letscher, Palsson,
J. theor. Biol. 203, 229 (2000)

8. Lecture SS 20005
Cell Simulations 13
keep pathways that do not change concentrations of internal metabolites
(2) Begin forming the new matrix T(x) by copying
all rows from T(x – 1) which contain a zero in the
column of ST that corresponds to the first
metabolite identified in step 1, denoted by index c.
(Here 3rd column of ST.)
Schilling, Letscher, Palsson, J. theor. Biol. 203, 229 (2000)
1 -1 1 0 0 0
1 0 -1 1 0 0
1 0 1 -1 0 0
1 0 0 -1 1 0
1 0 0 1 -1 0
1 0 0 -1 0 1
1 -1 1 0 0 0
T(0) =
T(1) =
+

8. Lecture SS 20005
Cell Simulations 14
balance combinations of other pathways
(3) Of the remaining rows in T(x-1) add together
all possible combinations of rows which contain
values of the opposite sign in column c, such that
the addition produces a zero in this column.
Schilling, et al.
JTB 203, 229
1 -1 1 0 0 0
1 0 -1 1 0 0
1 0 1 -1 0 0
1 0 0 -1 1 0
1 0 0 1 -1 0
1 0 0 -1 0 1
T(0) =
T(1) =
1 0 0 0 0 0 -1 1 0 0 0
0 1 1 0 0 0 0 0 0 0 0
0 1 0 1 0 0 0 -1 0 1 0
0 1 0 0 0 1 0 -1 0 0 1
0 0 1 0 1 0 0 1 0 -1 0
0 0 0 1 1 0 0 0 0 0 0
0 0 0 0 1 1 0 0 0 -1 1

8. Lecture SS 20005
Cell Simulations 15
remove “non-orthogonal” pathways
(4) For all of the rows added to T(x) in steps 2 and 3 check to make sure that no
row exists that is a non-negative combination of any other sets of rows in T(x) .
Schilling et al.
JTB 203, 229

8. Lecture SS 20005
Cell Simulations 16
repeat steps for all internal metabolites
(5) With the formation of T(x) complete steps 2 – 4 for all of the metabolites that do
not have an unconstrained exchange flux operating on the metabolite,
incrementing x by one up to . The final tableau will be T().
Note that the number of rows in T () will be equal to k, the number of extreme
pathways.
Schilling et al.
JTB 203, 229

8. Lecture SS 20005
Cell Simulations 17
balance external fluxes
(6) Next we append T(E) to the bottom of T(). (In the example here = 1.)
This results in the following tableau:
Schilling et al.
JTB 203, 229
T(1/E) =
1 -1 1 0 0 0
1 1 0 0 0 0 0
1 1 0 -1 0 1 0
1 1 0 -1 0 1 0
1 1 0 1 0 -1 0
1 1 0 0 0 0 0
1 1 0 0 0 -1 1
1 -1 0 0 0 0
1 0 -1 0 0 0
1 0 0 0 -1 0
1 0 0 0 0 -1

8. Lecture SS 20005
Cell Simulations 18
balance external fluxes
(7) Starting in the n+1 column (or the first non-zero column on the right side),
if Ti,(n+1) 0 then add the corresponding non-zero row from T(E) to row i so as to
produce 0 in the n+1-th column.
This is done by simply multiplying the corresponding row in T(E) by Ti,(n+1) and
adding this row to row i .
Repeat this procedure for each of the rows in the upper portion of the tableau so
as to create zeros in the entire upper portion of the (n+1) column.
When finished, remove the row in T(E) corresponding to the exchange flux for the
metabolite just balanced.
Schilling et al.
JTB 203, 229

8. Lecture SS 20005
Cell Simulations 19
balance external fluxes
(8) Follow the same procedure as in step (7) for each of the columns on the right
side of the tableau containing non-zero entries.
(In this example we need to perform step (7) for every column except the middle
column of the right side which correponds to metabolite C.)
The final tableau T(final) will contain the transpose of the matrix P containing the
extreme pathways in place of the original identity matrix.
Schilling et al.
JTB 203, 229
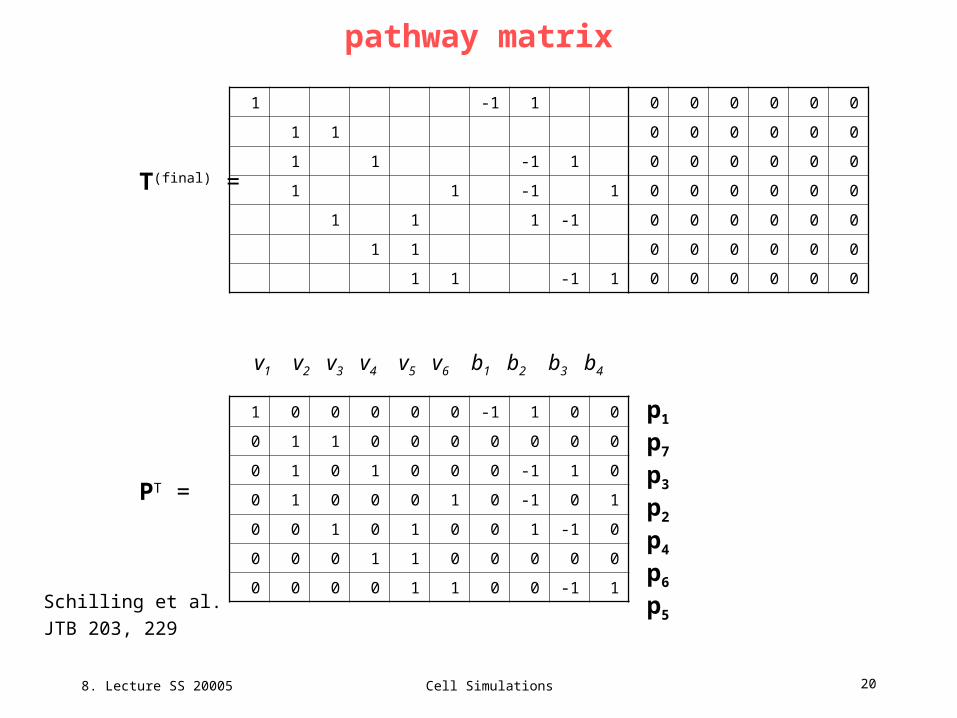
8. Lecture SS 20005
Cell Simulations 20
pathway matrix
T(final) =
PT =
Schilling et al.
JTB 203, 229
1 -1 1 0 0 0 0 0 0
1 1 0 0 0 0 0 0
1 1 -1 1 0 0 0 0 0 0
1 1 -1 1 0 0 0 0 0 0
1 1 1 -1 0 0 0 0 0 0
1 1 0 0 0 0 0 0
1 1 -1 1 0 0 0 0 0 0
1 0 0 0 0 0 -1 1 0 0
0 1 1 0 0 0 0 0 0 0
0 1 0 1 0 0 0 -1 1 0
0 1 0 0 0 1 0 -1 0 1
0 0 1 0 1 0 0 1 -1 0
0 0 0 1 1 0 0 0 0 0
0 0 0 0 1 1 0 0 -1 1
v1 v2 v3 v4 v5 v6 b1 b2 b3 b4
p1 p7 p3 p2 p4 p6 p5

8. Lecture SS 20005
Cell Simulations 21
Extreme Pathways for model system
Schilling et al.
JTB 203, 229
1 0 0 0 0 0 -1 1 0 0
0 1 1 0 0 0 0 0 0 0
0 1 0 1 0 0 0 -1 1 0
0 1 0 0 0 1 0 -1 0 1
0 0 1 0 1 0 0 1 -1 0
0 0 0 1 1 0 0 0 0 0
0 0 0 0 1 1 0 0 -1 1
v1 v2 v3 v4 v5 v6 b1 b2 b3 b4
p1 p7 p3 p2 p4 p6 p5
2 pathways p6 and p7 are not shown (right below) because all exchange fluxes with the exterior are 0.Such pathways have no net overall effect on the functional capabilities of the network.They belong to the cycling of reactions v4/v5 and v2/v3.

8. Lecture SS 20005
Cell Simulations 22
How reactions appear in pathway matrix
In the matrix P of extreme pathways, each column is an EP and each row
corresponds to a reaction in the network.
The numerical value of the i,j-th element corresponds to the relative flux level
through the i-th reaction in the j-th EP.
Papin, Price, Palsson,
Genome Res. 12, 1889 (2002)
PPP TLM

8. Lecture SS 20005
Cell Simulations 23
Papin, Price, Palsson, Genome Res. 12, 1889 (2002)
A symmetric Pathway Length Matrix PLM can be calculated:
where the values along the diagonal correspond to the length of the EPs.
PPP TLM
Properties of pathway matrix
The off-diagonal terms of PLM are the number of reactions that a pair of extreme
pathways have in common.

8. Lecture SS 20005
Cell Simulations 24
Papin, Price, Palsson, Genome Res. 12, 1889 (2002)
One can also compute a reaction participation matrix PPM from P:
where the diagonal correspond to the number of pathways in which the given
reaction participates.
TPM PPP
Properties of pathway matrix

8. Lecture SS 20005
Cell Simulations 25
Exp. distribution of metabolic fluxes in 137 B. subtilis mutants
Relative (a−e) and absolute (f−h, light gray arrows)
carbon fluxes during exponential growth on
glucose. Relative fluxes in a−c were analytically
quantified from the mass isotope distribution by
metabolic flux ratio analysis. They specify the
contribution of a given pathway or reaction to the
synthesis of a particular metabolite.
Wild-type values are indicated by asterisks.
CoA, coenzyme A; PP, pentose phosphate.
Observation:
Absolute fluxes in and out of the cell
varied by 31 – 55% around the wt.
Relative fluxes inside the cell varied
only by 3-8%!Fischer & Sauer, Nature Genetics 37, 636 (2005)

8. Lecture SS 20005
Cell Simulations 26
C13 flux analysis
Fischer & Sauer, Eur J Biochem 270, 880 (2003)

8. Lecture SS 20005
Cell Simulations 27
C13 flux analysis
Fischer & Sauer, Eur J Biochem 270, 880 (2003)

8. Lecture SS 20005
Cell Simulations 28
Calculation of metabolic flux ratios
Fischer & Sauer, Eur J Biochem 270, 880 (2003)

8. Lecture SS 20005
Cell Simulations 29
Calculation of metabolic flux ratios
Fischer & Sauer, Eur J Biochem 270, 880 (2003)

8. Lecture SS 20005
Cell Simulations 30
Effects of knockouts on absolute fluxes and optimality
Fischer & Sauer, Nature Genetics 37, 636 (2005)

8. Lecture SS 20005
Cell Simulations 31
Effects of knockouts on relative fluxes
(a) Relative fluxes through glycolysis
and the TCA cycle to the synthesis
of glyceraldehyde-3-phosphate and
oxaloacetate, respectively, as
obtained from flux ratio analysis.
The complementing fractions are
contributed by the pentose
phosphate pathway and the
anaplerotic reaction.
Fischer & Sauer, Nature Genetics 37, 636 (2005)
(b) Relative carbon fluxes to acetate and biomass formation.
Black circle: wild type.
Metabolic genes: the categories of central carbon metabolism, biosynthetic reactions and catabolic reactions.
Regulatory genes: the categories of transcriptional regulators and signal transduction.
The 10 mutants mentioned also have different ratios between catabolism and anabolism.
Extreme flux re-partitioning: critical reaction knockouts in TCA cycle (OdHA, SdhC, MdH), glycolysis (Pgi), or PPP (Zwf, GndA)Only 10 mutants without metabolic functions had altered intracellular fluxes.

8. Lecture SS 20005
Cell Simulations 32
Effects of knockouts on relative fluxes
(a) Relative fluxes through
glycolysis and the TCA cycle to
the synthesis of glyceraldehyde-
3-phosphate and oxaloacetate,
respectively, as obtained from
flux ratio analysis.
The complementing fractions are
contributed by the pentose
phosphate pathway and the
anaplerotic reaction.
Fischer & Sauer, Nature Genetics 37, 636 (2005)
Extreme repartioning in theperipheral biosynthetic network,pathway disruptions are mostlylethal; there are only a fewbypass reactions.
More redundancy exists withincentral carbon metabolism respond to food changes.

8. Lecture SS 20005
Cell Simulations 33
Effects of knockouts on absolute fluxes and optimality
Absolute molecular fluxes at the three key
divergent branch points of glucose catabolism:
(a) Glucose-6-phosphate (Glc6P),
(b) acetyl-coenzyme A (acetyl-coA) and
(c) the branching between anabolism and
catabolism.
A linear correlation between partitioned fluxes
shows a rigid branch point with a rate-
independent flux splitting. The wild type is
highlighted by a black circle.
(d) Growth optimality in 137 investigated
mutants. Lines indicate equal biomass
productivity (g (g glucose h)-1). The white area
indicates improved biomass productivity in the
mutant compared with the wild type (thick line).
Fischer & Sauer, Nature Genetics 37, 636 (2005)

8. Lecture SS 20005
Cell Simulations 34
Conclusions for B. subtilis
Systematic large-scale flux analysis shows that the control architecture of central
metabolism is designed to provide a rigid flux distribution that is largely independent
of the rate and yield of biomass formation.Key factor underlying the evolved robustness of metabolic networks to sustain
proliferation in the face of environmental and genetic perturbations.
Possible design principle:
Maintain B. subtilis in a standby mode that allows rapid responses to variations in
environmental conditions of its natural soil habitat.
Fischer & Sauer, Nature Genetics 37, 636 (2005)





























