Embed Size (px)
Citation preview

© 2021 Journal of Cancer Research and Practice | Published by Wolters Kluwer - Medknow 41
Abstract
Original Article
Background: Curcumin and its metabolite, tetrahydrocurcumin (THC), have been widely studied due to their compelling capabilities in the prevention of various diseases. However, these compounds face some shortcomings, including the bioavailability of curcumin and comparatively weaker anti-inflammatory effects of THC. The amination of natural compounds in the hosts’ colons has garnered attention because these aminated compounds retain and even increase their bioactivity relative to their original counterparts. Materials and Methods: The existence of THC-NH2 as a metabolite of THC in mice feces was analyzed by using LC-MS. Three-week DSS-induced colitis in mice experiment was designed to confirm the ameliorative effect of THC-NH2 on inflammatory bowel disease. The anti-inflammatory effect of THC-NH2 on LPS-treated murine macrophage RAW264.7 cell line was further clarified in vitro. Results: In this study, the metabolite 3-amino-3-deoxytetrahydrocurcumin (THC-NH2) was discovered in the feces of mice administered with THC. Compared to THC, THC-NH2 exhibits greater anti-inflammatory effects in terms of nitric oxide production. In a study of dextran sulfate sodium-induced colitis in which animal subjects were supplied with both THC and THC-NH2, each sample displayed encouraging but not compelling effects on inflammation reduction. In vitro research revealed that intervention using THC-NH2 could significantly reduce protein expression levels of nitric oxide synthase (iNOS) but cause the accumulation of cyclooxygenase-2 (COX-2). By using cycloheximide, THC-NH2 was found to retard the degradation of COX-2 by increasing its stability. However, mRNA levels of COX-2 and concentrations of prostaglandin E2 (PGE2) in spent medium indicated that COX-2 activity did not increase alongside its accumulated protein level, though no significant effects on the reduction of COX-2 activity were seen. Conclusion: As the current body of research is inadequate, in order to ensure that all things are considered, the efficacy and safety of THC-NH2 as a pharmaceutical drug require further investigation. Nevertheless, recent results showed that THC-NH2 can be used in multi-targeting anti-inflammation drugs to inhibit iNOS levels and reduce the side effects of COX-2 inhibitors by acting as a competitive inhibitor.
Keywords: 3-amino-3-deoxytetrahydrocurcumin, amination, cyclooxygenase-2, inflammation, tetrahydrocurcumin
Address for correspondence: Dr. Min‑Hsiung Pan, Institute of Food Science and Technology, National Taiwan University,
No. 1, Section 4, Roosevelt Road, Taipei 10617, Taiwan. E‑mail: [email protected]
Access this article online
Quick Response Code:Website: www.ejcrp.org
DOI: 10.4103/JCRP.JCRP_21_20
This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution‑NonCommercial‑ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non‑commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
For reprints contact: [email protected]
How to cite this article: Koh YC, Liu SY, Wu JC, Chou YC, Nagabhushanam K, Ho CT, et al. A new metabolite: The effects of aminated tetrahydrocurcumin on inducible nitric oxide synthase and cyclooxygenase-2. J Cancer Res Pract 2021;8:41-53.
A New Metabolite: The Effects of Aminated Tetrahydrocurcumin on Inducible Nitric Oxide Synthase and Cyclooxygenase‑2
Yen‑Chun Koh1, Sheng‑Yang Liu1, Jia‑Ching Wu1, Ya‑Chun Chou1, Kalyanam Nagabhushanam2, Chi‑Tang Ho3, Min‑Hsiung Pan1,4,5*1Institute of Food Sciences and Technology, National Taiwan University, Taipei, Taiwan
2Sabinsa Corporation, East Windsor, New Jersey, USA 3Department of Food Science, Rutgers University, New Brunswick, New Jersey, USA
4Department of Medical Research, China Medical University Hospital, China Medical University, Taichung City, Taiwan 5Department of Health and Nutrition Biotechnology, Asia University, Taichung City, Taiwan
Journal of Cancer Research and Practicejournal homepage: www.ejcrp.org
Submitted: 06-Jul-2020 Revised: 26-Jul-2020 Accepted: 27-Aug-2020 Published: 01-Jun-2021
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]

Highlights• 3‑Amino‑3‑deoxytetrahydrocurcumin (THC‑NH2) may be a natural aminated compound found in the colons of tetrahydrocurcumin (THC)‑administered
mice• Both THC and THC‑NH2 exhibit ameliorative effects on dextran sulfate sodium‑induced colitis in mice• THC‑NH2 inhibits the mRNA expression levels of nitric oxide synthase (iNOS) and cyclooxygenase‑2 (COX‑2) via repression of the phosphorylation
by the PI3K/Akt pathway• THC‑NH2 suppresses iNOS protein levels and nitric oxide production• THC‑NH2 retards the degradation of COX2 protein but does not lead to prostaglandin E2 (PGE2) elevation
Koh, et al.: Journal of Cancer Research and Practice (2021)
42 Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
IntRoductIon
Curcumin, also known as diferuloylmethane, is a yellow crystalline coloring agent isolated from Curcuma longa, and its capabilities in combating oxidative stress, inflammation, diabetes, virus infection, and neurodamage are widely known, discussed, and studied.[1] THC is a colorless/white metabolite of curcumin which is first identified by Holder et al., in 1978 as a major biliary metabolite in rats caused by orally administered curcumin, although it can occur naturally as a minor component of turmeric as well.[2,3] It has been suggested that as a reductive metabolite, THC shows higher stability in pH levels as compared to the major turmeric components, such as curcumin, demethoxycurcumin, and bisdemethoxycurcumin.[4] Unlike other biologically inactive curcumin metabolites, such as curcumin glucuronide and curcumin sulfate, THC is relatively active in its pharmacological activities, especially its sight catching antioxidative properties. However, due to its insufficient production of α,β-dienes after hydrogenation, THC is unable to form Michael adducts with cysteine-rich proteins.[1] Other major advantages of THC compared to curcumin are its bioavailability and lack of observable adverse effects at 400 mg/kg a day in rats.[5] Therefore, even though THC is not found to be as effective as curcumin, more studies find it necessary to propose and support its efficacy.
As a source of nitrogen for amino acid metabolism, protein synthesis, and maintenance of pH homeostasis, ammonia is an important component of many processes. However, high blood ammonia concentrations can lead to neurological dysfunction, cerebral edema, and seizures.[6] Therefore, ammonia generated in the intestine must be detoxified in the liver through the urea cycle to reduce neurotoxicity. In 2019, it was reported that, when facilitated by microbiota, green tea polyphenol (-)-epigallocatechin-3-gallate (EGCG) can form aminated metabolites. Surprisingly, it was found that aminated EGCG retains the bioactivities of EGCG, including inhibiting cancer cell growth and exhibiting anti-inflammatory effects. Furthermore, EGCG amination could also be used to trap ammonia, which explains reports that observed reductions in elderly fecal ammonia concentrations after the intake of tea catechins.[7] In addition to EGCG, Zhang et al. demonstrated that myricetin could be aminated in vitro via chemical reactions between myricetin quinone and ammonia.[8] Similarly, aminated myricetin also retained the same anti-inflammatory capacities as its parent compound. Therefore, amination of natural compounds may be a novel but viable strategy for the enhancement of biological activity.
Inf lammatory bowel disease (IBD) is a chronic immune-mediated disease that can be attributed to several factors, although the precise etiology has not yet been defined and is commonly known as a gut inflammation
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]

Koh, et al.: Journal of Cancer Research and Practice (2021)
43Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
disease with the potential to relapse.[9,10] Dextran sulfate sodium (DSS) is now widely used to induce colitis in animal models as this can successfully produce many clinical symptoms of IBD, especially the features of human ulcerative colitis, such as immune cell infiltration, damaging epithelial linings, and endotoxemia caused by bacteria-secreted lipopolysaccharides (LPSs).[11,12] A disease activity index (DAI) is commonly used to assess the severity of DSS-induced colitis by examining positively correlated characteristics, including weight loss, bloody feces (hematochezia), and stool consistency as a result of consecutive episodes of diarrhea.[13]
Nitric oxide (NO) is a short-lived signaling molecule produced by NO synthases (iNOSs). Of the three isoforms of iNOS, elevated expressions of inducible iNOS are most commonly accompanied by inflammation, and NO has thus become an indication of inflammatory response.[14] NO produced by iNOS is cytotoxic and can cause damage to DNA and RNA.[15] iNOS expression, in terms of mRNA and protein levels, increases colonic tissue as well as splenic and peritoneal macrophages in mice models with DSS-induced colitis.[9,16] cyclooxygenase-2 (COX-2) is an important target of many anti-inflammatory drugs because it is commonly induced by inflammatory conditions.[17] As the key enzyme in prostaglandin synthesis, COX-2 is responsible for the production of PGE2, resulting in enhanced proliferation of cells, inhibition of apoptosis, and promotion of angiogenesis.[18] Overexpression of COX-2 can lead to chemoresistance, cancer invasion (particularly breast cancer), and bone metastasis due to interleukin-8 stimulation.[17] iNOS and COX-2 are two major target proteins to evaluate with regard to pro-inflammatory response because they have been widely studied. Therefore, in this study, to assess the anti-inflammatory effects of the newly identified curcumin-related compound 3-amino-3-deoxytetrahydrocurcumin (THC-NH2), iNOS and COX-2 were selected as the primary indicators of inflammation.
THC-NH2 is a compound that occurs naturally in the colon, possibly as a product yielded from ammonia elimination, though it has not been discussed in previous studies. Therefore, the anti-inflammatory efficacy of THC-NH2 compared to THC was investigated and evaluated through animal and in vitro experimentation to uncover its potential value and the underlying molecular mechanisms of this newfound compound on inflammation-related disease.
MateRIals and Methods
Reagents and antibodiesAntibodies β-actin, ERK, p-ERK, JNK, p-JNK, p-38, p-p38, Akt, and p-Akt were purchased from Cell Signal Technology (Beverly, MA, USA). P-PI3K (Try508) and PI3K were purchased from Santa Cruz Biotechnology, Inc., (Dallas, TX, USA). Cycloheximide and chemical reagents were purchased from Sigma-Aldrich Corporation (St. Louis, MO, USA). Sample standard THC-NH2 was synthesized and generously provided by Sabinsa Corporation (East Windsor, NJ, USA).
Animal experiment design and animal careThe first animal study was designed to ensure that THC can be aminated after absorption. The experiment was conducted according to the protocol approved by National Taiwan University’s Institutional Animal Care and Use Committee (NTU106-EL-00163). Twenty Institute of Cancer Research (ICR) mice were purchased from the BioLASCO Experimental Animal Center (Taipei, Taiwan). The animals were randomly assigned into four groups of five mice each, fed with either a normal diet (ND, laboratory rodent diet 5001) or an ND with 2% L-glutamine. Among them, one ND group and one 2% L-glutamine group were administered with THC (250 mg/kg) dissolved in corn oil. Feces were collected 3 and 9 h after administration. THC and THC-NH2 were extracted from the feces and assessed using high-performance liquid chromatography (HPLC). In this process, the collected feces were treated with 1 mL of acetonitrile in ultrasonic oscillation for 10 min and then filtered using a 0.22 µm Polytetrafluoroethylene (PTFE) filter membrane. This process was repeated to collect 5 mL of extract. Finally, the extracts were diluted with 50% acetonitrile 200 times before HPLC analysis.
The second animal study consisted of six groups of six mice each, for a total of 36 5-week-old ICR mice. The experimental procedures were the same as those described above. The mice were housed under controlled conditions of 25°C ± 1°C, with a relative humidity of 50% and a light-dark cycle of 12 h. The animals were randomly assigned into control (ND, fed with standard diet), DSS (fed standard diet and 2% DSS solution as drinking water), and four experimental DSS-induced IBD groups fed with diets of 0.005% THC, 0.02% THC, 0.005% THC-NH2, and 0.02% THC-NH2, respectively. Two percent DSS solution was prepared by dissolving 2 g DSS in deionized water, resulting in 100 mL of final solution. The mice received the DSS solution for 7 days, and the sample
Table 1: The criteria of scores used for a disease activity index analysis
Score
0 1 2 3 4Weight loss (%) No weight loss 1–5 5–10 10–15 >20Stool consistency Normal - Loose stool - DiarrheaFeces bleeding No blood Hemoccult positive Hemoccult positive and visual pellet bleeding - Gross bleeding, blood around anus
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]

Koh, et al.: Journal of Cancer Research and Practice (2021)
44 Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
diets were administered a week early as a pretreatment. The mice were inspected daily, and their body weight changes, stool consistencies, and levels of feces bleeding were recorded as criteria scores, as shown in Table 1. The sum of scores was used as a DAI to predict DSS-induced colitis before dissections. Animals were sacrificed 3 days after induction. The entire colons were dissected from the cecum to the anus, and the lengths and weights of the colons were recorded.
L iqu id chromatography–mass spec t romet r y analysis parameters for tetrahydrocurcumin and 3‑amino‑3‑deoxytetrahydrocurcuminThe ACE C18 Column (2.1 mm ID × 150 mm, 2 µm) was chosen for the liquid chromatography, and the operation thermostat was set to 45°C. 0.1% formic acid in the deionized water comprised mobile phase A, while mobile phase B was composed of acetonitrile with 0.1% formic acid. The injection volume was set to 10 µL with a flow rate of 0.3 mL/min. For the mass spectrometry parameter, the source temperature was set to 150°C while the desolvation temperature was 400°C. The capillary voltage was 3000 V and the cone voltage was 10 V. Cone gas flow was maintained at 20 L/h while desolvation gas flow was 500 L/h. The precursor ions and product ions of standard samples are quantified Table 2.
Cell culture and measurement of cell viabilityA RAW 264.7 murine macrophage purchased from American Type Culture Collection (ATCC, Rockville, MD, USA) was cultured in Dulbecco’s modified Eagle medium supplemented with 10% heat-inactivated fetal bovine serum (FBS, GIBCO, Grand Island, NY, USA) and a 1% solution of penicillin and streptomycin under the conditions of a 5% CO2 atmosphere at 37°C. Cell viability was analyzed using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay. The cells were seeded on 96-well plates at a density of
1 × 106 cells/well for 12 h before being treated with THC-NH2 at various concentrations for 24 h both with and without LPS (100 ng/mL). Absorbance was measured at 570 nm to ensure that the sample concentration selected did not exhibit cytotoxicity before further analysis.
NO measurement by nitrite assayNitrogen oxide (NO) released from an LPS-induced RAW264.7 cell line is an indicative of inflammatory response, which can be measured in terms of nitrite concentrations using a nitrite assay. After seeding for 12 h, the cells were treated with samples at final concentrations of 10, 20, 30, and 40 µM and an LPS concentration of 100 ng/mL. After a treatment period of 24 h, 100 µL medium was mixed with an equivalent volume of Griess reagent to produce a pinkish azo compound in the presence of nitrite. Absorbancy was read with an enzyme-linked immune sorbent assay plate reader (ELISA reader, Dynatech MR-7000; Dynatech Laboratories, Chantilly, VA, USA) at 550 nm, and nitrite concentrations were calculated using a standard curve made with known concentrations of NaNO3.
RNA extraction, reverse transcription, and quantitative real‑time polymerase chain reactionRAW264.7 RNA was extracted using TRIzol reagent, and phase separation was performed by adding chloroform at a ratio of 1:5. After centrifugation, the aqueous phase on the top was carefully transferred to a new tube. Five hundred microliters isopropanol was added to the aqueous phase and incubated at room temperature for 10 min followed by the centrifugation for RNA precipitation. Organic reagents of the precipitate were removed using 75% ethanol. At last, ethanol was removed and precipitate was air-dried using laminar flow before re-suspending the pellet in autoclaved ddH2O to obtain an mRNA solution.
Before reverse transcribing the mRNA into cDNA, the quality and the concentration of RNA were analyzed using a NanoDrop 1000 Spectrophotometer. Reverse transcription was performed using a SensiFAST™ cDNA synthesis kit according to the manufacturer’s instructions. Reverse transcription was carried out under the following conditions: primer annealing at 25°C for 10 min, extension at 42°C for 15 min, and finally, enzyme inactivation at 85°C for 5 min. After cooling down, cDNA was stored at −20°C before use as a template for polymerase chain reaction (PCR).
mRNA levels were de termined v ia quant i ta t ive RT-PCR using a StepOnePlus RT-PCR system and SYBR Green Master Mix. The mRNA levels were measured using relative quantification compared to a constitutively expressed gene (glyceraldehyde 3-phosphate dehydrogenase [GAPDH]). The primer sequences of GAPDH, iNOS, and COX-2 are listed in Table 3.
Western blottingThe cells were washed with phosphate-buffered saline, collected, and centrifuged at 5000 × g for 10 min at 4°C. The
Table 3: Primer sequences of target genes
Target genes Primer sequencesGAPDH Forward: 5’-TCAACGGCACAGTCAAGG-3’
Reverse: 5’-ACTCCACGACATACTCAGC-3’iNOS Forward: 5’-ACCCTAAGAGTCACCAAAATGG-3’
Reverse: 5’-CCAGGGATTCTGGAACATTCT-3’COX-2 Forward: 5’-GGGAGTCTGGAACATTGTGAA-3’
Reverse: 5’-GCACATTGTAAGTAGGTGGACTGT-5’
Table 2: The quantitative product ions and precursor ions of standard samples
Compound Precursor ion (m/z)
Product ion (collision energy)
Quantitative ion Qualitative ionCurcumin 369 285 (10) 161 (20)THC 373 137 (15) 179 (15)THC-NH2 372 236 (15) 178 (15)THC: Tetrahydrocurcumin
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]

Koh, et al.: Journal of Cancer Research and Practice (2021)
45Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
supernatants were removed, and the pellets were lysed with a gold lysis buffer for 30 min and vortexed for a few seconds at 5-min intervals. The lysates collected after centrifugation were stored at under −20°C for western blotting. Protein concentration was measured using a Bio-Rad protein assay (Bio-Rad Laboratories, Inc., Hercules, CA, USA), and the western blotting process was conducted as follows. After separating protein samples with 10% sodium dodecyl sulfate-polyacrylamide gels, the proteins were transferred to a
polyvinylidene difluoride transfer membrane (Immobilon®-P, Merck Millipore Ltd., Tullagreen, County Cork, Ireland). The transferred membranes were blocked with a blocking solution for 1 h and then incubated with primary antibodies at under 4°C overnight. The membranes were washed with 0.2% phosphate buffered saline tween-20 (TPBS) three times before and after probing with secondary antibodies for 1 h. At last, the density of protein bands was quantified using ImageJ software (National Institutes of Health, USA).
Figure 1: Conversion of tetrahydrocurcumin into 3‑amino‑3‑deoxytetrahydrocurcumin in mice colon. (a) Experimental design to investigate the occurrence of amination on tetrahydrocurcumin. The representative figures of compounds potentially present in feces, analyzed with liquid chromatography/mass spectrometry, (b) standard compounds (tetrahydrocurcumin, 3‑amino‑3‑deoxytetrahydrocurcumin and curcumin), (e) tetrahydrocurcumin, and (f) aminated‑tetrahydrocurcumin extracted from feces. Quantified concentration (µg/g of feces) of (c) tetrahydrocurcumin and (d) 3‑amino‑3‑deoxytetrahydrocurcumin extracted from feces collected at 3 and 9 h after oral administration of tetrahydrocurcumin. Administration dosage of tetrahydrocurcumin was 250 mg/kg
e
d
b
a
f
c
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]

Figure 2: 3‑Amino‑3‑deoxytetrahydrocurcumin but not tetrahydrocurcumin reduced nitrite production of lipopolysaccharides‑induced murine macrophages RAW264.7 without causing cytotoxicity. Cell viability of tetrahydrocurcumin‑ and 3‑amino‑3‑deoxytetrahydrocurcumin (10–40 µM)‑treated RAW264.7 (a) without lipopolysaccharides and (c) with lipopolysaccharides inducement at concentration of (100 ng/mL). (b and d) Nitrite production of lipopolysaccharides‑induced RAW264.7 after being treated with either tetrahydrocurcumin or 3‑amino‑3‑deoxytetrahydrocurcumin. *** and **** indicate significant differences of P < 0.005 and P < 0.001 between the compared groups. Different lower cases indicate significant difference (P < 0.05) among the groups
db
ca
Koh, et al.: Journal of Cancer Research and Practice (2021)
46 Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
Protein translation inhibitionCycloheximide is a common reagent for protein synthesis inhibition as it can inhibit eukaryotic translation. To determine protein stability, cycloheximide was added to LPS-induced RAW264.7 18 h after inducement to inhibit protein translation. The cells were collected for protein isolation, and then, western blotting was subsequently conducted to investigate and compare protein levels.
Statistical analysisFor the purposes of statistical analysis, data were expressed as mean ± standard deviation. A Student’s t-test was used to assess the differences between the means of the sample groups and those of the control and induction groups, while a one-way ANOVA and Tukey’s multiple comparison test were selected to assess the significance of the multiple comparisons. The accepted level of statistical significance was P ≤ 0.05.
Results
3‑Amino‑3‑deoxytetrahydrocurcumin is a metabolite of tetrahydrocurcumin intertransformed in ICR miceThe aminated metabolite THC-NH2 was found in the feces of mice orally administered with THC. Both THC and THC-NH2 were detected in the feces of mice who received THC with and without having been fed diets supplemented with 2% L-glutamine, as shown in Figure 1, which indicates that a supplement of 2% L-glutamine may not be compulsory
for THC-NH2 production and the source of nitrogen could be provided primarily through diet. THC concentrations in the feces of mice that received L-glutamine and THC were lower than that those in the feces of mice that received only THC, which may suggest that L-glutamine increases the rate of THC conversion into aminated THC. However, no significant difference in terms of aminated THC concentration was observed between the two groups. Meanwhile, concentrations of THC-NH2 increased within 3 h after oral administration. After 9 h, the concentration of THC in feces sharply decreased due to absorption and conversion, while up to 21.15 µg of THC-NH2 remained in each gram of feces in both experimental groups with no significant difference found between them. These results indicate that conversion of THC into THC-NH2 naturally occurs when metabolites and nitrogen sources of amination are obtained from diet, and supplementation of L-glutamine may facilitate conversion effects.
3‑Amino‑3‑deoxytetrahydrocurcumin reduces NO production without exerting cytotoxic effects on RAW264.7To predict the anti-inflammatory capacity of THC-NH2, NO production was determined by treating THC-NH2 with LPS-induced RAW264.7. Before that, the cell viability of RAW264.7 treated with different concentrations of THC-NH2 was investigated. As illustrated in Figure 2a, neither THC nor THC-NH2 caused cytotoxicity at concentrations up to 40 µM. The NO production of THC and THC-NH2 was then analyzed
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]
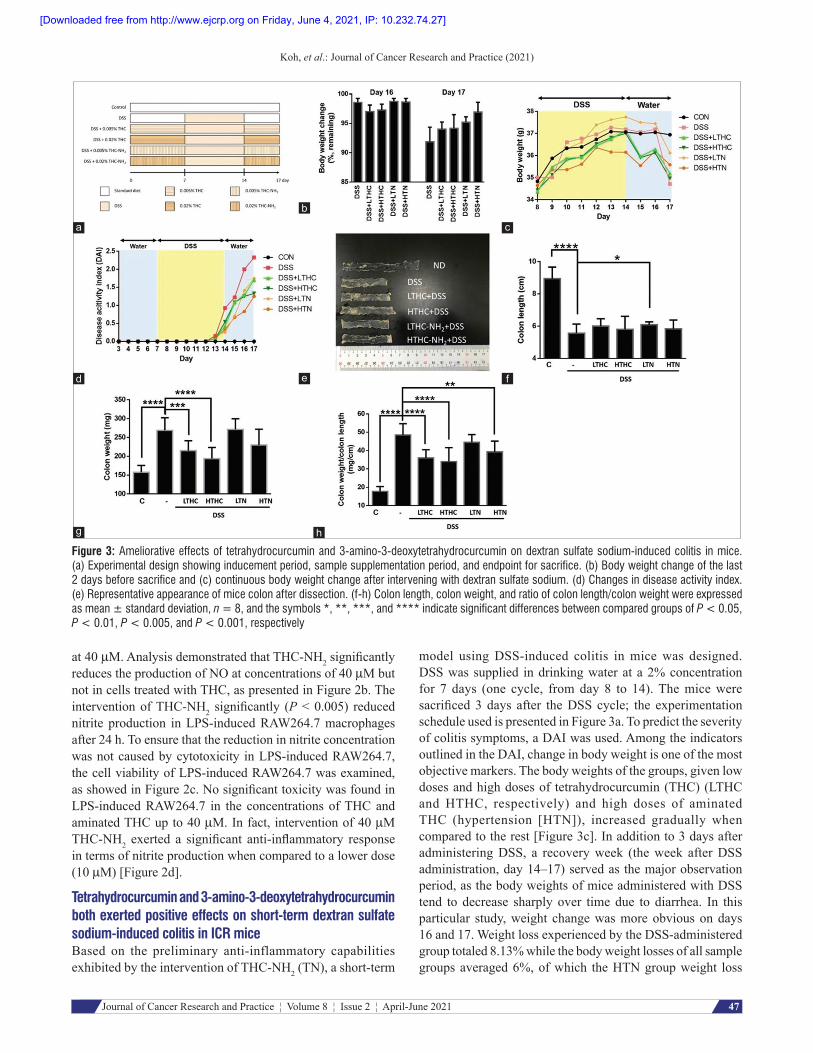
Figure 3: Ameliorative effects of tetrahydrocurcumin and 3‑amino‑3‑deoxytetrahydrocurcumin on dextran sulfate sodium‑induced colitis in mice. (a) Experimental design showing inducement period, sample supplementation period, and endpoint for sacrifice. (b) Body weight change of the last 2 days before sacrifice and (c) continuous body weight change after intervening with dextran sulfate sodium. (d) Changes in disease activity index. (e) Representative appearance of mice colon after dissection. (f‑h) Colon length, colon weight, and ratio of colon length/colon weight were expressed as mean ± standard deviation, n = 8, and the symbols *, **, ***, and **** indicate significant differences between compared groups of P < 0.05, P < 0.01, P < 0.005, and P < 0.001, respectively
d
h
c
g
b
f
a
e
Koh, et al.: Journal of Cancer Research and Practice (2021)
47Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
at 40 µM. Analysis demonstrated that THC-NH2 significantly reduces the production of NO at concentrations of 40 µM but not in cells treated with THC, as presented in Figure 2b. The intervention of THC-NH2 significantly (P < 0.005) reduced nitrite production in LPS-induced RAW264.7 macrophages after 24 h. To ensure that the reduction in nitrite concentration was not caused by cytotoxicity in LPS-induced RAW264.7, the cell viability of LPS-induced RAW264.7 was examined, as showed in Figure 2c. No significant toxicity was found in LPS-induced RAW264.7 in the concentrations of THC and aminated THC up to 40 µM. In fact, intervention of 40 µM THC-NH2 exerted a significant anti-inflammatory response in terms of nitrite production when compared to a lower dose (10 µM) [Figure 2d].
Tetrahydrocurcumin and 3‑amino‑3‑deoxytetrahydrocurcumin both exerted positive effects on short‑term dextran sulfate sodium‑induced colitis in ICR miceBased on the preliminary anti-inflammatory capabilities exhibited by the intervention of THC-NH2 (TN), a short-term
model using DSS-induced colitis in mice was designed. DSS was supplied in drinking water at a 2% concentration for 7 days (one cycle, from day 8 to 14). The mice were sacrificed 3 days after the DSS cycle; the experimentation schedule used is presented in Figure 3a. To predict the severity of colitis symptoms, a DAI was used. Among the indicators outlined in the DAI, change in body weight is one of the most objective markers. The body weights of the groups, given low doses and high doses of tetrahydrocurcumin (THC) (LTHC and HTHC, respectively) and high doses of aminated THC (hypertension [HTN]), increased gradually when compared to the rest [Figure 3c]. In addition to 3 days after administering DSS, a recovery week (the week after DSS administration, day 14–17) served as the major observation period, as the body weights of mice administered with DSS tend to decrease sharply over time due to diarrhea. In this particular study, weight change was more obvious on days 16 and 17. Weight loss experienced by the DSS-administered group totaled 8.13% while the body weight losses of all sample groups averaged 6%, of which the HTN group weight loss
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]

Figure 4: Effects of tetrahydrocurcumin and 3‑amino‑3‑deoxytetrahydrocurcumin on organ status of dextran sulfate sodium‑induced mice. (a‑c) Organ weight expressed as mean ± standard deviation, and significant differences among different groups (P < 0.05) are indicated by symbol *. (d) Representative appearance of liver, kidneys, and spleen. Size of each background square box is 1 cm2
d
cba
Figure 5: 3‑Amino‑3‑deoxytetrahydrocurcumin inhibits nitric oxide synthase expression in terms of protein level and mRNA level but leads to cyclooxygenase‑2. protein accumulation. (a) Relative protein expressions of nitric oxide synthase and cyclooxygenase‑2 of 24‑h lipopolysaccharides‑induced RAW264.7 treated with tetrahydrocurcumin or 3‑amino‑3‑deoxytetrahydrocurcumin, using β‑actin as internal control. (b and c) Relative protein expression of nitric oxide synthase and cyclooxygenase‑2 using β‑actin as internal control. (d‑e) Relative mRNA level of nitric oxide synthase and cyclooxygenase‑2 by using glyceraldehyde 3‑phosphate dehydrogenase as internal control. Symbols *, **, ***, and **** indicate significant differences between compared groups of P < 0.05, P < 0.01, P < 0.005, and P < 0.001, respectively
dc
ba
e
Koh, et al.: Journal of Cancer Research and Practice (2021)
48 Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
was the lowest (3.07%), indicating that all samples did exhibit positive impacts on DSS-induced colitis [Figure 3b].
Furthermore, the effects on samples in terms of DAI scoring were determined by assembling scores (from 0 to 3) which include assessment of stool characteristics, body weight, and bleeding levels, as presented in Table 1. DAI scores of all DSS-treated groups dramatically increased from day 12 onward and the DSS-only group had the highest DAI score at the end of day 17, followed by the groups that received low doses of both THC and
THC-NH2, while the groups that received high doses of THC and THC-NH2 had the lowest DAI scores among all induced groups [Figure 3d]. After colon dissection, colon length and colon weight become the primary indicators of the effects of DSS. DSS inducement significantly reduced colon length (P < 0.001) and increased colon weight (P < 0.001) in comparison to control groups [Figure 3e]. In the experimental groups, most samples did not exhibit positive results in terms of inhibiting colon shortening, but both LTHC and HTHC seemed to reverse colon thickening
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]
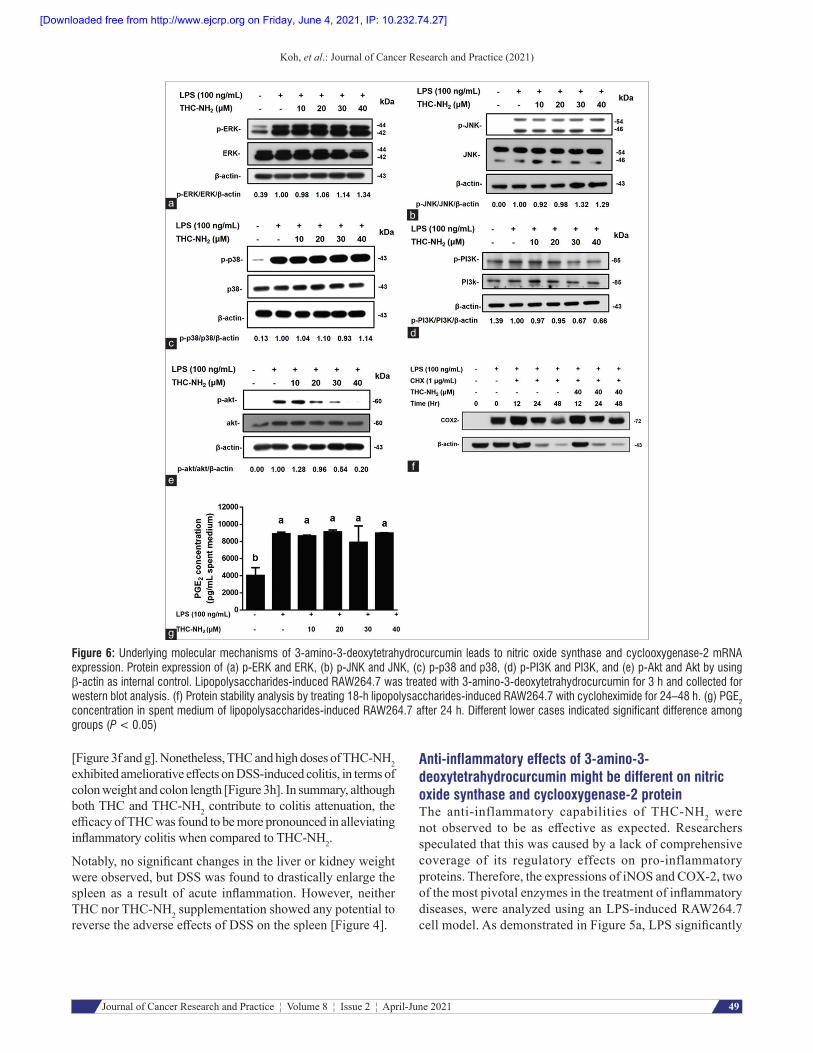
Figure 6: Underlying molecular mechanisms of 3‑amino‑3‑deoxytetrahydrocurcumin leads to nitric oxide synthase and cyclooxygenase‑2 mRNA expression. Protein expression of (a) p‑ERK and ERK, (b) p‑JNK and JNK, (c) p‑p38 and p38, (d) p‑PI3K and PI3K, and (e) p‑Akt and Akt by using β‑actin as internal control. Lipopolysaccharides‑induced RAW264.7 was treated with 3‑amino‑3‑deoxytetrahydrocurcumin for 3 h and collected for western blot analysis. (f) Protein stability analysis by treating 18‑h lipopolysaccharides‑induced RAW264.7 with cycloheximide for 24–48 h. (g) PGE2 concentration in spent medium of lipopolysaccharides‑induced RAW264.7 after 24 h. Different lower cases indicated significant difference among groups (P < 0.05)
e
d
a
c
g
b
f
Koh, et al.: Journal of Cancer Research and Practice (2021)
49Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
[Figure 3f and g]. Nonetheless, THC and high doses of THC-NH2 exhibited ameliorative effects on DSS-induced colitis, in terms of colon weight and colon length [Figure 3h]. In summary, although both THC and THC-NH2 contribute to colitis attenuation, the efficacy of THC was found to be more pronounced in alleviating inflammatory colitis when compared to THC-NH2.
Notably, no significant changes in the liver or kidney weight were observed, but DSS was found to drastically enlarge the spleen as a result of acute inflammation. However, neither THC nor THC-NH2 supplementation showed any potential to reverse the adverse effects of DSS on the spleen [Figure 4].
Anti‑inflammatory effects of 3‑amino‑3‑ deoxytetrahydrocurcumin might be different on nitric oxide synthase and cyclooxygenase‑2 proteinThe anti-inflammatory capabilities of THC-NH2 were not observed to be as effective as expected. Researchers speculated that this was caused by a lack of comprehensive coverage of its regulatory effects on pro-inflammatory proteins. Therefore, the expressions of iNOS and COX-2, two of the most pivotal enzymes in the treatment of inflammatory diseases, were analyzed using an LPS-induced RAW264.7 cell model. As demonstrated in Figure 5a, LPS significantly
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]

Koh, et al.: Journal of Cancer Research and Practice (2021)
50 Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
induced iNOS expression when compared to its control counterpart, and THC-NH2 was shown to reduce protein levels, especially in concentrations of 40 µM [Figure 5b]. This result was consistent with the results of LPS-induced nitrite production as shown in Figure 2d. In contrast, COX-2 protein levels unexpectedly increased with increases in THC-NH2 concentration. However, without the inducement of LPS, the upregulation of COX-2 expression did not occur in RAW264.7 treated with THC-NH2. Therefore, we suspect that THC-NH2 treatment does not induce COX-2 expression but does inhibit COX-2 degradation.
3‑Amino‑3‑deoxytetrahydrocurcumin reduces nitric oxide synthase and cyclooxygenase‑2 mRNA expression via the inhibition of phosphorylated PI3K/Akt pathway – but not othersAlthough THC-NH2 is unable to reduce COX-2 protein expression, it still exhibits repressive effects on inos and cox 2 mRNA levels [Figure 5d and e]. However, several pathways lead to the expression of inos and cox 2, including JNK, p38, ERK, and PI3K/Akt. As illustrated in the results shown in Figure 6a-c, phosphorylation of JNK, p38, and ERK did not decrease after the implementation of THC-NH2. In contrast, relative level of phosphorylation in PI3K and Akt sharply decreased from 0.56 to 0.20 and 0.67 to 0.54, respectively, when compared to their LPS-induced counterparts after the intervention of THC-NH2 in concentrations up to 30 and 40 µM [Figure 6d and e].
3‑Amino‑3‑deoxyte t rahydrocurcumin re tards cyclooxygenase‑2 protein degradation rates but increases neither mRNA levels nor prostaglandin E2 levelsTo further confirm our assumptions regarding THC-NH2, we examined its effects on mRNA expression. As presented in Figure 5d, inos mRNA levels significantly increased when treated with LPS and decreased with the intervention of THC-NH2. These results were congruent with our expectations for the expression of the protein. Surprisingly, mRNA levels of cox 2 did not elevate protein expression after THC-NH2 treatment in LPS-induced RAW264.7. These results only partly verified our assumptions.
To further investigate the properties of THC-NH2, two experiments were designed. Cycloheximide (a protein synthesis inhibitor) was used in concentrations of 1 pg/mL to elucidate the effects of THC-NH2 on COX-2 protein expression. As presented in Figure 6f, COX-2 protein levels gradually degraded in the 24–48-h period. However, COX-2 protein expression increased in the presence of THC-NH2, the effects of which became especially apparent at the 48-h mark. Likewise, results showed that THC-NH2 may retard COX-2 degradation. Thus, researchers hypothesized that if accumulated COX-2 proteins impact PGE2 production, then they will have positive pro-inflammatory effects. It was revealed that PGE2 concentration did not increase significantly along with protein expression, despite the direct relationship between mRNA levels and PGE2 concentration [Figure 6g]. Nevertheless, to
summarize the results above, COX-2 accumulation caused by THC-NH2 seems to pose no negative effects in terms of inflammation at this point in time. However, it is apparent that THC-NH2 can only reduce iNOS, but not COX-2, in the treatment of inflammation.
dIscussIon
The positive effects of curcumin on several diseases, including various cancers, have been well established, and both animal models and human studies have evidenced its safety in high doses.[19] The main reason curcumin is not commonly used as a therapeutic compound, is majorly due to its poor bioavailability, which is curtailed by poor absorption and rapid metabolic rates. Therefore, even though it has exhibited striking capabilities, it can only be classified as a Class II[20,21] or Class IV[22,23] drug in the Biopharmaceutics Classification System as the permeability of curcumin is still controversial. However, because curcumin quickly metabolizes into other substances, its metabolites have been widely studied in the hopes of discovering substituents that are as efficacious and effective as curcumin while maintaining sustainable levels of bioavailability. THC has been identified as one of the natural hepatic and intestinal metabolites of curcumin in both humans and rats.[24] The presence of THC coupled with the absence of curcumin in subcutaneous adipose tissue further substantiates its comparative bioavailability in vivo, and the discovery of THC but not curcumin in the stable plasma indicates its stability rather than curcumin.[1] Although THC exhibits compelling antioxidative capabilities and anticancer effects, the anti-inflammatory potential of THC has not yet been widely researched.[25] Our study revealed that processes occurring in the colon create an aminated product of THC, THC-NH2 that displays inhibitory effects on inflammation in terms of nitrite production and iNOS expression. In contrast to aminated EGCG, which possesses similar bioactive functions to EGCG,[7] THC-NH2 exhibits additional benefits compared to its original counterpart.
In the animal experiment designed in this study, our researchers expected THC-NH2 to display greater ameliorative effects on DSS-induced colitis than THC. However, results displayed no particularly prominent difference in effects between all experimental groups. Thus, some new assumptions have been created to explain the disparity between the results and the expectations of the experiment. First of all, THC, which displayed almost no effect on anti-inflammation in vitro, did exhibit positive effects in vivo in terms of preventing colon shortening and thickening when compared to the DSS-induced group. Upregulation of iNOS expression in both mucosa and surface epithelial cells, concomitant with elevated levels of serum nitrite caused by DSS, has been widely reported in previous studies.[26,27] Notably, mucus thickness was found to be negatively correlated to iNOS levels during the onset of colitis. Therefore, we speculate that the THC had already been metabolized, and the metabolites (which likely
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]

Koh, et al.: Journal of Cancer Research and Practice (2021)
51Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
include THC-NH2) exhibited anti-inflammatory effects on DSS-induced colitis in the early stages of inducement.
According to Ireson et al., who incubated curcumin with microsomes and cytosol from human and rat intestines and livers, THC can metabolize into hexahydrocurcuminol,[28] while Prasad et al. suggested that THC can metabolize into hexahydrocurcumin (HHC) and then hexahydrocurcuminol, which can break down into dihydroferulic acid and ferulic acid.[29] Among these substances, HHC exhibited the greatest inhibitory effects on NO production and COX-2 and p65 expression.[30] HHC has been reported to inhibit COX-2 in previous experiments.[31] It is important to note that, like THC, HHC did not demonstrate significant effects on COX-2 levels in vitro.[32] Comparatively, the cleavage of curcumin metabolites into degradation products is considered to be a minor occurrence.[33] Nevertheless, biodegraded compounds such as ferulic acid are undeniably potential anti-inflammatory compounds, as has been reported in previous research.[34,35] Therefore, although THC may not possess significant anti-inflammatory effects per se, its in vivo metabolites presumably make it a promising compound, as much as or even moreso than THC-NH2. This may explain why the experimental group supplemented with THC showed reversals in the negative effects of DSS as well [Figure 3g and h].
Second, THC-NH2 did not display ameliorative effects on inflammation-related colitis in experimental groups as expected, which could be explained by a few possible factors. Presumably, THC-NH2 may not be as stable as THC in the digestive tracts where degradation and transformation can occur. Furthermore, THC-NH2 might metabolize, therefore, reducing its bioactivity. Third, deamination of THC in the colon could contribute to colitis amelioration. According to Fung et al., high levels of ammonia have been reported to increase inflammatory lesions in rats. In addition, ammonia exposure also facilitates the growth of cancer cells.[36,37] Although these findings are controversial in that they have contradicted the suggestions of previous studies, ammonia production has been reported to significantly increase in patients with IBD.[38] Moreover, some studies have suggested that ammonia could be reduced in circulation, concomitant with increased short chain fatty acids, when supplemented with fermentable fiber in diet.[39] These findings imply that the elimination of ammonia in the colon can contribute to inflammation amelioration. When evaluating the effects of THC and those of THC-NH2, the latter substance demonstrated a diminished ability to eliminate ammonia compared to its original counterpart, which could also explain their disparity in colitis amelioration ability.
Relative to the structure of curcumin, THC comparatively lacks the Michael acceptors necessary to interact with some enzymes, such as HI1 integrase, HIV protease, and glyoxalase I.[40] However, according to Gupta et al., curcumin inhibits the enzyme activity of COX-2 via interaction of a methoxy group with Ala562 through hydrogen bonding and directly bonding to Cal523, Val116, Ala516, and Tyr355.
Furthermore, the inhibitory effects of THC on COX-2 were also observed, indicating that the absence of Michael acceptors may not positively influence COX-2 repression via direct binding or interaction.[41] In terms of COX-2 activity, the substitution of an amine group for a hydroxyl group might not dampen the binding of the methoxy group to proteins because PGE2 production did not increase as COX-2 accumulated. In contrast, our previous findings demonstrated that Se-allylselenocysteine (ASC) contains an amine group that exhibits similar effects on inflammation. ASC inhibits the expression of iNOS but enhances the expression of COX-2. Therefore, amine groups might have a regulative effect on proteins and mRNA levels of COX-2; however, further investigation is needed to determine this for sure.[42] In addition to structural binding, inhibition of proteasome activity might explain COX-2 accumulation. Aggarwal demonstrated curcumin’s inhibitive effects on proteasome activity at high dosages but not at low dosages, which implies that THC-NH2 might have similar effects on proteasome activity, resulting in the inactivation of COX-2 accumulation.[42] Studies have suggested that continuous exposure of COX-2 to arachidonic acid may prevent the degradation of COX-2 via cellular proteasomes but is disposed of by exosomes.[43] Therefore, the limited conversion of arachidonic acid into PGE2 could retard the degradation of COX-2. On the other hand, Shin et al. suggested that Nrf2 ubiquitination can be reduced by curcumin intervention.[44] Therefore, the presence of carbonyl carbons in curcumin, which are also a functional group in THC-NH2, might be a major source of proteasome inhibition. However, as far as the basic requirements for a therapeutic compound, the bioavailability of curcumin still poses a large problem, regardless of its greater biological activities.[45]
One of the major structural differences between THC and THC-NH2 is the number of available carbonyl groups that enable THC to transform. Therefore, THC can also exist in the form of keto–enol tautomers.[46] The second is the substitution of an amine group for a hydroxyl group. It is notable that in this study, THC did not inhibit iNOS expression and NO production as efficaciously as curcumin. However, intriguingly, amination endows THC with the capacity to reduce iNOS. Reduced iNOS expression decreasing NO production as a consequence conforms with the findings of previous studies[47] and might be due to the comparatively less repressive effects of THC on NF-κB compared to curcumin.[48] Moreover, as observed in a study performed by Ma et al., THC-NH2 reduces inflammatory response via PI3K/Akt pathways, but THC does not inhibit JNK pathways.[49,50] Therefore, structural disparities could lead to completely different effects on inflammatory response via different molecular mechanisms. On the other hand, according to Youn et al. (2006), curcumin inhibits the dimerization of receptor activation through Michael addition.[51] Therefore, it is speculated that without the presence of Michael acceptors, amine substituents might act as competitive inhibitors of TLR4 because animated THC had shown significant anti-inflammatory effect as compared to THC and
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]

Koh, et al.: Journal of Cancer Research and Practice (2021)
52 Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
both of them do lack of Michael acceptors. However, there is a lack of information about the effectiveness and safety of THC-NH2 compared to other curcumin metabolites. Therefore, further in-depth investigation of the effects of THC-NH2 on inflammation is needed before determining its efficacy.
conclusIon
Our study reveals that THC-NH2 may be a naturally occurring aminated compound of THC in vivo, even as a minor metabolite found in the colons of mice. It has the potential to overcome the problem of curcumin’s limited bioavailability as well as to address the shortcomings of THC in anti-inflammatory response. Synthetic THC-NH2 exhibited anti-inflammatory effects via PI3K/Akt/iNOS pathways but displayed no effects on COX-2 repression in vitro. Comparatively, both THC and THC-NH2 have shown positive but not exceptional effects on DSS-induced colitis in mice. We speculate that the effects of THC are mainly provided by its metabolites, which may include THC-NH2, while THC-NH2 exerts anti-inflammatory effects on its own. THC-NH2 may not be a particularly effective anti-inflammation drug, due to its diminished inhibition of COX-2. However, to some extent, THC-NH2 could potentially act as a supportive or augmentative component of some anti-inflammation/COX-2-inhibitory drugs, such as valdecoxib and celecoxib, to reduce their side effects and dosages. Nevertheless, further evaluation of THC-NH2 is imperative in both the near and distant futures.
Financial support and sponsorshipThis study was supported by Ministry of Science and Technology, Taiwan [105-2628-B-002-003-MY] and [108-2320-B-002-016-MY].
Conflicts of interestThere are no conflicts of interest.
RefeRences1. Aggarwal BB, Deb L, Prasad S. Curcumin differs from
tetrahydrocurcumin for molecular targets, signaling pathways and cellular responses. Molecules (Basel, Switzerland) 2014;20:185-205.
2. Holder GM, Plummer JL, Ryan AJ. The metabolism and excretion of curcumin (1,7-Bis-(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) in the rat. Xenobiotica 1978;8:761-8.
3. Liu YH, Lee TL, Han CH, Lee YS, Hou WC. Anti-glycation, anti-hemolysis, and ORAC activities of demethylcurcumin and tetrahydroxycurcumin in vitro and reductions of oxidative stress in D-galactose-induced BALB/c mice in vivo. Bot Stud 2019;60:9.
4. Sandur SK, Pandey MK, Sung B, Ahn KS, Murakami A, Sethi G, et al. Curcumin, demethoxycurcumin, bisdemethoxycurcumin, tetrahydrocurcumin and turmerones differentially regulate anti-inflammatory and anti-proliferative responses through a ROS-independent mechanism. Carcinogenesis 2007;28:1765-73.
5. Majeed M, Natarajan S, Pandey A, Bani S, Mundkur L. Subchronic and reproductive/developmental toxicity studies of tetrahydrocurcumin in rats. Toxicol Res 2019;35:65-74.
6. Matoori S, Leroux JC. Recent advances in the treatment of hyperammonemia. Adv Drug Deliv Rev 2015;90:55-68.
7. Zhang S, Zhao Y, Ohland C, Jobin C, Sang S. Microbiota facilitates the formation of the aminated metabolite of green tea polyphenol (-)-epigallocatechin-3-gallate which trap deleterious reactive
endogenous metabolites. Free Radic Biol Med 2019;131:332-44.8. Zhang S, Wang R, Zhao Y, Tareq FS, Sang S. Biotransformation of
myricetin: A novel metabolic pathway to produce aminated products in mice. Mol Nutr Food Res 2019;63:e1900203.
9. Mei Y, Fang C, Ding S, Liu X, Hu J, Xu J, et al. PAP-1 ameliorates DSS-induced colitis with involvement of NLRP3 inflammasome pathway. Int Immunopharmacol 2019;75:105776.
10. Hanić M, Trbojević-Akmačić I, Lauc G. Inflammatory bowel disease Glycomics perspective. Biochimica Biophys Acta 2019;1863:1595-601.
11. Lee BS, Lee C, Yang S, Park EK, Ku SK, Bae JS. Suppressive effects of pelargonidin on lipopolysaccharide-induced inflammatory responses. Chem Biol Interact 2019;302:67-73.
12. Jang J, Kim SM, Yee SM, Kim EM, Lee EH, Choi HR, et al. Daucosterol suppresses dextran sulfate sodium (DSS)-induced colitis in mice. Int Immunopharmacol 2019;72:124-30.
13. Park YH, Kim N, Shim YK, Choi YJ, Nam RH, Choi YJ, et al. Adequate dextran sodium sulfate-induced colitis model in mice and effective outcome measurement method. J Cancer Prev 2015;20:260-7.
14. Cai HA, Huang L, Zheng LJ, Fu K, Wang J, Hu FD, et al. Ginsenoside (Rg-1) promoted the wound closure of diabetic foot ulcer through iNOS elevation via miR-23a/IRF-1 axis. Life Sci 2019;233:116525.
15. Dong B, An L, Yang X, Zhang X, Zhang J, Tuerhong M, et al. Withanolides from Physalis peruviana showing nitric oxide inhibitory effects and affinities with iNOS. Bioorg Chem 2019;87:585-93.
16. El-Ashmawy NE, Khedr NF, El-Bahrawy HA, El-Adawy SA. Roflumilast, type 4 phosphodiesterase inhibitor, attenuates inflammation in rats with ulcerative colitis via down-regulation of iNOS and elevation of cAMP. Int Immunopharmacol 2018;56:36-42.
17. Hu KQ. Cyclooxygenase 2 (COX2)-prostanoid pathway and liver diseases. Prostaglandins Leukot Essent Fatty Acids 2003;69:329-37.
18. Montrose DC, Nakanishi M, Murphy RC, Zarini S, McAleer JP, Vella AT, et al. The role of PGE2 in intestinal inflammation and tumorigenesis. Prostaglandins Other Lipid Mediat 2015;116-117:26-36.
19. Anand P, Kunnumakkara AB, Newman RA, Aggarwal BB. Bioavailability of curcumin: Problems and promises. Mol Pharm 2007;4:807-18.
20. Hu L, Shi Y, Li JH, Gao N, Ji J, Niu F, et al. Enhancement of oral bioavailability of curcumin by a novel solid dispersion system. AAPS PharmSciTech 2015;16:1327-34.
21. Wan S, Sun Y, Qi X, Tan F. Improved bioavailability of poorly water-soluble drug curcumin in cellulose acetate solid dispersion. AAPS PharmSciTech 2012;13:159-66.
22. Wahlang B, Pawar YB, Bansal AK. Identification of permeability-related hurdles in oral delivery of curcumin using the Caco-2 cell model. Eur J Pharm Biopharm 2011;77:275-82.
23. John MK, Xie H, Bell EC, Liang D. Development and pharmacokinetic evaluation of a curcumin co-solvent formulation. Anticancer Res 2013;33:4285-91.
24. Weerawatanakorn M, Hsieh SC, Tsai ML, Lai CS, Wu LM, Badmaev V, et al. Inhibitory effect of tetrahydrocurcumin on dimethylnitrosamine-induced liver fibrosis in rats. J Funct Foods 2014;7:305-13.
25. Pan MH, Lin-Shiau SY, Lin JK. Comparative studies on the suppression of nitric oxide synthase by curcumin and its hydrogenated metabolites through down-regulation of IκB kinase and NFκB activation in macrophages. Biochem Pharmacol 2000;60:1665-76.
26. Zucker SD, Vogel ME, Kindel TL, Smith DL, Idelman G, Avissar U, et al. Bilirubin prevents acute DSS-induced colitis by inhibiting leukocyte infiltration and suppressing upregulation of inducible nitric oxide synthase. Am J Physiol Gastrointestinal Liver Physiol 2015;309:G841-54.
27. Schreiber O, Petersson J, Waldén T, Ahl D, Sandler S, Phillipson M, et al. iNOS-dependent increase in colonic mucus thickness in DSS-colitic rats. PloS One 2013;8:e71843-3.
28. Ireson CR, Jones DJ, Orr S, Coughtrie MW, Boocock DJ, Williams ML, et al. Metabolism of the cancer chemopreventive agent curcumin in human and rat intestine. Cancer Epidemiol Biomarkers Prev 2002;11:105-11.
29. Prasad S, Tyagi AK, Aggarwal BB. Recent developments in delivery,
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]

Koh, et al.: Journal of Cancer Research and Practice (2021)
53Journal of Cancer Research and Practice ¦ Volume 8 ¦ Issue 2 ¦ April-June 2021
bioavailability, absorption and metabolism of curcumin: the golden pigment from golden spice. Cancer Res Treat 2014;46:2-18.
30. Wicha P, Tocharus J, Janyou A, Jittiwat J, Changtam C, Suksamrarn A, et al. Hexahydrocurcumin protects against cerebral ischemia/reperfusion injury, attenuates inflammation, and improves antioxidant defenses in a rat stroke model. PloS One 2017;12:e0189211.
31. Srimuangwong K, Tocharus C, Yoysungnoen Chintana P, Suksamrarn A, Tocharus J. Hexahydrocurcumin enhances inhibitory effect of 5-fluorouracil on HT-29 human colon cancer cells. World J Gastroenterol 2012;18:2383-9.
32. Li F, Nitteranon V, Tang X, Liang J, Zhang G, Parkin KL, et al. In vitro antioxidant and anti-inflammatory activities of 1-dehydro-[6]-gingerdione, 6-shogaol, 6-dehydroshogaol and hexahydrocurcumin. Food Chem 2012;135:332-7.
33. Dei Cas M, Ghidoni R. Dietary curcumin: Correlation between bioavailability and health potential. Nutrients 2019;11: 2147.
34. Zhu H, Liang QH, Xiong XG, Chen J, Wu D, Wang Y, et al. Anti-inflammatory effects of the bioactive compound ferulic acid contained in oldenlandia diffusa on collagen-induced arthritis in rats. Evid Based Complement Alternat Med 2014;2014:573801.
35. Yin ZN, Wu WJ, Sun CZ, Liu HF, Chen WB, Zhan QP, et al. Antioxidant and anti-inflammatory capacity of ferulic acid released from wheat bran by solid-state fermentation of Aspergillus Niger. Biomed Environ Sci 2019;32:11-21.
36. Visek WJ. Diet and cell growth modulation by ammonia. Am J Clin Nutrit 1978;31:S216-20.
37. Fung KY, Ooi CC, Zucker MH, Lockett T, Williams DB, Cosgrove LJ, et al. Colorectal carcinogenesis: A cellular response to sustained risk environment. Int J Molec Sci 2013;14:13525-41.
38. Van Nuenen MH, Venema K, Van Der Woude JC, Kuipers EJ. The metabolic activity of fecal microbiota from healthy individuals and patients with inflammatory bowel disease. Digest Dis Sci 2004;49:485-91.
39. Kurtovic J, Liu Q, Duan ZP, Ha DK, Bengmark S, Riordan SM. Reply. Hepatology (Baltimore, Md) 2005;41:219.
40. Gupta SC, Prasad S, Kim JH, Patchva S, Webb LJ, Priyadarsini IK, et al. Multitargeting by curcumin as revealed by molecular interaction studies. Natl Product Rep 2011;28:1937-55.
41. Zhang ZB, Luo DD, Xie JH, Xian YF, Lai ZQ, Liu YH, et al. Curcumin’s metabolites, Tetrahydrocurcumin and Octahydrocurcumin,
possess superior anti-inflammatory effects in vivo through suppression of TAK1-NF-κB pathway. Front Pharmacol 2018;9:1181.
42. Aggarwal B. Targeting proteasomal pathways by dietary curcumin for cancer prevention and treatment. Curr Med Chem 2013;21:1583-94.
43. Saadi E, Tal S, Barki-Harrington L. Substrate-inactivated cyclooxygenase-2 is disposed of by exosomes through the ER-Golgi pathway. Biochem J 2018;475:3141-51.
44. Shin JW, Chun KS, Kim DH, Kim SJ, Kim SH, Cho NC, et al. Curcumin induces stabilization of Nrf2 protein through Keap1 cysteine modification. Biochem Pharmacol 2020;173:113820.
45. Liu W, Zhang Z, Lin G, Luo D, Chen H, Yang H, et al. Tetrahydrocurcumin is more effective than curcumin in inducing the apoptosis of H22 cells via regulation of a mitochondrial apoptosis pathway in ascites tumor-bearing mice. Food Funct 2017;8:3120-9.
46. Trivedi MK, Panda P, Sethi KK, Gangwar M, Mondal SC, Jana S. Solid and liquid state characterization of tetrahydrocurcumin using XRPD, FT-IR, DSC, TGA, LC-MS, GC-MS, NMR and its biological activities. J Pharm Analysis 2020;10:334-45.
47. Pae HO, Jeong SO, Kim HS, Kim SH, Song YS, Kim SK, et al. Dimethoxycurcumin, a synthetic curcumin analogue with higher metabolic stability, inhibits NO production, inducible NO synthase expression and NF-κB activation in RAW264.7 macrophages activated with LPS. Molecul Nutrit Food Res 2008;52:1082-91.
48. Murakami Y, Ishii H, Takada N, Tanaka S, Machino M, Ito S, et al. Comparative anti-inflammatory activities of curcumin and tetrahydrocurcumin based on the phenolic O-H bond dissociation enthalpy, ionization potential and quantum chemical descriptor. Anticancer Res 2008;28:699-707.
49. Ma F, Liu F, Ding L, You M, Yue H, Zhou Y, et al. Anti-inflammatory effects of curcumin are associated with down regulating microRNA-155 in LPS-treated macrophages and mice. Pharma Biol 2017;55:1263-73.
50. Cianciulli A, Calvello R, Porro C, Trotta T, Salvatore R, Panaro MA. PI3k/Akt signalling pathway plays a crucial role in the anti-inflammatory effects of curcumin in LPS-activated microglia. Int Immunopharmacol 2016;36:282-90.
51. Youn HS, Saitoh SI, Miyake K, Hwang DH. Inhibition of homodimerization of Toll-like receptor 4 by curcumin. Biochem Pharmacol. 2006;72:62-9.
[Downloaded free from http://www.ejcrp.org on Friday, June 4, 2021, IP: 10.232.74.27]





























