Embed Size (px)
Citation preview

REGULATED RIVERS: RESEARCH & MANAGEMENT, VOL. I I , 85- 104 (1995)
A PERSPECTIVE ON DRYLAND RIVER ECOSYSTEMS
K. F. WALKER, F. SHELDON AND J. T. PUCKRIDGE Department of Zoology, University of Adelaide, South Australia 5005, Australia
ABSTRACT The ecosystem concept should be reappraised as a basic model for rivers, with regard for flow as an organizing variable. This would facilitate comparisons between the large rivers of humid climates, where flow regimes are comparatively reg- ular, and those of arid and semi-arid areas, where river regimes are highly variable. Ecosystem processes might be mod- elled by combining the river continuum and flood pulse concepts, with refinements to accommodate a complex flood pulse (e.g. variations in stage amplitude, timing, duration, rates of rise and fall). Patch boundaries (ecotones) such as the riverine littoral zone warrant close study because they strongly influence the structure and dynamics of the ecosystem. The gen- eral model needs a quantitative basis, perhaps focused on the balance of processes involved in the physical transport and biological transformation of carbon. The ultimate test of such a model will be in its capacity to predict the effects of flow regulation. Further development, however, is limited by data. In both research and management monitoring pro- grammes need to be established to provide information and to develop a sustained, comprehensive approach to dryland rivers as ecosystems.
KEY WORDS dryland rivers; ecosystems; floodplains; flow regimes; river continuum concept; flood pulse concept; flow variability
INTRODUCTION
In its short history lotic ecology has gained much of its impetus from studies of streams. It has progressed from empiricism to theory and mixed with fluvial geomorphology, hydrology and geography, but still seeks a robust conceptual framework to accommodate large rivers on the scale of, say, the Mississippi. Although it may be possible to predict the gross effects of environmental changes such as dam construction, our under- standing of more subtle effects and the processes involved is still rudimentary. Progress is impeded by a lack of historical data and the difficulties of sampling. These impediments stem from problems of spatial and temporal scale that are common to virtually all environmental sciences (e.g. Kotliar and Wiens, 1990). There can be little scope for generalizations if ideas are scale-dependent.
Generalizations may come from a review of the effects of flow, the maestro that orchestrates pattern and process in rivers. Its effects pervade the physical and biological environments, determining the form of the channel and the nature of the sediments (Graf, 1987), affecting the physiology, distribution and abundance of organisms (Ward, 1992) and the dynamics of river and floodplain communities (Junk et al., 1989). Com- parisons of flow-related phenomena, perhaps in the context of hierarchy theory (O’Neill er al., 1986; Allen and Hoekstra, 1992), could elucidate the relationships between scale-dependent processes and, incidentally, reinforce the links between ecological concepts and those of hydrology and geomorphology.
Comparative analyses of flow regimes are usually related to perceived climatic or geographical regions (e.g. Gustard, 1992). Thus, the flows of rivers in arid and semi-arid regions are less regular than those of humid tropical and temperate regions, due partly to climatic phenomena such as the El Niiio-Southern Oscillation (ENSO) (cf. Richey et al., 1989; Molles et al., 1992). At a continental scale the most variable regimes are in the predominantly dry regions of Australia and southern Africa (Finlayson and McMahon, 1988; McMahon et al., 1992), but pockets of aridity occur in the Americas, China, India, the Mediterranean region and elsewhere. Indeed, arid and semi-arid regions occupy about one-third of the world’s land surface area (Thomas, 1989).
Junk et al. (1989) highlighted the role of the seasonal ‘flood pulse’ as a key variable in river-floodplain ecosystems. In dry regions floods are no less significant for riverine processes, but variability is also a
CCC OSS6-9375/95/050085-20 8 1995 by John Wiley & Sons, Ltd.
Received I S July 1994 Accepted 15 March 1995

86 K. F. WALKER, F. SHELDON AND J . T. PUCKRIDGE
dominant feature of the ‘physical habitat template’ (Poff and Ward, 1989; 1990). Junk et al. (1989: 122) sug- gested that the flood pulse concept is not easily applied where the pulse is variable, because ‘unpredictable pulses generally impede the adaptation of organisms’. However, life-history attributes such as opportunism and flexibility are arguably adaptations to unpredictability, and are typical of species from variable environ- ments (e.g. Baird et al., 1987). We believe that if its emphasis on the synchrony of hydrological and seasonal cycles is qualified, the flood pulse concept could be a useful model for lowland rivers in dry, as well as humid, regions.
A flow-based model would have an implicit capability to measure and predict the effects of regulation. It is remarkable how few current models have this capability, apart perhaps from the serial discontinuity concept (Ward and Stanford, 1983; 1995). The effects of flow regulation arise essentially because the processes of water supply and demand operate at different scales: daily and seasonal demands are imposed on a resource whose status is governed, in part, by non-seasonal factors. In dry regions regulation for agriculture may impose seasonal stability on rivers by amplifying the frequencies of some flows and tempering others, and there may be a partial reversal whereby summer-autumn flows are augmented and winter-spring Rows are depleted. In rivers used for hydropower, the regulated regime may include diel, as well as seasonal, changes (e.g. Petts, 1984). In either case the new space-time regime is likely to transform the physical, chemical and biological character of the river.
This paper outlines an ecological perspective on the large rivers of dryland (arid and semi-arid) regions. It is a speculative discussion rather than a review. We contend that dryland rivers are not well served by current concepts, mainly because they are governed by irregular flow patterns that have physical and biological sig- natures over a very wide spatio-temporal scale. We propose a reappraisal of the familiar ecosystem concept, based on analyses of hierarchical, flow-related phenomena, and integration of the river continuum and flood pulse models (Vannote et al., 1980; Junk et al., 1989) with an extended perspective to accommodate the regimes of dryland rivers and predict the impacts of regulation.
SPATIAL AND TEMPORAL SCALE
Environmental scientists seek to frame problems in space and time, ideally in a manner that is determined by the problem itself rather than by human perceptions. This is a challenge for all spheres of ecology (Kotliar and Wiens, 1990), and geomorphology (Schumm, 1988) and hydrology (Farquharson et al., 1992). It is especially problematic in the patchy, changeable environments of large dryland rivers.
For ecologists, some of the difficulties are traceable to ambiguities in the ecosystem concept (see later). These encourage perceptions of cause and effect relationships that make it difficult, if not impossible, to trace processes through an ecosystem. For example, schemes for assessing the impacts of regulation may consider biological taxa, assemblages and communities, qualitative and quantitative changes in water and sediment transport and other physical, chemical and biological factors (e.g. Ward and Stanford, 1979; Petts, 1984; 1987). These are valuable as classifications, but they may not help us to dissect complex systems into component subsystems. Hence they may discourage ‘operationalization’ of concepts (representing them by measurable patterns or processes: Peters, 1990). Table I shows a simple scheme wherein scales are related to orders of magnitude of space and time, and correlated with geomorphological, hydrological and biological phenomena.
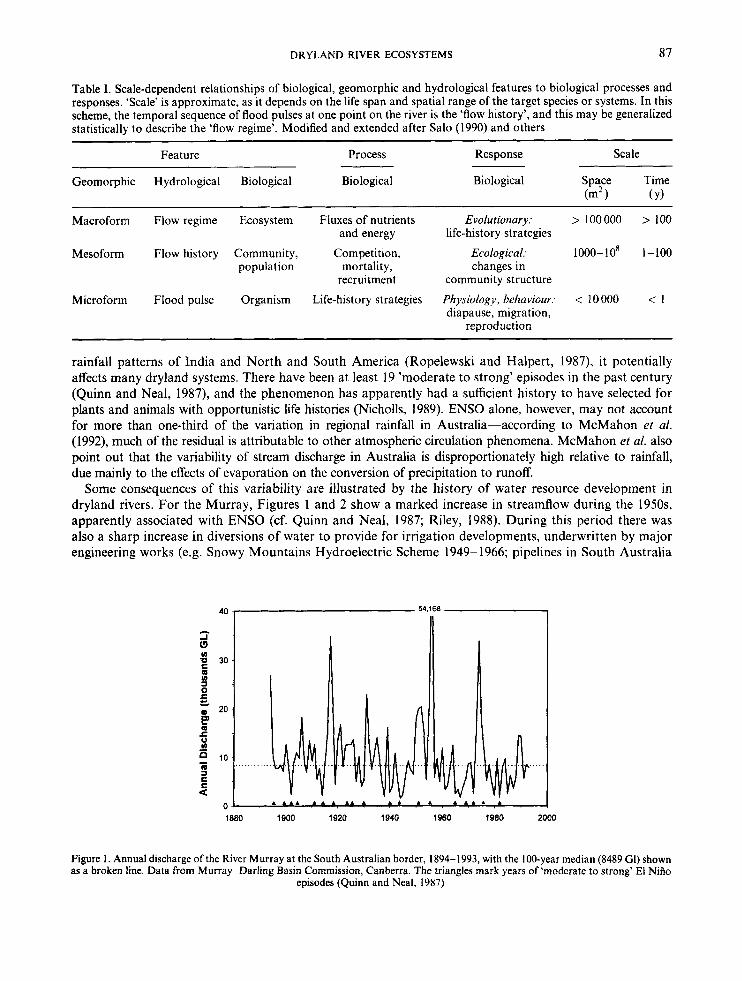
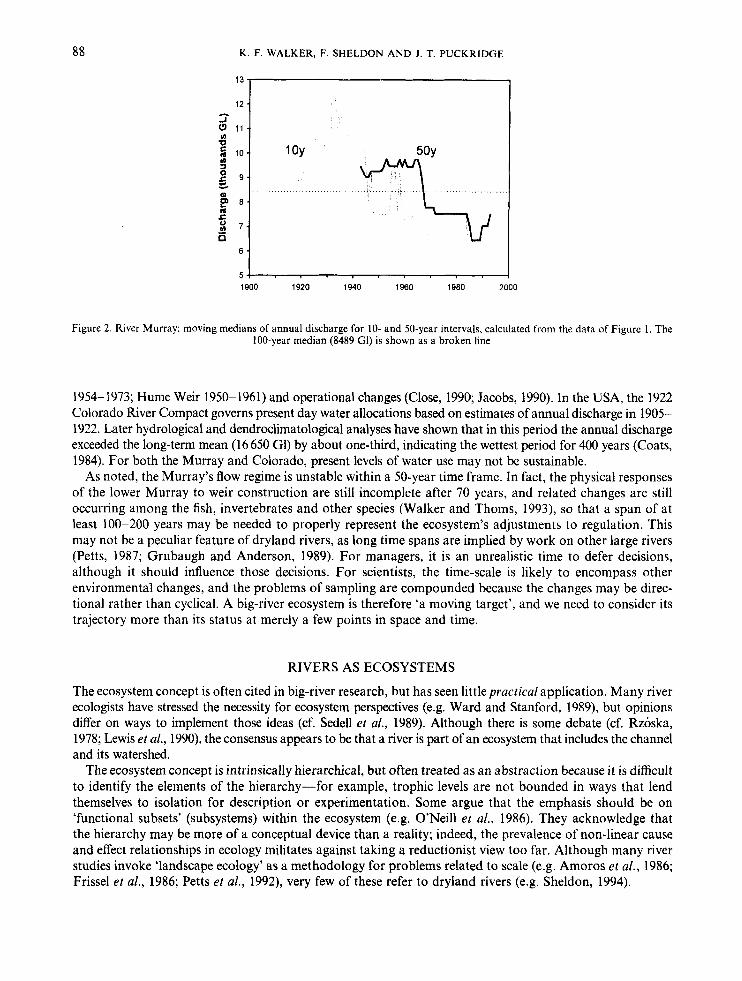
It is axiomatic that extended scales are needed to monitor variable systems. If, for example, river regimes in the Great Lakes region of the USA are adequately represented by a 10-year hydrograph (cf. Richards, 1990), dryland rivers may require a much longer time to demonstrate their repertoire of temporal behaviour. Figure 1 shows a 100-year time series (1894-1993) of annual discharge in the lower River Murray, a major river (catchment 1.073 x lo6 km2) in semi-arid south-eastern Australia. Figure 2 shows 10- and 50-year moving medians for the same data. The 100-year median (8489 G1) differs from the mean (10090 Gl), indicating skewness (IGI = 1091 = 106m3). The diagrams suggest that a 50-year record is not representative of the Murray’s regime, nor indicative of future trends.
There is a relationship between discharge in the Murray and Darling rivers and the sea surface tempera- ture anomalies that underlie ENSO (Simpson ef al., 1993; cf. Figure 1). As the ENSO signal also shows in the
DRYLAND RIVER ECOSYSTEMS 87
Table I . Scale-dependent relationships of biological, geomorphic and hydrological features to biological processes and responses. ’Scale’ is approximate, as it depends on the life span and spatial range of the target species or systems. In this scheme, the temporal sequence of flood pulses at one point on the river is the ‘flow history’, and this may be generalized statistically to describe the ‘flow regime’. Modified and extended after Salo (1990) and others
Feature Process Response Scale
Geomorphic Hydrological Biological Biological Biological Space Time (m2 ) (Y)
~~ ~
Macroform Flow regime Ecosystem Fluxes of nutrients Evolutionary: > 100000 > 100
Mesoform Flow history Community, Competition, Ecological: 1000-lo8 1-100
and energy life-history strategies
population mortality, changes in recruitment community structure
diapause, migration, reproduction
Microform Flood pulse Organism Life-history strategies Physiology, behaviour: < 10 000 < 1
rainfall patterns of India and North and South America (Ropelewski and Halpert, 1987), it potentially affects many dryland systems. There have been at least 19 ’moderate to strong’ episodes in the past century (Quinn and Neal, 1987), and the phenomenon has apparently had a sufficient history to have selected for plants and animals with opportunistic life histories (Nicholls, 1989). ENSO alone, however, may not account for more than one-third of the variation in regional rainfall in Australia-according to McMahon et al. (1992), much of the residual is attributable to other atmospheric circulation phenomena. McMahon et a[. also point out that the variability of stream discharge in Australia is disproportionately high relative to rainfall, due mainly to the effects of evaporation on the conversion of precipitation to runoff.
Some consequences of this variability are illustrated by the history of water resource development in dryland rivers. For the Murray, Figures 1 and 2 show a marked increase in streamflow during the 1950s, apparently associated with ENSO (cf. Quinn and Neal, 1987; Riley, 1988). During this period there was also a sharp increase in diversions of water to provide for irrigation developments, underwritten by major engineering works (e.g. Snowy Mountains Hydroelectric Scheme 1949- 1966; pipelines in South Australia
40 , 54,168
A A A A A A A A A A A A A A A A A A A
1900 1920 1940 1960 1980 2000
Figure 1 . Annual discharge of the River Murray at the South Australian border, 1894-1993, with the 100-year median (8489 GI) shown as a broken line. Data from Murray-Darling Basin Commission, Canberra. The triangles mark years of ‘moderate to strong’ El Nirio
episodes (Quinn and Neal, 1987)
88 K. F. WALKER, F. SHELDON A N D J. T. PUCKRIDGE
m -0
m 3 0
f 1 0 -
5 9 -
P 8 - a
s 7 - 6
- a!
S
5.1 1900 1920 1940 1960 1980 2000
Figure 2. River Murray: moving medians of annual discharge for 10- and 50-year intervals, calculated from the data of Figure I . The 100-year median (8489 GI) is shown as a broken line
1954-1973; Hume Weir 1950-1961) and operational changes (Close, 1990; Jacobs, 1990). In the USA, the 1922 Colorado River Compact governs present day water allocations based on estimates of annual discharge in 1905- 1922. Later hydrological and dendroclimatological analyses have shown that in this period the annual discharge exceeded the long-term mean (16 650 Gl) by about one-third, indicating the wettest period for 400 years (Coats, 1984). For both the Murray and Colorado, present levels of water use may not be sustainable.
As noted, the Murray’s flow regime is unstable within a 50-year time frame. In fact, the physical responses of the lower Murray to weir construction are still incomplete after 70 years, and related changes are still occurring among the fish, invertebrates and other species (Walker and Thoms, 1993), so that a span of at least 100-200 years may be needed to properly represent the ecosystem’s adjustments to regulation. This may not be a peculiar feature of dryland rivers, as long time spans are implied by work on other large rivers (Petts, 1987; Grubaugh and Anderson, 1989). For managers, it is an unrealistic time to defer decisions, although it should influence those decisions. For scientists, the time-scale is likely to encompass other environmental changes, and the problems of sampling are compounded because the changes may be direc- tional rather than cyclical. A big-river ecosystem is therefore ‘a moving target’, and we need to consider its trajectory more than its status at merely a few points in space and time.
RIVERS AS ECOSYSTEMS
The ecosystem concept is often cited in big-river research, but has seen little practical application. Many river ecologists have stressed the necessity for ecosystem perspectives (e.g. Ward and Stanford, 1989), but opinions differ on ways to implement those ideas (cf. Sedell et al., 1989). Although there is some debate (cf. Rzoska, 1978; Lewis et al., 1990), the consensus appears to be that a river is part of an ecosystem that includes the channel and its watershed.
The ecosystem concept is intrinsically hierarchical, but often treated as an abstraction because it is difficult to identify the elements of the hierarchy-for example, trophic levels are not bounded in ways that lend themselves to isolation for description or experimentation. Some argue that the emphasis should be on ‘functional subsets’ (subsystems) within the ecosystem (e.g. O’Neill et al., 1986). They acknowledge that the hierarchy may be more of a conceptual device than a reality; indeed, the prevalence of non-linear cause and effect relationships in ecology militates against taking a reductionist view too far. Although many river studies invoke ‘landscape ecology’ as a methodology for problems related to scale (e.g. Amoros et al., 1986; Frissel et al., 1986; Petts et al., 1992), very few of these refer to dryland rivers (e.g. Sheldon, 1994).
DRYLAND RIVER ECOSYSTEMS
1; 89
j
I Mesotime
1~ ............,...... ....
Macrotime f \ Microtime
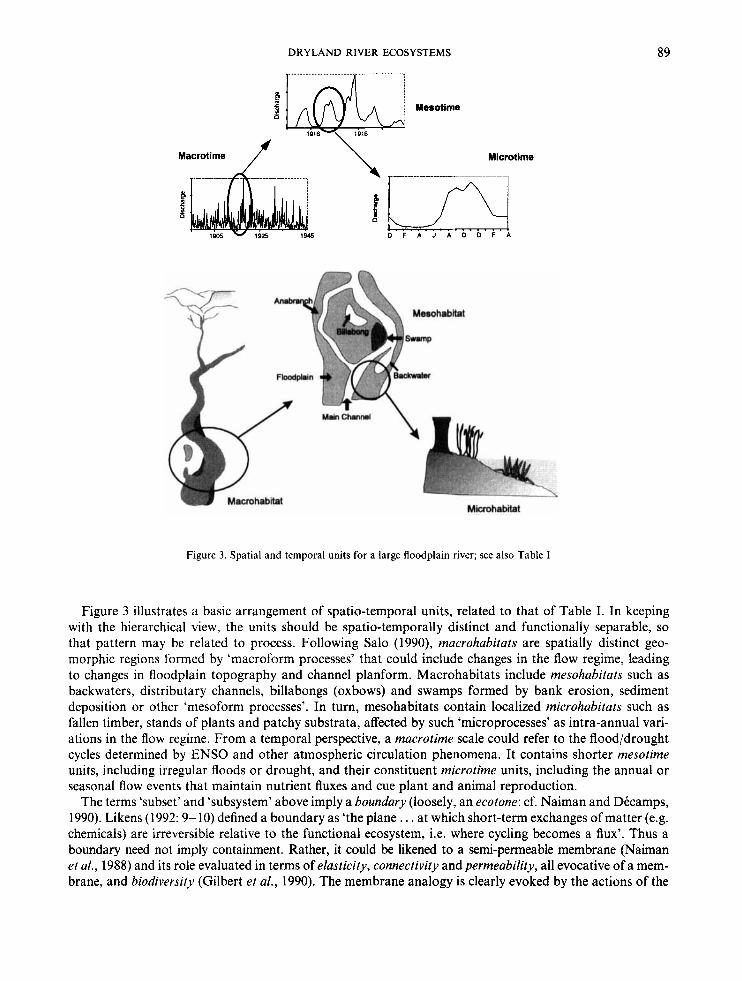
Figure 3. Spatial and temporal units for a large floodplain river; see also Table I
Figure 3 illustrates a basic arrangement of spatio-temporal units, related to that of Table I. In keeping with the hierarchical view, the units should be spatio-temporally distinct and functionally separable, so that pattern may be related to process. Following Salo (l990), macrohabitats are spatially distinct geo- morphic regions formed by ‘macroform processes’ that could include changes in the flow regime, leading to changes in floodplain topography and channel planform. Macrohabitats include mesohabitats such as backwaters, distributary channels, billabongs (oxbows) and swamps formed by bank erosion, sediment deposition or other ‘mesoform processes’. In turn, mesohabitats contain localized microhabitats such as fallen timber, stands of plants and patchy substrata, affected by such ‘microprocesses’ as intra-annual vari- ations in the flow regime. From a temporal perspective, a macrotime scale could refer to the flood/drought cycles determined by ENS0 and other atmospheric circulation phenomena. It contains shorter mesotime units, including irregular floods or drought, and their constituent microtime units, including the annual or seasonal flow events that maintain nutrient fluxes and cue plant and animal reproduction.
The terms ‘subset’ and ‘subsystem’ above imply a boundary (loosely, an ecotone: cf. Naiman and Decamps, 1990). Likens (1992: 9-10) defined a boundary as ‘the plane.. . at which short-term exchanges of matter (e.g. chemicals) are irreversible relative to the functional ecosystem, i.e. where cycling becomes a flux’. Thus a boundary need not imply containment. Rather, it could be likened to a semi-permeable membrane (Naiman et al., 1988) and its role evaluated in terms of elasticity, connectivity andpermeability, all evocative of a mem- brane, and biodiversity (Gilbert et al., 1990). The membrane analogy is clearly evoked by the actions of the

90 K. F. WALKER, F. SHELDON AND J . T. PUCKRIDGE
‘moving littoral’ during a flood (Junk et al., 1989). In general, the roles of boundaries in defining subsystems, and mediating exchanges between subsystems, suggest that they are ‘linkages between domains of scale’ (Gosz, 1993), acting to maintain structure in river-floodplain systems (Gregory et al., 1991). In some respects, therefore, patch boundaries may be a more appropriate focus for ecosystem-level studies than the patches themselves. An example of a boundary is the littoral zone of the River Murray (Walker et al., 1992), comparable with the ‘main-channel border habitat’ of the Mississippi (McConville et al., 1986).
Published work on dryland rivers (and big rivers in general) shows little evidence of the sustained, com- prehensive research and monitoring that is implied by ‘an ecosystem approach’. Following Likens (1992):
An empirical or natural history approach refers to the accumulation of descriptive data. Much of the published work on big rivers emphasizes simple variables and pattern recognition (e.g. Poff and Ward, 1989; Voelz and Ward, 1990; Davies et al., 1993), although these methods could be expected to founder where the observer’s perspective does not correspond to a functional spatio-temporal unit. A bulance or budgetary approach indicates elemental fluxes and cycling. There are few such data for dry- land rivers, compared with tropical and termperate rivers (e.g. Webb and Walling, 1992). Mass-budget data are easily referred to different scales of space and time and may provide a means to identify hier- archical effects and ecologically significant scales. An experimental approach, involving ecosystem-level manipulations of key variables, is scarcely repre- sented in published work on big rivers. One way to pursue this is as part of ‘adaptive management’ (Walters, 1990), wherein scientists and managers collaborate to develop their knowledge of exploited ecosystems. For example, much could be learned from monitoring the physical and biological parameters of controlled floods in rivers and floodplains. A comparative approach is also barely represented. Interbasin transfers, for example, present oppor- tunities to test ecological and other ideas on a grand scale (cf. O’Keeffe and de Moor, 1988; Davies et al., 1992). A modelling or computer simulation approach is ideally part of adaptive management. Many instream flow models are founded on work in mesic environments (e.g. Stalnaker, 1994), but their value for dryland rivers is open to question (e.g. Gan and McMahon, 1991).
FLOW REGIMES
Nature of :flow variability Many ‘dryland’ rivers, including the Murray, are allogenic: they rise in well-watered areas and flow for long
distances through arid or semi-arid country that contributes little runoff. Others, like Cooper Creek in central Australia (Kotwicki, 1986), rise in arid or semi-arid catchments and retain that hydrological character through- out their courses. Dryland rivers are therefore an ill-defined group: they may have features of both mesic and xeric systems, but are generally dominated by the latter. A quantitative index of xeric influences might be devel- oped, perhaps using ratios of mean annual precipitation and mean annual runoff (MAP/MAR) integrated over the drainage basin (e.g. Alexander, 1985). In the following, however, the term is used loosely.
Dryland rivers are hydrologically distinctive (Molles ef al., 1992):
0 Responses to variations in precipitation tend to be non-linear, compared with the near-linear responses of mesic catchments, and may amplify climatic changes.
0 Annual discharge may decrease with basin area, whereas that of mesic basins tends to increase (cf. Finlayson and McMahon, 1988).
0 Seasonal (and longer term) flow behaviour is strongly influenced by ENSO. 0 There may be a negative correlation between antecedent summer moisture and runoff (for example, dry
summers may precede high runoff in winter and spring).
The salient feature, however, is flow variability. Molles et al. (1992: 197) urged researchers and managers ‘to shift their focus from an equilibria1 to a non-equilibria1 view and concentrate their efforts on an exploration
DRYLAND RIVER ECOSYSTEMS 91
of the extent, causes and consequences of all major aspects of variation in stream and river flow'. The chal- lenge is unquestioned, but it is not clear how best to measure the variance of flows. The dimensions of the problem vary, depending on whether the observer is a hydrologist, geomorphologist or ecologist.
Hydrologists have developed sophisticated techniques for the analysis of flow frequency distributions, but these tend to reflect the problems of civil engineering (that is, storage and transport of water) rather than environmental science. The gap is narrowing (e.g. Falkenmark and Chapman, 1989; Gordon et al., 1992; Smith, 1992), but ecologists still make little use, for example, of analyses that specify recurrence times for flows of given magnitude, or probabilities of exceedance, although these should be useful in analyses of the ecological consequences of flow variability (cf. Maheshwari et af., 1993; 1995). Gaines and Denny (1993) showed that, given a sufficient time series, the probability distributions of extreme events (and hence recurrence times) can be described using maximum likelihood techniques. This could be useful in dryland river investigations because extreme events such as flood and drought are often of more immediate interest than measures of central tendency and dispersion.
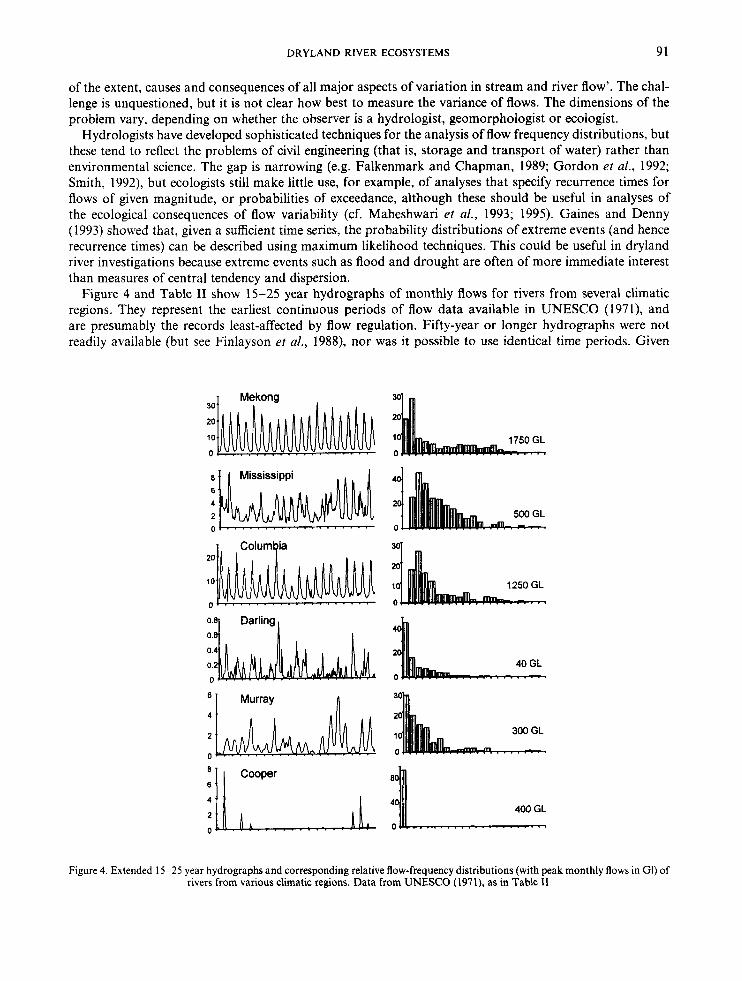
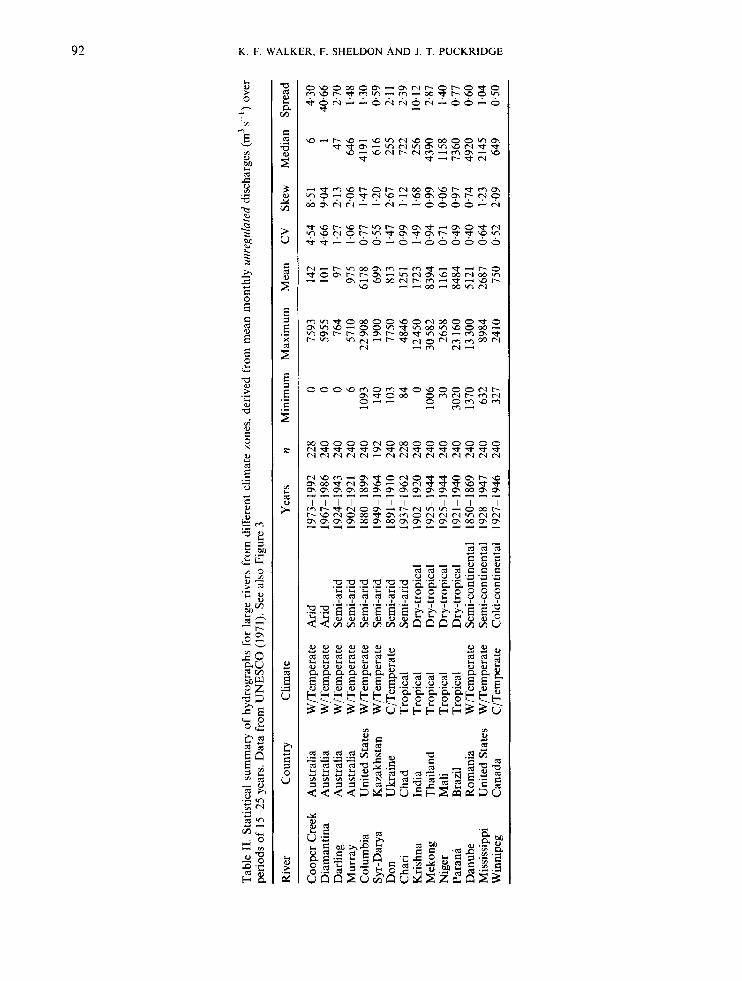
Figure 4 and Table I1 show 15-25 year hydrographs of monthly flows for rivers from several climatic regions. They represent the earliest continuous periods of flow data available in UNESCO (1971), and are presumably the records least-affected by flow regulation. Fifty-year or longer hydrographs were not readily available (but see Finlayson et al., 1988), nor was it possible to use identical time periods. Given
30
20
10
0
1 Columbia 20
10
0
0.9 Darling, 0.
0.
0.
0
d 1 Murray
Figure 4. Extended 15-25 year hydrographs and corresponding relative flow-frequency distributions (with peak monthly flows in GI) of rivers from various climatic regions. Data from UNESCO (1971), as in Table I1
Tabl
e 11
. Sta
tistic
al s
umm
ary
of h
ydro
grap
hs f
or la
rge
river
s fr
om d
iffer
ent c
limat
e zo
nes,
der
ived
fro
m m
ean
mon
thly
unr
egul
ated
dis
char
ges
(m3 s
-' )
over
pe
riods
of
15-2
5 ye
ars.
Dat
a fr
om U
NE
SCO
(197
1). S
ee a
lso
Figu
re 3
Riv
er
Cou
ntry
C
limat
e Y
ears
n
Min
imum
M
axim
um
Mea
n CV
Sk
ew
Med
ian
Spre
ad
Coo
per
Cre
ek
Dia
man
tina
Dar
ling
Mur
ray
Col
umbi
a Sy
r-D
arya
D
on
Cha
ri
Kri
shna
M
ekon
g N
iger
Pa
rana
D
anub
e M
issi
ssip
pi
Win
nipe
g
Aus
tral
ia
Aus
tral
ia
Aus
tral
ia
Aus
tral
ia
Uni
ted
Stat
es
Kaz
akhs
tan
Ukr
aine
C
had
Indi
a Th
aila
nd
Mal
i B
razi
l R
oman
ia
Uni
ted
Stat
es
Can
ada
W/T
empe
rate
W
/Tem
pera
te
W/T
empe
rate
W
/Tem
pera
te
W/T
empe
rate
W
/Tem
pera
te
C/T
empe
ra te
Tr
opic
al
Trop
ical
Tr
opic
al
Trop
ical
Tr
opic
al
W/T
empe
rate
W
/Tem
pera
te
C/T
empe
rate
Arid
A
rid
Sem
i-arid
Se
mi-a
rid
Sem
i-arid
Se
mi-a
rid
Sem
i-arid
Se
mi-a
rid
Dry
-tro
pica
l D
ry-t
ropi
cal
Dry
-tro
pica
l D
ry-t
ropi
cal
Sem
i-con
tinen
tal
Sem
i-con
tinen
tal
Col
d-co
ntin
enta
l
1973
- 19
92
1967
- 198
6 19
24-1
943
1902
-192
1
1880
-189
9 19
49- 1
964
I891
- I9
1 0
1937
- 19
62
1902
- 192
0 19
25-1
944
1925
-194
4 19
2 1 -
1940
18
50-1
869
1928
- 194
7 19
27- 1
946
228
0 24
0 0
240
0 24
0 6
240
1093
19
2 14
0 24
0 10
3 22
8 84
24
0 0
240
1006
24
0 30
24
0 30
20
240
1370
24
0 63
2 24
0 32
7
7593
59
55
764
5710
22
908
19
00
7750
48
46
1245
0 30
582
26
58
23 1
60
13 30
0 89
84
2410
142
4.54
8.
51
6 4.
30
101
4.66
9.
04
1 40
.66
97
1.27
2.
13
47
2.70
97
5 1.
06
2.06
64
6 1.
48
6178
0.
77
1.47
41
91
I .30
69
9 0.
55
1.20
61
6 0.
59
813
1.47
2.
67
255
2.11
12
51
0.99
1.
12
722
2.39
17
23
1.49
1.
68
256
10.1
2 83
94
0.94
0.
99
4390
2.
87
1161
0.
71
0.06
11
58
1.40
84
84
0.49
0.
97
7360
0.
77
5121
0.
40
0.74
49
20
0.60
26
87
0.64
1-
23
2145
1.
04
750
0.52
2.
09
649
0.50
v) F2 E; 0 z P
z U
L
9

DRYLAND RIVER ECOSYSTEMS 93
the apparent skewness, especially for the rivers in arid climates, it is curious that hydrologists usually favour parametric, rather than non-parametric, statistics to describe flow frequency distributions (cf. Agnew and Anderson, 1992). In general, flow distributions are not often normally distributed, although they may be log-normal (Gordon et al., 1992). Variability is often described by the mean and coefficient of variation (ratio of standard deviation to mean), although neither is reliable for non-normal distributions (cf. Table 11). Skewness probably exaggerates the coefficient of variation, but the effect is obscured because both the mean and standard deviation are affected. The non-parametric alternatives are rarely used-for example, the median and spread (ratio of interquartile range to median, e.g. Richards, 1989; 1990). Rodier (1985) defended the use of parametric data, pointing out that the mean may be realistic for long time series (say, more than 50 years). Nonetheless, if few data are available and the concordance of mean and median (and mode) is unknown, parametric measures should be used with caution. This is especially true for inves- tigations of dryland rivers, as mean and median flows are divergent roughly in relation to the degree of regio- nal aridity (Table 11). In general, the median and spread may be better descriptors in the presence of skewness, but are less amenable to modelling and statistical analysis.
Table I1 suggests that the variability of amplitude in the semi-arid Murray is higher than that of the Mekong River, but does not approach the extreme represented, for example, by Cooper Creek and the Diamantina River in arid central Australia (Kotwicki, 1986). Collectively, the regimes of dryland rivers tend not to show dominant, regular, annual or seasonal cycles. Irregularity, however, may simply reflect an observer’s perspective-non-seasonal trends appear less erratic at time-scales corresponding to ENS0 episodes (cf. Riley, 1988; Molles et al., 1992).
Colwell’s (1 974) indices of predictability, constancy and contingency are convenient measures of periodic phenomena. They were used to good effect in analyses of flow data by Poff and Ward (1989), but have some important limitations. For example, all are interdependent, rather than independent. Contingency and predict- ability (but not constancy) depend on the time class-interval underlying the frequency distribution. All three indices are vulnerable to skewness, which is an attribute of all flow distributions and pronounced in dryland rivers. For these reasons, Gan et al. (1991) suggested that the indices are more suited to discontinuous, qualitative data than to continuous, quantitative data.
In many rivers, modes in the flow frequency distribution may correspond with features of channel geometry (Graf, 1987). For example, the unregulated River Murray had in-channel benches that are now only partly preserved (Thoms and Walker, 1992) and its natural flow frequency distribution might be expected to show some corresponding modality. Spectral analysis or fractal modelling might be useful in this context.
Anatomy of a j o o d pulse Whether a flood hydrograph measures water level (stage) or flow (discharge), it is not an adequate basis to
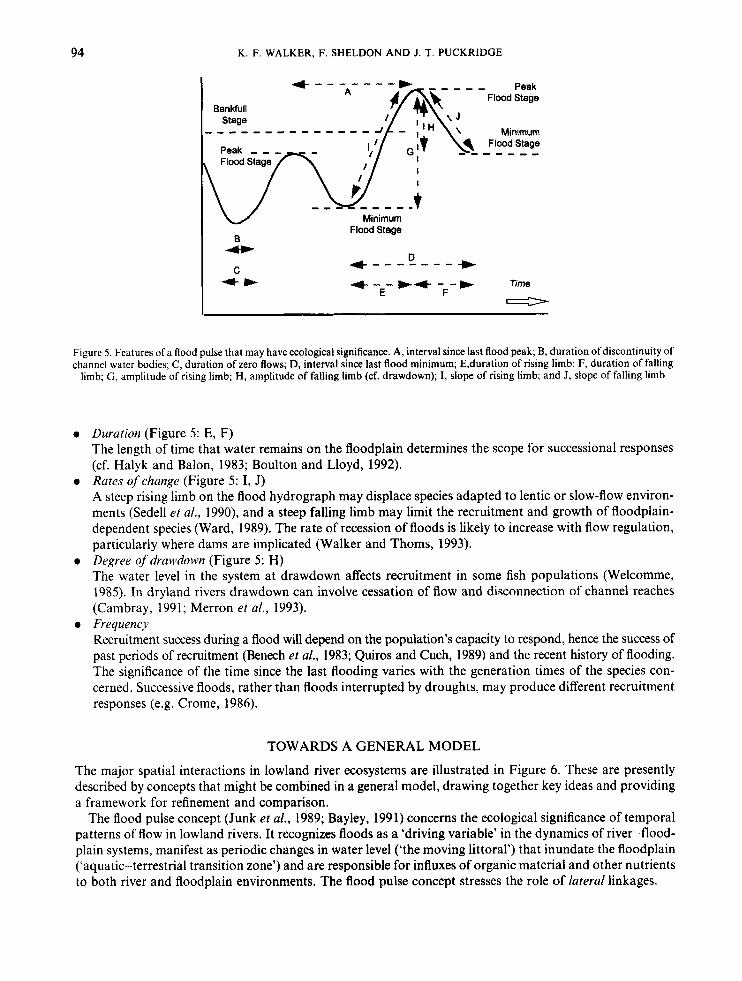
model ecological responses (Gustard, 1985). Figure 5 illustrates the potentially complex character of a flood pulse and implies that floods may differ in ways that could have diverse biological consequences. For example:
0 Stage amplitude (Figure 5 : G, H) Big floods are likely to promote extensive breeding and recruitment of river and floodplain species: for example, fishery yields tend to be correlated with flood magnitudes (Welcomme, 1985). Smaller (perhaps seasonal) floods may also be significant. Flood pulses within the channel release nutrients from sedi- ments and detritus, promoting plant growth and thereby providing food and habitats for animals. Small floods may also provide ‘bridging’ recruitment, maintaining the population’s capacity to endure drought and to respond to the less-frequent big floods. For example, although Murray cod (Percichthyidae: Muc- cullochellu peeli) breed seasonally and are not completely dependent on floods for spawning, recruitment is enhanced by flooding. Regulation of the Murray has curtailed spring flows and limited the incidence of all but the largest floods (Thomson, 1992), and so contributed to widespread declines of cod and other native fish in the last 30-40 years (Rowland, 1989).
0 Timing (Figure 5: A, D) Floods of sufficient magnitude and duration to sustain recruitment may not do so if their timing is decoupled from seasonal cycles of temperature and other spawning cues (Cambray, 1991).
94 K . F. WALKER. F. SHELDON AND J. T. PUCKRIDGE
Bankfull Stage
Minimum Flood Stage
B
- Peak Flood Stage
Minimum Flood Stage - - - - -
Figure 5. Features of a flood pulse that may have ecological significance. A, interval since last flood peak; B, duration of discontinuity of channel water bodies; C, duration of zero flows; D, interval since last flood minimum; E,duration of rising limb; F, duration of falling
limb; G , amplitude of rising limb; H, amplitude of falling limb (cf. drawdown); I, slope of rising limb; and J, slope of falling limb
0 Duration (Figure 5: E, F) The length of time that water remains on the floodplain determines the scope for successional responses (cf. Halyk and Balon, 1983; Boulton and Lloyd, 1992).
0 Rates ofchange (Figure 5: I, J) A steep rising limb on the flood hydrograph may displace species adapted to lentic or slow-flow environ- ments (Sedell et al., 1990), and a steep falling limb may limit the recruitment and growth of floodplain- dependent species (Ward, 1989). The rate of recession of floods is likely to increase with flow regulation, particularly where dams are implicated (Walker and Thoms, 1993).
0 Degree ofdrawdown (Figure 5 : H) The water level in the system at drawdown affects recruitment in some fish populations (Welcomme, 1985). In dryland rivers drawdown can involve cessation of flow and disconnection of channel reaches (Cambray, 1991; Merron et al., 1993).
Recruitment success during a flood will depend on the population’s capacity to respond, hence the success of past periods of recruitment (Benech et al., 1983; Quiros and Cuch, 1989) and the recent history of flooding. The significance of the time since the last flooding varies with the generation times of the species con- cerned. Successive floods, rather than floods interrupted by droughts, may produce different recruitment responses (e.g. Crome, 1986).
0 Frequency
TOWARDS A GENERAL MODEL
The major spatial interactions in lowland river ecosystems are illustrated in Figure 6. These are presently described by concepts that might be combined in a general model, drawing together key ideas and providing a framework for refinement and comparison.
The flood pulse concept (Junk et al., 1989; Bayley, 1991) concerns the ecological significance of temporal patterns of flow in lowland rivers. It recognizes floods as a ‘driving variable’ in the dynamics of river-flood- plain systems, manifest as periodic changes in water level (‘the moving littoral’) that inundate the floodplain (‘aquatic-terrestrial transition zone’) and are responsible for influxes of organic material and other nutrients to both river and floodplain environments. The flood pulse concept stresses the role of lateral linkages.

DRYLAND RIVER ECOSYSTEMS 95
headwaters
estuary groundwater
Figure 6. Spatial interactions in a lowland river ecosystem. After Ward and Stanford (1989)
The river continuum concept (Vannote et al., 1980; Minshall et al., 1985) suggests that lotic communities respond to a continuum of changes in the physical environment. Thus, terrestrial organic material is imported to the headwaters as litter (allochthonous carbon) and rendered into smaller fractions by hetero- trophic organisms as it is transported to the lower reaches where, as conditions become favourabie for instream production (autochthonous carbon), autotrophic organisms increase. The concept has gained most acceptance as a model for streams and upland rivers, notwithstanding arguments about its global applicability (e.g. Ward, 1992). A corollary, the serial discontinuity concept (Ward and Stanford, 1983; 1999, describes the consequences of an interruption to the continuum represented, for example, by an impoundment. In general, the river continuum concept emphasizes longitudinal linkages.
Although advocates of the river continuum concept have acknowledged the importance of lateral linkages (e.g. Sedell et al., 1989), the model is not easily applied to lowland systems where the carbon supply from upland reaches is less significant than that produced within the river-floodplain environment. The flood pulse concept addresses this problem, but does not accommodate upland rivers or rivers without a flood- plain (that is, with a confined channel). Thorp and Delong (1994) proposed the Riverine Productivity Model to highlight the significance of instream production and riparian inputs of organic material in periods not necessarily limited to flood pulses. Even where there are significant inputs from upstream or floodplain sources, Thorp and Delong argue that much of this material is refractory and less easily assimilated into riverine food webs than material from local instream and riparian sources.
The best general model for large rivers may be one that incorporates the longitudinal perspective of the river continuum concept in upland reaches and the lateral perspective of the flood pulse concept in lowland reaches. This should not obscure the importance of vertical (river-groundwater) linkages (e.g. Gilbert et al., 1990) or estuarine processes in many rivers (e.g. Limburg et al., 1989). The dominant linkages in any reach will be revealed in the direction of net movement of organic matter and nutrients. The hybrid model may not always apply-some rivers with a steep gradient or minor floodplain development may better reflect the river continuum model over their entire length (cf. Moisie River, Canada: Sedell et al., 1989), or they may derive a disproportionately high portion of their carbon supply from instream and riparian production (cf. upper Ohio River, USA: Thorp and Delong, 1994). Other rivers with extensive floodplains may be better described by the flood pulse model (e.g. Amazon River, Brazil: Junk et al., 1989).
Various refinements are needed, particularly to accommodate dryland rivers. One modification could address the point that a flood pulse need not necessarily be regular to have ecological and evolutionary sig- nificance. If, as suggested earlier, opportunistic and flexible life-history strategies are regarded as adaptations to unpredictable regimes, the original emphasis on a seasonal pulse could be relaxed (cf. Junk et al., 1989). A second, related point is that the flood pulse potentially has a complex character, demonstrated by variations in duration, rates of rise and fall, magnitude, timing and other ecologically significant ways. Finally, the hybrid model, like its constituents, needs a stronger quantitative basis to facilitate measurement, experiment

96 K. F. WALKER, F. SHELDON AND J . T. PUCKRIDGE
Littoral Zone Water Level
and analysis. Although the predictive capabilities of the river continuum model have been partly explored (e.g. Sedell et a!., 1989), the flood pulse model remains largely descriptive (but see Bayley, 1991). There is a dearth of predictive models concerning the ecological impact of flow regulation.
Moving Falling in Stable in Rising in Channel Channel Channel over Floodplain
RIVER-FLOODPLAIN INTERACTIONS
Measurements of river--floodplain interactions may provide for ecological comparisons between river ecosystems. For example, the relative abundance of lacustrine and riverine plankton species may be indicative. The plankton of rivers without significant floodplain wetlands tends to dominance by Rotifera, whereas that of floodplain riv- ers is dominated by Cladocera and Copepoda. Similarly, unregulated rivers tend to have a riverine (rotiferan) plankton, and regulated rivers have a lacustrine (microcrustacean) plankton. For example, the middle reaches of the regulated River Murray support a microcrustacean plankton, derived from impoundments and wetland areas, whereas the less regulated Darling River, which has few wetlands, has a rotiferan plankton (Shiel and Walker, 1985). Zooplankton from riparian wetlands also contribute to the plankton of the Blue Nile and Tigris rivers (Talling and Rzoska, 1967; Sabri et al., 1993). Comparable patterns are evident for phytoplankton in the Murray (Sullivan et al., 1988) and zoobenthos in the Colorado River (Ward et al., 1986). The balance may vary, of course, with different stages in the flood-drought cycle.
An alternative approach could be to measure the balance of physical transport and biological transformation in lowland river ecosystems (Figure 7). The transport-transformation dichotomy is implicit in both the river con- tinuum and flood pulse models, and recalls ‘supply-side ecology’ (Roughgarden et al., 1987) and the concept of resource spiralling (Newbold et al., 1982; Elwood et al., 1983). In a confined lowland reach not influenced by floodplain exchanges, imported material is transported downstream relatively quickly, with minimal processing by organisms. Where there are floodplain interchanges, nutrients carried by the river are deposited in forests and wetlands, subject to biological transformations, and returned to the river with the ebb of the flood (or subsequent floods). Thus, the floodplain is alternately a seasonal ‘sink’ and ‘source’ of material. Large lowland rivers are likely to have extensive two-way interactions with the riparian zone, including the floodplain, so that the parameters of the nutrient budget (and the boundaries for the ecosystem) should be sought at the watershed level. The interactions are mediated by flooding and are likely to show corresponding spatio-temporal variability
Allochthonous or Autochthonous
Inputs
Source or Sinkt 1
Transport or Transformation
I\ Floodplain Level
Figure 7. Processes of transport and transformation in relation to phases of the flood pulse for the lower reaches of a turbid, floodplain river with a shallow photic zone (see text for explanation)

DRYLAND RIVER ECOSYSTEMS 97
(cf. Junk et a/., 1989; Ward, 1989; Hillbricht-Ilkowska, 1993). Other indicator variables could be pollutants, or the composition of the plankton, benthos and littoral communities.
An obvious application of a general model would be to simulate the transport and transformation of carbon, the currency for most ideas about the dynamics of lotic ecosystems (cf. Hill et al., 1992). Figure 7 suggests that in the lower reaches of a turbid, floodplain river with a shallow photic zone, allochthonous inputs are likely to increase with the rise and fall of the flood pulse, and autochthonous inputs will increase when water levels (and the photic depth) are stable. Transport will occur to and from the channel and flood- plain during rising and falling floods, with transformation during periods of stable low or high flow. With the flood’s ebb the channel banks and floodplain floor will be a source of carbon, and the channel and floodplain (sediments and wetlands) will be a sink. Where flow regulation occurs, the frequency of exchanges between river and floodplain is decreased and, in the vicinity of impoundments, autochthonous carbon assumes more importance than allochthonous carbon. In effect, the longitudinal vector is strengthened at the expense of lateral vectors, and transport dominates over transformation. This rationale could be extended to describe the effects of regulation on a naturally confined river tract (cf. Thorp and Delong, 1994).
FLOW REGULATION
Nature of regulated regimes Flow variability is the feature of dryland rivers that most justifies regulation by dams and other structures.
Regulation is necessarily intensive, to satisfy demands for a reliable source of water in a climate that, in the short term, is inherently unreliable. It is difficult, however, to isolate the effects of regulation from those of a changeable climate, and more so in the absence of data for the period before regulation.
The mode of regulation determines the pattern of flow that is superimposed on the natural regime. Although reservoirs often have more than one purpose, the basic modes of operation are water supply (including irrigation), hydropower, navigation and flood control (e.g. Graf, 1987). The patterns of regulated flows, by definition, are regular and should submit more readily than natural regimes to analysis and general- ization. As suggested earlier, spectral analysis might help to describe the shifts in emphasis on daily, seasonal and annual components. The effects of regulation might then be compared, crudely, with a bandpass filter, in that daily or seasonal frequencies are filtered and modulated to approximate the regime(s) required by the regulating authority. A more intensive analysis is proposed by Richter et al. (in press). This involves 32 ‘indicators of hydrological alteration’, derived from flow data for the periods before and after regulation. The method recognizes the diverse consequences of hydrological changes and is a step towards the quanti- tative expression of ideas about the environmental impact of flow regulation. It also recognizes the inherent complexity of the flood pulse and should prove useful in investigations of dryland rivers.
The disparity between natural and regulated flow regimes is especially striking in dryland rivers. The Colorado River is a dramatic example, once prone to extreme flow variations and now perhaps the most closely controlled of any dryland system (e.g. Carlson and Muth, 1989). Following closure of Glen Canyon Dam in 1962 the mean annual discharge in the Colorado decreased from 2440 to 760m3 s-I, whereas the median annual discharge increased from 210 to 350m3 s-’ (Graf, 1987). The difference between mean and median indicates protracted low flows and that between unregulated and regulated flows demonstrates the immediate impact of regula- tion. Related changes in sediment transport are described by Graf (1987) and Lyons et a/. (1992).
In the River Murray also, patterns of flow are fundamentally altered (Thomson, 1992; Maheshwari et al., 1995). From 1930- 1991, annual water diversions from the Murray-Darling Basin (excluding Queensland) increased from 3000 to 11 000 G1. Whereas (simulated) natural annual flows in this period ranged from 2500-20000 G1 95% of the time, regulated flows were 0-15000 G1 95% of the time. Under regulation, the median natural flow (10968 G1) is exceeded only 8% of the time. Low flows (< 5000 G1) occurred 7 % of the time under natural conditions, but actually occurred 66% of the time, and high flows (> 25000 G1) occurred 5% of the time under natural conditions, but actually occurred 2% of the time. Changes in sediment transport are described by Thoms and Walker (1992). There are comparable data for dams on the Changjiang (Yangtse River) and Huanghe (Yellow River) in China (Liu and Zuo, 1987;

98 K . F. WALKER, F. SHELDON AND J . T. PUCKRIDGE
Liu, 1989; Liu and Yu, 1992), the Nile River in Egypt (e.g. Springuel and Murphy, 1991) and the Orange- Vaal system in South Africa (e.g. Davies et al., 1993).
Small weirs, barrages, causeways, levees and river training structures may be no less influential than dams, by virtue of their numbers and ubiquity. Their effects are compounded by offstream storages, selective manipulation of tributary flows and interbasin transfers, so that the cumulative effects may represent a far more extensive level of regulation than that suggested by dams alone (see, for example, Ferrar, 1979; Ebel et al., 1989; Storey et al., 1991; Davies et al., 1993; Walker and Thoms, 1993). Interbasin transfers (IBTs) have special significance for dry- land rivers and will become more important as demands increase and supplies dwindle. They are well advanced in South Africa, Australia, China and the USA (Liu and Zuo, 1987; Petitjean and Davies, 1988; Davies et al., 1992; 1994). In South Africa’s Eastern National Water Carrier Project the Kavango River will lose 47% of its mean annual flow during droughts (Davies et al., 1992). In the Orange River Project, flow in the upper Great Fish River has increased more than five-fold, and in the Lesotho Highlands Water Project the Ash River will receive an order of magnitude increase in its annual flow. Planning for these IBTs was not part of whole-catchment manage- ment; impacts were not considered and the ecological effects, including the breakdown of biogeographical boundaries, are scarcely known.
Changes in spatio-temporal domain The consequences of regulation are apparent throughout the space-time domain of a dryland river eco-
system, including the subsystem boundaries referred to earlier. Thus in the River Murray the impact of regulation is seen in patterns of water-level movements in the littoral zone (Walker et al., 1992; Maheshwari et al., 1993; Sheldon, 1994).
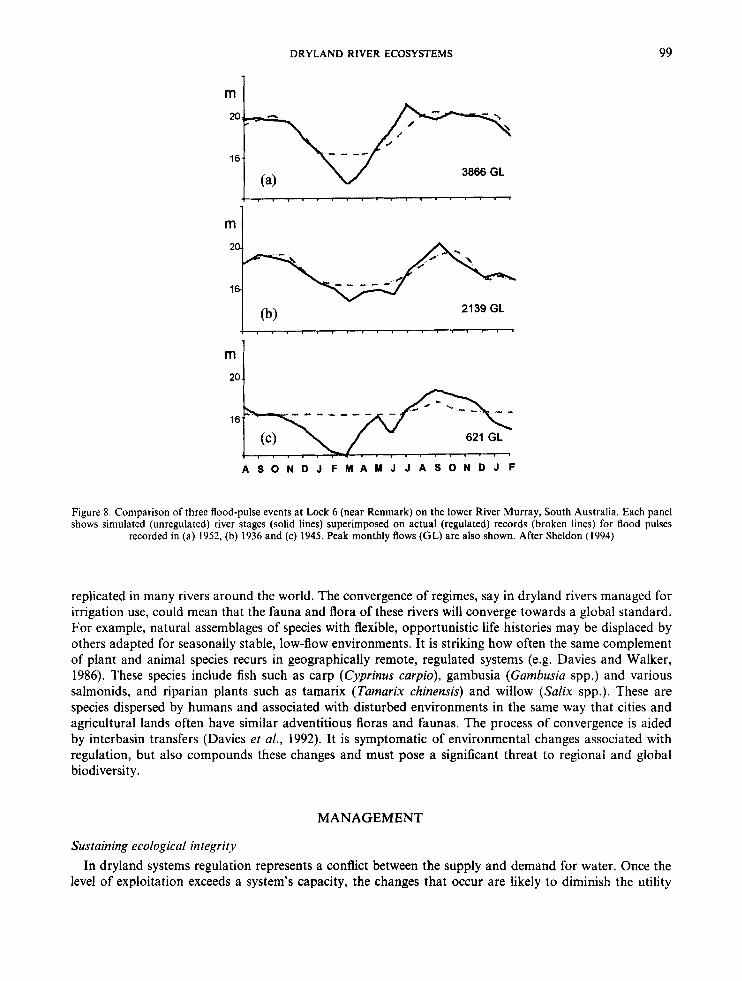
Figure 8 compares the behaviour of three dissimilar flood pulses in the lower Murray, which is controlled by upstream dams and diversions and a series of low-level weirs (Walker and Thoms, 1993). Each pulse is represented by actual stage measurements and data from the Murray-Darling Basin Commission’s monthly simulation model, which estimates natural flows by setting diversions to zero (e.g. Maheshwari et al., 1995). During a big, unregulated flood (Figure Sa), the river level moves continually over the banks and the floodplain. A moderate pulse (Figure 8b) shows the same change, but with more prolonged stable levels. For smaller, in-channel pulses (Figure Sc), regulation has eliminated any water-level response. In general, regulation has decreased the amplitudes of the rises and falls and prolonged the periods of stability.
In the lower Murray ecological processes concentrated in the rising and falling phases of the flood pulse are disrupted, whereas those active during periods of stable water are reinforced (cf. Figure 7). Regulation has modified the scale of littoral habitat patches by compressing lateral gradients and extending longitudinal gradients (hence alienating the river and floodplain), by reducing the frequency, amplitude and duration of floods, and by increasing the frequency of short-term (daily) water-level fluctuations. The littoral biota is generally dominated by emergent plants (Walker et al., 1995), decapod Crustacea, chironomids and other macroinvertebrates. Diversity is low-the atyid shrimp Paratya australiensis typically accounts for about two-thirds of the macroinvertebrate biomass (Sheldon, 1994).
Regulation has stabilized the river level at a seasonal scale by maintaining near-bankfull capacities, but also introduced daily level fluctuations in reaches immediately below the weirs (Walker et al., 1992). Thus regulation has decreased or increased variability, depending on the observer’s viewpoint. Sheldon ( 1 994) suggested that in the unregulated river sustained rises and falls in the water level and photic zone discouraged algal growth, and the littoral biofilms would have been dominated by heterotrophic bacteria and fungi. The overall effect of the temporal changes in water-level variations has been to promote the growth of fila- mentous algae such as Spirogyra in the near-surface water. The shift towards autotrophy appears to have reduced the nutritional quality of the biofilms for grazing invertebrates and thereby caused the local extinc- tion of 15 or more species of snails (Sheldon and Walker, 1993). A comparable heterotrophic-autotrophic shift is reported also for the Colorado River (Carlson and Muth, 1989).
Convergence of regulated river environments Flow variance is the hallmark of dryland rivers. Regulation redistributes the variance in space and time,
and although regulated flows are never entirely freed of external influences, they tend towards a model that is
DRYLAND RIVER ECOSYSTEMS 99
m 1
A S O N D J F M A M J J A S O N D J F
Figure 8. Comparison of three flood-pulse events at Lock 6 (near Renmark) on the lower River Murray, South Australia. Each panel shows simulated (unregulated) river stages (solid lines) superimposed on actual (regulated) records (broken lines) for flood pulses
recorded in (a) 1952, (b) 1936 and (c) 1945. Peak monthly flows (GL) are also shown. After Sheldon (1994)
replicated in many rivers around the world. The convergence of regimes, say in dryland rivers managed for irrigation use, could mean that the fauna and flora of these rivers will converge towards a global standard. For example, natural assemblages of species with flexible, opportunistic life histories may be displaced by others adapted for seasonally stable, low-flow environments. It is striking how often the same complement of plant and animal species recurs in geographically remote, regulated systems (e.g. Davies and Walker, 1986). These species include fish such as carp (Cyprinus carpio), gambusia (Gambusia spp.) and various salmonids, and riparian plants such as tamarix (Tumarix chinensis) and willow (Salix spp.). These are species dispersed by humans and associated with disturbed environments in the same way that cities and agricultural lands often have similar adventitious floras and faunas. The process of convergence is aided by interbasin transfers (Davies et al., 1992). It is symptomatic of environmental changes associated with regulation, but also compounds these changes and must pose a significant threat to regional and global biodiversity.
MANAGEMENT
Sustaining ecological integrity In dryland systems regulation represents a conflict between the supply and demand for water. Once the
level of exploitation exceeds a system's capacity, the changes that occur are likely to diminish the utility

100 K. F. WALKER, F. SHELDON AND J. T. PUCKRIDGE
of the resource and prejudice human use. Thus, insidious depletion of a resource at 2% annually will have doubled its effect in about 25 years, or roughly the generation time for humans (Paterson, 1984). The processes of degradation are not linear, however, and although dryland rivers have an extraordinary capacity to absorb change, a slow decline is likely to be followed by a sudden shift to a new condition (cf. Sparks et al., 1990). Human enlightenment in these matters is often stalled by factors from beyond the bounds of ecology.
Can the ecological integrity of dryland rivers be sustained if flow allocations are determined seasonally, rather than according to a long-term flow regime, and if the environment is considered a competitor for water rather than a guarantor? If there is a simple formula for sustainable exploitation, it may be to adjust the pattern of demand to that of supply. Users would be obliged to draw only water that is ‘surplus’ to the maintenance needs of the ecosystem, at times determined by again the natural regime. Such a proposition seems absurd to most irrigators, economists and politicians, but it is the basis of an ecological prescription for maintaining the integrity of large dryland rivers. If the prescription must be compromised, a positive step would be to review all aspects of the relationship between irrigated agriculture and its environment, includ- ing water requirements, pricing policies and irrigation technology (e.g. Agnew and Anderson, 1992).
Management, monitoring and research Monitoring, research and management are a triad: the goal of monitoring is to identify pattern, research is
to understand process, and management uses both kinds of information to balance supply and demand in the long term. As we are constrained by the paucity of historical data, future ecology will be so limited if we fail now to establish ongoing programmes to monitor environmental change. The responsibility for implemen- tation rests with managers and governments, but the tasks of design and review fall to researchers.
In the past decade there have been several surveys of the natural resources of the River Murray and its tributaries, leading to management plans for salinity, water quality, riparian vegetation, invertebrates, fish, nutrients, algae and wetlands (e.g. Walker, 1992; MDBC, 1993). Significantly, there is not a flow- management strategy-environmental flow allocations are effectively ad hoc, and the rules governing the distribution of water overwhelmingly favour irrigators. There is also a voluminous, management-oriented body of published work for the Colorado (e.g. Carlson and Muth, 1989) and Columbia systems in the USA (e.g. Ebel et al., 1989) and the Orange-Vaal and other systems in South Africa (e.g. Department of Water Affairs and Forestry, 1986; 1987-1990; Braune and Rogers, 1987; Ferrar, 1989; Chutter, 1992; Davies et al., 1993). However, there is no evidence that surveys and monitoring are driven by concepts, such as the ecosystem concept, that might provide for an understanding of processes and for inter-system com- parisons. Decisions about what, where and when to monitor ought to be guided by research and manage- ment, and subject to continual review. And if ecological perceptions of dryland rivers require a broad horizon, the same should be true for monitoring and management.
ACKNOWLEDGEMENTS
We are indebted to Dr M. L. Pace and Dr S. G. Findlay, Institute of Ecosystem Studies, New York Botanical Garden, for suggesting ideas about the transport and transformation of materials as a basis for a model of river ecosystem function, and to three anonymous reviewers for their comments and advice.
REFERENCES
Agnew, C. and Anderson, E. 1992. Water Resources in the Arid Realm. Routledge, London. 329 pp. Alexander, W. J. R. 1985. ‘Hydrology of low latitude Southern Hemisphere land masses’, Hydrohiologia, 125, 75-83. Allen, T. F. H. and Hoekstra, T. W. 1992. Toward a Unified Ecology. Columbia UP, New York. 384 pp. Amoros, C., Roux, A. L., Reygrobellet, J. L., Bravard, J.-P.. and Patou, G. 1986. ‘A method for applied ecological studies of fluvial
Baird, D. J., Linton, L. R., and Davies, W. 1987. ’Life-history flexibility as a strategy for survival in a variable environment’, Functional
Bayley, P. B. 1991. ‘The flood pulse advantage and the restoration of river-floodplain systems’, Regul. Riv., 6,75-86.
hydrosystems’, Regul. Riv., 1, 17-36.
E d . , 1,45-48.

DRYLAND RIVER ECOSYSTEMS 101
Benech, V., Durand, J.-R., and Quensiere, J. 1983. ‘Fish communities of Lake Chad and associated rivers and floodplains’ in Carmoye, J.-P., Durand, J.-R., and LtvCque, C. (Eds), Lake Chad: Ecology and Productivity of a Shallow Tropical Ecosystenl. Dr W. Junk, The Hague. pp 293-356.
Boulton, A. J. and Lloyd, L. N. 1992. ‘Flooding frequency and invertebrate emergence from dry floodplain sediments of the River Murray, Australia’, Regul. Riv., 7 , 137-1 5 1.
Braune, E. and Rogers, K. H. 1987. ‘The Vaal River catchment: problems and research needs’ S. Afr. Nat. Sci. Prog. Rep., 143,36 pp, Cambray, J. A. 1991. ‘The effects of fish spawning and management implications of impoundment water releases in an intermittent
Carlson, C. A. and Muth, R. T. 1989. ‘The Colorado River: lifeline of the American southwest’, Can. Spec. Publ. Fish. Aquat. Sci., 106,
Chutter, F. M. 1992. Prognosis of the Relevant Environmental Impact of the Lesotho Highlands Water Project on the Riverine Ecosysrems of the Orange River from the Lesotho Border to the P. K . Le Roux Dam. Department of Water Affairs and Forestry, Pretoria. I I pp.
Close, A. 1990. ‘The impact of man on the natural flow regime’ in Mackay, N. and Eastburn, D. (Eds), The Murray. Murray-Darling Basin Commission, Canberra, pp. 61-74.
Coats, R. 1984. ‘The Colorado River: river of controversy’, Environment, 26(2), 6-13, 36-39. Colwell, R. K. 1974. ‘Predictability, constancy, and contingency of periodic phenomena’, Ecology, 55, 1148-1 153. Crome, F. H. J. 1986. ‘Australian waterfowl do not necessarily breed on a rising water level’, Aust. Wildlife Res., 13,461-480. Davies, B. R. and Walker, K. F. (Eds) 1986. The Ecology of River Systems. Dr W. Junk, Dordrecht. 793 pp. Davies, B. R., Thorns, M. C. and Meador, M. 1992. ‘The ecological impact of inter-basin water transfers and their threats to river basin
integrity and conservation’, Aquar. Conserv. Mar. Freshwater Ecosyst., 2, 325-349. Davies, B. R., O’Keeffe, J. H. and Snaddon, C. D. 1993. ‘A synthesis of the ecological functioning, conservation and management of
South African river ecosystems’, Wut. Ref . Comm. (Pretoria) Rep. TT62/93, 232 pp. Davies, B. R., Thorns, M. C., Walker, K . F., O’Keeffe, J. H. and Gore, J. A. 1994. ‘Dryland rivers: their ecology, conservation and
management’ in Calow, P. and Petts, G. E. (Eds), The Rivers Handbook. Vol. 2. Blackwell Scientific, Oxford, pp. 484-512. Department of Water Affairs and Forestry 1986. Management of the Water Resources of the Republic of Sourh Africa. DWAF,
Pretoria. Department of Water Affairs and Forestry (1989-1990). Orange River Ecology: (I) Assessment of Environmental Water Requirement of
the Orange River between P . K . Le Roux Dam and the Mouth, ( 2 ) Assessment of Environmental Water Requirement,for the Orange River Mouth. Orange River Environmental Task Group, DWAF, Pretoria.
Ebel, W. J., Becker, C. D., Mullan, J. W. and Raymond, H. L. 1989. ‘The Columbia River-towards a holistic understanding’, Can. Spec. Publ. Fish. Aquat. Sci., 106, 205-219.
Elwood, J. W., Newbold, J. D., O’Neill, R. V. and Van Winkle, W. 1983. ‘Resource spiralling: an operational paradigm for analysing lotic ecosystems’ in Fontaine 111, T. D. and Bartell, S. M. (Eds), Dynamics of Lotic Ecosysrems. Ann Arbor Science, Ann Arbor,
Falkenmark, M. and Chapman, T. (Eds) 1989. Comparative Hydrology. An Ecological Approach to Land and Water Resources.
Farquharson, F. A. K., Meigh, J. R. and Sutcliffe, J. V. 1992. ‘Regional flood frequency analysis in arid and semi-arid areas’, J . Hydrol.,
Ferrar, A. A. (Ed.) 1979. ‘Ecological flow requirements for South African rivers’, S. Afr. Nat. Sci. Prog. Rep., 162, 118 pp. Finlayson, B. L. and McMahon, T. A. 1988. ‘Australia v the world: a comparative analysis of streamflow characteristics’ in Warner,
Finlayson, B. L., McMahon, T. A., Srikanthan, R., and Haines, A. 1986. ‘World hydrology: a new data base for comparative analyses’
Frissel, C. A,, Liss, W. J., Warren, C. E., and Hurley, M. D. 1986. ‘A hierarchical framework for stream habitat classification: viewing
Gaines, S. D. and Denny, M. W. 1993. ‘The largest, smallest, highest, lowest, longest and shortest: extremes in ecology’, Ecology, 74,
Gan, K. and McMahon, T. A. 1991. ‘Variability of results from the use of PHABSIM in estimating habitat area’, Regul. Riv., 5,233-239. Gan, K. C., McMahon, T. A. and Finlayson, B. L. 1991. ‘Analysis of periodicity in streamflow and rainfall data by Colwell’s indices’,
J . Hydrol., 123, 105-118. Gilbert, J. Dole-Olivier, M., Marmonier, P. and Vervier, P. 1990. ‘Surface water-groundwater ecotones’ in Naiman, R. J. and Dtcamps,
H. (Eds), The Ecology and Management of Aquatic- Terresirial Ecotones. UNESCO, Paris and Parthenon, Carnforth. pp. 199-226. Gordon, N. D., McMahon, T. A. and Finlayson, B. L. 1992. Stream Hydrology. An Introduction for Ecologists. Wiley, Chichester. 526 pp. Gosz, J. R. 1993. ‘Ecotone hierarchies’, Ecol. Appl., 3, 369-376. Graf, W. L. 1987. Fluvial Processes in Dryland Rivers. Springer-Verlag, Berlin. 346 pp. Gregory, S . V., Swanson, F. J., McKee, W. A. and Cummins, K. W. 1991. ’An ecosystem perspective of riparian zones’, BioScience, 41,
Grubaugh, J. W. and Anderson, R. V. 1989. ‘Long-term effects of navigation dams on a segment of the Upper Mississippi River’. Regui.
Gustard, A. 1985. ‘The characterisation of flow regimes for assessing the impact of water resource management on river ecology’ in
Gustard, A. 1992. ‘Analysis of river regimes’ in Calow, P. and Petts, G. E. (Eds), The Rivers Handbook. Vol. 1. Blackwell Scientific,
Halyk, L. C. and Balon, E. K. 1983. ‘Structure and ecological production of the fish taxocene of a small floodplain system’, Can. J . Zool.,
Hill, B. H., Gardner, T. J. and Ekisola, 0. F. 1992. ‘Predictability of streamflow and particulate organic matter concentration as indi-
South African river’, Regul. Riv., 6, 39-52.
220-239.
pp. 3-28.
UNESCO, Paris, 479 pp.
138,487-501.
R. F. (Ed.), Fluvial Geomorphology of Australia. Academic Press, New York. pp. 17-39.
in Hydrology and Water Res0urce.r Symposium. November 1986. Griffith University, Brisbane. pp. 288-296.
streams in a watershed context’, Environ. Manage., 10, 199-214.
1677-1692.
540-551.
Riv., 4, 97-104.
Lillehammer, A. and Saltveit, S. V. (Eds), Regulated Rivers. Universitetforslaget AS, Oslo. pp. 53-60.
Oxford. pp. 29-47.
61,2446-2464.
cators of stability in prairie streams’, Hydrobiologia, 242, 7- 18.

102 K. F. WALKER, F. SHELDON AND J. T. PUCKRIDGE
Hillbricht-llkowska, A. 1993. ‘Temperate freshwater ecotones: problem with seasonal instability’ in Gopal, B., Hillbricht-llkowska. A., and Wetzel, R. G . (Eds), Wetlands and Ecotones. Studies on Land- Water Interactions. National Institute of Ecology, New Delhi, pp. 17-34.
Jacobs, T. 1990. ‘River regulation’ in Mackay, N. and Eastburn, D. (Eds), The Murray. Murray-Darling Basin Commission, Canberra, pp. 39-60.
Junk, W. J. , Bayley, P. B., and Sparks, R. E. 1989. ‘The flood pulse concept in river-floodplain systems’, Can. Spec. Puhl. Fish. Aquat. Sci., 106, 110-127.
Kotliar, N. B. and Wiens, J. A. 1990. ‘Multiple scales and patch structure: a hierarchical framework for the study of heterogeneity’,
Kotwicki, V. 1986. Flood.? of Lake E y e . Engineering and Water Supply Department, Adelaide. Lewis, W. M. Jr, Weibezahn, F. H., Saunders I l l , J. F., and Hamilton, S. K. 1990. ‘The Orinoco River as an ecological system’, Infer-
Likens, G . E. 1992. The Ecosystem Approach: its Use And Abuse. Ecology Institute, Oldendorf/Luhe. 166 pp. Limburg, K . E., Levin, S. A,, and Brandt, R. E. 1989. ‘Perspectives on management of the Hudson River ecosystem’, Can. Spec. Puhl.
Liu, C. 1989. ‘Problems in management of the Yellow River’, Regul. Riv., 3, 361-370. Liu, C. and Zuo, D. 1987. ‘Environmental issues of the Three Gorges Project, China’, Regul. Riv., 1, 267-273. Liu, J . K. and Yu, Z. T. 1992. ‘Water quality changes and effects on fish populations in the Hanjiang River, China, following hydro-
electric dam construction’, Regul. Riv., 7, 359-368. Lyons, J. K., Pucherelli, M. J., and Clark, R. C. 1992. ‘Sediment transport and channel characteristics of a sand-bed portion of the
Green River below Flaming Gorge Dam, Utah, U S A , Regul. Riv., 7, 219-232. Maheshwari, B. L., Walker, K. F. and McMahon, T. A. 1993. The Impact of Flow Regulation on the Hydrology ofthe River Murray and
its Ecological Implications. Centre for Environmental Applied Hydrology, University of Melbourne, and River Murray Laboratory, Department of Zoology, University of Adelaide. 154 pp.
Maheshwari, 9. L., Walker, K. F., and McMahon, T. A. ‘Effects of regulation on the flow regime of the River Murray, Australia’, Re&.
McConville, D. R.. Anderson, D. D., Vose, R. N., and Wilcox, D. B. 1986. ‘The species composition, occurrence and temporal stability of submerged aquatic macrophyte patches along the main channel border of Pool 5A, Upper Missippi River’, Hydrobiologia, 136,77-84.
McMahon, T. A,, Finlayson, B. L., Haines, A. T., and Srikanthan, R. 1992. Global Runoff-Continental Comparisons of Annual Flows rind Peak Discharges. Catena Verlag, Cremlingen. 166 pp.
MDBC 1993. Algal Management Strategy. Background Papers. Murray-Darling Basin Commission, Canberra. 1 13 pp. Merron, G., Bruton, M., and la Hausse de Lalouviere, P. 1993. ‘Changes in fish communities of the Phongolo Floodplain, Zululand
(South Africa) before, during and after a severe drought’, Regul. Riv., 8, 335-344. Minshall, G. W., Cumniins, K. W., Petersen, R. C., Cushing, C. E., Bruns, D. A,, Sedell, J. R., and Vannote, R. L. 1985. ‘Developments
in stream ecosystem theory’, Can. J . Fish. Aquat. Sci., 42, 1045-1055. Molles, M. C. Jr., Dahm, C. N., and Crocker, M. T. 1992. ‘Climatic variability and streams and rivers in semi-arid regions’ in Robarts,
R. D. and Bothwell, M. L. (Eds), Aquatic Ecosystems in Semi-Arid Regions: Implications f o r Resource Management. NHRISymp. Ser. 7. Environment Canada, Saskatoon, pp. 197-202.
Naiman, R. J. and Dicamps, H. (Eds) 1990. The Ecology and Management of Aquatic-Terrestrial Ecotones. Unesco, Pans and Parthenon, Carnforth. 3 16 pp.
Naiman, R. J., Decamps, H., Pastor, J. and Johnston, C. A. 1988. ‘The potential importance of boundaries to fluvial ecosystems’, J . North Am. Benthol. Soc., 7, 289-306.
Newbold, J. D., Mulholland, P. J., Elwood, J. W., and O’Neill, R. V. 1982. ‘Organic carbon spiralling in stream ecosystems’, Oikos, 38, 266-272.
Nicholls, N. 1987. ‘How old is ENSO?’ Climate Cliange, 14, 1 1 1-115. O’Keeffe, J. H. and de Moor, F. C. 1988. ‘Changes in the physico-chemistry and benthic invertebrates of the Great Fish River, South
O’Neill, R. V., Angelis, D. L., Waide, J. B., and Allen, T. F. H. 1986. A Hierarchical Concept ofEcosystems. Princeton UP, Princeton. Patcrson, J. 1984. ‘Exploitation of natural systems and the existence of a sustainable yield solution’, Water, 11, 5. Peters, R. H. 1990. A Critiquefor Ecology. Cambridge UP, Cambridge. 366 pp. Petitjean, M. 0. G. and Davies, B. R. 1988. ‘Ecological impacts of inter-basin water transfers: some case studies, research requirements
and assessment procedures in southern Africa’, S. Afr. J . Sci., 84, 819-828. Petts, G. E. 1984. Impounded Rivers. Wiley, Chichester. 326 pp. Petts, G. E. 1987. ’Time scales for ecological change in regulated rivers’ in Craig, J . F. and Kemper, J. B. (Eds), Regulated Streams:
Advances in Ecology. Plenum Press, New York. pp. 257-266. Petts, G. E., Large, A. R. G., Greenwood, M. T. and Bickerton, M. A. 1992. ‘Floodplain assessment for restoration and conservation:
linking hydrogeomorphology and ecology’ in Carling, P. and Petts, G. E. (Eds), Lowland Floodplain Rivers: Geomarphological Perspectives. Wiley, Chichester. pp. 21 7-234.
Poff, L. N. and Ward, J . V. 1989. ‘Implications of stream flow variability and predicatability for lotic community structure: a regional analysis of stream flow patterns’, Can. J. Fish. Aqua?. Sci., 46, 1805-1818.
Poff, N. L. and Ward, J. V. 1990. ‘Physical habitat template of lotic systems: recovery in the context of historical patterns of spatio- temporal heterogeneity’, Environ. Manage., 14, 629-645.
Quinn, W. H. and Neal, V. T. 1987. ‘El Nitio occurrences over the past four and a half centuries’, J . Geophys. Res., 14,449-461. Quiros, R. and Cuch, S. 1989. ’The fisheries and limnology of the Lower Plata Basin’, Can. Spec. Publ. Fish. Aquat. Sci., 106,429-443. Richards, R. P. 1989. ‘Measures of flow variability for Great Lakes tributaries’ Environ. Monitoring Assessment, 12, 361-377. Richards, R. P. 1990. ‘Measures of flow variability and a new flow-based classification of Great Lakes tributaries’, J . Great Lakes Res.,
0iko.T. 59, 253-260.
ciencia, 15, 346-357.
Fish. Aquat. Sci., 106, 265-291.
R ~ K , 10, 15-38.
Africa, following an interbasin transfer of water’, Regul. Riv., 1, 39-56.
16.53-70.

DRYLAND RIVER ECOSYSTEMS 103
Richey, J. E., Nobre, C., and Deser, C. 1989. ‘Amazon River discharge and climate variability: 1903 to 1985’, Science. 12, 101-103. Richter, B. D., Baumgartner, J. V.. Powell, J. and Braun, D. P. ‘A method for assessing hydrologic alteration within ecosystems’,
Riley, S. J. 1988. ’Secular change in the annual flows of streams in the NSW section of the Murray-Darling Basin’ in Warner, R. F.
Rodier, J . A. 1985. ‘Aspects of arid zone hydrology’ in Rodda, J. C. (Ed.), Facets of Hydrology. Vol. 2. Wiley, Chichester. pp. 205-247. Ropelewski, C. F. and Halpert, M. S. 1987. ‘Global and regional scale precipitation patterns associated with the El Niiio/Southern
Roughgarden, J . , Gaines, S. D., and Pacala, S . W. 1987. ‘Supply-side ecology: the role of physical transport processes’ in Gee, J. H. R.
Rowland, S. J. 1989. ‘Aspects of the history and fishery of the Murray cod, Maccullochella peeli (Mitchell) (Percichthyidae)’ Proc.
Rzoska, J . 1978. On the Nature of Rivers, with Case Stories of Nile, Zake and Amazon. Dr W. Junk, The Hague. 67 pp, Sabri, A. W., Ali, Z. H., Shawkat, S. F., Thejar, L. A., Kassim, T. I . , and Rasheed, K. A. 1993. ‘Zooplankton population in the River
Tigris: effects of Samarra Impoundment’, Regul. Riv., 8, 237-250. Salo, J. 1990. ‘External processes influencing origin and maintenance of inland water-land ecotones’ in Naiman, R. J. and Decamps, H.
(Eds), The Ecology and Management of Aquatic-Terrestrial Ecotones. UNESCO Man and the Biosphere Series. Vol. 4. Parthenon, Paris. pp. 37-64.
Schumm, S. A. 1988. ‘Variability of the fluvial system in space and time’ in Standing Committee on Problems of the Environment (SCOPE), Scales and Global Change. Wiley, Chichester. pp. 225-250.
Sedell, J . R., Richey, J . E., and Swanson, F. J. 1989. ‘The River Continuum Concept: a basis for the expected ecosystem behaviour of very large rivers?’ Can. Spec. Publ. Fish. Aquat. Sci., 106, 49-55.
Sedell, J. R., Reeves, G. H., Hauer, F. R., Stanford, J. A., and Hawkins, C. P. 1990. ‘Role of refugia in recovery from disturbances: modern fragmented and disconnected river systems’, Environ. Manage., 14, 71 1-724.
Sheldon, F. 1994. ‘Littoral ecology of a regulated dryland river (River Murray, South Australia) with reference to the Gastropoda’, Unpublished PhD Thesis, University of Adelaide, South Australia.
Sheldon, F. and Walker, K. F. 1993. ‘Pipelines as a refuge for freshwater snails’, Regul. Riv., 8, 295-300. Shiel, R. J. and Walker, K. F. 1985. ‘Zooplankton of regulated and unregulated streams: the Murray-Darling river system, Australia’ in
Simpson, H. J., Cane, M. A., Herczeg, A. L., Zebiak, S. E., and Simpson, J. H. 1993. ‘Annual river discharges in southeastern Australia
Smith, I . R. 1992. Hydrocliniate-the Influence of Water Movement of Freshwater Ecology. Elsevier, Amsterdam. 285 pp. Sparks, R. E., Bayley, P. B., Kohler, S. L., and Osborne, L. L. 1990. ‘Disturbance and recovery of large floodplain rivers’, Environ.
Springuel, I. and Murphy, K. J . 1991. ‘Euhydrophyte communities of the River Nile and its impoundments in Egyptian Nubia’, Hydro-
Stalnaker, C. B. 1994. ‘Evolution of instream flow habitat modelling’ in Calow, P. and Petts, G. E. (Eds), The Rivers Handbook. Vol. 2.
Storey, A. W., Edward, D. H., and Gazey, P. 1991. ‘Recovery of aquatic macroinvertebrate assemblages downstream of the Canning
Sullivan, C., Saunders, J . , and Welsh, D. 1988. ‘Phytoplankton of the River Murray, 1980-1985’, Murray-Darling Basin Commission,
Conser. Biol., in press.
(Ed.), Fluvial Geomorphology of Australia. Academic Press, Sydney. pp. 245-266.
Oscillation’, Monthly Weather Rev., 115, 1606-1626.
and Giller, P. S . (Eds), Organization of Communities Past and Present. Blackwell Scientific, Oxford. pp. 491-518.
Linnean Soc. New South Wales, 11, 201-213.
Lillehammer, A. and Saltveit, S . (Eds), Regulated Rivers. Universitetsforlaget, Oslo. pp. 263-270.
related to El Niiio Southern Oscillation forecasts of sea surface temperatures’, War. Resour. Res., 29, 3671-3680.
Manage., 14,699-709.
biologia, 218, 35-47.
Blackwell Scientific, Oxford. pp. 276-286.
Dam, Western Australia’, Regul. Riv., 6, 21 3-224. . -
Canberra. Wat. Qual. Rep., 2, 61 pp. Talling, J. F. and Rzoska, J . 1967. ‘The development of plankton in relation to hydrological regime in the Blue Nile’, J . Ecol., 55,657-
662.
London and Halsted Press, New York. pp. 1-10,
A. and Petts, G. E. (Eds), Lowland Floodplain Rivers: Geomorphological Perspectives. Wiley, Chichester. pp. 234-249.
sion, Canberra, Tech. Rep. 9215.1, 1 1 pp.
large river ecosystems’, Oikos, 70, 305-308.
Thomas, D. S. G. 1989. ‘The nature of arid environments’, in Thomas, D. S . G. (Ed.), Arid Zone Geomorphology. Belhaven Press,
Thorns, M. C. and Walker, K. F. 1992. ‘Channel changes related to low-level weirs on the River Murray, South Australia’ in Carling, P.
Thomson, C. 1992. ‘The impact of river regulation on the natural flows of the Murray-Darling Basin’, Murray-Darling Basin Commis-
Thorp, J. H. and Delong, M. D. 1994. ‘The riverine productivity model: an heuristic view of carbon sources and organic processing in
UNESCO 1971. Discharge of Selected Rivers of the World. Vol. 2. UNESCO, Paris. Vannote, R. L., Minshall, G . W., Cummins, K. W., Sedell, J . R., and Cushing, C. E. 1980. ‘The river continuum concept’, Can. J . Fish.
Voelz, N. J. and Ward. J . V. 1990. ‘Macroinvertebrate responses along a complex regulated stream environment’, Regul. Riv., 5, 365-374. Walker, K. F. 1992. ‘A semi-arid lowland river: the River Murray, Australia’ in Calow, P. A. and Petts, G. E. (Eds), The Rivers Hand-
Walker, K. F. and Thorns, M. C. 1993. ‘A case history of the environmental effects of flow regulation on a semi-arid lowland river: the
Walker, K. F., Thorns, M. C. and Sheldon, F. 1992. ‘Effects of weirs on the littoral environment of the River Murray, South Australia’ in
Walker, K. F., Boulton, A. J., Thorns, M. C., and Sheldon, F. 1995. ‘Effects of water-level changes induced by weirs on the distribution
Walters, C. 1990. ‘Large scale management experiments and learning by doing’, Ecology, 71,2060-2068. Ward, J. V. 1989. ‘Riverine-wetland interactions’ in Sharitz, R. R. and Gibbons, J. W. (Eds), Freshwater Wetlands and Wildli&e.
Aquat. Sci., 37, 130- 137.
book. Vol. I . Blackwell Scientific, Oxford. pp. 472-492.
River Murray, South Australia’, Regul. Riv., 7, 103-1 19.
Boon, P. J. , Petts, G. E., and Calow, P. (Eds), River Conservation and Management. Wiley, Chichester. pp. 271-292.
of littoral plants along the River Murray, South Australia’, Aust. J . Mar. Freshwater Res. 45, 1421-1438.
USDOE Office of Scientific and Technical Information, Oak Ridge, Tennessee. pp. 385-400.

104 K. F. WALKER. F. SHELDON A N D J. T. PUCKRIDGE
Ward. J. V. 1992. Aquatic Inseci Ecology, 1. Biology and Habitat. Wiley, Chichester. 438 pp. Ward, J. V. and Stanford, J. A. 1979. ‘Limnological considerations in reservoir operation: optimization strategies for protection of biota
Ward, J. V. and Stanford, J. A. 1983. ‘The serial discontinuity concept of lotic ecosystems’ in Fontaine 111, T. D. and Bartell, S . M. (Eds).
Ward, J. V. and Stanford, J. A. 1989. ‘Riverine ecosystems: the influence of man on catchment dynamics and fish ecology’, CUB. Spec.
Ward, J. V. and Stanford, J. A. 1995. ‘The serial discontinuity concept: extending the model to floodplain rivers’, Re&. Riv., 10, 159-
Ward, J. V., Zimmerman, H. J. and Cline, L. D. 1986. ‘Lotic zoobenthos of the Colorado system’ in Davies, B. R. and Walker, K. F.
Webb, B. W. and Walling, D. E. 1992. ‘Water quality, 11. Chemical characteristics’ in Calow, P. and Petts, G. E. (Eds), The Rivers Hand-
Welcomme, R . L. 1985. ‘River fisheries’, F A 0 Fisheries Tech. Pap., 262,330 pp.
in the receiving stream’ in Proceedings of the Mitigation Symposium. US Dept Agriculture, Fort Collins. pp. 347-356.
Dynaniics of Lotic Ecosystems. Ann Arbor Science, Ann Arbor. pp. 29-42.
Publ. Fish. Aquat. Sci., 106, 56-64.
168.
(Eds), The Ecology of River Systems. Dr W. Junk, Dordrecht, pp. 403-424.
book. Vol. 2. Blackwell Scientific, Oxford, pp. 73-100.












![1 MONSOON DEPENDENT ECOSYSTEMS: IMPLICATIONS OF THE ... · with good representation of dryland ecohydrological triggers [Wang et al., 2012]. Arid and semiarid regions are characterized](https://img.pdfslide.net/doc/110x75/5f89872adddb812f8122e619/1-monsoon-dependent-ecosystems-implications-of-the-with-good-representation.jpg)
















