Embed Size (px)
Citation preview

A MarR-type regulator directly activates transcription 1
from the Brucella abortus virB promoter by sharing a 2
redundant role with HutC 3
4 Rodrigo Sieira1*, Gastón M. Arocena 1, Angeles Zorreguieta 1,3, Diego J. Comerci 2, and 5
Rodolfo A. Ugalde2†. 6
7
Author affiliation: 1 Fundación Instituto Leloir, IIBBA-CONICET, Av. Patricias 8 Argentinas 435, Buenos Aires C1405BWE, Argentina. 2 Instituto de Investigaciones 9 Biotecnológicas, Universidad Nacional de General San Martín, IIB-INTECH-10 CONICET, San Martín 1650, Buenos Aires, Argentina. 3 Facultad de Ciencias Exactas 11 y Naturales-Departamento de Química Biológica, Universidad de Buenos Aires, 12 Argentina. 13 14
15
16
17
18
19
20
21
22
23
24
25
*Corresponding author: Fundación Instituto Leloir (IIBBA-CONICET), Av. Patricias 26
Argentinas 435, Buenos Aires C1405BWE, Argentina. Tel. (54 11) 5238-7500. E-mail: 27
[email protected] 28 †: Deceased 17 august 2009. 29
30
31
32
Copyright © 2012, American Society for Microbiology. All Rights Reserved.J. Bacteriol. doi:10.1128/JB.01007-12 JB Accepts, published online ahead of print on 21 September 2012
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

2
Abstract 33
Type-IV Secretion Systems (T4SS) are multiprotein structures that direct the 34
translocation of specific molecules across the bacterial cell envelope. As in other 35
bacteria, pathogenicity of the genus Brucella essentially depends on the integrity of the 36
T4SS-encoding virB operon, whose expression is regulated by multiple transcription 37
factors belonging to different families. Previously, we identified IHF and HutC, two 38
direct regulators of the virB genes that were isolated from total protein extracts of 39
Brucella. Here, we report the identification of MdrA, a third regulatory element that was 40
isolated using the same screening procedure. This transcription factor, which belongs to 41
the MarR-family of transcriptional regulators, binds at two different sites of the virB 42
promoter and regulates expression in a growth phase-dependent manner. Like other 43
members of the MarR family, specific ligands were able to dissociate MdrA from DNA 44
in vitro. Determination of the MdrA-binding sites by DNase I Footprinting and analyses 45
of protein-DNA complexes by electrophoresis mobility shift assays (EMSA) showed 46
that MdrA competes with IHF and HutC for the binding to the promoter because their 47
target DNA sequences overlap. Unlike IHF, both MdrA and HutC bound to the 48
promoter without inducing bending of DNA. Moreover, these two latter transcription 49
factors activated virB expression to similar extents and in doing so, they are functionally 50
redundant. Taken together, our results show that MdrA is a regulatory element that 51
directly modulates activity of the virB promoter and is probably involved in 52
coordinating gene expression in response to specific environmental signals. 53
54
55
56
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

3
INTRODUCTION 57
Type-IV Secretion Systems (T4SS) are multiprotein structures that direct the 58
translocation of specific molecules across the bacterial cell envelope (3). During 59
evolution, these machineries have specialized to secrete different kind of substrates, 60
which led to the emergence of the different biological functions that T4SSs can exert. 61
Both the Escherichia coli Tra and the Agrobacterium tumefaciens VirB T4SSs are 62
responsible for secreting protein-DNA complexes during plasmid conjugation or 63
oncogenic T-DNA translocation, respectively (48, 54). Instead, other T4SSs such as the 64
Helicobacter pylori Cag system, the Legionella pneumophila Dot/Icm system, or the 65
Bordetella pertussis Ptl system, have evolved to deliver protein substrates towards the 66
eukaryotic host, where they subvert cellular processes and favor the development of the 67
bacterial infection (8, 44). As in all these pathogenic bacteria, virulence of Brucella is 68
also fully dependent on the integrity of a T4SS, since deletion of one or more T4SS-69
encoding virB genes completely abrogates the ability of this genus of facultative 70
intracellular bacteria to colonize its eukaryotic host (30, 47). 71
Brucella comprises many species which are the causative agent of a worldwide 72
distributed zoonotic disease known as brucellosis. The members of Brucella differ in 73
their host preference, affecting mammals that range from rodents to cetaceans (52). 74
Brucella abortus, the causal agent of bovine brucellosis, is one of the species that can be 75
transmitted from infected animals to humans through inhalation, contact with skin 76
wounds or mucosa, or by consumption of contaminated dairy products. In the natural 77
host, brucellosis produces sterility in males and abortion in pregnant females, whereas 78
in humans it causes debilitating symptoms and complications that can lead to death 79
(16). 80
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

4
The ability of Brucella to infect and persist within the mammalian host relies on 81
different mechanisms that allow these bacteria to overcome the innate and adaptive 82
immune host responses. Among all these strategies, bacterial survival and intracellular 83
replication into the host professional phagocytes is a key process underlying brucellosis. 84
After entering the host macrophages, the bacterium is located in the lumen of the so-85
called Brucella-containing vacuole (BCV) (32). During the first stages of the 86
intracellular infection, this compartment undergoes sustained interactions with 87
endoplasmic reticulum (ER) membranes. This process, which depends essentially on the 88
activity of the VirB system, is necessary for the biogenesis of an ER-derived bacterial 89
replication compartment, since abrogation of the early BCV-ER interactions prevents 90
Brucella from multiplying intracellularly and leads to bacterial degradation in 91
lysosomes (7). Recently, screening procedures based on different methods led to the 92
identification of proteins that are translocated into the host cell cytoplasm in a VirB-93
dependent manner (10, 22). Until now, the function of these substrates in the 94
establishment of the host cell infection have not yet been described except for RicA, a 95
protein that interacts with and recruits the small GTPase Rab2 into BCVs during the 96
intracellular trafficking of Brucella (9). 97
The first reported evidence of intracellular regulation of the VirB T4SS showed 98
that expression of the virB genes is induced after internalization of the bacterium in 99
macrophage cell lines (4). Such induction is dependent on the acidification of the BCV, 100
and reaches the maximal level of expression at 5 h post infection. Subsequently, 101
expression of the virB operon is rapidly turned off, which suggests that the system is 102
controlled by regulatory mechanisms that respond to specific signals sensed by the 103
bacterium within the host cell (46). 104
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

5
The main regulatory protein controlling expression of the Brucella virB genes is 105
VjbR, a Quorum Sensing (QS)-related regulator that binds the virB promoter (PvirB) at 106
position -94 relative to the transcription start site (2, 12). In addition, many other 107
regulators were shown to directly or indirectly influence virB expression, including the 108
stringent response mediator Rsh and transcription factors belonging to different families 109
(13, 18, 37). In addition, although their binding sites in the virB promoter have not yet 110
been reported, it was recently shown that both the Two-component regulator BvrR and 111
the second QS-related transcription factor BabR/BlxR participate in regulation of virB 112
expression by directly interacting with PvirB, as demonstrated by pull-down experiments 113
and electrophoresis mobility shift assays (EMSA), respectively (6, 25). 114
In previous works from our group, we identified two transcriptional regulators that 115
directly contribute to induction of virB expression: (i) IHF, a nucleoid associated protein 116
that binds to PvirB at position -162.5 and induces a DNA bend that is necessary for the 117
promoter activation (46); and (ii) HutC, a GntR-family regulator that binds at position -118
188 and links virB expression to histidine catabolism (45). Here, we report the 119
identification of MdrA, a third regulatory element that was isolated using the same 120
screening procedure. Our experiments showed that this transcription factor binds at two 121
different sites of PvirB and regulates virB expression in a growth phase-dependent 122
manner. Analysis of promoter activities under different mutant backgrounds revealed 123
that MdrA exerts its regulatory role in redundancy with the function of HutC. Besides, 124
consistently with both the architecture and localization of the MdrA-binding sites within 125
the promoter region, we also observed that this transcription factor competes with IHF 126
and HutC for the binding to PvirB. As is usually the case with MarR family members, our 127
experiments showed that the presence of a specific ligand dissociates MdrA from a 128
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

6
target DNA binding site. Such ligand-induced disruption of promoter binding indicates 129
that this process may allow MdrA to receive and respond to environmental signals. 130
. 131
132
133
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

7
MATERIALS AND METHODS 134
Media and culture conditions 135
Brucella strains were grown in triptic soy broth (TSB); or in minimal media Gerhardt-136
Wilson (15) or MM1 (45) at pH 4.5, 5.5, or 7.0. Bacteria were cultured at 37˚C in a 137
rotary shaker (250 rpm). Media were supplemented with kanamycin (50 μg ml-1) and/or 138
nalidixic acid (5 μg ml-1), as needed. Sodium Deoxycholate (Sigma) was eventually 139
added to the media at final concentration of 100, 250, 500, 750, or 1000 μM. 140
Escherichia coli strains were cultured in Luria-Bertani (LB) broth at 37°C in a rotary 141
shaker at 250 rpm. Media were supplemented with ampicilin (100 μg ml-1) or 142
kanamycin (25 μg ml-1), as needed. 143
144
Isolation and identification of MdrA 145
The protein detected by EMSA was isolated from exponential phase cultures (OD600 ~ 146
0.5) of the avirulent strain B. abortus Δpgm (51). Bacteria were harvested and disrupted 147
as described previously (46). After ultracentrifugation and filtration, the total protein 148
extract was fractionated by ammonium sulphate precipitation. The different fractions 149
were analyzed by EMSA using probe vir-up or the control probe B10. The positive 150
fraction (0-45% ammonium sulphate saturation) was suspended in 35 mM phosphate 151
buffer (pH 6.8), 3 mM β-mercaptoethanol and dialyzed against the same buffer O.N. at 152
4°C. The solution was loaded onto a Mono-S column and eluted with a linear gradient 153
of 35 mM phosphate buffer (pH 6.8), 3 mM β-mercaptoethanol, 1M NaCl. DNA-154
binding activity of the fractions was again analyzed by EMSA using the probes 155
described above. Positive fractions were subjected to affinity chromatography: the 156
biotinylated probe biot-vir-up, which corresponds to the -430 to -202 region of PvirB, 157
was constructed by PCR using Taq, the 5´-biotinylated primer pvd228, the primer pirup 158
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

8
and plasmid genomic DNA of B. abortus 2308 as template. The biotinylated control 159
probe was constructed as described previously (45). The biotinylated probes were 160
bound to streptavidin paramagnetic spheres (Promega) and a binding reaction was 161
performed using binding buffer and the positive fractions of the Mono-S column. After 162
two washes with binding buffer with 0.2 M NaCl, the DNA-bound proteins were eluted 163
with 0.85 M NaCl and analyzed by 12.5% SDS-PAGE. The gel was silver-stained with 164
a mass spectrometry compatible method for visualizing the protein bands. A band which 165
was observed with the probe biot-vir-up but was absent in samples from the control 166
probe was excised form the gel and analyzed by mass spectrometry in Vital probes Inc. 167
(PA, USA). 168
169
Construction of plasmids 170
pK18mob-sacB-ΔmdrA: two PCRs were carried out using Pfx (Invitrogen), genomic 171
DNA of B. abortus 2308 as template, and primers SpeMarA (5’- 172
GGACTAGTTGACGATATGTTTGCCGCAT-3’) and COMMarA (5’- 173
CTGCAAGAAGGAGAGTATCGAATGCAGGCGCTCGACAAGC-3’), or SpeMarB (5’- 174
GGACTAGTAACCGCTCTTGCGGCAATC-3’) and COMMarB (5’- 175
TCGATACTCTCCTTCTTGCAGTTTATGGCGATGAACAAGTCG-3’). Both PCR 176
products, corresponding to two 400-bp flanking regions of mdrA, were annealed and 177
used as templates for a PCR performed with primers SpeMarA and SpeMarB. The 178
product was digested with SpeI and cloned into the kanamycin-resistant plasmid 179
pK18mob-sacB (42). 180
Complementation plasmid pK18mob-sacB-Knock-in-mdrA: a PCR was carried out 181
using Pfx, genomic DNA of B. abortus 2308 as template, and primers SpeMarA (5’- 182
GGACTAGTTGACGATATGTTTGCCGCAT-3’) and SpeMarB (5’- 183
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

9
GGACTAGTAACCGCTCTTGCGGCAATC-3’). The PCR product, which contains the 184
wild type-sequence of the mdrA gene and two 400-bp flanking regions, was digested 185
with SpeI and cloned into the kanamycin-resistant plasmid pK18mob-sacB. 186
Expression vector pQE-31-mdrA: a PCR reaction was performed using Pfx, genomic 187
DNA of B. abortus 2308 as template, and primers BamMarII (5’-188
CGGGATCCGACCAATACCCAGCGCAAGAT-3’) and HindMar (5’- 189
GGTACCTCAGCCGCGAGATGGCGT-3’). The PCR product were digested with BamHI 190
and HindIII, and cloned into plasmid pQE-31 (Qiagen). 191
192
Construction of strain B. abortus ΔmdrA 193
Plasmid pK18mob-sacB-ΔmdrA was transferred to B. abortus 2308 by biparental 194
conjugation. Kanamycinr colonies were selected as single-homologous recombinants. 195
Selection with sucrose, excision of plasmids and generation of deletion mutants was 196
performed as described previously (46). PCR analyses of kanamycins colonies were 197
carried out with primers SpeMarA and SpeMarB to identify clones that contain the 198
deletion of mdrA. 199
200
Construction of the double mutant deletion strain B. abortus ΔhutC ΔmdrA 201
Plasmid pK18mob-sacB-ΔmdrA was transferred to the strain B. abortus ΔhutC (45) by 202
biparental conjugation. Kanamycinr colonies were selected as single-homologous 203
recombinants. After selection with sucrose and excision of the plasmid, a PCR analysis 204
of kanamycins colonies was carried out with primers SpeMarA and SpeMarB to identify 205
clones that contain the deletion of mdrA in the ΔhutC mutant background. 206
207
Construction of the Knock-in complemented strain B. abortus ΔhutC ΔmdrA-KI 208
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

10
Plasmid pK18mob-sacB-Knock-in-mdrA was transferred to the strain B. abortus 209
ΔhutC ΔmdrA by biparental conjugation. Kanamycinr colonies were selected as single-210
homologous recombinants. After selection with sucrose and excision of the plasmid, a 211
PCR analysis of kanamycins colonies was carried out with primers SpeMarA and 212
SpeMarB to identify clones that incorporated the wild type mdrA gene. 213
214
Construction of strains containing a single PvirB-lacZ chromosomal transcriptional 215
fusion. 216
Plasmid pK18mob-PvirB-lacZ was transferred by biparental conjugation into the wild 217
type strain B. abortus 2308 or into strain B. abortus ΔmdrA, B. abortus ΔhutC, B. 218
abortus ΔhutC ΔmdrA, or B. abortus ΔhutC ΔmdrA-KI. Kanamycinr colonies were 219
selected as single-homologous recombinants. 220
221
Electrophoresis Mobility Shift Assays (EMSA) 222
32P-labeling of probes, incubations and running conditions were performed as described 223
previously (46). The radiolabeled probe vir-up was constructed using Taq (Invitrogen), 224
primers pvirup (5’-ATGACAGGCATATTTCAAC-3’) and pvd228 (5’-225
GTGATTTTCATATTTTTGCGG-3’) and genomic DNA of B. abortus 2308 as 226
template. The radiolabeled probes vir-down and vir-down-ihf- were constructed by PCR 227
using primers pvd229 and pvirdownI, and genomic DNA of B. abortus 2308 or plasmid 228
pBluescript-PvirB-IHF-sacB/R as template, respectively, as described previously (46). The 229
radiolabeled control probe B10 was constructed with primers B10Qu and B10d2, as 230
described previously (45). The radiolabeled probe 286/59 was constructed using primers 231
pvu144 (5’-CGGGATCCGATTCTTGGTCGGGTTAC-3’) and pvirdownIII (5’-232
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

11
TGTTTAAGCCGATATATGGAC-3’), and genomic DNA of B. abortus 2308 as 233
template. 234
The unlabeled competitors were constructed by PCR using Taq, genomic DNA of B. 235
abortus 2308 as template, and primers pvirup and pvd228 for competitor up; primers 236
pvu77 (5’-CGGGATCCGATGCCGCCTAATGGAGC-3’) and pvd228 for competitor 237
77; primers pvu144 and pvd228 for competitor 144; and primers pvu229 and pvirdownI 238
for competitor down. The unlabeled competitors A1 and B1 were constructed as 239
described previously (46). Binding reactions were performed in binding buffer [15 mM 240
Tris-HCl (pH 8.0), 0.5 mM EDTA, 10 μg ml-1 bovine serum albumin (BSA), 1 mM 241
DTT, 30 mM KCl, 6% glycerol, 50 μg ml-1 salmon sperm DNA]. 242
243
DNase I Footprinting 244
Labelling reactions, purification of probes, binding reactions, DNase I digestion, 245
purification of digested fragments, and electrophoresis conditions were performed as 246
described previously (46). Probe vir-upII was constructed using Taq, primer pvd228, 247
the 32P-labeled primer pvirup, and genomic DNA of B. abortus 2308 as template. Probe 248
vir-downII was constructed using primer pvu144, the 32P-labeled primer pvirdownI, and 249
genomic DNA of B. abortus 2308 as template. Probe vir-upIII was constructed using 250
primer pvirup, the 32P-labeled primer pvirdownIII, and genomic DNA of B. abortus 251
2308 as template. 252
253
Expression and purification of recombinant proteins 254
Recombinant IHF and HutC were prepared as described previously (45-46). 255
Recombinant MdrA was prepared as follows. Plasmid pQE-31-mdrA was transferred 256
into E. coli M15 (pREP4) (Qiagen). Transformed bacteria were cultured in LB until 257
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

12
DO600 = 0.6 and induced with 1 mM IPTG. After 4 h, bacteria were harvested, 258
suspended in lysis buffer [20 mM Tris-HCl (pH 7.6), 1 mM PMSF] and disrupted by 259
sonication. After centrifugation NaCl was added to a final concentration of 0.5 M and 260
the sample was loaded into a Hi-Trap Nickel-chelating column (Amersham 261
Biosciences). After washing with buffer B [20 mM Tris-HCl (pH 7.6), 0.5 M NaCl, 50 262
mM imidazole] the column was equilibrated with buffer A [20 mM Tris-HCl (pH 7.6), 263
0.5 M NaCl] and eluted with a linear gradient of buffer C [20 mM Tris-HCl (pH 7.6), 264
0.5 M NaCl, 1 M Imidazole]. Eluates were analyzed in a 12.5% SDS-PAGE, and the 265
fractions containing the recombinant protein MdrA (purity near to 95%) were pooled 266
and dialized against buffer D [20 mM Tris-HCl (pH 7.6), 0.5 M NaCl, 3 mM β-267
mercaptoethanol]. 268
269
β-galactosidase activity determinations 270
β-Gal activity was determined in whole bacterial cells as described (46), and it was 271
expressed as Miller Units [A420 / volume × OD600] ×100. 272
273
274
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

13
RESULTS 275
Isolation and identification of MdrA 276
In previous studies, two transcription factors implicated in the control of 277
expression of the virB operon were isolated by affinity chromatography from B. abortus 278
total protein extracts (45-46). Such analyses were performed by using DNA-probes that 279
contained sequences corresponding to positions -201 to +24 relative to the transcription 280
start site of the virB promoter (PvirB). In order to identify possible additional regulatory 281
proteins interacting with further upstream regions, we performed EMSA analyses using 282
crude protein extracts and a radioactive-labeled probe corresponding to positions -430 283
to -202 of PvirB. As shown in Figure 1, a protein-DNA complex was detected using total 284
protein extracts of B. abortus grown in rich medium (TSB) until exponential phase, 285
whereas no signal was observed with stationary-phase extracts. To isolate the factor 286
responsible for the observed complex, proteins were fractionated by ammonium 287
sulphate precipitation and ion-exchange chromatography, followed by EMSA analysis 288
of each fraction during the purification process (see Materials and Methods). 289
Subsequently, the partially purified protein fractions were subjected to affinity 290
chromatography using a biotin-labeled probe vir-up or a non-related control probe 291
bound to streptavidin-coated paramagnetic spheres, and the corresponding eluates were 292
analyzed by SDS-PAGE. Using this procedure we observed a specific band, which was 293
excised from the gel and identified by mass spectrometry as YP_414216, a protein 294
belonging to the MarR family of transcriptional regulators (14). Because of its 295
biochemical properties and its role on regulation of virB expression (see below), we 296
called this protein MdrA (for MarR-like sodium deoxicholate-responsive activator). As 297
revealed by BLAST analyses, MdrA shares 29% identity and 44% similarity with the 298
Escherichia coli prototypical member MarR (24), whereas it displayed slightly higher 299
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

14
homology with other representative MarR members such as PecS, SlyA, and RovA 300
(33% identity and 54% similarity; 30% and 49%; 31% and 50%; respectively) (21, 27, 301
34). As shown in Figure 2, a multiple sequence alignment indicated that MdrA displays 302
a high degree of amino acid conservation at postions corresponding to the winged-helix 303
turn helix DNA-binding domain [β1-α3- α4(recognition helix)-β2-W1(wing)-β3], 304
which is characteristic of this family of transcriptional regulators (1). 305
306
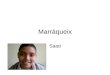
MdrA interacts with PvirB at two different binding sites 307
To study the MdrA-PvirB interaction, a His-tagged recombinant protein was 308
expressed and assayed by EMSA using different radioactive-labeled probes (see Fig. 309
3A). As shown in Fig. 3B, recombinant MdrA interacted with probe vir-up showing an 310
electrophoretic mobility similar to that of the native nucleoprotein complex, whereas no 311
signal was observed with the control probe B10. Incubation of MdrA with probe vir-312
down also resulted in the formation of a protein-DNA complex, which indicated that 313
this regulator specifically recognizes two separated regions of the promoter (Fig. 3B). 314
On the other hand, incubation of MdrA with a probe that lacks the IHF-binding site 315
produced no signal by EMSA (Fig. 3B), suggesting that the MdrA-binding site overlaps 316
with that of IHF. Competition experiments performed in the presence of an excess of 317
different unlabeled fragments indicated that the sequences recognized by MdrA in PvirB 318
are located at two regions residing between positions -353 and -286, and between -201 319
and -130 (Fig. 3C-D). Hereafter, we will refer to the downstream- and upstream-located 320
elements recognized by this protein in PvirB as the MdrA-binding sites I and II, 321
respectively. 322
323
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

15
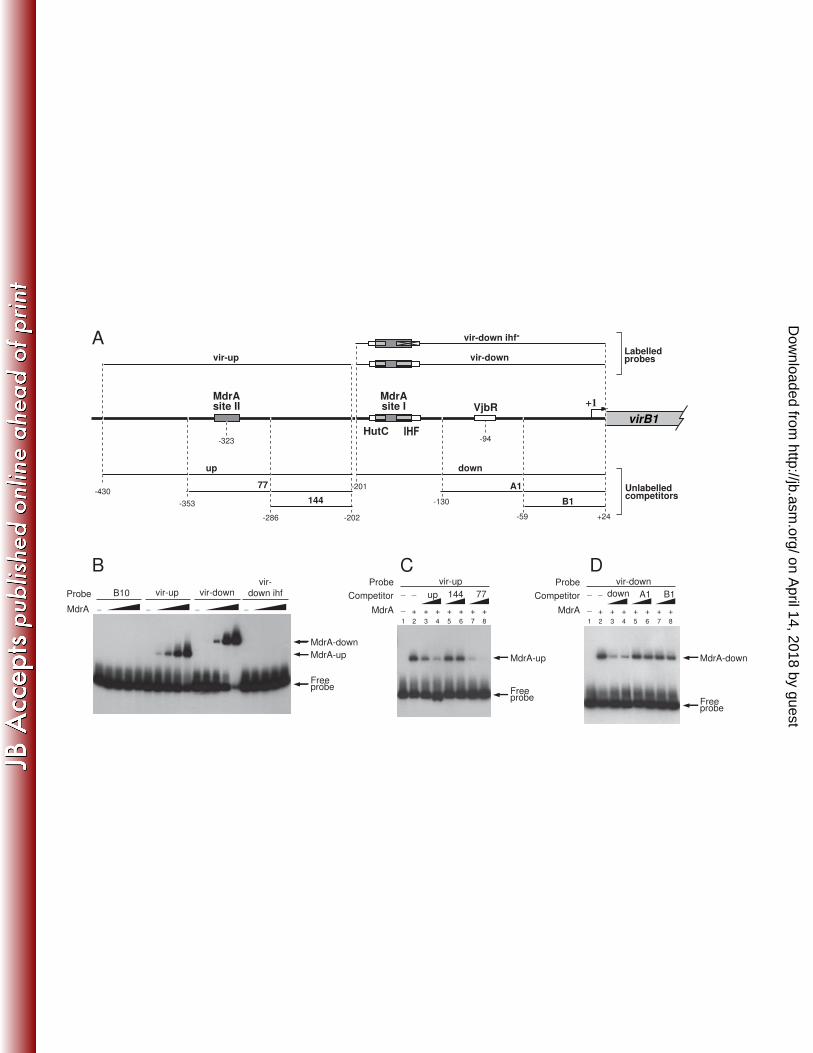
Analysis of the interaction of MdrA, IHF and HutC with the downstream region of 324
PvirB 325
The above experiments showed that MdrA specifically binds to a 71-bp region of 326
the promoter which also contains the binding sites for IHF and HutC. In previous 327
works, we found that IHF and HutC compete with one another during binding to PvirB 328
because their binding sites overlap (45-46). Accordingly, we next asked if binding of 329
MdrA to PvirB affects the interaction of IHF or HutC with the promoter. To address this 330
question, we incubated probe vir-down with each of the three pairs of proteins, or with 331
all three proteins together. We then carried out EMSA on the different protein 332
combinations. Figure 4B shows that, separately, each of the regulators generated a 333
signal corresponding to a binary protein-DNA complex, whereas no ternary complexes 334
were observed with any of the combinations of two or more proteins. These results 335
indicated that MdrA competes with IHF and HutC for the binding to the probe vir-336
down, suggesting that the MdrA-binding site I overlaps those of the two other 337
regulators. The same results were obtained when EMSAs were carried out with probe 338
289/59, whose length is the same as that of probe vir-down but the location of the 339
binding sites for MdrA, HutC and IHF are close to the center of the probe rather than 340
being near one end (data not shown).. However, the electrophoretic mobility of the 341
complex between IHF and probe 289/52 was substantially slower than that of the 342
complex between IHF and probe vir-down (Fig. 4B-C). The reduced mobility of the 343
IHF-289/52 complex is because IHF bends DNA as it binds to it, and the effect of 344
bending on migration is more pronounced when the binding site is located near the 345
center of the molecule (38). On the other hand, unlike IHF, the relative mobility of the 346
MdrA- and HutC-nucleoprotein complexes remained constant regardless of the probe 347
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

16
used, indicating that these two latter transcriptional regulators do not induce bending of 348
DNA upon binding (Fig. 4B-C). 349
350
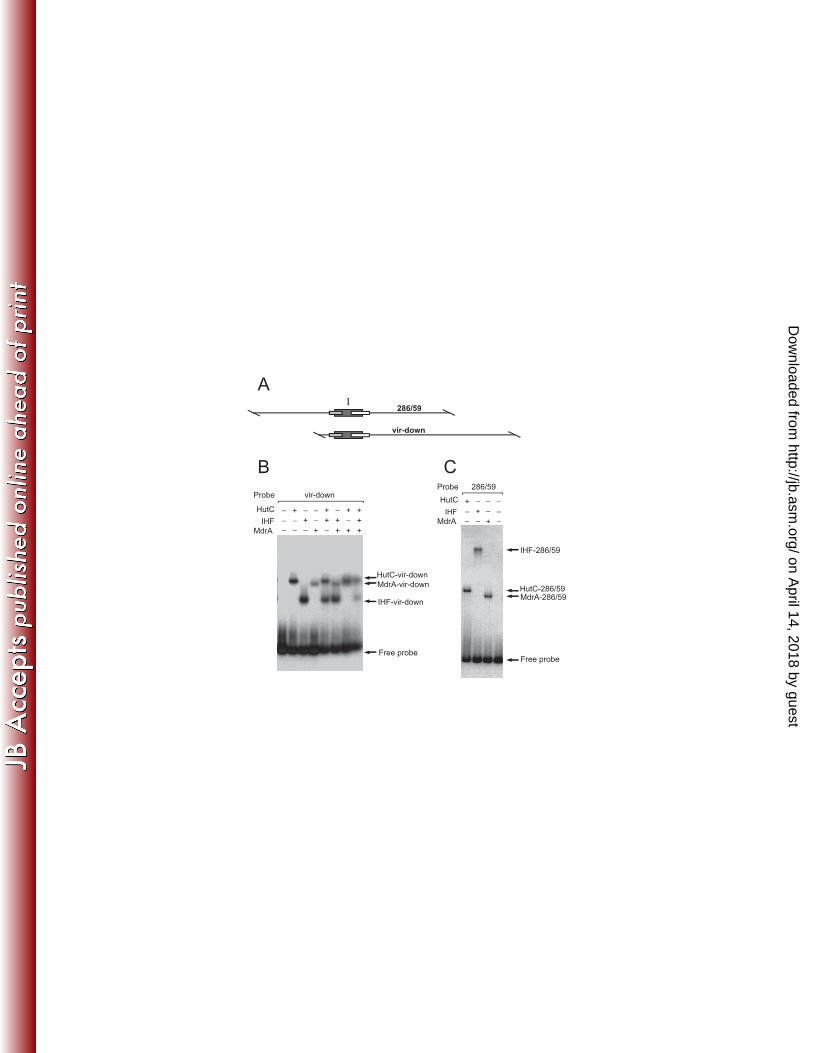
Determination of the MdrA-binding sites in PvirB 351
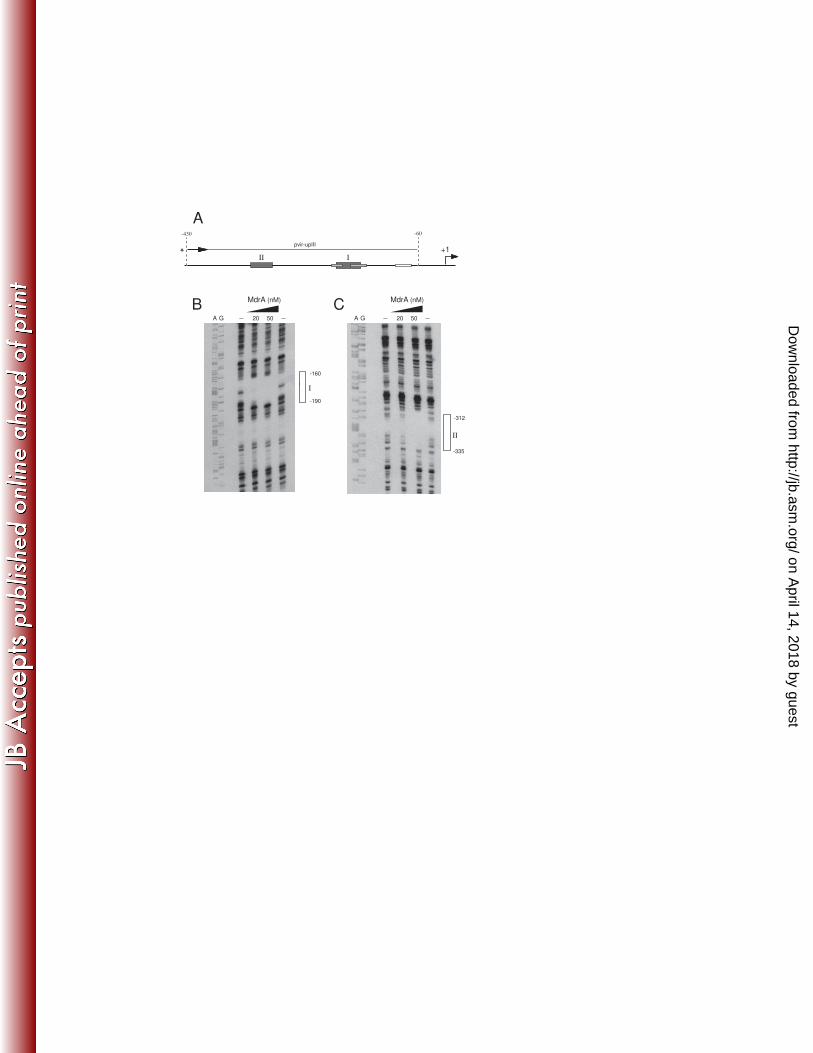
To identify both MdrA-binding sites I and II, we performed DNAse I Footprinting 352
experiments using probes corresponding to different regions of PvirB: (a) a 228-bp 353
fragment containing the upstream region of the promoter that was labeled at the 5’ end 354
of the top strand; and (b) a 312-bp fragment containing the downstream region that was 355
labeled at the 5’ end of the bottom strand (Fig. 5A). As shown in Fig. 5B, it was 356
observed that MdrA bound to a 30-bp region extending from position -190 to -160, 357
which corresponds to the MdrA-binding site I (Fig. 5B). DNase I Footprinting 358
experiments performed with the probe that contains sequences of the upstream region of 359
PvirB showed that MdrA protected a 24-pb region that extends from position -335 to -360
312, which corresponds to the MdrA-binding site II (Fig. 5C). When a 370-bp fragment 361
was end-labeled at the top strand and used as probe (Fig. 6A), it was observed that 362
MdrA protected two regions corresponding to the MdrA-binding sites I and II (Fig.6B-363
C), which indicated that this transcription factor is able to bind simultaneously to both 364
operator sequences. 365
The DNAseI Footprinting experiments showed that the MdrA-binding site I 366
overlaps those of both HutC and IHF (Fig. 5A), consistently with the observation that 367
MdrA competed with these two transcriptional regulators for the binding to probe vir-368
down (Fig. 4B). The analysis of the protected regions revealed the presence of two 369
partially conserved 11-bp motifs arranged as inverted repeats in the MdrA-binding site 370
II (Fig. 7B). In contrast, the analysis of sequences corresponding to the MdrA-binding 371
site I showed no obvious dyad symmetry. However, alignment of the 11-bp motifs 372
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

17
together with the downstream protected region revealed the presence of a similar 373
sequence located at the center of the MdrA-binding site I (Fig. 7A), suggesting that 374
MdrA specifically recognizes this sequence. In agreement with this hypothesis, MdrA 375
was unable to bind a probe lacking a sequence which is comprised within the 11-bp 376
motifs found in both MdrA-binding sites (Fig. 7C), which supports the notion that the 377
sequence TAAA is part of the motif recognized by MdrA in the promoter. 378
379
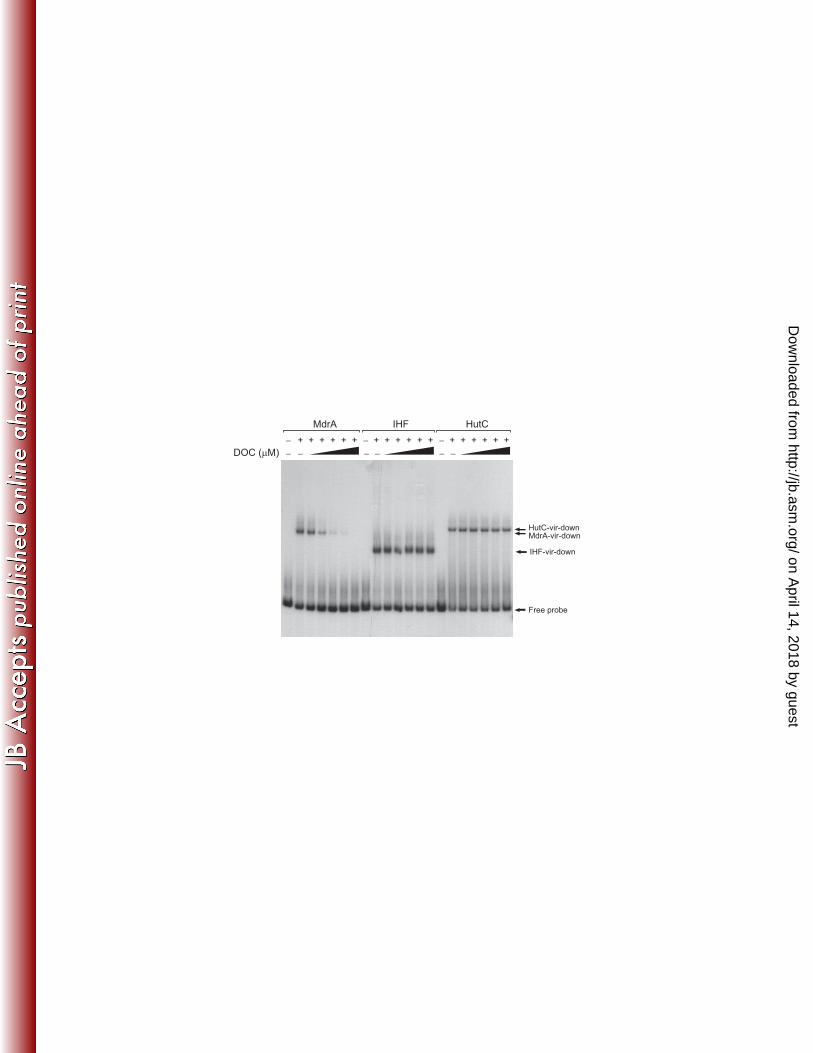
MdrA dissociates from DNA in the presence of sodium deoxycholate 380
Several members of the MarR family are capable of directly bind specific ligands, 381
which dissociate the regulator from DNA with a consequent modulation of gene 382
expression (17). In Salmonella typhimurium, it was recently found that sodium 383
deoxycholate (DOC), a component of bile, interacts with MarR and interferes with its 384
ability to bind the promoter of the marRAB operon (35). Accordingly, we investigated 385
whether the DNA-binding activity of MdrA is susceptible to be affected by total bile 386
salts or bile components. We observed by EMSA that formation of the MdrA-vir-down 387
complex was impaired in the presence of total bile salts (data not shown). When 388
individual components of bile were tested, we observed that binding of MdrA to PvirB 389
was impaired by 250 μM DOC, whereas the binding of IHF or HutC was not affected 390
by this bile salt at any of the assayed concentrations (Fig. 8). Therefore, these results 391
demonstrated that like other members of the MarR-family of transcriptional regulators, 392
the interaction between MdrA and its operator sequences can be modulated by specific 393
compounds, and such ability may be involved in transduction of environmental stimuli 394
to regulate MdrA-dependent gene expression. 395
396
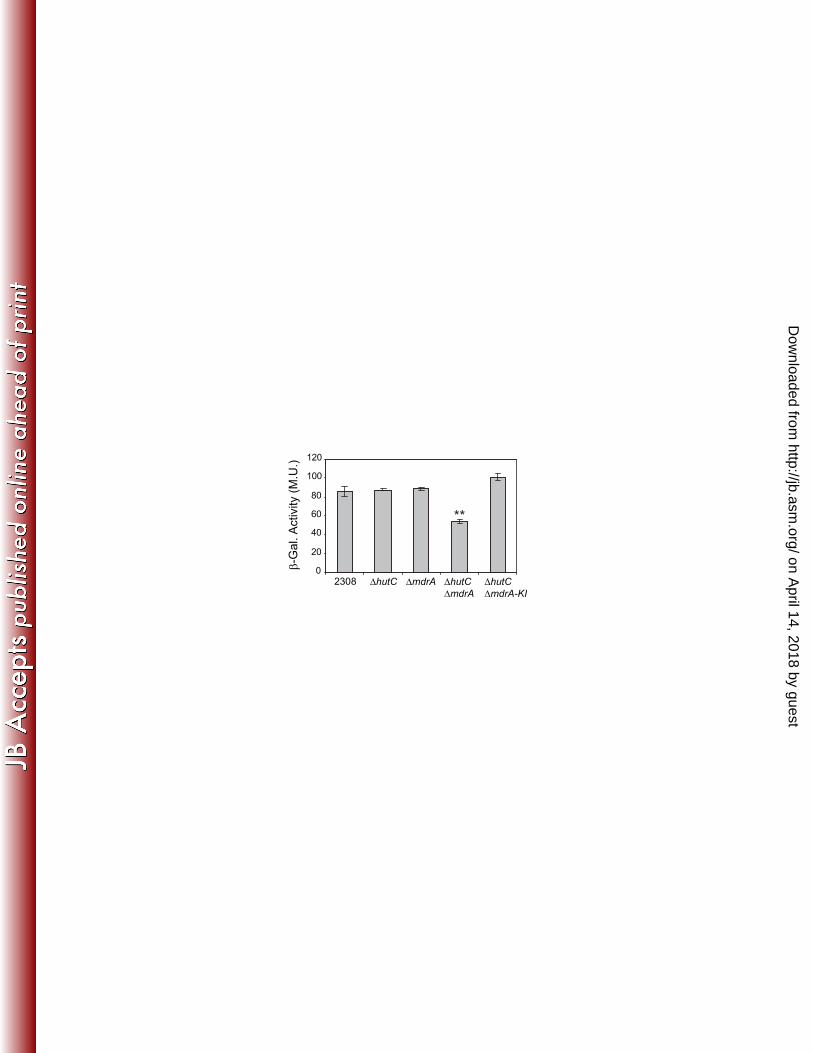
MdrA and HutC exert redundant roles on regulation of virB expression 397
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

18
In order to determine the role of MdrA on transcriptional regulation of the virB 398
genes, we introduced transcriptional fusions between PvirB and the lacZ reporter gene 399
into the B. abortus 2308 wild type strain or into a ΔmdrA deletion mutant. 400
Determination of β-Galactosidase activities showed no differences between these strains 401
after incubation of bacteria in rich medium (TSB) or in minimal media at different pH 402
values in the presence of different carbon sources (data not shown). Based on these 403
results, we hypothesized that the regulatory function of MdrA on PvirB is probably 404
redundant with that of other transcription factor. Given that both MdrA and HutC bound 405
to overlapping binding sites, and they appear to have similar structural roles since 406
neither of them induces DNA bending, we aimed to determine whether these two 407
proteins exert redundant roles on virB expression. To this end, β-Galactosidase activites 408
of the PvirB-lacZ fusions were also assayed in both B. abortus ΔhutC and the B. abortus 409
ΔhutC ΔmdrA double deletion mutant strain. Using all these genetic backgrounds, we 410
neither observed β-Galactosidase activity differences between strains when bacteria 411
were grown in TSB until stationary phase (OD600 ≥ 3.5) or in the above mentioned 412
minimal culture media (data not shown). However, in bacteria grown in TSB until 413
exponential phase (OD600 ~ 0.5-1.0), the ΔhutC ΔmdrA double deletion mutant showed 414
a decrease of about 45% in promoter activity, whereas both the ΔhutC and the ΔmdrA 415
single mutants showed the same levels of β-Galactosidase activity as the wild type 416
strain (Fig. 9). Complementation of mdrA by a knock-in strategy in the double mutant 417
restored the wild type β-Galactosidase activity levels (Fig. 9), thus confirming that 418
MdrA exerts a growth-phase dependent positive regulatory role on virB expression, 419
functionally redundant with HutC. The analyses performed in the presence of DOC 420
displayed the same pattern of β-Galactosidase activity as that observed without this bile 421
salt (data not shown), even at concentrations 3-fold higher than that dissociated MdrA 422
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

19
from the promoter. Thus, the sole presence of this bile salt was not a sufficient stimulus 423
for modulating virB expression under standard culture conditions, which suggests that 424
the capability of the MdrA-mediated regulation to respond to environmental signals 425
may be restricted to specific environmental conditions. 426
Taken together, these results revealed that MdrA controls expression of the virB 427
genes in a growth phase-dependent manner, and exerts a regulatory role functionally 428
redundant to a non-related factor. 429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
451
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

20
DISCUSSION 452
Expression of the virB operon is under the control of transcriptional regulatory 453
mechanisms that involve perception of environmental conditions related to the 454
intracellular lifestyle of Brucella (4). Over the last years, several studies revealed that 455
activity of the virB promoter is modulated by the direct action of many transcriptional 456
regulators belonging to different families, including a nucleoid associated protein (IHF), 457
QS-related transcription factors, and a regulator of histidine catabolism (6, 12, 25, 45-458
46). Here, using an approach that previously allowed us to isolate IHF and HutC, we 459
identified MdrA, a protein belonging to the MarR-family of transcription factors 460
involved in regulation of virulence genes, aromatic catabolic pathways, or bacterial 461
responses to environmental stress (53). The first evidence of interaction between MdrA 462
and PvirB indicated that this regulator binds to the promoter at sequences located far 463
upstream of the transcription start site (positions -430 to -202). Further EMSA analyses 464
showed that MdrA was also able to interact with the downstream region, indicating that 465
this regulator recognizes two distinct binding sites at PvirB (Fig. 3). These observations 466
were confirmed by DNase I Footprinting analyses that showed that MdrA is able to bind 467
simultaneously to both regions, which are centered at positions -175 and -323.5 (MarR-468
binding sites I and II, respectively). It is remarkable that all positive regulators 469
characterized so far in the MarR family act by means of displacing repressors (19, 40) 470
or through prototypical activation mechanisms by interacting with RNAP (28, 50). This 471
latter mode of regulation may not account for activity of MdrA on the virB promoter, 472
since it activates transcription from regions located far upstream of position +1. We 473
hypothesize that MdrA could interfere with the activity of an unknown repressor; a 474
possible candidate may be BabR, which is a negative regulator that affects virB 475
expression by 20% and whose binding site in the promoter has not yet been identified 476
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

21
(6). Alternatively, contact between MdrA and RNAP may be mediated by additional 477
elements which could introduce conformational changes in chromatin structure. Similar 478
mechanisms were previously suggested for other positive regulators that bind far 479
upstream from the transcription start site but whose mechanisms of activation have not 480
been yet elucidated (e.g., EspR, RutR, TodT, AlgR) (20, 26, 29, 39). It is worth 481
mentioning that the possible involvement of elements that introduce modifications of 482
the nucleoprotein structure of local sequences was also hypothesized for the LuxR-type 483
regulator VjbR, since it binds at position -94 and consequently could not activate 484
transcription of the virB promoter in a prototypical manner (2). 485
Our experiments revealed that the MdrA-binding site I overlaps those previously 486
identified for IHF and HutC, which was consistent with the fact that these three 487
transcription factors compete for binding to the promoter (Fig. 4). Similar structures 488
have been reported for other promoters regulated by members of the MarR-family (i.e, 489
CbaR, MarR, RovA), wherein the regulators bind to two or more operator sites (19, 24, 490
36). Furthermore, competition with nucleoid associated proteins (i.e, H-NS, IHF) or 491
with other global regulators (CRP) due to overlapping binding sites has also been 492
reported to be part of the regulatory mechanisms involving MarR-related regulators (19, 493
40, 43). An additional feature shared by MdrA and most of the MarR homologs is the 494
ligand-mediated modulation of its DNA-binding activity. However, it is important to 495
highlight that MdrA is the first ligand-responsive positive regulator characterized in this 496
family, since no ligand has yet been found for the other MarR-type transcriptional 497
activators described so far (14). Curiously, MdrA is the second positive regulator of the 498
virB promoter which dissociates from DNA in response to specific signals, since VjbR 499
was previously shown to respond to acyl-homoserine lactones (2, 12). Our results 500
indicated that either bile or micromolar concentrations of DOC specifically dissociated 501
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

22
MdrA from DNA without affecting neither IHF nor HutC (Fig. 8). These observations 502
have raised the hypothesis that after ligand recognition, MdrA could act as a signal 503
transducer that modulates expression of the virB genes in response to exposure of 504
Brucella to DOC within the host environment. Previous studies have described in 505
Brucella mechanisms that confer resistance to DOC (11, 23, 33). Moreover, it was 506
recently shown that Brucella is able to infect dendritic cells from murine Peyer´s 507
patches and to negatively modulate its activation through the action of the protein Btp1 508
(41). Although these lines of evidence support the notion that Brucella is able to 509
overcome the action of bile salts and to invade antigen-presenting cells in the host’s gut, 510
we failed to observe any effect of DOC on virB expression in exponential phase cultures 511
under standard growth conditions. However, it cannot be excluded that, under some 512
type of stress experienced by the bacterium within the host, this bile salt may permeate 513
the bacterial membrane more easily and act as an environmental signal to induce an 514
MdrA-mediated response. It is worth mentioning that it cannot be deduced any additional 515
potential ligand for MdrA from the few MarR-homologs reported so far in Rhizobiales, 516
since their known ligands do not share structural similarity with DOC (5, 31). Therefore, 517
further studies will be needed to determine whether this bile salt, or a structurally 518
similar compound, can interact with MdrA in vivo and modulate virB expression. 519
The results presented here showed that MdrA and HutC, two independently 520
isolated proteins belonging to different families of transcription factors, bind to PvirB at 521
overlapping target DNA-sequences and positively modulate expression of the virB 522
genes to similar extents. Moreover, both regulators have been shown to play redundant 523
roles since the regulatory activity of MdrA could not be determined unless a ΔmdrA 524
ΔhutC double deletion mutant background was used to measure virB promoter activity 525
(Fig. 9). In addition to these evidences, our EMSA experiments indicated that, unlike 526
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

23
IHF, neither MdrA nor HutC induced DNA-bending upon binding to PvirB (Fig. 4), 527
suggesting that these two transcriptional regulators exert similar structural roles in 528
agreement with their redundant activity on modulation of virB expression. 529
The data obtained from the present work showed that the function of MdrA leads to a 530
positive regulatory effect on virB expression which does not take place in an all-or-531
nothing manner. Deletion of mdrA affected virB expression by 45 %, which was a 532
magnitude of difference detectable by quantitative measurements of β-gal activity 533
(Figure 9) but not by Western blot experiments performed with an anti-VirB7 antibody 534
(data not shown). Our results indicated that, similarly to what was previously observed 535
with HutC, MdrA also acts as an activator which enhances virB promoter activity under 536
defined conditions and appears to play an accessory regulatory role, probably acting to 537
synchronize maximal virB expression with certain metabolic and/or environmental 538
signals. It was previously observed that HutC activates intracellular expression of the 539
virB genes within J774 macrophages (45). In contrast, MdrA does not appear to play 540
any role in this experimental model, since deletion of mdrA did not produce any 541
detectable intracellular effect, and the ΔmdrA ΔhutC double mutant affected 542
intracellular virB expression to the same extent as in the single ΔhutC deletion 543
background (Supplemental Fig. S1). On the other hand, MdrA was necessary for virB 544
expression in cultured bacteria at the exponential phase of the growth in rich medium, 545
whereas in stationary phase we did not observe any MdrA-dependent effect (Fig. 9). In 546
addition, it is worth to note that the growth phase wherein MdrA exerts its regulatory 547
activity is coincident with the only condition at which it could be isolated (Fig 1), 548
indicating that this regulator is probably under the control of mechanisms that modulate 549
its expression and/or its DNA-binding activity. Taken together, our observations 550
suggest that the MdrA-mediated regulation of the virB genes may be a result of an 551
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

24
adaptation to activate virB expression at stages of the infection process where HutC is 552
not acting. However, although functionality of MdrA was demonstrated, it still remains 553
to be determined which are the conditions wherein MdrA-dependent expression is 554
achieved in vivo. Besides, it will also be important to assess whether this MarR-related 555
transcriptional regulator is able to perceive environmental signals at specific 556
tissues/organs of the mammalian host during the course of the disease. 557
558
559
560
561
562
563
564
565
566
567
568
569
570
571
572
573
574
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

25
ACKNOWLEDGEMENTS 575
This work is dedicated to the memory of R.A.U. We thank Joseph Connolly for help 576
with mass spectrometry. This work was supported by grant PICT05-38207 to Rodrigo 577
Sieira, and grant PICT06-651 to Rodolfo A. Ugalde from Agencia Nacional de 578
Promoción Científica y Tecnológica, Buenos Aires, Argentina; grant PIP2011-00336 to 579
Rodrigo Sieira from the Argentinian Council of Research (CONICET); and grant 580
UBACyT X240 to Angeles Zorreguieta from the University of Buenos Aires, 581
Argentina. 582
583
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

26
REFERENCES 584
1. Alekshun, M. N., S. B. Levy, T. R. Mealy, B. A. Seaton, and J. F. Head. 585 2001. The crystal structure of MarR, a regulator of multiple antibiotic resistance, 586 at 2.3 A resolution. Nat Struct Biol 8:710-4. 587
2. Arocena, G. M., R. Sieira, D. J. Comerci, and R. A. Ugalde. 2010. 588 Identification of the Quorum Sensing target DNA-sequence and N-acyl 589 Homoserine Lactone responsiveness in the Brucella abortus virB promoter. J 590 Bacteriol. 591
3. Backert, S., and T. F. Meyer. 2006. Type IV secretion systems and their 592 effectors in bacterial pathogenesis. Curr Opin Microbiol 9:207-17. 593
4. Boschiroli, M. L., S. Ouahrani-Bettache, V. Foulongne, S. Michaux-594 Charachon, G. Bourg, A. Allardet-Servent, C. Cazevieille, J. P. Liautard, 595 M. Ramuz, and D. O'Callaghan. 2002. The Brucella suis virB operon is 596 induced intracellularly in macrophages. Proc Natl Acad Sci U S A 99:1544-9. 597
5. Caswell, C. C., J. E. Baumgartner, D. W. Martin, and R. M. Roop, 2nd. 598 2012. Characterization of the organic hydroperoxide resistance system of 599 Brucella abortus 2308. J Bacteriol. 600
6. Caswell, C. C., J. M. Gaines, and R. M. Roop, 2nd. 2012. The RNA 601 chaperone Hfq independently coordinates expression of the VirB type IV 602 secretion system and the LuxR-type regulator BabR in Brucella abortus 2308. J 603 Bacteriol 194:3-14. 604
7. Celli, J., C. de Chastellier, D. M. Franchini, J. Pizarro-Cerda, E. Moreno, 605 and J. P. Gorvel. 2003. Brucella evades macrophage killing via VirB-606 dependent sustained interactions with the endoplasmic reticulum. J Exp Med 607 198:545-56. 608
8. Covacci, A., J. L. Telford, G. Del Giudice, J. Parsonnet, and R. Rappuoli. 609 1999. Helicobacter pylori virulence and genetic geography. Science 284:1328-610 33. 611
9. de Barsy, M., A. Jamet, D. Filopon, C. Nicolas, G. Laloux, J. F. Rual, A. 612 Muller, J. C. Twizere, B. Nkengfac, J. Vandenhaute, D. E. Hill, S. P. 613 Salcedo, J. P. Gorvel, J. J. Letesson, and X. De Bolle. 2011. Identification of a 614 Brucella spp. secreted effector specifically interacting with human small GTPase 615 Rab2. Cell Microbiol 13:1044-58. 616
10. de Jong, M. F., Y. H. Sun, A. B. den Hartigh, J. M. van Dijl, and R. M. 617 Tsolis. 2008. Identification of VceA and VceC, two members of the VjbR 618 regulon that are translocated into macrophages by the Brucella type IV secretion 619 system. Mol Microbiol 70:1378-96. 620
11. Delpino, M. V., M. I. Marchesini, S. M. Estein, D. J. Comerci, J. Cassataro, 621 C. A. Fossati, and P. C. Baldi. 2007. A bile salt hydrolase of Brucella abortus 622 contributes to the establishment of a successful infection through the oral route 623 in mice. Infect Immun 75:299-305. 624
12. Delrue, R. M., C. Deschamps, S. Leonard, C. Nijskens, I. Danese, J. M. 625 Schaus, S. Bonnot, J. Ferooz, A. Tibor, X. De Bolle, and J. J. Letesson. 2005. 626 A quorum-sensing regulator controls expression of both the type IV secretion 627 system and the flagellar apparatus of Brucella melitensis. Cell Microbiol 628 7:1151-61. 629
13. Dozot, M., R. A. Boigegrain, R. M. Delrue, R. Hallez, S. Ouahrani-Bettache, 630 I. Danese, J. J. Letesson, X. De Bolle, and S. Kohler. 2006. The stringent 631 response mediator Rsh is required for Brucella melitensis and Brucella suis 632
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

27
virulence, and for expression of the type IV secretion system virB. Cell 633 Microbiol 8:1791-802. 634
14. Ellison, D. W., and V. L. Miller. 2006. Regulation of virulence by members of 635 the MarR/SlyA family. Curr Opin Microbiol 9:153-9. 636
15. Gerhardt, P. 1958. The nutrition of brucellae. Bacteriol Rev 22:81-98. 637 16. Godfroid, J., H. C. Scholz, T. Barbier, C. Nicolas, P. Wattiau, D. Fretin, A. 638
M. Whatmore, A. Cloeckaert, J. M. Blasco, I. Moriyon, C. Saegerman, J. B. 639 Muma, S. Al Dahouk, H. Neubauer, and J. J. Letesson. 2011. Brucellosis at 640 the animal/ecosystem/human interface at the beginning of the 21st century. Prev 641 Vet Med 102:118-31. 642
17. Grkovic, S., M. H. Brown, and R. A. Skurray. 2002. Regulation of bacterial 643 drug export systems. Microbiol Mol Biol Rev 66:671-701, table of contents. 644
18. Haine, V., A. Sinon, F. Van Steen, S. Rousseau, M. Dozot, P. Lestrate, C. 645 Lambert, J. J. Letesson, and X. De Bolle. 2005. Systematic targeted 646 mutagenesis of Brucella melitensis 16M reveals a major role for GntR regulators 647 in the control of virulence. Infect Immun 73:5578-86. 648
19. Heroven, A. K., G. Nagel, H. J. Tran, S. Parr, and P. Dersch. 2004. RovA is 649 autoregulated and antagonizes H-NS-mediated silencing of invasin and rovA 650 expression in Yersinia pseudotuberculosis. Mol Microbiol 53:871-88. 651
20. Lacal, J., A. Busch, M. E. Guazzaroni, T. Krell, and J. L. Ramos. 2006. The 652 TodS-TodT two-component regulatory system recognizes a wide range of 653 effectors and works with DNA-bending proteins. Proc Natl Acad Sci U S A 654 103:8191-6. 655
21. Ludwig, A., C. Tengel, S. Bauer, A. Bubert, R. Benz, H. J. Mollenkopf, and 656 W. Goebel. 1995. SlyA, a regulatory protein from Salmonella typhimurium, 657 induces a haemolytic and pore-forming protein in Escherichia coli. Mol Gen 658 Genet 249:474-86. 659
22. Marchesini, M. I., C. K. Herrmann, S. P. Salcedo, J. P. Gorvel, and D. J. 660 Comerci. 2011. In search of Brucella abortus type IV secretion substrates: 661 screening and identification of four proteins translocated into host cells through 662 VirB system. Cell Microbiol 13:1261-74. 663
23. Martin, F. A., D. M. Posadas, M. C. Carrica, S. L. Cravero, D. 664 O'Callaghan, and A. Zorreguieta. 2009. Interplay between two RND systems 665 mediating antimicrobial resistance in Brucella suis. J Bacteriol 191:2530-40. 666
24. Martin, R. G., and J. L. Rosner. 1995. Binding of purified multiple antibiotic-667 resistance repressor protein (MarR) to mar operator sequences. Proc Natl Acad 668 Sci U S A 92:5456-60. 669
25. Martinez-Nunez, C., P. Altamirano-Silva, F. Alvarado-Guillen, E. Moreno, 670 C. Guzman-Verri, and E. Chaves-Olarte. 2010. The two-component system 671 BvrR/BvrS regulates the expression of the type IV secretion system VirB in 672 Brucella abortus. J Bacteriol 192:5603-8. 673
26. Mohr, C. D., N. S. Hibler, and V. Deretic. 1991. AlgR, a response regulator 674 controlling mucoidy in Pseudomonas aeruginosa, binds to the FUS sites of the 675 algD promoter located unusually far upstream from the mRNA start site. J 676 Bacteriol 173:5136-43. 677
27. Nagel, G., A. Lahrz, and P. Dersch. 2001. Environmental control of invasin 678 expression in Yersinia pseudotuberculosis is mediated by regulation of RovA, a 679 transcriptional activator of the SlyA/Hor family. Mol Microbiol 41:1249-69. 680
28. Navarre, W. W., T. A. Halsey, D. Walthers, J. Frye, M. McClelland, J. L. 681 Potter, L. J. Kenney, J. S. Gunn, F. C. Fang, and S. J. Libby. 2005. Co-682
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

28
regulation of Salmonella enterica genes required for virulence and resistance to 683 antimicrobial peptides by SlyA and PhoP/PhoQ. Mol Microbiol 56:492-508. 684
29. Nguyen Ple, M., I. Bervoets, D. Maes, and D. Charlier. 2010. The protein-685 DNA contacts in RutR*carAB operator complexes. Nucleic Acids Res 38:6286-686 300. 687
30. O'Callaghan, D., C. Cazevieille, A. Allardet-Servent, M. L. Boschiroli, G. 688 Bourg, V. Foulongne, P. Frutos, Y. Kulakov, and M. Ramuz. 1999. A 689 homologue of the Agrobacterium tumefaciens VirB and Bordetella pertussis Ptl 690 type IV secretion systems is essential for intracellular survival of Brucella suis. 691 Mol Microbiol 33:1210-20. 692
31. Perera, I. C., and A. Grove. 2010. Urate is a ligand for the transcriptional 693 regulator PecS. J Mol Biol 402:539-51. 694
32. Pizarro-Cerda, J., S. Meresse, R. G. Parton, G. van der Goot, A. Sola-695 Landa, I. Lopez-Goni, E. Moreno, and J. P. Gorvel. 1998. Brucella abortus 696 transits through the autophagic pathway and replicates in the endoplasmic 697 reticulum of nonprofessional phagocytes. Infect Immun 66:5711-24. 698
33. Posadas, D. M., F. A. Martin, J. V. Sabio y Garcia, J. M. Spera, M. V. 699 Delpino, P. Baldi, E. Campos, S. L. Cravero, and A. Zorreguieta. 2007. The 700 TolC homologue of Brucella suis is involved in resistance to antimicrobial 701 compounds and virulence. Infect Immun 75:379-89. 702
34. Praillet, T., W. Nasser, J. Robert-Baudouy, and S. Reverchon. 1996. 703 Purification and functional characterization of PecS, a regulator of virulence-704 factor synthesis in Erwinia chrysanthemi. Mol Microbiol 20:391-402. 705
35. Prouty, A. M., I. E. Brodsky, J. Manos, R. Belas, S. Falkow, and J. S. Gunn. 706 2004. Transcriptional regulation of Salmonella enterica serovar Typhimurium 707 genes by bile. FEMS Immunol Med Microbiol 41:177-85. 708
36. Providenti, M. A., and R. C. Wyndham. 2001. Identification and functional 709 characterization of CbaR, a MarR-like modulator of the cbaABC-encoded 710 chlorobenzoate catabolism pathway. Appl Environ Microbiol 67:3530-41. 711
37. Rambow-Larsen, A. A., G. Rajashekara, E. Petersen, and G. Splitter. 2008. 712 Putative quorum-sensing regulator BlxR of Brucella melitensis regulates 713 virulence factors including the type IV secretion system and flagella. J Bacteriol 714 190:3274-82. 715
38. Robertson, C. A., and H. A. Nash. 1988. Bending of the bacteriophage lambda 716 attachment site by Escherichia coli integration host factor. J Biol Chem 717 263:3554-7. 718
39. Rosenberg, O. S., C. Dovey, M. Tempesta, R. A. Robbins, J. S. Finer-719 Moore, R. M. Stroud, and J. S. Cox. 2011. EspR, a key regulator of 720 Mycobacterium tuberculosis virulence, adopts a unique dimeric structure among 721 helix-turn-helix proteins. Proc Natl Acad Sci U S A 108:13450-5. 722
40. Rouanet, C., K. Nomura, S. Tsuyumu, and W. Nasser. 1999. Regulation of 723 pelD and pelE, encoding major alkaline pectate lyases in Erwinia chrysanthemi: 724 involvement of the main transcriptional factors. J Bacteriol 181:5948-57. 725
41. Salcedo, S. P., M. I. Marchesini, H. Lelouard, E. Fugier, G. Jolly, S. Balor, 726 A. Muller, N. Lapaque, O. Demaria, L. Alexopoulou, D. J. Comerci, R. A. 727 Ugalde, P. Pierre, and J. P. Gorvel. 2008. Brucella control of dendritic cell 728 maturation is dependent on the TIR-containing protein Btp1. PLoS Pathog 729 4:e21. 730
42. Schafer, A., A. Tauch, W. Jager, J. Kalinowski, G. Thierbach, and A. 731 Puhler. 1994. Small mobilizable multi-purpose cloning vectors derived from the 732
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

29
Escherichia coli plasmids pK18 and pK19: selection of defined deletions in the 733 chromosome of Corynebacterium glutamicum. Gene 145:69-73. 734
43. Schielke, S., C. Huebner, C. Spatz, V. Nagele, N. Ackermann, M. Frosch, O. 735 Kurzai, and A. Schubert-Unkmeir. 2009. Expression of the meningococcal 736 adhesin NadA is controlled by a transcriptional regulator of the MarR family. 737 Mol Microbiol 72:1054-67. 738
44. Segal, G., M. Purcell, and H. A. Shuman. 1998. Host cell killing and bacterial 739 conjugation require overlapping sets of genes within a 22-kb region of the 740 Legionella pneumophila genome. Proc Natl Acad Sci U S A 95:1669-74. 741
45. Sieira, R., G. M. Arocena, L. Bukata, D. J. Comerci, and R. A. Ugalde. 742 2010. Metabolic control of virulence genes in Brucella abortus: HutC 743 coordinates virB expression and the histidine utilization pathway by direct 744 binding to both promoters. J Bacteriol 192:217-24. 745
46. Sieira, R., D. J. Comerci, L. I. Pietrasanta, and R. A. Ugalde. 2004. 746 Integration host factor is involved in transcriptional regulation of the Brucella 747 abortus virB operon. Mol Microbiol 54:808-22. 748
47. Sieira, R., D. J. Comerci, D. O. Sanchez, and R. A. Ugalde. 2000. A 749 homologue of an operon required for DNA transfer in Agrobacterium is required 750 in Brucella abortus for virulence and intracellular multiplication. J Bacteriol 751 182:4849-55. 752
48. Stachel, S. E., and E. W. Nester. 1986. The genetic and transcriptional 753 organization of the vir region of the A6 Ti plasmid of Agrobacterium 754 tumefaciens. EMBO J 5:1445-54. 755
49. Thompson, J. D., D. G. Higgins, and T. J. Gibson. 1994. CLUSTAL W: 756 improving the sensitivity of progressive multiple sequence alignment through 757 sequence weighting, position-specific gap penalties and weight matrix choice. 758 Nucleic Acids Res 22:4673-80. 759
50. Tran, H. J., A. K. Heroven, L. Winkler, T. Spreter, B. Beatrix, and P. 760 Dersch. 2005. Analysis of RovA, a transcriptional regulator of Yersinia 761 pseudotuberculosis virulence that acts through antirepression and direct 762 transcriptional activation. J Biol Chem 280:42423-32. 763
51. Ugalde, J. E., C. Czibener, M. F. Feldman, and R. A. Ugalde. 2000. 764 Identification and characterization of the Brucella abortus phosphoglucomutase 765 gene: role of lipopolysaccharide in virulence and intracellular multiplication. 766 Infect Immun 68:5716-23. 767
52. Whatmore, A. M. 2009. Current understanding of the genetic diversity of 768 Brucella, an expanding genus of zoonotic pathogens. Infect Genet Evol 9:1168-769 84. 770
53. Wilkinson, S. P., and A. Grove. 2006. Ligand-responsive transcriptional 771 regulation by members of the MarR family of winged helix proteins. Curr Issues 772 Mol Biol 8:51-62. 773
54. Winans, S. C., and G. C. Walker. 1985. Conjugal transfer system of the IncN 774 plasmid pKM101. J Bacteriol 161:402-10. 775
776 777
778
779
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

30
LEGENDS TO FIGURES 780
Figure 1: Identification of MdrA. EMSA performed with probe vir-up and total crude 781
protein extracts from exponential (E) or stationary-phase (S) cultures of B. abortus. 782
Arrows indicate position of the free probe or a protein-DNA complex. 783
784
Figure 2: Sequence alignment of MdrA and different representative members of the 785
MarR-family of transcriptional regulators. Relative position of secondary structure 786
elements is shown as a schematic representation of α-helices, β-sheets and the wing 787
region (W1), according to the Escherichia coli MarR crystal structure described by 788
Alekshun et al. (2001) (1). White letters identify residues identical in all (highlighted in 789
black), or in six out of the seven MarR homologs considered (highlighted in dark gray). 790
Black letters identify residues identical in five or four (highlighted in gray), or in three 791
out of the seven MarR homologs (highlighted in light gray). The sequence alignment 792
was performed using ClustalW (49). 793
794
Figure 3: Analysis of the interaction between MdrA and the virB promoter. A. 795
Schematic representation of genomic sequences corresponding to the regulatory region 796
of the virB operon, radiolabeled probes used for EMSA, or unlabeled competitors. 797
Positions relative to the transcription start site are indicated. MdrA, HutC, IHF and 798
VjbR-binding sites are indicated as boxes. Crossed box indicates the non-related 799
sequences that replace the IHF-binding site in probe vir-down-ihf-. B. EMSA performed 800
with the indicated probes and 0, 10, 20, 40 or 60 nM MdrA. C. EMSA performed with 801
probe vir-up, MdrA, and the indicated unlabeled DNA fragments as competitors. 802
Protein concentration was as follows: line 1, no protein; lines 2-8, 30 nM. Concentration 803
of unlabelled DNA competitors was as follows: lanes 1 and 2, no competitor DNA; 804
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

31
lanes 3, 5, and 7, 150 ng; lanes 4, 6 and 8, 360 ng. D. EMSA performed with probe vir-805
down, MdrA, and the indicated unlabeled DNA fragments as competitors. 806
Concentration of protein and unlabelled DNA competitors was as in (C). 807
808
Figure 4: Analysis of the interaction between MdrA, HutC and/or IHF with the 809
downstream region of the virB promoter. A. Schematic representation of probes vir-810
down and 286/59. Rectangles indicate position of the binding sites of HutC, IHF, and 811
MdrA. B. Co-incubation experiment performed by EMSA with probe vir-down and 812
different combinations of 20 nM MdrA, 20 nM HutC or 40 nM IHF. C. Co-incubation 813
experiment performed by EMSA with probe 286/59 and different combinations MdrA, 814
HutC or IHF. Protein concentration was as in (B). 815
816
Figure 5: Determination of the MdrA-binding sites in the virB promoter. A. Schematic 817
representation of probes vir-up and vir-downI used for DNase I Footprinting 818
experiments. Positions relative to the transcription start site of PvirB are indicated. MdrA, 819
HutC, IHF and VjbR-binding sites are indicated as boxes. Arrows and asterisks indicate 820
the primer used for construction of each 5’ radiolabeled probe. B. DNase I Footprinting 821
experiment performed with probe vir-downI and MdrA at the indicated concentrations. 822
Lanes A and G show sequence reactions performed by the Sanger method with primer 823
vir-downI. The protected region of the MdrA-binding site I is indicated with an open 824
rectangle. C. DNase I Footprinting experiment performed with probe vir-up and MdrA 825
at the indicated concentrations. Lanes A and G show sequence reactions performed by 826
the Sanger method with primer vir-up. The protected region of the MdrA-binding site II 827
is indicated as in (B). 828
829
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

32
Figure 6: MdrA binds simultaneously to binding sites I and II. A. Schematic 830
representation of probe vir-upIII used for DNase I Footprinting experiments shown in 831
(B) and (C). Positions relative to the transcription start site of PvirB are indicated. 832
Rectangles indicate MdrA, HutC, IHF and VjbR-binding sites. Arrow and asterisk 833
indicate the primer used for construction of the 5’ radiolabeled probe vir-upIII. B and C 834
are different runs of a DNase I Footprinting experiment performed with probe vir-upIII 835
and MdrA at the indicated concentrations. Lanes A and G show sequence reactions 836
performed by the Sanger method with primer pvirup. The protected region of the MdrA-837
binding sites I and II are indicated with open rectangles. 838
839
Figure 7: Sequence analysis of the MdrA-protected regions in the virB promoter. A. 840
Schematic representation of the MdrA-protected sequence in the binding site I. The 841
rectangle indicates regions protected from DNase I cleavage in experiments performed 842
with probe vir-upIII and vir-downII. Positions relative to the transcription start site are 843
indicated. Sequences corresponding to the HutC and IHF-binding sites are underlined. 844
Arrow indicates position of the 11-bp MdrA-binding consensus motif. Nucleotides that 845
match the 11-bp MdrA-binding consensus motif are highlighted in grey. B. Schematic 846
representation of the MdrA-protected sequence in the binding site II. The open rectangle 847
indicates the region protected from DNase I cleavage. Positions relative to the 848
transcription start site are indicated. Arrows indicate position of the 11-bp MdrA-849
binding consensus motifs. Nucleotides that match the 11-bp MdrA-binding consensus 850
motifs are highlighted in grey. Arrows indicate dyad symmetry. C. Schematic 851
representation of sequences corresponding to probe vir-down-ihf-. The HutC-binding 852
site is indicated as in (A). Nucleotides that match the 11-bp MdrA-binding consensus 853
motif are highlighted in grey. Nucleotides that replaced the IHF-binding site by a non-854
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

33
related sequence are indicated in italic. Position of nucleotides of the 11-bp MdrA-855
binding consensus motif that are not present in probe vir-down-ihf- is indicated by an 856
open rectangle. D. Alignment of sequences corresponding to the 11-bp MdrA-binding 857
consensus motif found at the protected regions of the MdrA-binding sites I and II. 858
859
Figure 8: Effect of DOC on the binding activity of MdrA. EMSA performed with probe 860
vir-down and 30 nM MdrA, 50 nM IHF or 30 nM HutC, in the presence of 100, 250, 861
500, 750, or 1000 μM DOC. 862
863
Figure 9: Role of MdrA in regulation of the virB promoter activity. Strains B. abortus 864
PvirB-lacZ (wt), B. abortus ΔhutC PvirB-lacZ, B. abortus ΔmdrA PvirB-lacZ, B. abortus 865
ΔhutC ΔmdrA PvirB-lacZ, or B. abortus ΔhutC ΔmdrA-KI PvirB-lacZ were grown in rich 866
medium (TSB) until exponential phase (OD600 0.5-1). Subsequently, bacteria were 867
harvested, and β-Galactosidase activites were determined. Values are means ± standard 868
deviations of duplicate wells from a representative of three experiments. **, P < 0.01. 869
870
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

_ E SExtract
Freeprobe
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

α1 α2 β1 α3
α4 β2 β3w1
α5
α6
Escherichia coli MarR
Salmonella typhimurium SlyA
Yersinia enterocolitica RovA
Erwinia chrysanthemi PecS
Pseudomonas aeruginosa MexR
Brucella abortus MdrA
Sinorhizobium meliloti ExpG
MarR
SlyA
RovA
PecS
MexR
MdrA
ExpG
MarR
SlyA
RovA
PecS
MexR
MdrA
ExpG
MarR
SlyA
RovA
PecS
MexR
MdrA
ExpG
---------------------------------------MKSTSDLFNEIIPLGRLIHMV 21
---------------------------------------------MKLESPLGS-DLARL 14
-----------------------------------------------MESTLGS-DLARL 12
-------------------------MARYLEVSDIVQQWRNERPDLDVEPMLVIGTLSRV 35
-------------------------------------------MNYPVNPDLMPALMAVF 17
MRNKPPHKTGRELHRGFTKCGSGREAKEMTNTQRKMDASAPFMAPQTALRSLYLEALQLV 60
----MERGMNHRILYPFADFGDTVAILPANETQRKGLDTPVDDRDGDDSLVTYFELARVM 56
NQKKDRLLNEY------LSPLDITAAQFKVLCSIRC---AACITPVELKKVLSVDLGALT 72
VRIWRALIDHR------LKPLELTQTHWVTLHNIHQ--LPPDQSQIQLAKAIGIEQPSLV 66
VRVWRALIDHR------LKPLELTQTHWVTLHNINR--LPPEQSQIQLAKAIGIEQPSLV 64
SLLIDRALDKV------FSKYKLSAREFDILATLRRRGAPYAISPSQIVNALMINNSTLT 89
QHVRTRIQSELD-----CQRLDLTPPDVHVLKLIDE---QRGLNLQDLGRQMCRDKALIT 69
ERLHRRLLDVVKDEFDRNGRSDINATQALLLFNIGN----SELTAGELRSRGYYLGSNVS 116
ERASRRFSGLLRAELTKLGVEDIGPAQAMVLLAIGE----AELSVGELLDRGHYVGSNIS 112
RMLDRLVCKGWVERLPNPNDKRGVLVKLTTSGAAICEQCHQLVGQDLHQELTKN-LTADE 131
RTLDQLEDKGLISRQTCASDRRAKRIKLTEKAEPLIAEMEEVIHKTRGEILAG--ISSEE 124
RTLDQLEEKGLITRHTCANDRRAKRIKLTEQSSPIIEQVDGVICSTRKEILGG--ISPDE 122
SRLDRLEQAGWLRRMPIEGDRRSVNIQLTDEGFALINRVVEEHVENERDILSP--FSEEE 147
RKIRELEGRNLVRRERNPSDQRSFQLFLTDEGLAIHQHAEAIMSRVHDELFAP--LTPEE 127
YNLKKLVEMGFIHHQRSRVDRRSVRVSLTDKGNEIANQVASLYERHITSIEQVGGIQVEE 176
YYLKQLADGDYIDRIASQRDKRSARIRLSEKGRQLCAGLRQAAKGYERALSHGD-QDRRN 171
VATLEHLLKKVLP---------- 144
IELLIKLIAKLEHNIMELHSHD- 146
IELLSGLIDKLERNIIQLQSK-- 143
KTQLRALLGRVEKHLVNNR---- 166
QATLVHLLDQCLAAQPLEDI--- 147
FMAMNKSLQRLDRFWNDSIAYRL 199
LETAFQTLHRLELVWGNAARYGI 194
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
B10 vir-upProbe
MdrA
vir-downvir-
down ihf
Free probe
MdrA-up
MdrA-down
C D vir-up
Competitor
MdrA
__
_ + +++++
Probe
up 77144
B
+1 8764 532
Free probe
MdrA-up
Free probe
MdrA-down
vir-down
Competitor
MdrA
__
_ + +++++
Probe
down B1A1
+1 8764 532
____
-430
-353
-286 -202
-201
up
77
144
vir-up
-94
-130
+24
down
A1
-59
B1
virB1
vir-down ihf-
vir-down Labelledprobes
Unlabelledcompetitors
+1VjbR
IHFHutC
MdrAsite I
-323
MdrAsite II
A
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
_
_
+
+_
_ _
+_ _
++
+_
_ +
+
+
+
+
+
_
_
_
Free probe
IHF-vir-down
B C
vir-downProbe
A
MdrA-vir-down
HutC-vir-down
MdrA-286/59HutC-286/59
Free probe
IHF-286/59
HutC
MdrA
IHF
286/59
_
_
+
_
+_
+_
__
_
_
_
Probe
HutC
MdrA
IHF
vir-down
286/59I
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
A G _ _40 60
MdrA (nM)
2010AG _ _10 20 1206040
MdrA (nM)B
+1III
A
*
*-335
-312
-190
-160
C
I
II
-430 -202
-286 +24
vir-upII
vir-downII
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
+1III
A
pvir-upIII
A G _ _20 50A G _ _20 50
CB
-190
-160
-335
-312
MdrA (nM)MdrA (nM)
II
I
*
-430 -60
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

agcaagttcggtaaaaatttagccagcttgga
tcgttcaagccatttttaaatcggtcgaacct
-335 -312
cgcataccacttgtatataagattttgttaaaaaagaattttctaatagagcgtatggtgaacatatattctaaaacaattttttcttaaaagattatct
IHFHutC
cgcataccacttgtatataagattttgtgccaggtgccaactgacagggagcgtatggtgaacatatattctaaaacacggtccacggttgagctgtcct
-190 -160
MdrA-binding site I
MdrA-binding site II
attttgttaaa
agttcggtaaa
agctggctaaa
MdrA-binding site I
MdrA-binding site II
B
C D
A
HutC
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
DOC (µM)
MdrA
___ ++_ __ __ _
IHF HutC
MdrA-vir-downHutC-vir-down
Free probe
IHF-vir-down
++ ++ ++++ ++ ++++ ++
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
0
20
40
60
80
100
120
2308 ∆hutC ∆mdrA ∆hutC
∆mdrA
∆hutC
∆mdrA-KI
β-G
al. A
ctivity (
M.U
.)
**
on April 14, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
![Ethylene Response Factor ERF11 Activates BT4 Transcription to … · Ethylene Response Factor ERF11 ActivatesBT4 Transcription to Regulate Immunity to Pseudomonas syringae1[OPEN]](https://img.pdfslide.net/doc/110x75/60d5b6fb3b5e1745a96489d9/ethylene-response-factor-erf11-activates-bt4-transcription-to-ethylene-response.jpg)








![The TCP4 Transcription Factor Directly Activates ... · The TCP4 Transcription Factor Directly Activates TRICHOMELESS1 and 2 and Suppresses Trichome Initiation1[OPEN] Batthula Vijaya](https://img.pdfslide.net/doc/110x75/5fb0eee496a7d621cf56e262/the-tcp4-transcription-factor-directly-activates-the-tcp4-transcription-factor.jpg)