Embed Size (px)
Citation preview

A gnotobiotic transgenic mouse model for studying interactions between small intestinal enterocytes
and intraepithelial lymphocytes*
Indira U. Mysorekar1, Robin G. Lorenz2*, and Jeffrey I. Gordon1
Departments of 1Molecular Biology and Pharmacology, and 2Pathology and Immunology, Washington
University School of Medicine, St. Louis, MO 63110
Correspondence to:
Jeffrey I. GordonDepartment of Molecular Biology and PharmacologyBox 8103Washington University School of Medicine660 South Euclid Ave.St. Louis, MO 63110Phone: 314-362-7243FAX: 314-362-7047e-mail: [email protected]
Running title: Cross-talk between intestinal enterocytes and intraepithelial lymphocytes
Copyright 2002 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on July 22, 2002 as Manuscript M205300200 by guest on M
arch 26, 2019http://w
ww
.jbc.org/D
ownloaded from

2
Summary
The mouse intestinal epithelium undergoes continuous renewal throughout life.
Intraepithelial lymphocytes (IELs) represent a significant fraction of this epithelium and play an
important role in intestinal mucosal barrier function. We have generated a germ-free transgenic
mouse model to examine the effects of a genetically-engineered proliferative abnormality in the
principal epithelial cell lineage (enterocytes) on IEL census and on IEL-enterocytic cross-talk.
SV40 large T antigen (TAgWt), or a mutant derivative (TAgK107/8) that does not bind pRB , were
expressed in small intestinal villus enterocytes under the control of elements from the intestinal
fatty acid binding protein gene (Fabpi). Quantitative immunohistochemical and flow cytometric
analyses of conventionally raised and germ-free FVB/N Fabpi-TAgWt, Fabpi-TAgK107/8, and
non-transgenic mice disclosed that forced re-entry of enterocytes into the cell cycle is
accompanied by an influx of thymically-educated αβTCR+ CD4+ and αβ TCR+ CD8αβ+ and a
decrease in intestinally-derived γδ TCR+CD8αα IELs. Real time quantitative RT-PCR studies of
jejunal villus epithelium recovered from germ-free transgenic and normal mice by laser capture
microdissection, and γδ TCR+ jejunal IELs purified by flow cytometry, disclosed that the
proliferative abnormality is accompanied by decreased expression of enterocytic IL-7 as well as
IEL IL-7Rα and TGFβ3. The analysis also revealed that normal villus epithelium expresses fms-
like tyrosine kinase 3 (Flt3), a known regulator of hematopoietic stem cell proliferation and
neuronal survival, and its ligand (Flt3L). Epithelial expression of this receptor and its ligand is
reduced by the proliferative abnormality while IEL expression of Flt3L remains constant.
Together, these findings demonstrate that changes in the proliferative status of the intestinal
epithelium affects maturation of γδTCR+ IELs, and produces an influx of αβTCR+ IELs even in
the absence of a microflora.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

3
Introduction
The adult mouse small intestine is a complex, spatially diversified ecosystem that
maintains distinctive cephalocaudal differences in its various functions. This regional variation in
function is accompanied by regional differences in the differentiation programs of its four
continuously renewing epithelial cell lineages, in the composition of its mucosal immune system,
and in the composition of its resident society of commensal/symbiotic microorganisms (reviewed
in refs. 1-3). A full understanding of how this ecosystem is organized and functions in health,
and how it is reorganized or disorganized in various disease states, requires knowledge about the
nature and regulation of interactions between its microflora, epithelium and gut-associated
lymphoid tissue (GALT) (1, 4, 5). The molecular nature and significance of the signals
exchanged between these components have been difficult to decipher because of the dynamic
nature and complexity of the system. One way of approaching this problem is to simplify the
ecosystem using inbred strains of mice with defined microbiological status (gnotobiotic animals).
For example, comparative functional genomics studies of mice containing no resident micro-
organisms (germ-free), conventionally raised mice harboring an complete microflora, and germ-
free animals that have been colonized with single species from the normal microflora (ex-germ-
free), have shown that indigenous commensal bacteria play an important role in regulating host
nutrient processing, fortifying the epithelial barrier, and organizing/educating the mucosal
immune system (5, 6).
The intestine contains a large population of intraepithelial lymphocytes (IELs1),
equivalent in size to the population of peripheral lymphocytes that resides in the spleen (7). IELs
are distributed throughout the epithelium that overlies small intestinal villi (average of one IEL
for every six to ten epithelial cells; ref. 8). Virtually all small intestinal IELs are T-cells, but they
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

4
are heterogeneous with respect to their surface phenotype. The majority are CD3+ and can be
divided into αβ T cell receptor positive (TCR+) and γδTCR+ subsets (8). They can be further
subdivided based on expression of CD8 (αα homodimer or αβ heterodimer) or CD4 coreceptors:
i.e., (i) γδTCR+ CD8α-CD8β−; (ii) γδTCR+ CD8α+CD8β- (abbreviated γδ+TCR+CD8αα); (iii)
αβTCR+ CD4+; (iv) αβTCR+ CD8α+CD8β- (αβTCR+ CD8αα); and (v) αβTCR+ CD8α+CD8β+
(αβTCR+CD8αβ).
Studies of Rag1-/-mice injected with bone marrow from nude mice or peripheral lymph
node T cells from euthymic mice, demonstrated that generation of αβTCR+ CD4+ and CD8+
IELs is thymus-dependent, while γδTCR+ CD8αα+ IELs appeared in the absence of a thymus
(9). One site of extra-thymic maturation may be the crypts of Lieberkuhn. These distinct mucosal
invaginations surround the base of each villus and contain long-lived multipotent stem cells (10)
that gives rise to the four epithelial lineages of the small intestine - enterocytes, goblet and
enteroendocrine cells which differentiate as they migrate from the crypt up adjacent villi, and
Paneth cells which differentiate and remain at the crypt base (11-15). Crypts possess structures
(cryptopatches) that contain clusters of c-kit+ interleukin-7 receptor (IL-7R)+ Thy1+ lymphocytes
(16). Mice with a truncated mutation of the common cytokine receptor chain (17) lack these
cryptopatches, do not have γδTCR+CD8αα+ IELs, but contain thymus-dependent αβTCR+ CD4+
and αβTCR+ CD8+ IELs, suggesting a role for cryptopatches in maturation of extrathymically-
derived γδTCR+ IELs (18-20).
The epithelium also appears to play a direct role in regulating IEL development.
Epithelial cells produce stem cell factor (SCF) (21), a ligand for the c-kit receptor expressed on
the surface of γδTCR+ IELs (22). Mice deficient in either SCF or c-kit have reduced numbers of
γδTCR+ IELs (22). Furthermore, thyrotropin-releasing hormone stimulation of enterocytes
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

5
results in local release of thyroid stimulating hormone (TSH) which interacts with IEL-based
TSH-R to promote IEL development (23): e.g., hyt/hyt mice, which have a loss-of-function TSH-
R mutation, have disrupted IEL maturation (24, 25).
Epithelial-based IL-7 provides another regulatory signal for IEL proliferation (26).
Studies of mice that lack IL-7 or the IL-7R have demonstrated that IL-7R-mediated signaling is
essential for γδ TCR+ IEL development (26, 27). Moreover, Laky and coworkers (28) used
transcriptional regulatory elements from the rat intestinal fatty acid binding protein (Fabpi) to
express IL-7 in villus enterocytes of Il-7-/- mice. γδ TCR+ IELs were restored in the intestinal
epithelium but remained absent from all other tissues, indicating that local production of IL-7
was sufficient for proper development/survival of this IEL subset.
Interactions between intestinal epithelial cells and IELs are reciprocal: IELs can influence
epithelial cell biology. One illustration of this reciprocity is provided by TCRδ− subunit-deficient
mice. These animals have reduced numbers of dividing cells in their crypts of Lieberkuhn,
reduced crypt cellularity (29), and exhibit more severe intestinal epithelial damage following
infection with the parasite Eimeria vermiformis (30). γδ TCR+ IELs produce keratinocyte growth
factor (KGF) which affects epithelial cell growth and repair (31). These findings raise the
question of whether γδTCR+ IELs form part of a homeostatic surveillance mechanism that can
detect and respond to perturbations in intestinal epithelial proliferation in order to maintain
steady state cellular census in crypts and their associated villi.
Some workers have proposed that IELs are key elements in a ‘mucosal intranet’ where
they function to control epithelial integrity and immunologic homeostasis (32). Recent
comparative DNA microarray-based studies of gene expression in γδTCR+ IELs harvested from
the small intestine of conventionally raised adult C57Bl6/J mice and αβ+ T-cells harvested from
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

6
their mesenteric lymph nodes have provided a list of candidate factors, preferentially expressed
by γδ TCR+ IELs, that may support this mucosal intranet (33, 34).
In the present study, we examine the cross-talk between IELs and epithelium using
transgenic mice that express Simian Virus 40 large T Antigen (TAgWt) in their villus-associated
enterocytes. The rationale for our experimental approach was as follows. Fabpi-directed
expression of TAgWt produces a proliferative abnormality restricted to villus enterocytes: Fabpi-
reporter transgenes are not expressed in the IELs. Expression of the viral oncoprotein in post-
mitotic enterocytes induces their re-entry into the cell cycle (35), an associated p53-independent
apoptosis (36), and not accompanied by evidence of dysplasia during the 1-2 d interval that they
take to complete their migration to the cellular extrusion zone located at the villus tip (36, 37).
Fabpi-directed expression of a mutant TAg containing Glu→Lys substitution at residues 107 and
108 (TAgK107/8) disrupts pRB binding, but does not produce this proliferative abnormality. Thus,
a three way comparison of FVB/N Fabpi-TAgWt and Fabpi-TAgK107/8 transgenic mice, and their
age-matched nontransgenic littermates would allow direct assessment of whether a proliferative
abnormality limited to the predominant intestinal epithelial lineage is accompanied by changes in
the fractional representation of extrathymically-educated or thymically-derived IEL subsets. By
performing this analysis in conventionally raised and germ-free mice, we could also determine
whether the microflora contributed to any observed changes in IELs. Finally, by using laser
capture microdissection (LCM) of small intestinal cryosections to harvest villus epithelium, flow
cytometry to retrieve their IELs, the DNA microarray-based datasets of IEL gene expression to
direct quantitative RT-PCR measurements of the levels of specified mRNAs in each cell
population, we could use this environmentally well-defined system to identify enterocytic gene
products affected by proliferative status that may impact on IEL development/survival.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

7
Our results show that the engineered proliferative abnormality is accompanied by a
microflora-independent reduction in extra-thymically educated γδ TCR+CD8αα+ IELs. This
change is accompanied by coordinate changes in the expression of enterocytic and γδ TCR+IEL
gene products that likely help legislate the observed change in IEL representation.
Experimental Procedures
Generation and maintenance of conventionally-raised and germ-free transgenic
mice - FVB/N mice hemizygous for a transgene containing nucleotides -1178 to +28 of rat Fabpi
linked to TAgWt or TAgK107/8 are described in earlier reports (35, 36, 38). Conventionally raised
animals were maintained in microisolators in a specified pathogen free state.
Normal and transgenic mice were re-derived as germ-free by Caesarian section of
transgenic mothers and transfer of their E19 fetuses to plastic gnotobiotic isolators (Standard
Safety Equipment Co.) containing germ-free foster mothers. The protocol used for this re-
derivation is described in a recent publication (6). Both conventionally raised and germ-free mice
were given sterilized BeeKay Autoclavable Diet (B&K Universal Inc.) ad libitum. All animals
were maintained under a strict light cycle (lights on at 0600h and off at 1800h). Animals were
genotyped using primers, tail DNA and PCR conditions described in ref. 36. Some mice received
an intraperitoneal injection of an aqueous solution of 5-bromo-2’-deoxyuridine (BrdU, 120
mg/kg) and 5'-fluoro-2'-deoxyuridine (12 mg/kg) (Sigma), 90 min prior to sacrifice. Only male
mice were studied.
Quantitative immunohistochemical analysis of the IEL subsets - FVB/N transgenic
mice and their wild type littermates were sacrificed at 6-8 weeks of age (n = 3 conventionally
raised or germ-free animals/genotype/experiment; n = 3 independent experiments). The middle
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

8
third of their small intestine (arbitrarily defined as jejunum) was immediately flushed with PBS,
and subdivided into 5 equal length segments. All were segments placed together in a tissue
cassette, overlaid with OCT (Miles Scientific) and frozen in Cytocool (Stephens Scientific). 100
serial 5µm thick sections were cut parallel to the cephalocaudal axes of the segments. For each
antibody surveyed, every 10th section was fixed for 20 min in methanol (at -20oC), washed 3
times in PBS (3 min/cycle), and treated with PBS-blocking buffer (1% bovine serum albumin,
0.05% Triton X-100 in PBS) for 2h at room temperature. Sections were subsequently treated
three times with TNT wash buffer (0.1 M Tris, pH 7.5/0.15M NaCl/0.05%Tween-20; 3 cycles; 5
min/cycle), and then incubated overnight at 4oC with each of the following monoclonal
antibodies [all from Pharmingen, each diluted 1:1000 in TNB-blocking buffer (0.1M Tris (pH
7.5), 0.15M NaCl, and 0.5% blocking reagent from Perkin Elmer)]: (i) rat anti-mouse CD4
(clone H129.19); (ii) rat anti-mouse CD8α (clone 53-6.7); (iii) rat anti-mouse CD8β (clone Ly-
32); (iv) hamster anti-mouse αβ TCR (β chain; clone H57-597); (v) hamster anti-mouse
γδ TCR (δ chain; clone GL3); and (vi) hamster anti-mouse CD103 (integrin αIEL chain; clone
2E7).
Following incubation with these primary antibodies, sections were washed in TNT buffer
(3 cycles, each 5 min). Biotin-conjugated mouse anti-rat IgG1/IgG2a (PharMingen), or biotin-
conjugated mouse anti-hamster IgG cocktail (PharMingen) was added (final dilution of each =
1:100 in TNB blocking buffer). After a 30 min incubation with the secondary antibodies at room
temperature, sections were treated 3 times with TNT wash buffer (5 min/wash cycle). The
sections were then incubated for 30 min at room temperature with streptavidin-horseradish
peroxidase (Perkin Elmer; 1:1000 in TNB) followed by 3 washes of 5 min each in TNT buffer.
The final steps consisted of (i) adding biotinyl-tyramide (Perkin Elmer Lifesciences; diluted
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

9
1:100 in 1X amplification diluent from the same manufacturer) for 10 min; (ii) washing three
times with TNT buffer (5 min/cycle); (iii) incubating the section with Cy3-conjugated
streptavidin (Perkin Elmer Lifesciences; diluted 1:500 in TNB) for 30 min, and (iv) performing 3
final rinses in TNT buffer. Two controls were performed to verify the specificity of the signals
produced: (i) direct amplification of endogenous peroxidase activity alone without addition of
primary or secondary antibodies but with addition of biotinyl-tyramide; (ii) direct amplification
of endogenous peroxidase activity followed by omission of each primary antibody but with
inclusion of all other steps and reagents.
Only well-oriented jejunal crypt-villus units were scored. ‘Well-oriented’ was defined as
sectioned parallel to the crypt-villus axis with an unbroken epithelial column extending from the
crypt base to the villus tip. The data for each unit was compiled as the number of IELs of a
particular type per 1000 villus epithelial cells, or per 100 crypt epithelial cells. A minimum of
100 jejunal crypt-villus units were scored per mouse. Data obtained with each antibody from all
mice of a given genotype (germ-free or conventional) were averaged and a standard deviation
calculated.
Multilabel immunohistochemical studies were performed on sections of normal and
transgenic jejunums using rabbit anti-TAg (1:1000 in PBS-blocking buffer; kindly provided by
Doug Hanahan, University of California, San Francisco, CA), and goat anti-BrdU (1:1000; refs.
38, 39). Antigen-antibody complexes were detected with indocarbocyanine (Cy3)-labeled
donkey anti-rabbit and fluorescein isothiocyanate (FITC)-labeled donkey anti-goat (1:500;
Jackson ImmunoResearch).
FACS analysis of IELs - Six to eight week old transgenic mice and their normal
littermates were sacrificed and their jejunums recovered (n =3 germ-free and 3 conventionally-
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

10
raised mice/genotype/experiment; 3 independent experiments). Peyer’s patches were identified
by inspecting the serosal surfaces of the jejunal segment and were then excised. Each jejunal
segment was subsequently opened with a longitudinal incision, washed in PBS, and cut into 1 cm
pieces that were placed in 40 ml of ice cold sterile PBS. The pooled segments from all three
animals/genotype/experiment were washed 5 times in PBS (vigorous shaking), allowed to settle
by gravity, and resuspended in 25 ml of R2 medium [RPMI 1640 buffer containing 5 % fetal calf
serum (Sigma), 1 mM sodium pyruvate, 1 mM sodium bicarbonate, 1% non-essential amino
acids (Sigma), and 0.1% 2-mercaptoethanol]. The mixture was shaken gently for 30 min at 37oC
and then rigorously for 2 min at room temperature. The intestinal segments were allowed to
settle by gravity, and the supernatant was collected and passed through a Nytex filter (Becton
Dickinson). The flow-through, containing IELs and epithelial cells, was passed over a column of
dimethyldichlorosilane-treated glass wool fiber (0.5g/10 ml syringe) pre-equilibrated in R2
medium. The flow-thru was spun at 1500 x g for 5 min and the resulting cell pellet, highly
enriched for IELs, was resuspended in 10 ml of R2 medium. The suspension was centrifuged at
1500 x g for 5 min, and the pellet resuspended to a final concentration of 107 cells/ml of FACS
staining buffer [RPMI, 1% bovine serum albumin (Sigma), 1mg/ml human IgG (Sigma)].
IELs were stained with the following antibodies in various combinations (all from
Pharmingen; all diluted 1:100 in FACS staining buffer): (i) phycoerythrin (PE)-conjugated
hamster anti-mouse αβ TCR (β chain; clone H57-597); (ii) PE-conjugated hamster anti-mouse
γδ TCR (δ chain; clone GL3); (iii) PE-conjugated rat anti-mouse CD8β.2 (clone 53-5.8); (iv)
FITC-conjugated rat anti-mouse CD8α (clone 53-6.7); (v) FITC- or PE-conjugated rat anti-
mouse CD4 (clone RM4-5); (vi) FITC-conjugated rat anti-mouse CD45 (clone 30-F11); and (vii)
biotinylated hamster anti-mouse CD103 (integrin αIEL chain; clone 2E7). Biotinylated primary
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

11
antibodies were visualized with FITC-streptavidin or PE-streptavidin (PharMingen). Idiotype as
well as secondary antibody alone controls were also performed. Following incubation with these
reagents (60-90 min on ice), cells were spun for 5 min at 1500x g, washed with sterile ice cold
PBS, and examined by flow cytometry (FACScalibur, Becton Dickinson).
Isolation of RNA from γγγγδδδδTCR+ IELs - γδ TCR+ IELs were isolated from jejunal
segments that had been recovered from 6 week old germ-free male transgenic mice and their
normal littermates. The γδ TCR+, CD103+ lymphocyte population was sorted (FACS Vantage,
Becton Dickinson), collected in sterile cold PBS, and recovered by centrifugation (1000 x g for 5
min at room temperature). RNA was isolated using the RNAeasy kit (Qiagen) (5 mice/IEL
preparation; n = 10 preparations/genotype). RNA was also isolated from intact jejunal segments
(n =10 germ-free mice/genotype).
Laser-capture microdissection (LCM) of jejunal villus epithelium - LCM was
conducted using jejunal cryosections that had been stained briefly with eosin Y and methyl
green. Dissection of villus epithelium was restricted to well-oriented crypt-villus units and was
accomplished using the PixCell II system (Arcturus, 7.5-µm-diameter laser spot), CapSure HS
LCM Caps (Arcturus), and protocols described in ref. 40. ~10,000 jejunal villus epithelial cells
were recovered from each germ-free normal and TAg mouse (n=3 animals/group). RNA was
prepared from captured cells from each mouse in each group using the PicoPure RNA Isolation
Kit (Arcturus). The concentration of each preparation was defined (RiboGreen RNA quantitation
kit; Molecular Probes) and equal size aliquots from members of a group of animals pooled.
Analysis of previously published DNA microarray datasets - Datasets of gene
expression profiles from γδTCR+ IELs and the αβ T-cells were a generous gift from Aude Fahrer
and Y-H Chien (Dept. Microbiology and Immunology, Stanford University) (33). These datasets
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

12
were obtained using an early manufactured version of a high density, oligonucleotide-based
DNA microarray containing probesets representing 6,352 mouse genes or EST clusters (Mu6K
GeneChip, Affymetrix).
We used GeneChip software (v4.0; Affymetrix) to compute an average fluorescence
intensity across all probe sets on the GeneChips prior to conducting pairwise chip-to-chip
comparisons (41) of γδ TCR+ IEL versus αβ T-cell transcript levels (the αβ T-cell GeneChip
was designated as ‘Baseline’). We then extracted all mRNAs fulfilling the following selection
criteria: (a) called ‘Present’ in either the baseline or partner chip; (b) ≥2-fold difference in
transcript level in the two RNA populations (increased or decreased); and (c) the increase or
decrease reproduced in duplicate GeneChip comparisons (cRNAs were independently generated
from the same starting RNAs and each preparation used to interrogate a separate Mu6K
GeneChip).
SYBR Green-based real time quantitative PCR (qRT-PCR) - qRT-PCR was used to
examine changes in levels of selected mRNAs in RNAs prepared from the intact jejunums, LCM
villus epithelium and/or sorted γδ TCR+ IELs harvested from 6-8 week old male germ-free
normal and TAgWt mice. cDNAs were generated from each pooled RNA preparation (see above)
using reagents and protocols described in ref. 40. cDNA was added to 25 µl qRT-PCR reactions
containing 12.5 µl of 2X SYBR Green master mix (Applied Biosystems), 900 nM gene-specific
primers (see Table in on-line supplemental material) and 0.25 units UDP-N-glycosidase (Life
Technologies). A melting curve was used to identify a temperature where only the amplicon, and
not primer dimers, accounted for the SYBR Green-bound fluorescence (6). Assays were
performed in triplicate with an ABI Prism 7700 Sequence Detector (Applied Biosystems). All
data were normalized to an internal standard (glyceraldehyde 3-phosphate dehydrogenase
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

13
(Gapdh) mRNA; ∆∆CT method, User Bulletin #2 Applied Biosystems). For the γδ ΤCR+ IEL
mRNA analysis, 18S rRNA was used as the internal standard.
Results and Discussion
Forced expression of TAgWt in villus enterocytes causes a change in the representation of
IEL subsets within the small intestinal epithelium
As noted in the Introduction, transcriptional regulatory elements from the Fabpi gene
were used to direct expression of TAgWt in small intestinal villus enterocytes of adult FVB/N
transgenic mice (Fig. 1A). There was no detectable TAgWt in the crypt epithelium, the
mesenchyme underlying crypt-villus units (Fig 1A), the organized gut-associated lymphoid
tissue (Peyer’s patch lymphocytes plus smaller submucosal lymphoid aggregates), or in the
spleen and thymus (data not shown). Other than villus enterocytes, the only other site of
transgene expression was the follicle-associated epithelium (FAE) overlying Peyer’s patches
(Fig. 1B). An identical pattern of transgene expression was observed in FVB/N mice from the
reference control pedigree containing Fabpi-TAgK107/108 (data not shown).
Age-matched 6-8 week old Fabpi-TAgWt and Fabpi-TAgK107/108 male mice, as well as
their nontransgenic littermates, were given an intraperitoneal injection of BrdU, 1.5h prior to
sacrifice (n = 2-3 mice/genotype). Expression of the wild type viral oncoprotein induced villus
enterocytes to re-enter the cell cycle (Fig. 1A). In contrast, the jejunal villus epithelium and FAE
were not labeled with BrdU in either wild type or Fabpi-TAgK107/108 mice (data not shown).
To determine whether the proliferative abnormality induced by TAgWt caused a change in
the composition or spatial organization of IELs, these cells were isolated from the jejunal
epithelium of each group of conventionally-raised mice and subjected to flow cytometry (n = 3
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

14
mice/genotype/experiment; n = 3 independent experiments). There were no statistically
significant differences in the purity of the lymphocyte preparations from each group of mice:
>80% of the gated lymphocytes expressed the IEL-specific marker, CD103 (Fig. 2A). The total
yield of lymphocytes was similar in each group (5-7 x 107).
The majority of the IELs were also positive for CD45, a pan-lymphocyte marker (Fig
2B). However, there was a statistically significant increase in the fractional representation of
αβ TCR+ IELs in Fabpi-TAgWt mice compared to their normal littermate controls (p<0.05;
Student’s t-test) and a statistically significant decrease in γδTCR+ IELs (p<0.05) (Fig. 2C,D). In
contrast, there were no differences in the percentages of these IEL subsets in Fabpi-TAgK107/8
versus normal animals (Fig. 2C,D).
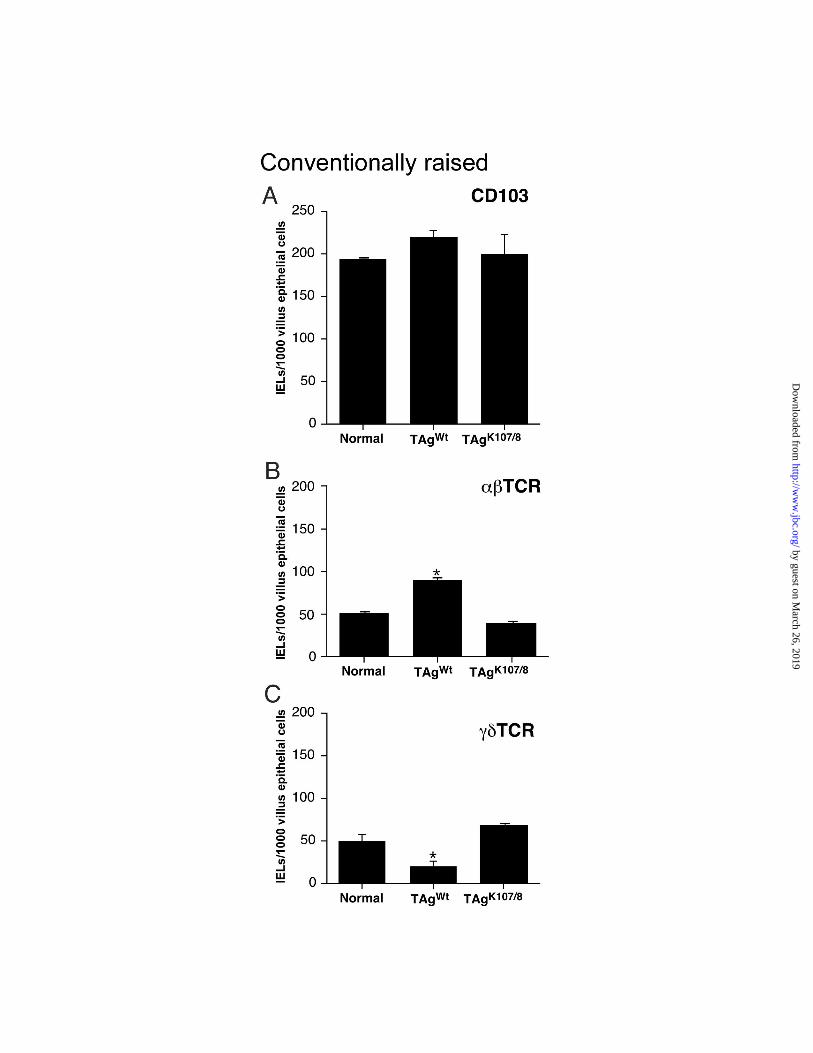
We performed a quantitative immunohistochemical analysis of jejunal crypt-villus units
to determine whether the change in αβ ΤCR+ and γδ TCR+ IEL representation in Fabpi-TAgWt
mice were restricted to the villus epithelium where the proliferative abnormality was evident, or
whether the change extended to the crypt epithelium, where there was no change in proliferative
status. Surveys of sections of jejunum indicated that there were no significant differences in the
total number of CD103+ IELs per 1000 villus epithelial cells between age matched Fabpi-TAgWt,
Fabpi-TAgK107/108, and normal FVB/N mice (Fig. 3A). However, there was a significant increase
in the density of αβ TCR+ IELs, and a significant reduction in γδ TCR+ IELs in TAgWt mice
compared to the other two groups (p<0.05) (Fig. 3B,C).
In the crypt epithelium of conventionally raised normal male 6-8 week old FVB/N mice,
the densities of CD103+, αβ TCR+, and γδ TCR+ lymphocytes are 10±1, 5±1, and 4±1 per 100
epithelial cells, respectively. There were no statistically significant differences in the numbers of
these cells among the three groups of mice, indicating that the proliferative abnormality
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

15
produced by TAgWt had a ‘local’ effect on villus IELs that did not extend to the crypt.
The increase in ααααββββ TCR+ and decrease in γγγγδδδδ TCR+ IELs observed in conventionally raised
TAgWt transgenics is recapitulated in germ-free mice
One question raised by these finding is whether the intestinal microflora was exerting an
influence on the composition of the villus IEL population: e.g., from a potential epithelial barrier
disruption associated with the engineered proliferative abnormality, or as a direct consequence of
a cross-talk between components of the microbiota and the epithelium. To address this question,
we re-derived our pedigrees of Fabpi-TAgWt and Fabpi-TAgK107/108 transgenic mice, and their
normal littermates, as germ-free. The cellular patterns of expression of TAgWt and TAgK107/108
were not affected when the microflora was removed. An epithelial proliferative abnormality
extending from the base to the tips of the villi was evident in 6-8 week old germ-free FVB/N
Fabpi- TAgWt but not in Fabpi-TAgK107/108 or normal animals (Fig. 4A plus data not shown).
Quantitative immunohistochemical surveys also disclosed that Fabpi-TAgWt mice, like
their conventionally raised counterparts, had a reduction in the density of their villus γδ TCR+
IELs when compared to age- and gender-matched FVB/N Fabpi-TAgK107/108 or normal mice
(p<0.05; Fig. 4B-D) (n = 3 mice assayed/genotype). There was also a modest increase in
αβTCR+ IELs associated with the TAgWt-induced proliferative abnormality, although the
differences were not statistically significant compared to the other two groups of mice (Fig. 4E).
These quantitative immunohistological studies indicated that the density of all IELs (i.e.,
CD103+ cells) in the jejunal villus epithelium was similar in all three groups of germ-free mice
(Fig. 3F) but several-fold less than in conventionally raised animals (compare Figs. 3A and 4F).
As in conventionally raised mice, production of TAgWt in the villus epithelium did not result in
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

16
any changes in the number of crypt-associated CD103+, αβ TCR+, or γδ TCR+ IELs (data not
shown).
FACS analysis of germ-free jejunal IELs confirmed the results of our quantitative
immunohistologic survey, and established that TAgWt expression produced a statistically
significant increase in αβ TCR+, and a significant decrease in γδ TCR+ IELs (p<0.05 in
comparison to age matched normal or TAgK107/108 mice; n = 3 mice/genotype/experiment; 3
independent experiments) (Fig. 5A-C).
Based on these findings, we concluded that the alterations in these two IEL populations
occurred independently of the microflora, and were ascribable to the proliferative effects of
TAgWt rather than to other functions mediated by regions of the viral oncoprotein located outside
of its pRB pocket protein binding domain.
Expression of TAgWt leads to a decrease in accumulation of intestinally-derived
γγγγδδδδTCR+CD8αααααααα IELs and an increase in thymically-derived ααααββββTCR+CD8ααααββββ IELs
As noted in the Introduction, intestinal IELs are derived from two sources. The vast
majority of αβTCR+CD4+ and αβTCR+CD8αβ+ IELs are thymically-derived while all CD8αα+
cells, whether they express αβTCR or γδTCR, are derived from extra-thymic sites (42). The
phenotype produced by TAgWt-induced proliferation of villus enterocytes in germ-free mice
could reflect subtle disruptions of epithelial barrier function with resulting presentation of non-
microbial luminal antigens (e.g., from the diet) to components of the gut-associated lymphoid
tissue. If this were the case, one would expect an increased influx of thymically-derived, antigen-
induced αβTCR+ IELs.
We addressed this hypothesis in two ways. First, germ-free Fabpi-TAgWt mice and their
normal littermates were given an intraperitoneal injection of BrdU 1.5h prior to sacrifice to label
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

17
intestinal epithelial cells in S-phase. Sections of jejunal crypt-villus units were then stained with
antibodies to BrdU and E-cadherin, the principal epithelial cadherin and an important regulator
of cell adhesion in this system (43, 44). Expression of TAgWt and/or entry of jejunal enterocytes
into the cell cycle produced no detectable changes in the steady state cellular levels or
intracellular compartmentalization of E-cadherin (data not shown; n = 2 germ-free
mice/genotype). Second, FACS analysis of jejunal IELs demonstrated that the TAgWt-associated
increase in αβTCR+ IELs involved both the CD4 and CD8αβ subsets (Fig. 5D,E). There were no
changes in the CD8αα subtype of αβTCR IELs (data not shown). These findings confirm that
the proliferative abnormality engineered in enterocytes is associated with an influx of thymically
derived IELs.
FACS analysis also established that expression of TAgWt, but not TAgK107/108, in germ-
free villus epithelium leads to a significant (p<0.05) reduction in intestinally-derived γδTCR+
CD8αα IELs compared to normal littermate controls (Fig. 5F). Immunostaining of intestinally-
derived γδ TCR+ CD8αα IELs and thymically-derived αβ TCR+ CD4+ and CD8αβ subsets
obtained by flow cytometry revealed that they did not contain detectable levels of TAg (data not
shown plus see below).
Taken together, these findings demonstrate that TAgWt-dependent re-entry of villus
enterocytes into the cell cycle produces a specific decrease in the γδ TCR+ CD8αα IEL
populations that normally develop in the intestine.
qRT-PCR analysis of TAgWt-dependent regulation of IL-7 expression
Previous studies have established that the majority of intestinal IELs are maintained in G0
of the cell cycle (45). In addition, some reports have suggested that epithelial cells may act as
antigen presenting cells for induction and activation of these resting IELs (46, 47). Thus, the
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

18
proliferative abnormality produced by TAgWt could result in suppression of critical trophic
factors necessary for the appropriate development and activation of γδ TCR+ CD8αα IELs
leading to their diminution in the epithelium.
IL-7 is a trophic factor known to be produced by the epithelium and required for
generation of mature γδ TCR+ IELs (see Introduction). γδ TCR+ IELs express the receptor for
this cytokine, IL-7R (48). We tested the hypothesis that TAgWt-induced re-entry of villus
enterocytes into the cell cycle is accompanied by reduced IL-7 expression by performing SYBR-
Green-based real time quantitative RT-PCR (qRT-PCR) analysis of total cellular RNAs isolated
from intact jejunum as well as laser capture microdissected (LCM) jejunal villus epithelium (Fig.
6A). The results revealed a 12-fold lower steady state concentration of IL-7 mRNA in the intact
jejunum of germ-free Fabpi-TAgWt mice compared to germ-free normal littermates, and a 4-fold
reduction in levels in their LCM villus epithelium (Fig. 6B). Control qRT-PCR assays of LCM
epithelial RNA documented a 2-fold reduction in TCRδ mRNA (Fig. 6C) - consistent with the
reduced representation of γδTCR+ IELs in transgenic mouse jejunum documented by quantitative
immunohistochemical and flow cytometry analyses (Figs. 4D,5F).
To address the question of whether the TAgWt-induced proliferative abnormality in villus
enterocytes produced changes in γδ TCR+ IEL gene expression, we purified these cells, using
flow cytometry, from the jejunums of 6-8 week old germ-free male Fabpi-TAgWt and normal
mice (n = 50 mice/group). qRT-PCR studies indicated that the IELs from transgenic mice did not
contain detectable levels of TAg mRNA – in agreement with the results of our
immunohistochemical studies (see above). IL-7Rα mRNA levels were significantly decreased in
γδ TCR+ IELs from transgenic compared to normal mice (5.5 fold; p<0.05; Fig. 7).
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

19
Fujihashi and coworkers used IL-7 knockout mice to show that IL-7 signaling is
necessary for IL-7R expression in γδ TCR+ IELs and for their subsequent activation and cell
division (24). Therefore, we concluded that TAgWt expression in villus enterocytes results in
decreased epithelial expression of IL-7, leading to a concomitant decrease in expression of the
IL-7 receptor in γδ TCR+ IELs, and impeded intestinal development of γδ TCR+ CD8αα IELs.
TAgWt expression in enterocytes is associated with reduced levels of other γγγγδδδδ TCR++++ IEL-
derived factors that may affect IEL:epithelial cross-talk
Analysis of published DNA microarray-based expression profiles of γδ TCR+ IELs
purified from conventionally raised C57Bl/6 mice (33) allowed us to identify factors whose
expression is enriched in γδ TCR+ IEL relative to αβ T-cells, and that may affect epithelial
barrier functions and/or important interactions between the epithelium and its population of
IELs.
TGFβ3 - Recent reports have shown that IL-7 regulates TGFβ3 production in fibroblasts
(49). Increased expression of TGFβ3 leads to enhanced intestinal epithelial cell migration across
wound edges in an in vitro model. Neutralizing antibodies to TGFβ3 inhibit this process (50).
TGFβ3 also functions as a signaling factor that induces apoptotic cell death during involution of
the mammary epithelium (51), suggesting that it may help regulate epithelial cell census.
qRT-PCR studies disclosed a 10-fold decrease in the steady state level of TGFβ3 mRNA
in LCM villus epithelium from germ-free Fabpi-TAgWt compared to germ-free normal
littermates (Fig. 6C). Furthermore, enterocytic expression of TAgWt is associated with a 262-fold
decrease in the concentration of γδTCR+ IEL TGFβ3 mRNA (Fig. 7). Together, these results
indicate that one consequence of reduced enterocytic IL-7 expression is reduced γδTCR+ IEL-
derived TGFβ3. Loss of TGFβ3 may alter the integrity of the epithelial barrier, contribute to the
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

20
observed influx of thymically-derived αβ TCR+ IELs, or help regulate the extent of the p53-
independent apoptotic response that occurs in villus enterocytes undergoing unscheduled,
TAgWt-induced re-entry into the cell cycle.
Flt3L – The DNA microarray studies revealed that the mRNA encoding the ligand for
fms-like tyrosine kinase-3 (Flt3) is enriched in γδ TCR+ IELs compared to αβ T-cells (33). The
function of Flt3L in γδ TCR+ IELs is not known. Flt3 was initially identified in hematopoietic
stem cells (52). It is member of the class III receptor tyrosine kinases that includes c-kit. Flt3
ligand stimulates proliferation of quiescent as well as cytokine-stimulated hematopoietic
progenitors (e.g., refs. 53-55). However, this proliferative response is not shared by other
progenitors: Flt3 ligand inhibits EGF- and FGF2-stimulated division of neuronal stem cells (56).
There is very little information about the regulation of expression of Flt3 ligand and its receptor,
or their functions, in epithelia. One report indicated that Flt3 mRNA is present in mouse bile duct
epithelium (57) while another identified the transcript in dividing neuroepithelial cells (56).
Our LCM/qRT-PCR studies revealed that the receptor is expressed in normal jejunal
villus epithelium. Moreover, expression is down-regulated by the engineered proliferative
abnormality: mRNA levels are reduced 7.5-fold in LCM TAgWt compared to non-transgenic
epithelium (Fig. 6C).
The qRT-PCR/LCM analysis indicated that the mRNA encoding Flt3 ligand is also
reduced in TAgWt epithelium (Fig. 6C). qRT-PCR assays disclosed that TAgWt expression in
enterocytes does not have a discernible effect on IEL Flt3 ligand expression (Fig. 7). Since the
extent of the reduction in Flt3 ligand mRNA in TAgWt epithelium was several fold greater than
the reduction of γδTCR+ IEL number (5- versus 2-fold), and since IEL Flt3 ligand expression is
unaffected by enterocytic TAgWt expression, we concluded that the proliferative abnormality
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

21
reduces epithelial expression of the ligand. The response of Flt3 and its ligand to changes in the
proliferative status of enterocytes raises the possibility that signaling through this system may
normally serve to help suppress cell division as members of this lineage execute their terminal
differentiation program.
Prospectus
These studies reveal that an engineered proliferative abnormality in post-mitotic
enterocytes impedes intestinal development of γδ TCR+ CD8αα IELs, and promotes
accumulation of thymically-educated CD4 and CD8αβ subsets of αβ TCR+ IELs. Our findings
highlight the interdependent contributions of enterocytes and γδ TCR+ IELs to intestinal mucosal
biology – a point illustrated by the diminution in enterocytic IL-7 expression associated with
TAgWt production. The resulting diminution in intestinal maturation of γδ TCR+ IELs ‘robs’ the
epithelium of IEL-derived factors known or postulated to support epithelial barrier function (e.g.,
TGFβ3). Gnotobiotic FVB/N Fabpi-TAgWt mice provide an environmentally and genetically
defined, ‘sensitized’ model system for genetic or pharmacologic tests of the role of enterocyte-
derived factors postulated to promote maturation of γδ TCR+ IELs, of IEL-derived factors that
may affect epithelial barrier function, and/or of microbes or microbially-derived products that
may influence mucosal biology.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

22
Acknowledgements
We are indebted to Aude Fahrer and coworkers for generously providing us with the
Mu6K GeneChip datasets of IEL gene expression, our colleague Jason Mills for his help in
analyzing the datasets, and David O’ Donnell and Maria Karlsson for their superb technical
assistance in generating and maintaining the germ-free mice used in this study.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

23
References
1. Falk, P. G., Hooper, L. V., Midtvedt, T., and Gordon, J. I. (1998) Microbiol Mol Biol Rev
62, 1157-1170.
2. Hooper, L. V., and Gordon, J. I. (2001) Science 292, 1115-1118.
3. McCracken, V. J., and Lorenz, R. G. (2001) Cell Microbiol 3, 1-11.
4. Gordon, J. I., Hooper, L. V., McNevin, M. S., Wong, M., and Bry, L. (1997) Am J
Physiol 273, G565-570.
5. Hooper, L. V., Wong, M. H., Thelin, A., Hansson, L., Falk, P. G., and Gordon, J. I.
(2001) Science 291, 881-884.
6. Hooper, L.V., Mills, J. C., Roth, K.A., Stappenbeck, T.S., Wong, M. H., and Gordon, J. I.
(2002) Methods in Microbiology: Molecular Cellular Microbiology (P. J. Sansonetti, A.
Zychlinsky, Eds), Academic Press, London, 559-589.
7. Kaufmann, S. H. (1996) Proc Natl Acad Sci U S A 93, 2272-2279.
8. Beagley, K. W., and Husband, A. J. (1998) Crit Rev Immunol 18, 237-254.
9. Rocha, B., Vassalli, P., and Guy-Grand, D. (1992) Immunol Today 13, 449-454.
10. Wong, M. H., Saam, J. R., Stappenbeck, T. S., Rexer, C. H., and Gordon, J. I. (2000)
Proc Natl Acad Sci U S A 97, 12601-12606.
11. Cheng, H., and Leblond, C. P. (1974) Am J Anat 141, 537-561.
12. Cheng, H., and Leblond, C. P. (1974) Am J Anat 141, 461-479.
13. Cheng, H., and Leblond, C. P. (1974) Am J Anat 141, 503-519.
14. Nichols, D. B., Cheng, H., and Leblond, C. P. (1974) J Histochem Cytochem 22, 929-
944.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

24
15. Cheng, H., Merzel, J., and Leblond, C. P. (1969) Am J Anat 126, 507-525.
16. Kanamori, Y., Ishimaru, K., Nanno, M., Maki, K., Ikuta, K., Nariuchi, H., and Ishikawa,
H. (1996) J Exp Med 184, 1449-1459.
17. Ohbo, K., Asao, H., Kouro, T., Nakamura, M., Takaki, S., Kikuchi, Y., Hirokawa, K.,
Tominaga, A., Takatsu, K., and Sugamura, K. (1996) Int Immunol 8, 951-960.
18. Oida, T., Suzuki, K., Nanno, M., Kanamori, Y., Saito, H., Kubota, E., Kato, S., Itoh, M.,
Kaminogawa, S., and Ishikawa, H. (2000) J Immunol 164, 3616-3626.
19. Suzuki, K., Oida, T., Hamada, H., Hitotsumatsu, O., Watanabe, M., Hibi, T., Yamamoto,
H., Kubota, E., Kaminogawa, S., and Ishikawa, H. (2000) Immunity 13, 691-702.
20. Lambolez, F., Azogui, O., Joret, A. M., Garcia, C., von Boehmer, H., Di Santo, J., Ezine,
S., and Rocha, B. (2002) J Exp Med 195, 437-449.
21. Godfrey, D. I., Zlotnik, A., and Suda, T. (1992) J Immunol 149, 2281-2285.
22. Puddington, L., Olson, S., and Lefrancois, L. (1994) Immunity 1, 733-739.
23. Wang, J., Whetsell, M., and Klein, J. R. (1997) Science 275, 1937-1939.
24. Stein, S. A., Zakarija, M., McKenzie, J. M., Shanklin, D. R., Palnitkar, M. B., and
Adams, P. M. (1991) Thyroid 1, 257-266.
25. Stein, S. A., Oates, E. L., Hall, C. R., Grumbles, R. M., Fernandez, L. M., Taylor, N. A.,
Puett, D., and Jin, S. (1994) Mol Endocrinol 8, 129-138.
26. Fujihashi, K., McGhee, J. R., Yamamoto, M., Peschon, J. J., and Kiyono, H. (1997) Eur J
Immunol 27, 2133-2138.
27. Maki, K., Sunaga, S., Komagata, Y., Kodaira, Y., Mabuchi, A., Karasuyama, H.,
Yokomuro, K., Miyazaki, J. I., and Ikuta, K. (1996) Proc Natl Acad Sci U S A 93, 7172-
7177.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

25
28. Laky, K., Lefrancois, L., Lingenheld, E. G., Ishikawa, H., Lewis, J. M., Olson, S.,
Suzuki, K., Tigelaar, R. E., and Puddington, L. (2000) J Exp Med 191, 1569-1580.
29. Komano, H., Fujiura, Y., Kawaguchi, M., Matsumoto, S., Hashimoto, Y., Obana, S.,
Mombaerts, P., Tonegawa, S., Yamamoto, H., Itohara, S., and et al. (1995) Proc Natl
Acad Sci U S A 92, 6147-6151.
30. Roberts, S. J., Smith, A. L., West, A. B., Wen, L., Findly, R. C., Owen, M. J., and
Hayday, A. C. (1996) Proc Natl Acad Sci U S A 93, 11774-11779.
31. Boismenu, R., and Havran, W. L. (1994) Science 266, 1253-1255
32. Yamamoto, M., Fujihashi, K., Kawabata, K., McGhee, J. R., and Kiyono, H. (1998) J
Immunol 160, 2188-2196.
33. Fahrer, A. M., Konigshofer, Y., Kerr, E. M., Ghandour, G., Mack, D. H., Davis, M. M.,
and Chien, Y. H. (2001) Proc Natl Acad Sci U S A 98, 10261-10266.
34. Shires, J., Theodoridis, E., and Hayday, A. C. (2001) Immunity 15, 419-434.
35. Chandrasekaran, C., Coopersmith, C. M., and Gordon, J. I. (1996) J Biol Chem 271,
28414-2821.
36. Coopersmith, C. M., Chandrasekaran, C., McNevin, M. S., and Gordon, J. I. (1997) J
Cell Biol 138, 167-179.
37. Falk, P., Roth, K. A., and Gordon, J. I. (1994) Am J Physiol 266, G987-1003
38. Kim, S. H., Roth, K. A., Moser, A. R., and Gordon, J. I. (1993) J Cell Biol 123, 877-893.
39. Cohn, S. M., and Lieberman, M. W. (1984) J Biol Chem 259, 12456-12462.
40. Stappenbeck, T.S., Hooper, L.V., Manchester, J.K., Wong, M.H., and Gordon, J.I. (2002)
Methods in Enzymology, 31, 559-589.
41. Mills, J. C., and Gordon, J. I. (2001) Nucleic Acids Res 29, E72.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

26
42. Bandeira, A., Itohara, S., Bonneville, M., Burlen-Defranoux, O., Mota-Santos, T.,
Coutinho, A., and Tonegawa, S. (1991) Proc Natl Acad Sci U S A 88, 43-47.
43. Hermiston, M. L., and Gordon, J. I. (1995) Am J Physiol 268, G813-822.
44. Hermiston, M. L., and Gordon, J. I. (1995) Science 270, 1203-1207.
45. Yamamoto, M., Fujihashi, K., Beagley, K. W., McGhee, J. R., and Kiyono, H. (1993) J
Immunol 150, 106-114.
46. Kaiserlian, D., Vidal, K., and Revillard, J. P. (1989) Eur J Immunol 19, 1513-1516.
47. Hoyne, G. F., Callow, M. G., Kuo, M. C., and Thomas, W. R. (1993) Immunology 80,
204-208.
48. Moore, T. A., von Freeden-Jeffry, U., Murray, R., and Zlotnik, A. (1996) J Immunol 157,
2366-2373.
49. Huang, M., Sharma, S., Zhu, L. X., Keane, M. P., Luo, J., Zhang, L., Burdick, M. D., Lin,
Y. Q., Dohadwala, M., Gardner, B., Batra, R. K., Strieter, R. M., and Dubinett, S. M.
(2002) J Clin Invest 109, 931-937.
50. McKaig, B. C., Makh, S. S., Hawkey, C. J., Podolsky, D. K., and Mahida, Y. R. (1999)
Am J Physiol 276, G1087-1093.
51. Nguyen, A. V., and Pollard, J. W. (2000) Development 127, 3107-3118.
52. Rosnet, O., Marchetto, S., deLapeyriere, O., and Birnbaum, D. (1991) Oncogene 6, 1641-
1650.
53. Lyman, S. D., James, L., Vanden Bos, T., de Vries, P., Brasel, K., Gliniak, B.,
Hollingsworth, L. T., Picha, K. S., McKenna, H. J., Splett, R. R., et al. (1993) Cell 75,
1157-1167.
54. Lyman, S. D., and Jacobsen, S. E. (1998) Blood 91, 1101-1134.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

27
55. Shah, A. J., Smogorzewska, E. M., Hannum, C., and Crooks, G. M. (1996) Blood 87,
3563-3570.
56. Brazel, C. Y., Ducceschi, M. H., Pytowski, B., and Levison, S. W. (2001) Mol Cell
Neurosci 18, 381-393.
57. Omori, M., Omori, N., Evarts, R. P., Teramoto, T., and Thorgeirsson, S. S. (1997) Am J
Path 150, 1179-1187.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

28
Footnotes
*This work was supported in part by a grant from the National Institutes of Health (DK30292).
R.L.’s present address is Dept. of Pathology, University of Alabama at Birmingham,
Birmingham, AL 35233.
1Abbreviations used include: IEL, intraepithelial lymphocyte; TCR, T cell receptor; IL-7,
interleukin-7; IL-7R, interleukin-7 receptor; TAg, SV40 large T Antigen; TAgK107/8; mutant TAg
with Glu→Lys substitition at positions 107 and 108; FACS, fluorescence activated cell sorting;
LCM, laser capture microdissection; qRT-PCR real time quantitative reverse transcriptase-PCR;
FAE, follicle associated epithelium; Flt3, fms-like tyrosine kinase 3 receptor Flt3L, ligand for
fms-like tyrosine kinase 3 receptor.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

29
Figure legends
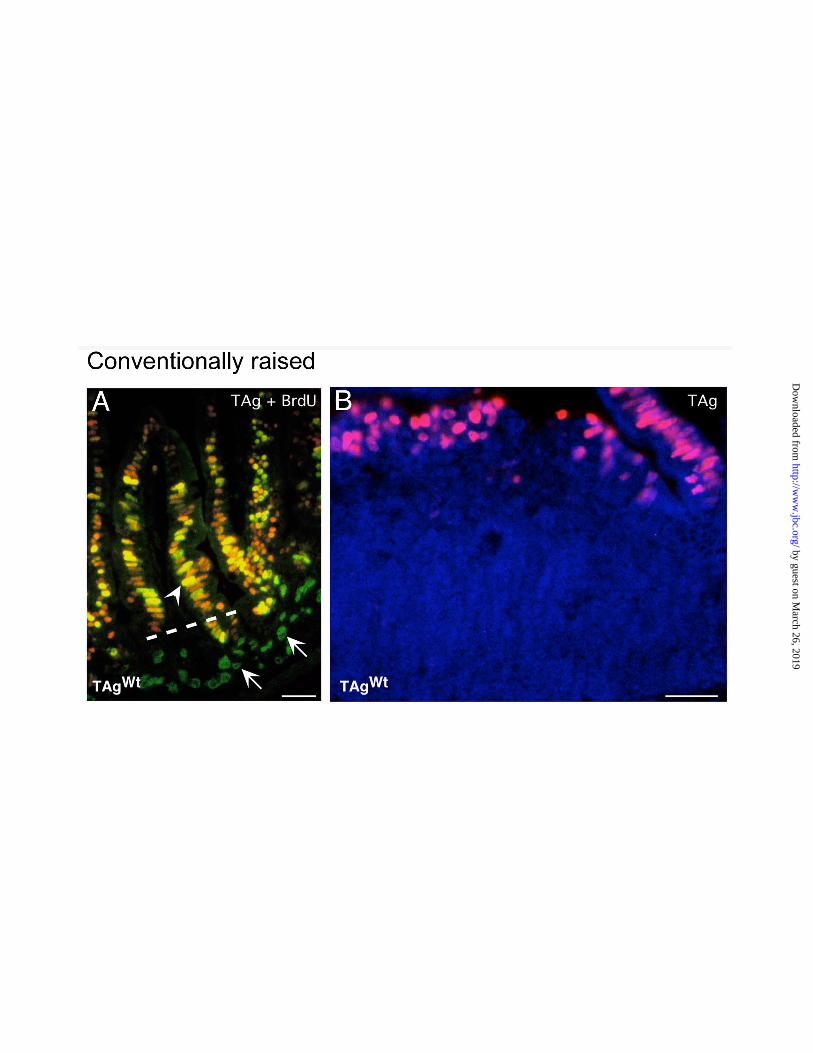
Figure 1. TAgWt expression in the jejunual villus epithelium of conventionally raised adult
FVB/N Fabpi-TAgWt transgenic mice. Multilabel immunohistochemical study of a 6 week old
mouse that had received an intraperitoneal injection of BrdU 90 min prior to sacrifice. (A)
Section of jejunum stained with rabbit antibodies to TAg, Cy3-labeled donkey anti-rabbit Ig,
goat anti-BrdU, and FITC-conjugated donkey anti-goat Ig. TAgWt-positive nuclei appear
red/orange. BrdU-positive nuclei appear green. Co-expression of TAg and BrdU produces
yellow staining of nuclei (e.g. arrowhead). TAg is not expressed in the crypt epithelium (all
nuclei are green; e.g., arrows). (B) TAgWt expression in the follicle-associated epithelium (FAE)
overlying Peyer’s patches. The section was incubated with antibodies to TAgWt and Cy3-donkey
anti-rabbit Ig, resulting in pink colored TAgWt-positive nuclei in the FAE. The lymphoid
population underlying the FAE does not express detectable levels of TAgWt: their nuclei appear
blue after counter-staining with bis-benzidine. Bars = 25 µm.
Figure 2. Expression of TAgWt results in an increase in ααααββββTCR+ and a decrease in γγγγδδδδ TCR+
IELs. IELs from the jejunums of conventionally raised 6-8 week old FVB/N Fabpi-TAgWt,
Fabpi-TAgK107/8, and normal mice were analyzed by flow cytometry (n=3
mice/group/experiment; n=3 experiments). Mean values ± S.E. are plotted. (A) Gated
lymphocytes positive for the IEL-specific marker, CD103. The results reveal no statistically
significant differences in the purity of the lymphocyte preparations between groups. (B) Sorted
IELs double positive for CD103 and CD45, a pan-lymphocyte marker. The total yield of
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

30
lymphocytes is similar in each group. (C) Results showing a statistically significant increase in
the percentage of αβTCR+ IELs in Fabpi-TAgWt mice (asterisk; p<0.05 relative to normal mice).
(D) Evidence for a statistically significant decrease in γδ TCR+ IELs in Fabpi-TAgWt
transgenics.
Figure 3. Quantitative immunohistochemical studies of ααααββββ TCR++++ and γγγγδδδδ TCR++++ IELs in
conventionally raised transgenic and normal mice. Sections, prepared from the jejunums of 6-
8 week old male Fabpi-TAgWt, Fabpi-TAgK107/108 and normal FVB/N animals, were stained with
antibodies to CD103, the β chain or the δ chain of TCR. (A) Evidence that there no significant
differences in the total number of CD103+ IELs per 1000 villus epithelial cells among the three
groups. (B) Data indicating that there is a statistically significant increase in the density of
αβTCR+ IELs in Fabpi-TAgWt mice (asterisk, p<0.05 when compared to normal FVB/N mice).
(C) Results showing a statistically significant reduction in γδ TCR+ IELs in Fabpi-TAgWt
animals.
Figure 4. Quantitative immunohistochemical studies of IEL populations in germ-free
transgenic and normal mice. (A) A section of jejunum, harvested from a 6-8 week old male
germ-free Fabpi-TAgWt mouse, was stained exactly as described in the legend to Fig. 1.
Enterocytes distributed from the base to the tips of jejunal villi have re-entered the cell cycle
(BrdU; red, TAg, green; colocalization, yellow). (B,C) Immunohistochemical study using
antibodies to the δ chain of TCR, showing that Fabpi-TAgWt mice have a marked reduction in
the density of jejunal villus γδ TCR+ IELs (pink) compared to normal litteremates. Nuclei are
stained blue with bis-benzimide. (D-F) Sections of jejunum were stained antibodies specific for
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

31
CD103, the β chain or the δ chain of TCR and the number of CD103+, γδTCR+ and αβTCR+
IELs scored per 1000 villus epithelial cells. Mean values ± S.E. for each subset are plotted
(asterisk, p<0.05 when compared to normal mice; n = 3 animals/group/experiment; 3
experiments).
Figure 5. FACS analysis of IELs harvested from the jejunums of germ-free Fabpi-TAgWt
mice demonstrates an increase in thymically-derived CD4+, CD8ααααββββ++++ ααααββββTCR IELs and a
decrease in intestinally-derived γγγγδδδδTCR+CD8αααααααα IELs compared to normal littermates.
Mean values ± S.E. are plotted for each group of mice (n = 3 mice/group/experiment; 3
experiments).
Figure 6. qRT-PCR studies of gene expression in laser capture microdissected jejunal villus
epithelium obtained from germ-free Fabpi-TAgWt mice and their normal littermates. (A)
LCM of jejunal villus epithelium. 5 µm-thick cryosections were prepared from jejunal segments
of a normal mouse, fixed in 70% ethanol, and stained with eosin Y and methyl green.
Bar = 25µm. (B) qRT-PCR showing that forced expression of TAgWt in villus enterocytes is
associated with a reduction in IL-7 mRNA levels. Mean values ± S.E. are plotted. Transcript
levels were first normalized to Gapdh mRNA. The normalized values were then referenced to
levels of IL-7 mRNA (arbitrarily set at 1) in LCM jejunal villus epithelial RNA obtained from
normal littermate controls. (C) qRT-PCR analysis of the effects of TAgWt expression on levels of
TCRδ, TGFβ3, Flt3 ligand and Flt3 receptor mRNAs in LCM villus epithelium. (n=3
mice/experiment; 2 independent experiments in both B and C).
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

32
Figure 7. qRT-PCR analysis of IL-7Rαααα, TGFββββ3, and Flt3 ligand expression in
γγγγδδδδ TCR++++ IELs purified from 6 week old germ-free Fabpi-TAgWt male transgenic mice and
their normal FVB/N littermates. IELs were harvested from 50 mice per group. Each IEL RNA
preparation was assayed in triplicate. Mean values ± S.D. are plotted.
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

33
Supplemental on-line material
Table 1 - Gene-specific primers used for SYBR-Green real time quantitative RT-PCR
studies
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Primers for real time quantitative RT-PCR
GeneGenBank Accession #
Forward Primer (5'-3') Reverse Primer (5'-3')amplicon
length (bp)
amplicon
Tm (oC)Gapdh NM008084 TGGCAAAGTGGAGATTGTTGCC AAGATGGTGATGGGCTTCCCG 155 8018S rRNA AY083001 CATTCGAACGTCTGCCCTATC CCTGCTGCCTTCCTTGGA 136 80TGFβ3 M32745 GGAAATGGGTCC ACGAACCTA TCCAAGCACCGTGCTATGG 101 82Flt3L U29875 GCAGGGTCTAAGATGCAAACG ACGAATCGCAGACATTCTGGTA 101 80Flt3 NM010229 TCCTTTGCTTTGCGTACCAA GTGGGTGACCAACACATTCCT 101 82IL-7 NM008371 GGAATTCCTCCACTGATC TGTCTTTAATGTGGCACTCAGATGAT 64 75IL-7Rα AF078906 CAGAAATAGGCCTCCTGTGTATCAA GGATTCCCGACTGGAAAGG 104 81TCRδ L36135 GTGGGCTGACATGAGGAGACTAC GGTGGTGAAGGGTCAGTCTGA 109 80
by guest on March 26, 2019http://www.jbc.org/Downloaded from

Indira U. Mysorekar, Robin G. Lorenz and Jeffrey I. Gordonintestinal enterocytes and intraepithelial lymphocytes
A gnotobiotic transgenic mouse model for studying interactions between small
published online July 22, 2002J. Biol. Chem.
10.1074/jbc.M205300200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on March 26, 2019
http://ww
w.jbc.org/
Dow
nloaded from