Embed Size (px)
Citation preview

Segunda aula (T2)
Texto adaptado de:MOORE, J. A. Science as a Way of Knowing -Genetics. Amer. Zool. v. 26: p. 583-747, 1986.
A DESCOBERTA DA CÉLULA
O nascimento da Citologia pode ser fixado comconsiderável precisão. No dia 15 de abril de 1663,Robert Hooke (1635-1703) colocou um pedaçode cortiça sob seu microscópio e mostrou sua es-trutura a seus colegas da Royal Society de Londres.
A Royal Society havia sido fundada no anoanterior com o intuito de melhorar o conheci-mento sobre a natureza. Ela reunia uns poucoshomens cultos de Londres que se encontravamregularmente, em geral semanalmente, paradiscutir assuntos científicos e como o conheci-mento poderia ser usado para melhorar asatividades práticas. A inspiração para a formaçãoda Royal Society veio de uma sugestão anteriorde Francis Bacon.
Hooke, um matemático de excepcional habili-dade, era um membro muito ativo da Royal So-ciety. Era costume entre os membros não apenasdiscutir mas também realizar experimentos e fazerdemonstrações. Havia um grande interesse nonovo microscópio que Hooke havia construído eele deixou que os membros da sociedade olhas-
sem partes de um musgo em seu microscópio nodia 8 de abril de 1663. No dia 15 daquele mês o“ Sr. Hooke apresentou dois esquemasmicroscópicos, uma representação dos poros dacortiça, cortados transversal e perpendicular-mente ...”. Esse era o começo de dois séculos deobservações e experimentações que estabelecerama Teoria Celular.
As várias observações de Hooke foram reunidase publicadas em 1665 com o título de Micrographia,sob os auspícios da Royal Society. Essa foi a pri-meira visão geral de uma parte da natureza até entãodesconhecida. Hooke descreveu e ilustrou muitosobjetos em sua publicação: a cabeça de um alfinete,muitos insetos pequenos e suas partes, penas,“enguias” [nematódeos] do vinagre, partes de muitasplantas, cabelo, bolores, papel, madeira petrificada,escamas de peixe, seda, areia, flocos de neve, urina,e, é claro, aquele pedaço de cortiça. (Fig. 1)
Hooke imaginou que a cortiça consistia de inú-meros tubos paralelos com divisões transversais:“Estes poros, ou células, não eram muito fundos,mas consistiam de um grande número de peque-nas caixas, separadas ao longo do comprimento
14
AS ORIGENS DA BASE CITOLÓGICADA HEREDITARIEDADE
Objetivos 1. Descrever como e quando foi descoberta a célula. 2. Explicar a idéia central e a importância da teoria celular. 3. Discutir as dificuldades para se identificar os gametas
como células. 4. Descrever os passos que levaram à compreensão da im-
portância do núcleo celular. 5. Identificar as dificuldades para a compreensão do proces-
so de divisão celular. 6. Descrever o raciocínio dedutivo que levou à conclusão de
que a mitose não seria o único tipo de divisão celular. 7. Descrever as meioses masculina e feminina em Ascaris. 8. Explicar o papel da meiose e da fertilização no ciclo de
vida dos organismos. 9. Listar os principais argumentos que levaram alguns cito-
logistas no final do século XIX a defender a idéia que oscromossomos seriam a base física da herança.

dos tubos por uma tipo de diafragma.” Ele obser-vou estruturas semelhantes em muitos outrostipos de plantas. Muitos pensam que Hooke des-creveu aquelas caixas como vazias e parou poraí. Isso não é verdade, ele observou cortes deplantas vivas e verificou que as caixas microscó-picas eram preenchidas por um suco.
A presença de células na cortiça e em outrasplantas poderia ser uma característica geral oupoderia ser restrita a uns poucos tipos de organis-mo. A continuação das pesquisas iria mostrar queas plantas consistiam inteiramente ou quase intei-ramente de estruturas parecidas, semelhantes acaixas. Um outro membro da Royal Society,Nehemiah Grew (1641 - 1712), publicou umamonografia em 1682 que contém muitas pranchasbelíssimas mostrando a estrutura microscópicadas plantas. Com o tempo, a idéia de que os seresvivos são formados por células foi estendida paraos animais. Hooke havia feito uma observaçãointeressante que não foi importante na sua época– ela se tornou uma descoberta importante muitomais tarde, em função de pesquisas posteriores.
Mais de dois séculos foram necessários parase chegar à conclusão que o conhecimento dascélulas era essencial para a compreensão da here-ditariedade. Podemos ter certeza que, quandoRobert Hooke sentou-se à frente de seu micros-cópio, ele não estava interessado em descobriros mistérios da herança. Não havia maior razãopara acreditar que as células tivessem algo a vercom a hereditariedade do que, por exemplo, ascerdas que ele descreveu em detalhe sobre ocorpo de uma pulga.
O ESTABELECIMENTO DA TEORIA CELULAR
As células se tornaram verdadeiramente impor-tantes somente quando foi proposta a hipótese deque os corpos de todos os organismos eram consti-tuídos apenas de células ou de produtos de células.Essa hipótese foi formulada e testada no começodo século XIX e está associada principalmente atrês cientistas: R. J. H. Dutrochet, Matthias JacobSchleiden e Theodor Schwann.
Mas como alguém poderia provar que “oscorpos de todos os organismos são constituídosapenas de células ou de produtos de células?”
Ao tentar responder essa questão pode-seaprender algo muito importante sobre ciência. Aresposta é, obviamente, que não há nenhuma pos-sibilidade dessa afirmação ser comprovada. Comoalguém poderia estudar todos os organismos? Amaioria já se extinguiu há muito tempo e não serianem mesmo possível estudar um indivíduo decada uma das espécies viventes. Qual seria suaresposta se alguém lhe perguntasse se os corposdos dinossauros eram constituídos de células?Mas lembre-se, tudo o que se pode desejar emciência é que uma afirmação seja “verdadeiraacima de qualquer suspeita.”
Após as observações iniciais de Hooke, foiverificado que as células eram uma caracte-rística comum das plantas. Mais e mais plantasde uma quantidade crescente de espécies fo-ram estudadas e todas apresentavam estruturassemelhantes a células. Foi observado que essasestruturas microscópicas não tinham todas aforma de caixa como as células da cortiça.Descobriu-se que as células podiam ter diversasformas e tamanhos. Não podemos esquecer queesses microscopistas pioneiros não estavamobservando células como as entendemos hoje,eles observavam paredes celulares.
Schwann e as células nos animais
Com poucas exceções, o corpo dos animaisnão continha estrutura alguma que se parecessecom “células”, isto é, com as paredes celularesdas plantas. Assim, foi necessário muito trabalhoe imaginação arrojada até tornar óbvio que o con-ceito de célula podia ser aplicado com sucessoaos animais. Isso foi conseguido principalmentepor Theodor Schwann (1810-1882) em suamonografia de 1839, publicada quando ele tinha29 anos de idade. Algumas de suas ilustraçõesestão reproduzidas na figura 2.
15
Figura 1. Desenhos de cortes de cortiça ao micros-cópio publicados por Hooke em 1665.

Figura 2. Algumas das ilustrações apresentadas por Schwann em sua mono-grafia de 1839: A.) células de cebola; B.) de notocorda de um peixe; C.)de cartilagem de rã; D.) de cartilagem de girino; E.) de músculo de feto deporco; F.) de embrião de porco; G.) de gânglio de rã; H.) de um vasocapilar da cauda de girino; I.) de embrião de porco. Note que o núcleo e osnucléolos estão mostrados em quase todas as células.
Schwann enfatizou a gran-de diferença entre as célulasdas plantas e o que ele acredi-tava serem as células dosanimais, mas sugeriu que elasrepresentavam fundamental-mente a mesma coisa.
Por que chamar todas essasestruturas tão diversas decélulas?
Procure examinar fotomi-crografias de diversos tipos decélulas de plantas e especial-mente de animais, ou melhor,caso tenha oportunidade, ob-serve preparações citológicasdesses tipos no microscópio.Como é possível dizer quecérebro, músculos, rins, pul-mões, sangue, cartilagens,ossos, parede intestinal etc.são feitos de um mesmo tipode elemento? Já que essasestruturas são obviamente tãodiferentes, por que afirmarque elas são constituídas pelosmesmos tipos de elementos?Qual seria a vantagem em se afirmar que as“células” animais correspondiam àquelasestruturas com aspecto tão diferente presentesnas plantas?
Schwann nos fornece a resposta, “Se, noentanto, analisarmos o desenvolvimento dessestecidos, então parece que todas essas diversasformas de tecidos são constituídas apenas porcélulas e são análogas às células das plantas ...O objetivo do presente tratado é provar essa idéiapor meio da observação.”
Isto é, apesar da grande diversidade, todas asestruturas que Schwann propunha chamar decélulas tinham em comum a característica de sedesenvolverem a partir de estruturas muito maissimples que podiam ser melhor comparadas comas células das plantas. Mas, como se poderiadefinir “célula”?
Se um neurônio e um leucócito são células, elesdevem ter algo em comum para serem reunidosem uma mesma categoria. Schwann encontrouum critério: a presença de núcleo, que ele achava
mais importante do que a origem de células alta-mente diferenciadas a partir de células simples.
Apenas seis anos antes, em 1833, Robert Brown(1773 - 1858), o mesmo que descreveu o poste-riormente denominado “movimento Browniano”,havia descrito a presença de uma auréola circular,ou núcleo, em células de orquídeas e de muitosoutros tipos de plantas. Antes dele, outros observa-dores já haviam visto e desenhado essas estruturasem suas publicações, mas não atribuíram nenhumaimportância a elas. Brown verificou que muitos tiposde célula continham núcleo mas não especulou sobreseu significado.
Schwann então mudou as regras para definircélula. Ao invés de se basear na forma, que nasplantas correspondia à estrutura da parede, eleescolheu como base para a definição, a presençade um núcleo.
Embora Schwann fosse um observador cuida-doso, sua principal contribuição não foi o queele viu mas como ele interpretou as observações.Seus antecessores haviam enfatizado as “caixas”;
16
A B C
D
H
EF
I
G

Schwann deu ênfase ao que estava dentro das“caixas”. Para ele a célula animal era uma porçãode matéria viva envolta por uma membrana econtendo um núcleo, enquanto que as célulasvegetais eram ainda envoltas por uma parede.
O que essa nova visão de célula tem a ver comhereditariedade? Muito pouco, tem que se admitir.Seriam necessárias outras duas informações antesque as células pudessem ser consideradas impor-tantes para a hereditariedade: a descoberta de queos gametas são células e o reconhecimento de quecélulas só se originam de células pré-existentes.
O reconhecimento dos gametas como células
Schwann reconheceu os óvulos como células,uma vez que eles apresentavam a estrutura reque-rida por sua definição de célula – o núcleo. Anatureza do espermatozóide era menos clara. Seunome, que significa “animais do esperma”, indica-va essa incerteza. Em 1667, Antonie van Leeu-wenhoek havia descoberto e comunicado à RoyalSociety de Londres que o fluido seminal continhacriaturas microscópicas que ele imaginou queentrassem no óvulo causando sua fertilização.Essa hipótese foi muito contestada e alguns cien-tistas imaginaram que os espermatozóides fossemparasitas. Na décima segunda edição do livroSystema Naturae (1766 - 1768), Linnaeus tentouclassificar os “animais” encontrados no espermapor Leeuwenhoek, mas concluiu que a determi-nação de seu lugar correto no sistema de clas-sificação deveria ser deixado para quando elestivessem sido mais pesquisados.
Cerca de um século mais tarde, em 1784, Spal-lanzani realizou importantes experimentos com oobjetivo de determinar a função do sêmen nareprodução de rãs. Durante o acasalamento, osmachos abraçam as fêmeas e, como sabemos atual-mente, depositam esperma sobre os óvulos à medidaque estes saem pela abertura cloacal. De início,Spallanzani não sabia disso, foi ele quem descobriu.Um outro pesquisador com quem ele se corres-pondia havia tentado, sem muito sucesso, descobriro papel das rãs machos vestindo-as com calças. Spal-lanzani repetiu esse experimento e verificou que,quando o sêmen ficava retido nas calças, os ovosnão se desenvolviam. No entanto, se os ovos fossemcolocados em contato com o sêmen retirado dascalças, em um processo de fecundação artificial, elespassavam a se desenvolver. Em um outro experi-
mento, Spallanzani filtrou o sêmen e verificou que,com isso, ele perdia seu poder fecundante. Ele obser-vou o que hoje chamamos de espermatozóides, masnão os considerou essenciais para a reprodução.
Foi somente em 1854 que George Newport,usando rãs, forneceu boas evidências de que osespermatozóides entram no óvulo durante a fecun-dação (Nesse caso, como em muitos outros, é difícildar crédito ao cientista que descobriu um importantefenômeno biológico. Afinal, o descobridor do esper-matozóide, Leeuwenhoek, havia pensado que oespermatozóide era o agente da fertilização. Outrosantecessores de Newport eram da mesma opinião,mas foi Newport quem fez as primeiras observaçõesconvincentes. Em 1841, Kölliker estudou ahistologia dos testículos verificando que algumasdas células testiculares eram convertidas em esper-matozóides. Os espermatozóides tinham umaaparência tão estranha que não eram consideradoscélulas. No entanto, quando se pôde demonstrarque eles se originavam de células típicas, suaverdadeira natureza tornou-se evidente. Os esper-matozóides passaram então a ser considerados comocélulas altamente modificadas.
Vejamos o que se pode concluir dessa análise:1. Os gametas são a única ligação física entre as
gerações, pelo menos em muitos organismose possivelmente em todos.
2. Portanto, os gametas devem conter toda ainformação hereditária.
3. Uma vez que óvulos e espermatozóides sãocélulas, toda informação hereditária precisaestar contida nestas células sexuais. Portanto,a base física da herança são as células sexuais.
Isto não permite concluir que todas as célulascontenham informação hereditária. Poderíamosainda pensar que os gametas são célulasespecializadas onde os fatores responsáveis pelaherança, talvez as gêmulas, entram. Nós aindanecessitamos de uma segunda informação: “Qualé a origem das células?”
Omnis cellula e cellula
A divisão celular foi observada em 1835, mas,nessa época, não se concluiu que fosse um fenômenogeral. Schwann, por exemplo, acreditava que ascélulas podiam surgir espontaneamente por agluti-nação de substâncias amorfas. Essa hipótese assumiaque a origem das células é um evento episódico no
17

ciclo de vida dos organismos. Se isso fosse verdade,a unidade da hereditariedade seria o organismo todoe não a célula.
A hipótese de Schwann sobre a origem dascélulas foi logo rejeitada, uma vez que a divisãocelular estava sendo observada com freqüênciaem uma variedade de organismos e em diferentesépocas do desenvolvimento. Mais e mais investi-gadores começavam a suspeitar que a divisãocelular era o único mecanismo para a produçãode novas células.
Essa foi uma hipótese muitíssimo difícil de seprovar acima de qualquer suspeita. Os micros-cópios e as técnicas para se estudar as células, nocomeço do século XIX, eram muito inadequadose foi preciso muita observação em diferentes tiposde organismos e de tecidos antes que RudolphVirchow pudesse, em 1855, cunhar sua famosafrase omnis cellula e cellula (“toda célula vemde célula”) e que ela fosse amplamente aceita.Em uma conferência proferida em 1858 ele apre-sentou a idéia de que uma célula só surge de outracélula pré-existente.
É claro que nem todos concordaram com aidéia de Virchow de que todas as células e todosos organismos originavam-se de células e de orga-nismos pré-existentes. Muitos pesquisadorescontinuavam a acreditar que células podiam seoriginar de novo e apresentavam o que pareciamser observações acuradas para provar isso. Algunsacreditavam até mesmo que organismos comple-tos podiam se originar de novo. Pasteur e a acei-tação geral de que geração espontânea não podeocorrer ainda estavam no futuro. Mesmo assim,as duas hipóteses apoiadas por Virchow foramtestadas em um número crescente de pesquisase, gradualmente, elas se estabeleceram como umaverdade acima de qualquer suspeita.
Não restando, portanto, dúvida alguma de quea hereditariedade está baseada na continuidadecelular, podemos trabalhar agora com a hipótesede que toda informação hereditária está contidanão apenas nas células germinativas mas também,muito provavelmente, nas células a partir dasquais elas se formam – e em todas elas até ozigoto. Igualmente possível é a hipótese de quetodas as células contenham a informação heredi-tária necessária para o desenvolvimento do indiví-duo e à sua transmissão, via células sexuais, paraa geração seguinte.
O DESENVOLVIMENTO TECNOLÓGICO
E A CITOLOGIA
Durante a maior parte da história da humani-dade as pessoas se basearam quase que inteira-mente em seus órgãos dos sentidos para obterinformações sobre o ambiente. Cada um de nossosórgãos sensoriais detecta apenas uma pequenaporção da ampla gama de estímulos possíveis.Nossos olhos, por exemplo, só conseguem res-ponder à porção do espectro eletromagnéticoentre o violeta e o vermelho de modo que sóconseguimos ver os comprimentos de onda en-tre essas duas cores. Para detectar comprimentosde onda menores, como luz ultra-violeta, raios-X e raios cósmicos, ou comprimentos de ondamaiores, como luz infra-vermelha e ondas derádio, precisamos utilizar instrumentos especiais.
A olho nu não conseguimos visualizar emdetalhe nem mesmo objetos em movimentaçãorápida. As lâminas de um ventilador em movi-mento rápido são vistas como um círculo contínuoe uma bala que sai de um rifle é totalmente invisí-vel para nós. Também não conseguimos verobjetos muito pequenos. A aparente uniformi-dade de uma ilustração com meios-tons resultado fato de os pontos individuais estarem tão jun-tos que o olho humano não consegue distingui-los. Os faróis de um automóvel aparecem comoum único ponto de luz até uma certa distância; àmedida que o automóvel se aproxima, somoscapazes de resolver o ponto único de luz em dois.
O poder de resolução do olho humano, ou seja,sua capacidade de distinguir dois pontos muitopróximos, é da ordem de 100 micrômetros a umadistância normal de leitura; a maioria das pessoascom visão normal distingue dois objetosseparados por um espaço de um milímetro a umadistância de 10 metros. Uma afirmação mais geralé que o olho humano pode distinguir dois objetosseparados por um arco de 1 minuto. Esse valorfoi determinado por Robert Hooke que estavapreocupado em saber qual seria a menor distânciaentre duas estrelas para que elas fossem vistascomo dois objetos separados. Quando elasestavam a uma distância menor do que um arcode 1 minuto, a maioria das pessoas as via comoum único ponto de luz. Algumas pessoas podemver melhor do que isso, mas o poder máximo deresolução de nosso olho é de 26 segundos de arco.
18

Quase todas as células são muito pequenaspara serem vistas a olho nu, de modo que a Cito-logia não foi possível, nem mesmo teoricamente,antes da invenção do microscópio – o que ocorreuprovavelmente na última década do século XVI.Um tempo relativamente longo se passou entreessa invenção e 1663, quando Hooke fez ademonstração daqueles pedaços de cortiça paraos membros da Royal Society. Na verdade, fo-ram poucos os trabalhos sérios e contínuos commicroscópios antes do século XIX. Durante amaior parte de sua história, os microscópios nãopassaram de brinquedos de adultos.
O pequeno tamanho das células não é o únicoproblema que dificulta seu estudo. A maioria dosanimais e de seus tecidos é opaca e, uma vez quea observação através do microscópio compostoé mais efetiva quando os objetos são iluminadoscom luz transmitida, o objeto a ser estudadoprecisa ser muito fino ou cortado em fatias bemfinas de modo que a luz possa atravessá-lo. Ima-gine tentar cortar fígado, por exemplo, em fatiascom cerca de 10 micrômetros de espessura, paraque fosse possível estudá-lo no microscópio.Além da quase impossibilidade de se fazer isso,as células hepáticas, constituídas principalmentede água, iriam secar rapidamente, tornando-seuma massa enrugada. Esse é um problema espe-cial com as células animais que não possuem umaparede de suporte como as células das plantas.
Métodos muito especiais tiveram que serdesenvolvidos pelos microscopistas do início doséculo XIX quando eles quiseram aprender sobrea natureza celular dos organismos e, mais tarde,sobre a estrutura interna das próprias células.Tornou-se uma prática comum tentar preservaros tecidos de tal maneira que a estrutura de suascélulas permanecesse intacta e que eles pudessemser cortados em fatias finíssimas.
O primeiro passo foi a fixação. Ela consistiaem tratar o material com álcool, com formaldeído,ou com soluções de ácido pícrico, de bicromatode potássio, de cloreto de mercúrio ou de tetró-xido de ósmio. Essas substâncias matam eendurecem as células, em geral, por coagular suasproteínas. Esperava-se, é claro, que isso aconte-cesse de tal forma que as partes das células conti-nuassem a guardar uma certa semelhança com asda célula viva.
O tecido fixado podia então ser embebido emparafina para ganhar sustentação e ser fatiado com
lâminas cortantes ou em um instrumentoconstruído para essa finalidade – o micrótomo.Mesmo essas fatias finíssimas podiam revelarmuito pouco; as células e seus conteúdos internoseram indistinguíveis. Mas aqueles microscopistasinventivos tentavam de tudo e logo descobriramque alguns corantes tingiam certas partes dascélulas mas não outras.
Em 1858, Gerlach descobriu que uma soluçãodiluída de carmim corava mais intensamente onúcleo do que o citoplasma das células. Essasubstância era obtida dos corpos secos da fêmeade um inseto (Coccus cacti), conhecido popular-mente como cochonilha-do-carmim, que vive emcactos na América Central e sudoeste dos EstadosUnidos. Em 1865, Böhmer descobriu que ahematoxilina, extraída do tronco de uma árvore(Haematoxylon campechianum) da América Cen-tral, também tinha maior afinidade pelo núcleodo que pelo citoplasma.
Mais tarde foi sintetizada uma grandevariedade de anilinas para a indústria téxtil e, entre1875 e 1880, muitas delas mostraram-se úteispara corar células. Uma dessas anilinas era aeosina, que mostrou ter uma grande afinidade porproteínas citoplasmáticas. Um procedimento decoloração citológica corriqueiro até hoje usahematoxilina e eosina (HE) e cora o núcleo emazul e o citoplasma em laranja.
Da mesma forma, houve melhoria dos micros-cópios disponíveis para pesquisas citológicas, princi-palmente no final do século XIX. Muitas delas fo-ram introduzidas por Ernst Abbe (1840-1905) e pelaindústria óptica Zeiss em Jena, na Alemanha. Abbefoi, durante a maior parte de sua vida, professor deFísica na Universidade em Jena e o principalprojetista de lentes da companhia Zeiss, da qual setornou dono. Em 1878, ele desenvolveu a objetivade imersão em óleo e, em 1886, a objetiva apocro-mática. Essas melhorias nas mãos de um microsco-pista habilidoso tornava possível a obtenção deampliações de até 2500 vezes. O microscópiofotônico estava chegando ao limite de seu poder deresolução teórico. Esse limite é imposto pela próprianatureza da luz; isto é, dois objetos só podem serresolvidos se a distância entre eles for, pelo menos,igual à metade do comprimento da onda utilizado.
Oportunidades adicionais para se estudar aestrutura fina das células estavam ainda para vircom a invenção do microscópio de contraste-de-fase e do microscópio eletrônico, no século XX.
19

Veremos a seguir como os microscopistas do últimoterço do século XIX foram capazes de usar atecnologia disponível na época e estabelecer, comouma hipótese altamente provável, que a base físicada hereditariedade está no núcleo da célula, ou maisespecificamente nos cromossomos.
Não devemos ficar imaginando que essesinvestigadores não faziam outra coisa senãoexaminar células vivas e fixadas com o melhorequipamento óptico disponível, descrevendo domodo mais preciso o que viam. Um problemaconstante era se uma dada estrutura observadaem uma preparação citológica refletia ou não algopresente na célula viva, ou se era um simples arte-fato resultante do drástico tratamento a que ascélulas eram submetidas para poderem serobservadas no microscópio.
Uma preparação citológica realmente refletea estrutura de uma célula viva? A resposta é “Nãomuito”; mas se o tratamento produz sempre omesmo resultado é possível imaginar como eramas preparações quando vivas. Apesar disso,nenhuma descoberta importante em Citologia noséculo XIX foi aceita de imediato. As observaçõeseram repetidas e as conclusões originais confir-madas por uns e contestadas com veemência poroutros. Uma interpretação errada podia fazer comque muitos citologistas perdessem meses natentativa de repetir as observações. Aconteciamdebates intermináveis sobre a estrutura fina doprotoplasma uma vez que, como era admitido,estava-se olhando para a base fundamental davida. Muitos citologistas acreditavam que oprotoplasma fosse granular, ou um retículofibroso, ou alveolar (composto de gotas) oualguma combinação disso.
A Citologia como um caminho para o conheci-mento, especialmente no século XIX, nos mostraque a ciência não progride de maneira ordenadamas por meio de testes e retestes constantes dasobservações, dos experimentos e das hipóteses.Longe de ser uma linha direta em direção àverdade, esse caminho assemelha-se mais àqueleretículo que alguns viam como a estrutura básicado protoplasma. (Devemos ressaltar que o termoprotoplasma é raramente utilizado nos dias dehoje. Uma vez que ele significa nada mais do que“substância viva”, Hardin [1956] sugeriu quepoderíamos passar sem ele.)
O QUE EXISTE NAS CÉLULAS?
Durante a última metade do século XIX, ahipótese de que os animais e plantas sãocompostos somente de células e produtoscelulares estava estabelecida como uma verdadeacima de qualquer suspeita nas mentes damaioria dos microscopistas competentes. Nóspodemos falar, então, da teoria celular, usando otermo “teoria” como um corpo completo dedados, hipóteses e conceitos relativos a um impor-tante fenômeno natural. Até hoje a teoria celularé o mais importante conceito relacionado com aestrutura de animais e plantas e no século XX elefoi sendo gradualmente aceito também como omais importante conceito relativo ao funciona-mento dos organismos.
Essa enorme importância da teoria celulardecorre do fato de ela estabelecer que as célulassão as unidades básicas de estrutura e função,que elas são as menores unidades capazes de tervida independente, isto é, são capazes de usarsubstâncias obtidas do meio para manter eproduzir o estado vivo. A célula é o denominadorcomum da vida.
Existia uma outra razão importante para seestudar as células: a análise dos níveis mais sim-ples de organização contribuem para o enten-dimento dos níveis mais complexos. As interaçõesdas substâncias químicas são melhor entendidasquando se conhece sua estrutura molecular. Osmovimentos do corpo humano podem serestudados em muitos níveis. Pode-se observar edescrever os complexos movimentos de umbailarino ou de um arremessador de beisebol. Acompreensão aumenta quando se obtéminformações sobre os diversos músculos e seuslocais de ligação, que tornam os movimentospossíveis. Outros tipos de entendimento surgemquando se estuda os músculos no nível molecu-lar. E, finalmente, mais informações ainda sãoobtidas quando se aprende sobre a atividade damiosina, da actina e de outras moléculas que parti-cipam da movimentação dos músculos.
O conhecimento obtido em cada nível deorganização contribui para um entendimento dofenômeno como um todo, enquanto cada nívelmantém seu próprio valor. Entender a arte de umbailarino ou de um esportista meramente com o
20

conhecimento sobre actina e miosina seria tãoimpossível quanto predizer as propriedades da águaa partir do conhecimento sobre os elementoshidrogênio e oxigênio. No entanto, pode-se conhecermelhor os níveis mais complexos se conhecermosos mais simples. Assim, os biólogos do século XXjá pensavam que poderiam saber mais sobre a vidase conhecessem melhor as células.
Quando examinavam as células, aqueles citolo-gistas pioneiros encontravam todo tipo de esferas,de grânulos e de fibras. Como seria possíveldeterminar qual dessas estruturas teria um papelna hereditariedade? Ou melhor, como seriapossível determinar a função de qualquer estru-tura presente nas células?
Essa é uma questão difícil e os citologistasdaquela época não conseguiam respondê-la. Elesnão podiam fazer outra coisa senão investigar ascélulas de modo aleatório. Este foi um estágionecessário no desenvolvimento da Citologia – aidentificação de estruturas nas células e, quandopossível, descobrir alguma coisa sobre seucomportamento. Aparentemente se pesquisavamcélulas de todo animal e planta disponível àprocura de exemplos de estruturas celulares e,um a um, todos os reagentes disponíveis nasestantes dos químicos foram colocados sobre ascélulas e suas conseqüências observadas – emgeral matavam as células. Esse período da Cito-logia foi de “procura e destruição.”
O núcleo efêmero
Como mencionado anteriormente, as dificul-dades em se analisar células vivas fizeram daspreparações fixadas e coradas o material ideal deestudo. Nesse tipo de preparação, a estrutura maisproeminente é o núcleo descrito por Brown.Muitos corantes, especialmente os corantesbásicos como o carmim e a hematoxilina, coravamo núcleo profundamente; isto, juntamente com aaparente presença universal do núcleo, sugeriaque ele tivesse um papel importante.
Mas qual seria a origem do núcleo da célula?Levou mais de meio século de observações eexperimentações por parte de numerosos pesqui-sadores para que essa questão fosse respondida.Em 1835, Valentin sugeriu que o núcleo seriaformado pela precipitação de substâncias no in-terior da célula. Três anos mais tarde, Schleidene, em seguida, Schwann também sugeriram queo núcleo podia se originar de novo. Até por volta
de 1870, alguns pesquisadores famosos acre-ditavam que pelo menos alguns núcleos podiamter uma origem não-nuclear.
Nessa mesma época, outros pesquisadoresigualmente competentes estavam clamando quetodos os núcleos surgiam de núcleos pré-existentes. Diversos processos foram sugeridos– em geral alguma forma de partição em dois oufragmentação, um mecanismo que mais tarde foidenominado amitose.
Não havia nenhuma razão, é claro, porque osnúcleos teriam de surgir por apenas um tipo demecanismo. Considerando a enorme variedade defenômenos naturais, não seria surpresa sehouvesse diversas maneiras de surgimento denúcleos. No entanto, os cientistas procuram regu-laridades na natureza e seria mais satisfatóriointelectualmente se houvesse um mecanismoconstante para a origem do núcleo.
A DESCOBERTA DA DIVISÃO CELULAR
Em 1873, A. Schneider publicou o que agorapode ser tomado como a primeira descriçãorazoável das complexas alterações nucleares, hojechamadas de mitose, que ocorrem durante adivisão da célula. Neste ano, Otto Bütschi e Her-mann Fol fizeram descrições semelhantes.
A descrição de Schneider foi a mais completa;seu objetivo era descrever a morfologia deMesostoma sp., um platelminto. Quase todo seutrabalho é dedicado à estrutura desse verme mas,sendo um observador cuidadoso, ele descreveutudo o que viu. A fertilização em Mesostoma sp.é interna e o início do desenvolvimento ocorreem um útero. As ilustrações do que ele viu estãomostradas na figura 3.
Os primeiros desenhos mostram o ovorodeado por células foliculares. Na região bemcentral está o pequeno núcleo com seu pequenonucléolo. As estruturas espirais são espermato-zóides. O ovo é a área clara central da ilustraçãoe os glóbulos menores ao seu redor são as célulasfoliculares, que não foram representadas nosdesenhos seguintes. Pouco antes da célula se divi-dir o limite do núcleo se torna indistinto. Schnei-der, no entanto, verificou que com a adição deum pouco de ácido acético ele se tornava visível,apesar de dobrado e enrugado. Mais tarde onucléolo desaparecia e tudo o que restava do nú-cleo era uma área clara na região central da célula.
21

No entanto, o tratamento com ácido acético mos-trava uma massa de filamentos delicados e curvos.O segundo desenho mostra esses filamentos, oscromossomos (um termo que só seria propostoem 1888, por Waldeyer) alinhados em uma placaequatorial. A quantidade de filamentos parecia
aumentar e quando a célula se dividia eles iampara as células-filhas.
O que alguém faria com essas observações?A resposta está longe de ser clara. Se não erapossível ver os filamentos nas células vivas e, seeles apareciam repentinamente quando as células
22
Figura 3. Ilustrações de Schneider (1873) das alterações nucleares durante a clivagem do ovode Mesostoma. À esquerda, desenho de um ovo (zona clara central, onde se vê o núcleo comum nucléolo) rodeado por células foliculares. As outras figuras mostram os filamentos, hojechamados de cromossomos, e seus movimentos durante a divisão da célula.
Figura 4. Ilustrações deFlemming de mitoses emcélulas fixadas e coradas deembrião de salamandra. A.)Duas células em intérfase:não existem cromossomosvisíveis. B.) Célula em pró-fase: os nucléolos já desapa-receram, mas a membrananuclear continua intacta; ocitoplasma não está mostra-do. C.) Célula em início demetáfase: a membrana nu-clear desapareceu e os cen-trossomos se separaram. D.)Uma preparação deexcelente qualidade, onde sevê os cromossomos meta-fásicos duplos, isto é, com-postos por duas cromátides.E.) As cromátides se sepa-ram e se movem para ospólos do fuso. F.) Célula emfinal de divisão com oscromossomos dos núcleos-filhos sendo envolvidos pelamembrana nuclear. (Flem-ming, 1882)
A B
C D
E F

eram tratadas com ácido acético, não seriarazoável pensar que eles fossem um artefato? Noentanto, o fato de os filamentos serem obser-vados repetidamente, e de eles parecerem sofrerestranhos movimentos, sugeria que já estivessempresentes na célula viva, numa forma invisível.Flemming teve sucesso em determinar que oseventos nucleares observados na célula em divisãoem materiais fixados e corados tinham suacontrapartida na célula viva. Apesar de não terdescoberto a mitose, devemos a ele mais do quea qualquer outro o conceito de mitose que temoshoje; apenas detalhes do processo foram adicio-nados à sua descrição. (Fig. 4 e 5)
O sucesso de Flemming foi conseguido graçasa alguns fatores: material que ele selecionou paraseu estudo; ter sido cuidadoso em procurar nascélulas vivas as estruturas observadas nas célulasfixadas e coradas; ter à sua disposição microscó-
pios muito melhores do que os existentes ante-riormente. O uso de células vivas, além de dar aconfiança de que o observado era real e nãoartefato, permitiu também determinar a seqüênciados eventos.
As fases da mitose
Costuma-se dizer que um núcleo que não estásofrendo divisão encontra-se em repouso. Esseé um termo infeliz pois sugere inatividade e hojenós sabemos que a maior atividade fisiológica donúcleo acontece durante esse período. Flemmingnão viu cromossomos nos núcleos em “repouso”de células vivas. Esses núcleos pareciam não ternenhuma estrutura interna. Quando essas célulaseram fixadas e coradas via-se que seus núcleoscontinham uma rede densa com grande afinidadepor certos corantes, além de um ou dois grânulosesféricos, os nucléolos.
Figura 5. Ilustrações de Flemming de mitose em células vivas de larva de salamandra. Os desenhos estãoorganizados em seqüência, começando com a prófase, no canto superior à esquerda, e terminando com duascélulas, na fileira inferior. Os dois últimos desenhos mostram os cromossomos vistos do pólo da célula e umatelófase em vista lateral, respectivamente. O desenho mais à direita na segunda fileira mostra que oscromossomos estão duplos. (Flemming, 1882)
23

Mudanças no núcleo são as primeiras evidên-cias que a mitose está a caminho. No núcleo vivo,aparentemente desprovido de estruturas, apare-cem longos e delicados fios. Quando eles podemser vistos, é o começo da prófase. (A mitose éum processo contínuo; ela é dividida em fasespelos citologistas apenas com o intuito de facilitarsua descrição.) Esses fios se condensam emcromossomos que se posicionam no meio dacélula na metáfase, época em que a membrananuclear já desapareceu. Em células coradas pode-se ver que os cromossomos estão presos a umelaborada estrutura fibrosa – o fuso. Célulascoradas podem mostrar também a presença deminúsculos grânulos nas extremidades do fuso –os centríolos. Elas podem mostrar também umoutro conjunto de fibras, os raios astrais, queirradiam dos centríolos. Durante a anáfase dascélulas vivas, os cromossomos se separam emdois grupos que se movem através do fuso parapólos opostos da célula. Quando os cromossomosatingem as extremidades do fuso, é a telófase.Os cromossomos nas células vivas se tornam cadavez menos distintos e a membrana nuclear serefaz. O núcleo está de novo em “repouso”. Oque se pode concluir desse processo?
É óbvio que todas as estruturas celularesprecisam ser reproduzidas para que as células-filhas sejam idênticas à célula-mãe. Flemming foicapaz de explicar como isso acontece para oscromossomos. Se os cromossomos de uma célulavão ser divididos igualmente entre as células-filhas, eles precisam dobrar em número em algumestágio do ciclo celular. Flemming observou que,quando os cromossomos aparecem pela primeiravez no início da prófase eles já estão duplos;assim, em algum momento entre seu desapareci-mento na telófase e seu reaparecimento na pró-fase, cada cromossomo deve ter se duplicado.
Hoje, é claro, nós consideramos os cromos-somos como estruturas permanentes nas célulasmesmo sendo eles visíveis apenas na mitose. Nóstambém reconhecemos a individualidade doscromossomos, isto é, que eles existem em geralem pares homólogos, cada par contendo umconjunto específico de genes. Essas conclusõespoderiam ter sido tiradas a partir das observaçõesde Flemming? Na verdade não. E as hipóteses aseguir, poderiam ser refutadas?
Você poderia argumentar o seguinte: como oprocesso mitótico assegura que cada célula-filha
receba seu lote de cromossomos isto deve indicar,sem muita dúvida, que um mecanismo tão elabo-rado e preciso para duplicação e distribuição éde importância fundamental. E o que pode sermais importante do que assegurar que os ele-mentos controladores da hereditariedade e da vidade cada célula cheguem até elas?
Mas alguém pode responder que, sendo ascélulas-filhas idênticas à célula-mãe, todos osprodutos celulares são reproduzidos. Pode-seargumentar que é mero acidente que o processode reprodução e distribuição seja mais facilmenteobservado nos cromossomos. Não existe razão,portanto, para não assumirmos que cromosso-mos, membranas celulares e todos aquelesgrânulos e glóbulos observados no citoplasmapossam ter igual chance de estarem envolvidosna hereditariedade.
A DESCOBERTA DA MEIOSE
Flemming e muitos outros citologistas seuscontemporâneos estavam considerando que asdivisões mitóticas do núcleo aconteciam em todadivisão celular. A reunião de inúmeras obser-vações em células de um grande número deespécies de plantas e animais permitia que sefizesse esta afirmação geral. Note que isso é umbom exemplo de indução.
Nós podemos agora usar essa afirmaçãogeral como uma hipótese a ser testada. Isto é,nós podemos partir para um raciocínio dedu-tivo. Por exemplo: se a hipótese de que onúcleo sempre divide por mitose for verdadeira,então o número de cromossomos deve dobrara cada geração. Isso seria inevitável. Como osnúcleos do óvulo e do espermatozóide se unemna fertilização, caso eles se formassem pormitose, o zigoto deveria ter duas vezes onúmero de cromossomos de seus genitores.
Mas isso não acontece: Flemming e outroscitologistas estavam cientes de que o número decromossomos parecia ser o mesmo em todos osindivíduos e em todas as gerações de uma espécie.
Obviamente existe um problema com essahipótese. Deveria haver algum mecanismo quereduziria o número de cromossomos antes oudurante a fertilização. Seria possível supor que,quando os núcleos do óvulo e do espermatozóidese fundiam na fertilização, os cromossomos tam-bém se fundiriam uns com os outros, sendo quemetade de cada um deles seria destruída. Uma
24

hipótese alternativa é que ocorresse redução donúmero de cromossomos durante a formação dosóvulos e dos espermatozóides nas gônadas.
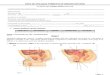
O significado dos corpúsculos polares.
Vários pesquisadores tinham descrito, emdiversas espécies animais, a eliminação de minús-culas esferas na região do pólo animal do óvulo,por ocasião da fertilização. Essas esferas logodesapareciam e, como pareciam não ter funçãoalguma, foram denominadas corpúsculos polares.Observou-se também que na partenogênese for-mava-se um único corpúsculo polar, mas que nosóvulos fertilizados eles sempre pareciam ser dois.Em algumas espécies, um corpúsculo era formadoantes da fertilização e um segundo, depois daentrada do espermatozóide. Em outras espécies,os dois corpúsculos polares eram formados apósa fertilização. (Fig. 6)
Em 1887, August Weismann propôs umahipótese para explicar a constância da quanti-dade de material hereditário de uma geraçãopara outra. Com base na observação de muitoscitologistas, ele diz: “pelo menos um certoresultado sugere que exista uma substânciahereditária, um material portador detendências hereditárias, e que esta substância
está contida no núcleo das células germinativas,no filamento enovelado no interior do núcleo[alguns citologistas pensavam que oscromossomos formavam um fio contínuo ouespirema durante a interfase], que em certosperíodos aparece na forma de alças ou barras[estes eram os cromossomos nos estágiosmitóticos]. Nós podemos, além disso, considerarque a fertilização consiste no fato de um númeroigual de alças [cromossomos] de cada genitorser colocado lado a lado, e que o núcleo do zigotoé composto desta maneira. No que diz respeito aesta questão, não tem importância se as alças[cromossomos] dos dois pais se misturam maiscedo ou mais tarde ou se permanecem separadas.A única conclusão essencial necessária à nossahipótese é que a quantidade de substânciahereditária fornecida por cada um dos genitoresseja igual ou aproximadamente igual entre si.Se for assim, as células germinativas dosdescendentes conterão os germoplasmas de am-bos os pais unidos, isso implica que tais célulassó podem conter metade do germoplasmapaterno, como estava contido nas células ger-minativas do pai, e metade do germoplasmamaterno, como estava contido nas célulasgerminativas da mãe.”
25
Figura 6. Ilustrações da meiose em fêmea de Ascaris. Anteriormente às etapas mostradas nos desenhos, oscromossomos haviam se duplicado e se emparelhado, formando duas tétrades. Estas se separaram na primeira divisãomeiótica e duas díades foram para o primeiro corpúsculo polar, enquanto que as outras duas permaneceram nosóvulos. Isto é mostrado no desenho 33 de Boveri. No desenho 36, as díades estão em rotação antes da sua separaçãona segunda divisão meiótica. O segundo corpúsculo polar pode ser visto na posição correspondente à das duas horas.Os desenhos 42 e 43 mostram as díades se separando. No desenho 45, a segunda divisão já terminou e o segundocorpúsculo polar com seus dois cromossomos aparece na superfície do óvulo; o primeiro corpúsculo polar está acimadele. Os dois cromossomos no óvulo estão para formar o pró-núcleo feminino. O desenho 46 mostra o primeirocorpúsculo polar na posição correspondente à das 3 horas, o segundo corpúsculo polar na superfície do óvulo, naposição correspondente à das 12 horas, o pró- núcleo.
Fig. 33 Fig. 36 Fig. 42
Fig. 43 Fig. 45 Fig. 46

Weismann acreditava que a redução à metadedo material hereditário da mãe, necessária à suahipótese, ocorria quando o segundo corpúsculopolar era formado. Diz ele: “Minha opinião sobreo significado do segundo corpúsculo polar é, empoucas palavras, esta: a redução do germo-plasma acontece na sua formação, uma reduçãonão só em quantidade, mas sobretudo na comple-xidade de sua constituição. Por meio da segundadivisão nuclear, evita-se o acúmulo de diferentestipos de tendências hereditárias ou germoplas-mas que, sem isso, seria necessariamente produ-zido em excesso pela fertilização. Com o núcleodo segundo corpúsculo polar são removidos doóvulo tantos tipos diferentes de idioplasmas [umtermo usado na época para designar o materialhereditário] quantos serão posteriormenteintroduzidos pelo núcleo do espermatozóide;assim, a segunda divisão do núcleo do óvuloserve para manter constante o número dediferentes tipos de idioplasmas, que compõem ogermoplasma durante o curso das gerações.”
E, se a constância é mantida de geração ageração, Weismann supõe que um processo simi-lar precisaria ocorrer no macho. Ele diz: “Se onúmero de germoplasmas ancestrais contido nonúcleo do óvulo destinado para a fertilizaçãodeve ser reduzido à metade, não pode haverdúvida que uma redução semelhante tambémdeve ocorrer, em alguma época e de algumamaneira, nos germoplasmas das células germi-nativas do macho.”
Na época em que essas surpreendentespredições foram feitas (surpreendentes porque semostraram essencialmente corretas) os citolo-gistas estavam encontrando evidências que asapoiavam. A observação mais importante estavasendo feita no verme nematóide Ascaris, que tema grande vantagem de possuir poucos cromos-somos e de grande tamanho, o que os torna fáceisde serem estudados.
Meiose na fêmea de Ascaris
Na penúltima década do século XIX, foramfeitas importantes contribuições para o entendi-mento da formação dos gametas e da fertilização.Três citologistas merecem referência especial:Edouard van Beneden (1846-1912), TheodorBoveri (1862-1915) e Oskar Hertwig (1849-1922). Eles descobriram que ocorrem duas
divisões celulares diferentes durante a formaçãodos gametas, as quais resultam na redução donúmero de cromossomos à metade – como Weis-mann previu que deveria acontecer. Estas duasdivisões são divisões mitóticas modificadas e fo-ram denominadas divisões meióticas – os nomessão tão parecidos que continuam a causarproblema para os estudantes até hoje. Nadescrição que se segue, utilizaremos aterminologia moderna.
O ovário de Ascaris começa a se formar noinício do desenvolvimento e o extraordinárioaumento no número de suas células é conseqüên-cia de divisões mitóticas. Cada núcleo tem quatrocromossomos, o número diplóide, e pode-se notarque cada cromossomo aparece duplo já no inícioda prófase. Nessa fase, eles estão formados porduas cromátides, indicando que eles se duplicaramantes do início da divisão. Na mitose, as oitocromátides são divididas entre as duas células-filhas, o que resulta em quatro cromossomos emcada uma delas.
À medida que a fêmea de Ascaris amadurece,seu ovário passa a conter células aumentadas, asovogônias, ainda com o número diplóide decromossomos. A célula reprodutiva femininapermanece diplóide até ser libertada do ovário epenetrada pelo espermatozóide. É somente entãoque a meiose começa e os corpúsculos polaressão formados. A figura 6 (de Boveri, 1887)mostra o que acontece.
No começo da meiose, cada um dos quatrolongos cromossomos da ovogônia encurta e tomao aspecto de uma pequena esfera. Estes quatrocromossomos então se juntam em pares, umprocesso conhecido como sinapse. Quando issoocorre, cada um dos cromossomos já está dupli-cado, pois a duplicação ocorreu na intérfaseprecedente. Assim, a célula em início de meioseterá dois grupos com quatro cromátides cada.Cada um desses grupos é denominado tétrade.As tétrades se separam em uma divisão celularaltamente desigual que resulta em um pequenocorpúsculo polar e uma célula grande, o futuroóvulo. Cada uma dessas duas células contém doiscromossomos duplicados. Portanto, cada tétradefoi dividida em duas díades.
Na segunda divisão meiótica, observa-se umacaracterística essencial da meiose: os cromos-somos não são duplicados. Então, cada díade seliga ao fuso e, na anáfase, suas duas cromátides
26

vão para pólos opostos. A célula se divide nova-mente de modo desigual. O resultado é umminúsculo corpúsculo polar com dois cromossomose um grande óvulo também com dois cromossomos.
Assim, no decorrer das duas divisões quecompõem a meiose, o número diplóide de quatrocromossomos da célula feminina foi reduzido aonúmero monoplóide de dois cromossomos. Ahipótese de Weismann provou ser verdadeira, pelomenos para as fêmeas de Ascaris.
Meiose no macho de Ascaris
A previsão de Weismann para os machos tambémmostrou-se correta. Quando os testículos foramestudados, verificou-se que, durante o início do
desenvolvimento, as células aumentam em númeropor divisões mitóticas, isto é, as células que seoriginam desse processo tem o número diplóide dequatro cromossomos (Bauer, 1893).
No entanto, no testículo maduro, as duasúltimas divisões antes de as células se diferen-ciarem em espermatozóides são diferentes. Équando ocorrem as divisões meióticas no macho.No que se refere aos cromossomos, os eventossão os mesmos que os da fêmea, mas quando seconsidera a célula como um todo, existemdiferenças entre as divisões meióticas de machoe de fêmea. (Fig. 7)
Durante a meiose do macho, os quatrocromossomos já duplicados se juntam dois a dois,formando dois pares, ou duas tétrades. Na
27
Células que irão formarespermatozóides possuem 4cromossomos, o númerodiplóide.
No início da meiose oscromossomos homólogosse emparelham.
Os cromossomosemparelhados formamtétrades, cada uma com 4cromátides.
Os cromossomoshomólogos se separamna primeira divisão dameiose.
Cada célula-filha daprimeira divisãocontém duas díades.
Não há duplicaçãocromossômica antes dasegunda divisão da meiosee as duas cromátides decada díade se separam.
Cada uma das células originadas na meiosediferencia-se em um espermatozóide.
Em Ascaris, a fecundaçãoé que desencadeia ameiose feminina.
No início da meiose, ocorre oemparelhamento dos
cromossomos homólogos.
Os cromossomos emparelhadosformam tétrades, cada uma com 4cromátides.
Os cromossomos homólogos separam-se na primeira divisão da meiose.
O primeiro glóbulo polar e o futuroóvulo contêm duas díades cada.
A formação do segundo glóbulo polardeixa o óvulo com um número
haplóide de cromossomos.
Os núcleos de origempaterna e materna seaproximam um do outro.
Cada pró-núcleo contribui comdois cromossomos para o
zigoto, restabelecendo o númerodiplóide da espécie.
➤➤
➤
➤
➤
➤➤ ➤ ➤ ➤
➤ ➤ ➤
➤
➤
➤
➤
➤
➤
➤
➤
➤
Figura 7. Representação esquemática da meiose masculina, à esquerda, e da meiose feminina, à direita emascaris de cavalos.

primeira divisão meiótica os cromossomos decada par se separam, indo uma díade (cromos-somo duplicado) para cada pólo. No entanto, aocontrário da primeira divisão meiótica da fêmea,no macho são produzidas duas células de igualtamanho. Na segunda divisão meiótica não háreplicação cromossômica, as díades se dividem ecada célula-filha termina com dois cromossomos.
Assim, a partir de cada célula diplóide origi-nal, com quatro cromossomos, formam-se quatrocélulas, cada uma com dois cromossomos, onúmero háplóide. Não há outras divisões dessascélulas e cada uma se diferencia em umespermatozóide.
Uma diferença fundamental entre a mitose e ameiose é que na mitose há uma duplicação decada cromossomo para cada divisão celular; nameiose há somente uma duplicação de cadacromossomo para duas divisões sucessivas.Então, a mitose é um mecanismo que mantéma constância do número cromossômico nasdivisões celulares enquanto a meiose reduz essenúmero à metade.
Fertilização e restabelecimento do númerodiplóide de cromossomos
O fato básico da fertilização, de que é o esper-matozóide e não o líquido seminal o desenca-
deador do desenvolvimento do ovo, foi desco-berto por J. L. Prévost e J. B. Dumas em 1824.No entanto, o real papel do espermatozóide nãofoi estabelecido nesse trabalho. Como relatadoanteriormente, foi George Newport (1854) quemdemonstrou que o espermatozóide penetra noóvulo das rãs. Mas o que acontece então?
Ascaris mostrou-se um material muito bompara o estudo dos detalhes da fertilização, nova-mente devido ao fato de possuir poucos e grandescromossomos. Van Beneden e Boveri descre-veram o processo em detalhe. (Fig.8)
A figura 8 mostra desenhos de Boveri, publi-cados em 1888. A primeira ilustração (A) é deum corte de um ovo logo após a entrada do esper-matozóide. O pró-núcleo paterno aparece noquadrante inferior direito. As duas massasirregulares fortemente coradas são os doiscromossomos – dois é o número haplóide. Aestrutura formando uma cápsula dobradaimediatamente acima do pró-núcleo paterno é oacrossomo, que é a parte da cabeça do esperma-tozóide composta por material do aparelho deGolgi. A massa granular escura no centro doóvulo é o centrossomo, que também se originado espermatozóide. Existem quatro corposescuros, aproximadamente na posição correspon-dente à das 12 horas; os dois superiores são oscromossomos do segundo corpúsculo polar. Os
28
AB C
D E F
Figura 8. Ilustrações de Boveri da fertilização em Ascaris Veja explicação no texto. (Boveri, 1888)

29
dois de baixo são os cromossomos, em númerohaplóide, do pró-núcleo materno. O segundocorpúsculo polar aparece nos cortes de embriõesmostrados em (C) e (E).
Em (B), os pró-núcleos materno e paterno jáse aproximaram um do outro e seus cromossomostornaram-se indistintos. Em (C) os cromossomosalongaram-se bastante e, embora agora saibamosque existem apenas dois cromossomos em cadapró-nucleo, isto não pode ser visto na ilustração(este é um exemplo claro da grande dificuldadeenfrentada pelos citologistas para chegarem àconclusão de que o número de cromossomos dequalquer espécie é constante e que os cromos-somos são individualmente únicos – na maioriadas vezes eles pareciam tão emaranhados comoum prato de espaguete). Pode-se distinguir doisgrânulos escuros no centrossomo: os centríolos.
Em (D), os cromossomos tornaram-se distin-tos uma vez mais [de (B) a (C) eles estavam emum estágio modificado de “repouso”] e vê-se doisem cada pró-núcleo. O centrossomo dividiu-seem dois, cada um deles com um centríolo nocentro. Este processo continua através de (E).Em (F), os quatro cromossomos, dois de cada pró-núcleo, estão alinhados no fuso e, logo depois disso,pode-se ver que cada um está duplicado, isto é, com-posto por duas cromátides. As cromátides irão seseparar para formar cromossomos independentesque vão para pólos opostos da célula.
A forma do aparelho mitótico está bemmostrada em (F). Em cada extremidade do fusoencontra-se um minúsculo centríolo, cercado poruma área granular escura – o centrossomo. Emmaterial bem preservado, pode-se observar fibrasirradiando de cada centrossomo, formando umáster. Outras fibras se estendem de um centros-somo ao outro, formando o fuso. Em (F), célulaem metáfase da primeira divisão embrionária comos cromossomos alinhados na placa equatorial.
Significado da formação dos gametas e dafertilização
Tudo estava acontecendo como previsto porWeismann. As células que iriam, muitas geraçõesmais tarde, formar os gametas, tanto no ovário comono testículo de Ascaris, começavam com quatrocromossomos, o número diplóide. E, estas célulasse dividiam repetidamente, sempre por mitose.
Nos machos, as duas últimas divisões das célulasformadoras de espermatozóides no testículo eram,
no entanto, meióticas e não mitóticas. Durante estasduas divisões, as células se dividiam duas vezes masos cromossomos se replicavam somente uma vez.Essas divisões celulares eram iguais, isto é,produziam, cada uma delas, duas células de mesmotamanho. Isso resultava em quatro células de igualtamanho, cada uma com dois cromossomos, onúmero haplóide. As quatro células, então, sediferenciavam nos espermatozóides.
Depois de muitos ciclos celulares de divisõesmitóticas, algumas das células ovarianas aumen-tavam bastante em tamanho, formando osovócitos. Como no caso dos machos, ocorriamduas divisões meióticas do material nuclear comsomente uma única replicação dos cromossomos.A primeira divisão da célula era tão desigual quea maior parte do material permanecia na célulaque iria originar o óvulo e só uma ínfimaquantidade era incluída no primeiro corpúsculopolar. Isto se repetia na segunda divisão,produzindo um minúsculo segundo corpúsculopolar e um grande óvulo. Todavia, os núcleos dosegundo corpúsculo polar e do óvulo eramidênticos – cada um continha um número haplóidede dois cromossomos.
A meiose em Ascaris produzia, portanto, oespermatozóide monoplóide e o óvulomonoplóide. A união deles originava o zigotodiplóide – o início de um novo verme nematóide.Os processos estão sumarizados na figura 7.
Estava claro pelo trabalho de van Beneden,Boveri e outros que cada genitor transmite o mesmonúmero de cromossomos ao zigoto. Além disso, oscromossomos no núcleo materno e paterno pare-ciam ser idênticos. Estas duas observações podiamajudar a explicar o que já se acreditava há algumtempo: que a contribuição hereditária de cada geni-tor é aproximadamente a mesma.
Este era um campo de pesquisa excitante eimportante e logo muitos pesquisadores estavamestudando uma grande variedade de plantas eanimais. Com poucas exceções, o que seencontrou em Ascaris mostrou-se verdadeiro paratodos os outros organismos. Certamente existiampequenas variações, mas um estudo intenso serviusomente para aumentar a profundidade do nossoentendimento do processo global. Um conceitode aplicação universal havia sido descoberto.
As extraordinárias observações sobre ocomportamento dos cromossomos na mitose,meiose e fertilização, feitas entre 1870 e 1890,

30
PARTE A: REVENDO CONCEITOS BÁSICOS
Preencha os espaços em branco nas frases de1 a 6 usando o termo abaixo mais apropriado.
(a) anáfase (c) metáfase (e) prófase(b) meiose (d) mitose (f) telófase
EXERCÍCIOS
principalmente na Alemanha, forneceram umquadro geral para a transmissão de geração ageração das estruturas fundamentais responsáveispela herança.
Pode-se argumentar, com razão, que essesestudos não forneceram qualquer evidência cru-cial de que os cromossomos seriam, de fato, abase física da hereditariedade. Eles apenas suge-riam que os cromossomos poderiam desempenhartal papel. Nem mesmo durante as últimas duasdécadas do século XIX se chegou próximo àdescoberta de como seria possível estabelecer opapel de uma estrutura celular na herança. Tantoa Citologia como o que atualmente chamamosde Genética estavam no estágio de ciência nor-mal Kuhniana, esperando pela chegada de umnovo paradigma. Isso iria ocorrer, de uma formadramática, no ano de 1900.
Mas antes de entrarmos no século XX,podemos concluir com este sumário de E. B.Wilson sobre o que havia sido estabelecido du-rante o florescimento da Citologia no últimoquarto do século XIX.
“ O trabalho da Citologia neste período deestabelecimento de seus fundamentos construiu umabase ampla e substancial para as nossas concep-ções mais gerais de hereditariedade e seu substratofísico. Foi demonstrado que a base da heredita-riedade é uma conseqüência da continuidadegenética das células pela divisão e que as célulasgerminativas são o veículo da transmissão de umageração para outra. Acumularam-se fortes evidên-cias de que o núcleo da célula desempenha umpapel importante na herança.
Descobriu-se o significativo fato de que emtodas as formas ordinárias de divisão celular onúcleo não divide “em massa” mas que primeiroele se transforma em um número definido decromossomos; estes corpos, originalmente forma-dos por longos fios, dividem-se longitudinalmentepara efetuar uma divisão merismática da subs-tância nuclear inteira.
Provou-se que toda fertilização do óvulo,envolve a união ou estreita associação de doisnúcleos, um de origem materna e o outro deorigem paterna. Está estabelecido o fato, algumasvezes chamado de “lei de van Beneden” em home-nagem ao seu descobridor, que estes núcleosgerminativos primários dão origem a grupossemelhantes de cromossomos, cada um contendometade do número encontrado nas células docorpo. Demonstrou-se que quando novas célulasgerminativas são formadas cada uma volta areceber apenas metade do número característicode cromossomos das células do corpo.
Acumularam-se evidências, especialmentepelos estudos admiráveis de Boveri, que oscromossomos de sucessivas gerações decélulas, que normalmente não são vistos nosnúcleos em repouso, na realidade, não perdemsua individualidade, ou que de uma maneiramenos óbvia eles se adaptam ao princípio dacontinuidade genética. Desses fatos, tirou-seuma conclusão de que os núcleos das célulasdo corpo são diplóides ou estruturas duplas,descendentes igualitários dos grupos decromossomos de origem materna e paterna doovo fertilizado. Esses resultados, continua-mente confirmados pelos trabalhos dos últimosanos [isto é, ciência normal], gradualmentetomaram um lugar central na Citologia; [...]Uma nova era de descobertas agora se abre[um novo paradigma]. Assim que o fenômenomendeliano tornou-se conhecido ficouevidente que em linhas gerais, ele forma umcomplemento para aqueles fenômenos que aCitologia já tinha tornado conhecido arespeito dos cromossomos.”
Esta citação é parte da famosa Croonian Lec-ture to the Royal Society of London proferida porWilson em 1914. Nessa época as relações entrecromossomos e herança já haviam sido testadase se mostrado verdadeiras acima de qualquersuspeita..
1. ( ) é um tipo de divisão nuclear em que osnúcleos-filhos conservam o mesmo número decromossomos do núcleo original.
2. A migração dos cromossomos para os pólosocorre na ( ).

31
3. ( ) é um tipo de divisão nuclear que reduz àmetade o número de cromossomos nosnúcleos-filhos.
4. Cromossomos arranjados na região equa-torial da célula caracteriza a fase da divisãochamada ( ).
5. ( ) é a primeira fase da divisão celular, naqual os cromossomos se tornam evidentes.
6. ( ) é a fase final da divisão celular, em queos núcleos se reorganizam.
Preencha os espaços em branco nas frases de7 a 11 usando o termo abaixo mais apropriado.
(a) corpúsculo polar (c) haplóide (e) ovogônia(b) diplóide (d) ovócito
7. ( ) é uma célula que está sofrendo meiose edará origem a um gameta feminino.
8. Uma célula animal feminina que irá sofrermeiose é chamada ( ).
9. ( ) é o termo usado para designar uma célulaque possui dois conjuntos de cromossomos.
10. ( ) é o nome das minúsculas células que seformam no decorrer da meiose feminina.
11. ( ) é o termo que designa uma célula quepossui apenas um conjunto de cromossomos.
Preencha os espaços em branco nas frases de12 a 17 usando o termo abaixo mais apropriado.
(a) cromátide (d) pró-núcleo(b) díade (e) sinapse(c) fertilização (f) tétrade
12. Dois cromossomos emparelhados no inícioda meiose formam um(a) ( ).
13. O emparelhamento de cromossomos nameiose é chamado ( ).
14. ( ) é o nome do núcleo do espermatozóideou do óvulo imediatamente antes de eles sefundirem para formar o núcleo do zigoto.
15. Um cromossomo duplicado, formadoportanto por dois filamentos idênticos, échamado ( ).
16. ( ) é nome que se dá a cada um dos doisfilamentos que formam um cromossomoduplicado.
17. A fusão de dois gametas com formação deum zigoto é chamada ( ).
PARTE B: L IGANDO CONCEITOS E FATOS
Utilize as alternativas abaixo para completar asfrases de 18 a 21.
a. fertilização b. meiose c. mitose
18. Na ( ) ocorre uma replicação cromossômicapara duas divisões celulares.
19. Na ( ) ocorre uma replicação cromossômicapara uma divisão do citoplasma.
20. Apenas células diplóides se dividem por ( ).
21. Células diplóides e haplóides se dividempor ( ).
Utilize as alternativas abaixo para completar asfrases de 22 a 25.
a. células diplóidesb. células haplóides
22. Ovogônias são sempre ( ).
23. Gametas são sempre ( ).
24. Zigotos são sempre ( ).
25. Meiose produz sempre ( ).
26. Dos cinco eventos listados a seguir,quatro ocorrem tanto na mitose quanto nameiose. Indique qual deles acontece es-sencialmente na meiose?a. Condensação dos cromossomos.b. Formação do fuso.c. Emparelhamento dos cromossomos.d. Migração dos cromossomos.e. Descondensação dos cromossomos.
Utilize as alternativas abaixo para completar asfrase de 27 e 28.
a. Antonie van Leeuwenhoekb. Nehemiah Grewc. Robert Brownd. Robert Hooke
27. ( ) é considerado o descobridor da célula.
28. ( ) foi o descobridor do núcleo celular.
Utilize as alternativas abaixo para completar asfrases de 29 a 33.
a. August Weismann c. Theodor Schwannb. Rudolph Virchow d. Walther Flemming
29. A idéia de que a formação do corpúsculo polaré uma estratégia para a redução do materialhereditário do óvulo foi lançada em 1887 por ( ).

32
30. Um dos formuladores da teoria celular foi ( ).
31. A célebre frase “omnis cellula e cellula”,indicando que toda célula provém da divisãode outra célula, foi cunhada em 1855 por ( ).
32. ( ) é considerado o descobridor da mitosepelo fato de ter demonstrado que os eventoscromossômicos observados em células fixadase coradas ocorriam nas células vivas.
33. A teoria celular mostrou que, apesar dasdiferenças visíveis a olho nu, todos os seresvivos são iguais em sua constituição básica, poisa. são capazes de se reproduzirsexuadamente.b. são formados por células.c. contêm moléculas.d. se originam de gametas.
34. Os vírus não são exceções à teoriacelular poisa. são formados por células.b. formam gametas.c. são organismos vivos.d. só conseguem se reproduzir no interior deuma célula viva.
PARTE C: QUESTÕES PARA PENSAR E DISCUTIR
O número diplóide de cromossomos da espéciehumana é 46. Essa informação deve ser usadapara responder as questões 35 e 36.
35. Determine o número de filamentoscromossômicos (cromátides) presentes em umnúcleo celular humana em
a. prófase mitótica. d. telófase mitótica.b. prófase I da meiose. e. telófase I da meiose.c. prófase II da meiose. f. telófase II da meiose.
Obs. No caso das telófases, considere apenasum dos núcleos em formação.
36. Determine o número de filamentoscromossômicos (cromátides) presente em cadaum dos tipos celulares relacionados a seguir.
a. Espermatozóide.b. Óvulo.c . Primeiro corpúsculo polar.d. Segundo corpúsculo polar.e. Ovócito primário.f . Ovócito secundário.
37. Qual é a hipótese central da teoria celular?
38. Por que foi difícil estender o conceito decélula para os animais?
39. Que critério Schwann utilizou paraestabelecer relações de semelhança entre asunidades microscópicas que compõem o corpodos animais e das plantas?
40. Que tipo de observação permitiu concluirque espermatozóides eram células?
41. Por que não se deve chamar o núcleointerfásico de núcleo em repouso, como faziamos antigos citologistas? Por que era usadoaquele nome?
42. O que Weismann imaginou ser necessáriopara manter a constância do número decromossomos através das gerações?
43. Identifique as principais diferenças entremitose e meiose.
44. Analise os tipos de argumento usados parajustificar a idéia de que a informação hereditáriaestaria contida nos gametas.
45. Que tipo de raciocínio dedutivo levou osantigos citologistas a concluir que a mitose nãopoderia ser o único tipo de divisão celular?
46. Qual é o significado da meiose e dafertilização no ciclo de vida dos organismos?
47. Que importantes paradigmas direcionaramas pesquisas citológicas nos primeiros doisterços e no último terço do século XIX,respectivamente?

Terceira aula(T3)
Texto adaptado de:MOORE, J. A. Science as a Way of Knowing -Genetics. Amer. Zool. v. 26: p. 583-747, 1986.
M ENDELISMO : AS LEIS DA SEGREGAÇÃO
E DA SEGREGAÇÃO INDEPENDENTE
AS ORIGENS DA GENÉTICA
O ano de 1900 marca o início da GenéticaModerna. Foi quando um modesto, subestimadoe esquecido trabalho de um monge Agostiniano,falecido em 1884, tornou-se conhecido pelacomunidade científica em geral. A área decruzamento animal e vegetal tinha passado porum período longo e não-excitante de “ciêncianormal” kuhniana, mas em 1900 estava paraacontecer uma notável mudança de paradigma ea Genética estava a caminho de se tornar umaCiência com ampla capacidade de explicar fatose de fazer previsões. O novo paradigma surgiucom a descoberta de um trabalho longo e poucoconhecido, Versuche über Pflanzen-Hybriden,baseado em conferências que Gregor Mendelproferiu na Sociedade de História Natural deBrünn, na Áustria (hoje Brno, na atual RepúblicaTcheca) em 8 de fevereiro e 8 de março de 1865,publicado em 1866.
A história é familiar aos professores de Bio-logia e uma fonte apropriada de documentos bási-cos é fornecida por Stern & Sherwood (1966).Dois cientistas, o holandês Hugo de Vries (1900)e o alemão Carl Correns (1900) são consideradoscomo os primeiros a compreender a importânciado que Mendel tinha feito. Um terceiro cientista,o austríaco Erick von Tschermark, é usualmente
incluído como um dos redescobridores mas Sterne Sherwood (1966, p. X-XI) dão razões de por-que ele não merece esse reconhecimento.
De Vries havia cruzado numerosas “espécies”e variedades de plantas durante a última décadado século XIX. Naquela época, o termo “espécie”era às vezes aplicado para plantas cultivadas quediferiam entre si por um ou poucos alelos capazesde produzir grandes alterações fenotípicas e quehoje são considerados como pertencentes à mes-ma espécie. De Vries adotou o ponto de vista deque estas diferentes “espécies” poderiam serconsideradas “como constituídas de fatores inde-pendentes” ou unidades e que, “As unidadesdeterminantes dos caracteres específicos dasespécies devem ser consideradas, nesse sentido,como entidades nitidamente separadas e deve-riam ser estudadas como tal. Elas deveriam sersempre tratadas como independentes umas dasoutras, uma vez que não há razão para ser deoutro modo. Em todo experimento de cruza-mento, apenas um caráter, ou um número defi-nido deles, deve ser considerado.”
De Vries se referiu a caracteres em estadosantagônicos mas observou que somente umdesses estados se expressava nos híbridos (istoé, em F
1). Contudo quando os grãos-de-pólen e
óvulos eram formados nos híbridos “os doisestados antagônicos das características se
Objetivos1. Identificar as épocas da descoberta e redescoberta
das leis fundamentais da hereditariedade, assimcomo os pesquisadores envolvidos.
2. Identificar as vantagens da ervilha para o estudoda hereditariedade.
3. Descrever os experimentos de Mendel.4. Explicar como Mendel testou suas hipóteses
sobre hereditariedade.5. Enunciar a primeira e a segunda leis de Mendel.
33

separavam, seguindo na maioria das vezes asleis simples da probabilidade”.
De Vries verificou que essas conclusões essen-ciais a que havia chegado tinham sido apresentadas35 anos antes por Mendel, cujo trabalho tinha sidoesquecido e seu significado não compreendido.
A história de como De Vries tomou conheci-mento do trabalho de Mendel é bem interessante.Ele não o descobriu por meio de pesquisa biblio-gráfica, mas por um desses acasos extraordináriosque parecem ser de grande importância nas desco-bertas científicas. Um cientista holandês, Profes-sor Beyerinck, sabia que De Vries estava traba-lhando com hibridação de plantas e escreveu per-guntando se ele não estaria interessado em umaantiga publicação sobre o assunto. Era o trabalhode Mendel. A carta e a publicação chegaram àsmãos De Vries em 1900, justamente quando eleestava preparando seus próprios resultados parapublicação. Ele estava preparado para compre-ender que estava confirmando os experimentosanteriores e mais completos de Mendel.
A história sobre Correns é igualmente inte-ressante. Ele também tinha realizado experimen-tos de cruzamento com plantas e estava tentandodesenvolver uma hipótese para explicar os resul-tados obtidos. No outono de 1899, a solução veioa ele num “estalo” que, com mais freqüência doque se imagina, parece ser a origem de rupturasverdadeiramente importantes em Ciência. Poucotempo depois ele encontrou o trabalho de Mendele o leu. Ele publicou seus próprios resultados emostrou sua semelhança com os de Mendel.
A famosa obra de Gregor Mendel não é umtrabalho científico no sentido convencional, masum conjunto de conferências que ele apresentouna Sociedade de História Natural de Brünn em1865. Os dados completos nunca foram publi-cados mas, a parte que ele incluiu, juntamentecom sua extraordinária análise dos resultados,coloca sua contribuição no mesmo nível da deCharles Darwin.
Mendel estava plenamente consciente dosexperimentos de cruzamentos de plantas, normal-mente chamados de hibridação, que haviam sidorealizados durante anos por muitos cientistasfamosos. Nenhuma lei geral havia sido proposta,como já vimos ao considerar o insucesso de Dar-win no livro Variation ... publicado somente doisanos depois do trabalho de Mendel .
Mendel começou seus experimentos sobrehereditariedade alguns anos antes da publicação,em 1859, da obra On the Origin of Species e umadas razões para o que ele se propunha a fazer,como ele próprio diz, era a necessidade de “sechegar a uma solução para a questão, cujo signi-ficado para a história evolutiva da vida não deveser subestimado.” Assim, o trabalho de Mendelcomeçou como “Ciência normal” dentro do para-digma da Teoria da Evolução. Somente mais tardeé que ele iria se tornar o início de um novo para-digma – A Genética mendeliana. Este é um pontointeressante, pois mostra como uma descobertaem um determinado campo da Ciência pode serde grande importância para outro campo.
M ATERIAL E MÉTODOS USADOS
POR MENDEL
Características favoráveis da ervilha
Na metade do século XIX, as pessoas interes-sadas em hibridação de plantas tinham umariqueza de material à sua disposição. Numerosasvariedades de uma mesma espécie, tanto deplantas comestíveis como ornamentais, tinhamsido selecionadas. Muitas das variedades diferiambastante entre si; algumas eram tão diferentes quechegavam a ser consideradas espécies distintas erecebiam nomes científicos próprios.
Mendel decidiu trabalhar com ervilhas e come-çou com 34 variedades. Ele as cultivou por duasestações para estar seguro de que elas eram capazesde se cruzar e produzir descendência fértil. Nofinal reduziu o número a 22 variedades. (Fig. 9)
As ervilhas tinham vantagens importantes.Além de existir muitas variedades disponíveis,como já mencionamos, elas eram fáceis de secultivar e seu tempo de geração era curto. Osdescendentes obtidos por cruzamento entre asvariedades eram férteis. A estrutura da flortambém era importante. Os estames e pistilosficavam encobertos pelas pétalas e, se as floresfossem cobertas para evitar a ação dos insetos,elas se autofecundavam, isto é, o pólen caía sobreo estigma da mesma flor.
Apesar disso, era possível fazer cruzamentosexperimentais removendo as anteras antes damaturação da flor e, mais tarde, colocando pólende outra planta sobre o estigma. Assim, Mendelpodia cruzar qualquer uma das suas variedades
34

ou, se ele deixasse as flores intactas, a próximageração seria resultado da autofecundação.
A análise matemática
Aqueles que já ensinaram Genética mendelianasabem que muitos estudantes têm dificuldade coma Matemática. Isso poderá se tornar mais fácilquando chegarmos a 1903 e colocarmos as unida-des hereditárias nos cromossomos, mas vale fazerum pequeno esforço agora para entender algunsaspectos críticos do modelo mendeliano como,por exemplo, a proporção 3 : 1 para um par deestados contrastantes de um caráter pode ser ex-pandida para a proporção 9 : 3 : 3 : 1 quando setrata de dois pares de estados de dois caracteres.
Em minha experiência, um dos princípios maisdifíceis para os estudantes é aprender que 1/4 de1/4 não é 1/2 nem 1/8, mas 1/16. Além disso, osestudantes devem perceber que a proporção 3 : 1 é
AB
C
D
E
Pétalas que for-mam a carena
AnterasEstigma
Carena
➤
Estandarte
Ala
➤
Estandarte
Estandarte
Estilete
Filete
Óvulo
Ala
Carena➤
➤
➤
➤
➤
➤
➤
➤
➤
➤
➤➤
Óvulo
➤
➤
Ala ➤
Sépalas
Figura 9. Representação esquemática da flor de ervilha (Pisumsativum). A. Flor inteira. B. Corte longitudinal da flor. C. Pétalasisoladas. D. Estames envolvendo o pistilo. E. Pistilo.(Redesenhada de Rawitscher, F. Elementos básicos de Botânica.São Paulo: Nacional, 1967)
o mesmo que dizer que 3/4 da amostraé de um tipo e 1/4 da amostra é de outrotipo ou então que 75% é de um tipo e25% de outro.
Na discussão do experimento deMendel que se segue procurou-seapresentar o tópico de uma maneiraque auxilie a compreensão e permitauma apreciação a altura do queMendel fez.
OS RESULTADOS DE MENDEL
As características estudadas
Por meio de cruzamentos gené-ticos tenta-se descobrir a base heredi-tária das diferenças; ao mesmo tempoas informações obtidas podem nosajudar a entender porque indivíduosde uma mesma espécie se parecemtanto entre si. Não se cruzam indiví-duos geneticamente idênticos naesperança de se descobrirem leis daherança. Por isso Mendel escolheuvariedades de ervilhas que diferiamentre si. A diferença era em relaçãoaos estados das características entreas diversas variedades. Assim,algumas de suas variedades tinhamsementes lisas enquanto que emoutras as sementes eram rugosas
(angulosa seria a melhor tradução do termo usadopor ele); algumas das suas variedades tinhamsementes amarelas e outras, verdes. Ao todo, eleusou 7 caracteres, que apresentavam estadoscontrastantes, como se segue:
Variedades apresentando estados contrastantesde caracteres foram cruzadas por meio da remo-ção das anteras ainda imaturas das flores de umavariedade e colocação de pólen de outra varie-
CARÁTER AFETADO ESTADO
Textura da semente lisa ou rugosaCor da semente amarela ou verdeRevestimento da semente colorido ou brancoTextura da vagem inflada ou enrugadaCor da vagem verde ou amarelaPosição da flor axilar ou apicalComprimento do caule longo ou curto
35

dade, sobre seus estigmas. A primeira geraçãohíbrida, ou F
1 ( abreviatura para first Filial gen-
eration) para usarmos um termo introduzido maistarde, dava um resultado uniforme: todos asplantas F
1 exibiam o estado da característica de
um dos genitores. Mendel chamou de dominanteo estado da característica que aparecia nas plantasF
1, em contraste com o estado da característica
que não aparecia, por ele chamado de recessivo.Estes resultados, tão familiares hoje em dia,
eram bastante inesperados naquela época.Embora outros casos semelhantes ao obtido porMendel já houvessem sido descritos, a regra geralera a de que os indivíduos de F
1 apresentassem
estados de caracteres intermediários aos dos pais.E na maioria dos casos, isso ocorria por umasimples razão: se as variedades diferirem emdiversas características os indivíduos F
1 serão, em
geral, mais ou menos intermediários entre os tiposparentais. Mas Mendel concentrou-se na análiseda herança dos detalhes, ou seja, dos caracteresisolados, e não do indivíduo como um todo. Nessesentido, ele esqueceu da planta como um todo equestionou somente se as ervilhas tinham semen-tes lisas ou rugosas, se eram altas ou baixas etc.
As plantas F1 foram protegidas para não serem
polinizadas pelos insetos e, em conseqüência seautofecundaram. De novo, os resultados foramuniformes. Para cada um dos sete tipos de cruza-mento originais entre plantas com estados decaráter contrastantes, a descendência F
2 asse-
melhava-se a um ou outro genitor da geração P.Ele nunca encontrava intermediários.
A proporção 3 : 1
Enquanto que a maioria dos cultivadores deplantas tinha descrito somente o reaparecimentode ambas as variedades em F
2 ( abreviatura para
second Filial generation), Mendel fez uma coisasimples e extraordinária. Ele contou o númerode indivíduos com cada característica. Os resul-tados para os sete tipos de cruzamentos foramos mesmos: a proporção de três plantas com a carac-terística dominante para uma com a recessiva.Ou nós podemos dizer 3/4 (75%) de F
2 apresen-
tava o estado dominante e 1/4 (25%), o estadorecessivo da característica analisada (Tab. 1).
Estas proporções e porcentagens eramderivadas dos dados brutos observados. Por
exemplo, no caso docruzamento de plantaspuras de sementes lisascom plantas puras desementes rugosas, Mendelobteve 7324 sementes F
2
das quais 5474 eram lisase 1850, rugosas – umaproporção de 2,96 para 1.O cruzamento de plantas“puras” de sementes ama-relas com plantas “puras”de sementes verdes produ-ziu 8023 sementes F
2 das
quais 6022 eram amarelase 2001, verdes – umaproporção de 3,01 para 1.Podemos ver que Mendeltinha razão em suspeitar quea resposta teórica deveria se3 : 1 e não 3,01 para 1. Esteseram cruzamentos mono-íbridos, isto é, envolviam sóum carácter com doisestados contrastantes.
36
1. Textura das sementesLisa X Rugosa
Sementes lisas
2. Cor das sementesAmarela X Verde
3. Cor da cascadas sementesCinza X Branca
4. Textura das vagensInflada X Comprimida
5. Cor das vagensVerde X Amarela
6. Posição das floresAxilar X Terminal
7. Comprimento do cauleLongo X Curto
Sementesamarelas
Semente decasca cinza
Vagens infladas
Vagens verdes
Flores axilares
Caule longo
Lisa X Lisa
Amarela X Amarela
Cinza X Cinza
Inflada X Inflada
Verde X Verde
Axilar X Axilar
Longo X Longo
5474 lisas1850 rugosas
7324 (total)6022 amarelas2001 verdes
8023 (total)
705 cinzas224 brancas
929 (total)
882 infladas 299 comprimidas
1181 (total)428 verdes152 amarelas
580 (total)
651 axilaress207 terminais
858 (total)
787 longos 277 curtos
1064 (total)
2,96 : 1
3,01 : 1
3,15 : 1
2,95 : 1
2,82 : 1
3,14 : 1
2,84 : 1
Tipos de cruzamentoentre plantas “puras”
Característicasdas plantas F
1
Autofecundaçãode F
1
Plantas F2
Razão entreos tipos F
2
Tabela I. Resultados obtidos por Mendel em cruzamentos entre variedadesde ervilha.

A proporção 9 : 3 : 3 : 1
Quando Mendel seguiu a herança de duascaracterísticas com dois pares de estados contras-tantes, no cruzamento diíbrido, foram obtidosnovamente, resultados uniformes. As plantas dageração F
1 exibiam os dois estados dominantes das
características analisadas e, em F2, apareciam quatro
tipos de planta na proporção de 9 : 3 : 3 : 1. Isto é,9/16 de F
2 mostrava ambos os estados domi-
nantes, 3/16 mostrava um dominante e outrorecessivo, 3/16 mostrava o outro dominante e oprimeiro recessivo e 1/16 tinha ambos os estadosrecessivos dos caracteres.
Assim, quando as plantas cruzadas na geraçãooriginal P eram do tipo liso-amarelo e rugoso-verde, todos os F
1 apresentavam sementes do tipo
liso-amarelo. Na geração F2, Mendel obteve 315
sementes do tipo liso-amarelo, 108 liso-verde,101 rugoso-amarelo e 32 rugoso-verde. Para umtotal de 556, a proporção dos diferentes tipos foide 9,8 : 3,4 : 3,2 : 1. Estas proporções represen-tam os dados reais, mas Mendel propôs a hipótesede que o resultado teórico esperado deveria ser9 : 3 : 3 : 1.
Agora nosso problema é analisar como aproporção 3 : 1 está relacionada com a 9 : 3 : 3 : 1.
No cruzamento amarelo com verde, a geraçãoF
2 seria 3/4 amarela e 1/4 verde; no cruzamento
liso e rugoso, F2 seria 3/4 lisa e 1/4 rugosa.
Uma pergunta que se pode fazer nesse pontoé se com base nessas informações seria possívelprever a proporção de 9 : 3 : 3 : 1 obtida em F
2?
Para muitos essa pergunta parecerá um problemainsolúvel, mas a análise a seguir resolve a questão.
Quando duas ou mais características comestados contrastantes estão envolvidas, pode-severificar que a proporção 3 : 1 ainda é mantidase considerarmos cada característica individual-mente. Verifique no cruzamento mencionadoacima (liso-amarelo x rugoso-verde) que aproporção 3 : 1 é mantida para cada um dos paresde estados das características consideradas.
[Não devemos nos esquecer que Mendel era professor deFísica; teria havido aqui uma influência do pensamentode Galileu? “Para Galileu, o objetivo da investigação erao conhecimento da Lei, captada na própria natureza, pelaobservação dos fenômenos, confirmada pelaexperimentação e matematicamente quantificada”]
Isso pode ser verificado também matematica-mente. Considere a proporção 9/16 liso-amarelo:3/16 liso-verde: 3/16 rugoso-amarelo: 1/16rugoso-verde. Considerando o par liso/rugososeparadamente, temos 9/16 + 3/16 = 12/16 desementes do tipo liso e 3/16 + 1/16 = 4/16 dotipo rugoso. Desde que 12/16 = 3/4 e 4/16 = 1/4,nós observamos a proporção 3:1 para esse parde características. O mesmo é verdadeiro para opar amarelo-verde.
Assim, se nós perguntarmos em que freqüên-cias aparecerão os indivíduos F
2 de um cruza-
mento diíbrido, a resposta pode ser obtida comuma simples multiplicação de frações. Assim, dos3/4 que serão lisos, 3/4 serão também amarelos e1/4 será verde; portanto 3/4 x 3/4, ou 9/16, serãolisos e amarelos e 3/4 x 1/4, ou 3/16, serão lisose verdes. Do 1/4 de F
2 que será rugoso, 3/4 serão
também amarelos e 1/4 será verde. Logo, 1/4 x3/4 (ou 3/16) serão rugosos-amarelos e 1/4 x 1/4(ou 1/16) será rugoso-verde. Esta é a derivaçãoda proporção 9 : 3 : 3 : 1.
Estas regularidades foram observadas porMendel em todos os seus cruzamentos. Por isso,ele achou que devia haver um princípio funda-mental responsável por elas. E havia.
M ODELO PARA CRUZAMENTO MONOÍBRIDO
A figura 10 é um modelo que explica a hipóteseque Mendel propôs para os cruzamentos mono-íbridos. Tanto o esquema quanto a terminologiaseriam padronizados meio século mais tarde, noinício do século XX.
A primeira coisa que um leitor atento notariaé um “erro” nos genótipos da geração P. Elesestão mostrados como haplóides ao invés dediplóides e isto levanta um ponto muito impor-tante em nosso estudo sobre conceitosgenéticos. Mendel usou os símbolos de genótipopara indicar os tipos de fatores hereditários, enão seu número por gameta. Os gametasparentais lisos poderiam conter inúmeros fatoresR, e não somente um, como se acredita hoje. Aautopolinização de linhagens puras lisasproduzia apenas descendência lisa, pois seusgametas só podiam conter fatores R. (As letrasmaiúsculas e minúsculas indicam que um aleloé dominante ou recessivo, respectivamente.)
37

A pureza dos gametas
No cruzamento entre plantas “puras” desementes lisas com plantas de sementes rugosaspoderia haver apenas um tipo de descendência(Rr) já que há somente um tipo de pólen e umtipo de óvulo. Quando estas plantas F
1 amadure-
ceram, cada flor produziu óvulos e grãos de pólen.Agora, surge um dos aspectos mais importantesdo modelo de Mendel: ele assumiu que umgameta poderia apresentar fatores de heredi-tariedade de apenas um tipo, isto é, um gametaproduzido pela planta F
1 iria ter R ou r, mas nunca
ambos (Fig. 10).
Com certeza, esta idéia seria de difícil aceita-ção pelos geneticistas do início do século – suasmentes estavam influenciadas pelo conceito de inú-meras gêmulas. Como um gameta poderia ser“puro”, ou seja, ter gêmulas apenas do tipo R ou r?
Mendel considerou, então, que os indivíduosde F
1 produziam gametas que continham ou o
fator R ou o r, mas nunca ambos. Em seguida,ele assumiu que óvulos e grãos de pólen seriamcombinados aleatoriamente e que as freqüênciasdos diferentes tipos de descendentes seriam
Liso X Rugoso
R r
➤ ➤R r
X
➤ ➤
➤ ➤
R
r R
r
Liso
Rr
LisoRR
Liso Rr
Liso
Rrrugoso
rr
➤
Rugoso
Rr
➤ ➤
➤
P1
F2
Fenótipo
Fenótipose
Genótipos
Genótipo
Genótipo dosgametas
F1
Fenótipo
Genótipo
Genótipo dosgametas
Figura 10. Modelo de um cruzamento mendelianomonoíbrido. Os genótipos dos indivíduos da geraçãoP estão representados como mostrado por Mendel emseu trabalho. O quadro de cruzamento abaixo mostraa origem do F2 (genótipos como representado atual-mente) a partir de pólen e óvulos de F
1.
determinadas pelas freqüências dos diferentestipos de gametas formados.
Deve-se enfatizar nesse ponto que a aparentesimplicidade do esquema da figura 10, sobre aorigem de F
2 a partir de gametas de F
1, advém do
fato de as classes genotípicas de óvulos e grãos depólen estarem em freqüências iguais. Isto é, cadaindivíduo produz 50% de gametas R (pólen ouóvulo) e 50%, r . As linhas representam todas ascombinações que podem ocorrer – e elas ocorremem freqüências iguais. O gameta R da esquerda,por exemplo, supondo que seja um grão de pólen),tem igual oportunidade de se combinar com umóvulo R ou r . O mesmo é válido para o pólen r.
O quadro inferior da figura 10 é um outromodo convencional de apresentação que facilitao entendimento de como um cruzamentomonoíbrido, pela hipótese de Mendel, resulta naproporção de 3 : 1 em F
1.
O modelo é válido para todos os cruzamentosque envolvem apenas um par de estados contras-tantes de uma característica. No entanto, omodelo explicará os resultados apenas nas seguin-tes condições:1. Em cada par de unidades hereditárias contras-
tantes, um membro do par é dominante e ooutro recessivo. Dominância e recessividadesão definições operacionais – determinadaspelo fenótipo de um indivíduo que apresentaambos os tipos de unidades hereditárias.
2. As unidades hereditárias dominantes e reces-sivas, quando presentes juntas, não modificamuma à outra de modo permanente. Assim, emF
1 o fator r do parental rugoso, no cruzamento
da figura 10, está combinado com o fator Rdo parental liso. Não há expressão do fator rem F
1, mas em F
2 1/4 dos indivíduos são
rugosos e tão enrugados quanto um de seus avós.3. Por algum mecanismo desconhecido por
Mendel, os dois tipos de fatores segregam detal maneira que cada gameta contém apenasum dos tipos. Assim, no exemplo, os gametasconterão R ou r.
4. Ainda por outro mecanismo tambémdesconhecido por Mendel, os gametas quecontêm R e aqueles que contêm r sãoproduzidos em números iguais.
5. Combinações entre os grãos de pólen e osóvulos são totalmente ao acaso e a proporçãoda descendência resultante irá depender daproporção das diferentes classes de gametas.
38

O teste da hipótese
Deve ser enfatizado que a concordância entreos dados e o modelo não é casual. Apesar dositens 1 e 2 acima poderem ser considerados fatos,os itens de 3 a 5 são totalmente hipotéticos –foram propostos para explicar os dados. Esse éum procedimento científico perfeitamenteaceitável. Este modelo deve ser consideradocomo uma tentativa de explicação, uma hipótese,que será conirmada ou não através de testes dasdeduções feitas a partir dela.
Um teste decisivo era fácil de ser realizado. Ageração F
2 do cruzamento representado na figura
10 era composta por três plantas com a carac-terística dominante lisa para cada planta recessivarugosa. No entanto, se a hipótese fosse verda-deira, as sementes lisas deveriam ser de dois tipose em proporções previsíveis. Assim, para cadasemente com genótipo R (lembre-se que estamosusando ainda o esquema de Mendel e, portanto,não falamos em RR), haveria duas Rr.
Não era possível distinguirvisualmente as sementes R eRr, mas se elas fossem planta-das e ocorresse autofertili-zação de suas flores, a descen-dência daria a resposta. Nestecaso, as plantas com genótipoR produziriam apenas semen-tes lisas, enquanto as plantascom genótipo Rr dariamorigem a sementes lisas erugosas, na proporção de 3:1.Mendel deixou que as plantasF
2 se autofecundassem e obte-
ve o resultado esperado deacordo com sua hipótese.
MODELO PARA
CRUZAMENTO DIÍBRIDO
A figura 11 mostra omodelo de cruzamento di-íbrido já discutido anterior-mente. A linhagem pura deplantas amarelo-lisas foicruzada com a linhagem puraverde-rugosa. Os indivíduosresultantes em F
1 foram
39
Liso - amarelo X Rugoso - amarelo
R V r v
X➤ ➤
RrVv➤ ➤
P1
F2
Fenótipos
Genótipos
Genótipos
Genótiposdos gametas
F1
Fenótipos
Genótipos
Genótiposdos óvulos
➤ ➤
RV rv
Liso - amarelo Liso - amarelo
➤➤
RrVv
RV
rv
rV
Rv
RV
rv
rV
Rv
➤
➤
➤
➤
➤
➤
Genótipos dosgrãos de pólen RRVV
RRVvRRVv
RRvv RrVVRrVV
rrVV
rrVv rrVv
rrvv
RrVvRrVv RrVvRrVv
RrvvRrvv
Figura 11. Modelo de um cruzamento mendeliano diíbrido. Os genótiposda geração P foram representados como Mendel os teria representado. Oquadro de cruzamento mostra a origem de F2 a partir de grãos de pólen eóvulos de F1 (os genótipos como representados atualmente).
uniformes: expressavam o fenótipo dominantedos estados contrastantes das duas caracte-rísticas analisadas.
Na formação de gametas pelos indivíduos deF
1, Mendel assumiu, como no cruzamento
monoíbrido, que cada gameta receberia apenasum tipo de cada uma das duas unidades contras-tantes - R ou r . O mesmo foi assumido em relaçãoa V e v. Neste ponto, outra condição teve que serassumida para explicar os resultados obtidos:deveria haver uma distribuição independente dasunidades de cada par. Assim, cada gameta deveriater ou R ou r, além de V ou v. Deveriam, portanto,ser formadas quatro classes, tanto de grãos depólen quanto de óvulos: RV, Rv, rV e rv. Omodelo também exigia que estas classes deveriamaparecer em igual freqüência - 25 % cada.
Simulação com moedas
Um jogo simples com duas moedas diferentesnos ajudará a entender a origem das quatro classes

de gametas. Cada moeda representará um genee, em uma delas, “cara” representará, porexemplo, o alelo dominante R enquanto “coroa”será o alelo recessivo r. Na outra moeda, “cara”representará o alelo dominante V e “coroa”, oalelo recessivo v. As moedas são então lançadase os resultados anotados. Se arremessossuficientes forem feitos, espera-se 1/4 de “caras”para ambas as moedas (ou seja, a categoria RVacima); 1/4 de “coroas” para ambas as moedas(o que corresponde à categoria rv); 1/4 será“cara” para uma das moedas e “coroa” para aoutra (= Rv) e 1/4 será o contrário (= vR).
Quando há quatro classes de gametas não éprático usar as linhas indicativas utilizadas nafigura 10; assim, na parte inferior da figura 11 háum quadro indicando todas as possíveiscombinações de grãos de pólen e óvulos naprodução de F
2 (para simplificar, todos os
genótipos foram representados como diplóides).Há 16 combinações possíveis e, se considerarmosos fenótipos, 9 das 16 são amarelo-lisas, 3 sãoverde-lisas, 3 são amarelo-rugosas e 1 é verde-rugosa. Isto representa uma proporção de9:3:3:1.
Note que somente os fenótipos amarelo-lisoe verde-rugoso estavam presentes nas geraçõesP e F
1. O modelo predizia, no entanto, que dois
novos tipos de sementes deveriam aparecer:verde-lisas e amarelo-rugosas.
A seguir estão os resultados registrados porMendel para F
2:
Os números observados correspondiam aosobtidos experimentalmente por Mendel e osnúmeros esperados correspondiam à proporçãoexata de 9:3:3:1. A concordância entre os doisvalores é extraordinária, como Weldon e Fishernotariam anos mais tarde.
Outros testes da hipótese
O modelo do cruzamento diíbrido permitiutestes ainda mais requintados para a hipótese. A
40
Fenótipo Obtido Esperado
lisa - amarela 315 313lisa - verde 108 104rugosa - amarela 101 104rugosa - verde 32 35
hipótese predizia que, exceto as 32 sementesverde-rugosas, todas as outras classes consisti-riam de indivíduos geneticamente diferentes,apesar de iguais na aparência. Isto pôde sertestado, plantando-se as sementes F
2, permitindo
a autopolinização das plantas adultas e, então,contando-se as sementes F
3.
Primeiramente, considere as 32 sementesverde-rugosas. O modelo prevê que estasmanteriam suas características, se autofecun-dadas. Tais sementes foram plantadas e 30atingiram a fase adulta. Todas comprovaram serverde-rugosas.
As 101 sementes amarelo-rugosas eram idên-ticas quanto à aparência. Podemos ver, noentanto, a partir do modelo da figura 11, que doisgenótipos estão aí representados dentre os 3/16desta categoria: um dos três é rrVV e os outrosdois são rrVv ( Mendel usaria rV, ao invés derrVV , mas, para facilitar, estou usando a mesmarepresentação da figura 11). Desta forma, umaem cada três destas sementes, o grupo rrVV ,manteria suas características, produzindo apenassementes amarelo-rugosas, por autofecundação.Já o grupo rrVv ( duas das três), deveria produzirsementes em uma proporção de 3 amarelo-rugosas para 1 verde-rugosa. As 101 sementesforam plantadas, sendo que 96 atingiram amaturidade. Destas, 28 (onde o esperado era 32)produziram apenas sementes amarelo-rugosas,enquanto 68 (o esperado era 64) produziramtanto sementes amarelo-rugosas como verde-rugosas em uma proporção de 3:1. Assim, adedução a partir da hipótese foi confirmada.
A mesma análise foi realizada com as sementesverde-lisas. A proporção de 3/16 desta classedeveria ser composta por 1/3 RRvv e 2/3 Rrvv.O 1/3, representado por RRvv, deveria produzirprole idêntica. Os 2/3 restantes deveriam produzirsementes em uma proporção de 3 verde-lisas para1 verde-rugosa. As 108 sementes foram plantadase 102 atingiram a maturidade. O número esperadode indivíduos que originariam descendentes todosverde-lisos era 34, enquanto 68 resultariam naproporção de 3 : 1. Os números observados fo-ram 35 e 67, respectivamente.
O teste mais complexo para a hipótese baseou-se nos 9/16 de F
2 que eram amarelo-lisos. A
distribuição no quadro é a seguinte: 1 dos 9 éRRVV, 2 são RRVv, 2 são RrVV e 4 são RrVv.Assim, somente 1/9, o grupo RRVV, deveria

41
produzir prole ‘pura’. A descendência de RRVvestaria em uma proporção de 3 amarelo-lisas para1 verde-lisa. Os RrVV deveriam produzir 3amarelo-lisas para 1 amarelo-rugosa. E final-mente, os RrVv, que são iguais aos F
1 na figura
11, deveriam resultar na proporção de 9:3:3:1.As 315 sementes foram plantadas e 301 chegaramà fase adulta. O modelo predizia que os númerosesperados em cada classe seriam, na ordem apre-sentada acima, 33, 67, 67 e 134. Por exemplo,1/9 ou 33 sementes deveriam manter suas caracte-rísticas, uma vez que o modelo previa que essenúmero de sementes corresponderia à RRVV.Mendel observou o seguinte resultado: 38, 65,60 e 138.
O fato de F2 resultar em uma F
3 que não difere
significativamente da hipótese, em testes tão preci-sos quanto estes, é um argumento muito forte emfavor da validade da hipótese. Em todos os casosos números observados foram bem próximos dosesperados. E os valores esperados basearam-se naprobabilidade dos gametas se comportarem deacordo com regras precisas. No entanto, os núme-ros observados e esperados nunca foram idênticos.A probabilidade disto ocorrer equivaleria à proba-bilidade de obtermos 5 “caras” e 5 “coroas” em 10lançamentos de moedas.
AS CONCLUSÕES DE MENDEL
Os experimentos de Mendel com cruzamentosde variedades de ervilha e a notável análise queele fez dos resultados levaram a oito importantesconclusões, arroladas a seguir. Deve-se ressaltarque, em 1865, tais conclusões eram válidas paraervilhas e somente para ervilhas. Para ter certeza,Mendel realizou alguns cruzamentos preliminarescom feijão, mas os resultados foram confusos.1. A conclusão mais importante era a de que a
herança parecia seguir regras definidas erelativamente simples. Mendel propôs ummodelo que poderia explicar os dados de todosos seus cruzamentos. Além disto, o modelo tinhaum grande poder de previsão – um objetivo detodas as hipóteses e teorias em Ciência.
2. Quando dois tipos diferentes de plantas eramcruzados, não havia mistura dos estados dascaracterísticas individuais. Nos sete pares deestados contrastantes de caracteres estudados,um estado era dominante e o outro recessivo.Isto significava que em um cruzamento de uma
linhagem pura de plantas com umacaracterística dominante com outra purarecessiva, o híbrido formado seria idêntico aoparental dominante e uniforme na aparência.
3. Já que o híbrido descrito acima era idênticoem aparência ao parental puro dominante, erapossível concluir que não havia relação exataentre genótipo e fenótipo. Assim, o fenótipoliso poderia ser conseqüência tanto dogenótipo RR (Mendel diria R) quanto do Rr.
4. Os fatores da hereditariedade responsáveispelas condições de dominância e recessividadenão eram modificados quando ocorriam jun-tos no híbrido. Se dois destes híbridos fossemcruzados, a descendência seria constituídatanto por indivíduos manifestando o estadodominante da característica quanto por aquelesapresentado o estado recessivo, não havendoevidência de que os fatores hereditáriosresponsáveis por eles fossem modificados porsua associação nos híbridos parentais.Qualquer indivíduo recessivo de F
2 deveria ser
fenotipicamente idêntico à geração recessivaoriginal P.
5. Quando híbridos tais como Rr eramcruzados, os dois tipos de unidade heredi-tária (R e r) segregavam um do outro erecombinavam-se aleatoriamente na fertili-zação. A proporção fenotípica na descen-dência seria de 3 : 1, enquanto que genotipi-camente, usando a nomenclatura atual, seriade 1 RR, 2 Rr e 1 rr . A segregação dosfatores de um mesmo par ficou conhecidacomo “Primeira Lei de Mendel”.
6. Essa proporção somente poderia ocorrer secada gameta recebesse apenas um tipo de fatorde hereditariedade – no caso, R ou r.
7. Quando um cruzamento envolvia dois paresde unidades hereditárias contrastantes, talcomo RrVv x RrVv, cada par se comportavaindependentemente. Isto é, os diferentes tiposde unidades hereditárias associavam-se inde-pendentemente uns dos outros. Então, osgametas podiam ser somente de quatro tipos:RV, Rv, rV ou rv. Desta maneira, todas ascombinações possíveis seriam obtidas, combase estritamente na regra de que cada gametapodia apresentar somente um tipo de cada umdos pares de unidades hereditárias. Asdiferentes classes de gameta estariam emfreqüências iguais. Este fenômeno de associa-

42
ção independente ficou conhecido como“Segunda Lei de Mendel”.
8. A hipótese mendeliana, e sua formulação emum modelo, era tão específica que deduçõespodiam ser feitas e testadas pela observação eexperimentos. Nenhum outro campo daBiologia experimental havia atingido equiva-lente nível de desenvolvimento em 1865.
O trabalho de Mendel não é compreendido
Mas, como vimos, naquela época nenhumbiólogo parecia ter consciência de que esse era ocaso. Certamente, o trabalho de Mendel não seriatão importante, se ele valesse apenas para ervilhas,do mesmo modo que se a descoberta das célulaspor Hooke se aplicasse apenas à cortiça.
O campo do conhecimento relativo aoscruzamentos de plantas estava cheio de dadosque não permitiam conclusões gerais. Mendelescreveu cartas para um grande estudioso naárea, Nägeli, explicando seus resultados. Nägelideve ter considerado os dados para ervilhasapenas como mais um exemplo da enormevariação nos resultados obtidos em experi-mentos de hibridação.
Nägeli sugeriu que Mendel utilizasse outraplanta, a chicória ( Hieracium sp.). Mendel assim
o fez e falhou em encontrar regras consistentespara a herança. (Ocorre que Mendel não estavarealizando os experimentos que acreditava estar).Era extremamente dificil realizar experimentosde hibridação com as pequenas flores deHieracium. Contudo, Mendel acreditou terconseguido em muitos casos, e surpreendeu-secom a falta de uniformidade dos resultados. Oproblema estava, na realidade, com Hieracium,e não com Mendel. Muito depois de sua morte,descobriu-se que, em Hieracium, ocorre um tipode desenvolvimento partenogenético, em que háa formação de um novo indivíduo a partir de umóvulo não fecundado. Assim, nenhuma proporçãouniforme era de se esperar se parte da prole eraresultado de fertilização e parte, de apomixia.
Deste modo, o próprio Mendel passou aacreditar que seus primeiros resultados tinhamaplicação restrita, e o fato é que seu modelo foiignorado nas três últimas décadas do século XIX.Durante esse período, os principais estudiososde hereditariedade abandonaram o paradigma docruzamento experimental e se concentraramprincipalmente no comportamento dos cromos-somos na meiose, mitose e fertilização. Eles acre-ditavam estar construindo uma base física para aherança, e pesquisas posteriores viriam a mostrarque eles estavam corretos.
EXERCÍCIOS
PARTE A: REVENDO CONCEITOS BÁSICOS
Preencha os espaços em branco nas frasesde 1 a 8 usando o termo abaixo maisapropriado.
(a) autofecundação (e) hibridação(b) espécie (f) híbrido(c) fecundação cruzada (g) óvulo(d) grão-de-pólen (h) variedade
1. A estrutura da flor que contém os gametasmasculinos, ou núcleos gaméticos, é o ( ).
2. O ( ) das plantas angiospermas é umaestrutura multicelular onde se forma o gametafeminino, a oofera.
3. Na ervilha ocorre ( ) , isto é, o gametamasculino fecunda o gameta feminino damesma flor.
4. O desenvolvimento do pólen de uma plantano estigma da flor de outra planta é um (a) ( ).
5. A ( ) em ervilha é feita removendo-se asanteras de uma flor, antes de sua maturação e,mais tarde, colocando-se pólen de outra plantasobre o seu estigma.
6. O termo ( ), empregado por Mendel, refere-se ao indivíduo proveniente do cruzamentoentre duas plantas de linhagens diferentes.
7. ( ) era o termo usado para designar plantascultivadas que diferiam entre si quanto a umaou algumas características contrastantes, trans-missíveis aos descendentes.
8. ( ) é o termo usado para designar gruposde populações naturais capazes de se cruzar,produzindo descendência fértil.

43
Preencha os espaços em branco nas frases de9 a 16 usando o termo abaixo mais apropriado.
(a) dominante (d) geração F1
(g) monoíbrido(b) fenótipo (e) geração P (h) recessivo(c) genótipo (f) geração F2
9. As duas linhagens puras com uma ou maiscaracterísticas em seus estados contrastantesque são cruzadas em um experimento genéticoconstituem o (a) ( ).
10. Mendel chamou de ( ) o estado dacaracterística que aparecia em todas as plantasda primeira geração híbrida.
11. O estado da característica que não aparecianos indivíduos híbridos foi denominado porMendel de ( ).
12. A primeira geração híbrida, ou seja, aquelaresultante do cruzamento entre indivíduos devariedades diferentes, é chamada ( ).
13. A descendência resultante da autofecunda-ção da primeira geração híbrida é chamada ( ).
14.O termo ( ) é empregado para designar ascaracterísticas apresentadas por um indivíduo,sejam elas morfológicas, fisiológicas, oucomportamentais.
15. O termo ( ) refere-se à constituição genéticado indivíduo, isto é, aos fatores hereditários,ou genes, que ele possui.
16. Um cruzamento ( ) envolve indivíduos quediferem apenas quanto a um caráter, com doisestados contrastantes.
Preencha os espaços em branco nas frases de17 a 21 usando o termo abaixo mais apropriado.(a) associação dos fatores(b) haplóide(c) diplóide(d) pureza dos gametas(e) segregação dos fatores
17. Gameta é uma célula ( ) pois apresentaapenas um lote de cromossomos.
18. O zigoto, por possuir dois lotes cromos-sômicos, é uma célula ( ).
19. A expressão ( ) significa que um gametacontém apenas um fator para cada caracte-rística hereditária.
20. A ( ) ocorreria, segundo Mendel, naformação dos gametas e seria responsável porsua pureza.
21. Uma das premissas do modelo mendelianode monoibridismo é que a ( ) se dêaleatoriamente na fertilização, por meio dacombinação ao acaso dos gametas masculinose femininos.
PARTE B: L IGANDO CONCEITOS E FATOS
Utilize as alternativas abaixo para completar asfrases de 22 a 26.
(a). 1 : 1 (d). 1 : 1 : 1 : 1(b). 1 : 2 : 1 (e). 9 : 3 : 3 : 1(c). 3 : 1
22. Nos cruzamentos envolvendo apenas umpar de caracteres contrastantes, Mendel obtevena geração F2 indivíduos com característicasdominante e recessiva, na proporção de ( ).
23. Quando Mendel seguiu a herança de duascaracterísticas, isto é, em cruzamentos diíbridos,encontrou na geração F2 indivíduos com ambosos estados dominantes para as duascaracterísticas analisadas , com um dominante eo outro recessivo, com o primeiro recessivo e ooutro dominante e com ambos recessivos,respectivamente, na proporção de ( ).
24. A proporção de ( ) na geração F2
corresponderia, segundo o modelo domonoibridismo, aos genótipos dos indivíduosdominantes puros, híbridos e recessivos,respectivamente.
25. Um organismo heretorozigótico quanto a umpar de fatores formará gametas na ( ).
26. Um organismo duplo-heterozigótico quantoa dois pares de fatores com segregação inde-pendente formará gametas na proporção ( ).
27. Mendel selecionou 22 ( ) de ervilha capazesde se cruzar e produzir descendência fértil, paraseus experimentos de hibridação.a. espécies c . populaçõesb. fatores d. variedades
28. De acordo com o modelo do monoibridismo,os indivíduos da geração F2
a. são todos puros.b. são todos híbridos.c . são metade puros e metade híbridos.d. são 3/4 puros e 1/4 híbridos.
29. Se você tem uma drosófila com umadeterminada característica dominante, que tipode teste poderia ser feito para se determinarse ela é pura (AA ) ou híbrida (Aa)?

Utilize as alternativas abaixo para responder àsquestões 30 e 31.
a. associaçãob. dominânciac. segregação independented. segregação
30. A pureza dos gametas é resultado da ( )dos fatores de cada par na formação dosgametas.
31. Os quatro tipos de gametas que um diíbridoforma resulta da ( ) dos fatores dos dois pares.
PARTE C: QUESTÕES PARA PENSAR E DISCUTIR
32. No que a análise de Mendel diferiu da deseus predecessores que trabalharam comhibridação de plantas?
33. Quais as razões que levaram Mendel aescolher ervilha como material para os seusexperimentos?
34. Qual foi a hipótese levantada por Mendelpara explicar a proporção 3 : 1 obtida emcruzamentos monoíbridos?
35. Como pode ser derivada a proporção9 : 3 : 3 : 1, típica de um cruzamento diíbrido, apartir da proporção 3 : 1, típica de um cruza-mento monoíbrido?
36. Quais das condições necessárias para queo modelo mendeliano fosse válido podiam serconsideradas fatos?
37. Como se explica o fato de Mendel não terencontrado as proporções previstas por suasleis em cruzamentos de plantas de Hieracium?
38. Admitindo-se a segregação independente,quantos tipos de gameta cada indivíduo abaixoproduz?I. AaBbCCdd; II. AaBbcc; III. AAbbCCDDEe
39. Enuncie a primeira e a segunda leis deMendel.
40. Com base nas hipóteses e observações deMendel, esquematize os resultados esperadosnos seguintes cruzamentos:a. ervilha alta homozigótica (dominante) comervilha anã.b. F
1 do cruzamento (a) entre si.
c. F1 do cruzamento (a) com a ervilha anã original.
41. O biólogo francês Cuenot, no início doséculo, cruzou camundongos selvagens de corcinza, com camundongos brancos (albinos). Naprimeira geração todos os indivíduos tinham cor
44
cinza. O cruzamento desses últimos indivíduosentre si produziu uma geração F2 constituídapor 198 camundongos cinzas e 72 brancos.
a. Proponha uma hipótese para explicar essesresultados.
b. Com base na sua hipótese faça um diagramado cruzamento e compare os resultadosobservados com os esperados de acordo como diagrama.
42. Um dos diferentes tipos de albinismo queocorrem na espécie humana é determinado porum fator recessivo a.
a. Do casamento entre uma mulher portadorado fator para albinismo (Aa) e um homem al-bino, qual a proporção esperada de filhos albi-nos?
b. Do casamento entre dois portadores (Aa),qual a proporção esperada de filhos albinos?
43. Em algumas variedades de gado bovino, aausência de chifres é condicionada por um fatordominante (C). Um touro sem chifres foi cruzadocom três vacas. No cruzamento com a vaca I,portadora de chifres, foi produzido um bezerrosem chifres. No cruzamento com a vaca II,portadora de chifres, foi produzido um bezerrocom chifres. No cruzamento com a vaca III, semchifres, foi produzido um bezerro com chifres.
a. Proponha uma hipótese para explicar essesresultados.
b. Com base na sua hipótese faça um diagramado cruzamento e compare os resultadosobservados com os esperados de acordo como diagrama.
44. Uma planta de flores longas e brancas foicruzada com outra de flores curtas e vermelhas.Os descendentes obtidos, todos de floreslongas e vermelhas, foram autofecundadosproduzindo os seguintes tipos de descendente:63 com flores longas e vermelhas,21 com flores longas e brancas,20 com flores curtas e vermelhas e 8 com flores curtas e brancas.
a. Proponha uma hipótese para explicar essesresultados.
b. Com base na sua hipótese faça um diagramado cruzamento e compare os resultadosobservados com os esperados pelo diagrama.

A REDESCOBERTA E A EXPANSÃO DO MENDELISMO
Texto adaptado de:MOORE, J. A. Science as a Way of Knowing -Genetics. Amer. Zool. v. 26: p. 583-747, 1986.
Quarta aula(T4)
Objetivos da Unidade
1. Identificar os motivos que fizeram com que o trabalhode Mendel permanecesse ignorado por 35 anos.
2. Explicar os motivos do antagonismo entre mendelistase biometristas.
3. Conceituar os seguintes termos: alelo, homozigoto,heterozigoto, loco gênico e gene.
4. Resolver problemas sobre variações nas proporçõesmendelianas.
MENDELISMO , UM NOVO PARADIGMA
Muita importância é dada ao fato de que otrabalho de Mendel foi publicado em uma obscurarevista de uma sociedade também obscura, de talmodo que foi esquecido ou desconhecido por 35anos, durante os quais floresceram a Citologia eum enorme interesse pela hereditariedade. Umjulgamento mais acurado, eu acredito, nos levariaà conclusão de que sua publicação foi desconside-rada, e não desconhecida. Essa publicação erado conhecimento de Focke (1881), que a discutiubrevemente em seu criterioso tratado de hibrida-ção em plantas, e também foi mencionada maistarde por Bailey (1895). Como já mencionado,Mendel se correspondia com um dos mais proemi-nentes estudiosos de hereditariedade daquelestempos, Karl Wilhelm von Nägeli, o qual não semostrou impressionado com os resultados doscruzamentos em ervilhas.
A explicação de Bateson, em sua introduçãoà publicação de Mendel (Mendel, 1902, p.2), foia seguinte: “Pode parecer surpreendente que umtrabalho de tal importância não tenha tido reco-nhecimento por tanto tempo, e não tenha sedifundido no mundo científico. É bem verdadeque a revista em que ele foi publicado era poucodivulgada, mas circunstâncias como esta rara-mente tem retardado o reconhecimento de umtrabalho pela comunidade. A causa será, sem
dúvida, encontrada na negligência em relaçãoaos estudos experimentais sobre o problema deEspécie, decorrente da aceitação geral dasdoutrinas darwinianas. O problema da Espécie,conforme concebido por Gartner, Kölreuter,Naudin, Mendel e os demais hibridistas daprimeira metade do século XIX, não mais atraiaos pesquisadores. A questão, acreditava-se, jáhavia sido respondida e o debate concluído.Ninguém mais tinha interesse no assunto [umamudança de paradigma!]. Várias outras linhasde pesquisa haviam sido repentinamente abertase, em 1865, os pesquisadores mais entusiastasnaturalmente consideraram esses novos métodosmais atraentes do que as observações tediosasdos hibridistas, cujos questionamentos tinham,além do mais, levado a resultados inconclusivos.Entretanto, se pretendemos avançar no estudoda hereditariedade e levar adiante o problemade “O que é uma Espécie?” como uma questãodiferente de “Como uma espécie sobrevive?”devemos voltar e retomar a linha de questiona-mento exatamente como Mendel a tomou.”
E, como podemos ver, foi exatamente o queBateson fez.
O trabalho de Mendel com ervilhas não é oúnico exemplo de uma importante descoberta nãocompreendida pela comunidade científica de seutempo. Novos paradigmas dificilmente sãoprontamente identificados e adotados. A maioria
45

dos cientistas de qualquer época, estará ocupadafazendo sua ciência normal, seguindo os para-digmas preexistentes. A dificuldade em mudar oque se faz e como se pensa gera resistência àsnovas idéias e ao empreendimento de novos pro-gramas de pesquisa.
Isto não foi problema para de Vries e Corrensem 1900. A razão pela qual eles entenderam aimportância das conclusões de Mendel foi teremfeito o mesmo tipo de trabalho e desenvolvidohipóteses semelhantes, antes mesmo de lerem apublicação de Mendel. Eles estavam trabalhandono novo paradigma antes de conhecerem seuparadigmático autor.
O mesmo pode ser dito de Bateson. Ele haviaestudado variação e hibridação em plantas poranos e, apesar de nunca ter observado as regulari-dades do modelo mendeliano, ele sabia os tipos deexperimentos que deveriam ser feitos. Considere oparágrafo a seguir, sobre as idéias de Bateson.
Em 11 e 12 de julho de 1899, uma terça e umaquarta-feira, a “Royal Horticultural Society”realizou uma “Conferência Internacional sobreHibridação (cruzamento entre espécies) e sobreCruzamento de Variedades” em Chiswick,Londres. O volume 24 da revista da Sociedadeconsiste do registro da conferência. Assim, temosacesso às opiniões de importantes pesquisadoresem hibridações com plantas, da época imediata-mente antes de Mendel mudar sua Ciência. Amaioria dos artigos da revista descrevemresultados de cruzamentos, mas Bateson preferiuum discurso mais teórico, como podemos notarno trecho seguinte (Bateson, 1900a): “O queprecisamos inicialmente saber é o que acontecequando uma variedade é cruzada com outrasvariedades próximas. Se pretendemos que oresultado tenha algum valor científico, é quaseabsolutamente necessário que a descendência detais cruzamentos seja então examinada estatisti-camente. Deve ser registrado quantos descen-dentes se parecem com cada um dos parentais equantos mostram o caráter intermediário entreeles. Se os parentais diferem em vários carac-teres, a descendência deve ser examinada estatis-ticamente e classificada, considerando cada umdos caracteres separadamente.”
É como se Bateson estivesse aconselhando umestudante de graduação, que atendia pelo nomede Mendel, como planejar o programa de pesquisapara seu Ph.D. (doutorado em filosofia)!
46
Há vários aspectos da história de Mendel quesão interessantes. Um deles é a atenção quase uni-versal que é dada ao cientista que fez a descoberta.Até recentemente, cientistas, especialmentebiologistas, raramente poderiam ter a expectativade fazer fortuna como cientistas – isto é, ganharuma grande soma de dinheiro. A recompensa paraum cientista era o próprio prazer de investigar anatureza e a aprovação de seus pares pela pesquisabem feita e pela formulação de hipóteses arrojadase imaginativas. Hoje, cientistas vêem o trabalho deMendel com admiração. Como poderia ter eleavançado tanto em relação aos paradigmas da épocae feito observações que, logo após sua morte,revolucionariam as Ciências Biológicas?
Outro ponto interessante é que, repetidamentena história, parece que quando o campo está“pronto” a descoberta será feita. Se Mendel nãotivesse nascido, o caminho da Genética não seriamuito diferente. Ao redor de 1900, alguém chega-ria às mesmas conclusões. Apenas aconteceu deserem de Vries e Correns. Tschermak estava tãoperto da resposta que geralmente é incluído comde Vries e Correns como co-redescobridor.Bateson, em mais ou menos um ano, poderia terdescoberto independentemente as regras mende-lianas da herança. Como parece, o progresso daCiência é inevitável.
OPOSIÇÃO INICIAL AO MENDELISMO
No decorrer da história de Mendel, podemos serlevados a acreditar que em 1900, com a publicaçãodos artigos de de Vries e Correns, a “Ciência pura”tinha finalmente triunfado. Mas não foi bem assim.Havia vigorosa, muitas vezes sarcástica, oposiçãoàs conclusões de Mendel (Provine, 1971). Esseduelo científico envolveu principalmente trêsingleses - William Bateson versus Karl Pearson eW. F. R. Weldon, cada lado apoiado por um grupode seguidores. As duas escolas eram fundamen-talmente diferentes em suas propostas. Batesonprocurava informações sobre herança através decruzamentos experimentais. Weldon, Francis Galtone Karl Pearson procuravam aplicar métodos mate-máticos, especialmente estatísticos, aos problemasbiológicos. A oposição destes biometristas é real-mente surpreendente, se lembrarmos que Mendelse baseou na matemática.
A disputa básica começou antes de 1900 eestava relacionada com a temática evolutiva. Mais

47
uma vez era o caso de paradigmas conflitantes.Weldon, Galton, Pearson e outros eram segui-dores de Darwin, acreditando que a evolução dosseres está baseada em alterações filogenéticasgraduais. Em populações naturais a variaçãotende a ser contínua. Quando organizados pelotamanho ou outra característica qualquer, os indi-víduos de uma espécie parecem mostrar umavariação contínua. Não surpreende que se consi-derasse a evolução envolvendo mudanças tãopequenas que, apenas após longo intervalo detempo, seria possível observar qualquer diferença.
A lei da herança ancestral de Galton
Uma importante asserção desta escola davariação contínua era a “lei da herança ancestral”de Galton. Ele considerava a herança na sua tota-lidade e realçava que cada característica hereditáriade um indivíduo não provinha apenas dos pais,mas também dos ancestrais mais remotos. Galton(1897) propôs sua famosa lei, baseando-se emcuidadoso estudo de genealogias de cães basset.
“A lei a ser examinada pode parecer à primei-ra vista muito artificial para ser verdadeira, masuma análise mais acurada mostra a predisposi-ção advinda dessa impressão superficial é infun-dada. O assunto será mencionado novamente,por enquanto a lei deve ser enunciada. Ela dizque os dois genitores contribuem ambos comaproximadamente metade, ou (0,5) do total daherança da descendência; os quatro avós, comum quarto, ou (0,5)2; os oito bisavós, com umoitavo, ou (0,5)3, e assim por diante. Assim, asoma da contribuição ancestral é expressa pelasérie {(0,5) + (0,5)2 + (0,5)3 + ....)}, que, sendoigual a 1, dá conta de toda a hereditariedade.”
Assim, traços de nossos mais remotos ances-trais poderiam ser encontrados em nós e estaherança passada frearia mudanças repentinas. Asexigências darwinianas de um mecanismo paramudanças imperceptíveis e lentas na evoluçãoestariam então satisfeitas.
Mas, como funcionaria a Lei de Galton? Atual-mente, a noção de que nossos genes são herdadosapenas dos nossos pais, exatamente metade decada, está tão fixada em nossas mentes, que nãopodemos imaginar um mecanismo de herança deancestrais veneráveis, sobrepondo-se à paterna.A resposta é que Galton estava se referindo afenótipos, e não genótipos. Já era bem conhecido
que um indivíduo pode expressar certas caracte-rísticas fenotípicas que seus pais não expressam.
Variação contínua e descontínua em debate
A noção de variação contínua em evolução e,claro, em herança, foi contestada por Bateson eoutros. Ele havia estudado evolução e hereditarie-dade por muitos anos. Em 1894, produziu oimenso volume Materials for the Study of Varia-tion Treated with Especial Regard to Disconti-nuity in the Origin of Species (“Materiais para oEstudo da Variação voltado especialmente paraa Descontinuidade na Origem das Espécies”). Eledesejava descobrir se as evidências sugeriam quea evolução é produto de variação contínua oudescontínua, e concluiu que esta última é possível.Sobre isso, escreveu, em 1900: “Nós aprendemosque Evolução é um processo muito lento, aconte-cendo por passos infinitesimais. Na horticulturaé raro acontecer algo desse tipo .... O melhora-mento de organismos anda a galope. Nesse caso,portanto, em que pode ser mostrado que a varia-ção dos organismos é descontínua, não é maisnecessário supor que para sua produção foramnecessárias inúmeras gerações de seleção eacúmulo gradual das diferenças, e o processode Evolução torna-se assim mais fácil de serentendido. De acordo com o que pode ser descritocomo a visão mais aceita, esse processo consistena transição gradual de um tipo normal para outrotipo normal. Essa suposição implica na hipótesequase impossível de que cada tipo intermediárioapareceu sucessivamente entre as duas formasextremas. Se existir descontinuidade essa hipóteseé totalmente dispensável.”
Não é de admirar que Bateson acreditou ser oparadigma mendeliano tão aceitável. Diferençashereditárias poderiam ser marcantes, isto é, asvariações pareciam ser descontínuas. ParaBateson e seus seguidores o modelo de Mendelera compatível com o seu paradigma.
(Como um aparte, seria interessante notar queo motivo deste debate ainda hoje é pertinente.Alguns evolucionistas acreditam que o padrãoprincipal seguido pela evolução é baseado emmudanças pequenas. Outros acreditam ser opadrão comum de pequenas mudanças, ocor-rendo durante longos períodos de tempo, seguidaspor mudanças bruscas de curta duração –equilíbrio pontuado. Parte da confusão é conse-

48
qüência da dificuldade de definição quanto aotamanho da mudança, para considerá-la grandeou pequena. Além disso, o que é exatamente umlongo ou um curto período de tempo? Algunscurtos períodos são de 10 mil anos. A respostaserá, provavelmente, que algumas linhagenscaracterizam-se por mudanças lentas e relativa-mente constantes ao longo das eras, outras porperíodos de estase e de saltos, e outras pormudanças pequenas em períodos muito longosde tempo. As duas escolas opostas seriam, então:“Evolução Gradual” e “Evolução em Saltos”).
A veemência com que estes debates sederam indica claramente que os partidários doantigo paradigma da variação contínua senti-ram-se ameaçados. Mesmo aqueles que, dooutro lado, viram a proposta de Mendel comouma grande promessa, isto é, a idéia da variaçãodescontínua, tinham que admitir que as conclu-sões de Mendel não poderiam explicar osresultados de cruzamentos para todos os orga-nismos e para todas as características.
A conseqüência foi que Bateson e os hibri-distas continuaram a realizar experimentos quemostravam até que ponto os princípios de Mendelpoderiam ser aplicados, e Weldon e outros conti-nuaram a apontar que nem tudo poderia ser expli-cado a partir da hipótese original de Mendel.
Weldon (1902) resumiu as conclusões deMendel e escreveu: “É evidente a importânciade se testarem essas afirmações incomuns pormeio de um estudo cuidadoso dos resultadosnuméricos, e pela aplicação de tais testes sempreque possível. Tenho a impressão que ao negligen-ciarem essas precauções alguns escritoresdeixaram de apreciar como os resultados deMendel concordam de modo surpreendentementeconsistente com sua teoria.”
A concordância entre o esperado e o obtidopor Mendel
Weldon submeteu as proporções obtidas porMendel a testes estatísticos e concluiu que: “se osexperimentos fossem repetidos uma centena devezes, nós esperaríamos obter um resultado piorcerca de 95 vezes, ou as chances contra um resul-tado igual ou melhor que esse é de 20 para 1 .”
Anos mais tarde, ainda outro cientista incli-nado à área matemática, R. A. Fisher (1936), veioa considerar a questão dos dados de Mendelserem “tão bons”. De qualquer modo, havia abun-
dante confirmação disto. Sinnott e Dunn (1925,p. 47) relacionaram as proporções obtidas porMendel e por outros seis experimentadores quetentaram conferir os resultados dele, entre 1900e 1909. No caso de amarelo x verde, por exemplo,o número total de sementes foi 179399. Destas,134707 eram amarelas (75,09%) e 44692(24,91%) eram verdes. Mendel havia registrado75,05% e 24,95%, respectivamente. Aparente-mente, não era muito difícil a obtenção de dados“tão bons”.
Mendel nunca publicou seu trabalho completo.Seu artigo de 1865 foi baseado em palestras epareceu razoável a ele selecionar os dados doscruzamentos que melhor ilustravam a hipóteseque estava propondo. Em conferências, normal-mente, os cientistas não descrevem todos os seusexperimentos, nem apresentam todos os seusresultados – mesmo quando parecem fazê-lo.Então, Mendel também estava seguindo os proce-dimentos de sua época, não os atuais. Mas, apóstodas as verificações e discussões, comprovou-se que Mendel estava certo.
S. Wright (1966) estudou novamente os dados,e concluiu “Estou convicto, entretanto, de quenão houve esforço deliberado em falsificação”.
A questão da dominância e da recessividade
Weldon continuou a questionar a noção dedominância e recessividade. Ele cometeu o erro,que Mendel insistiu em evitar, de assumir que umtal fenótipo implica sempre em um mesmo genó-tipo. Weldon conhecia muitas variedades deervilhas que tinham características semelhantesàquelas que Mendel usou. Mas elas nem sempredavam os mesmos resultados quando cruzadas.Weldon parecia não entender que um determinadofenótipo em uma variedade poderia não ter amesma base genotípica que um fenótipo aparente-mente idêntico, em outra variedade. Além disso,Weldon parecia não entender a importância de seconhecer previamente a constituição dos pais -se, por exemplo, o fenótipo era produzido porum genótipo homozigótico ou heterozigótico. Noentanto, foi capaz de citar casos onde a dominân-cia não era completa e os “híbridos” eram interme-diários. Isto seria comprovado em vários casos,como Correns e outros iriam descobrir mais tarde.
Weldon atacava a idéia de que as “afirmaçõesde Mendel eram universalmente válidas” eresumiu, “Eu penso que nós só podemos concluir

49
que aquela segregação dos caracteres dassementes nos cruzamentos de ervilha não é deocorrência universal, e que quando ocorre, podeou não seguir as leis de Mendel. A lei da segrega-ção, como a lei da dominância, parece portantose aplicar a raças com ancestralidades particu-lares... . O erro fundamental que invalida todotrabalho baseado no método de Mendel é negli-genciar a ancestralidade, e tentar considerar oefeito completo de um genitor particular sobre adescendência, como devido à existência nogenitor de caracteres estruturais particulares;enquanto que os resultados contraditóriosobtidos por aqueles que observaram a descen-dência de genitores aparentemente idênticos emcertos caracteres mostram claramente que nãoapenas os pais, mas também sua raça, isto é,sua ancestralidade, devem ser levados em contaantes que os resultados de um cruzamento entreeles possam ser previstos.”
A última objeção é um tanto estranha.Mendel tomou muito cuidado para se assegurarque suas variedades originais sofressem auto-fertilização. Esta é uma das razões pelas quaisele obteve sucesso, quando tantos falharam. Oúltima frase da citação mostra que Weldonainda achava que a Lei da Herança Ancestralde Galton deveria ser considerada.
O mendelismo estava em clara competiçãocom a Lei de Galton e, portanto, não surpreendeque os biometristas estivessem ansiosos paracontestar seus resultados e conclusões – foi issoque Weldon fez. Algumas indicações da altaconsideração desse grupo por Galton estãopresentes nas seguintes citações de Pearson(1898): “Em resumo, se a lei do Sr. Galton podeser firmemente estabelecida, ela é uma soluçãocompleta, em qualquer caso, para todo o pro-blema de hereditariedade. Ela baseia a questãoda herança em duas constantes, que podem deuma vez por todas ser determinadas; daí suaimportância fundamental.”
E Pearson fecha seu artigo com esta arrogantenota: “No momento eu meramente registrariaminha opinião pessoal que, com todas as reservasdevidas, me parece que a Lei da Herança An-cestral irá provar ser uma das mais brilhantesdescobertas do Sr. Galton; é altamente provávelque é seu enunciado descritivo simples que reúnesob o mesmo foco todas as complexas linhas dahereditariedade. Se a evolução darwiniana é
seleção natural combinada com hereditariedade,então a afirmação simples que abarca todo ocampo da hereditariedade deverá se tornar parao biólogo uma referência quase do mesmo nívelque a lei da gravitação é para o astrônomo.”
Bateson versus Weldon
Esta área desenvolveu-se muito rapidamenteno anos 1900 - 1903. O artigo de deVries (1900)sobre a ‘redescoberta’ havia sido submetido àpublicação em 14 de março de 1900 e o deCorrens (1900) em 26 de abril. Essas publicaçõesgeraram muita discussão. Logo depois, Bateson(1900b) discutiu tais artigos em um encontro da“Royal Horticultural Society”. Subseqüente-mente, foi feita uma tradução do artigo de Mendel(1902), possibilitando uma análise por todo omundo científico – poucas bibliotecas deveriampossuir o original de 1865. O artigo anti-Mendelde Weldon (1902) foi recebido pelos editores daBiometrika em 9 de dezembro de 1901.
Bateson iniciou imediatamente a produção deum livro, Mendel’s Principles of Heredity: A de-fense (“Os Princípios da Hereditariedade deMendel: Uma Defesa”) (1902). Este livro nãoapenas apresenta uma tradução dos artigos deMendel sobre Pisum e Hieracium, como tambémdiscute seus dados com clareza.
Bateson, então, documenta a visão errônea edistorcida de Weldon sobre o trabalho de Mendel.Weldon, vendo seu paradigma severamente contes-tado, respondeu-lhe de uma maneira que gostaría-mos de acreditar não ocorresse em Ciência. Eletrouxe o descrédito para si mesmo e para os biome-tristas como um grupo. Bateson conclui com oseguinte: “Eu acredito que o que escrevi convenceuo leitor de que nós estamos [como conseqüênciado trabalho de Mendel] finalmente começando anos mover. O Professor Weldon declara que ele“não deseja depreciar a importância dasrealizações de Mendel”; que ele deseja “simples-mente chamar a atenção para uma série de fatosque para ele parecem sugerir linhas frutíferas deinvestigação”. Eu me aventuro a auxiliá-lo nessepropósito, pois eu penso que, da maneira como eleestá desamparado - citando uma frase de HoraceWalpole, está tão próximo de acender fogo compano molhado quanto de estimular o interesse pelasdescobertas de Mendel com sua apreciaçãohumorada. Se eu contribui um pouco para essacausa, meu tempo não foi perdido.

Nestas páginas eu apenas toquei as fronteirasdesse novo campo que está se descortinandodiante de nós, de onde em um período de dezanos deveremos olhar para trás, para o cativeirodos dias atuais. Logo, toda a ciência que lidacom animais e plantas estará repleta de desco-bertas que o trabalho de Mendel tornou possí-veis. O criador, seja de plantas ou de animais,não mais se desgastando nos velhos caminhostradicionais, será superado apenas pelo químicoem recursos e em capacidade de previsão. Cadaconcepção de vida da qual fizer parte a heredita-riedade - e qual delas estaria isenta? - deverámudar antes do desencadeamento de fatos queestá por vir.”
A previsão de Bateson sobre o que se veriadez anos mais tarde estava correta - Morgan esta-beleceria as bases para um desenvolvimentoespantoso em Genética.
Bateson, o campeão e o profeta, fez muito paraproteger e promover o mendelismo em sua faseinicial. Ele agiu de maneira similar a seu compatriota,Thomas Henry Huxley, que meio século antes, haviasido vigoroso e efetivo defensor do darwinismo.
DEFININDO ALGUNS TERMOS GENÉTICOS
No ano de 1902 surgiu uma outra publicaçãode fundamental importância - a primeira dascomunicações ao Comitê de Evolução da RoyalSociety, realizada por Bateson e pela senhoritaSaunders (1902). Em 1897 eles deram início auma série de cruzamentos em uma grandevariedade de plantas e animais. Seu objetivo inicialera aprender mais sobre os fenômenos de herançacontínua e descontínua, e também sobre o fenô-meno da “predominância”, que mais tarde seriadenominado dominância. Naquela época elespensaram que, “Do que foi averiguado até omomento sobre o fenômeno da hereditariedade,não se pode evitar a inferência de que não háuma lei universal, mas que, por meio do estudode diversos casos específicos, diferentes leispodem ser descobertas.”
Um empreendimento muito grande para Dar-win, Nägeli, Weismann e Galton! Por outro lado,Bateson e Saunders, antes de publicarem seusresultados, perceberam que “todo o problema dahereditariedade estava sofrendo uma completarevolução”. Essa visão permitiu que eles usassem oparadigma mendeliano para explicar seus resultados.
50
Bateson (em Bateson e Saunders, 1902)forneceu-nos algumas das terminologias básicaspara a Genética mendeliana: “Esta pureza dalinhagem germinativa, e sua incapacidade detransmitir simultaneamente as duas caracterís-ticas antagônicas, é o fato central do trabalhode Mendel. Nós chegamos assim à concepçãode unidades de caráter existentes em pares anta-gônicos. Nós propomos para tais caracteres adenominação de alelomorfos, e para o zigotoformado pela união de gametas portadores dealelomorfos antagônicos, a denominação deheterozigótico. De modo semelhante, o zigotoformado pela união de gametas portadores dealelomorfos semelhantes deve ser chamadohomozigótico”. Mais tarde, o termo alelomorfofoi encurtado para alelo.
Distinguindo gene e alelo
Nesse ponto irei explicar como planejo usaros termos gene, loco gênico, e alelo. Osprofessores com larga experiência no ensino deGenética em cursos introdutórios sabem o quantoé difícil para os estudantes lidarem com essaterminologia. Esta dificuldade se deve mais aomodo inexato como muitos geneticistas usamestes termos, do que à falta de competência doaluno. E, atualmente, apareceu um complicadoradicional: quanto mais se sabe a respeito das basesmoleculares de um loco gênico, mais complicadose torna o conceito de gene. No momento, vamosignorar o presente e discutir os genes como eleseram concebidos nos Anos Dourados da GenéticaClássica: pequenas contas de um colar – naverdade, não exatamente isso.
O principal problema é o uso freqüente dostermos alelo e gene como sinônimos. Eu tentareinão fazer isso, mas como muitos de meus amigosgeneticistas, eu poderei algumas vezes incorrernesse erro. Gene será a menor porção do cromos-somo (os átomos da hereditariedade!) que produzum efeito definido que, é claro, precisa ser detec-tável (ou nós nunca poderiamos saber de suaexistência). A posição que o gene ocupa nocromossomo será seu loco. Alelos serão as dife-rentes variantes detectáveis de um dado gene.Cada gene deve ter pelo menos dois alelos – deoutro modo nós não saberíamos de sua existência.Um gene revela sua existência quando ele mutade tal modo que o novo alelo mutante tenha umefeito detectável.

51
VARIAÇÕES NAS PROPORÇÕES
MENDELIANAS
Os resultados dos cruzamentos entre varie-dades de ervilhas - dominância e recessividade,segregação e segregação independente, com suasconse-qüências, ou sejam, a proporção 3 : 1 emF
2 de um cruzamento monoíbrido e a proporção
9 : 3 : 3 : 1 no F2 de um cruzamento diíbrido -
exibiram um alto grau de uniformidade e, porisso, levantaram a questão da universalidadedos resultados.
Um caso típico de monoibridismo
Em sua primeira comunicação ao Comitê deEvolução, Bateson e Saunders (1902) descreve-ram vários cruzamentos, muitos dos quais inicia-dos antes deles tomarem conhecimento dotrabalho de Mendel. Saunders descreveu seusexperimentos com plantas de espécies selvagensdo gênero Lychnis. Algumas das espécies sãopubescentes, isto é, com pêlos, e outras sãoglabras, isto é, sem pêlos.a. Cruzamentos de plantas pubescentes com
plantas glabras produziram em F1 1006 plantas
pubescentes e nenhuma glabra.b. Quando indivíduos F
1 foram cruzados entre
si, obteve-se um F2 constituído por 408 plantas
pubescentes e 126 glabras. Esses resultadospodem ser explicados assumindo-se a existên-cia de um gene condicionante do caráter, quese apresenta sob a forma de um alelo domi-nante, condicionante do traço pubescente, ede um alelo recessivo, condicionante do traçoglabro. Essa conclusão se baseia no fato de ocaráter glabro não ter aparecido em F
1, mas
ter reaparecido em 1/4 dos indivíduos de F2.
c. Quando um indivíduo F1 foi cruzado com uma
planta pubescente pura, a descendência continha41 plantas pubescentes e nenhuma glabra.
d. Quando um indivíduo F1 foi cruzado com
uma planta glabra pura, a descendênciaobtida era formada por 447 plantas pubes-centes e 433 glabras.
Esses tipos de cruzamento, em que o indivíduoé cruzado com um ou outro dos tipos parentaisrecebe o nome de retrocruzamento. O fato deno retrocruzamento com o parental recessivo (d)ter sido produzido 50% de indivíduos pubescentese 50% glabros corrobora a hipótese da produção
de dois tipos de gameta (nas mesmas proporções)pela geração F
1, a qual é, portanto, formada por
indivíduos heterozigóticos.Com base nesse raciocínio, quando se deseja
determinar o genótipo de um indivíduo comfenótipo dominante faz-se o cruzamento dele comum indivíduo com fenótipo recessivo. No casodos descendentes serem todos de fenótipo domi-nante, conclui-se que o indivíduo em teste formouapenas um tipo de gameta, sendo, portanto,homozigótico. No caso de se obter os dois fenó-tipos na progênie, conclui-se que o indivíduo emteste formou dois tipos de gameta, sendo,portanto, heterozigótico. Esse tipo de cruza-mento, com o intuito de se determinar o genótipode um indivíduo, é chamado cruzamento-teste.
Herança da forma da crista em Gallus gallus
Na mesma publicação Bateson descreveu seusprimeiros experimentos com Gallus gallus. Eleestudou muitos tipos de características, incluindoa forma da crista desses galináceos. (Fig. 12)
Um dos tipos de crista era ervilha e o outro,simples.a. Quando ervilha foi cruzada com simples, todo
o F1 apresentou crista ervilha.
Figura 12. Tipos de crista em Gallus gallus: A. cristasimples; B. crista ervilha; C. crista rosa; D. crista noz.
A
B
C
D

b. Quando indivíduos F1 foram cruzados entre si,
a descendência foi composta por 332 indivíduoscom crista ervilha e 110 com crista simples.
Esses resultados podem ser explicados assu-mindo-se a existência de um gene condicionante docaráter, que se apresenta sob a forma de um alelodominante, condicionante de crista ervilha, e de umalelo recessivo, condicionante de crista simples.
Em uma segunda publicação (Bateson et al.,1905), Bateson relatou outros experimentos sobreherança da forma da crista em Gallus gallus.c. Quando indivíduos com um terceiro tipo de
crista, denominada rosa, foram cruzadas comindivíduos de crista simples obteve-se um F
1
totalmente composto por rosa.d. Quando os F
1 foram cruzados entre si, obteve-
se um F2 formado por 221 rosa e 83 simples.
e. Quando indivíduos F1 foram cruzados com
indivíduos de crista simples, a descendência foicomposta de 449 rosa e 469 simples. (Fig. 13)Esses resultados podem ser explicados assu-
mindo-se a existência de um gene condicionante docaráter, que se apresenta sob a forma de um alelodominante, condicionante de crista rosa, e de umalelo recessivo, condicionante de crista simples.
Assim, rosa e ervilha são ambos dominantessobre simples. Mas como pode haver três alelos:simples, rosa e ervilha?
52
Rosa X Simples
RR rr
➤ ➤
R r
X
➤ ➤
➤ ➤
R
r
Rosa
Rr
RosaRR
Rosa Rr
Rosa Rr
simples rr
➤
Rosa
Rr
➤ ➤
➤
P1
F2
Fenótipo
Fenótipose
Genótipos
Genótipo
Genótipo dosgametas
F1
Fenótipo
Genótipo
Genótipo dosgametas
R
r
Ervilha X Simples
EE ee
➤ ➤
E e
X
➤ ➤
➤ ➤
E
e
Ee
EE
Ee
Eesimples
ee
➤
Ee
➤ ➤
➤
P1
F2
Fenótipo
Fenótipose
Genótipos
Genótipo
Genótipo dosgametas
F1
Fenótipo
Genótipo
Genótipo dosgametas
E
e
Ervilha Ervilha
Ervilha
Ervilha
Ervilha
Figura 13. Diagramas genéticos de cruzamentos entre galináceos com diferentes tipos de crista.
f. A situação se tornou mais complicada com oresultado do cruzamento entre animais de umalinhagem pura de crista ervilha com animaisde uma linhagem pura de crista rosa. A geraçãoF
1 foi uniforme mas todos os animais
apresentavam um tipo de crista diferente dados tipos parentais – crista noz. Esse tipo decrista já era conhecido de outros cruzamentos.
g. Quando os F1 com crista noz foram cruzados
entre si, obteve-se, em F2, 99 noz, 26 rosa, 38
ervilha e 16 simples.Pode-se perceber que a proporção em F
2 é
aproximadamente 9 : 3 : 3 : 1. Esta proporção podeser tomada como significando o envolvimento dedois genes cada um deles com um par de alelos.Estes alelos estão cumprindo as regras mendelianasde dominância, segregação e segregação indepen-dente. A situação é enigmática apenas porque am-bos os pares estão afetando a mesma característica,a forma da crista. (Figs. 13 e 14)h. Quando indivíduos noz foram cruzados com
simples, a descendência foi de 139 noz, 142rosa, 112 ervilha e 141 simples.
Esse foi um cruzamento teste, onde umindivíduo recessivo (crista simples) foi cruzadocom um indíviduo cujo genótipo se desejadeterminar (crista noz). O fato de terem sidoproduzidos quatro tipos de descendente nas

53
mesmas porcentagens indica que o indivíduo emteste produziu quatro tipos de gameta na propor-ção de 1 : 1 : 1 : 1. Isso corrobora o envolvimentode dois genes com segregação independente.Casos como esse, em que dois ou mais genesinteragem na determinação de um mesmo caráter,são conhecidos como interação gênica.
Herança da cor da flor em Salvia
Em seu segundo trabalho (Bateson et al.,1905), Bateson apresentou os resultados de várioscruzamentos com plantas de sangue-de-adão(gênero Salvia). Foram usadas linhagens comflores rosa e branca.a. Quando rosa é cruzado com branco todo o F
1
é violeta.b. Em um cruzamento entre plantas F
1 foram
obtidas 59 plantas de flores violetas, 25 rosase 34 brancas. Em outro cruzamento, foramobtidas 225 violetas, 92 rosas e 114 brancas.
Note que o F1 apresenta um fenótipo diferente
dos de ambos os tipos parentais. Com base na
Ervilha X Rosa
EErr eeRR
X➤ ➤
➤ ➤
P1
F2
Fenótipos
Genótipos
Genótipos
Genótiposdos gametas
F1 Fenótipos
Genótipos
Genótiposdos óvulos
➤ ➤
Er eR
Noz
➤➤
EeRr
ER
er
eR
Er
➤
➤
➤
➤
➤
➤
Genótipos dosespermatozóides
Noz
EeRr
ER
Er
eR
er
EERR
EERr EERr
EErrEeRR EeRR
EeRr EeRr EeRr EeRr
Eerr EerreeRR
eeRr eeRr
eerr
Figura 14. Diagrama genético de um cruzamento entre galináceos de crista
ervilha e de crista rosa, mostrando a produção de indivíduos de crista noz.
proporção fenotípica obtidaem F
2 pode-se explicar esse
caso supondo que a cor da florseja determinada por um únicogene com dois alelos. Umaplanta homozigótica para umdos alelos produz flor rosa, ahomozigótica para o outroalelo produz flor branca e aheterozigótica produz florvioleta [Nota do tradutor :Posteriormente, este e outroscasos semelhantes, no quaisa proporção fenotípica era1 : 2 : 1 e não 3 : 1 passaram aser denominados de padrão deherança monofatorial comcodominância]
Herança da cor da flor emLathyrus e em Matthiola
Os cruzamentos a seguirmostraram outra variação dasproporções mendelianas e sãomais difíceis de seremcompreendidos.
Bateson e colaboradores(1906) haviam realizado numerosos cruzamentoscom ervilhas-de-cheiro (gênero Lathyrus) e comcrucíferas (gênero Matthiola), obtendo em am-bos os casos resultados confusos.a. Quando duas variedades diferentes com flores
brancas foram cruzadas, todas as plantas deF
1 foram coloridas.
b. Quando os F1 foram cruzados entre si, pareceu
à primeira vista, que havia um número igualde flores brancas e coloridas em F
2. Entretanto,
estudos posteriores mostraram que a propor-ção em F
2 era na verdade de 9 coloridas para
7 brancas.A figura 15 apresenta o diagrama de Bateson
para os resultados de F2. Ele e seus co-autores
concluíram que a cor branca das flores das duasvariedades originais não era determinada pelomesmo genótipo. Um genótipo era CCrr e o outroccRR. Para que as flores fossem coloridas eranecessário que a planta possuísse pelo menos umC e um R. Em F
1, por essa hipótese, todos seriam
CcRr, e portanto, coloridos. Casos como esse,em que alelos de um gene mascaram a expressão

54
Branca X
X➤ ➤
CcRr➤ ➤
P1
F2
Fenótipos
Genótipos
Genótipos
Genótiposdos gametas
F1 Fenótipos
Genótipos
Genótiposdos óvulos
➤ ➤
Cr cR
Colorida
➤➤
CR
cr
cR
Cr
CR
cr
cR
Cr
➤
➤
➤
➤
➤
➤
Genótipos dosgrãos de pólen CCRR
CCRrCCRr
CCrr CcRRCcRR
ccRR
ccRr ccRr
ccrr
CcRrCcRr CcRrCcRr
CcrrCcrr
Bran
Colorida
CcRr
CCrr ccRR
Figura 15. Diagrama que mostra a natureza da razão 9 : 7 em F2 de umcruzamento entre variedades de ervilha-doce. O estado colorido do carátercor da flor aparece apenas quando os alelos C e R se encontram. Cadaquadro corresponde a um zigoto e as letras em seu interior mostram seugenótipo. Os quadros escuros representam as plantas com flores coloridase os claros, as com flores brancas.
de outro gene, são conhecidos como casos e inte-ração gênica com epistasia.
Esses poucos exemplos de cruzamentos gené-ticos indicam o tipo de problemas que se podeencontrar em estudos genéticos. Há inúmerosoutros, é claro. Os livros indicados na bibliografiacomplementar trazem diversos exemplos. Muitosdos problemas nestes livros apresentam apenasas proporções observadas e não os números reaisde indivíduos em cada classe fenotípica. Terámaior valor se os estudantes manipularem osdados reais porque eles podem chegar a umaconclusão importante; quando somente asproporções são dadas, esta conclusão importantejá foi alcançada.
CONCLUSÃO
Este quadro da Genética dosprimeiros anos do século XXnos conscientizou da comple-xidade e confusão que existianaquela época. Aqueles quedesejavam que o mundo vivose comportasse como o deli-neado pelas ervilhas de Mendellogo verificaram que isto nãoera verdadeiro. Isto não signifi-ca que a história original deMendel estivesse “errada”. Istosignifica apenas que ela estavaincompleta e que estava sendosubstituída por uma compre-ensão mais profunda danatureza da hereditariedade.Nenhuma das regras originaisde Mendel mostrou-se verda-deira para todos os casos.
Pode ser argumentado queo significativo progresso daGenética foi baseado numaatitude que pode ser vista como“não-científica”. Ou seja, desdeo momento em que as hipótesesde Mendel se tornaram conhe-cidas, ficou claro que elas nãose aplicavam a todos os organis-
mos. Entretanto, os que realmente acreditavam nelasignoraram as exceções e lentamente encontraramexplicações para esses casos que podiam sercolocadas em termos originais do mendelismo. Àmedida que o conhecimento sobre experimentos decruzamentos em diferentes espécies aumentava, foipossível expandir a teoria para acomodar os novosresultados. Foi possível compreender mais e maisas exceções.
Descobriu-se que alguns dos problemas maisdifíceis tinham base cromossômica. Um dessesproblemas foi o dos “cromossomos acessórios” ou“cromossomos X”. Para discutir esse casoprecisamos ver agora o que os citologistas estavamfazendo nos primeiros anos do século vinte.

55
PARTE A: REVENDO CONCEITOS BÁSICOS
Preencha os espaços em branco nas frases de1 a 6 usando o termo abaixo mais apropriado.
(a) biometrista (d) hibridista(b) evolução gradual (e) variação contínua(c) evolução em saltos (f) variação descontínua
1. A escola ( ) se caracterizava por procurarobter informações sobre hereditariedade pormeio de cruzamentos experimentais.
2. A escola ( ) se caracterizava por procurarcompreender hereditariedade por meio daaplicação de métodos matemáticos.
3. Um caráter com ( ) apresenta diferençaspequenas e graduais entre os indivíduos deuma população.
4. Um caráter com ( ) apresenta diferençascontrastantes, facilmente identificáveis, entre osindivíduos de uma população.
5. Pequenas mudanças, lentas e relativamenteconstantes ao longo do tempo caracteriza a ( ).
6. Períodos de mudanças lentas e graduaisseguido de períodos curtos de mudançasbruscas caracteriza o ( ).
Preencha os espaços em branco nas frases de7 a 10 usando o termo abaixo mais apropriado.
(a) alelo ou alelomorfo (c) loco(b) gene (d) mutação
7. ( ) pode ser definido como a porção do cro-mossomo (um segmento da molécula dahereditariedade) que produz um efeitodetectável no indivíduo.
8. Uma alteração hereditária em umacaracterística é chamada ( ).
9. Cada uma das formas detectáveis de um ( )é chamada ( ).
10. A posição que um determinado ( ) ocupano cromossomo é seu (sua) ( ).
Preencha os espaços em branco nas frases de11 a 15 usando o termo abaixo mais apropriado.
(a) dominância(b) interação gênica com epistasia(c) heterozigótico
EXERCÍCIOS
(d) homozigótico(e) interação gênica sem epistasia(f) codominância
11. O zigoto, e conseqüentemente o indivíduo,resultante da união de gametas portadores deum mesmo tipo de alelo é chamado ( ).
12. O zigoto, e conseqüentemente o indivíduo,resultante da união de gametas portadores dealelos diferentes de um mesmo gene édenominado ( ).
13. O fenômeno de dois ou mais genes atuaremna determinação de uma mesma característicaé chamado ( ).
14. O fenômeno de a expressão de um alelode um gene mascarar o efeito de um alelo deum outro gene é chamado ( ).
15. O fenômeno de um alelo de um genemascarar o efeito de um outro alelo do mesmogene é chamado ( ).
PARTE B: L IGANDO CONCEITOS E FATOS
Indique os termos que completam ourespondem as questões de 16 a 28.
16. Para Weldon e Pearson, seguidores deGalton, o mendelismo era incompatível com ateoria darwinista e por isso o combatiam. Essepensamento era defendido pela chamada escolaa. evolucionista. c . hibridista.b. biometrista. d. darwinista.
17. A “lei da herança ancestral” de Galton dizia quea. as características adquiridas eram hereditárias.b. as regras da herança podiam ser expressasmatematicamente pelas fórmulas 3:1 e 9:3:3:1.c . a contribuição ancestral na formação de umindivíduo era de 1/2 dos pais, 1/4 dos avós, 1/8dos bisavós e assim por diante.d. as mudanças hereditárias eram lentas egraduais e dependiam da ancestralidade.
18.Para Bateson a explicação mendeliana paraa hereditariedade não se contrapunha aodarwinismo, pois o processo de evolução po-dia envolver característicasa. adquiridas.b. com variação contínua.c . com variação descontínua.d. com efeito aditivo.

Utilize as alternativas abaixo para completar asfrases de 19 e 20.a. dominância c. interação gênicab. epistasia d. variação contínua
19. Em galinhas, como observado por Batesone colaboradores no início do século, ocruzamento entre indivíduos de crista nozproduz descendência com crista tipo noz, rosa,ervilha e simples, na proporção de 9:3:3:1,respectivamente. Trata-se de ( ).
20. Em ervilha-de-cheiro, Bateson e colabora-dores obtiveram, no cruzamento entre plantashíbridas com flores coloridas, plantas com florescoloridas e plantas com flores brancas naproporção de 9:7. Trata-se de ( ).
21. O cruzamento de um indivíduo com fenótipodominante com outro de fenótipo recessivo,com o objetivo de se determinar o genótipo doprimeiro, é denominadoa. cruzamento teste. c. monoibridismo.b. diibridismo. d. retrocruzamento.
Utilize as alternativas abaixo para responder àsquestões de 22 e 25.a. diplóide. c. heterozigótico.b. haplóide. d. homozigótico.
22. No cruzamento entre um indivíduo com umcaráter hereditário dominante eoutro com ocaráter recessivo, todos os descendentesapresentaram o caráter dominante. Pode-sedizer, portanto, que, muito provavelmente, o tipoparental dominante é ( ).
23. Em um cruzamento entre um indivíduo comum caráter hereditário dominante e outro como caráter recessivo, foram produzidos des-cendentes com o caráter dominante e descen-dentes com o caráter recessivo. Esse resultadopermite concluir que o tipo parental dominanteé ( ).
24. Um indivíduo que apresenta em suas célulasapenas um alelo de cada gene é ( ).
25. Um indivíduo que apresenta em suas célulasum par de alelos de cada gene é ( ).
Utilize as alternativas abaixo para responder àsquestões de 26 a 28.a. 1 : 2 : 1 c. 9 : 7b. 9 : 3 : 4 d. 12 : 3 : 1
26. Nos casos em que alelos diferentes de umgene se expressam na condição heterozigótica,a proporção fenotípica esperada em umcruzamento entre dois híbridos é ( ).
27. No caso de uma interação entre dois genescom segregação independente, em que o alelodominante de um dos genes mascara ou inibea expressão do outro gene, a proporçãofenotípica esperada no cruzamento de doisduplo-heterozigotos é ( ).
28. No caso de uma interação entre dois genescom segregação independente, em que o parde alelos recessivos de um dos genes mascaraou inibe a expressão do outro gene, a proporçãofenotípica esperada no cruzamento de doisduplo-heterozigotos é ( ).
PARTE C: QUESTÕES PARA PENSAR E DISCUTIR
29. Qual a explicação que se costuma dar parao fato de o trabalho de Mendel não ter sidoentendido durante 35 anos?
30. Por que Moore admite que o antagonismoentre o mendelista Bateson e os biometristasWeldon e Pearson era uma questão de conflitode paradigmas?
31. Por que algumas pessoas levantaram aquestão de que Mendel pode ter falsificado seusresultados?
32. Como surgem os diferentes alelos de umagene?
33. Quais serão as proporções fenotípicasesperadas em um cruzamento entre dois duplo-heterozigotos em que os dois genesapresentam segregação independente einteragem das seguintes maneiras:
a. um dos genes na condição homozigóticarecessiva, independentemente da condição dooutro gene, condiciona o fenótipo A ; emqualquer outra situação o fenótipo do indivíduoserá B.
b. os alelos recessivos não produzem nenhumtipo de pigmento e cada alelo dominantecondiciona a produção de uma quantidade Xde um mesmo pigmento.
c. basta ter um alelo dominante de qualquerdos dois genes para apresentar o fenótipo A;no caso dos dois genes estarem na condiçãohomozigótica recessiva, o fenótipo será B.
34. Em rabanetes, a forma da raiz pode serarredondada, ovalada ou alongada. Cruza-mentos entre plantas de raiz alongada e plantasde raiz arredondada produziram apenas indi-víduos com raiz ovalada. Em cruzamentosdesses indivíduos entre si foram obtidos 400
56

descendentes, dos quais 100 apresentaram raízesalongadas, 195 apresentaram raízes ovaladas e105 apresentaram raízes arredondadas.
a. Proponha uma hipótese para explicar essesresultados.
b. Com base na sua hipótese faça um diagramado cruzamento e compare os resultadosobservados com os esperados de acordo como diagrama.
c. Os resultados obtidos estão de acordo comas leis mendelianas? Explique
d. De acordo com a hipótese, se cruzássemosrabanetes de raiz ovalada com rabanetes deraiz arredondada, quais as proporções fenotí-pica e genotípica esperadas na descendência?
35. Em abóboras, a forma do fruto pode serdiscóide, esférica ou alongada. Uma variedadepura de frutos discóides foi cruzada com umavariedade pura de frutos alongados. A geraçãoF
1 foi inteiramente constituída por plantas de
frutos discóides. A autofecundação das plantasF1 produziu 80 descendentes, dos quais 30tinham frutos esféricos, 5 tinham frutos alon-gados e 45 tinham frutos discóides.
a. Proponha uma hipótese para explicar essesresultados.
b. Com base na sua hipótese faça um diagramado cruzamento e compare os resultadosobservados com os esperados de acordo como diagrama.
c . Os resultados obtidos estão de acordo comas leis mendelianas? Explique.
36. Em uma certa linhagem de animais foramidentificados dois genes com segregaçãoindependente afetando a massa. Os alelosdominantes condicionam, cada um, umacréscimo de 10 gramas à massa básica de 50gramas do duplo-recessivo. Determine a massamáxima e a mínima na descendência de umcruzamento Aabb X AaBB .
37. A pigmentação da plumagem em uma certalinhagem de galinhas é condicionada por doisgenes com segregação independente. O alelodominante de um dos genes (C) condiciona aprodução de pigmento enquanto o alelorecessivo (c) é inativo, condicionando cor bran-ca. O alelo dominante do outro gene (I) inibe aformação de pigmento, enquanto o alelorecessivo (i) não o faz. Determine a proporçãofenotípica em um cruzamento entre doisindivíduos duplo-heterozigóticos.
57