Embed Size (px)
Citation preview

Biological Journal of the Linnean Society (1992), 47: 1-21
A nutritional analysis of foraging in the Malabar giant squirrel (Ratufa indica)
RENEE M. BORGES*
Department of Biology, University of Miami, Coral Gables, Florida 33124, U.S.A.
Received 5 3tih 1990, accepted f o r publication on I0 December 1990
The effect of nutrients and their availability on the diet of the herbivorous Malabar giant squirrel Ratufu indicu (Sciuridae) was investigated at Magod and Bhimashankar in western India. The daily consumption of food items (percent wet diet) and the intake rates of these items and the contained nutrients (wet g s-I) were determined by continuous observation of focal animals. Water content was a significant positive predictor of relative food item consumption while mineral contents in general had the opposite effect. The intake rates of water and more digestible nutrients such as non- structural carbohydrates, as well as the intake rate of minerals such as zinc (at Magod), were significant positive predictors of the relative contribution of an item to the daily diet, thus indicating considerations of feeding costs versus benefits. Tannins, some alkaloids, and other secondary metabolites may negatively influence food choice. Protein content, relative to digestibility reducers, influenced food item consumption only at Bhimashankar. There was seasonal variation in daily biomass consumption. At Bhimashankar, daily biomass consumption increased with the proportion of fruit in the diet while this did not occur at Magod. This is perhaps a result of the higher water content and the lower content of some soluble nutrients within fruit at Bhimashankar relative to Magod. There was also intra-month variation between individual squirrels in daily biomass of food consumed. Squirrels consumed ephemera1 food items opportunistically and non-ephemeral items such as mature leaves and bark on a regular daily basis. Squirrels probably obtained minerals and nitrogen from both fruit (especially seeds) and non-fruit sources (mature leaves and bark in the case of minerals, young and mature leaves in the case of nitrogen). However, they obtained lipid and non-structural carbohydrates mostly from fruit. Squirrels were selective in their utilization of tree species on a monthly basis, but this selectivity was not evident over a longer time period because individuals were constrained to use different phenological stages of tree species present within their territories. Large body size in this squirrel permits dietary flexibility and enables an overall generalist feeding strategy.
KEY WORDS:-Feeding strategy - generalist herbivore - India - nutrients - plant chemistry - Ratufa - tropical forest - tropical squirrel.
CONTENTS
Introduction . . . . . . . . . Materials and methods. . . . . . .
Study sites . . . . . . . . Observations on feeding activity of squirrels Food analyses . . . . . . . Tree phenology . . . . . . . Statistical analyses . . . . . .
Results . . . . . . . . . . Factors influencing food consumption . . Nutrient profiles of food items . . . .
. . . . . . . . . . . 2
. . . . . . . . . . . 3
. . . . . . . . . . . 3
. . . . . . . . . . . 3
. . . . . . . . . . . 4
. . . . . . . . . . . 5
. . . . . . . . . . . 5
. . . . . . . . . . . 5
. . . . . . . . . . . 5
. . . . . . . . . . . 1 0
*Current address: l04jC Aradhana, G.D. Ambekar Road, Naigaum, Bombay 400 014, India I
0024-4074/92/090001+ 2 1 SOS.OO/O 0 1992 The Linnean Society of London

R. M. BORGES
Seasonal variation in food and nutrient consumption Food consumption relative to food availability . . Tree species selectivity. . . . . . . .
Discussion . . . . . . . . . . . . Factors influencing feeding . . . . . . Profitability of item classes . . . . . . Seasonal variation in food consumption . . . Concordance between squirrels in food consumption Selectivity in tree species use . . . . . . Need for dietary flexibility. . . . . . . Limitations of this study . . . . . . .
Acknowledgements . . . . . . . . . References . . . . . . . . . . .
. . . . . . . . . 1 0
. . . . . . . . . I 1
. . . . . . . . . 1 2
. . . . . . . . . I 3
. . . . . . . . . 1 3
. . . . . . . . . I 6
. . . . . . . . . 1 6
. . . . . . . . . I 7
. . . . . . . . . 1 7
. . . . . . . . . 1 8
. . . . . . . . . 1 8
. . . . . . . . . I 9
. . . . . . . . . 1 9
INTRODUCTION
Herbivore food choice is influenced by nutritional and non-nutritional factors. Nutritional factors include positive nutrients such as calories, non-structural carbohydrates, protein, fat, minerals such as calcium and sodium (see Robbins, 1983 for review), and negative nutrients (sensu Westoby, 1978) such as digestibility reducers (fibre and tannins) and toxins (Freeland & Janzen, 1974; Westoby, 1978; Rosenthal & Janzen, 1979). Negative nutrients can reduce the availability of positive nutrients within a food resource. For example, the availability of protein, an item important in food choice, depends on the ratio of protein to digestibility reducers such as tannins and fibre (Milton, 1979). Tannins act as digestibility reducers because they bind to proteins (Van Sumere et al., 1975) and also inhibit the digestion of cellulose (Rhoades & Cates, 1976) and starch (Feeny, 1970). Herbivores, therefore, must balance their diets in terms of positive nutrients despite the presence of negative nutrients in potential food items (Westoby, 1974, 1978). Non-nutritional factors that can influence herbivore diets include the spatial and temporal heterogeneity of food resources and the social and reproductive status of the herbivores, among other parameters (Borges, 1989).
Studies of the nutritional aspects of food choice in tropical arboreal herbivores have been largely restricted to primates (Oates, Swain & Zantovska, 1977; Oates, 1978; Milton, 1979; Oates, Waterman & Choo, 1980; Glander, 1981, McKey et al., 1981; Baranga, 1982; Calvert, 1985; Davies, Bennett & Waterman, 1988). These studies have shown that food choice is complex and multifactorial. For example, McKey et al. (1981) showed that food choice in black colobus monkeys Colobus satanus could generally be explained by the ratio of nutrients (protein and minerals) to digestion inhibitors such as tannins and lignin. Avoidance of food items with high nutrientldigestion inhibitor ratios was generally found to be due to the presence of toxic secondary compounds.
Most foraging studies have been developed around the theoretical framework of optimal foraging theory (reviewed by Pyke, 1984; Stephens & Krebs, 1986) which makes the assumption that animals maximize fitness by foraging optimally. One way in which optimal foraging can occur is by maximizing the net rate of energy gain per unit feeding time. However, the application of optimal foraging models to herbivores which have to balance diets despite negative nutrients in potential food sources has not been completely successful (Belovsky, 1981, 1984). In another approach, Schoener (1969, 1971) attempted to categorize feeding strategies into two main types-time-minimization and

FORAGING IN RATUFA INDICA 3
energy maximization. However, Hixon (1982) urged the judicious use of this terminology because it assumes that many of the underlying nutritional and non- nutritional constraints on feeding behaviour are already known. Among these constraints are factors such as energetic requirements (Kleiber, 1932; McNab, 1986), thermal tolerance (Viljoen, 1984; Borges, 1989) , digestive capability (Milton, 1981), predation risk minimization (Lima, Valone & Caraco, 1985), seasonal fluctuations in resource availability due to resource phenology (Robinson, 1986), lack of access to certain resources due to territoriality (Borges, 1989) and limitations imposed by the reproductive or social status of the animal (Schaik & Noordwijk, 1985; Fleming, 1988). Therefore, it appears that empirical studies of foraging behaviour must proceed piecemeal until a realistic model of foraging behaviour for a particular species can be constructed.
Foraging in tree squirrels, especially in the tropics, has received scant attention. In the temperate zone, a few studies have interpreted tree squirrel food preferences from an optimal foraging viewpoint (Smith & Follmer, 1972; Lewis, 1980). Lewis (1982) argued for an inclusion of nutrient constraints and risk minimization as important factors in the food choice of the grey squirrel, Sciurus carolinensis. With regard to secondary compounds, Smith & Follmer (1972) felt that tannin content was not an important factor in the food choice of fox and grey squirrels. Smallwood & Peters (1986) found evidence for the fact that grey squirrels are able to detoxify tannins. In the tropics, however, Whitten & Whitten (1987) found that the hydrolysable tannin content of the bark negatively influenced bark feeding in the south-east Asian tree squirrel Sundasciurus lowii.
This paper analyses the foraging of the Malabar giant squirrel Ratufu indicu (Sciuridae) . This squirrel is a tropical, large, arboreal, facultatively frugivorous, generalist herbivore. The major portion of the daily diet of this species consisted of seeds whenever they were available. The fruit pulp of only a few species was consumed. During non-fruiting periods, alternative food sources consisted of young and mature leaves, flowers, pith and bark (Borges, 1989).
In this paper, the effect of the nutrient content of food resources, nutrient intake rates, association between nutrients within resources, resource phenology and tree species selectivity on determining diet composition is investigated. Some effects of the spatial and temporal heterogeneity of food resource availability on the diets of individual giant squirrels have already been described (Borges, 1989).
MATERIALS AND METHODS
Study sites
I observed squirrels at two sites in the Sahyadri mountains (Western Ghats) of India. Magod is a moist deciduous forest at an elevation of 665 m while Bhimashankar is an evergreen seasonal cloud forest situated at 900 m. Both sites receive seasonal monsoon rains from June through September. Detailed description of the study sites is available in Borges (1989).
Observations on feeding activip of squirrels
I followed ten individual squirrels each month from nest exit a t dawn to nest entry at dusk, a single squirrel on each observation day. I conducted intensive

4 R. M. BORGES
observations from January 1985 to September 1985 (inclusive) at Magod and from January 1986 to June 1986 (inclusive) a t Bhimashankar. I continuously timed the onset and duration of each activity to the nearest second (see Borges, 1989, for details of observational protocol). I repeated observation days if a focal squirrel was out of sight for a maximum of 60 minutes before noon (as concentrated feeding bouts occur before that time) or 90 minutes during the day. I recorded total counts of food items (in case of non-bark food items) and total volume (in case of bark food items) consumed each day. I estimated total volume of bark consumed by careful, close (5-10 m) observation of completely habituated squirrels and from bark and twigs dropped by these squirrels while feeding. I calculated the percent contribution of each item to the daily diet (PCDD) per squirrel per observation day in terms of wet mass. I converted volume into wet mass using average masses of bark pieces of known dimensions for that species. I recorded intake rates (g s-I) of each type of item for each focal squirrel per observation day either for 2 min at 5-min intervals throughout the day for small food items or by actually timing the consumption of the entire item in the case of a large food item (search and handling times included). I calculated intake rates for each nutrient from each item using the equality: nutrient intake rate (g s-I) = (g food item consumed s-I) x (g nutrient g food item-').
Food analyses
Food item samples were collected at the same time and at the same apparent stage of acceptability as those consumed by squirrels. A total of 63 items at Magod and 68 items at Bhimashankar was collected and prepared for analysis (see Borges, 1989, for list of items). These totals are 92.6% and 94.2% of all items consumed by squirrels a t Magod and Bhimashankar, respectively. Only items that were consumed by one or two individuals or that were rare enough to make collection difficult or impossible were not collected for analysis. Samples of a single item were pooled to obtain a singie nutrient value for the item. Food samples were dried to constant mass at 40°C to 50°C in kerosene ovens in the field and stored for laboratory analysis.
Food items were analysed for gross energy, ash, fat, nitrogen, total non- structural carbohydrates (TNC), minerals (Ca, Na, Mg, K, Fe, Zn), tannins, alkaloids and fibre in terms of neutral detergent fibre (NDF), acid detergent fibre (ADF) and crude lignin. NDF measures total plant cell wall content while ADF measures lignocellulose (Goering & Van Soest, 1970). The nutrient contents of food items are available in Borges (1989). Gross energy was estimated by bomb calorimetry. Ash was estimated by heating the sample to 600°C in a muffle furnace (Horwitz, 1980). Total nitrogen was measured by the Kjeldahl method (Horwitz, 1980). T N C was estimated using the modified Weinmann method (Smith, 1969; Milton, 1979). Total lipid content was determined by Soxhlet extraction using diethyl ether (Horwitz, 1980). NDF, ADF and lignin were determined by the procedure of Goering & Van Soest (1970). Mineral analyses (Ca, Mg, Na, K, Fe, Zn) were performed using an atomic absorption spectrophotometer (Varian Techtron AA 1275). Samples were prepared for mineral analyses by a procedure provided by Doran Hughes and Arnold Blum (personal communication) (see Borges, 1989, for details).

FORAGING IN RATUFA INDICA 5
Tannins were determined using a bovine serum albumin (BSA) assay (Wilson, 1984). This is a modification of the standard assay of Martin & Martin (1982). The protein precipitation capacity of the tannins was recorded as mg BSA precipitated per mg of dry tissue extract as in Martin & Martin (1982). Samples were tested qualitatively for the presence of alkaloids with Dragendorffs and Meyer’s reagents using extraction procedures and tests for false positive reactions recommended by Gartlan et al. ( 1980). Only samples that gave positive reactions with both Dragendorffs and Meyer’s reagents after false positive reactions were discounted were considered to contain alkaloids.
Tree phenology
The phenology of the forests was monitored on a regular basis (see Borges, 1989, for details). No attempt was made to estimate the relative proportions of a particular phenophase on a tree. Therefore, an individual tree could have been counted in more than one phenophase at a given time.
Throughout this paper, an item class refers to items within either an ephemeral (e.g. flush leaves and fruit) or a non-ephemeral feeding category (e.g. mature leaves and bark). In this paper, ‘bark’ is the generic term used to refer to feeding on either cambium, xylem, phloem or a combination of these tissues or woody twigs. Outer bark was never consumed.
Statistical analyses
Statistical analyses were generally performed using non-parametric procedures (Siegel, 1956). Bonferroni significance levels (Miller, 1966) were used throughout whenever multiple univariate tests were performed. Data were arcsine transformed for multiple regression analysis. All statistical analyses were performed on a wet mass basis since this takes into account wet bulk which is related to gut capacity and digestive ability of the animal.
RESULTS
Factors injuencing food consumption
Nutrient contents I examined univariate correlations (Kendall’s rank correlation tests) between
the percent contribution of individual food items to the daily diet (PCDD) and the nutrient content of the items (measured as percent wet mass) for all analysed nutrients (positive and negative) (see Borges, 1989, for detailed results). In general, squirrels did not consume items with higher nutrient content to a greater extent relative to those with a lower nutrient content. The only positive correlation obtained at both sites was between PCDD and water content (Kendall’s z = 0.12, jV = 569, P < 0.0001 at Magod; z = 0.22, N = 517, P < 0.0001 at Bhimashankar). In addition, at Magod potassium content was positively correlated with PCDD (z = 0.10, N = 516, P < 0.001).
Relative food item consumption was significantly negatively correlated with gross energy content at both sites and with the contents of Na, Ca and Fe at Magod. In addition, at Bhimashankar, with the exception of fat and TNC

6 R. M. BORGES
TABLE 1. Stepwise multiple linear regression of the nutrient contents of food items against the percent contribution of food items to the daily diet (wet mass
basis)
Magod Bhimashankar
Variable Variable entered entered ( N = 569) Multiple R ( N = 51 7) Multiple R
A: All nutrients in analysis
Zn f0.30999 P < 0.0005 Nitrogen -0.33163 P < 0.0004 Water f0.38892 P < 0.00001 Water f0.34408 P < 0.03
-
Na -0.26128 P < 0.00001 Ash -0.29500 P < 0.00001
Ca -0.40541 P < 0.01 K +0.35822 P < 0.01 TNC +0.41791 P < 0.03
B: Gross energy, ash, ADF, NDF, and lignin deleted from analysis Na -0.26353 P i 0.00001 Water +0.26323 P < 0.00001 Zn +0.30510 P < 0.001 Nitrogen -0.29765 P < 0.001
TNC f0.39801 P < 0.01 TNC f0.34591 P < 0.01 Water f0.38096 P < 0.00001 Na -0.32859 P < 0.001
Ca -0.40961 P < 0.03
TNC = Total non-structural carbohydrates. Variables not mentioned did not enter the analysis.
content (for which non-significant correlations were obtained), significant negative correlations were obtained for all other nutrients.
A stepwise multiple linear regression of all measured positive and negative nutrients against PCDD indicated that at both sites water and mineral contents were significant predictors of PCDD, water being a positive factor and minerals in general having the opposite effect with the exception of zinc at Magod (Table 1A). The multiple regression yielded more similar results for the two sites when the highly intercorrelated nutrients such as gross energy, ash and fibre contents were deleted from the analysis (Table 1B). In addition, the TNC content seems to be an important positive factor at both sites.
Feeding rates If nutrient intake rates are used as a currency in a cost-benefit feeding
analysis, then squirrels should consume greater amounts of food items from which they can obtain nutrients at a faster rate and therefore a t a lower cost relative to items with a lower feeding rate. I, therefore, initially calculated univariate correlations between the PCDD of food items and the daily average feeding rates (g s-I) of nutrients within the items (search and handling times included) (see Borges, 1989, for Kendall’s z values). The observations were pooled for each site. The PCDD of food items was positively correlated with the intake rate of gross energy. Significant positive correlations were also obtained with three calorigenic positive nutrients (lipid, crude protein and TNC) and one calorigenic negative nutrient, fibre (NDF and ADF at Magod and NDF only at Bhimashankar) . With regard to mineral content at Magod, significant positive correlations were obtained for intake rates of total ash, K, Mg, Ca and Zn. At Bhimashankar, positive correlations were obtained for intake rates of K, Na and Fe. Non-significant correlations were obtained for all other nutrients.
A stepwise multiple linear regression of feeding rates of nutrients from food items against the PCDD of the items showed that the results are similar to those

FORAGING IN RATUFA INDICA
2. Stepwise multiple linear regression of the intake rates of nutrients from food items against the percent contribution of food items to the
daily diet (wet mass basis) as the independent variable
7
TABLE (g s-'1
Magod
Variable entered ( N = 569) Multiple R
Zn +0.27316 P < 0.00001
Water +0.38448 P < 0.001 Nitrogen -0.45624 P < 0.00001
Ash -0.35212 P < 0.00001
Bhimashankar
Variable entered ( N = 517) Multiple R
TNC +0.26363 P < 0.00001
Water f0.30760 P < 0.004 Ash -0.28123 P < 0.02
~ ~~ ~~ ~ ~
TNC = Total non-structural carbohydrates. Variables not mentioned did not enter the analysis.
obtained with nutrient contents and indicates that feeding rates of water and minerals are the important predictors of PCDD (Table 2). The intake rate of some minerals such as zinc at Magod (which is highly correlated with minerals such as sodium and calcium) also appears to be important in determining PCDD (also see Borges, 1989, 1990).
(When nutrients are expressed in terms of dry mass, the results of the univariate and multivariate analyses presented above are similar to those obtained with wet mass, and allow the same conclusions to be drawn. The positive effects of water, TNC and minerals such as zinc (at Magod) and the negative effects of ash and fibre on PCDD are evident. Details of these analyses are available from the author.)
Tannins I examined the relationship between the PCDD of food items and tannin
activity measured as mg BSA precipitated per mg of tissue extract. The correlations were statistically significant neither at Magod ( P > 0.165) nor at Bhimashankar ( P > 0.08 1) . However, there is some preliminary evidence that tannins may negatively influence food selection. At Bhimashankar, nutrient contents were measured for ten items that were rejected by squirrels. These were specific parts of either flowers, fruits or leaves (e.g. the outer syconium wall of Ficus glomerata (Moraceae) or the petals of Garcinia talbotii (Clusiaceae)), that were consistently discarded by the squirrels. When the nutrient content of the eaten and discarded portions were compared, the only significant difference between them was in their tannin activity levels (Wilcoxon matched pairs signed ranks tests). The portions consumed were significantly lower in tannin activity than the portions discarded ( P < 0.01).
Ratio of protein to digestibility reducers I examined the relationship between the PCDD of food items and various
ratios of protein to digestibility reducers and found that at Magod, neither fibre alone nor tannin +fibre apparently reduced the nutritional value of an item significantly (Table 3) . In contrast, a t Bhimashankar, ADF, lignin and tannin content relative to protein content had a significant negative effect on the contribution of a food item to the diet. At this site the most significant relationship was obtained with the protein/ADF ratio.

8 R. M. BORGES
TABLE 3. Kendall’s rank correlation tests between percent contribution of food items to the daily diet and various ratios of protein to digestibility reducers (wet mass basis)
Kendall’s T Kendall’s T Ratio (Magod) N = 492 (Bhimashankar) N = 503
6.25 x N ADF+NDF+Lignin -0‘0499 (’= 0“04) 0.0600 ( P < 0.05)
6.25 x N ADF
6.25 x N NDF
6.25 x N Lignin
6.25 x N ADF + Tannin
-0.0149 ( P = 0.628) 0.1230 (P < 0.0001)*
-0.0618 ( P < 0.05) 0.0426 (P = 0.161)
-0.0266 ( P = 0.385) 0.0995 (P < 0.001)*
-0.0371 ( N = 466) ( P = 0.239) 0.1034 (Af = 408) ( P < 0.002)* ~~~~
*Indicates P-values significant after Bonferroni’s correction for multiple tests. X = Number of bod items.
Alkaloids and other secondary metabolites Sixteen food items (25.4% of analysed items) a t Magod and seven food items
(10.8% of analysed items) a t Bhimashankar tested positive for alkaloids. The most common tree at Magod was Aporosa lindleyana (Euphorbiaceae) (22.5% of all trees in study site). Its young leaves contained alkaloids as did those of the most common riverine tree at Bhimashankar, Mangifera indica (Anacardiaceae) (17.3% of all trees in the study site). I t appears, then, that the most common sources of young leaves for squirrels at both Magod and Bhimashankar contain secondary compounds such as alkaloids that squirrels are probably able to detoxify.
At Magod squirrels consumed only the ripe fruit pulp and seeds of Vitex altissima (Verbenaceae) and avoided its bark, leaves and flowers which contain the flavonoid vitexin (Rao, 1965). Squirrels did not eat the seeds of Olea dioica (Oleaceae) which are intensely bitter (Kirtikar & Basu, 1935). The wood of Terminalia paniculata (Combretaceae) is reported to contain ellagic acid-3,3‘- dimethyl ether, the corresponding glucoside and flavellagic acid-3,3’,4-trimethyl ether (Lowry, 1968) and squirrels did not consume any part of this species although it was fairly common in the core study area at Magod (6.8% of trees). The seeds of Strychnos nux-vomica (Loganiaceae) were never consumed probably because they contain the alkaloids strychnine, brucine, struxine, vomicine and colubrine as well as the glycoside loganin (Willaman & Schubert, 1961).
Although apparently unable to deal with some toxins, squirrels are able to detoxify certain other secondary compounds. For example, they fed extensively on the seeds, fruit pulp and wood of Holigarna arnottiana (Anacardiaceae). The seeds of this species contain a toxic lactone (Gibbs, 1974). The bark, wood and fruit (especially pulp) have intensely vesicant properties which may be caused by compounds such as laccol (Gibbs, 1974) , cardol, anacardic acid and anacardol (Symes & Dawson, 1953). The fruit pulp of only one fruit of S. nux-vomica was consumed on three different occasions. Since the fruit pulp contains the glycoside loganin (Willaman & Schubert, 1961), this suggests that an ability to detoxify these secondary metabolites, if present, is limited.
TA
BL
E
4A. C
orre
latio
n m
atri
x of
nut
rien
t con
tent
s w
ithin
foo
d ite
ms
at M
agod
(K
enda
ll's
rank
cor
rela
tion
coef
ficie
nts)
GE
-0
.823
8 A
sh
-0.2
2 0.
18
Fat
-0.2
8'*
0.38
'* 0.
20
N
-0.2
5"
0.33
'* 0.
24"
0.48
'*
TN
C
-0.2
4"
0.22
0.
10
0.10
0.
07
K
-0.1
0 0.
13
0.19
0.
27'*
0.
32'*
0.16
M
g -0
.11
0.07
0.
403*
0.
09
0.07
-0
.09
0.03
N
a -0
.24"
0.
24"
0.22
0.
07
0.12
0.
01
-0.1
6 0.
09
Ca
-0.3
0'*
0.25
" 0.4
6'.
0.10
0.
05
0.06
-0
.13
0.26
" 0.
56'*
Fe
-0.2
0 0.
18
0.28
'*
0.14
0.
11
-0.0
9 -0
.14
0.19
0.
58'*
0.48
?*
ND
F
-0.6
63*
0.63
3*
0.20
0.
08
0.11
0.
18
-0.0
2 0.
10
0.29
'*
0.32
'*
0.17
0.
503*
Z
n -0
.50'
8 0.
53'*
0.26
" 0.
34**
0.
30'*
0.
11
0.11
0.
10
0.41
3*
0.42
'* 0.
45'*
AD
F
-0.4
7'*
0.46
3*
0.24
-0
.04
-0.0
1 -0
.08
-0.1
2 0.
09
0.37
'* 0.
423*
0.
25"
OM
3*
0.803*
Lig
nin
-0.4
03*
0.41
3*
0.21
0.
07
0.06
0.
09
-0.0
1 0.
10
0.25
0.
34'*
0.15
0.
37'*
0.57
?*
0.70
3*
Wat
er
GE
Ash
Fa
t N
T
NC
K
M
g N
a C
a Fe
Z
n N
DF
A
DF
> z 0 2 9 5 2
N, N
itrog
en; G
E, g
ross
ene
rgy.
"P
Q 0
.01;
'P
Q 0
.003
3 =
Bon
ferr
oni
sign
ifica
nce
leve
l; 'P Q
0.0
01; 'P
< 0.
0001
. *V
alue
s si
gnifi
cant
afte
r B
onfe
rron
i's c
orre
ctio
n fo
r m
ultip
le t
ests
.
Y
b
TA
BL
E
4B. C
orre
latio
n m
atri
x of
nut
rien
t con
tent
s w
ithin
food
ite
ms
at B
him
asha
nkar
(K
enda
ll's
rank
cor
rela
tion
coef
ficie
nts)
8 G
E -0
.86'
* A
sh
-0.5
4'*
0.53
'* Fa
t -0
.35'
* 0.
39'*
0.39
?*
3 b N
-0
.44'
8 0.
50'8
0.
503*
0.
27'*
K
-0.4
6'8
0.46
'8 0.
603*
0.
34'*
0.42
'*
0.03
-0
.38'
8 0.
403*
0.
48'*
0.31
5~~
0.30
'*
0.02
0.
42'*
-0
.30'
* 0.
27'8
0.
28'*
0.
16
0.17
0.
1 1
0.15
0.
22"
Na
Ca
-0.5
3'8
0.53
'8
0.64
3*
0.30
2*
0.42
?*
0.01
0.
44'*
0.
41'*
0.
48'*
Fe
-0.3
7'*
0.35
'8 0.
32'8
0.23
" 0.
312*
0.
13
0.25
'*
0.25
'*
0.58
'*
0.49
?*
Zn
-0.4
8'*
0.45
'* 0.
41'8
0.23
" 0.
41'8
0.11
0.
30'*
0.41
'*
0.55
'*
0.62
'*
0.61
3*
ND
F
-0.6
03*
0.62
'* 0.
52?*
0.
35'*
0.
36'8
-0.0
6 0.
49'*
0.37
'* 0.
15
0.50
3*
0.23
" 0.
312*
A
DF
-0.4
5?*
0.50
'8
0.57
'*
0.32
'8
0.35
?*
-0.1
6 0.
453*
0.
413*
0.
19
0.52
'*
0.22
" 0.
31"
0.73
'*
Lig
nin
-0.3
73*
0.44
'*
0.49
'*
0.29
'* 0.
32'8
-0
.17
0.40
3*
0.38
'*
0.12
0.
43'.
0.20
0.
26"
0.62
'* 0.
833*
TN
C
-0.2
0 0.
17
-0.0
7 0.
16
-0.0
8
Mg
Wat
er
GE
A
sh
Fat
N
TN
C
K
Mg
Na
Ca
Fe
Zn
ND
F
AD
F
Col
umn
code
s as
in T
able
4A
.
10 R. M. BORGES
Nutrient profiles o f food items
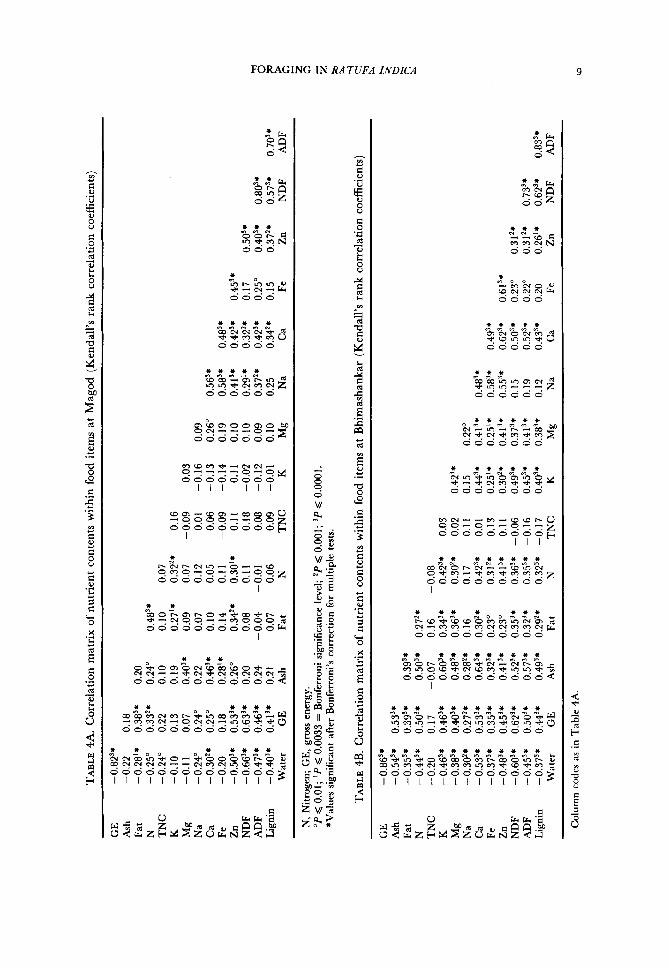
Xutrient inter-correlations The inter-correlations of nutrient contents within food items are presented in
the form of correlation matrices ( N ranges from 51 to 63 at Magod and from 59 to 68 at Bhimashankar) (Tables 4A, B). The results show that gross energy content is correlated with fat, nitrogen and fibre a t both sites. Nitrogen and ash are significantly correlated with fibre at Bhimashankar but not a t Magod. Minerals such as Ca, Fe and Zn are highly correlated with fibre at both sites.
Between-site comparison No significant difference was found between the nutrient content of food items
at the two sites except that gross energy and fibre contents of food items were higher at Magod than at Bhimashankar (Mann-Whitney U two-tailed tests; U = 1559, P < 0.001 for gross energy; U = 1195, P < 0.0001 for ADF).
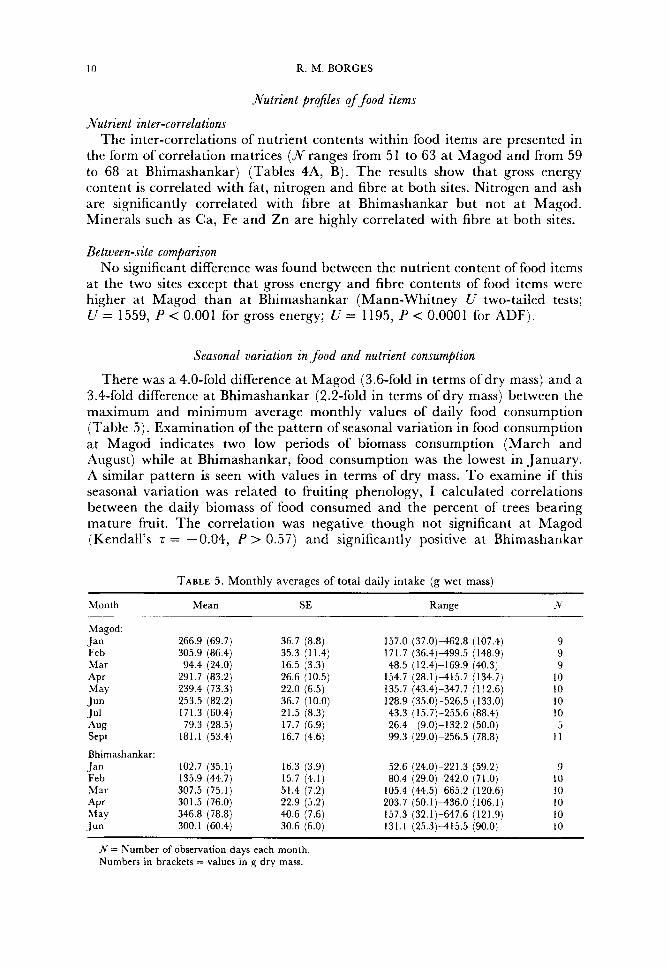
Seasonal variation in food and nutrient consumption
There was a 4.0-fold difference at Magod (3.6-fold in terms of dry mass) and a 3.4-fold difference at Bhimashankar (2.2-fold in terms of dry mass) between the maximum and minimum average monthly values of daily food consumption (Table 5). Examination of the pattern of seasonal variation in food consumption at Magod indicates two low periods of biomass consumption (March and August) while a t Bhimashankar, food consumption was the lowest in January. A similar pattern is seen with values in terms of dry mass. T o examine if this seasonal variation was related to fruiting phenology, I calculated correlations between the daily biomass of food consumed and the percent of trees bearing mature fruit. The correlation was negative though not significant at Magod (Kendall’s z = -0.04, P > 0.57) and significantly positive at Bhimashankar
TABLE 5 . Monthly averages of total daily intake (g wet mass) ~
Month Mean SE Range
Magod:
Fe b Mar
J a n
*Pr May JUn J U I
*us Sept
Bhimashankar: Jan Feb Mar Apr May J un
266.9 (69.7) 305.9 (86.4) 94.4 (24.0)
291.7 (83.2) 239.4 (73.3) 253.5 (82.2) 171.3 (60.4) 79.3 (28.5)
181.1 (53.4)
102.7 (35.1) 135.9 (44.7) 307.5 (75.1) 301.5 (76.0) 346.8 (78.8) 300.1 (60.4)
36.7 (8.8) 35.3 (1 1.4) 16.5 (3.3) 26.6 (10.5) 22.0 (6.5) 36.7 (10.0) 21.5 (8.3) 17.7 16.9) 16.7 (4.6)
16.3 (3.9) 15.7 (4.1) 51.4 (7.2) 22.9 (5.2) 40.6 (7.6) 30.6 (6.0)
157.0 (37.0)-462.8 (107.4) 171.7 (36.4)-499.5 (148.9) 48.5 (12.4)-169.9 (40.3)
154.7 (28.1)-415.7 (134.7) 135.7 (43.4)-347.7 ( 1 12.6) 128.9 (35.0) -526.5 (133.0) 43.3 (15.7)-255.6 (88.4) 26.4 (9.0)-132.2 (50.0) 99.3 (29.0)-256.5 (78.8)
52.6 (24.0)-221.3 (59.2) 80.4 (29.0)-242.0 (71.0)
105.4 (44.5)-665.2 (120.6) 203.7 (50.1)-436.0 (106.1) 157.3 (32.1)-647.6 (121.9) 131.1 (25.3)-415.5 (90.0)
9 9 9
10 10 10 10 5
1 1
9 10 10 10 10 10
X = Number of observation days each monlh Numbers in brackets = values in g dry mass.

FORAGING IN RATUFA INDICA
TABLE 6. Kendall’s rank correlation tests between the total daily intake of nutrients and the percent of non-fruit items in the daily diet (wet mass basis)
11
Kendall’s T Kendalls’s T Nutrient (Magod) N = 83 (Bhimashankar) N = 59
Gross energy -0.0626 ( P < 0.405) -0.3159 ( P < 0.0001)* Ash 0.1168 ( P i 0.12) -0.0662 ( P < 0.46) Fat -0.4114 ( P < 0.0001)* -0.3687 ( P < 0.0001)* Nitrogen 0.0638 ( P < 0.396) -0.051 ( P < 0.569) TNC -0.1807 ( P < 0.016) -0.4132 ( P < 0.0001)*
*Indicates values significant after Bonferroni’s correction for multiple tests. N = Number of observation days.
(Kendall’s z = 0.48, P < 0.0001). Also, the correlation between daily biomass consumption and the percent of fruit in the daily diet was non-significant at Magod (P > 0.365) and significantly positive at Bhimashankar ( P < 0.0001). Thus, at Bhimashankar squirrels consumed a greater daily food biomass in fruiting periods and daily food intake increased with the proportion of fruit in the diet; this did not occur at Magod.
There was considerable variation in daily biomass consumption between individual squirrels each month (e.g. as much as a 6.0-fold difference between maximum and minimum intramonthly values at Magod in July and a 6.3-fold difference at Bhimashankar in March).
There was significant monthly variation in daily intake of gross energy, ash, fat, nitrogen and TNC at both sites (Kruskal-Wallis one-way analysis of variance; Borges, 1989). To determine whether this was due to a seasonal variation in the intake of fruit versus non-fruit items. I examined correlations between the total daily consumption of these nutrients and the percent of diet contributed by non-fruit sources (Table 6). For gross energy I obtained a non- significant correlation at Magod and a significant negative correlation at Bhimashankar. I obtained non-significant correlations for ash and nitrogen at both sites. Significant negative correlations were obtained for TNC at Bhimashankar and for fat at both sites. Therefore, gross energy and TNC at Bhimashankar, and fat at both sites, are probably obtained from fruit sources.
Food consumption relative to food availability
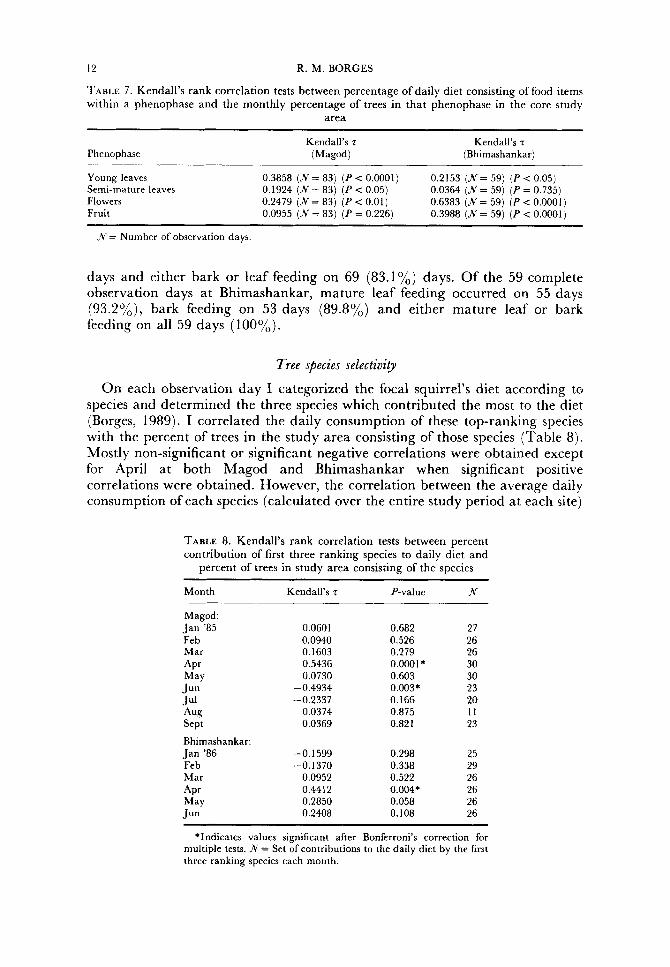
Unlike north temperate squirrels, these squirrels did not cache food at the study site and, therefore, their food consumption has to be analysed on a daily basis relative to available resources. I estimated the proportion of four major ephemeral item classes (flush leaves, semi-mature leaves, flowers and fruit) in squirrel daily diets and correlated these with the percent of trees bearing each item class (Table 7 ) . At Magod, positive correlations were obtained with all ephemeral phenological categories except fruit while at Bhimashankar significant positive correlations were obtained with all categories except semi- mature leaves. Therefore, in general, the diets of squirrels tracked the phenology of the forest.
Squirrels were consistent in their consumption of constantly available items such as mature leaves and bark. Of the 83 complete observation days at Magod, mature leaf feeding occurred on 49 (59%) days, bark feeding on 41 (49.4%)
12 R. M. BORGES
TABLE 7. Kendall’s rank correlation tests between percentage of daily diet consisting of food items within a phenophase and the monthly percentage of trees in that phenophase in the core study
area
Phenophase Kendall’s 7
(Bhimashankar)
Young leaves Semi-mature leaves Flowers Fruit
0.3858 ( N = 83) ( P < 0.0001) 0.1924 ( N = 83) (P < 0.05) 0.2479 (A’ = 83) ( P < 0.01) 0.0955 ( N = 83) (P = 0.226)
0.2153 ( N = 59) ( P < 0.05) 0.0364 ( N = 59) (P = 0.735) 0.6383 ( N = 59) ( P < 0.OOOl) 0.3988 ( N = 59) (P < 0.OOOl)
~ ~ ~~~~
JV = Number of observation days
days and either bark or leaf feeding on 69 (83.1%) days. Of the 59 complete observation days at Bhimashankar, mature leaf feeding occurred on 55 days (93.2%), bark feeding on 53 days (89.8%) and either mature leaf or bark feeding on all 59 days (100%).
Tree species selectivity
O n each observation day I categorized the focal squirrel’s diet according to species and determined the three species which contributed the most to the diet (Borges, 1989). I correlated the daily consumption of these top-ranking species with the percent of trees in the study area consisting of those species (Table 8). Mostly non-significant or significant negative correlations were obtained except for April at both Magod and Bhimashankar when significant positive correlations were obtained. However, the correlation between the average daily consumption of each species (calculated over the entire study period at each site)
TABLE 8. Kendall’s rank correlation tests between percent contribution of first three ranking species to daily diet and
percent of trees in study area consisting of the species
Month Kendall’s 7 P-value x Magod: Jan ’85 Feb Mar *Pr May J u n Ju l *ug Sept
Bhimashankar: Jan ’86 Fe b Mar
May *Pr
J u n
0.0601 0.0940 0.1603 0.5436 0.0730
-0,4934 -0.2337
0.0374 0.0369
-0.1599 - 0.1370
0.0952 0.4412 0.2850 0.2408
0.682 0.526 0.279 0.0001* 0.603 0.003* 0.166 0.875 0.82 1
0.298 0.338 0.522 0.004* 0.058 0.108
27 26 26 30 30 23 20 I I 23
25 29 26 26 26 26
*Indicates values significant after Bonferroni’s correction for multiple tests. N = Set of contributions to the daily diet by the first three ranking species each month.

FORAGING IN RATUFA INDICA 13
and the percent of trees at each site consisting of the species was significant at both Magod (Kendall's z = 0.66, P < 0.0001) and at Bhimashankar (Kendall's z = 0.43, P < 0.05).
DISCUSSION
The Malabar giant squirrel Ratufa indica is a facultatively frugivorous, generalist herbivore. Below I discuss my findings in the context of other herbivores and propose that within certain dietary constraints, dietary flexibility is an important feature of the foraging strategy of this squirrel.
Factors inzuencing feeding
Nutrient contents Water was the only nutrient which was positively correlated with the percent
contributions of food items to the daily diet (PCDD) at both study sites. Similarly the mature leaves of Gomphandra coriacea (Icacinaceae) which had the highest water content compared with other available leaf species, constituted the most important item in the diet of the south Indian leaf monkey Presbytisjohnii (Oates et al . , 1980). Water content was also a strong positive predictor of the cellulase digestibility of leaves in an African montane forest (Waterman et al., 1983). This suggests a relationship between water content and the profitability of an item in terms of soluble nutrients such as TNC.
The PCDD of food items and nutrient content were not positively correlated indicating that nutrient content per se is not the criterion for the relative consumption of food items. This has also been found for several primates such as the black colobus monkey Colobus satanus in which nutrient content relative to digestibility was important in food choice (McKey et al., 1981). Also, the strong negative correlation seen at both sites between PCDD and gross energy could perhaps be due to the correlation between energy and fibre. A strong negative correlation between gross energy and number of feeding records has also been observed in C. satanus (McKey et al., 1981).
At both sites a stepwise linear regression of nutrient contents against the PCDD of items indicated that water made a significant positive contribution whilst minerals in general made a negative contribution to the regression equation. Since mineral content is positively correlated with fibre content at both sites (Table 4), a negative effect of minerals may perhaps be interpreted as the negative effect of fibre. The positive contribution of TNC to the equation at Magod and the negative contribution of nitrogen at Bhimashankar (which at this site is strongly positively correlated with fibre within food items) further strengthens this view. When the results of the multivariate analysis are combined with those of the univariate tests, it appears that squirrels at both sites may have used water content as one possible cue in the selection of items with greater digestibility (such as young leaves) and thereby greater profitability.
The importance of water itself as a nutrient should not be overlooked (Maynard & Loosli, 1969; Glander, 1978; Wrangham, 1981), particularly since squirrels did not drink water at the study sites and obtained their water requirements entirely from plant material. Investigation of water balance in these squirrels should provide more insight into these phenomena.

14 R. M. BORGES
Feeding rates At both study sites, univariate statistics revealed that squirrels appeared to
consume items in proportion to the intake rates of gross energy and minerals from the items. Since gross energy was correlated with protein, fat and TNC contents (Table 4), squirrels consuming items with a high rate of energy intake automatically ingested items rich in positive nutrients. Similarly, Kenward (1982, 1983) and Kenward & Parish (1986) reported that the extent of bark stripping of beech, sycamores and maples by grey squirrels, S. carolinensis, in the British Isles was positively correlated with the volume of sap flow or phloem width per unit area and not with sugar concentration.
Multiple regression analysis also showed that intake rates of water and TNC were important positive predictors of the PCDD of items at both sites. At Magod, intake rates of minerals such as zinc (and, therefore, of associated minerals such as calcium) appear to be important. Since such minerals are obtained largely from fibrous item classes such as mature leaves and bark (Ernst, 1975; Borges, 1989, 1990), the intake rates of items with lower digestibilities could also influence diet composition at Magod. In the case of female squirrels at Magod, there was a significant positive correlation between the PCDD of food items and the calcium intake rates while this correlation was neither significant for males at the same site nor for either sex at Bhimashankar (see Borges, 1990, for a discussion of this phenomenon). As suggested by Borges (1990), an explanation for the absence of a sexual difference in calcium consumption at Bhimashankar is the existence of higher calcium intake rates and the probable presence of more easily digestible calcium at this site relative to, Magod. The fact that zinc content and zinc intake rates are strong predictors of PCDD at Magod while no such pattern is evident at Bhimashankar may indicate that in general minerals such as calcium and zinc may be more easily available at Bhimashankar than at Magod.
The ability of these squirrels to digest fibre is unknown although species within the genus Ratufa have enlarged caeca (Payne, 1979). Squirrels consumed considerable quantities of mature leaves and bark when young leaves, flowers and fruit were not available, suggesting that a fermentation capacity exists. I have recorded a maximum of 53.3% bark and 85.5% mature leaves in the daily diet of individuals of this squirrel species (Borges, 1989).
Tannins The effect of tannins on squirrel food choice is not clear. There appeared to
be a negative effect of tannins on food consumption at Bhimashankar as shown by lower tannin activity levels (as measured by astringency) of ten selected versus rejected food items. The non-significant correlation obtained between PCDD of items and tannins can be interpreted in two ways. (1) Squirrels only consumed items with tannin levels below a certain maximum, so that within the diet there was no obvious relationship between the consumption of items and tannins. (2) Squirrels are able to detoxify tannins and hence their food choice was not determined by tannins. With currently available data, it is not possible to choose between these hypotheses because firstly, the ability of these squirrels to detoxify tannins is unknown, and secondly, a systematic survey of tannins in items that were not eaten by squirrels a t both sites was not conducted. Therefore, i t is not known if there is a threshold tannin content or astringency beyond

FORAGING IN RATUFA INDICA 15
which food items are unacceptable to squirrels. Only extensive comparison of the tannins of selected and non-selected items will resolve this issue.
I did not estimate the levels of condensed and hydrolysable tannins within food items but used a protein precipitating assay instead, since the astringency of tannins rather than their concentration is considered to be biologically relevant (Bate-Smith, 1973). However, according to Zucker (1983), precipitation of proteins such as enzymes may not destroy their activity. This opens the question of astringency as a measure of the defensive properties of tannins. According to Waterman (1983, 1984), tannins were not an absolute deterrent in food selection in most primates studied and the role of tannins in food selection may have been over-emphasized (see also Bernays, 1981). The fact that temperate squirrels may have the ability to detoxify tannins (Smith & Follmer, 1972; Smallwood & Peters, 1986), seems to strengthen this view.
Ratio of protein to digestibility reducers The digestibility of protein is positively associated with the digestibility of the
non-protein components of food (Glover & Duthie, 1958). Therefore, a low protein/digestion inhibitor ratio could also mean a low value of available, digestible nutrients in general. At Bhimashankar, but not at Magod, ADF, lignin and tannin content relative to protein content had a significant negative relationship with the PCDD of a food item. The significance of these differences between the sites cannot be interpreted until data on protein/fibre ratios of selected versus non-selected items at both sites are compared. With available data, it is not possible to decide if squirrels at Magod and Bhimashankar differ in their feeding criteria with respect to these ratios or whether the fibre content of the vegetation at Magod is generally higher than at Bhimashankar resulting in lower protein/fibre ratios. The fact that food items at Magod were higher in fibre content than at Bhimashankar suggests this is the case, but more data on the phytochemistry of both sites are needed to resolve this issue. The negative effect of digestion inhibitors on food choice has also been observed in several species of primates (Milton, 1979; Oates et al., 1980; McKey et al., 1981), ungulates (Belovsky, 1981, 1984) and rodents (Batzli & Cole, 1979).
Alkaloids and other secondary metabolites The available evidence indicates that while R. indica is capable of detoxifying
some alkaloids and other secondary metabolites that occur within common tree species at both study sites, it avoided the secondary compounds and alkaloids in other foods. Hladik (1977) and Waterman & Choo (1981) found no correlation between the presence of alkaloids in food items and their consumption in certain primates. However, alkaloids can vary tremendously in their toxicity (Levin & York, 1978). According to McKey et al. (1981), the seeds of Strychnos tricalysioides are avoided by C. satanus probably because of their alkaloid content. Therefore, the mere presence of alkaloids within food items may not be a good indication of their anti-herbivore properties.
The relationship of secondary chemistry to food choice in generalist non- ruminant herbivores such as R . indica is bound to be constrained by detoxification ability, especially since they do not have a complex microbial stomach flora to assist in the detoxification process (Freeland & Janzen, 1974).

16 R. M. BORGES
Prof;tability of item classes
Nitrogen sources Squirrels can obtain nitrogen from young and mature leaves, judging from the
higher contents of nitrogen within these item classes relative to other classes (Borges, 1989). Moreover, young leaves had higher nitrogen contents than mature leaves, as also seen in other studies (Milton, 1979; McKey et al., 1981).
Mineral sources Squirrels may consistently obtain minerals from mature leaves and bark. This
is supported by the fact that although the extent of the mature leaf and bark feeding varied with individuals and with months (Borges, 1989), yet ( 1 ) mature leaves and/or bark usually contributed to the daily diet even in fruiting periods, (2) mature leaves and bark were highest in mineral content (Borges, 1989, 1990) and ( 3 ) the mineral content, especially of calcium and sodium, increases with the maturity of the leaf (Ernst, 1975; Baranga, 1983; Borges, 1989, 1990). As described in Borges (1989), squirrels at Bhimashankar fed on only the central portion of the mature leaves of Memecylon umbellatum (Melastomataceae) and Diploclisia macrocarpa (Menispermaceae) whose mineral contents were greater than those of the outer leaf rims. However, the fact that mature leaves and bark could also be consumed consistently to provide roughage (since they are rich in fibre; cannot be ruled out.
TNC and lipid sources Seeds may be a profitable, easily digestible source of energy and lipid judging
from their higher energy, TNC and fat contents and lower fibre contents relative to other item classes (Borges, 1989). Since the calorigenic value of fat is twice that of non-structural carbohydrates (Maynard et al., 1979), seeds can be extremely rich energy sources. Also, their gross energy value probably closely matches their energy digestibilities because of their low fibre contents. It is not surprising, therefore, that squirrels should be facultative seed-eaters during fruiting periods. In other squirrels also (e.g. grey and fox squirrels) choice of seeds was related to their lipid contents and digestibilities (Smith & Follmer, 1972).
Judging from the correlations between total daily intake of non-fruit items and the total daily intake of ash, fat, nitrogen and TNC, it appears that at both sites, squirrels can obtain ash and nitrogen from both fruit (especially seeds) and non-fruit sources (mature leaves and bark in the case of ash, young and mature leaves in the case of nitrogen). However, they obtain lipid and TNC mostly from fruit sources. This has also been found for primates (Milton, 1979; McKey et al., 198 1 ; Calvert, 1985).
Seasonal variation in . food consumption
There was seasonal variation in daily biomass consumption at both sites. The correlation between daily biomass consumption and fruiting phenology was non- significant at Magod and significantly positive at Bhimashankar. Examination of the seasonal pattern of consumption of different food item classes (Borges, 1989) shows that at Magod, the early high values of biomass consumption (January

FORAGING IN RATUFA INDICA 17
and February) are due mostly to the consumption of young leaves and flowers, and the later high values (April-July, September) are due mainly to the consumption of fruit. The two low values of biomass consumption at Magod (March and August) result from the consumption of semi-mature and mature leaves, bark and pith, all of which have high fibre contents. I t is possible that at Magod, the correlation between biomass consumption and fruiting phenology is not significant because young leaves and flowers contribute mainly to the early high values of biomass consumption. However, at Bhimashankar, the high values of biomass consumption (March-June) are due mostly to fruit consumption, while the lower values (January-February) result mostly from the consumption of mature leaves, bark and flowers (Borges, 1989). These patterns of food item consumption closely track phenological events at both sites with the exception of fruit consumption at Magod.
At Bhimashankar, there was a significant correlation between daily biomass consumption and the proportion of fruit in the diet while this relationship was not significant at Magod. The fact that fruit (both pulp and seeds) at Bhimashankar was higher in water content ( P < O.OOOl), lower in gross energy ( P < 0.0016), fat ( P < 0.04) and protein ( P < 0.002), but not in TNC ( P > 0.63) than at Magod (Mann-Whitney two-tailed tests) may explain this finding.
Concordance between squirrels in food consumption
Borges (1989) found that squirrels a t both sites were concordant in their monthly consumption of food item classes. However, they were not concordant in their consumption of individual food items. This is a result of territoriality- based restriction of access to certain food items (Borges, 1989) which also explains the considerable intramonth variation in biomass consumption between individual squirrels. Squirrels consumed non-ephemeral item classes such as mature leaves and bark with consistency throughout the study period and ephemeral items of greater profitability in an opportunistic manner as indicated by the positive correlations between contributions of young leaves and flowers to the diet and the phenology of these items at both sites. However, a non-significant correlation was obtained at Magod between daily fruit consumption and fruit phenology while a positive correlation was obtained at Bhimashankar. This difference can be explained by the fact that fruit at Magod was obtained extensively from tree species that were rare, e.g. Artocarpus hirsutus (Moraceae) (1.8% of trees a t the study site), Garcinia indica (Clusiaceae) (0.6:/,) and Carallia brachiata (Rhizophoraceae) (0.2%) as well as from common species such as Aporosa lindleyana (22.5%) and Olea dioica (13.4%) whereas at Bhimashankar, fruit feeding was dominated by one species, Mangifera indica, which was a common riverine species (1 7.3% of trees at the study site) accessible to all individual squirrels.
Selectivity in tree species use
Squirrels appeared to be selective in their use of tree species on a monthly basis as indicated by the non-significant and negative correlations obtained between daily consumption of the three species that contributed most to the daily diet

18 R. M. BORGES
and the percent of trees in the core area consisting of those species (Table 8) . Since all individual trees of a species did not enter the different phenophases (Borges, 1989), the above correlations are conservative in that they assumed that all individuals contributed to a particular phenophase. However, positive correlations were obtained for diets in April at both Magod and Bhimashankar. This is because in these months at both sites, fruit of the two most common tree species was available and consumed (A. lindleyana and 0. dioica at Magod together constituting 35.9% of trees, and M . indica and 0. dioica at Bhimashankar together constituting 2 1 .O% of trees). However, the average daily consumption of a species (irrespective of the plant part) calculated over the entire study period was positively correlated with tree species abundance. Therefore, on a time scale greater than one month, e.g. the six or nine month duration of the study at the two sites, the large contribution and consequent importance of rare species to the daily diet was no longer detected. I n howler monkeys Alouatta palliata, Glander (1981) found the opposite result, i.e. over the entire study period, percent feeding time on a tree species by a troop was negatively correlated with tree species abundance. I believe that this difference is to be expected when comparing a solitary territorial generalist herbivore such as R. indica with a group-territorial generalist such as the howler monkey in which all individuals within the troop have equal access to the forest space occupied by the troop. Individuals of R. indica are forced to utilize many different phenological stages of most tree species that occur within individual territories and therefore cannot exhibit overall tree species selectivity over a long time scale.
Need f o r dietary Jexibility
The ability to utilize resources of very low profitability when unavoidable and greater amounts of profitable resources when available indicates a flexible, generalist feeding strategy. As suggested by Borges (1989), the large body size of this squirrel (median body mass = 1364 g, range = 909-2316 g, British Museum Collection) and its proportionately longer gut retention time (Demment, 1983; Demment & Van Soest, 1985), which leads to greater digestibility, may serve as insurance against shortages of the more highly digestible items and may be an important component in the ecology of food scarcity in this species. The spatiotemporal heterogeneity of resources is unique for each individual squirrel, making dietary flexibility an essential feature of the feeding ecology of this species.
Limitations o f this study
This study has limitations in the extent to which food ‘values’ can be assigned to food items. This is because digestibilities of items were not estimated and no data on the digestive ability of these squirrels are currently available. The extensive consumption of bark and mature leaves on occasion seems to indicate fermentation ability. However, the fact that the PCDD of items was influenced positively by water content and intake rates of more highly digestible nutrients seems to indicate that fermentation capacity is limited. Another limitation of this study is that analysis of items not consumed by squirrels was not performed (with few exceptions). Therefore, thresholds for selection of nutrients within potential food items cannot be identified at present. An additional limitation is that the

FORAGING IN R A T U F A I N D I C A 19
differential costs of feeding on items are not known. The fact that intake rates of nutrients are significant feeding criteria indicate that feeding costs versus feeding benefits are important considerations. Data on digestibility of food items, nutrient analysis of uneaten food items and on squirrel energetics, will help to resolve many questions raised by this study.
ACKNOWLEDGEMENTS
I am grateful to T. H. Fleming, W. E. Glanz, C. C. Horvitz, D. B. McKey, A. P. Smith and anonymous reviewers for useful comments on this manuscript. I thank Mahadeva, Asawale, Soma, Ganpat, and Kisan for help with fieldwork. I thank the Karnataka Power Corporation and the Maharashtra Tourism Development Corporation for logistic support in the field. I thank J. C. Daniel and the Bombay Natural History Society for collaboration. I am grateful to ITALAB Pvt. Ltd., and St. Xavier’s College, Bombay, and the Department of Animal Science, Cornell University, for assistance with nutrient analyses. I thank R. M. Syren for assistance with data management. This research was conducted in partial fulfilment of the requirements for a Ph.D. degree from the University of Miami, Coral Gables, USA. Funding for this research was provided by the United States Fish and Wildlife Service and by the Chicago Zoological Society.
REFERENCES
BARANGA, D., 1982. Nutrient composition and food preferences of colobus monkeys in Kibale forest,
BARANGA, D., 1983. Changes in chemical composition of food parts in the diet of colobus monkeys. Ecology,
BATE-SMITH, E. C., 1973. Haemanalysis of tannins: the concept of relative astringency. Phytochemisty
BATZLI, G. 0. & COLE, F. R., 1979. Nutritional ecology of microtine rodents: digestibility of forage. journal
BELOVSKY, G. E., 1981. Food selection by a generalist herbivore: the moose. Ecology, 62: 1020-1030. BELOVSKY, G . E., 1984. Herbivore optimal foraging: a comparative test of three models. American Naturalist,
BERNAYS, E. A,, 1981. Plant tannins and herbivores: an appraisal. Ecological Entomology, 6: 353-360. BORGES, R. M., 1989. Resource Heterogeneity and the Foraging Ecology of the Malabar Giant Squinel Ratufa indica.
Unpublished Ph.D. Thesis, University of Miami. BORGES, R. M., 1990. Sexual and site differences in calcium consumption by the Malabar Giant Squirrel
Ratufa indica. Oecologia 85: 80-86. CALVERT, J. J,, 1985. Food selection by western gorillas (G. g . gorilla) in relation to food chemistry. Oecologia
(Berl in) , 6 5 : 236-246. DAVIES, A. G., BENNETT, E. L. & WATERMAN, P. G., 1988. Food selection by two south-east Asian
colobine monkeys (Presbytis rubicmda and Presbytis melalophos) in relation to plant chemistry. Biological Journal of the Linnean Society, 34: 33-56.
DEMMENT, M. W., 1983. Feeding ecology and the evolution of body size in baboons. AJrican Journal OJ
DEMMENT, M. W. & VAN SOEST, P. J., 1985. A nutritional explanation for body-size patterns of
ERNST, W., 1975. Variation in the mineral content of leaves of trees in miombo woodland in South Central
FEENY, P., 1970. Seasonal changes in oak leaf tannins and nutrients as a cause of spring feeding by winter
FLEMING, T. H., 1988. The Short-tailed Fruit Bat: a Study in Plant-animal Interactions. Chicago: Chicago Uniuersity
FREELAND, W. J . & JANZEN, D. H., 1974. Strategies in herbivory by mammals: the role of plant secondary
Uganda. African journal of Ecology, 20: 113-121.
64: 668-673.
(Oxford ) , 12: 907-912.
of Mammalogy, 60: 740-750.
124: 97-1 15.
ECOIOQ, 21: 219-233.
ruminant and non-ruminant herbivores. American Naturalist, 125: 641-672.
Africa. Journal oJ Ecology, 63: 801-807.
moth caterpillars. Ecology, 51: 565-581.
Press.
compounds. American Naturalist, 108: 269-289.

20 R. M. BORGES
GARTLAN, J . S., h4cKEY, D. B., WATERMAN, P. G., MBI, C. N. & STRUHSAKER, T . T., 1980. A comparative study of the phytochemistry of two African rain forests. Biochemical Systematics B Ecolqpy, 8:
GIBBS, R. D., 1974. Chemotaxonomy of Flowering Plants. Montreal and London: McGill-Queen's University Press.
GLANDER, K . E., 1978. Drinking from arboreal water sources by mantled howling monkeys (Allouatta palliata). Folia Primatologica, 29: 206-2 17.
GLANDER, K . E., 1981. Feeding patterns in mantled howling monkeys. In A. C. Kamil & T . D. Sargent (Eds), Foraging Behauior: Ecological, Ethological, and Psychological Approaches: 23 1-257. New York: Garland STPM Press.
GLOVER, J. & DUTHIE, D. W. 1958. The nutritive ratioicrude protein relationship in ruminant and non- ruminant digestion. Journal of Agricultural Science, 50: 227-229.
GOERING, H. K. & VAN SOEST, P. J., 1970. Forage Fiber Analyses. (Apparatus, Reagents, Procedures and Some Applications). Agriculture Handbook Number 379. Washington, D.C.: United States Department of Agriculture.
HIXON, M. A., 1982. Energy maximizers and time minimizers: theory and reality. American Naturalist, 119:
HLADIK, C. M., 1977. Chimpanzees of Gabon and chimpanzees of Gombe: some comparative data on the
HORM'II'Z, W., 1980. Ojicial Methods of Analysis of the Association of OJicial Analytical Chemists, 13th edn.
40 1-422.
596-599.
diet. In '1. H. Clutton-Brock (Ed.) , Primate Ecology: 481-503. London: Academic Press.
Washington D.C.: AOAC. 0
tiENWARD, R. E., 1982. Bark stripping by grey squirrels-some recent research. Quarterly j%urnal of Foresty, 76: 108-121.
KENWARD, R. E., 1983. The causes of damage by red and grey squirrels. lcfammaf Review, 13: 159-166. KENWARD, R. E. & PARISH, T., 1986. Bark stripping by grey squirrels. Journal o f ~ o o l o ~ r (London), 210:
KIRTIKAR, K. R. & BASU, B. D., 1935. Indian Medicinal Plants, 2nd edn. Revised by E. Blatter, J . F. Caius
KLEIBER, M., 1932. Body size and metabolism. Hil,gardia, 6: 315-353. LE\'IN, D. A. & YORK, B. M., 1978. The toxicity of plant alkaloids: an ecogeographic perspective.
LEiYIS, A. R.. 1980. Patch use by gray squirrels and optimal foraging. Ecology, 61: 1371-1379. LEWIS, A. R. , 1982. Selection of nuts by gray squirrels and optimal foraging theory. American Midland
LIMA, S . L., VALONE, T. J. & CARACO, 'l., 1985. Foraging efficiency-predation risk trade-off in the grey
LOWRY, J. B., 1968. The distribution and potential taxonomic value of alkylated ellagic acids. Phytochemistry,
MARI'IN, J . S . & MARTIN, M. M., 1982. Tannin assays in ecological studies: lack of correlation between phenolics, proanthocyanidins and protein-precipitating constituents in mature foliage of six oak species. Oecologia ( Berlin), 54: 205-2 1 1.
473-48 1.
and K. S. Mhaskar. Calcutta: Prabasi Press.
Biochemical Systematics €9 Ecology. 6: 61-76.
.?;aturaiistsl, 107: 250-257.
squirrel. Animal Behauiour, 33: 155-165.
7: 1803-1813.
MAYN.4RD, L. A. & LOOSLI, J. K., 1969. Animal Nutrition. New York: McGraw-Hill. MAYNARD. L. A., LOOSLI, J. K., HINTZ, H . F. & WARNER, R. G., 1979. Animal Nutrition, 7th edn. New
York: McGraw-Hill. McKEY, D. B., GARTLAN, J. S., WATERMAN, P. G. & CHOO, G. M., 1981. Food selection by black
colobus monkeys (Colobus satanas) in relation to plant chemistry. Biological Journal of the Linnean Sociely, 16; 115-~146.
SfcNAB, B. K., 1986. The influence of food habits on the energetics of eutherian mammals. Ecological Monographs, 56: 1-19.
.MILLER, R. G., Jr., 1966. Simultaneous Statistical Injierence. New York: McGraw-Hill. MILTON, K., 1979. Factors influencing leaf choice by howler monkeys. A test of some hypotheses of food
SIILTOK, K., 1981. Food choice and digestive strategies of two sympatric primate species. ilmerican .Vaturalist,
OATES, J. F., 1978. Water-plant and soil consumption by guereza monkeys (Colobus guerera): a relationship
OATES, J . F., SWAIN, T . & ZANTOVSKA, J., 1977. Secondary compounds and food selrction by colobus
OATES, J. F., WATERMAN, P. G. & CHOO, G. M., 1980. Food selection by the South Indian leaf-monkey,
PAYNE, J. B., 1979. Synecology of Malayan tree squirrels, with particular reference to the genus Ratufa. Unpublished
I'YKE, G. H., 1984. Optimal foraging theory: a critical review. Annual Review of Ecology @ Systematics, 15:
RAO, 1). s., 1965. ~~aturwissen~chaf ten , 52: 262.
selection by generalist herbivores. American Naturalist, 114: 362-378.
117: 496-505.
with minerals and toxins in the diet? Biotropica, 10: 241--253.
monkeys. Biochemical Systematics B Ecology, 5: 3 17-32 1 .
PresbytiJ johnii, in relation to leaf chemistry. Oecologia (Ber l in) , 45: 45-56.
Ph.D. Thesis, Univerqity of Cambridge.
523--575.

FORAGING IN R A T U F A I N D I C A 21
RHOADES, D. F. & CATES, R. G., 1976. Toward a general theory of plant anti-herbivore chemistry. In J. W. Wallace & R. L. Mansell (Eds), Biochemical Interactions Between Plants and Insects, Recent Advances in Phytochemistry, 10: 168-213. New York: Plenum Press.
ROBBINS, C. T., 1983. Wild& Feeding and Nutrition. N e w York B London: Academic Press. ROBINSON, J. G., 1986. Seasonal variation in use of time and space by the wedge-capped capuchin monkey,
Cebus olivaceus: implications for loraging theory. Smithsonian Contributions to <oology, Number 43 1. Washington, D.C.: Smithsonian Institution Press.
ROSENTHAL, G. A. & JANZEN, D. H., 1979. Herbivores, their Interaction with Secondary Plant Metabolites. New York & London: Academic Press.
SCHAIK, C. P. VAN & NOORDWIJK, M. A. VAN, 1985. Interannual variability in fruit abundance and the reproductive seasonality in Sumatran long-tailed macaques (Macaca fasicularis). Journal of ~ o o l o g y (London), 206: 533-549.
SCHOENER, 'r. W., 1969. Optimal size and specialization in constant and fluctuating environments: an energy-time approach. Brookhaven Symposium of Biology, 22: 103-1 14.
SCHOENER, T . W., 1971. Theory of feeding strategies. Annual Review of Ecology B Systematics, 2: 369-404. SIEGEL, S., 1956. Nonparametric Statistics f o r the Behavioral Sciences. New York: McGraw-Hill. SMALLWOOD, P. D. & PETERS, W. D., 1986. Grey squirrel food preferences: the effects of tannin and fat
SMITH, C. C. & FOLLMER, D., 1972. Food preferences of squirrels. Ecology, 53: 82-91. SMITH, D., 1969. Removing and anahzing total non-structural carbohydrates f rom plant tissue. Research Report No. 41.
Research Division, Wisconsin: College of Agricultural and Life Sciences, University of Wisconsin, Madison. STEPHENS, D. W. & KREBS, J . R., 1986. Foraging Theory. Monographs in Behavior and Ecology. New Jersey:
Princeton University Press. SYMES, W. F. & DAWSON, C. R., 1953. Cashew nut shell liquid. IX. The chromatographic separation and
structural investigation of the olefnic components of methyl-cardanol. Journal of the American Chemical Society, 75: 49524957.
VAN SUMERE, C. F., ALBRECHT, J., DEDONDER, A,, DEPOOTER, H. & PE, I., 1975. Plant proteins and phenolics. In J. B. Harborne and C. F. Van Sumere (Eds), Annual Proceedings sf the Phytochemical Sociely,
VILJOEN, S., 1984. Comparative thermoregulatory adaptations of southern African tree squirrels from four different habitats. South African Journal of Ecology, 20: 28-32.
WATERMAN, P. G. , 1983. Distribution of secondary metabolites in rain forest plants: towards an understanding of cause and effect. In S. L.. Sutton, T . C. Whitmore & A. C. Chadwick (Eds), Tropical Rain Forest: Ecology and Management: 167-1 79. Oxford: Blackwell Scientific Publications.
WATERMAN, P. G . , 1984. Food acquisition and processing as a function of plant chemistry. In D. J. Chivers, B. A. Wood & A. Bilsborough (Eds), Food Acqu on and Processing in Primates: 177-2 1 1. New York: Plenum Press.
WATERMAN, P. G. & CHOO, G. M., 1981. The effect ofdigestibility-reducing compounds in leaves on food selection by some Colobinae. Malaysian Applied Biology, 10: 147-162.
WATERMAN, P. G., CHOO, G . M., VEDDER, A. L. & WATTS, D., 1983. Digestibility, digestion- inhibitors and nutrients of herbaceous foliage and green stems from an African montane flora and comparisons with other tropical flora. Oecologia (Berl in) , 60: 246249.
WESTOBY, M., 1974. An analysis of diet selection by large generalist herbivores. American Naturalist, 108:
WESTOBY, M., 1978. What are the biological bases of varied diets? American .Naturalist, 112: 627-631. WHITTEN, J. E. J. & WHITTEN, A. J., 1987. Analysis of bark eating in a tropical squirrel. Biotropica, 19:
WILLAMAN, J. J. & SCHUBERT, B. G., 1961. Alkaloid-bearing Plants and their Contained Alkaloids. Technical Bulletin 1234, Washington, D.C.: United States Department of Agriculture.
WILSON, M. F., 1984. Comparison of tannin levels in developing fruit buds of two orchard pear varieties using two techniques, Fohn-Denis and protein precipitating assays. Journal of Chemical Ecology, 10: 493498.
WRANGHAM, R. W., 1981. Drinking competition in vervet monkeys. Animal Behaviour, 29: 904-910. ZUCKER, W. V., 1983. Tannins: does structure determine function? An ecological perspective. American
concentration. Ecology, 67: 168-1 74.
11: 211-264.
290-304.
107-1 15.
Naturalist. 121: 335-365.





























