Embed Size (px)
Citation preview

Ecology, 90(2), 2009, pp. 465–475! 2009 by the Ecological Society of America
A link between water availability and nesting successmediated by predator–prey interactions in the Arctic
NICOLAS LECOMTE,1,3 GILLES GAUTHIER,1 AND JEAN-FRANCOIS GIROUX2
1Departement de Biologie and Centre d’Etudes Nordiques, Universite Laval, Quebec, Canada2Groupe de Recherche en Ecologie Comportementale et Animale, Departement des Sciences Biologiques,
Universite du Quebec a Montreal, Quebec, Canada
Abstract. Although water availability is primarily seen as a factor affecting foodavailability (a bottom-up process), we examined its effect on predator–prey interactionsthrough an influence on prey behavior (a top-down process). We documented a link betweenwater availability, predation risk, and reproductive success in a goose species (Chencaerulescens atlantica) inhabiting an Arctic environment where water is not considered alimited commodity. To reach water sources during incubation recesses, geese nesting in mesictundra (low water availability) must move almost four times as far from their nest than thosenesting in wetlands, which reduced their ability to defend their nest against predators and ledto a higher predation rate. Nesting success was improved in high rainfall years due toincreased water availability, and more so for geese nesting in the low water availability habitat.Likewise, nesting success was improved in years where the potential for evaporative water loss(measured by the atmospheric water vapor pressure) was low, presumably because females hadto leave their nest less often to drink. Females from water-supplemented nests traveled ashorter distance to drink, and their nesting success was enhanced by 20% compared to thecontrol. This shows that water availability and rainfall can have a strong effect on predator–prey dynamics and that changes in precipitation brought by climate change could have animpact on some Arctic species through a top-down effect.
Key words: Arctic-nesting Snow Geese; Bylot Island, Nunavut, Canada; Chen caerulescens; incubation;predator–prey interactions; resource limitation; top-down control; water availability and limitation.
INTRODUCTION
Water can be a limiting factor for ecophysiologicalprocesses (e.g., Bartholomew and Cade 1963, Klaassen2004, Williams and Tieleman 2005). However, wateravailability can also affect demographic processes suchas survival and reproduction (Newton 1998, Tieleman etal. 2004). For instance, increased water levels andflooding can lead to a complete breeding failure andreduced survival in ground-breeding species (e.g., Myerset al. 1985, Thompson and Furness 1991, Ratcliffe et al.2005). Reproductive success can also be positively linkedwith the amount of precipitation because of increasedwater availability at the onset of the breeding period(Coe and Rotenberry 2003, Ladia et al. 2007). This linkis usually assumed to be mediated through an increase infood availability rather than water per se (bottom-upcontrol; Blancher and Robertson 1987, Gibbs and Grant1987, Ogutu et al. 2008).Water availability can also affect predator–prey
interactions (top-down control; Ostfeld and Keesing2000). In arid ecosystems, where water limitations are
ubiquitous, individuals are distributed according toavailability of surface water, often trading off theirnutritionnal requirements against water use (Redfern etal. 2003, Tielman et al. 2004). The concentration ofindividuals at water holes can considerably increase theirvulnerability to predation (Ferns and Hinsley 1995,Doody et al. 2007, Valeix et al. 2007). However, theeffect of water availability on predation–prey interac-tions has been largely ignored in environments wherewater is not usually considered limiting (Morrison andBolger 2002). Yet, recent evidence suggests that thespatial distribution of water or precipitation regimes canaffect predator efficiency, with potentially strong conse-quences for predator–prey dynamics and ecosystemprocesses (e.g., Hilton et al. 1999, Post et al. 1999,Lecomte et al. 2008a). Although prey behavior couldalso be affected by water availability and rainfall, weknow very little about how it can influence predationrisk, and hence predator–prey dynamics, in non-aridenvironments.
In birds, nest predation can have a considerableimpact on reproductive success (Martin 1995). In specieswhere only one sex incubates, incubating birds mustbalance the conflicting needs of foraging and protectingtheir nest because, when they leave to feed, predationrisk of unattended eggs often increases (Thompson andRaveling 1987, Samelius and Alisauskas 2001). Change
Manuscript received 31 January 2008; revised 17 June 2008;accepted 23 June 2008. Corresponding Editor: E. G. Cooch.
3 Present address: Department of Biology, University ofTromsø, N-9037, Tromsø, Norway.E-mail: [email protected]
465

in resource availability during incubation could thenaffect egg predation rate if this influences the time spentoff the nest by the sole incubating parent (thereaftercalled recess). Much emphasis has been placed on foodavailability or feeding behavior during incubation(Martin and Ghalambor 1999), but constraints imposedby water requirements have been largely ignored, eventhough they can be significant in species that mostly fastduring incubation (Le Maho et al. 1981, Boismenu et al.1992, Reed et al. 1995). Conditions that favor evapora-tive water loss (e.g., low atmospheric water vaporpressure) or reduce water availability around the nest(e.g., low rainfall) may increase recess frequency or thedistance traveled by the birds to drink, thereby exposingtheir nest to increased predation. Therefore, such asystem is ideally suited for examining how wateravailability can affect a predator–prey interactionthrough its influence on prey behavior.We investigated how water availability affected
nesting success in an Arctic-nesting species, the SnowGoose (Chen caerulescens; see Plate 1). In this gynepar-ental incubating species, predation is the primary causeof nest failure and occurs mostly when females leavetheir nest to drink or feed during incubation (Cooke etal. 1995, Bety et al. 2001; and this study). Althoughmales can limit egg losses by attacking and chasing offpredators (Samelius and Alisauskas 2005), they mostoften follow their mate during recesses, leaving the nestunprotected. Unattended Snow Goose nests suffer amuch higher predation rate than attended ones (Same-lius and Alisauskas 2001) and this risk increases withdistance from the nest by the parents (Bety et al. 2002).We tested the hypothesis of a link between water
availability and nesting success mediated throughpredation. We first predicted that geese nesting in mesic
tundra where water is scarce should be more vulnerableto predation than those in wetlands because they had totravel a greater distance to drink during recesses (Fig. 1).Distance to water is taken as an index of local wateravailability and predators should thus find moreunattended nests in mesic tundra than in wetlands. Wetested these predictions by monitoring the behavior ofincubating females during recesses and the foragingbehavior of predators in the two habitats during fourbreeding seasons. We also hypothesized that rainfallshould increase water availability and thus decreasepredation risk as distance to drinking sources is reducedafter rainfall, especially in mesic tundra. We thereforepredicted that difference in nesting success betweenwetland and mesic tundra should vary with rainfall,increasing in dry years and decreasing in wet years. Wefurther hypothesized that atmospheric water vaporpressure should affect evaporative water loss by geeseand influences their need to leave the nest to drink.Thus, we predicted that nesting success should increasein years where atmospheric water pressure was high.Finally, to eliminate the confounding factor of differ-ences in food quality between wetlands and mesic tundra(i.e., wetlands have more food plants preferred by geese;Gauthier et al. 1996), we experimentally manipulatedwater availability by supplementing nesting pairs withwater. We predicted that pairs that were provided withwater should stay closer to their nest during recesses andachieve higher nesting success than controls.
METHODS
Study site
Data were collected between 1995 and 2005 on BylotIsland, Nunavut, Canada (728530 N, 798540 W). Thisisland is the most important breeding site for GreaterSnow Geese (Chen caerulescens atlantica), with morethan 20 000 breeding pairs, the majority of which arenesting in the same colony (Reed et al. 2002).The habitat is a mosaic of wetlands and mesic tundra.
Wetlands typically occur in small patches (average size:3.4 ha) embedded into a landscape dominated by mesictundra, which covers over 10 times more area thanwetlands (Lecomte et al. 2008b). Wetlands are mostlyfound in polygon-patterned ground created by frostaction in lowland tundra (Gauthier et al. 1996).Polygons often form a heterogeneous, intricate networkof lakes, ponds, and narrow water channels that hamperthe movement of walking predators (Fig. 1; Lecomte etal. 2008a). Wetlands have a rich cover of graminoids, thepreferred food plants of geese (Gauthier et al. 1996).Mesic tundra is found in drier lowlands, gentle slopes ofsmall hills, and low-altitude plateaus. Standing water isscarce in mesic tundra and is only found in isolatedsmall ponds or streams (Fig. 1) and between hummocks(low mounds of earth formed by winter freezing)following a rainfall. This habitat is dominated byprostrate shrubs and sparse forbs and graminoids.
FIG. 1. Constraints imposed by habitat on water supply in aSnow Goose colony. This figure is based on an aerialphotograph of a portion of the colony showing hypotheticalgoose nests (white circles) and movements during incubationrecesses (white arrows) toward permanent water sources (black)either in wetlands (dark gray) or in mesic tundra (light gray).
NICOLAS LECOMTE ET AL.466 Ecology, Vol. 90, No. 2

The predator community of Bylot Island is relativelysimple with only one terrestrial predator, the Arctic fox(Vulpex lagopus; hereafter fox), and several avianpredators, such as Parasitic Jaegers (Stercorarius para-siticus), Glaucous Gulls (Larus hyperboreus), andCommon Ravens (Corvus corax; Bety et al. 2001). Foxesare the most important predator, depredating 72% of allgoose eggs (Bety et al. 2002). All predators used bothhabitats and easily moved between them becausewetland patches are much smaller than their homerange size (Lecomte et al. 2008a).
Climatic data
At our study site, an automated weather station hasbeen recording hourly air temperature since 1995 andrelative humidity since 1999 using a Vaisala HMP35CFprobe (Vaisala, Blainville, Quebec, Canada). Mean dailytemperature and relative humidity during the study was3.78C (annual range: 1.0–7.78C) and 81.2% (annualrange: 77.7–85.8%; typical value for High Arctic areas).Each year, we manually recorded daily rainfall (mm)from 1 June to 15 August with a calibrated rain gauge.
We estimated the partial water vapor pressure of the air,p(H2O) in hectopascals (hPa), which drives evaporationrate in animals (Gates 1980), using the Goff-Gratchequation (Goff and Gratch 1946), which is based ontemperature and relative humidity.
Behavioral observations
From 2002 to 2005, we carried out behavioralobservations of foraging predators and incubating geese.We observed predators and geese throughout the 24-hdaylight period of the Arctic during the incubationperiod (9 June to 6 July; 600 hours of observationsduring 120 days). From three blinds, we used binoculars(103) and spotting scopes (20–603) to cover an area of2.8 km2 where both nesting habitats (wetlands and mesictundra) were present and where nest densities wererepresentative of those encountered in the whole colony.The number of goose nests under observation rangedannually from 100 to 500. We continuously scannedincubating females, and every time a female was seendeparting from a nest, we started a stopwatch andobserved it continuously until she resumed incubation.
PLATE 1. Greater Snow Goose pair protecting their nest at the Bylot Island goose colony, Nunavut, Canada, in 2005. Whenthey are present close or on the nest, the predation risk is at its lowest level for the eggs. Photo credit: N. Lecomte.
February 2009 467WATER LIMITATION IN ARCTIC-NESTING GEESE

Upon departure, we measured the distance traveled tothe nearest water source to drink. Distance was eitherestimated visually using 250 georeferenced sticks locatedin the colony, or a posteriori with a measuring tape(median precision of 1.5 m). For each drinking event, werecorded whether the water source was permanent(ponds, rivers) or temporary (small depression at thesurface of the tundra). We recorded all nest attacks bypredators during the absence of the female, noted thepredator involved, and whether it was successful (i.e., atleast one egg taken) or not. We defined an attack as anyattempt by a predator to take goose eggs. At least sevento eight foxes were identified individually by thedistinctive patterns of their fur (shedding from winterto summer fur) each year (Lecomte et al. 2008a).
Goose nest monitoring
We systematically searched for goose nests in differentareas of the colony during egg laying and earlyincubation. Nests located either in the central portionof the colony or in random plots scattered throughoutthe colony were found to differ little in terms of nestingparameters (Reed et al. 2005); thus, all nests were pooledand considered representative of the colony as a whole.For each nest, we determined the habitat (wetland ormesic tundra) and revisited them once during incubationand again at hatching to monitor their fate. Wedetermined laying date of the first egg (nest initiation)following Bety et al. (2001). We defined total clutch sizeas the maximum number of eggs found in a nest after thestart of incubation. At hatch, we counted the totalnumber of goslings in the nest. Although partialpredation occurs, only about 10% of nests surviving tohatch lost some eggs (Lepage et al. 2000) and mostpredation events usually resulted in the loss of a fullclutch. We therefore focused mostly on whole-nestsuccess in subsequent analyses and we defined a nestas successful if at least one egg hatched.
Water supplementation experiment
During the period when geese were incubating in2005, we supplemented 40 randomly chosen nests in thecolony with water, split equally between wetlands andmesic tundra. For each nest, we used two dark brownplastic containers with a volume of ;8 L (11.7 cm wide3 20.8 cm deep3 34.1 cm long). We buried them in theground in opposite directions from the nest at distancesranging from three to six meters. This distance waschosen because Bety et al. (2002) showed that the successof predator attacks on goose nests increased with thedistance between the parents and their nest, and wasvery high when parents were !10 m from the nest. Thisdistance also ensured that water supplementation wasrestricted to parents of focus nests as geese are territorialin the vicinity of their nest (Cooke et al. 1995). We filledcontainers with water from neighboring ponds for thefirst time one or two days after the onset of theincubation period and we refilled all of them on the
same day at mid-incubation. Since no containers wereempty at the time of refilling, this suggests that femaleshad access to supplementary water throughout theirincubation period. We used as control a sample ofnonmanipulated nests observed and monitored in thesame year. The spatial distribution of control andexperimental nests overlapped in the colony.We conducted behavioral observations at the 40
experimental nests using the same methods previouslydescribed, and we monitored their fate until hatch. Wealso recorded presence of males at the experimental nestsand at 40 control nests randomly chosen in the colony.Male presence was defined as the percentage of totaltime spent close to the nest ("10 m) during periods ofobservations (see also Samelius and Alisauskas [2001]).We detected no widowed females as males were seen atleast once at all nests under observation.
Data analysis
Behavior of incubating females and predators.—Wefirst explored the effect of water on predation risk bycomparing the distance traveled by females from theirnest to a drinking site between years and habitats withtwo-way ANOVAs. Second, we compared this distancebetween days following significant rainfall (i.e., presenceof .20 mm of rainfall in the preceding three days) andthose that did not. We also used a v2 statistic to comparethe success of predators’ attacks in the two nestinghabitats during recesses.Nesting success.—We examined the effects of habitat
and covariates such as rainfall and p(H2O) on goosenesting success with the daily nest survival (S ) procedureof program MARK Version 4.2 (Dinsmore et al. 2002).We included the age of the nest at the time of discovery(date found minus estimated laying date) as anindividual covariate in the model because predation ratecan vary during the nesting period (Bety et al. 2002). Wefirst considered the model with full time (i.e., years; t),habitat (h), day of the season (linear effect; d ), and ageeffects with relevant interactions as the most generalmodel. To select the most parsimonious model, we usedthe Akaike’s Information Criterion modified for smallsample size (AICc) and the relative weight of evidence infavor of a particular model (xAICc; Burnham andAnderson 1998). In presence of uncertainty in modelselection (DAICc , 4), we estimated model-weightedaverage parameter values and their unconditionalstandard errors (Burnham and Anderson 1998). Weestimated nesting success as the product of daily nestsurvival for the mean duration of laying and incubation(27 d).To examine the effect of rainfall and p(H2O), we
substituted time (i.e., years) by these covariates. Weparameterized rainfall and p(H2O) as binary variables tocontrast years with higher (wet years# 1) and lower (dryyears # 0) rainfall or p(H2O) than average. However,p(H2O) data were available for only seven years (1999–2005) compared to 11 years (1995–2005) for rainfall. We
NICOLAS LECOMTE ET AL.468 Ecology, Vol. 90, No. 2
thus ran two separate analyses with covariates, a firstone using only rainfall for 1995–2005 and a second usingp(H2O) and rainfall for 1999–2005. We determined theproportion of variation in nest survival explained bythese time-dependent covariates by calculating the ratioin difference in deviance between the constant modeland the model with covariates vs. the difference betweenthe constant and the full time-dependent (i.e., year)model (analogous to an r2; Agresti 2002). Finally, weformally tested the effect of rainfall or p(H2O) on nestsurvival using the analysis of deviance (ANODEV;Agresti 2002), which contrasts the amount of devianceaccounted for by a model with covariate vs. the totalamount of deviance accounted by the full time-dependent model.Water supplementation experiment.—We determined
the effect of habitat on the use of water containers vs.natural sources of water with a generalized estimatingequation (GEE) model and a logit link function (Hardinand Hilbe 2003). This procedure accounts for the non-independence of multiple observations on the sameindividuals (see Bety et al. [2002] for details on theprocedure). The same procedure was used to determinethe effects of habitat and experiment on male presence atthe nest. We used a generalized linear mixed regressionmodel to examine the effects of habitat and experimentaltreatment (fixed effects) on distance traveled to drink byfemales during recesses, adding female identity as arandom effect. We compared nesting success betweenexperimental and control nests following the sameprocedure presented earlier (Dinsmore et al. 2002). Wecompared the mean number of hatchlings per successfulnest between the two habitats and between experimentalvs. control nests with a two-way ANOVA. Because thedependant variable was not normally distributed, weranked transformed data and we tested interactionsusing the aligned rank test procedure as recommended
by Salter and Fawcett (1993). Finally, to test for possibleconfounding factors affecting nesting success, wecompared laying date and clutch size between habitatsand experimental and control nests with two-wayANOVAs, again using rank-transformed variables.
General statistics procedures.—Statistical analysesother than nest survival were performed using SASsoftware, Version 9 (SAS Institute 2002). We consideredeach goose nest as independent because (1) individualgeese can defend their nests against predators, (2) thefate of a nest is independent of the fate of its nearestneighbors (Bety et al. 2001), and (3) several foxes (7–8per year) were known to forage within the study area(Lecomte et al. 2008a). For all data, normality waschecked with the Kolmogorov-Smirnov test (Lillieforsoption). All probabilities are two-tailed and the level wasset to a # 0.05. Unless mentioned otherwise, values arereported as mean 6 SE.
RESULTS
Rainfall and water availability
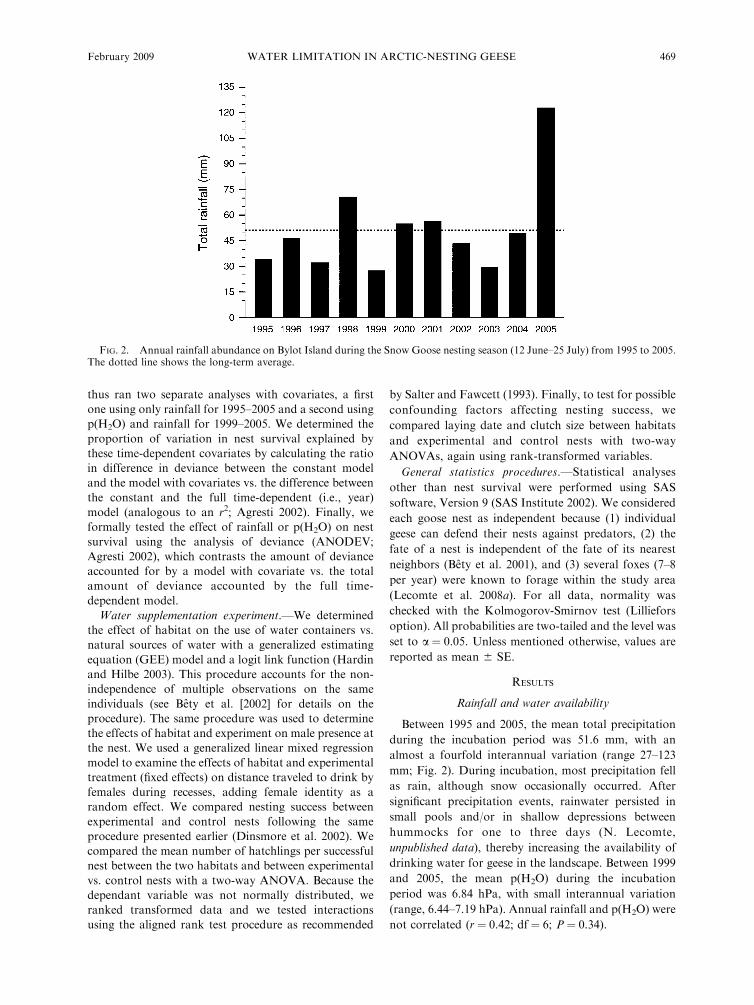
Between 1995 and 2005, the mean total precipitationduring the incubation period was 51.6 mm, with analmost a fourfold interannual variation (range 27–123mm; Fig. 2). During incubation, most precipitation fellas rain, although snow occasionally occurred. Aftersignificant precipitation events, rainwater persisted insmall pools and/or in shallow depressions betweenhummocks for one to three days (N. Lecomte,unpublished data), thereby increasing the availability ofdrinking water for geese in the landscape. Between 1999and 2005, the mean p(H2O) during the incubationperiod was 6.84 hPa, with small interannual variation(range, 6.44–7.19 hPa). Annual rainfall and p(H2O) werenot correlated (r # 0.42; df # 6; P # 0.34).
FIG. 2. Annual rainfall abundance on Bylot Island during the Snow Goose nesting season (12 June–25 July) from 1995 to 2005.The dotted line shows the long-term average.
February 2009 469WATER LIMITATION IN ARCTIC-NESTING GEESE

Use of water by geese and predator attacks
We observed 102 incubation recesses for 96 nests.After departure from the nest, drinking was the firstactivity performed by females in 89.3% of the cases (n#91) whereas feeding behavior was the first activity in8.8% of recesses (n # 9), and preening was the firstactivity in 1.9% (n # 2). Only 10.8% (n # 11) of allrecesses involved no drinking by females and occurredwhen the birds had to precipitately return to their nestdue to presence of a predator. Globally, drinking,feeding, and preening were the most important activitiesduring these recesses. Moreover, several drinking boutswere required by females to restore their hydric balancesince they drank on average seven times per recess (n #15).During recesses, females nesting in mesic tundra
traveled 3.8 times farther from their nest to drink thanthose nesting in wetlands (mesic, 37 6 3 m; wetland, 9 63 m; F1,98# 57.7; P , 0.001), with no difference betweenyears (F1,98 # 0.001; P # 0.97). During the three daysfollowing a significant rainfall, distance traveled to drinkdecreased in both habitats (mesic tundra before rain, 396 3 m; after rain, 196 5 m; wetlands before rain, 136 3m; after rain 6 6 1 m; F1,74 # 13.5; P , 0.001). Thisoccurred because females drank in temporary waterdepressions rather than in permanent water sources(ponds, rivers).Compared to wetlands, females nesting in mesic
tundra were 13.9 times more likely to exceed the 10-mthreshold distance where the success of predator attacksincreases considerably during incubation recesses (Betyet al. 2002). As a result, both avian and terrestrialpredators were more successful in robbing nests in mesictundra (89% of attacks were successful; n# 200 predatorattacks) than in wetlands (79% of attacks weresuccessful; n # 77 predator attacks; v2 # 4.3; df # 1; P# 0.038) during incubation recesses.
Water availability and nesting success
Between 1995 and 2005, we monitored the fate of 1331and 1693 nests in wetlands and mesic tundra, respec-
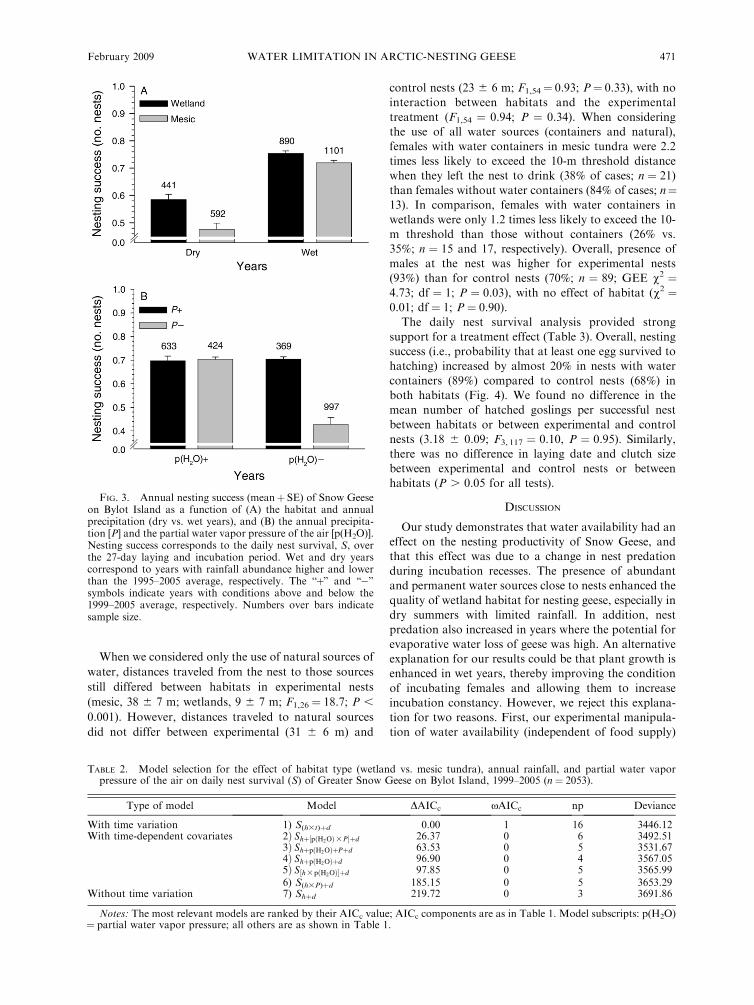
tively. During these years, daily nest survival differedamong habitats and years, increased with the day of theseason, and possibly with the age when a nest wasfound, as suggested by the four top models in Table 1.When time was substituted by rainfall in the model, thisvariable in interaction with habitat types (Model 6,Table 1) explained 34% (r2DEV) of the annual variation innest survival, a significant result (ANODEV of Model 6vs. Models 1 and 8 in Table 1; F2,18 # 4.54; P # 0.025).Overall, nesting success of geese was higher in wet thanin dry years, and the difference in success betweenhabitats was higher during dry years (11%, on average)than during wet ones (3%; Fig. 3A). This suggests thatrainwater was relatively more beneficial for pairs nestingin mesic tundra than for those nesting in wetlands wheresuccess was higher.When we considered simultaneously the effect of
rainfall and p(H2O) for the subset of years when bothmeasurements were available, we found that bothvariables in interaction explained 81% of the annualvariation in nest survival, a significant result (ANODEVof Model 2 vs. Models 1 and 7 in Table 2; F3,10 # 14.3;P , 0.001). Overall, nesting success of geese was highwhen either rainfall or p(H2O) was high, but success wasreduced by almost 39% when both of these variableswere low (Fig. 3B). The latter case corresponds tosituations where potential evaporative water loss forgeese was high and drinking water availability aroundthe nest was low.
Water supplementation and goose behavior
In 2005, we observed 44 incubation recesses byfemales for the 40 nests equipped with water containers,and 30 recesses for 40 control nests. Both females andmales used water containers, either for drinking orbathing. Females in wetlands used nearly four timesmore natural sources of water to drink (n# 15) than thewater containers (n # 4), whereas geese in mesic tundraused containers more often (11 times in natural sourcesvs. 14 in containers; GEE (generalized estimatingequation) v2 # 5.1; df # 1; P # 0.02).
TABLE 1. Model selection for the effect of habitat type (wetland vs. mesic tundra) and annual rainfall on daily nest survival (S ) ofGreater Snow Geese on Bylot Island, 1995–2005 (n # 3024).
Type of model Model DAICc xAICc np Deviance
With time variation 1) S(h3t)$d 0.00 0.54 23 5133.872) S(h3t)$d$age 1.84 0.21 24 5133.723) Sh$t$d 2.49 0.15 13 5156.384) Sh$t$d$age 3.65 0.08 14 5155.545) St$d$age 22.42 0.00 13 5176.31
With time-dependent covariates 6) S(h3P)$d 273.71 0.00 5 5443.587) Sh$P$d 308.36 0.00 4 5480.23
Without time variation 8) Sh$d 425.87 0.00 3 5599.77
Notes: The most relevant models are ranked by their AICc value. For each model, the difference in AICc values is given inrelation to the most parsimonious model (DAICc), the model’s AICc weight (xAICc), the number of estimable parameters (np), andthe deviance. Model subscripts are: age# age of the nest when found (date found minus the nest initiation date), d# linear effect ofday of the season, h# habitat (wetlands vs. mesic), P# precipitation (dry vs. wet years), t# year (‘‘$’’ indicates additive effect; ‘‘3’’indicates interactive effect.
NICOLAS LECOMTE ET AL.470 Ecology, Vol. 90, No. 2
When we considered only the use of natural sources ofwater, distances traveled from the nest to those sourcesstill differed between habitats in experimental nests(mesic, 38 6 7 m; wetlands, 9 6 7 m; F1,26 # 18.7; P ,0.001). However, distances traveled to natural sourcesdid not differ between experimental (31 6 6 m) and
control nests (23 6 6 m; F1,54# 0.93; P# 0.33), with nointeraction between habitats and the experimentaltreatment (F1,54 # 0.94; P # 0.34). When consideringthe use of all water sources (containers and natural),females with water containers in mesic tundra were 2.2times less likely to exceed the 10-m threshold distancewhen they left the nest to drink (38% of cases; n # 21)than females without water containers (84% of cases; n#13). In comparison, females with water containers inwetlands were only 1.2 times less likely to exceed the 10-m threshold than those without containers (26% vs.35%; n # 15 and 17, respectively). Overall, presence ofmales at the nest was higher for experimental nests(93%) than for control nests (70%; n # 89; GEE v2 #4.73; df # 1; P # 0.03), with no effect of habitat (v2 #0.01; df # 1; P # 0.90).
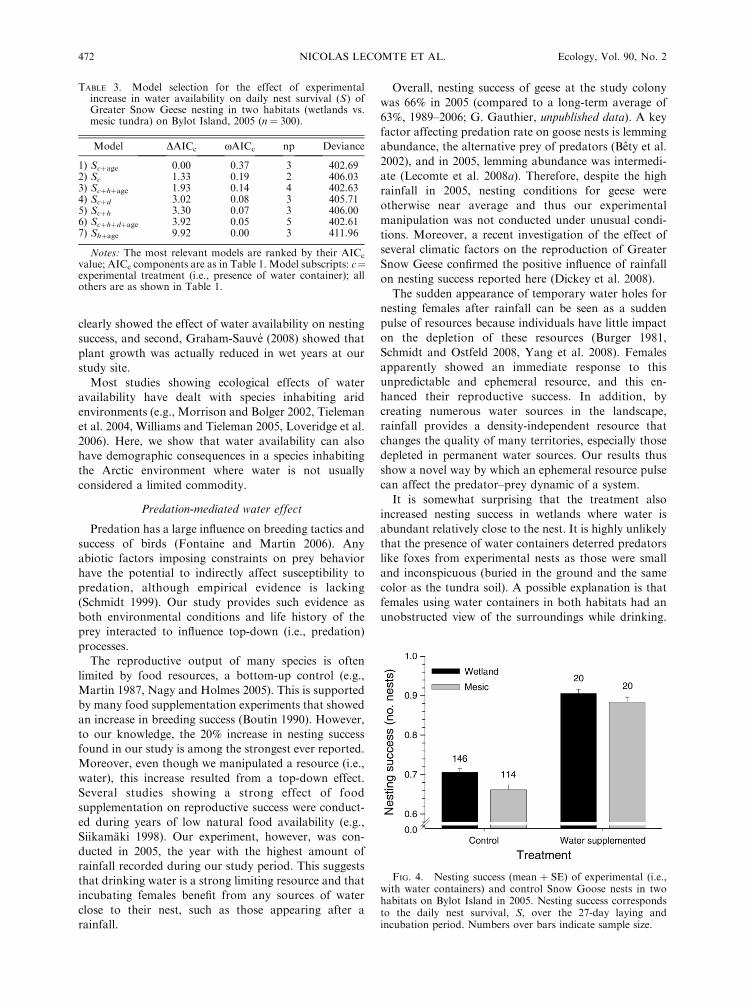
The daily nest survival analysis provided strongsupport for a treatment effect (Table 3). Overall, nestingsuccess (i.e., probability that at least one egg survived tohatching) increased by almost 20% in nests with watercontainers (89%) compared to control nests (68%) inboth habitats (Fig. 4). We found no difference in themean number of hatched goslings per successful nestbetween habitats or between experimental and controlnests (3.18 6 0.09; F3, 117 # 0.10, P # 0.95). Similarly,there was no difference in laying date and clutch sizebetween experimental and control nests or betweenhabitats (P . 0.05 for all tests).
DISCUSSION
Our study demonstrates that water availability had aneffect on the nesting productivity of Snow Geese, andthat this effect was due to a change in nest predationduring incubation recesses. The presence of abundantand permanent water sources close to nests enhanced thequality of wetland habitat for nesting geese, especially indry summers with limited rainfall. In addition, nestpredation also increased in years where the potential forevaporative water loss of geese was high. An alternativeexplanation for our results could be that plant growth isenhanced in wet years, thereby improving the conditionof incubating females and allowing them to increaseincubation constancy. However, we reject this explana-tion for two reasons. First, our experimental manipula-tion of water availability (independent of food supply)
TABLE 2. Model selection for the effect of habitat type (wetland vs. mesic tundra), annual rainfall, and partial water vaporpressure of the air on daily nest survival (S) of Greater Snow Geese on Bylot Island, 1999–2005 (n # 2053).
Type of model Model DAICc xAICc np Deviance
With time variation 1) S(h3t)$d 0.00 1 16 3446.12With time-dependent covariates 2% Sh$&p'H2O%3P($d 26.37 0 6 3492.51
3% Sh$p'H2O%$P$d 63.53 0 5 3531.674% Sh$p'H2O%$d 96.90 0 4 3567.055% S&h3 p'H2O%($d 97.85 0 5 3565.996) S(h3P)$d 185.15 0 5 3653.29
Without time variation 7) Sh$d 219.72 0 3 3691.86
Notes: The most relevant models are ranked by their AICc value; AICc components are as in Table 1. Model subscripts: p(H2O)# partial water vapor pressure; all others are as shown in Table 1.
FIG. 3. Annual nesting success (mean$ SE) of Snow Geeseon Bylot Island as a function of (A) the habitat and annualprecipitation (dry vs. wet years), and (B) the annual precipita-tion [P] and the partial water vapor pressure of the air [p(H2O)].Nesting success corresponds to the daily nest survival, S, overthe 27-day laying and incubation period. Wet and dry yearscorrespond to years with rainfall abundance higher and lowerthan the 1995–2005 average, respectively. The ‘‘$’’ and ‘‘)’’symbols indicate years with conditions above and below the1999–2005 average, respectively. Numbers over bars indicatesample size.
February 2009 471WATER LIMITATION IN ARCTIC-NESTING GEESE
clearly showed the effect of water availability on nestingsuccess, and second, Graham-Sauve (2008) showed thatplant growth was actually reduced in wet years at ourstudy site.Most studies showing ecological effects of water
availability have dealt with species inhabiting aridenvironments (e.g., Morrison and Bolger 2002, Tielemanet al. 2004, Williams and Tieleman 2005, Loveridge et al.2006). Here, we show that water availability can alsohave demographic consequences in a species inhabitingthe Arctic environment where water is not usuallyconsidered a limited commodity.
Predation-mediated water effect
Predation has a large influence on breeding tactics andsuccess of birds (Fontaine and Martin 2006). Anyabiotic factors imposing constraints on prey behaviorhave the potential to indirectly affect susceptibility topredation, although empirical evidence is lacking(Schmidt 1999). Our study provides such evidence asboth environmental conditions and life history of theprey interacted to influence top-down (i.e., predation)processes.The reproductive output of many species is often
limited by food resources, a bottom-up control (e.g.,Martin 1987, Nagy and Holmes 2005). This is supportedby many food supplementation experiments that showedan increase in breeding success (Boutin 1990). However,to our knowledge, the 20% increase in nesting successfound in our study is among the strongest ever reported.Moreover, even though we manipulated a resource (i.e.,water), this increase resulted from a top-down effect.Several studies showing a strong effect of foodsupplementation on reproductive success were conduct-ed during years of low natural food availability (e.g.,Siikamaki 1998). Our experiment, however, was con-ducted in 2005, the year with the highest amount ofrainfall recorded during our study period. This suggeststhat drinking water is a strong limiting resource and thatincubating females benefit from any sources of waterclose to their nest, such as those appearing after arainfall.
Overall, nesting success of geese at the study colonywas 66% in 2005 (compared to a long-term average of63%, 1989–2006; G. Gauthier, unpublished data). A keyfactor affecting predation rate on goose nests is lemmingabundance, the alternative prey of predators (Bety et al.2002), and in 2005, lemming abundance was intermedi-ate (Lecomte et al. 2008a). Therefore, despite the highrainfall in 2005, nesting conditions for geese wereotherwise near average and thus our experimentalmanipulation was not conducted under unusual condi-tions. Moreover, a recent investigation of the effect ofseveral climatic factors on the reproduction of GreaterSnow Geese confirmed the positive influence of rainfallon nesting success reported here (Dickey et al. 2008).The sudden appearance of temporary water holes for
nesting females after rainfall can be seen as a suddenpulse of resources because individuals have little impacton the depletion of these resources (Burger 1981,Schmidt and Ostfeld 2008, Yang et al. 2008). Femalesapparently showed an immediate response to thisunpredictable and ephemeral resource, and this en-hanced their reproductive success. In addition, bycreating numerous water sources in the landscape,rainfall provides a density-independent resource thatchanges the quality of many territories, especially thosedepleted in permanent water sources. Our results thusshow a novel way by which an ephemeral resource pulsecan affect the predator–prey dynamic of a system.It is somewhat surprising that the treatment also
increased nesting success in wetlands where water isabundant relatively close to the nest. It is highly unlikelythat the presence of water containers deterred predatorslike foxes from experimental nests as those were smalland inconspicuous (buried in the ground and the samecolor as the tundra soil). A possible explanation is thatfemales using water containers in both habitats had anunobstructed view of the surroundings while drinking.
TABLE 3. Model selection for the effect of experimentalincrease in water availability on daily nest survival (S) ofGreater Snow Geese nesting in two habitats (wetlands vs.mesic tundra) on Bylot Island, 2005 (n # 300).
Model DAICc xAICc np Deviance
1) Sc$age 0.00 0.37 3 402.692) Sc 1.33 0.19 2 406.033) Sc$h$age 1.93 0.14 4 402.634) Sc$d 3.02 0.08 3 405.715) Sc$h 3.30 0.07 3 406.006) Sc$h$d$age 3.92 0.05 5 402.617) Sh$age 9.92 0.00 3 411.96
Notes: The most relevant models are ranked by their AICc
value; AICc components are as in Table 1. Model subscripts: c#experimental treatment (i.e., presence of water container); allothers are as shown in Table 1.
FIG. 4. Nesting success (mean $ SE) of experimental (i.e.,with water containers) and control Snow Goose nests in twohabitats on Bylot Island in 2005. Nesting success correspondsto the daily nest survival, S, over the 27-day laying andincubation period. Numbers over bars indicate sample size.
NICOLAS LECOMTE ET AL.472 Ecology, Vol. 90, No. 2

In contrast, small ponds or water channels in wetlandsare often located in depressions surrounded by polygonrims, which hinder the vision of drinking females,thereby limiting their detection of approaching eggpredators (e.g., Kahlert 2003). Finally, higher wateravailability near the nest not only reduced the distancetraveled by females during incubation recesses but alsoincreased the presence of males at the nest. However,male presence was enhanced only when females werealready at the nest, because when females leave the nestto drink, males almost invariably follow them closely;presumably because it is more important to protect thefemale than the eggs from attacks by predators orneighboring territorial males. Nonetheless, because nestdefense against foxes is more efficient by both membearsof the pair than by a single mate (Samelius andAlisauskas 2001), the increased presence of males atthe nest may have contributed to the higher success ofmanipulated nests.Given the benefits accrued to birds having access to
drinking water close to their nest, we would expect thatproximity to predictable water resources should be anabiotic factor influencing nest site selection. Not only isdrinking water more available in wetlands than in mesictundra but the intricate network of water channels inwetlands also impedes movements of terrestrial preda-tors like foxes (Lecomte et al. 2008a). In accordancewith these observations, we have found that femaleSnow Geese prefer to nest in wetlands (Lecomte et al.2008b). Although rainfall events are unpredictable, theyare not rare as in arid ecosystems and could allowindividuals nesting in lower quality territories (i.e., inmesic tundra) to attenuate, to some extent, predationrisk.
Water economy in the Arctic
Comparative studies show a link between incubationconstancy and body mass in waterfowl (Thompson andRaveling 1987). For example, large goose speciesgenerally take fewer recesses than smaller ones due totheir relatively lower mass-specific metabolic rate andgreater amount of nutrient reserves (Afton and Paulus1992). Although these authors have typically empha-sized energetic constraints to explain interspecificvariations in incubation patterns, we argue that waterrequirement may be an additional constraint on recessfrequency. Although we did not have data on recessfrequency per se, we found a positive link betweenpartial water vapor pressure in the air and nestingsuccess. We suggest that when the potential forevaporative water loss of geese is high, females have toleave their nest more often to drink, thereby increasingexposure to predation. Geese are likely to face highwater needs during reproduction. First, animals sufferan increased tendency to dehydrate in the Arctic becausecold temperatures result in low atmospheric water vaporpressure (Willmer et al. 2000). Strong winds and highsolar radiation, conditions that are frequent in the
Arctic, should further enhance the potential for evapo-rative water loss. Second, birds face major physiologicalchallenges to regulate their water balance while fastingduring incubation (Le Maho et al. 1981). In Eider Ducks(Somateria molissima L.), Criscuolo et al. (2000)suggested that birds adjusted their drinking behaviorduring incubation recesses to regulate their body waterwhile fasting. Anecdotal evidence also suggests thatSnow Geese have high evaporative water loss, and hencehigh water requirements. J. Larochelle (personal com-munication) and Ratte (1998) reported high water loss bygosling Greater Snow Geese during experiments inmetabolic chambers. In addition, geese feeding infarmlands typically return to roosting ponds in mid-day to drink under sunny conditions, though not underovercast skies or during rain (Bechet et al. 2003). Wetherefore contend that the water economy of birds,which has received little attention in comparison to foodhabits even after the seminal paper of Bartholomew andCade (1963), has been hitherto underrated in reproduc-tive studies of geese and should be investigated in otherbird species as well.
In most biomes, change in rainfall abundanceresulting from climate warming could be at least asgreat as change in air temperature, but future trends inprecipitations are especially difficult to predict (Allenand Ingram 2002). Simulations predict a mean increaseof 10% in the next 20 years (6 mm/month) around theCanadian Arctic, a larger increase than for the world asa whole (IPCC 2001, ACIA 2005). Given that goosenesting success is positively linked to rainfall, we couldexpect some changes in goose productivity in the futureunder such a scenario. The importance of water in thefunctioning of tundra ecosystems is only beginning toemerge in the literature (Hodkinson et al. 1999) and ourstudy provides a novel illustration of it. How muchArctic birds will balance their water requirements withnest predation risk in face of changing climaticconditions remains an open question for the moment.
Conclusion
Our study provides a clear demonstration that abioticfactors such as water availability and rainfall can have astrong effect on predator–prey dynamics through aninfluence on prey behavior, even in a non-arid environ-ment such as the tundra. Such relations may havesignificant consequences in tundra ecosystems where it isbecoming increasingly evident that predator–prey inter-actions play a key role (Bety et al. 2002, Gilg et al. 2003,Gauthier et al. 2004). Our results also highlight thecomplexity of these relations due to the fact that rainfallcreates unpredictible pulses of resources (water holes),and that the benefit of this resource for the prey isheterogeneous at the habitat scale. This is an emergingperspective in the literature (Holt 2008, Schmidt andOstfeld 2008), where the spatial component of ephem-eral resources is predicted to fine tune predator–preyinteractions. When variability in prey productivity is
February 2009 473WATER LIMITATION IN ARCTIC-NESTING GEESE

linked to the dynamics of the pulse, this could allowpersistance of the prey in lower quality habitats and itsexploitation by predators during interpulse intervals.
ACKNOWLEDGMENTS
Funding was provided by grants from the Natural Sciencesand Engineering Research Council of Canada to G. Gauthier,the Arctic Goose Joint Venture (Canadian Wildlife Service), theFonds Quebecois pour la Nature et les Technologies, ArcticNet,and the Northern Ecosystem Initiative (Environment Canada).Universite Laval, the Centre d’Etudes Nordiques, the SocieteProvancher, Le Fonds, Richard Bernard, and the Departmentof Indian Affairs and Northern Development providedadditional financial assistance to N. Lecomte. Logistic supportwas generously provided by the Polar Continental Shelf Project(PCSP). We thank all the people who participated in thefieldwork. We are grateful to S. D. Cote, M.-A. Giroux, M.Humphries, J. Larochelle, and J. Williams for their commentson this manuscript. We are indebted to the Hunters andTrappers Association of Pond Inlet and to Parks Canada forallowing us to work on Bylot Island. Finally, we thank M.Lambrechts for encouraging us to manipulate water availabil-ity. All methods described in the paper were approved by theAnimal Care Committee of Universite Laval following guide-lines of the Canadian Animal Care Council. This is PCSPcontribution No. 026-08.
LITERATURE CITED
ACIA. 2005. Impacts of a warming Arctic: Arctic climateimpact assessment. Cambridge University Press, New York,New York, USA.
Afton, A. D., and S. L. Paulus. 1992. Incubation and broodcare. Pages 62–108 in B. D. J. Batt, A. D. Afton, M. G.Anderson, C. D. Ankney, D. H. Johnson, J. A. Kadlec, andG. L. Krapu, editors. Ecology and management of breedingwaterfowl. University of Minnesota Press, Minneapolis,Minnesota, USA.
Agresti, A. 2002. Categorial data analysis. Second edition. JohnWiley, New York, New York, USA.
Allen, M. R., and W. J. Ingram. 2002. Constraints on futurechanges in climate and the hydrologic cycle. Nature 419:224–232.
Bartholomew, G. A., and T. J. Cade. 1963. The water economyof land birds. Auk 80:504–539.
Bechet, A., J.-F. Giroux, G. Gauthier, J. D. Nichols, and J. E.Hines. 2003. Spring hunting changes the regional movementsof migrating Greater Snow Geese. Journal of AppliedEcology 40:553–564.
Bety, J., G. Gauthier, J.-F. Giroux, and E. Korpimaki. 2001.Are goose nesting success and lemming cycles linked?Interplay between nest density and predators. Oikos 93:388–400.
Bety, J., G. Gauthier, E. Korpimaki, and J.-F. Giroux. 2002.Shared predators and indirect trophic interactions: lemmingcycles and Arctic-nesting geese. Journal of Animal Ecology71:88–98.
Blancher, P. J., and R. J. Robertson. 1987. Effect of foodsupply on the breeding biology of Western Kingbirds.Ecology 68:723–732.
Boismenu, C., G. Gauthier, and J. Larochelle. 1992. Physiologyof prolonged fasting in Greater Snow Geese (Chen caeru-lescens atlantica). Auk 109:511–521.
Boutin, S. 1990. Food supplementation experiments withterrestrial vertebrates: patterns, problems, and the future.Canadian Journal of Zoology 68:203–220.
Burger, J. 1981. A model for the evolution of mixed-speciescolonies of Ciconiiformes. Quarterly Review of Biology 56:143–167.
Burnham, K. P., and D. R. Anderson. 1998. Model selectionand inference: a practical information-theoretic approach.Springer-Verlag, New York, New York, USA.
Coe, S. J., and J. T. Rotenberry. 2003. Water availability affectsclutch size in a desert sparrow. Ecology 84:3240–3249.
Cooke, F., R. F. Rockwell, and D. B. Lank. 1995. The SnowGeese of La Perouse Bay, natural selection in the wild.Oxford University Press, New York, New York, USA.
Criscuolo, F., M. Gauthier-Clerc, G. W. Gabrielsen, and Y.Le Maho. 2000. Recess behaviour of the incubating CommonEider Somateria mollissima. Polar Biology 23:571–574.
Dickey, M.-H., G. Gauthier, and M.-C. Cadieux. 2008.Climatic effects on the breeding phenology and reproductivesuccess of an Arctic-nesting goose species. Global ChangeBiology, in press.
Dinsmore, S. J., G. C. White, and F. L. Knopf. 2002. Advancedtechniques for modeling avian nest survival. Ecology 83:3476–3488.
Doody, J. S., R. A. Sims, and M. Letnic. 2007. Environmentalmanipulation to avoid a unique predator: drinking holeexcavation in the agile wallaby, Macropus agilis. Ethology113:128–136.
Ferns, P. N., and S. A. Hinsley. 1995. Importance oftopography in the selection of drinking sites by sandgrouse.Functional Ecology 9:371–375.
Fontaine, J. J., and T. E. Martin. 2006. Parent birds assess nestpredation risk and adjust their reproductive strategies.Ecology Letters 9:428–434.
Gates, D. M. 1980. Biophysical ecology. Springer-Verlag, NewYork, New York, USA.
Gauthier, G., J. Bety, J.-F. Giroux, and L. Rochefort. 2004.Trophic interactions in a High Arctic Snow Goose colony.Integrative and Comparative Biology 44:119–129.
Gauthier, G., L. Rochefort, and A. Reed. 1996. Theexploitation of wetland ecosystems by herbivores on BylotIsland. Geoscience Canada 23:253–259.
Gibbs, H. L., and P. R. Grant. 1987. Ecological consequencesof an exceptionally strong El-Nino event on Darwin’s finches.Ecology 68:1735–1746.
Gilg, O., I. Hanski, and B. Sittler. 2003. Cyclic dynamics in asimple vertebrate predator–prey community. Science 302:866–868.
Goff, J. A., and S. Gratch. 1946. Low-pressure properties ofwater from )160 to 2128 F. Pages 95–122. 52nd annualmeeting of the American Society of Heating and VentilatingEngineers, New York, New York, USA.
Graham-Sauve, M. 2008. Effets en cascade du climat et desinteractions trophiques indirectes sur les plantes de latoundra par l’oie des neiges. M.S. Thesis. Universite duQuebec, Rimouski, Canada.
Hardin, J. W., and J. M. Hilbe. 2003. Generalized estimatingequations. Chapman and Hall/CRC, Boca Raton, Florida,USA.
Hilton, G. M., G. D. Ruxton, and W. Cresswell. 1999. Choiceof foraging area with respect to predation risk in redshanks:the effects of weather and predator activity. Oikos 87:295–302.
Hodkinson, I. D., N. R. Webb, J. S. Bale, and W. Block. 1999.Hydrology, water availability and tundra ecosystem functionin a changing climate: the need for a closer integration ofideas? Global Change Biology 5:359–369.
Holt, R. D. 2008. Theoretical perspectives on resource pulses.Ecology 89:671–681.
IPCC. 2001. Climate change 2001: synthesis report. Acontribution of working groups I, II, and III to the thirdassessment report of the intergovernmental panel on climatechange. Cambridge University Press, Cambridge, UK.
Kahlert, J. 2003. The constraint on habitat use in wing-moulting Greylag Geese Anser anser caused by anti-predatordisplacements. Ibis 145:E45–E52.
NICOLAS LECOMTE ET AL.474 Ecology, Vol. 90, No. 2

Klaassen, M. 2004. May dehydration risk govern long-distancemigratory behaviour? Journal of Avian Biology 35:4–6.
Lada, H., J. R. Thomson, R. Mac Nally, G. Horrocks, andA. C. Taylor. 2007. Evaluating simultaneous impacts of threeanthropogenic effects on a floodplain-dwelling marsupialAntechinus flavipes. Biological Conservation 134:527–536.
Le Maho, Y., H. Vu Van Kha, H. Koubi, G. Dewasmes, J.Girard, P. Ferre, and M. Cagnard. 1981. Body composition,energy expenditure, and plasma metabolites in long-termfasting geese. American Journal of Physiology 241:E342–E354.
Lecomte, N., V. Careau, G. Gauthier, and J.-F. Giroux. 2008a.Predator behaviour and predation risk in the heterogeneousArctic environment. Journal of Animal Ecology 77:439–447.
Lecomte, N., G. Gauthier, and J.-F. Giroux. 2008b. Breedingdispersal in a heterogeneous landscape: the influence ofhabitat and nesting success in Greater Snow Geese.Oecologia 155:33–41.
Lepage, D., G. Gauthier, and S. Menu. 2000. Reproductiveconsequences of egg-laying decisions in Snow Geese. Journalof Animal Ecology 69:414–427.
Loveridge, A. J., J. E. Hunt, F. Murindagomo, and D. W.Macdonald. 2006. Influence of drought on predation ofelephant (Loxodonta africana) calves by lions (Panthera leo)in an African wooded savannah. Journal of Zoology 270:523–530.
Martin, T. E. 1987. Food as a limit on breeding birds—a life-history perspective. Annual Review of Ecology and System-atics 18:453–487.
Martin, T. E. 1995. Avian life history evolution in relation tonest sites, nest predation, and food. Ecological Monographs65:101–127.
Martin, T. E., and C. K. Ghalambor. 1999. Males feedingfemales during incubation. I. Required by microclimate orconstrained by nest predation? American Naturalist 153:131–139.
Morrison, S. A., and D. T. Bolger. 2002. Variation in asparrow’s reproductive success with rainfall: food andpredator-mediated processes. Oecologia 133:315–324.
Myers, P., L. L. Master, and R. A. Garrett. 1985. Ambienttemperature and rainfall: an effect on sex ratio and litter sizein deer mice. Journal of Mammalogy 66:289–298.
Nagy, L. R., and R. T. Holmes. 2005. Food limits annualfecundity of a migratory songbird: an experimental study.Ecology 86:675–681.
Newton, I. 1998. Population limitation in birds. AcademicPress, San Diego, California, USA.
Ogutu, J. O., H. P. Piepho, H. T. Dublin, N. Bhola, and R. S.Reid. 2008. Rainfall influences on ungulate populationabundance in the Mara-Serengeti ecosystem. Journal ofAnimal Ecology 77:814–829.
Ostfeld, R. S., and F. Keesing. 2000. Pulsed resources andcommunity dynamics of consumers in terrestrial ecosystems.Trends in Ecology and Evolution 15:232–237.
Post, E., R. O. Peterson, N. C. Stenseth, and B. E. McLaren.1999. Ecosystem consequences of wolf behavioural responseto climate. Nature 401:905–907.
Ratcliffe, N., S. Schmitt, and M. Whiffin. 2005. Sink or swim?Viability of a Black-tailed Godwit population in relation toflooding. Journal of Applied Ecology 42:834–843.
Ratte, J. 1998. Thermoregulation et croissance chez les oisonsde la grande oie des neiges, Anser caerulescens atlantica. M.S.Thesis. Universite Laval, Quebec, Canada.
Redfern, J. V., R. Grant, H. Biggs, and W. M. Getz. 2003.Surface-water constraints on herbivore foraging in theKruger National Park, South Africa. Ecology 84:2092–2107.
Reed, A., R. J. Hughes, and H. Boyd. 2002. Patterns ofdistribution and abundance of Greater Snow Geese on BylotIsland, Nunavut, Canada 1983–1998. Wildfowl 53:53–65.
Reed, A., R. J. Hughes, and G. Gauthier. 1995. Incubationbehavior and body-mass of female Greater Snow Geese.Condor 97:993–1001.
Reed, E. T., G. Gauthier, and R. Pradel. 2005. Effects of neckbands on reproduction and survival of female Greater SnowGeese. Journal of Wildlife Management 69:91–100.
Salter, K. C., and R. Fawcett. 1993. The ART test ofinteraction: a robust and powerful rank test of interactionin factorial models. Communications in Statistics: Simulationand Computation 22:137–153.
Samelius, G., and R. T. Alisauskas. 2001. Deterring Arctic foxpredation: the role of parental nest attendance by LesserSnow Geese. Canadian Journal of Zoology 79:861–866.
Samelius, G., and R. T. Alisauskas. 2005. Sex-biased costs innest defence behaviours by Lesser Snow Geese (Chencaerulescens): consequences of parental roles? BehavioralEcology and Sociobiology 59:805–810.
SAS Institute. 2002. SAS guide for personal computers. Version9. SAS Institute, Cary, North Carolina, USA.
Schmidt, K. A. 1999. Foraging theory as a conceptualframework for studying nest predation. Oikos 85:151–160.
Schmidt, K. A., and R. S. Ostfeld. 2008. Numerical andbehavioral effects within a pulse-driven system: consequencesfor shared prey. Ecology 89:635–646.
Siikamaki, P. 1998. Limitation of reproductive success by foodavailability and breeding time in Pied Flycatchers. Ecology79:1789–1796.
Thompson, K. R., and R. W. Furness. 1991. The influence ofrainfall and nest-site quality on the population-dynamics ofthe Manx Shearwater Puffinus puffinus on Rhum. Journal ofZoology 225:427–437.
Thompson, S. C., and D. G. Raveling. 1987. Incubationbehavior of Emperor Geese compared with other geese:interactions of predation, body size, and energetics. Auk 104:707–716.
Tieleman, B. I., J. B. Williams, and G. H. Visser. 2004. Energyand water budgets of larks in a life history perspective:parental effort varies with aridity. Ecology 85:1399–1410.
Valeix, M., H. Fritz, R. Matsika, F. Matsvimbo, and H.Madzikanda. 2007. The role of water abundance, thermo-regulation, perceived predation risk and interference. AfricanJournal of Ecology 46:402–410.
Williams, J. B., and B. I. Tieleman. 2005. Physiologicaladaptation in desert birds. Bioscience 55:416–425.
Willmer, P., G. Stone, and I. A. Johnston. 2000. Environmentalphysiology of animals. Blackwell Scientific Publications,Oxford, UK.
Yang, L. H., J. L. Bastow, K. O. Spence, and A. N. Wright.2008. What can we learn from resource pulses? Ecology 89:621–634.
February 2009 475WATER LIMITATION IN ARCTIC-NESTING GEESE





























