Embed Size (px)
Citation preview

Microenvironment and Immunology
A Novel Model of Dormancy for Bone Metastatic BreastCancer Cells
Rebecca Marlow1,2, Gabriella Honeth1, Sara Lombardi1, Massimiliano Cariati3, Sonya Hessey1, AikateriniPipili1, VeronicaMariotti1, Bharath Buchupalli1,2, Katie Foster5, Dominique Bonnet5, AgamemnonGrigoriadis4,Pranela Rameshwar6, Anand Purushotham2,3, Andrew Tutt2,3, and Gabriela Dontu1,3
AbstractMortality of patients with breast cancer is due overwhelmingly to metastatic spread of the disease. Although
dissemination is an early event in breast cancer, extended periods of cancer cell dormancy can result in longlatency of metastasis development. Deciphering the mechanisms underlying cancer cell dormancy and subse-quent growth at themetastatic site would facilitate development of strategies to interfere with these processes. Achallenge in this undertaking has been the lack of models for cancer cell dormancy. We have established novelexperimental systems that model the bone microenvironment of the breast cancer metastatic niche. Thesesystems are based on 3D cocultures of breast cancer cells with cell types predominant in bone marrow. Weidentified conditions in which cancer cells are dormant and conditions in which they proliferate. Dormant cancercells were able to proliferate upon transfer into supportive microenvironment or uponmanipulation of signalingpathways that control dormancy. These experimental systems will be instrumental for metastasis studies,particularly the study of cellular dormancy. Cancer Res; 73(23); 6886–99. �2013 AACR.
IntroductionMortality of patients with breast cancer is largely due to
metastatic spread of the disease. There is compelling evi-dence that dissemination of breast cancer cells at distantsites is an early event, usually occurring before detection ofthe primary tumor (1). Disseminated tumor cells (DTC)survive in a state of cell-cycle arrest for long periods oftime, a phenomenon defined as cellular dormancy (1–3). Incontrast, tumor dormancy is the result of equal rates of cellproliferation and cell death or slow rate of proliferation dueto immune surveillance or poor blood perfusion (3, 4). In asubset of patients with breast cancer, macrometastasesdevelop as late as 25 years after the initial diagnosis, pre-sumably from DTCs or micrometastases. Approximately halfof patients with DTC in bone marrow will never developmacrometastasis (5, 6). There is evidence that various tissues
can harbor dormant DTC, but macrometastases developwith different frequencies in various tissues, depending oncancer type (4, 7). In breast cancer, particularly in theestrogen receptor (ER)–positive subtype, the most frequentsite of metastasis is the bone (5, 6). It has also been proposedthat bone marrow is the origin of secondary cancer-celldissemination to viscera, even when bone macrometastasesare not detectable (8, 9). These clinical characteristics indi-cate that targeting the early steps of extravasation anddissemination may not be as efficacious as targeting DTC(3, 4). Furthermore, they offer opportunity for interventionaimed to prevent metastasis development, provided themechanisms underlying growth at the metastatic site arebetter understood. Despite their significance for progressionand outcome of disease, these aspects of cancer biology arerelatively underinvestigated, mostly due to lack of in vitroexperimental systems and scarcity of in vivo models.
Two hematopoietic stem cell (HSC) niches have beendefined in the bone marrow: the endosteal niche, definedby bone-lining osteoblasts, maintains HSC in quiescence,and the vascular niche, in bone marrow stroma, defined byendothelial, mesenchymal, and reticular cells, sustains pro-liferation and mobilization of HSC (10). Studies of breast andprostate bone metastasis have demonstrated that cancercells compete for occupancy of these HSC niches duringlocalization to the bone marrow (11, 12). Reasoning that thecomplex interactions between cancer cells and the micro-environment are most likely critical for cellular dormancyand proliferation, we established in vitro models of DTCbased on cocultivation of breast cancer cell lines (BCC) withbone marrow cells in a 3D-collagen biomatrix. We identifiedconditions in which BCCs are dormant (inhibitory niche)
Authors' Affiliations: 1Stem Cell Group, 2Breakthrough Breast CancerResearch Unit, 3Research Oncology, King's College London School ofMedicine, Guy's Hospital; 4Department of Craniofacial Development andStem Cell Biology, King's College London, Guy's Hospital; 5CancerResearchUK, LondonResearch Institute, Lincoln's InnFields Laboratories,London, United Kingdom; and 6Rutgers New Jersey Medical School,Newark, New Jersey
Note: Supplementary data for this article are available at Cancer ResearchOnline (http://cancerres.aacrjournals.org/).
Corresponding Authors: Gabriela Dontu and Rebecca Marlow, ResearchOncology, King's College London, Bermondsey Wing, 3rd Floor Guy'sHospital, London SE19RT, United Kingdom. Phone: 44-0207-1885-594;Fax: 44-0207-1889-986; E-mail: [email protected]; and RebeccaMarlow, E-mail: [email protected]
doi: 10.1158/0008-5472.CAN-13-0991
�2013 American Association for Cancer Research.
CancerResearch
Cancer Res; 73(23) December 1, 20136886
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

and conditions in which they proliferate (supportive niche).In this study, we define cellular dormancy as cell-cycle arrestof cancer cells (4). We show that cell-cycle arrest of BCCs inour experimental system is reversible either by changing themicroenvironment or by inhibiting signaling pathways pre-viously shown to have role in dormancy, such as p38 andTGF-b (3, 13). We validated these findings in vivo, in axenotransplantation model in which 3D-biomatrix seededwith BCCs and bone marrow stromal cells were implantedsubcutaneously. Tumors developed only from 3D-biomatrixbearing bone marrow stromal cells that represented thesupportive niche and not from those bearing stromal cellsthat represented the inhibitory niche. Importantly, BCCsrecovered from the inhibitory niche implant retained abilityto proliferate. These experimental systems will be instru-mental for investigating the mechanisms that control dor-mancy of cancer cells, and for developing strategies to targetthese cells or prevent their "re-awakening" during diseaseprogression.
Materials and MethodsCell linesPrimary human bone marrow stromal cells (BMSC) at
second passage and primary human umbilical vein endothelialcells (HUVEC) were obtained from Lonza and maintained asdirected by supplier: BMSC were propagated in Myelocultmedium (STEMCELL Technologies) and HUVEC in EGM-2medium (Lonza). Primary cells were used in experiments up tosix to eight passages. The immortalized human cell lines HS-5and hFOB were obtained from American Type Culture Col-lection (ATCC) and maintained in Dulbecco's Modified EagleMedium (DMEM) and DMEM-F12 supplemented with 10%FBS respectively (Invitrogen). These lines were used in thelaboratory in early passages, no more than 6 months afterreceipt from ATCC.Breast cancer cell lines were grown using the recommended
culture conditions, as described previously (SUM159, SUM149,MDA-MB-231, MDA-MB-435, and BT474; ref. 14). The BCCsMCF7, T47D, ZR75-1, and the cell lines 293T human embryonickidney, U2OS, and MG63 osteosarcoma cell lines were main-tained in DMEM supplemented with 10% FBS. All breast celllineswere authenticated bymeans of short tandem repeat (STR)analysis (PowerPlexW 1.2 System; Promega) as previously des-cribed (16). STRprofileswerematched to theGermanCollectionof Microorganisms and Cell Cultures (DSMZ) database (www.dsmz.com). The MDA-MB-231 organ-specific metastatic clones1833 BoM and 4175 LM2 were kindly provided by Prof. J.Massague, Memorial Sloan-Kettering Cancer Center, New York,NY (15). These lines were maintained in DMEM supplementedwith 10% FBS and used within 6 months of receipt.
Lentiviral infection of cell linesAhighly efficient lentiviral expression system (pLentiLox 3.7;
http://www.med.umich.edu/vcore/) was used to generate E2-Crimson-(Clontech), dsRED-, or GFP-expressing lentivirus. Allcell lines used in cocultures were infected with lentiviruses asdescribed previously (17) to express the fluorescent proteinsE2-Crimson, dsRED, or GFP.
Flow sorting for individual colorsLive cell sorting for E2-Crimsonþ, GFPþ and dsREDþ cells
was done on FACS-Aria II (BD Biosciences) using a 130-mmnozzle. Cell viability was assessed by ViViD dye incorporation(Invitrogen), used as directed by manufacturer. Cells weregated on viability, side and forward scatter in all experiments.
3D coculture3D-biomatrix (GELFOAM; Pfizer; refs. 18–20) was cut into
5-mm diameter circular pieces (3 mm in thickness) using asterile hole-punch and scalpel. 3D-biomatrix pieces wereplaced into ultra-low attachment 96-well plates (CoStar), wetwith Dulbecco's PBS (DPBS; Gibco) for 15 minutes, after whichDPBS was removed. Bone marrow cells were plated in 20-mLvolume into the 3D-biomatix by capillary action in the follow-ing ratios: for supportive niche, 100,000 BMSC cells per well,and for inhibitory niche, 40,000 cells per line (HS-5, hFOB, andHUVEC—120,000 cells in total per well). Media used for all 3Dcocultures were DMEM-F12 supplementedwith 5%FBS. Plateswere incubated at 37�C 5% CO2 for 2 hours to allow incorpo-ration of cells into 3D-biomatrix. Wells were then filled with200 mL of media.
3D cultures were incubated overnight. Bone marrow celllines (BMCL) or BMSCconfluencywas approximately 90%nextday as observed by fluorescence microscopy (Zeiss AxiovertS100 microscope fitted with AxioCam camera and imagesacquired using Axiovsion 4.8).
After 24 hours, BCCs were plated in a similar fashion at a celldensity of 10,000 cells per well in 10-mL volume after removal ofmedium from wells. To account for the heterogeneity of cloneproliferation within each cell line, theminimum cell number ofBCCs without significant differences in clonogenicity betweenreplicates was optimized (10,000 cells per well). There was nosignificant difference in readouts frommore than ten replicatewells of this cell number, which also represented theminimumnumber of cells detected reliably using the fluorescence platereader. 3D cocultures were incubated at 37�C 5% CO2 for 2hours. Wells were then filled with 200 mL of DMEM-F12 5%.
For 3Dmonoculture controls, BCCs were plated at a densityof 100,000 cells per 3D-biomatrix, using the same techniquedescribed earlier for BMCL and BMSC.
Cultures were maintained in mClear white 96-well plates(Greiner) for ease of fluorescent intensity reading after platingBCCs. Every 24 hours, 100 mL of medium in each well wasreplaced with fresh medium. Cell growth was monitored bymicroscopy and followed for 7 to 10 days using the fluorescentplate reader FLUOstar Omega (BMG Labtech Firmware 1.32,Software 1.20). Readings were taken every 24 hours subsequentto plating after changing media in wells. Fluorescent intensitywas read using bottom optic readings with orbital averaging of3 mm, GFP 485/520 nm, dsRED 544/590 nm, E2-Crimson 610/650 nm (excitation/emission nm).
InhibitorsThe following inhibitors were used in the culture medium of
the 3D-biomatrix cocultures: SB431542 used at 10 mmol/L(Tocris), SB203580 used at 2 mmol/L (Tocris), and S1042 (Sell-eck) used at 2-mmol/L final concentration (21–23). Dimethyl
Breast Cancer Metastatic Niche
www.aacrjournals.org Cancer Res; 73(23) December 1, 2013 6887
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

sulfoxide (DMSO; Sigma) was used as a vehicle. DMSO wasused in control experiments at the same dilution factor asinhibitors (1:1,000). Inhibitors were either added into culturemedium at plating of BCCs or after 4 days of culture.
Dissociation of 3D cocultures3D-biomatrix was incubated with Collagenase/Hyaluroni-
dase solution (STEMCELL Technologies) for 5 minutes at 37�C(50 mL/well) until dissociated to single cells. Cells were recov-ered by centrifugation.
Flow sorting for viable cells after dissociationLive cell sorting for GFPþ and dsREDþ cells after dissoci-
ation of 3D cocultures was done on FACS-Aria II (BD Bio-sciences) as described earlier for cell lines. Cells were sorteddirectly into 96-well plates (Greiner; 2,000 cells per well) or intoFACS tubes (BD Biosciences).
Proliferation assayCellTiter-Blue (Promega) was used according to supplier's
instructions. Fluorescent intensity was read using bottomoptic readings with orbital averaging of 5 mm (560/590 nm).
Colony formationDissociated single-cell cultures were plated at clonogenic
density in 6-well dishes (CoStar; 1,000 and 500 cells per well).Cultures were maintained for 14 days in DMEM-F12 5%,changing media every 4 to 5 days. Colonies were fixed informalin for 15 minutes then stained with 0.05% crystal violetfor 15 minutes, rinsed in running water, and left to air-dry.Colony formation was counted by eye.
Flow cytometry on fixed cells for 5-ethynyl-20-deoxyuridine detection
SUM159 dsREDþ cells were incubated with 10 mmol/L EdU(5-ethynyl-20-deoxyuridine)-containing medium for 24 hoursbefore plating into coculture. Alternatively, SUM159 dsREDþ
were plated into 3D cocultures as described earlier and incu-bated with medium containing 10 mmol/L EdU for 24 hoursafter plating. Dissociated single cells from cocultures werefixed with 4% paraformaldehyde (Thermo Scientific) for 10minutes. EdU incorporated into DNA detected using Click-iTEdU kit 647 (Invitrogen) according to manufacturer's instruc-tions. Cells were gated on forward scatter and analyzed fordsRED and EdU (647) labels using LSRFortessa flow analyzer(BD Biosciences).
CytospinFor immunofluorescent analysis, dissociated single-cell
cultures were fixed overnight in ice-cold 70% ethanol andcytospun onto Plus slides (Thermon Shandon) using cytospin(ShandonCytospin3) at 800 rpm for 3minutes. Slideswere thenfixed with 4% paraformaldehyde (Thermo Scientific) for 10minutes, before immunofluorescent staining was carried out.
ImmunofluorescenceAntibodies used are described in the Supplementary Mate-
rials and Methods. Nuclei were counterstained with 40,6-dia-
midino-2-phenylindole (DAPI; Invitrogen). Slides were imagedusing Zeiss Axio Imager A1 with AxioCam MRm and imagesacquired using AxioVision 4.8 software.
Subcutaneous implantation modelAll animal experiments were approved by the Institutional
Committees on Animal Welfare and in compliance with theUnited Kingdom Home Office Animals Scientific ProceduresAct, 1986. Of note, 21-day-old female nonobese diabetic/severecombined immunodeficient (NOD/SCID) mice were obtainedfrom Harlan UK Ltd. Procedures were carried out after 28 daysof age. 3D-biomatrix cocultures were prepared as describedearlier. Following anesthesia, 3D-biomatrices were implantedsubcutaneously into mice using a trocar and single incision inthe back of the neck. Implants were harvested after 1 to 8weeks.
Inhibitor treatment in vivo3D-biomatrices were implanted as earlier. Twenty-four
hours after implantation, mice were injected daily with inhi-bitors by intraperitoneal injection for 6 weeks. DMSOwas usedas a vehicle control and inhibitors were dosed at the followingconcentration: SB431542 10 mg/kg, SB203580 15 mg/kg, andS1042 60 mg/kg. These doses were chosen from literature thatdemonstrates a biologic effect from treatment of differentmouse models with these inhibitors (24–26). 3D-biomatriximplants were harvested for histologic examination after 6weeks of treatment.
Tissue fixation3D-biomatrix scaffoldswere dissected andfixed overnight in
formalin (Solmedia Lab Supplies). The tissue was processedand paraffin-embedded using routine protocols. 3-mmsectionswere cut and stained with hematoxylin and eosin (H&E; Dako).
ImmunohistochemistryStandard immunohistochemistry (IHC) protocols were used
(14). Antibodies used are described in the SupplementaryMaterials andMethods. Antigen retrieval was carried out usingcitric acid buffer pH 6 (Dako). For visualization of nuclei,hematoxylin counterstain was used. Secondary antibodiesconjugated to horseradish peroxidase (HRP; The Jackson Lab-oratory) were used for visualization with 3,30-diaminobenzi-dine (DAB; Dako), according to manufacturer's protocol.
Statistical analysisData were analyzed using software GraphPad Prism v5.0a.
Mann–Whitney, t test, 1-way or 2-way ANOVA statistical testswere used where appropriate. P values less than 0.05 wereconsidered statistically significant.
ResultsCocultivation of breast cancer cells with bone marrowcells in 3D-collagen biomatrix
We established an in vitro system that recapitulates in partthe complexity of the bone metastatic niche in vivo, and isamenable to a variety of experimental manipulations and
Marlow et al.
Cancer Res; 73(23) December 1, 2013 Cancer Research6888
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

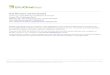
quantitative read-out assays in vitro. 3D-collagen biomatrixarrayed in a 96-well plate format was used as a scaffold forcocultures of BCCs and bonemarrow stromal cells (Fig. 1A andSupplementary Fig. S1A and S1B). Type I collagen, the main
component of this 3D-biomatrix, is also the predominantextracellular matrix (ECM) component in the bone marrow(6). The limited volume of cell culture prevents diffusion ofparacrine factors. The same 3D-biomatrix was used in
A 3D collagen biomatrix
Seed with
BM cells
BMCL GFP+Seed with
BCC
BMCL GFP+
BCC dsRED+Brightfield
B
MC
F7 o
n
BM
CL
MC
F7 o
n
BM
SC
24 h 3 day 6 day
SUM159
****
*** *
******
MDA-MB-231
SUM149
*
***
**
BT474
H BMCL BMSCCell line
SUM159SUM149MCF7MDA-MB-231MDA-MB-453BT474ZR-75-1T47D
–
–
–
–
–
–
–
–
++
+++
+
++
++
+++
+++
++
293TU2OSMG63
+++ +++
++ ++
++ ++
Key– <2 fold increase
+ 2–5-fold increase
++ 5–10-fold increase
+++ >10 fold increase
**
MCF7
D E
GF
24 h
Monitor for
6 days
24 h
Parental 231
1833 BoM
4175 LM2
– ++
++ ++
++–
C
DayF
old
in
cre
ase
Day
Fo
ld in
cre
ase
Day
Fo
ld in
cre
ase
Day
Fold
incre
ase
Day
Fold
incre
ase
Figure 1. Bone marrow (BM) metastatic niche models: primary BMSC supported proliferation of breast cancer cells (supportive niche), whereas a mix ofosteoblasts, mesenchymal, and endothelial cell lines (BMCL) did not support proliferation of breast cancer cells (inhibitory niche). A, diagram of bonemarrowniche model set-up. 3D-biomatrix is seeded with GFPþ BMCL or BMSC when bone marrow cells reach subconfluency; dsREDþ BCCs are seeded inlow numbers. B, representative images of dsREDþ MCF7 cells grown into GFPþ BMCL (bottom) or BMSC (top) over 6 days. Scale, 50 mm. C–G,total fluorescence of BCCs (MCF7, SUM159, MDA-MB-231, SUM149, and BT474) was monitored. BCCs seeded in 3D-biomatrix without stromal cells(3D monoculture, 3D) and BCCs plated in standard 2D conditions (2D) were used as controls. Fluorescence is expressed as the fold-increase from 24 hoursafter seeding (n ¼ 6 experiments, 5 replicates in each experiment). Error bars, SEM. �, P < 0.05; ��, P < 0.01; ���, P < 0.001; 2-way ANOVA withBonferroni post test. H, proliferation rates of cell lines in 3D coculture. Only BCCs were found to be growth-arrested in the inhibitory niche (BMCL). ERþ BCCare indicated in bold. The fetal kidney cell line, 293t, and the osteosarcoma cell lines, U2OS and MG63, proliferate in the BMCL coculture. The subline ofMDA-MB-231 1833 BoM proliferates in BMCL coculture, the parental and the 4175 LM2 lines do not.
Breast Cancer Metastatic Niche
www.aacrjournals.org Cancer Res; 73(23) December 1, 2013 6889
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

physiologically relevant in vitro and in vivo models of osteo-genesis and hematopoiesis (19, 20).
The 3D-biomatrix was seeded either with human primaryBMSCs or immortalized lines representing cell types foundin human bone marrow: osteoblasts (human fetal osteo-blasts, hFOBs), mesenchymal cells of bone marrow origin(HS-5), and endothelial cells (primary HUVEC), in a mix(BMCL). Specific in vitro assays confirmed the differentiationpotential of individual cell lines (Supplementary Fig. S1C andS1D). The arrayed 3D-biomatrix was preseeded with eitherGFP-labeled BMSC or BMCL, expanded to subconfluencyand subsequently seeded with dsRED-labeled BCCs at lowdensity, to model the high stromal cells:cancer cells ratio(20:1) associated with DTC (Fig. 1A). We used four ER-positive BCCs (MCF7, T47D, ZR75-1, and BT474) and fourER-negative BCCs (SUM149, SUM159, MDA-MB-231, andMDA-MB-453). The proliferation of cancer and bone marrowcells in 3D-biomatrix was monitored over 7 to 10 days byquantifying the total fluorescence from the labeled cells.Bone marrow cells (BMSC and BMCL) did not proliferatesignificantly after reaching confluence within 24 hours ofcoculture (Supplementary Fig. S1E–S1J). The sensitivity ofthe read-out assay for BCCs is shown in Supplementary Fig.S2A and S2B. All BCCs proliferated in coculture with BMSCof human or mouse origin (Supplementary Fig. S2C–S2E) orin control 3D monoculture (Supplementary Fig. S2G–S2I).Most ER-positive BCCs proliferated at higher rates than ER-negative BCCs (Fig. 1C–G). None of the BCC lines prolifer-ated in coculture with BMCLs, although other cell lines(293T, fetal kidney, U2OS and MG63, and osteosarcoma)proliferated equally in coculture with BMCLs and BMSCs(Fig. 1H and Supplementary Fig. S2J). We further used BCCsselected specifically for their ability to grow as bone metas-tasis in vivo and compared their proliferation in 3D cocul-ture with BMSC and BMCL. We used sublines of MDA-MB-231 generated by Kang and colleagues in the laboratory ofProf. J. Massague's group that metastasize to the bone (1833BoM) and lung (4175 LM2; ref. 15). We found that althoughin 3D monoculture the metastatic sublines proliferated morethan the parental line, in 3D coculture with BMSC theyproliferated equally (Fig. 1H and Supplementary Figure S2K–S2M). Importantly, the 1833 BoM line was the only BCC linethat proliferated in 3D coculture with BMCL, indicating thatthey possess intrinsic properties that overcome the inhibi-tion exerted by these culture conditions.
The 3D coculture of BCCs with BMCLs will henceforth betermed "inhibitory niche" and the coculturewith BMSCswill betermed "supportive niche." Additional control experimentsassessing the individual stromal cell lines as well as the effectof paracrine factors from cancer cells are shown in Supple-mentary Fig. S3.
Breast cancer cells in the inhibitory niche were inreversible cell-cycle arrest
The lack of proliferation observed in inhibitory niche couldbe due to cell death, equal cell death and cell-proliferationrates, or cell-cycle arrest. To investigate, we recovered singlecells by enzymatic dissociation. The viability of BCCs and bone
marrow cells recovered from inhibitory niche and supportiveniche were similar (>90% viable assessed by flow cytometry,data not shown). We also examined recovered BCCs and bonemarrow cells for apoptosis markers, active-capase-3 andAnnexin V. Apoptotic cancer cells and bone marrow cells wereobserved very infrequently in either coculture (SupplementaryFig. S4A and S4B).
To investigate if BCCs in the inhibitory niche were under-going cell-cycle arrest, we examined their ability to dilute theDNA label EdU, and their colocalization with markers forproliferation (Ki67) and cell-cycle inhibitors (p21 and p27).BCCs were labeled with EdU before seeding into inhibitoryniche or supportive niche coculture. Inhibitory niche andsupportive niche biomatrices were collected at different timepoints after seeding, dissociated to single cells, and EdU levelsin dsRED-positive BCCs were assessed by flow cytometry. TheEdU signal was rapidly lost in BCCs cultured in standard 2Dconditions or in supportive niche coculture (to <5% in EdU-positive cells). In contrast, the EdU label persisted in BCCscocultured in the inhibitory niche (�40%) showing that pro-liferation of BCCswasmarkedly reduced in the inhibitory niche(Fig. 2A). Addition of EdU to cocultures for 24 hours imme-diately after BCC plating did not result in incorporation ofthe label into BCCs when cocultured in the inhibitory niche(Fig. 2B). In contrast, the label was incorporated into BCCscocultured in the supportive niche, 2D, and 3D monoculture(label present for 24 hour immediately after plating) and lostrapidly when culture medium was changed and BCCs prolif-erated (Fig. 2B).
This was further supported by analysis of Ki67 expression inBCCs cocultured in the 3D-biomatrix systems. Ki67 is presentduring all active phases of the cell cycle (27). Single cells wererecovered fromdissociated biomatrices 5 days after BCCswereseeded. Colocalization of Ki67 and dsRED was assessed oncytospins by immunofluorescence. The frequency of Ki67-positive BCCs was more than 75% in the supportive nicheculture, but was significantly lower (<30%) in BCCs recoveredfrom the inhibitory niche culture (Fig. 2C–G).
To test if BCCs in the inhibitory niche coculture wereundergoing active cell-cycle arrest, we examined the expres-sion of cyclin-dependent kinase inhibitor proteins, p27 andp21, which indicate a block in cell-cycle progression (28).Single-cell immunofluorescence analysis indicated that dou-ble-positive p27/dsRED BCCs, or p21/dsRED BCCs wereobserved in cells recovered from the inhibitory niche coculture(>60%) and were absent in cells recovered from the supportiveniche coculture (Fig. 2H–M). Therefore, we conclude that BCCsare undergoing an active cell-cycle arrest when cocultured inthe inhibitory niche model.
Dormant breast cancer cells were capable to proliferatewhen removed from the inhibitory niche
By definition, dormant cells, unlike terminally differentiatedor senescent cells, are capable of reentering the cell cycle whenconditions become favorable. To test if BCCs cocultured in theinhibitory niche model were bona fide dormant cells, weinvestigated their ability to proliferate after removal frominhibitory niche. We dissociated cocultures after 5 days and
Marlow et al.
Cancer Res; 73(23) December 1, 2013 Cancer Research6890
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

recovered single-cell suspensions from which we sorteddsRED-positive BCCs by flow cytometry (Fig. 3A). dsRED-positive BCCswere subsequently plated in standard 2D culture
and fluorescence was monitored over 7 days. We observed nodifference in proliferation between BCCs recovered frominhibitory niche versus supportive niche by either total
J
M
An
ti-p
21/
an
ti-d
sR
ED
SNIN
SUM159
An
ti-p
27/
an
ti-d
sR
ED
IN SN
SUM159
G
SNIN
An
ti-K
i67
/
an
ti-d
sR
ED
MCF7
An
ti-K
i67
/
an
ti-d
sR
ED
IN
An
ti-K
i67
/
an
ti-d
sR
ED
SN
SUM159F
D E
**
MCF7
***
SUM159***
MDA-MB-231C
% K
i67
+/d
sR
ED
+ B
CC
% K
i67
+/d
sR
ED
+ B
CC
% K
i67
+/d
sR
ED
+ B
CC
H
K
I
L
SUM159
***
SUM159**
MCF7
**
***
MCF7
% p
27
+/d
sR
ED
+ B
CC
% p
27
+/d
sR
ED
+ B
CC
% p
21
+/d
sR
ED
+ B
CC
% p
21
+/d
sR
ED
+ B
CC
A B
0 h
24 h
120
h
EdU la
bel
BCC plated
in 3D culture
*** *** ***
% d
sR
ED
+ B
CC
EdU
+
0 h
24 h
96 h
EdU la
belBCC plated
in 3D culture
% d
sR
ED
+ B
CC
EdU
+
*****
***
Figure 2. The inhibitory niche induced cell-cycle arrest in breast cancer cells. A, EdU persistence wasmonitored by fluorescence-activated cell sorting (FACS)analysis. Graph represents the percent dsREDþ SUM159 cells that were EdUþ (n ¼ 2, 3 replicates in each experiment). Error bars, SEM. ���, P < 0.001;2-way ANOVA with Bonferroni post test. B, EdU incorporation following plating into 3D culture was monitored by FACS analysis. Graph represents thepercent dsREDþSUM159cells thatwereEdUþ (n¼2, 3 replicates in eachexperiment). Error bars,SEM. ���,P<0.001; 2-wayANOVAwithBonferroni post test.C–E, quantification of Ki67 anddsRED immunofluorescence (n¼ 3). As control, BCCs grown in 3D-biomatix alone (3D), without stromal cells, were used. Errorbars, SEM; if no bar is present the value is 0. ��, P < 0.01; ���, P < 0.001; 1-way ANOVA. F and G, five-day 3D-biomatrix cocultures were dissociatedto single cells, fixed, and cytospun onto slides. Representative images of immunofluorescence staining for anti-Ki67 (red) and anti-dsRED (green) are shown.Scale, 50 mm. Arrows, double-positive Ki67þ dsREDþ BCCs. Arrowheads, single-positive cancer cells (dsREDþ). H and I, quantification of p27 anddsRED immunofluorescence (n ¼ 3). Error bars, SEM; if no bar is present the value is 0. ��, P < 0.01; ���, P < 0.001; 1-way ANOVA. J, five-day 3D-biomatrixcocultures were dissociated to single cells, fixed, and cytospun onto slides. Representative images of immunofluorescence staining for anti-p27 (red),along with anti-dsRED (green) are shown. Scale, 50 mm. Arrows, double-positive p27þ/dsREDþ BCCs. Arrowheads, single-positive cancer cells (dsREDþ).K and L, quantification of p21 and dsRED immunofluorescence (n ¼ 3). Error bars, SEM. ��, P < 0.01; 1-way ANOVA. M, representative images ofimmunofluorescence staining for anti-p21 (red), along with anti-dsRED (green) are shown. Scale, 50 mm. Arrows, double-positive p21þ/dsREDþ BCCs.Arrowheads, single-positive BCCs (dsREDþ). IN, inhibitory niche; SN, supportive niche.
Breast Cancer Metastatic Niche
www.aacrjournals.org Cancer Res; 73(23) December 1, 2013 6891
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

fluorescence or in cell viability assay (Fig. 3B–E and Supple-mentary Fig. S4C–S4E). Colony formation assay (Fig. 3F)showed no difference in clonogenicity of BCCs recovered frominhibitory niche coculture compared with those recoveredfrom the supportive niche (Fig. 3G–I and Supplementary Fig.S4F–S4H). Taken together, these results indicate that cancercells in the inhibitory niche coculture were in a state ofreversible cell-cycle arrest, or bona fide dormant state.
Moreover, BCCs cocultured in the inhibitory niche wereable to proliferate when removed and placed into thesupportive niche coculture (Fig. 4A–C). Conversely, BCCproliferation was halted when removed from the supportiveniche coculture and seeded into the inhibitory niche (Fig.4A–C). BCCs seeded in 3D-biomatrix without stromal cellsand BCCs plated in standard 2D conditions were used ascontrols and these displayed the same behavior: cells wentinto cell-cycle arrest when transferred into the inhibitoryniche, and proliferated when transferred into the supportiveniche (Fig. 4A–C). These results indicate that cell-cyclearrest of BCCs in the inhibitory niche coculture and theirproliferation in supportive niche coculture are induced by
the microenvironment, and not by changing the cancer cells'intrinsic properties. Further supporting this conclusion,conditioned medium from the inhibitory niche coculturesignificantly decreased proliferation of BCCs in all cultureconditions tested: standard 2D monolayer, 3D monoculture,and 3D supportive niche coculture (Fig. 4D–E and Supple-mentary Fig. S5A–S5I). Conditioned medium from support-ive niche did not overcome the inhibitory effect of theinhibitory niche microenvironment (Fig. 4E and Supplemen-tary Fig. S5H and S5I). This latter observation indicates thatproliferation of BCCs in the inhibitory niche coculture isactively suppressed through secreted components, ratherthan being due to lack of factors needed for proliferation.Because the supportive niche and inhibitory niche cocul-tures have different cellular components and are not directlycomparable, we added each of the lines representing theinhibitory niche (hFOB, HS-5, or HUVEC cells) to the sup-portive niche and assessed the effect on BCCs proliferation.We found that hFOB and HS-5 cells, but not HUVEC, had aninhibitory effect on proliferation of BCCs (SupplementaryFig. S5J and S5K).
A
B C D
NS
FReplate into 2D
at clonogenic density
NS
G H I
NS
MCF7
NS
MDA-MB-231
NS
BT474
ETiter Blue 24 h Titer Blue 4 d
Flow sorting for
dsRED+ BCC
Replate into
2D culture
3D coculture
Dissociation to single cells
3D coculture
Dissociation to single cells
** **
3D coculture before sortingSUM159
2D culture after sorting
NS
DayDay
SUM159
3 days
6 days
Figure 3. Dormant breast cancer cells were capable of proliferating when removed from the inhibitory niche. A, schematic representation of flow sorting fordsREDþ BCCs and replating into standard 2D conditions. 3D monoculture of BCCs (3D) was used as control. B, fluorescence monitored before flowsorting (n ¼ 2; 40 replicate wells per condition). Error bars, SEM; ��, P < 0.01; 1-way ANOVA. C, fluorescence monitored after sorting (n ¼ 2; 10 replicatewells per condition). Error bars, SEM; NS, not significant; 1-way ANOVA. D and E, Titer Blue proliferation assay at 24 hours and 4 days after platinginto standard tissue culture (n¼ 2; 10 replicatewells per condition). Error bars, SEM; NS, not significant; 1-way ANOVA. F, schematic representation of colonyformation assay. G–I, quantification of colony formation (n ¼ 4; 6 wells plated per experiment). Error bars, SEM. NS, not significant Mann–Whitney test,inhibitory niche (IN) versus supportive niche (SN).
Marlow et al.
Cancer Res; 73(23) December 1, 2013 Cancer Research6892
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

Cell-cycle arrest of breast cancer cells in the inhibitoryniche was prevented by inhibition of specific signalingpathwaysTo generate proof-of-concept that these models can be used
to identify the molecular mechanisms that control breastcancer cell dormancy, we tested if inhibiting pathways previ-ously associated with cellular dormancy can reverse cell-cyclearrest in the inhibitory niche coculture. We used the small-molecule inhibitors SB203580 targeting p38 MAPK, SB431542targeting Alk5, the type-I TGF-b receptor, and SU11248(Sutent) targeting receptor tyrosine kinases (RTK; SU11248targets include VEGFR2, PDGFRb, c-Kit, and FLT3). p38MAPKsignaling has been implicated in cellular dormancy in modelsusing melanoma and head and neck cancer cell lines (29–31).TGF-b has dual properties and can induce both proliferationand dormancy in disseminated cells (13, 32). Specific inhibitionof RTK, Alk5, and p38 signaling in growth-arrested BCCs in theinhibitory niche induced their proliferation, while not inducingchanges in the stromal cells of inhibitory niche or supportiveniche (Fig. 5A–C and Supplementary Fig. S6A–S6E). Impor-tantly, an opposite effect was seen in BCC 2D monocultures(Fig. 5C and Supplementary Fig. S6C and S6E), underscoringthe importance of microenvironmental and 3D configuration.To confirm that the increase in fluorescence observed didresult in proliferation of BCCs, we examined dissociated singlecells from 3D cocultures with immunofluorescence for Ki67.Inhibitory niche cocultures treated with inhibitors had signif-icantly more Ki67-positive cells than control-treated (Fig. 5Dand E). Addition of Alk5, p38, and RTK inhibitors to coculturesat day 4 after BCC plating in inhibitory niche culture had no
effect on cell proliferation, indicating that activation of thesepathways prevent cells from entering quiescence and addi-tional factors contribute to quiescence maintenance (Supple-mentary Fig. S6F–S6H). Treatment with small-molecule inhi-bitors against additional RTK (ABT-869 and MGCD-265)completely blocked cell proliferation and/or induced cell deathin all the culture systems described earlier (data not shown).These results indicate that the microenvironment influencesthe cancer-cell response to molecular signals and differentculture conditions favor signaling that compensates or ispermissive for the inhibitory effect of drugs.
In vivo validation of the inhibitory and supportive nichemodels
To assess if the effects of inhibitory niche and supportiveniche coculture on BCC proliferation are maintained in vivoover longer periods of time and in the presence of systemicfactors, we implanted the 3D-biomatrix subcutaneously intoNOD/SCID mice. We first assessed if integrity of the 3D-biomatrix is maintained in vivo. 3D-biomatrix seeded withbone marrow cells only (BMCL or BMSC) maintained itsintegrity for up to 6 weeks, became vascularized, and GFP-labeled bone marrow cells were retained in the structures (Fig.6A–D). Subsequently, we assessed proliferation of BCCs inbiomatrix implanted subcutaneously. 3D-biomatrices werepreseeded with bone marrow cells to generate the inhibitoryniche and supportive niche and then seeded with low numbersof MDA-MB-231 cells. After a 24-hour incubation in vitro, theinhibitory niche and supportive niche biomatrices wereimplanted contralaterally into NOD/SCID mice (Fig. 6E). After
A B C
**
3D coculture before sorting
]*
3D coculture after sorting
SN
IN
SUM159 dsRED+
Flow sorting for
dsRED+ SUM159 cells
SN
IN
D E
] NS
***
MDA-MB-231
3D coculture with conditioned medium
SNIN
Conditioned medium
Control Control
Day
Fold
incre
ase
Day
Fold
incre
ase
Day
Fold
in
cre
ase
Figure 4. Paracrine signals induced cell-cycle arrest in breast cancer cells. A, schematic representation of flow sorting and replating into alternative 3Dcoculture. Standard 2D and 3D monoculture of BCCs (3D) were used as controls. B, fluorescence monitored before flow sorting (n ¼ 2; 60 replicatewells per condition). Error bars, SEM. ��,P < 0.01; 1-wayANOVA.C, fluorescencemonitored after replating into 3Dcoculture (n¼ 2; 5 replicates per condition).Error bars, SEM; �, P < 0.05; 2-way ANOVA with Bonferroni post test. D, schematic representation of conditioned media swap in 3D cocultures. Conditionedmedium was replaced every 24 hours. E, fluorescence of BCCs monitored after conditioned media swap (n ¼ 3; 5 replicates per condition). Error bars,SEM. �, P < 0.05; ��, P < 0.01; ���, P < 0.001 1-way ANOVA. IN, inhibitory niche; SN, supportive niche.
Breast Cancer Metastatic Niche
www.aacrjournals.org Cancer Res; 73(23) December 1, 2013 6893
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

8 weeks, tumors formed at the site of implantation of sup-portive niche, but not at the site of inhibitory niche biomatrices(6 mice total implanted in two experiments; tumors formed at5/6 supportive niche implantation sites, and 0/6 at inhibitoryniche implantation sites; Fig. 6F). IHC confirmed that thetumors were formed by dsRED-positive MDA-MB-231 cells(Fig. 6G). Rare dsRED-positive cancer cells were observed ininhibitory niche implants, at a density comparable with theoriginal seeding density (Fig. 6G). To test if these were indeeddormant MDA-MB-231 cells, we dissected and dissociatedbiomatrices to recover single cells (Fig. 6H). We plated all cellsrecovered (including BCCs, stromal cells, and cells of mouseorigin) at low density in 2D culture and analyzed the coloniesgenerated. MDA-MB-231 colonies were observed in cultures ofcells recovered from both inhibitory niche and supportiveniche biomatrices (Fig. 6I). These colonies had the similarmorphology and size; however, as expected, much fewercolonies overall were formed by BCCs recovered from theinhibitory niche as these BCCs had not proliferated in vivo.These results demonstrate that the rare dsRED-positive cellsobserved by IHC in inhibitory niche biomatrices in vivo wereindeed bona fide dormant MDA-MB-231 cells and are capableof proliferating in favorable conditions. To test if dormancycould be prevented in vivo by interfering with signaling involv-ing RTK, p38, Alk5, we implanted NOD/SCID mice with inhib-itory and supportive niche (supportive niche) 3D-biomatrixcocultures, as described earlier. Mice were injected daily with
inhibitors by intraperitoneal injection, starting 24 hours afterimplantation, for 6 weeks. DMSOwas used as a vehicle control.Palpable masses were observed in all supportive niche implan-tation sites, and in the majority of mice implanted withinhibitory niche and treated with inhibitors, but not in theimplantation sites of inhibitory niche in control animals,treated with vehicle only. Histologic analysis of sectionsthrough biomatrices collected and embedded in paraffin con-firmed that cancer-cell proliferation occurred in 44% (4/9) ofinhibitory niche implants upon treatment with SB431542, 67%(6/9) of inhibitory niche implants upon treatment withSB203580 and 56% (5/9) of inhibitory niche implants upontreatment with SU11248 (Fig. 7). The extent of BCC growth wascomparable with that observed in the supportive nicheimplants from control DMSO vehicle treatment and thatobserved in supportive niche implants from inhibitor treat-ment. Taken together, these results demonstrate that BCCs canbe manipulated using our experimental systems to be undermicroenvironmentally induced cell-cycle arrest (inhibitoryniche) or to be proliferating (supportive niche) in coculturewith bone marrow cell types and this quiescence is reversible,both by inhibiting cellular signaling pathways and altering themicroenvironment.
DiscussionSignificant progress has been made in elucidating the
molecular machinery of the multi-step metastasis cascade.
E
A CBBT474 - SU11248 (RTK)
*
SUM159 - SB203580 (p38)
*
*
MCF7 - SB431542 (Alk5)
*
D DMSO SB431542
SU11248 SB203580
Anti-K
i67/
anti-d
sR
ED
Anti-K
i67/
anti-d
sR
ED
***
MDA-MB-231
Day
Fold
in
cre
ase
Day
Fold
in
cre
ase
Day
Fold
in
cre
ase
Figure 5. Cell-cycle arrest in breast cancer cells was reversible upon treatmentwith small-molecule inhibitors against RTK, Alk5, and p38. A–C, fluorescence ofBCCs monitored after 3D coculture in the inhibitory niche (IN) after incubation with small-molecule inhibitors S1042 (RTK), SB431542 (Alk5), and SB203580(p38; n ¼ 6; 5 replicates per condition per experiment) immediately upon plating into 3D coculture. 2D monoculture (2D) is used as a control. Errorbars, SEM. �, P < 0.05 paired t test. D, five-day 3D-biomatrix cocultures of SUM159 cocultured in IN were dissociated to single cells, fixed, and cytospunonto slides. Representative images of immunofluorescence staining for anti-Ki67 (red) and anti-dsRED (green) are shown. Scale, 50 mm. Arrows,double-positive Ki67þ dsREDþ cancer cells. Arrowheads, single-positive cancer cells (dsREDþ). E, quantification of Ki67 and dsRED immunofluorescence(n ¼ 2). Error bars, SEM. ���, P < 0.001; 1-way ANOVA.
Marlow et al.
Cancer Res; 73(23) December 1, 2013 Cancer Research6894
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

However, the mechanisms underlying metastatic cell dor-mancy and relapse from dormancy remains one of the mostchallenging, but also most clinically relevant questions(3, 8, 13). Approximately one third of patients with breastcancer have detectable DTCs in bone marrow in early stagesof disease (1). Only half of these develop metastasis, some-times decades after diagnosis of primary disease. This sug-gests that microenvironmental and/or systemic changes arecritical for the "re-awakening" of dormant DTCs or micro-metastases. Elucidating the molecular framework of theseinteractions is critical for developing strategies to interfere
with the process and for predicting in which patients DTCswill progress to macrometastases. The biggest obstacle inthis endeavor is represented by the scarcity of experimentalsystems appropriate for study of DTCs, in particular in vitrosystems.
Several types of experimental models have been used so farto study interactions between metastatic cancer cells and themicroenvironment (33): knockout or transgenic mice, ortho-tropic, intracardiac, or intravenous xenotransplantation mod-els using metastatic cancer cell lines, and subcutaneous xeno-transplantation models using bioengineered implants or
BA
G
Anti-G
FP
FESubcutaneous implantation model
IN SNMDA-MB-231 MDA-MB-231
IN SNH
&E
8 w
eeks in
viv
o
H
7 days in vivo
C D
Anti-d
sR
ED
IN SN
Dissection and dissociation of biomatrix from in vivo
Recovered cells replate into 2D
at clonogenic density
I
IN
H&
E
IN
Anti-P
EC
AM SNSN
MDA-MB-231 coloniesIN SN
IN SN
80
60
40
20
0No.
of co
lonie
s/
10,0
00 c
ells
pla
ted
Figure 6. Breast cancer cells were dormant in inhibitory niche (IN) and proliferated in supportive niche (SN) in vivo. A, representative images of 3D-biomatrix(seeded with BMCL) subcutaneously implanted in vivo for 7 days (brightfield and fluorescent imaging). Scale, 0.25 mm. B, representative image ofanti-GFP IHC on 3-mm section from 3D-biomatrix (BMCL). Scale, 50 mm. White arrow, GFPþ BMCL. C, representative images of vascularization of theimplanted 3D-biomatrix, IN and SN. Brightfield images indicate blood vessel growth into 3D-biomatrix of both IN and SN. D, representative images ofH&E and Anti-PECAM staining on 3-mm section from 3D-biomatrices. Scale, 50 mm. Arrows, blood vessels penetrating the 3D-biomatrix of IN andSN implants (inset, magnification, �5). E, subcutaneous implantation model of implantation of 3D-biomatrix. IN and SN 3D-biomatrices were implantedcontralaterally. F, representative images of 3D-biomatrix implanted subcutaneously in vivo for 8 weeks. Scale, 0.25 mm. Tumors formed in n ¼ 5 outof 6 animals implanted with SNþMDA-MB-231. Representative H&E images from each 3D-biomatrix. Scale, 50 mm. Arrowheads, 3D-biomatrix. G,representative images of anti-dsRED IHC on 3-mm section from 3D-biomatrix seeded with BMCL or BMSC. Scale, 50 mm. Arrows, dsREDþ cells(inset, magnification,�5). H, dissociation of cells from 3D-biomatrix implanted in vivo and replating into 2D culture in vitro. I, representative images of crystalviolet–stained MDA-MB-231 colonies shown and quantification of colony formation. Scale, 50 mm. MDA-MB-231 cells recovered from IN dissociationwere able to proliferate in 2Dculture. Total number of cells recovered from INdissociation generated fewer cancer-cell colonies, asmajority of these cells werestromal cells. However, the colonies formed by BCCs have the same morphology and size as those formed by BCCs from SN dissociations.
Breast Cancer Metastatic Niche
www.aacrjournals.org Cancer Res; 73(23) December 1, 2013 6895
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

DBCC growth in IN implanted in vivo
(total IN implanted)
DMSO SB431542 SB203580 SU11248
0 (3) 4 (9) 6 (9) 5 (9)
Control (DMSO) SN
AControl (DMSO) IN
SU11248 IN
C
B
BCC
Biomatrix
Stromal cells
Key
BCC growth in SN implanted in vivo
(total SN implanted)
DMSO SB431542 SB203580 SU11248
3 (3) 3 (3) 3 (3) 3 (3)
E
Figure 7. Treatment with small-molecule inhibitors induced growth of BCCs in the subcutaneous implantation model. A and B, representative images of H&Esections of inhibitory niche (IN) and supportive niche (SN) implants from vehicle treated (DMSO) animals. Scale bar, 50 mm. C, representative imagesof H&E sections of IN implants from animals treated with the small-molecule inhibitor S1042. Dashed line, area of BCC growth. Scale bar, 50 mm.D, table representing the number of IN subcutaneously implanted in vivo and the number of implants with BCC growth observed by histologic examination ofexcised implants 6 weeks following daily dosing with small-molecule inhibitors, SB431542 (Alk5), SB203580 (p38), and S1042 (RTK). None (0/3) of thecontrol implants hadevidenceofBCCgrowth. 44% (4/9) of IN implants hadBCCgrowth following treatmentwithSB431542, 67% (6/9) of IN implants hadBCCgrowth following treatment with SB 203580 and 56% (5/9) of IN implants had BCC growth following treatment with S1042. The extent of BCC growth wascomparable with that observed in the SN implants from control DMSO vehicle treatment and that observed in SN implants from inhibitor treatment.In all images, arrows indicate BCCand arrowheads indicate stromal cells and dashed line delineates areas of BCCgrowth. E, table representing the number ofSN subcutaneously implanted in vivo and the number of implants with BCCgrowth observed by histologic examination of excised implants 6weeks followingdaily dosing with small-molecule inhibitors, SB431542 (Alk5), SB203580 (p38), and S1042 (RTK). All SN implants had comparable areas of BCC growth.
Marlow et al.
Cancer Res; 73(23) December 1, 2013 Cancer Research6896
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

human bone fragments inserted subcutaneously. Manipulat-ing the osteoclast/osteoblast balance in the bone marrowenvironment dramatically changes metastatic tumor burdenin several transgenic mouse models (6). In models of lungmetastasis, ECM components such as periostin (34), tenascin-C (35), and the BMP inhibitor, Coco (36), have been shown toinduce a metastatic niche conducive to proliferation of breastcancer cells. Manipulation of human cancer cell lines xeno-transplanted in chick chorioallantoic membranes and immu-nodeficient mice identified several signalingmolecules with keyroles in inducing or maintaining dormancy of cancer cells. Suchexamples are uPAR,GAS6, p38a/b, and the balancebetweenp38and ERK, Kiss1, BMP7, and TGF-b (28–32, 37, 38). A number ofmodels aimed at recreating the human bone marrow environ-ment in vitro and in vivo using subcutaneous xenotransplanta-tion of bioengineered materials for osteogenesis (39–42) orhematopoiesis studies (19, 20, 43).Models of thebonemetastaticniche have used silk scaffolds seeded with human BMSC andhuman bone fragments implanted subcutaneously (44, 45).Finally in vitro models used 2D coculture of cancer cells withfibroblasts, osteoclasts or mesenchymal stem cells, and osteo-blasts (46, 47). These in vitro and in vivo studies have demon-strated the importance of the microenvironment in controllingcancer cell growth at the dissemination site (46, 47).Several conclusions emerge when reviewing these models of
the metastatic niche. A single model cannot recapitulate thecomplexity of interactions at themetastatic site. In vivomodelshave the advantage of increased complexity and significance,but are more appropriate for hypothesis-driven studies. The invitromodels lack complexity, are oversimplified, and capture alimited number of interactions between a few cellular compo-nents at the metastatic site (coculture systems) or betweencancer cells and the ECM (mineralized bone, scaffold treatedwith BMP), but they are amenable to exploratory screeningstudies. A significant obstacle is the absence of in vitromodelsfor cellular dormancy. Toourknowledge, few suchmodels exist.One such model is based on culture of MCF7 cells in recom-binant basement membrane (48, 49). Until recently, there wereno in vitro experimental systems that specifically modeledDTCs: a small number of isolated cancer cells in a microenvi-ronment occupied predominantly by normal cells of a differenttissue type. A compelling study from Ghajar and colleaguesused in vitro organotypic models based on coculture of endo-thelial cells, mesenchymal cells of lung and bonemarrow originto investigate dormancy of DTCs (50) and identify the perivas-cular niche as the regulator of cancer cell dormancy.We used a similar approach in developing novel 3D exper-
imental systems that specifically model DTCs in the bonemarrow environment and found that interactions with osteo-balsts and mesenchymal bone marrow cells (HS5) promotecancer cell dormancy. We developed experimental systemsconducive to either proliferation or dormancy of cancer cells.We demonstrated that eight BCC lines remained quiescent inthe inhibitory environment (inhibitory niche) but proliferate inthe supportive one (supportive niche). ER-positive cell linesproliferated at higher rate than ER-negative cell lines, possiblyreflecting properties that account for the higher frequency ofbone metastasis in ER-positive compared with patients with
ER-negative breast cancer. The eight BCC lines were capable toproliferate in bone marrow in vivo, in an intratibial implanta-tionmodel; ER-positive cell lines had a higher rate of success inour hands than ER-negative cell lines (Supplementary Fig. S8).These characteristics are reflected by cultivation in the sup-portive niche system. However, ER-positive BCCs did not gen-erate metastases in the intracardiac injection model of metas-tasis,whereas ER-negativeBCCsdid, similar topreviousfindingsfrom us and others (Supplementary Fig. S8; refs. 14, 51). Theseobservations underscore the necessity of using a combination ofdifferent experimental systems for studying the metastaticprocess. The lack of cell proliferation in the inhibitory nichewas specific for BCCs and involves paracrine signals fromstromal cells. The only BCCs tested that proliferated in theinhibitory niche culture were the 1833 BoMMDA-MB-231 cells,selected for their ability tometastasize to the bone. All the otherBCCs undergo cell-cycle arrest in inhibitory niche coculture, butare able to proliferate when removed from the inhibitory niche.On the basis of all these findings, we argue that the experimentalsystems we developed model the supportive or inhibitory effectofmicroenvironment on BCC proliferation, while also reflectingintrinsic properties of the cancer cells.
As proof-of-concept that these models could be used todissect the cellular and molecular mechanisms underlyingproliferation or dormancy in the bone marrow, we were ableto manipulate and monitor the different cellular componentsof the coculture. Moreover, inhibitors of pathways known tosupport cell dormancy induced cancer-cell proliferation in theinhibitory niche. Importantly, we showed that treatment withthese small-molecule inhibitors elicited a different responsefrom cancer cells in these complex in vitro systems comparedwith classic 2D tissue culture or cancer-cellmonoculture in 3D-biomatrix.We validated these results in vivo, in a subcutaneousimplantation model, in which cancer cells implanted in sup-portive niche generated tumors, whereas those implanted ininhibitory niche failed to do so. Isolated dormant cancer cellswere present in the inhibitory niche implanted in vivo, asdemonstrated by their ability to proliferate when removedfrom this environment. Relevantly, the inhibitors that pre-vented cell-cycle arrest in vitro had the same effect in vivo.
The models we established facilitate studies of dynamicinteractions between cancer cells and several cell types presentin bonemarrow stroma. As with any experimental system, theyhave limitations: the lack of cell components such as osteo-cytes, osteoclasts, immune cells, neural cells,mineralized bone,and ECMcomponents.We aimed to reproduce in vitro the highratio between bone marrow cells and DTC observed inpatients. We also aimed to make these systems amenable toa variety of read-out assays, such as microscopy, total fluo-rescence reading, IHC of sectioned biomatrix, flow cytometryand immunofluorescence analysis of cells recovered from 3D-biomatrix. Different cell components of the coculture can bemanipulated individually or recovered as live cells for furtheranalysis. These assays are quantitative and can be adapted forsmall- or medium-throughput screening studies using RNAinterference (RNAi), antibody or small-molecule inhibitors,therefore permitting an exploratory approach in the study ofDTC and dormancy. Importantly, this is novel in vitromodel of
Breast Cancer Metastatic Niche
www.aacrjournals.org Cancer Res; 73(23) December 1, 2013 6897
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

DTC that induces dormancy of cancer cells in a bonemarrow–like environment. We envision that these models could facil-itate exploratory screening studies to identify the cellularfactors that control cancer-cell dormancy in the bone andmay reveal biomarkers indicative of those DTCs with adverseprognostic impact. This may produce potential targets fortherapy and therapeutics in both tumor cells and the cellularcomponents of the bone marrow niche.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: R. Marlow, A. Purushotham, A. Tutt, G. DontuDevelopment of methodology: R. Marlow, G. Honeth, M. Cariati, D. Bonnet,P. Rameshwar, G. DontuAcquisition of data (provided animals, acquired and managed patients,provided facilities, etc.):M. Cariati, S. Hessey, A. Pipili, B. Buchupalli, K. Foster,A. Grigoriadis, A. Purushotham, A. Tutt
Analysis and interpretation of data (e.g., statistical analysis, biostatistics,computational analysis): R. Marlow, G. DontuWriting, review, and/or revision of themanuscript: R.Marlow, S. Lombardi,M. Cariati, D. Bonnet, A. Grigoriadis, P. Rameshwar, A. Purushotham, A. Tutt,G. DontuAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): S. Lombardi, V. Mariotti, B. BuchupalliStudy supervision: R. Marlow, G. Dontu
AcknowledgmentsThe authors thank the staff at the flow core facilities and breast tissue bank at
King's College London.
Grant SupportThisworkwas supported by Breakthrough Breast Cancer, SusanG. Komen for
the Cure, and by the National Institute for Health Research (NIHR) BiomedicalResearch Centre at Guy's and St Thomas' NHS Foundation Trust and King'sCollege London.
Received April 10, 2013; revised August 16, 2013; accepted September 11, 2013;published OnlineFirst October 21, 2013.
References1. Husemann Y, Geigl JB, Schubert F, Musiani P, Meyer M, Burghart E,
et al. Systemic spread is an early step in breast cancer. Cancer Cell2008;13:58–68.
2. FehmT,Muller V, Alix-PanabieresC, Pantel K.Micrometastatic spreadin breast cancer: detection, molecular characterization and clinicalrelevance. Breast Cancer Res 2008;10 Suppl 1:S1.
3. Aguirre-Ghiso JA. Models, mechanisms and clinical evidence forcancer dormancy. Nat Rev Cancer 2007;7:834–46.
4. Klein CA. Framework models of tumor dormancy from patient-derivedobservations. Curr Opin Genet Dev. 2011;21:42–9.
5. Patel LR, Camacho DF, Shiozawa Y, Pienta KJ, Taichman RS.Mechanisms of cancer cell metastasis to the bone: a multistep pro-cess. Future Oncol. 2011;7:1285–97.
6. Weilbaecher KN, Guise TA, McCauley LK. Cancer to bone: a fatalattraction. Nat Rev Cancer 2011;11:411–25.
7. Uhr JW, Pantel K. Controversies in clinical cancer dormancy. ProcNatlAcad Sci USA 2011;108:12396–400.
8. Patel SA, Dave MA, Murthy RG, Helmy KY, Rameshwar P. Metastaticbreast cancer cells in the bone marrow microenvironment: novelinsights into oncoprotection. Oncol Rev 2011;5:93–102.
9. Goss PE, Chambers AF. Does tumour dormancy offer a therapeutictarget? Nat Rev Cancer 2010;10:871–7.
10. Ema H, Suda T. Two anatomically distinct niches regulate stem cellactivity. Blood 2012;120:2174–81.
11. Rao G, Patel PS, Idler SP, Maloof P, Gascon P, Potian JA, et al.Facilitating role of preprotachykinin-I gene in the integration ofbreast cancer cells within the stromal compartment of the bonemarrow: a model of early cancer progression. Cancer Res 2004;64:2874–81.
12. Shiozawa Y, Pedersen EA, Havens AM, Jung Y, Mishra A, Joseph J,et al. Human prostate cancer metastases target the hematopoieticstem cell niche to establish footholds in mouse bone marrow. J ClinInvest 2011;121:1298–312.
13. Bragado P, Sosa MS, Keely P, Condeelis J, Aguirre-Ghiso JA. Micro-environments dictating tumor cell dormancy. Recent Results CancerRes 2012;195:25–39.
14. Charafe-Jauffret E, Ginestier C, Iovino F, Wicinski J, Cervera N, FinettiP, et al. Breast cancer cell lines contain functional cancer stem cellswith metastatic capacity and a distinct molecular signature. CancerRes 2009;69:1302–13.
15. Kang Y, Siegel PM, Shu W, Drobnjak M, Kakonen SM, Cordon-CardoC, et al. A multigenic program mediating breast cancer metastasis tobone. Cancer Cell 2003;3:537–49.
16. Grigoriadis A, Mackay A, Noel E, Wu PJ, Natrajan R, Frankum J, et al.Molecular characterisation of cell linemodels for triple-negative breastcancers. BMC Genomics 2012;13:619.
17. Liu S, Ginestier C, Ou SJ, Clouthier SG, Patel SH, Monville F, et al.Breast cancer stem cells are regulated by mesenchymal stem cellsthrough cytokine networks. Cancer Res 2011;71:614–24.
18. Rohanizadeh R, SwainMV,MasonRS. Gelatin sponges (Gelfoam) as ascaffold for osteoblasts. J Mater Sci Mater Med 2008;19:1173–82.
19. Schneider A, Taboas JM, McCauley LK, Krebsbach PH. Skeletalhomeostasis in tissue-engineered bone. J Orthop Res 2003;21:859–64.
20. Song J, Kiel MJ, Wang Z,Wang J, Taichman RS, Morrison SJ, et al. Anin vivo model to study and manipulate the hematopoietic stem cellniche. Blood 2010;115:2592–600.
21. Aguirre-Ghiso JA, Liu D,Mignatti A, Kovalski K, Ossowski L. Urokinasereceptor and fibronectin regulate the ERK(MAPK) to p38(MAPK) activ-ity ratios that determine carcinoma cell proliferation or dormancy invivo. Mol Biol Cell 2001;12:863–79.
22. Giampieri S,ManningC,Hooper S, Jones L, Hill CS, Sahai E. Localizedand reversible TGFbeta signalling switches breast cancer cells fromcohesive to single cell motility. Nat Cell Biol 2009;11:1287–96.
23. Kodera Y, Katanasaka Y, Kitamura Y, Tsuda H, Nishio K, Tamura T,et al. Sunitinib inhibits lymphatic endothelial cell functions and lymphnode metastasis in a breast cancer model through inhibition of vas-cular endothelial growth factor receptor 3. Breast Cancer Res 2011;13:R66.
24. Nyati S, Schinske K, Ray D, Nyati MK, Ross BD, Rehemtulla A.Molecular imaging of TGFbeta-induced Smad2/3 phosphorylationreveals a role for receptor tyrosine kinases in modulating TGFbetasignaling. Clin Cancer Res 2011;17:7424–39.
25. Conley SJ, Gheordunescu E, Kakarala P, Newman B, Korkaya H,Heath AN, et al. Antiangiogenic agents increase breast cancer stemcells via the generation of tumor hypoxia. Proc Natl Acad Sci U S A2012;109:2784–9.
26. Sukhtankar D, Okun A, Chandramouli A, Nelson MA, Vanderah TW,Cress AE, et al. Inhibition of p38-MAPK signaling pathway attenuatesbreast cancer induced bone pain and disease progression in a murinemodel of cancer-induced bone pain. Mol Pain 2011;7:81.
27. Scholzen T, Gerdes J. The Ki-67 protein: from the known and theunknown. J Cell Physiol 2000;182:311–22.
28. Kobayashi A, Okuda H, Xing F, Pandey PR, Watabe M, Hirota S, et al.Bonemorphogenetic protein 7 in dormancy andmetastasis of prostatecancer stem-like cells in bone. J Exp Med 2011;208:2641–55.
29. Ranganathan AC, Adam AP, Aguirre-Ghiso JA. Opposing roles ofmitogenic and stress signaling pathways in the induction of cancerdormancy. Cell Cycle 2006;5:1799–807.
30. Ranganathan AC, Adam AP, Zhang L, Aguirre-Ghiso JA. Tumor celldormancy induced by p38SAPK and ER-stress signaling: an adaptiveadvantage for metastatic cells? Cancer Biol Ther 2006;5:729–35.
Marlow et al.
Cancer Res; 73(23) December 1, 2013 Cancer Research6898
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

31. Sosa MS, Avivar-Valderas A, Bragado P, Wen HC, Aguirre-Ghiso JA.ERK1/2 and p38alpha/beta signaling in tumor cell quiescence: oppor-tunities to control dormant residual disease. Clin Cancer Res 2011;17:5850–7.
32. Massague J. TGFbeta signalling in context. Nat Rev Mol Cell Biol2012;13:616–30.
33. Psaila B, Lyden D. The metastatic niche: adapting the foreign soil. NatRev Cancer 2009;9:285–93.
34. Malanchi I, Santamaria-Martinez A, Susanto E, Peng H, Lehr HA,Delaloye JF, et al. Interactions between cancer stem cells and theirniche govern metastatic colonization. Nature 2012;481:85–9.
35. Oskarsson T, Acharyya S, Zhang XH, Vanharanta S, Tavazoie SF,Morris PG, et al. Breast cancer cells produce tenascinCasametastaticniche component to colonize the lungs. Nat Med 2011;17:867–74.
36. Gao H, Chakraborty G, Lee-Lim AP, Mo Q, Decker M, Vonica A, et al.The BMP inhibitor Coco reactivates breast cancer cells at lung met-astatic sites. Cell 2012;150:764–79.
37. Shiozawa Y, Pedersen EA, Patel LR, Ziegler AM, Havens AM, Jung Y,et al. GAS6/AXL axis regulates prostate cancer invasion, proliferation,and survival in the bone marrow niche. Neoplasia 2010;12:116–27.
38. Nash KT, Phadke PA, Navenot JM, Hurst DR, Accavitti-Loper MA,Sztul E, et al. Requirement of KISS1 secretion for multiple organmetastasis suppression and maintenance of tumor dormancy. J NatlCancer Inst 2007;99:309–21.
39. de Barros AP, Takiya CM, Garzoni LR, Leal-Ferreira ML, Dutra HS,Chiarini LB, et al. Osteoblasts and bonemarrowmesenchymal stromalcells control hematopoietic stem cell migration and proliferation in 3Din vitro model. PLoS ONE 2010;5:e9093.
40. Gurkan UA, Kishore V, Condon KW, Bellido TM, Akkus O. A scaffold-free multicellular three-dimensional in vitro model of osteogenesis.Calcif Tissue Int 2011;88:388–401.
41. Heinemann C, Heinemann S, Worch H, Hanke T. Development of anosteoblast/osteoclast co-culture derived by human bone marrowstromal cells and human monocytes for biomaterials testing. Eur CellMater 2011;21:80–93.
42. Pallotta I, Lovett M, Rice W, Kaplan DL, Balduini A. Bone marrowosteoblastic niche: a new model to study physiological regulation ofmegakaryopoiesis. PLoS ONE 2009;4:e8359.
43. Nichols JE, Cortiella J, Lee J, Niles JA, Cuddihy M, Wang S, et al. Invitro analog of human bone marrow from 3D scaffolds with biomi-metic inverted colloidal crystal geometry. Biomaterials 2009;30:1071–9.
44. Goldstein RH, Reagan MR, Anderson K, Kaplan DL, Rosenblatt M.Human bone marrow-derived MSCs can home to orthotopic breastcancer tumors and promote bone metastasis. Cancer Res 2010;70:10044–50.
45. Moreau JE, AndersonK,Mauney JR,Nguyen T, KaplanDL, RosenblattM. Tissue-engineered bone serves as a target for metastasis of humanbreast cancer in a mouse model. Cancer Res 2007;67:10304–8.
46. Lu X, Mu E, Wei Y, Riethdorf S, Yang Q, Yuan M, et al. VCAM-1promotes osteolytic expansion of indolent bone micrometastasis ofbreast cancer by engaging alpha4beta1-positive osteoclast progeni-tors. Cancer Cell 2011;20:701–14.
47. Sethi N, Dai X,Winter CG, KangY. Tumor-derived JAGGED1promotesosteolytic bone metastasis of breast cancer by engaging notch sig-naling in bone cells. Cancer Cell 2011;19:192–205.
48. Barkan D, El Touny LH, Michalowski AM, Smith JA, Chu I, Davis AS,et al.Metastatic growth fromdormant cells inducedby a col-I-enrichedfibrotic environment. Cancer Res 2010;70:5706–16.
49. Barkan D, Kleinman H, Simmons JL, Asmussen H, Kamaraju AK,Hoenorhoff MJ, et al. Inhibition of metastatic outgrowth from singledormant tumor cells by targeting the cytoskeleton. Cancer Res2008;68:6241–50.
50. Ghajar CM, Peinado H, Mori H, Matei IR, Evason KJ, Brazier H, et al.The perivascular niche regulates breast tumour dormancy. Nat CellBiol 2013;15:807–17.
51. Charafe-Jauffret E, Ginestier C, Iovino F, Tarpin C, Diebel M, Esterni B,et al. Aldehyde dehydrogenase 1-positive cancer stem cells mediatemetastasis and poor clinical outcome in inflammatory breast cancer.Clin Cancer Res 2010;16:45–55.
Breast Cancer Metastatic Niche
www.aacrjournals.org Cancer Res; 73(23) December 1, 2013 6899
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991

2013;73:6886-6899. Published OnlineFirst October 21, 2013.Cancer Res Rebecca Marlow, Gabriella Honeth, Sara Lombardi, et al. CellsA Novel Model of Dormancy for Bone Metastatic Breast Cancer
Updated version
10.1158/0008-5472.CAN-13-0991doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2013/10/17/0008-5472.CAN-13-0991.DC1
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/73/23/6886.full#ref-list-1
This article cites 51 articles, 16 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/73/23/6886.full#related-urls
This article has been cited by 10 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/73/23/6886To request permission to re-use all or part of this article, use this link
on September 23, 2020. © 2013 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
Published OnlineFirst October 21, 2013; DOI: 10.1158/0008-5472.CAN-13-0991