Embed Size (px)
Citation preview

A New Genus of Gerbillid Rodent from the Early Pleistocene of the Middle EastAuthor(s): Eitan Tchernov and Rosine ChetbounSource: Journal of Vertebrate Paleontology, Vol. 4, No. 4 (Dec., 1984), pp. 559-569Published by: Taylor & Francis, Ltd. on behalf of The Society of Vertebrate PaleontologyStable URL: http://www.jstor.org/stable/4523016 .
Accessed: 11/07/2014 21:27
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The Society of Vertebrate Paleontology and Taylor & Francis, Ltd. are collaborating with JSTOR to digitize,preserve and extend access to Journal of Vertebrate Paleontology.
http://www.jstor.org
This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions

This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions

Bar-Yosef & Tchernov, 1972). Hence, the 'Ubeidiya antedates the middle Pleistocene tectonic movement of the Jordan Rift Valley, but was deposited well after the early Pleistocene major tectonic movement, which roughly agrees with the former results cited above. Five paleomagnetic samples extracted from the Li and Lu members (Fig. 1) by G. Kukla (pers. comm.) have all yielded "reversed" results, placing the 'Ubeidiya For- mation within the Matuyama reversed epoch. In con- clusion, a late early Pleistocene age (1,000,000 ybp) is suggested for the 'Ubeidiya sequence. The following fossiliferous layers in the 'Ubeidiya Formation yielded the remains of gerbillids (Fig. 1): II-22, 11-27, 11-36, I- 15, 1-16, 111-12, 111-20. In layer II-23 and II-24 they constitute, after the microtines, the second most dom- inant mammal.
EARLY GERBILLIDS
Leakeymis ternani (Lavocat, 1964) from the middle Miocene of Fort Ternan, Kenya, may be considered the earliest known gerbillid. Its transverse crests gen- erally resemble those seen in Tatera (Tables 1 and 2 in Appendix, and Fig. 2). Lavocat (1964) stressed that this early form represents a clear divergence from the Myocricetodontini ("... molaires a plan g6n6ral cri-
c6todont, mais ou la crete longitudinale est absente ou tres reduite .. ."), but could still be considered a highly evolved cricetid by some because of the presence of the external cingular crest which unites the prelobe with the first full lobe of the first lower molar.
Because of the characteristics of the zygomatic plate, the posterior palatine foramen, and the tooth pat- tern, Jaeger (1969, 1977) concluded that the Myo- cricetodontini can be considered the ancestral group of the gerbillids. He (ibid.) considered Protatera al- gerensis (Amana II, Algeria; Tables 1 and 2 in Appen- dix, and Fig. 2), which is a myocricetodontine, as rep- resenting a possible ancestor of Tatera.
The middle Pliocene Pseudomeriones (Teilhard de Chardin, 1926) and Epimeriones (Daxner-H6ck, 1972) from Asia and Europe (Tables 1 and 2 in Appendix, and Fig. 2) may be indicative of an Asiatic origin of the genus Meriones. Indeed, Pseudomeriones was first recorded from the Pontian of China (Teilhard de Char- din, 1926; Young, 1927; Schaub, 1934), and subse- quently from the upper Ruscinian of Rhodes (Bruijn et al., 1970) and Anatolia (Sen, 1977; Sen & Bruijn, 1977). The genus Pseudomeriones was based on the combined cricetid (alternating fold in M') and gerbillid (three-looped M' and M1 and reduced pattern of M3) features. The Mio-Pliocene Epimeriones (Daxner- H6ck, 1972) (Tables 1 and 2 in Appendix, and Fig. 2) is already much more specialized, with an incisor that has but a single groove, advanced stage of hypsodonty (which shows progress with time; Kowalski, 1974), prismatic pattern of the loop, and absence of any cin- gulum on M2. Weerd et al. (1977) suggested that Epi- meriones is closely related to Blancomys neglectus (Mio- Pliocene of Spain and southern France), having elon-
gated M2, M2, M3 and M3 in relation to M, and a similar reentrant labial fold. None of those forms, nor other gerbillids, which are at present well represented in the arid and semiarid regions of Africa and Asia, may as yet be linked with the fossil form from the 'Ubeidiya Formation.
SYSTEMATIC PALEONTOLOGY
Family GERBILLIDAE
PARAMERIONES, new genus
Generic Diagnosis--The anterior palatine foramen extends posteriorly to the middle alveoli of M'. (In Meriones it extends anterior to alveolar row.) The an- teriormost margin of the posterior palatine foramen begins at the level of the posterior alveoli ofM1, where- as in Meriones it begins at the level of the mesio-lingual
SECTIONS WEST EAST
MEMBER K III I II
Fault FU 92 NAHARAIM FORMATION
86 EROSION
85 42 51 56 33 =43
48-55 28-32 =41-42 47 26-27 =37-40
25 =36* 21 33
FI 26 20 32 29-30 =23-25
22 17-19 =28-31 top 15*-16* =26*"-27* main 15* =22*-25*
13-14 21 20 =20* 6-12
19 =19 19-20 18 17-18
14-17 11-16 LI13 9c,d-10
12* 10-11 9a,b
9 =1 =8 4-8 2-5 =2-7
B- ASE OF 'UBEIDIYA FORMATION NOT EXPOSED - -
FIGURE 1. The four members (ascending) through the sequence of'Ubeidiya Formation (after Picard & Baida, 1966) and correlations between the geological sections at 'Ubeidiya (modified after Bar-Yosef & Tchernov, 1972). Abbrevia- tions: FU, upper fluviatile; LU, upper limnic; FI, lower flu- viatile; LI, lower limnic. = established correlations; * layers containing Parameriones obeidiensis.
560 JVP 4(4), December 1984
This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions

AFRICA ASIA EUROPE East North Near + Middle East Far East
C C
o o
. • Gerbillus + Meriones 0
Parameriones Z Tatera +Gerbillus (Ternifine,Algeria) (Ubeidiya ,Israel) 2 - "(Ubeidiya,lIsrael)
C (Olduvai,Tanzania) C
o >
7 - Gerbillus + Merione Pseudomeriones Pseudomeriones P seudomeriones Epimerionesp.)
8-- _C
S(rhud "Ocre, Algeria)(Romania) 0
4 - C
17- Peoeoe PeoroeoPeocos Emrn
-- Moorrocc
o
e (A n IeBlancomysra
a M
C
(r (Fort TernanKenya) (Beni Mellal, Mroc
8
0c
Morrocco )
FIGURE 2. General distribution of early gerbillids and gerbillid-like cricetids (*) in the Neogene and Pleistocene of Eurasia and Africa. Time scale modified after Berggren & Van Couvering (1974).
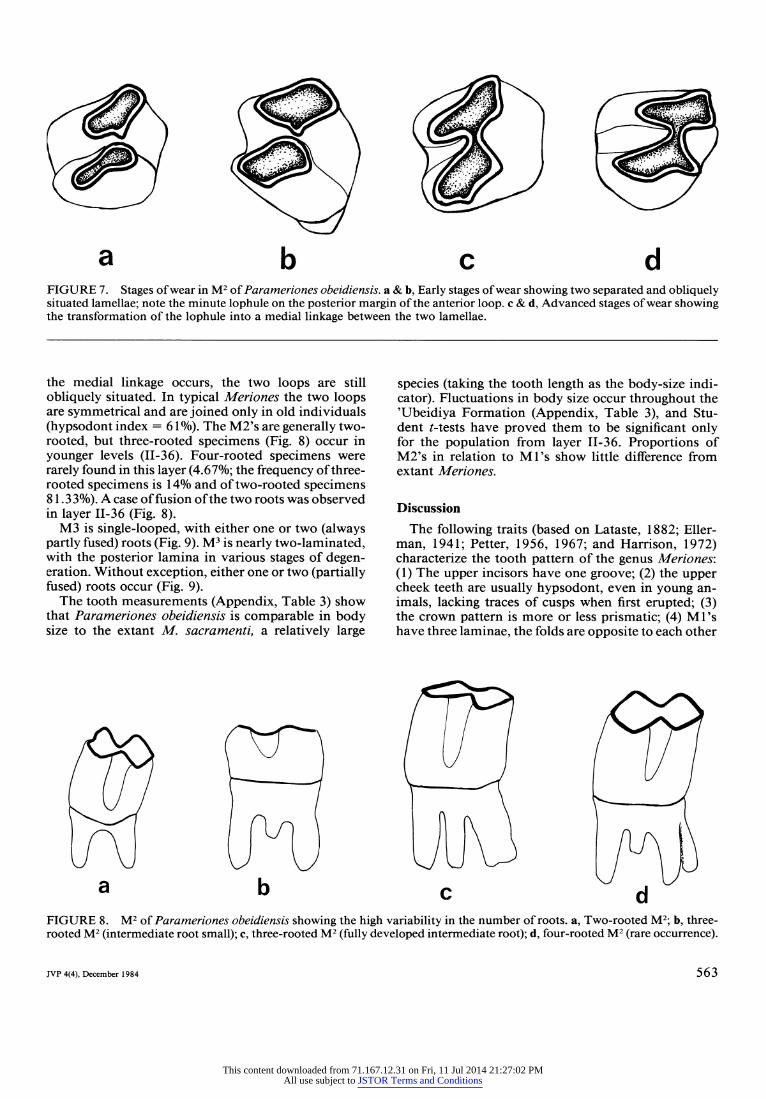
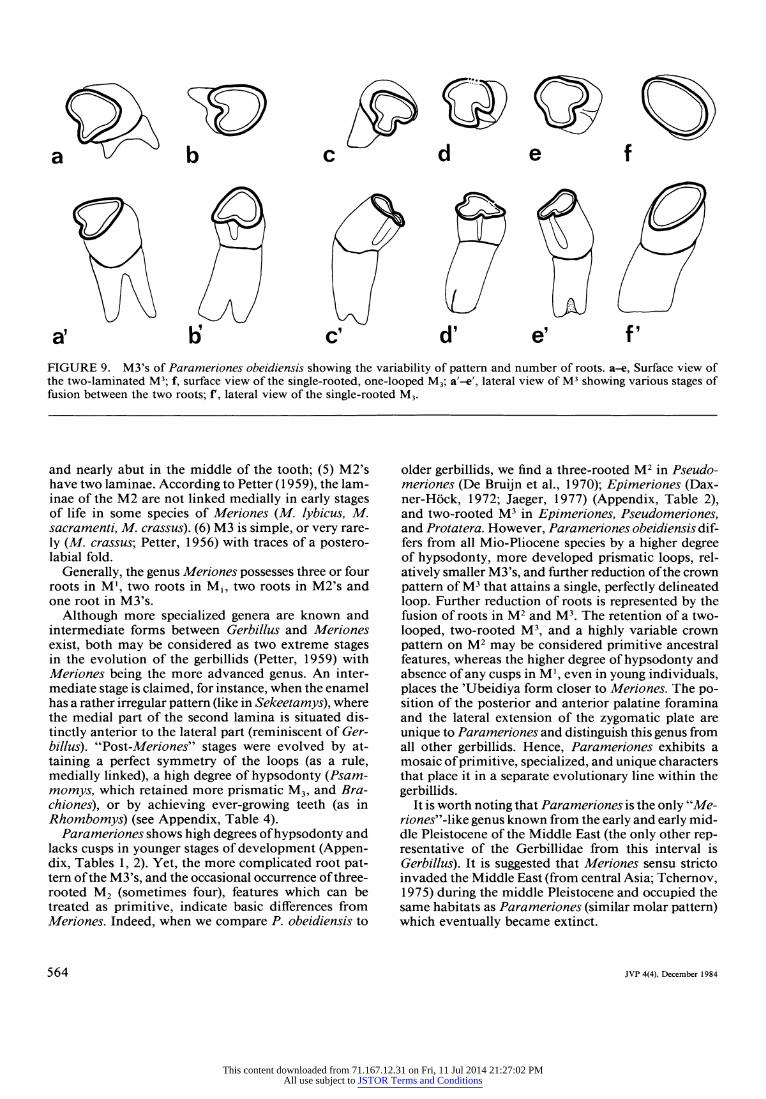
root. This foramen is situated more anteriorly com- pared with Meriones. Parameriones possessess single- grooved upper incisors and hypsodont molars with prismatic loops. M3's are reduced. Two-looped M2 generally two rooted, but occasionally with three roots. Delaminated M2 in young individuals, two-looped M3's with two, more or less fused, roots. One-looped and one-rooted M3.
Parameriones obeidiensis (Haas, 1966) (Figs. 3-9, Tables 1-4)
Holotype--Hebrew University, Jerusalem (HUJ). P.U. 1001 (Fig. 3), an incomplete mandible with M1, M2, M3 (Haas, 1966, pl. XII, fig. 28).
Paratypes-HUJ. P.U. 1002, an incomplete left ra- mus with M1, M2 (Haas, 1966, pl. XIII, fig. 26a, b, c). HUJ. P.U. 1003, an incomplete left ramus with M,, M2 (Haas, 1966, pl. XII, fig. 29). HUJ. P.U. 1004, a
toothless fragment of the right maxillary with alveoli. HUJ. P.U. 1005, fragment of the left maxillary with M', M2, M3 (Fig. 4).
Diagnosis-As for genus. Occurrence--'Ubeidiya, central Jordan Valley, Is-
rael: 'Ubeidiya Formation, late early Pleistocene. La- tamne, Syria: Mindel-Riss, middle Pleistocene, ac- cording to P. Mein, in verbis. A karst fissure filling at Giveat Shaul, near Jerusalem, Isreal: Riss.
Description The zygomatic plate is heavy and projects laterally,
whereas in Gerbillus, Tatera, Meriones, and Psam- momys it extends parallel to the maxilla. The anterior palatine foramen extends posteriorly to the level of mid-M' alveolus, while in Meriones it extends anterior to the alveolar row. The anteriormost margin of the posterior palatine foramen begins at the level of the posterior alveolus of M' and extends to the level of
JVP 4(4), December 1984 561
This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions

This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions
This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions
a b c d e f
a' b' c' d' e' f' FIGURE 9. M3's of Parameriones obeidiensis showing the variability of pattern and number of roots. a-e, Surface view of the two-laminated M3; f, surface view of the single-rooted, one-looped M3; a'-e', lateral view of M3 showing various stages of fusion between the two roots; f', lateral view of the single-rooted M3.
and nearly abut in the middle of the tooth; (5) M2's have two laminae. According to Petter (1959), the lam- inae of the M2 are not linked medially in early stages of life in some species of Meriones (M. lybicus, M. sacramenti, M. crassus). (6) M3 is simple, or very rare- ly (Ai. crassus; Petter, 1956) with traces of a postero- labial fold.
Generally, the genus Meriones possesses three or four roots in M', two roots in M,, two roots in M2's and one root in M3's.
Although more specialized genera are known and intermediate forms between Gerbillus and Meriones exist, both may be considered as two extreme stages in the evolution of the gerbillids (Petter, 1959) with Meriones being the more advanced genus. An inter- mediate stage is claimed, for instance, when the enamel has a rather irregular pattern (like in Sekeetamys), where the medial part of the second lamina is situated dis- tinctly anterior to the lateral part (reminiscent of Ger- billus). "Post-Meriones" stages were evolved by at- taining a perfect symmetry of the loops (as a rule, medially linked), a high degree of hypsodonty (Psam- momys, which retained more prismatic M3, and Bra- chiones), or by achieving ever-growing teeth (as in Rhombomys) (see Appendix, Table 4).
Parameriones shows high degrees of hypsodonty and lacks cusps in younger stages of development (Appen- dix, Tables 1, 2). Yet, the more complicated root pat- tern of the M3's, and the occasional occurrence of three- rooted M2 (sometimes four), features which can be treated as primitive, indicate basic differences from Meriones. Indeed, when we compare P. obeidiensis to
older gerbillids, we find a three-rooted M2 in Pseudo- meriones (De Bruijn et al., 1970); Epimeriones (Dax- ner-Hick, 1972; Jaeger, 1977) (Appendix, Table 2), and two-rooted M3 in Epimeriones, Pseudomeriones, and Protatera. However, Parameriones obeidiensis dif- fers from all Mio-Pliocene species by a higher degree of hypsodonty, more developed prismatic loops, rel- atively smaller M3's, and further reduction of the crown pattern of M3 that attains a single, perfectly delineated loop. Further reduction of roots is represented by the fusion of roots in M2 and M3. The retention of a two- looped, two-rooted M3, and a highly variable crown pattern on M2 may be considered primitive ancestral features, whereas the higher degree of hypsodonty and absence of any cusps in M', even in young individuals, places the 'Ubeidiya form closer to Meriones. The po- sition of the posterior and anterior palatine foramina and the lateral extension of the zygomatic plate are unique to Parameriones and distinguish this genus from all other gerbillids. Hence, Parameriones exhibits a mosaic of primitive, specialized, and unique characters that place it in a separate evolutionary line within the gerbillids.
It is worth noting that Parameriones is the only "Me- riones"-like genus known from the early and early mid- dle Pleistocene of the Middle East (the only other rep- resentative of the Gerbillidae from this interval is Gerbillus). It is suggested that Meriones sensu stricto invaded the Middle East (from central Asia; Tchernov, 1975) during the middle Pleistocene and occupied the same habitats as Parameriones (similar molar pattern) which eventually became extinct.
564 JVP 4(4), December 1984
This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions

This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions

TABLE 2. Dental characteristics of lower molars in fossil and Recent gerbillids. * After P. Mein (pers. comm.), ** After Petter (1956).
Blancomys neglectus Epimeriones Pseudomeriones (Van de Weerd (Daxner-Hock, 1972; (Schaub, 1934)
Leakeymis ternani Protatera algerensis et al., 1977) Kowalski, 1974) China, Rhodes, Meriones Geographic (Lavocat, 1964) (Jaeger, 1977) Spain & southern Austria, Germany, Greece, Turkey, Parameriones Africa and distribution Kenya Algeria France Poland, Hungary Afghanistan Israel, Syria* Asia
M Loops - - 3 3 (prismatic) 3 3 (prismatic) 3 (prismatic) Laminae 3 transverse 3 subtransverse - - - - Sinus 2 transverse 2 obliquely situat- medially linked medially linked medially linked medially linked medially linked
ed Root - 2 - 2 2 2 2
M2 Length - 63% of M, 90% of M 68% of M, 58% of M, 62% of M, 63%of M Loops - - 2 2 (prismatic) 2 2 (in advanced 2 (in adult stage)
wear) Laminae 2 transverse 2 transverse -- - 2 (in young indi- 2 (in very young
viduals) individuals)** Sinus 1 transverse I medially linked I medially linked 1 medially linked 1 laterally linked I medially linked 1 medially linked
with wear with advanced in young stage wear
Root - 2 - 2 2 2 2
M3 Length - 42% of M, 71% of M, 58% of M, 45% of M, 37% of M1 30% of M 66% of M2 79% of M2 85% of M2 78% of M2 60% of M2 53% ofM2
Loops - 1 - 2 prismatic 1 1 1 Laminae 12
.- Sinus - 1 labial cingulum 1 medially linked - - - Root -- 1 - 2 1 1
"4•
This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions
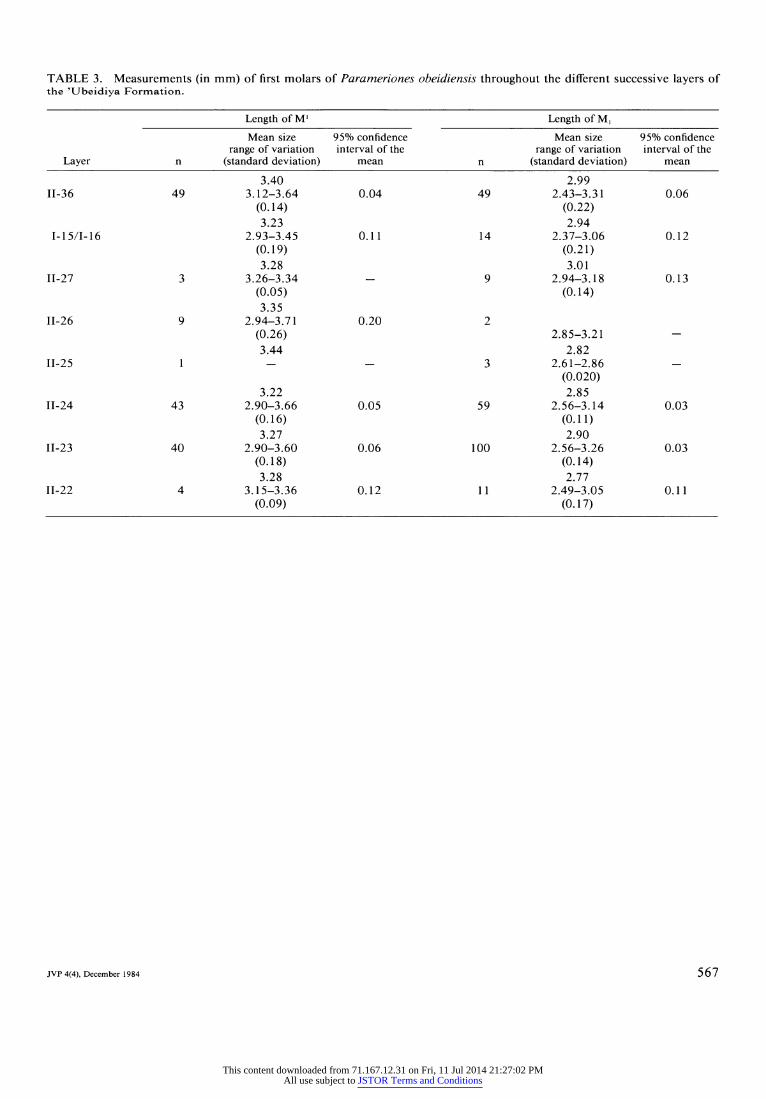
TABLE 3. Measurements (in mm) of first molars of Parameriones obeidiensis throughout the different successive layers of the 'Ubeidiya Formation.
Length of M' Length of M,
Mean size 95% confidence Mean size 95% confidence range of variation interval of the range of variation interval of the
Layer n (standard deviation) mean n (standard deviation) mean
3.40 2.99 II-36 49 3.12-3.64 0.04 49 2.43-3.31 0.06
(0.14) (0.22) 3.23 2.94
I-15/I-16 2.93-3.45 0.11 14 2.37-3.06 0.12 (0.19) (0.21) 3.28 3.01
II-27 3 3.26-3.34 - 9 2.94-3.18 0.13 (0.05) (0.14) 3.35
11-26 9 2.94-3.71 0.20 2 (0.26) 2.85-3.21 - 3.44 2.82
11-25 1 - - 3 2.61-2.86 -
(0.020) 3.22 2.85
II-24 43 2.90-3.66 0.05 59 2.56-3.14 0.03 (0.16) (0.11) 3.27 2.90
II-23 40 2.90-3.60 0.06 100 2.56-3.26 0.03 (0.18) (0.14) 3.28 2.77
II-22 4 3.15-3.36 0.12 11 2.49-3.05 0.11 (0.09) (0.17)
JVP 4(4), December 1984 567
This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions

TABLE 4. Tooth pattern in several modern gerbillids compared with Parameriones obeidiensis. * One groove in early stage (1 month; Petter, 1957), ** with a minute posterolabial fold (Petter, 1957).
Parameriones obeidiensis Tatera Gerbillus Meriones Psammomys Brachiones Rhombomys
Upper incisor one groove one groove one groove one groove ungrooved* one groove two grooves General pattern hypsodont round- brachyodont cus- brachyodont cus- hypsodont pris- hypsodont angu- hypsodont angu- evergrowing
of molars ed prismatic pidate (in pidate (in matic loop lated prismatic lated prismatic rounded pris- loop young); lami- young); lami- loop loop matic loop
nate (in adult) nate (in early subadult)
M' 3 loops 3 laminae 3 loops 3 loloops 3oops 3 loops 3 loops 3 roots 4 roots 3 roots 3-4 roots 2-3 roots 2 roots no roots
M2 2 loops 2 laminae 3 loops 2 loloops 2oops 2 loops 2 loops 2-3 roots 3 roots 3 roots 2 roots 2 roots 2 fused roots no roots
M3 2 loops 2 laminae 1-2 loops 1 loop 2 loops 1 loop 2 loops 1-2 roots 2 fused roots 1 root I root I root 1 root no roots
M, 3 loops 3 laminae 3 loops 3 loops 3 loops 3 loops 3 loops 2 roots 4 roots 2 roots 2-4 roots 2 roots 2 roots no roots
M2 2 loops 2 laminae 2 loops 2 loloops 2oops 2 loops 2 loops 2 roots 2 roots 2 roots 2 roots 2 fused roots 2 fused roots no roots
M3 1 loop 1 loop 1 loop 1 loop 1 loop 1 loop 1 loop 1-2 roots 2 fused roots 1 root 1 root 1 root I root no roots
00 oo
CD C0 .9b
oo
This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions

REFERENCES
Bar-Yosef, O. & Tchernov, E. 1972. On the palaeoecol- ogical history of the site of 'Ubeidiya. The Israel Acad- emy of Sciences and Humanities (Jerusalem), 35 pp.
Berggren, W. A. & Van Couvering, J. A. 1974. The late Neogene. Palaeogeography, Palaeoclimatology, Palaeo- ecology 16:1-216.
Daxner-Hick, G. 1972. Die Wirbeltierfauna aus dem Alt- Pliozdin (Pont) vom Eichk6gel bei Modling (Nieder6s- terreich). IV. Gerbillinae (Rodentia, Mammalia). An- nalen des Naturhistorischen Museums (Vienna) 76:149- 160.
De Bruijn, H., Dawson, M. R. & Mein, P. 1970. Upper Pliocene Rodentia, Lagomorpha and Insectivora (Mam- malia) from the Isle of Rhodes (Greece). Koninklijke Nederlandse Akademie van Vetenschappen, Proceed- ings (B) 73(5):535-564.
Ellerman, J. R. 1941. The Families and Genera of Living Rodents. British Museum (Natural History), London, 347 pp.
Haas, G. 1966. On the vertebrate fauna of the lower Pleis- tocene site of 'Ubeidiya. Israel Academy of Sciences and Humanities (Jerusalem), 68 pp.
Harrison, D. L. 1972. The Mammals of Arabia, Vol. III. Ernest Ben Limited, London, 272 pp.
Horowitz, A. 1973. Development of the Hula Basin, Israel. Israel Journal of Earth Sciences 22:107-139.
1979. The Quaternary of Israel. Academic Press, New York & London, 394 pp.
Jaeger, J. J. 1969. Les rongeurs de Pleistocene moyen de Ternifine (Alg6rie). Comptes-rendus hebdomadaires des
s6ances de l'Acad6mie des Sciences de Paris (D) 269: 1492-1495.
1977. Les rongeurs de Miocene moyen et sup6rieur du Maghreb. Palaeovertebrata 8(1):1-140.
Kowalski, K. 1974. Remains of Gerbillinae (Rodentia, Mammalia) from the Pliocene of Poland. Acad6mie Po- lanaise des Sciences, Ser. biol., Bulletin 22(9):591-595.
Lataste, F. 1882. Mammif'eres nouveaux d'Alg6rie. Le Nat- uraliste 16:1-126.
Lavocat, R. 1964. Fossil rodent from Fort Ternan, Kenya. Nature 201(4937):1131.
Leakey, L. 1967. Preliminary survey of the cultural ma- terial from Beds I and II, Olduvai Gorge, Tanzania; pp. 417-446 in Bishop, W. W. & Clark, J. D. (eds.), Back- ground to Evolution in Africa. University of Chicago Press.
Petter, F. 1959. Evolution du dessin de la surface d'usure des molaires de Gerbillides. Mammalia 23(3):304-315.
Picard, L. 1963. The Quaternary in the northern Jordan
Valley. Israel Academy of Sciences and Humanities, Sec- tion of Sciences, Proceedings 1(4): 1-34.
1965. The geological evolution of the Quaternary in the central and northern Jordan Graben. American Geological Society, Special Paper 84:337-366.
- & Baida, U. 1966. Stratigraphic position of the 'Ubeidiya Formation. Israel Academy of Sciences and Humanities, Section of Sciences, Proceedings 4:1-16.
Schaub, S. 1934. Lieber einige fossile Simplicidentaten aus China und der Mongolei. Schweizerischen Palaeonto- logischen Gesellschaft, Abhandlungen 54:1-40.
Sen, S. 1977. Le fauna de rongeurs Pliocene de yalta (An- kara, Turquie). Mus6um National d'Histoire Naturelle (Paris), Bulletin (3) 465:89-171.
& De Bruijn, H. 1977. Comparaison des rongeurs du N6og6ne terminal de Maritsa, (alta et Tourkobour- nia-l, Mediterran6e orientale. Koninklijke Nederlandse Akademie van Wetenschappen, Proceedings (B) 80(3): 222-227.
Siedner, G. & Horowitz, A. 1974. Radiometric ages of late Cenozoic basalts from northern Israel: chronostrati- graphic implications. Nature 250(5461):23-26.
Stekelis, M. 1966. Archaeological excavations at 'Ubeidiya, 1960-1963. Academy of Sciences and Humanities (Je- rusalem), pp. 1-32.
- , Bar-Yosef, O. & Schick, T. 1969. Archaeological excavations at 'Ubeidiya, 1964-1966. Israel Academy of Sciences and Humanities (Jerusalem), pp. 1-29.
Tchernov, E. 1968. A Pleistocene faunule from a karst fissure filling near Jerusalem. Verhandlungen Naturfor- schung Gesellschaft 79(2): 161-185.
1973. On the Pleistocene molluscs of the Jordan Valley. Israel Academy of Sciences and Humanities (Je- rusalem), pp. 1-50.
1975. Rodent faunas and environmental changes in the Pleistocene of Israel; pp. 331-362 in Prakash, I. & Ghosh, P. K. (eds.), Rodents in Desert Environments. W. Junk, The Hague.
1980. The Pleistocene birds of 'Ubeidiya, Jordan Valley. Israel Academy of Sciences and Humanities (Je- rusalem), pp. 1-83.
Teilhard de Chardin, P. 1926. Note sur quelques rongeurs des Terres Rouges Pontiennes de Kansu. Annales de Paleontologie 15:46-52.
Weerd, A. van de, Adrover, A. R., Mein, P. & Soria, E. 1977. A new genus and species of the Cricetidae (Mammalia, Rodentia) from the Pliocene of southwestern Europe. Koninklijke Nederlandse Akademie van Wetenschap- pen, Proceedings (B) 80(5):429-439.
Young, C. C. 1927. Fossile Nagetiere aus Nord-China. Pa- laeontologia Sinica 5(3): 1-82.
JVP 4(4), December 1984 569
This content downloaded from 71.167.12.31 on Fri, 11 Jul 2014 21:27:02 PMAll use subject to JSTOR Terms and Conditions





























