Embed Size (px)
Citation preview

This article was downloaded by: [Ams/Girona*barri Lib]On: 08 October 2014, At: 08:45Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Journal of Sustainable ForestryPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/wjsf20
A Review of the Theories to ExplainArctic and Alpine Treelines Around theWorldAndrew D. Richardson a & Andrew J. Friedland ba Complex Systems Research Center , Institute for the Study ofEarth, Oceans and Space, University of New Hampshire , Durham,New Hampshire, USAb Environmental Studies Program , Dartmouth College , Hanover,New Hampshire, USAPublished online: 26 Feb 2009.
To cite this article: Andrew D. Richardson & Andrew J. Friedland (2009) A Review of the Theoriesto Explain Arctic and Alpine Treelines Around the World, Journal of Sustainable Forestry, 28:1-2,218-242, DOI: 10.1080/10549810802626456
To link to this article: http://dx.doi.org/10.1080/10549810802626456
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

218
Journal of Sustainable Forestry, 28:218–242, 2009Copyright © Taylor & Francis Group, LLC ISSN: 1054-9811 print/1540-756X onlineDOI: 10.1080/10549810802626456
WJSF1054-98111540-756XJournal of Sustainable Forestry, Vol. 28, No. 1, Dec 2008: pp. 0–0Journal of Sustainable Forestry
A Review of the Theories to Explain Arctic and Alpine Treelines Around the World
Theories to Explain Arctic and Alpine TreelinesA. D. Richardson and A. J. Friedland
ANDREW D. RICHARDSON1 and ANDREW J. FRIEDLAND2
1Complex Systems Research Center, Institute for the Study of Earth, Oceans and Space, University of New Hampshire, Durham, New Hampshire, USA
2Environmental Studies Program, Dartmouth College, Hanover, New Hampshire, USA
Forest growth is restricted at high latitudes and high elevations,and the limits of tree growth in these environments are dramati-cally marked by the treeline transition from vertical, erect treestems to prostrate, stunted shrub forms. However, after 4 centuriesof research, there is still debate over the precise mechanism thatcauses Arctic and alpine treelines. We review the various theoriesfor treeline, including excessive light, low partial pressure of CO2,snow depth, wind exposure, reproductive failure, frost drought,and temperature. Some of these theories are very old and are nolonger held in high esteem; while they may help to explain treelinephysiognomy or local variation in treeline position, they generallyfail as global explanations. Temperature-based theories appear tobe the most reasonable, since cold temperature is really the onlytrait that is universally characteristic of treelines around theworld. Temperature may limit a variety of physiological processes,such as carbon fixation, cuticular ripening, or new tissue develop-ment, and theories invoking these mechanisms are discussed.The vertical growth habit of trees is unfavorable to growth in thishostile environment: Low-profile vegetation enjoys a far morefavorable microenvironment for growth. Microsite enviroment and
This is a contribution to the Festschrift in honor of Professor Graeme Berlyn and hislong career at the Yale School of Forestry and Environmental Studies. Our choice of topic isa tribute to Graeme’s lifelong interest in Arctic and alpine ecosystems, and his participationin numerous studies of the mountain environment (see Figure 1). We thank Jim Kellner forhelpful comments on the manuscript, and Bill Smith and Matt Germino for drawing to ourattention some of the recent literature on regeneration at treeline.
Address correspondence to Andrew D. Richardson, University of New Hampshire,Complex Systems Research Center, Institute for the Study of Earth, Oceans and Space,Durham, NH 03824. E-mail: [email protected]
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 219
ecological facilitation have been shown to be essential for successfulregeneration, which is a prerequisite for upward advancement oftreeline. Recent evidence supports a theory based on “sink limitation,”i.e., that new tissue development is restricted not by carbon avail-ability but by cold treeline temperatures which limit cell division,and that this situation is exacerbated by arborescent growth(aboveground meristems coupled to cold ambient air temperatures)and self-shading (which keeps soil temperatures cold and restrictsbelowground activity).
KEYWORDS alpine, Arctic, biogeography, climate change, elevation,forest limit, Krummholz, latitude, montane forests, sink limitation,sub-alpine, treeline
INTRODUCTION
Treelines are phenomena that occur at the transition from forestland tosome other type of vegetation, such as between forest and grassland, or atthe margins of a swamp or bog: Treelines are therefore a “tension zone”(Griggs, 1934). Causes of treeline can be varied, but include abiotic factors(e.g., temperature, salinity, drought, waterlogging, or soil nutrients), and dis-turbance, both natural (e.g., fire), and anthropogenic (e.g., timber harvesting,development, or agriculture).
One of nature’s most dramatic examples of treeline is the bordermarking the upper limit of forest growth, i.e., the alpine (high elevation)or Arctic (high latitude) treelines that will be the focus of this reviewpaper (Figure 1). These treelines may be sudden or indistinct (Figure 2),they may be wavy or straight, and they may advance or recede over time.Here our goal is to review the theories, both historic and current, thatexplain alpine and Arctic treelines. We will limit our discussion to thosetreelines that are climate-driven, and ignore anomalies such as the grassbalds of the southern Appalachians, for which there are other causes(Mark, 1958).
Air cools as it rises and consequently the air temperature decreases at arate intermediate between 0.50°C/100 m (the saturated adiabatic lapse rate)and 0.98°C/100 m (the dry adiabatic lapse rate). On average, the lapse rateis usually about 0.60°C/100 m increase in elevation (Barry, 1992; Körner,1999; Richardson, Lee, & Friedland, 2004). Temperature also generallydecreases with increasing latitude. This occurs because at higher latitudesthe sun’s radiation is less direct. Richardson, Berlyn, and Duigan (2003)calculated that mean annual temperature in Alaska decreases by 1.15°C perdegree increase in latitude, and based on these data, they concluded that a1°C increase in latitude was roughly comparable to a 192-m increase in
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

220 A. D. Richardson and A. J. Friedland
elevation. Humbolt’s Law, which dates to 1817, proposes that in terms oftemperature, latitude compensates for altitude, and treeline thus occurs at alower elevation as one moves from equator to Arctic (Salisbury & Ross,1992).
There are, however, differences between alpine and Arctic (Billings,1973), and there are similar differences between sub-alpine and sub-Arctic.Generally the sub-alpine zone has greater light intensity and more UV, withless annual day length variation but more diurnal temperature variation,
FIGURE 1 Professor Graeme Berlyn and the second author, near treeline on Mt. Moosilaukein the White Mountains of New Hampshire, summer 2001.
FIGURE 2 View of the summit ridge on Mt. Mansfield, in the Green Mountains of Vermont,illustrating a somewhat indistinct treeline transition from forest to patchy Krummholz. Notealso the rocky, shallow soils which are typical of high elevation.
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 221
than the sub-Arctic. However, both sub-alpine and sub-Arctic frequentlyhave short growing seasons, low temperatures and high winds (Berlyn,1993). Low temperature is one of the few factors globally common to both.
Many authors differentiate between Arctic and alpine treelines; Arctictreelines being associated with increasing latitude, and alpine treelinesbeing associated with increasing altitude. However, in some of the world’shigh-latitude mountain ranges, it is difficult to distinguish between the two.An example of this is the Richardson Mountains (Yukon-NWT border, 67°–70° Nin Canada), where the forest limit is determined both by high latitude andaltitude. In most cases, the distinction between Arctic and alpine is moreeasily made.
The scientific study of treeline has a long history, as according to Troll(1973), it began in the Swiss Alps during the 16th and 17th centuries.Despite four centuries of research, there is not yet a consensus theory toexplain Arctic and alpine treelines universally around the world. However,some explanations are more valid than others at a universal level, whereassome theories seem to provide good explanations for local variation intreeline positions. The purpose of this article is to assess the arguments bothfor and against the theories that have been popular in the past and thosethat are popular now. We will begin by offering some definitions of treelineand describing the physiognomy of different treelines around the world,and then proceed to an assessment of the different theories. In recent years,there has been a renewed interest in treeline research, spurred on byconcerns about human-induced climate change and potentially dramaticchanges in high-mountain vegetation patterns. A better understanding ofthe controls over treeline location will enable better predictions to bemade about the effects of climate change on these climate-sensitive indicatorecosystems.
DEFINITION OF TREELINE
Different definitions of tree, forest, timber, and line complicate things some-what. For example, although most authors agree that trees are erect, woodystems, Troll (1973) insists on a minimum height of 5 m, whereas Körner(1998) specifies 3 m, and Wardle (1974) settles for 2 m. These authors thereforedistinguish between “trees” and the prostate, stunted “Krummholz” (fromthe German for “elfin wood”) found at the extreme limits of growth.
Körner defined timberline to be the limit of closed forest, but notedthat this transition is rarely abrupt: With increasing elevation or latitude, treesize generally decreases gradually while at the same time the canopy opensup. Wardle (1965) defined timberline to be the upper limit of tall, erect timber-sized trees. Treeline marks the highest patches of forest across slopes ofsimilar aspect according to Körner (1998), but in Daubenmire’s (1954)
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

222 A. D. Richardson and A. J. Friedland
nomenclature, the forestline is the upper edge of continuous forest andtreeline is the elevation of the highest “tree,” whether Krummholz or not.
We acknowledge that these terms all have somewhat different connotations.For the purpose of this article, however, we wish to keep a broad view oftreeline, for the delineation of any line or limit is inherently subjective. It isimportant to focus on the phenomenon of interest (rather than semantics);namely the vegetation change, be it gradual or sudden, from tall trees toprostrate shrubs that occurs at high elevations and high latitudes. We followWardle’s (1974) definitions, but will use the term treeline rather than Wardle’stree limit. Thus: treeline is the extreme limit of trees and tall shrubs (morethan 2 m in height). Trees growing closely together make a forest and thereforedefine the forest limit. In some places, forest limit and treeline are the same(e.g., most Nothofagus in New Zealand). In other systems, there may be azone of parkland between forest limit and treeline, or the trees between theforest limit and treeline might be stunted and deformed Krummholz: In thiscase, treeline is the point where Krummholz with flagged stems changes toprostrate Krummholz. We consider vegetation immediately below thetreeline to be sub-alpine (or, as the case may be, sub-Arctic) whereas vege-tation above the treeline is alpine (or Arctic).
GENERAL PATTERNS OF TREELINE
Treeline elevations range from near sea level, as in northern Canada andAlaska, up to 4,700 m in Tibet and 5,000 m in the Andes of Bolivia andChile (Troll, 1973). Treeline elevation generally increases as one movesfrom the poles to the equator, but there is a wide variation in treeline eleva-tion at a given latitude. Aspect, prevailing winds, soil quality, and the heightof surrounding mountains are thought to cause local variation in treeline(Daubenmire, 1954). Human use, grazing, avalanches, and fire are all distur-bances which can also have indirect effects on treeline elevations (Wardle,1974). At night, cold air drainage onto valley floors may create what isknown as an “inverted treeline.” In the humid tropics, treeline is generallyhigher in valleys or gullies rather than on ridges, but the reverse is oftentrue in the temperate zone (Troll, 1973).
There is more variability in treeline elevations in the northern hemi-sphere than the southern hemisphere. This may be due to the prevalence ofinterior or continental mountains in the northern hemisphere, which aremore or less absent in the southern hemisphere (Körner, 1999). This differenceis significant because of the large mountain mass effect, “Massenerhebungseffekt,”whereby the adiabatic lapse rate is generally lower on large mountains (orin the middle of a mountain range) than small mountains (or at the edges ofa mountain range). Treeline thus occurs at higher elevations in the middleof a range compared to the edges, due to less moisture, more sun, and less
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 223
exposure—all of which contribute to warmer temperatures (Daubenmire,1943; Barry, 1992; Körner, 1999). Coastal mountain ranges similarly havelower treelines than inland ranges. For example, treeline in the CascadeRange (Pacific Northwest) or White Mountains (New England) occurs atabout 1500 m, compared to 3000 m in the Rocky Mountains (Colorado).
Nevertheless, within a mountain range there is a fairly consistent latitude-elevation relationship for treeline between 35° and 70° N latitude, as demon-strated by Daubenmire (1954). The y-axis (elevation) intercept changessomewhat among mountain ranges, but the slope is relatively constant: Theelevation of treeline decreases by approximately 110 m for each 1° increasein latitude. Below 35° N, the relationship flattens. The relationship is a bitdifferent in the southern hemisphere, but the slope appears similar. Using alarger data set, Körner (1999) reports that from 45° N to 70° N, the ratio is45 m for every 1° latitude. From 30° N to 50° N, the slope is almost threetimes as great: 130 m for every 1° latitude. In the northern Appalachians,the alpine treeline decreases from 1480 m at 44° N to 550 m at 55° N. Thiscorresponds to an 83-m decrease in treeline elevation for every 1° increasein latitude (Cogbill & White, 1991).
Due to the fact that treeline can vary significantly both among moun-tains in the same range and even at different points on the same mountain,it seems unlikely that any single treeline theory can be applied to the entireworld’s Arctic and alpine treelines, regardless of latitude or continent. Rather,we should perhaps look for a general theory that predicts the approximatetreeline location, and then consider what factors might be responsible forlocal or smaller scale variation (Körner & Paulsen, 2004).
Assessment of treeline theories is complex because treeline positioncan change over time with climatic change. It is important to ask whetherpresent treelines indicate current climatic conditions, or past climatic conditions?Trees are long-lived and fairly resilient to environmental change, whereasgerminating seeds and seedlings may die during even relatively short spellsof unfavorable weather (Slatyer & Noble, 1992; note that the age structure attreeline tends to be “top heavy,” see Stevens & Fox, 1991), so the answer ismore likely past climatic conditions (Körner, 1998). For example, whentreeline is advancing, changes in treeline will lag behind changes in climatebecause seedlings must not only become established (out competing existingvegetation), but then survive long enough, and grow large enough, to beidentified as treeline individuals. Furthermore, when treeline recedes, existingtrees have to die, which may also take time. Treeline position and climateare therefore not likely to be in perfect equilibrium. The timescale of thismismatch is not entirely clear, but it is more likely in the years-to-decadesrange, rather than centuries. These observations are significant because onlyif a treeline is stable and truly representative of current conditions can validexperimental tests of treeline hypotheses actually be carried out (Beaman,1962; Stevens & Fox, 1991). It is also important to note that the variability in
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

224 A. D. Richardson and A. J. Friedland
climate from year to year may mask climate-treeline relationships unless thedata record is long enough.
As pointed out by Griggs (1934), the treeline is a “mobile migrationfront” rather than a “static climatic boundary.” In Kodiak, AK, treeline is verysudden (Griggs, 1934), with no crippled or deformed trees, no Krummholz,and no dead trees at the margin between Sitka spruce forest and aldergrassland. Cone production is abundant, and there is significant reproductionbeyond treeline. Griggs deduced that the forest border at Kodiak is actuallyadvancing, and that it has advanced at a rate of about 15 m/yr for at leastthe last few centuries. Fonda and Bliss (1969) describe alpine meadow inva-sion by Abies lasiocarpa in the Olympic Mountains, WA. Forest reversion totundra has been noted in the central Yukon Territory (Cwynar & Spear,1991). Climatic warming has been associated with the advance of thetreeline in the Coast Range, BC (Brink, 1959). In parts of Scotland, thetreeline is at 600 m, but stumps dated to 5,000–9,000 years ago have beenfound in the peat at 800 m. Similarly, there is evidence from Scandinaviathat the treeline has dropped by 200 m over the last 8,600 years (Grace,1989). Changes in treeline position appear strongly correlated with changesin temperature. Using historical data, a 1°C change in annual temperaturewas shown to translate to a 100-m change in treeline elevation (Grace,1989). Various temperature-based theories about treeline will be discussedin greater detail below.
TRENDS IN ABIOTIC FACTORS WITH REGARD TO ELEVATION AND LATITUDE
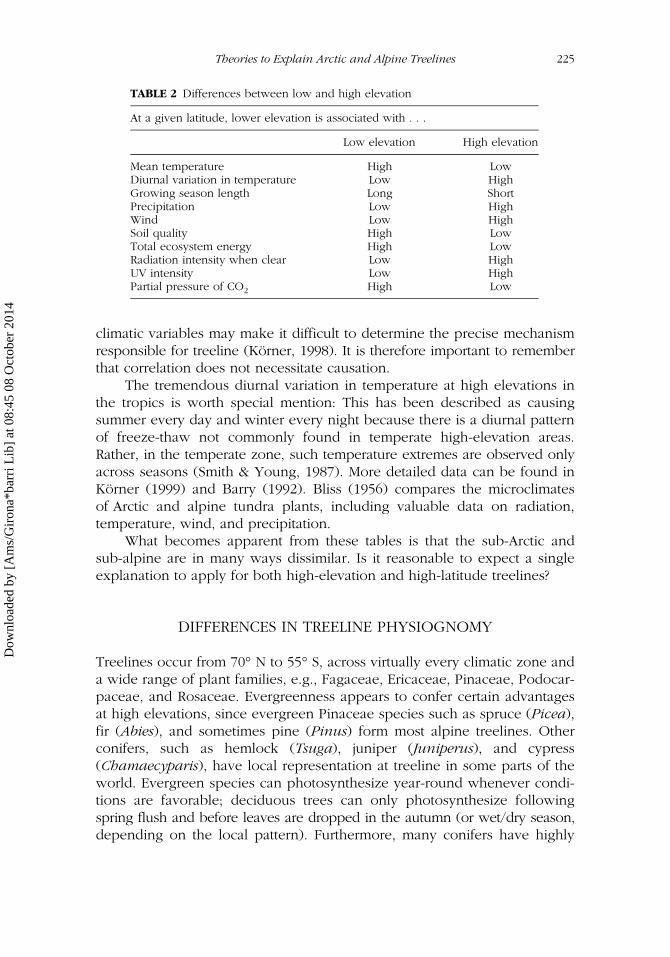
It may be dangerous to make broad generalizations, but in an attempt tocharacterize high-elevation and high-latitude treelines, we offer Tables 1and 2, which highlight some of the important differences between low andhigh elevation and low and high latitude. Note that autocorrelation among
TABLE 1 Differences between low and high latitude
At a given altitude, lower latitude is associated with . . .
Low latitude High latitude
Mean temperature High LowTotal ecosystem energy High LowSeasonal temperature amplitude Low HighDiurnal temperature amplitude High LowGrowing season length Long ShortSolar angle & peak radiation High LowCloudiness High Low
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4
Theories to Explain Arctic and Alpine Treelines 225
climatic variables may make it difficult to determine the precise mechanismresponsible for treeline (Körner, 1998). It is therefore important to rememberthat correlation does not necessitate causation.
The tremendous diurnal variation in temperature at high elevations inthe tropics is worth special mention: This has been described as causingsummer every day and winter every night because there is a diurnal patternof freeze-thaw not commonly found in temperate high-elevation areas.Rather, in the temperate zone, such temperature extremes are observed onlyacross seasons (Smith & Young, 1987). More detailed data can be found inKörner (1999) and Barry (1992). Bliss (1956) compares the microclimatesof Arctic and alpine tundra plants, including valuable data on radiation,temperature, wind, and precipitation.
What becomes apparent from these tables is that the sub-Arctic andsub-alpine are in many ways dissimilar. Is it reasonable to expect a singleexplanation to apply for both high-elevation and high-latitude treelines?
DIFFERENCES IN TREELINE PHYSIOGNOMY
Treelines occur from 70° N to 55° S, across virtually every climatic zone anda wide range of plant families, e.g., Fagaceae, Ericaceae, Pinaceae, Podocar-paceae, and Rosaceae. Evergreenness appears to confer certain advantagesat high elevations, since evergreen Pinaceae species such as spruce (Picea),fir (Abies), and sometimes pine (Pinus) form most alpine treelines. Otherconifers, such as hemlock (Tsuga), juniper (Juniperus), and cypress(Chamaecyparis), have local representation at treeline in some parts of theworld. Evergreen species can photosynthesize year-round whenever condi-tions are favorable; deciduous trees can only photosynthesize followingspring flush and before leaves are dropped in the autumn (or wet/dry season,depending on the local pattern). Furthermore, many conifers have highly
TABLE 2 Differences between low and high elevation
At a given latitude, lower elevation is associated with . . .
Low elevation High elevation
Mean temperature High LowDiurnal variation in temperature Low HighGrowing season length Long ShortPrecipitation Low HighWind Low HighSoil quality High LowTotal ecosystem energy High LowRadiation intensity when clear Low HighUV intensity Low HighPartial pressure of CO2 High Low
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

226 A. D. Richardson and A. J. Friedland
xeromorphic leaves that are very resistant to the desiccation that is commonat high elevations and high latitudes. However, deciduousness may alsohave some advantages in extreme environments, such as reduced water lossduring times of low potential photosynthetic activity, or reduced trapping ofsnow, which could minimize broken branches. Indeed, the deciduous larch(Larix gmelini), a conifer, is known to form the Arctic treeline in Siberia, at72° 30′ N. Some birch (Betula) species form the treeline in Japan, Scotland,Russia, and parts of Scandinavia; and other angiosperms, such as alder(Alnus), beech (Fagus), mountain ash (Sorbus), and poplar (Populus) arealso found at treeline in other parts of the world.
The physiognomy of treeline varies among regions around the world.In temperate regions, there is generally a progression from closed forests oflarge timber (broadleaf species at the base of a mountain, conifers higherup), to smaller and more widely-spaced trees (the Kampfzone), to shrubby,deformed, and often prostrate Krummholz, to treeless alpine vegetation(Berlyn, 1993). However, this pattern is by no means universal. For example,in the Rocky Mountains of Alberta, tall Picea and Abies define the forestlimit, and although somewhat stunted trees of these two species continue tothe treeline, there is generally no evidence of flagging or deformation (Wardle,1965). On the other hand, in Colorado, similar species form the irregular butclearly-defined forest limit and less-clearly-defined treeline, but beyond thetreeline there are often many flagged, prostrate, and deformed Krummholzthickets (Griggs, 1946; Wardle, 1965). Griggs (1946) observed many seedlingsabove treeline: He suggests they germinated from seeds blown up from lowelevations, since the highest elevation trees seldom produce cones or viableseed. At high elevation in the northern Appalachians, dwarfing of balsam firbegins as much as 600 m below treeline (Griggs, 1946). This is in contrast tothe pattern in the northern Rockies, where tall, large-diameter trees arefound right up to treeline (Griggs, 1938). In the Appalachians, Griggs reportedthat flagging occurs when an individual stem is higher than surrounding stems.However, the vertical growth habit is maintained and stem density remainsextremely high all the way to treeline. Above treeline, islands of Krummholzscrub continue to within 300–600 m of the summit of Mt. Washington (1916 mabove sea level), although some individual stems survive in sheltered loca-tions within 25 m of the summit.
In the southern hemisphere, the pattern is usually quite different. First,most treeline species are broadleaf evergreens from many different families.There is much more diversity in species mixture than in the northern hemi-sphere. A variety of mountain beech (Nothofagus) and eucalypt (Eucalyptus)species form treeline in different parts of New Zealand and Australia, wheretreeline tends to be very abrupt and clearly-defined, and there is noKrummholz (Wardle, 1965). On the other hand, at high elevation in thehumid tropics there is often a very sudden treeline without Krummholz,rather just a rapid transition to Páramo grasslands (high Andes) or Afro-alpine
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 227
(east and central Africa) vegetation (Troll, 1973). In South America, high-elevation vegetation and may include “megaphytes,” or large arborescentherbs, such as Espeletia, which may or may not be considered trees.Similar ambiguous examples, sometimes involving tree ferns (e.g.,Cyathea) are common in the mountains of both Africa and New Guinea(Wardle, 1974).
These differences in treeline physiognomy around the world are significantbecause they once again bring into question the existence of a single treeline-determining factor that can be considered universal across both alpine andArctic treelines.
THEORIES TO EXPLAIN TREELINE
We will now review and critique the major theories advanced to explain thelocation of treeline. Quite a few of these are more than half a century oldand they are not currently held in high esteem. Nevertheless, presenting themhere provides historical context and information upon which the newertheories are based. As will be seen, few theories actually enable directprediction of treeline location. Most theories fail to account for differencesin seasonality between temperate and tropical treelines. Other theories failto account for the sudden shift, by a single species, from vertical to prostrategrowth form that is found at most treelines.
Excessive Light
On clear days, radiation intensity at high elevations can be severe in somemountain ranges, and one early theory for treeline suggested that treelinemarked the point at which light intensity became too high for trees to withstand(reviewed by Daubenmire, 1954). Unfortunately, this theory doesn’t explain thepresence of treelines in foggy coastal mountains, like the Cascades and CoastRanges of the Pacific Northwest. In the White Mountains of New Hampshire,incoming solar radiation differs little between low and high-elevation,despite frequent clouds at high elevation, and yet the treeline is as low asanywhere in the coterminous United States (Richardson et al., 2004). Nordoes the theory give any explanation for why alpine shrubs and herbs cantolerate the high light intensity, but trees cannot.
However, sky exposure has been implicated as a factor limiting seedlingregeneration at treeline: Germinants that receive cover from neighboringplants have a higher survival rate than exposed seedlings that experiencefull sunlight (Germino, Smith, & Resor, 2002; Maher & Germino, 2006). Inthis regard, it is suggested that intense solar radiation further aggravates thecold temperature and drought stresses to which high-elevation seedlings aresubjected (Germino et al., 2002).
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

228 A. D. Richardson and A. J. Friedland
Low CO2 Partial Pressure
Because the barometric pressure decreases with increasing elevation, thepartial pressure of CO2 is lower at high elevations. Treeline has been attrib-uted to the effects of reduced CO2 on photosynthetic rates (reviewed byDaubenmire, 1954). However, Gale (1972) argued on theoretical groundsthat the depressed partial pressure of CO2 may have little effect on the avail-ability of CO2 for photosynthesis in plants, because the diffusivity of CO2increases at roughly the same rate as the partial pressure decreases; this hasbeen further debated by others (Cooper, 1986; Gale, 1986; Lamarche, Graybill,Fritts, & Rose, 1986; Terashima, Masuzawa, Ohba, & Yokoi, 1995). In anycase, it cannot explain the decrease in treeline elevation that occurs withincreasing latitude, since changes in barometric pressure with elevation aremore or less independent of latitude.
Snow Depth
Snow depth may modify treeline elevations to some degree since snowpackthat lingers to midsummer can significantly shorten the already-short grow-ing season at high elevations and high latitudes. For example, in parts of theSelkirk Mountains, BC, snow accumulates in depressions and remains therethrough the end of the summer: This prevents establishment of seedlings inhollows, while nearby snow-free ridges at the same elevation often havemature stands of timber (Daubenmire, 1954). Fonda and Bliss (1969) similarlyattribute local depression of treeline in the Olympic Mountains, WA, to latesnowmelt. But this theory cannot explain why snow-free ridges at highelevations don’t have trees, nor why some tropical mountains with little orno annual snowfall still have treelines. Variation in snow pack probably is ahighly significant factor in local treeline variation in some parts of theworld.
Wind
Wind appears to play a role in many northern hemisphere treelines, sinceflagging and desiccation of high-elevation trees are common (Griggs, 1938;Lindsay, 1971; Hadley & Smith, 1983) and may cause breakage or bucklingof exposed branches and even trunks. Strong prevailing winds may causeeccentric stems to develop (in the direction of wind loading), and may, inextreme cases, result in the formation of reaction wood (G. P. Berlyn, personalcommunication). In many instances, only sheltered individuals can surviveat highest elevations, and treeline is often higher on leeward slopes. Neigh-boring trees (and Krummholz mats) provide protection from wind abrasion,which promotes height growth, and thus facilitation as a positive factoroutweighs competition as a negative factor (Smith, Germino, Hancock, &
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 229
Johnson, 2003). Wind is thought to be a major factor determining treeline inthe northern Appalachians, since summer treeline temperatures are consid-erably warmer than have been reported at other treeline sites around theworld, and Krummholz showing evidence of wind damage is abundant(Cogbill & White, 1991). Based on studies with Picea and Abies, Hadley andSmith (1983) concluded that winter snow might protect prostrate Krummholzmats from desiccation by high winds. They showed that exposed shootssuffered from wind abrasion that resulted in low cuticular resistance. How-ever, since wind abrasion does not appear to be a factor near the equator orin the southern hemisphere (Daubenmire, 1954), it seems clear that whilewind may cause local variation in treelines, it is not the driving force behindArctic and alpine treelines in general. Furthermore, Marchand (1996) hasargued that winter wind will actually decrease transpiration, and others havefound little evidence for a wind effect (Baig & Tranquillini, 1980; Sowell,Koutnik, & Lansing, 1982). With the above arguments in mind, however,wind seems to be a convincing explanation for some of the variation inphysiognomy seen in the world’s treelines.
Reproduction
For treeline to advance upwards, successful germination and establishmentof new seedlings above the existing treeline is necessary (Smith et al.,2003). Reproduction rates at treeline are often quite low: Seed development,dispersal, germination, and seedling establishment are all limited by cold(although dispersal is enhanced by wind). It has been hypothesized thatreproductive failure or recruitment limitation may be a defining characteristicof high-elevation and high-latitude treelines (see Stevens & Fox, 1991, anddiscussion by Körner, 1998). While Tranquillini (1979) and Körner (1998)questioned this hypothesis, other evidence points to seedling survivivalbeing a major factor in determining treeline location (Germino et al., 2002).In the case of a treeline that is advancing (e.g., examples given above), it isto be expected that seedlings would be found above treeline, as reportedby Griggs (1946). However, Wardle’s experiments (1965, 1971) showed thatyoung seedlings are especially susceptible to the abiotic stresses characterizingthe above-treeline environment (see also Johnson, Germino, & Smith, 2004).In particular, there is a climatic constraint imposed by getting out of thewarm boundary layer found near ground level (Körner, 1998; Smith et al.,2003); the prostate growth form of Krummholz may be an adaptiveresponse to remain within this boundary layer.
Just as the Krummholz mat is a form of self-facilitation (providing shelterand enhancing winter survival by trapping snow, and enhancing photosyn-thesis during cooler weather by creating a more favorable thermal environ-ment), so too is microsite facilitation critical for seedling survival at treeline(Smith et al., 2003). Both Germino et al. (2002) and Maher and Germino
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

230 A. D. Richardson and A. J. Friedland
(2006) reported that seedling survival at a number of treeline sites in theRocky Mountains was highest when neighboring vegetation provided shel-ter, which reduced both sky and wind exposure. Sky exposure is correlatedwith warmer needle temperatures during the day and colder needle temper-atures at night, so seedlings growing in the shelter of mature trees experiencea narrower range of temperature extremes (Smith et al., 2003). The combi-nation of warmer nighttime temperatures and reduced daytime irradiancesreduces the likelihood of photoinhibition, while cooler daytime tempera-tures reduce the likelihood of water stress (Germino et al., 2002). Both factorscontribute to a better carbon balance and, ultimately, improved seedling sur-vival (but note that Johnson and Smith, 2005, reported high seedling mortalityunder a fully closed canopy, where shading was excessive and light levels toolow to maintain a positive carbon balance). Smith et al. (2003) have hypothe-sized a positive feedback loop resulting from facilitation resulting in betterseedling survivorship and subsequent improved growth, which leads to evengreater facilitation for subsequent generations of seedlings, and have sug-gested that this facilitation is a prerequisite for upward treeline migration tooccur. Unfortunately, this hypothesis does not include a mechanism (otherthan carbon limitation) by which climatic limitations ultimately exceed thebenefits of facilitation, and thus prevent further advancement of the treeline.
Winter Desiccation: Frosttrocknis
Frost drought, or Frosttrocknis, occurs during late winter when the soil isfrozen but skies are clear and solar radiation is high: As exposed foliagewarms in direct sun, a strong vapor pressure deficit is created, evapotranspi-ration from the leaf is high, and desiccation occurs. Frost drought isaccepted as a major cause of shoot dieback at timberline in the European(Daubenmire, 1954) and Japanese (Sakai, 1970) Alps. Although this mayplay a role in determining treeline in these ranges, it is not thought to be auniversal phenomenon (Marchand & Chabot, 1978; Kincaid & Lyons, 1981;Körner, 1998). For example, in a study of three conifers in the elevationsjust below treeline in New England, Vostral, Boyce, and Friedland (2002)did not find evidence of winter desiccation stress in any of the species.Furthermore, winter frost drought does not occur in aseasonal tropicalmountain ranges. Interestingly, frost drought is not likely to happen in thecoldest part of the year, which may be why minimum temperature does notseem correlated with treeline position.
Unfortunately, testing any sort of winter damage hypothesis is difficultbecause the damage is only obvious considerably after it has occurred.Often it is difficult to differentiate between desiccation and frost damage(Perkins, Adams, & Klein, 1991). However, there is some evidence thatfrozen soil is not the real cause of frost drought, although for many years itwas assumed that frozen soil was the cause. Investigating the problem in
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 231
New England, Marchand (1996) studied winter water relations in Picearubens and Abies balsamea. Marchand demonstrated that even when it wasimpossible for water replenishment to occur (due to a severed stem orfrozen soil), foliar desiccation did not occur until late spring. Under normalconditions, there is enough water stored in the tree’s stem to survive moderatecuticular transpiration throughout the winter. Therefore, frozen soil alone isnot the cause of winter drought: elevated needle temperatures resulting indramatically increased cuticular transpiration must occur, and then thewater reserves in the stem must be exhausted. Similarly, Sowell, McNulty,and Schilling (1996) have shown that stem water can recharge needlesthroughout the winter. It is clear that while some winter damage may contributeto local fine-tuning of the treeline, frost drought due to frozen soil is notitself the determinant of treeline positions around the world.
Temperature
There is considerable evidence for a direct link between temperature andtreeline. For example: (a) south-facing slopes have a higher treeline thannorth-facing slopes (in the Northern hemisphere); (b) treeline is higher inthe Rockies (clear, warm summers) than New England (cloudy, cool summers);and (c) treeline decreases in elevation with increasing latitude. Indeed, oneof the oldest explanations for treeline is that it is determined solely bytemperature. A good correlation between treeline position and the 10°Cmean July isotherm was demonstrated in early work (Daubenmire, 1954;but see more recent analyses by Körner, 1998 and Virtanen et al., 2004).Certainly, both Arctic and alpine treelines have cold, inhospitable climates,with mean growing season air and soil temperatures consistently falling inthe 5–8°C range (Körner, 1999; Körner & Paulsen, 2004). However, neithergrowing season length, which ranges from 92 days in the Sierra Nevada ofCalifornia, to year-round (or close to it) in the tropics, nor accumulated heatsums, nor temperature extremes, correlates globally with treeline location(Körner & Paulsen, 2004). Interestingly, although air temperature heat sums(daily means above 10°C) differ between Arctic (temperature sum of 600–700°C)and alpine (200–300°C) timberlines, it has been observed that leaf tempera-ture heat sums are similar (800°C) at both high-latitude and high-elevationtimberlines (Davitaja & Melnik 1962, cited in Tranquillini, 1979). To the bestof our knowledge, this analysis has not been repeated by other researchersin other treeline ecosystems, but it could potentially provide the foundationfor a theory that simultaneously explains both high-elevation and high-latitudetreeline positions.
Thus, it is not entirely clear what the temperature-based mechanism mightbe: Does temperature limit photosynthesis, reproduction, growth, or someother physiological process? When are temperatures most limiting? Are meantemperatures, extreme temperatures, air temperatures, soil temperatures, or
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

232 A. D. Richardson and A. J. Friedland
perhaps even leaf temperatures, critical? We will now compare a number oftemperature-based theories for treeline.
It is interesting to note that, by very nature of their vertical growthhabit, trees expose themselves to a microenvironment that is unfavorablefor growth. The degree to which shoot meristems are coupled with theambient environment depends on radiation, wind speed, and vegetationheight. Boundary layer effects mean that increased radiation and reducedwind speed dramatically increase the meristem temperatures of low profilevegetation (e.g., Krummholz, as well as alpine cushion plants) relative to airtemperature, and this can be a significant advantage in cold environments(Grace, 1989). For tall trees, meristems are more tightly coupled to theambient air temperature, which is a disadvantage when temperatures arebelow the photosynthetic temperature optimum. Closely spaced trees alsoshade and insulate their own soil; soil temperature under a forest stand attreeline is generally close to the mean annual air temperature (Grace, 1989;Körner, 1998). By comparison, the open spacing of Krummholz islands permitsmuch more soil warming to occur (Körner & Paulsen, 2004). These factorscombine to make the prostrate growth form far more suitable than a vertical,arborescent form for survival at high elevations and high latitudes (butnote that these arguments are counter to those of Smith et al., 2003, whoproposed that neighboring trees improved the microclimate for survival, asdiscussed above for seedling survival).
Temperature: Carbon Balance
With increasing elevation, lower temperatures result in lower maximal ratesof photosynthesis and a shortened growing season. As reviewed by Grace(1989), high-elevation trees photosynthesize at a 30–50% lower rate thanlow-elevation trees, and low temperatures decrease the quantum yield ofphotosystem II because turnover of the protein electron acceptor Qb isrestricted in cold environments, which results in photoinhibition (see alsoRichardson & Berlyn, 2002). The carbon balance theory suggests that treelineoccurs at the point where carbon fixation by photosynthesis, frequentlyrestricted to a short growing season, is offset by respiratory losses and theproduction of foliage, with no carbon left over for wood production(Stevens & Fox, 1991). For example, large reductions in growth incrementof Nothofagus have been observed across an elevational difference of ≈300 mand a temperature difference of only a few °C (Wardle, 1985).
It is thought to be more difficult for large trees to have a positive carbonbalance at high elevation than it is for small trees, which may explain whytree size decreases with increasing elevation, as well as the transition fromvertical “tree” to prostrate Krummholz. Trees are a relatively inefficientgrowth form in that production of woody stems and roots both require largeamounts of carbon. The ratio of photosynthetic tissue to total biomass
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 233
decreases with increasing tree size (Daubenmire, 1954; Stevens and Fox,1991) and respiration of non-photosynthesizing tissue may impose a signifi-cant carbon burden. On the other hand, prostrate Krummholz and othershrubs can survive at higher elevations because of a more favorable micro-climate near ground level and more efficient carbon allocation patterns(Daubenmire, 1954; Stevens & Fox, 1991; Smith et al., 2003).
The carbon balance theory neglects a number of important facts. First,moisture status is generally favorable to photosynthesis during the growingseason at high elevations—often more favorable than it is at lower elevations(Körner, 1998). Trees at high elevation may have lower maximal rates ofphotosynthesis, but when integrated over the growing season, total carbonfixation may actually be quite high. Second, satisfactory growth has beenshown to occur in Nothofagus seedlings at 300 m above local treeline,which suggests that carbon fixation at treeline is not inadequate (Wardle,1971), although carbon budgets for mature trees could be quite different. Infact, of the total carbon production of Pinus cembra at treeline in Austria,only one third was required for respiration and biomass production (seereference in Wardle, 1974). Third, most tropical high-elevation ecosystemsare marked by little or no seasonality, and thus while carbon balance maybe a factor when the growing season is short, it is probably not the decidingfactor in tropical mountain ranges with a year-round growing season.
Temperature: Cuticle Ripening Theory
Wardle (1965, 1968, 1971) resurrected the decades-old hypothesis, advancedby Michaelis (1934a, 1934b; cited by Wardle, 1965; and Tranquillini, 1979)in Ökologische Studien an der alpinen Baumgrenze (“Ecological studies ofthe alpine timberline”). This is another temperature-based explanationfor treelines; it also combines elements of the frost drought theory. Wardlesuggested that at high elevations, newly formed tissue might be unable to“ripen” due to the short growing season. According to Wardle, ripening hasseveral components, including anatomical (cuticle development and lignifica-tion) and physiological (ability to withstand low temperatures and desiccation)aspects. Inadequately ripened shoots cannot withstand winter desiccation,and Frosttrocknis results. Since low-lying Krummholz vegetation is oftenprotectively covered by snow during late winter when frost drought is liableto be most severe, only those shoots which stick up above the snow arelikely to be killed off (Wardle, 1974). Only the prostrate form can thussurvive above a certain elevation.
Wardle published a series of papers which provided evidence insupport of the ripening hypothesis: his 1965 comparison of timberlines inNew Zealand and North America; his 1968 study of Picea in Colorado; andhis 1971 study of Nothofagus in New Zealand. In the first paper, he relateddeath of high-elevation seedlings to desiccation and the failure of some
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

234 A. D. Richardson and A. J. Friedland
shoots to develop fully. In the second paper, he hypothesized that winterdesiccation of needles is influenced by events in preceding seasons, andthat this resulted in inadequate hardening against winter. In the third paper,Wardle demonstrated that growing season length was well correlated withthe degree of tissue maturation and with elevation. Bud flush occurred laterat higher elevations, and so the time available for ripening thus decreased.Wardle concluded that the short growing season caused incomplete devel-opment of the cuticle, epicuticular wax, and lignin. Foliar mortality resultedfrom excessive desiccation due to unripe cuticles. Many other authors haveprovided evidence that cuticle thickness at treeline is impaired, and thatthese trees may be prone to high levels of cuticular transpiration (Platter,1976; Baig & Tranquillini, 1976, 1980; Delucia & Berlyn, 1984; Hansen-Bristow,1985; Berlyn et al., 1993).
The strength of the cuticle ripening hypothesis is that it offers a decentmechanism for the sudden change from erect tree to prostrate Krummholz:During the summer, elevated crown tissue is in a less favorable microclimate,so it is unable to ripen. In the winter, any foliage not close to ground leveldoes not get covered in snow, and is subject to desiccation. However, likemany other treeline hypotheses, the cuticle ripening theory fails to be a uni-versal explanation because tropical mountains don’t have seasonality thatwould interfere with cuticular ripening. Even where winter is a factor, cuticlesmay still ripen properly at treeline. Grace (1990) demonstrated that in themountains of Scotland, there was no evidence of poor cuticle developmenton high elevation Pinus sylvestris, as cuticle mass per unit of leaf surfacearea was not related to elevation (however, it is worth noting that mostother authors have looked at cuticle thickness, which could be easier tomeasure accurately). Although high-elevation needles transpired more thanlow-elevation needles, Grace attributed this to stomatal dysfunction resultingfrom mechanical damage from blowing ice. Needles that were visibly damagedhad especially high rates of transpiration.
Other studies have shown that winter desiccation is not universal. Thetissue ripening theory requires that lethal desiccation through the cuticleoccur. As described above, it has been clearly shown that the cause of winterdrought is not frozen soil; there is also considerable evidence that winterdrought does not occur everywhere, and that even where it does occur, thecause may not be poorly developed cuticles (Hadley & Smith, 1986; Perkinset al., 1991).
Temperature: Tranquillini Integration
Tranquillini basically extended Wardle’s work. Tranquillini (1979) assertedthere are three possible causes of timberlines: an unfavorable carbon balance,an arrested phenological cycle, and inadequate plant resistance to climaticstress. Through a chain of cause and effect, the short growing season results
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 235
in immature plant tissues, particularly the cuticle. Trees then enter winterwith reduced resistance to transpiration. Finally, excessive water loss occursduring late winter as a result of both accelerated cuticular water loss andfrozen soil.
Although it proposes a slightly more complex mechanism, the Tran-quillini theory fails for the same reasons as both the Wardle theory and thetraditional frost drought theory. However, it is hard to dispute that in someparts of the world there appears to be a striking relationship between increas-ing altitude/latitude and decreasing cuticle thickness/resistance. Although thismay not actually be the cause of timberlines, it should definitely be a keyfactor to look at in order to understand how plants in such extreme habitatsfunction in relation to their environment.
Temperature: Impaired Sink Theory
The most recent comprehensive treeline theory centers on growth limita-tion. Körner (1998, 1999) proposed that cell division and differentiation intofunctional tissue is shut off at temperatures below a certain threshold, whichis thought to be between 3°C and 10°C, probably in the 5.5–7.5°C range.Körner (1998, 1999) hypothesized that even though there is enough carbonbeing fixed at treeline, temperatures are too low for carbon sinks likegrowth and renewal, since cellular development is impaired. Cold nights forcanopies coupled to the ambient atmosphere may mean that shoot expansioncannot occur at night. Trees shade and insulate their own root zone (Körner &Paulsen, 2004) and cold soils probably impair root growth. The resultingreduced meristem activity, both aboveground and belowground, means thatinhibited sinks rather than sources are the limiting factor for growth of tree-likeforms at high elevations. There is evidence for this sort of threshold temperaturein the root and shoot growth of Pinus cembra. A literature survey suggeststhat freezing is not involved with this threshold. Rather, it appears that celldoubling time rises asymptotically at ≈1–2°C (Körner, 1999). Furthermore,because photosynthesis at these temperatures is only reduced by ≈70% relativeto the temperature optimum, it is clearly possible for sink limitation to occurat temperatures sufficient for an adequate carbon supply.
This theory is supported by work reported by Tranquillini (1979) whichshowed that growth rates were reduced by as much as four-fold with onlysmall (<300 m) increases in elevation: This is much greater than any changewe would expect in incident radiation or a temperature-related photosyn-thesis effect. Furthermore, Körner (1998) noted that ring widths frequentlyare doubled during a growing season that is only modestly warmer (2–3°C) attreeline, and growth of Picea abies seedlings has been shown to be dramaticallyreduced at low temperatures. Radial growth has also been shown to belinearly correlated with temperature above a minimum temperature of ≈7–8°C(Grace, 1989). Grace, Berninger, and Nagy (2002) found zero growth of
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

236 A. D. Richardson and A. J. Friedland
Pinus sylvestris at 5°C, despite the fact that photosynthetic rates at thattemperature were still ≈30% of the rate at 20°C.
As suggested by other temperature-based theories, trees may, by thevery nature of their growth form, render themselves unsuitable for growth attreeline. Although Körner’s theory relies on the existence of a very suddenthreshold temperature at which cellular growth is reduced to zero, it can beimagined how, because of differences in microenvironment, tall trees couldfall on one side (the wrong side) of this threshold, whereas low-lying, prostatevegetation, in a more favorable microclimate, is not subject to temperaturelimitations. The main advantage of this theory is that it would appear toapply universally from the equator to high latitudes.
RECENT FIELD STUDIES
In recent years, a number of novel experiments have been conducted withthe hopes of determining which (if any) of the above theories for treeline iscorrect. Research has focused on the carbon limitation and impaired sinkhypotheses. Körner (2003) proposed that tissue concentrations of nonstruc-tural carbohydrates (NSC) offer insight into the balance between carbonsupply and demand, i.e., between photosynthetic production and therequirements for growth. A recent study found that Pinus cembra at thehighest elevations in the Alps accumulated more NSCs than those at lowerelevations (Hoch, Popp, & Körner, 2002), and a larger study with data fromPinus spp. at sub-tropical (Mexico) and sub-Arctic (Sweden) latitudes gaveadditional support for this pattern (Hoch & Körner, 2003). From lowlandtropics to temperate treeline, there appears to be a generally inverse rela-tionship between growing season temperature and NSC concentrations(Körner, 2003). Thus, from these results it could be concluded that thesupply of carbon does not limit growth at treeline. Rather, sink inhibition(i.e., tissue formation) limits growth (this conclusion has, however, beencriticized; Johnson et al., 2004, for example, suggest that maintaining a largepool of carbon reserves may simply be a strategy to increase survival in astressful and unpredictable environment; see also Chapin, Schulze, & Mooney,1990). Interestingly, however, Körner (2003) found that in a variety of differentecosystems (including three high-elevation treeline forests, as well as atemperate lowland forest and a semi-deciduous tropical forest), NSC con-centrations tended to be high throughout the year, even during periods oflow growth. Thus, it appears that growth of forest trees around the world,and not just at treeline, may be sink-limited, rather than carbon-limited.(Slow rates of phloem transport have been implicated as a cause of slowgrowth in some Pinus caribaea individuals; see Anoruo and Berlyn, 1993.Since phloem transport would be inhibited by cold temperatures, this problemcould be exacerbated at treeline.)
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 237
Results of a recent high-elevation free-air CO2 enrichment (FACE) study,led to the opposite conclusion (Hättenschwiler et al., 2002). Because growthof both Larix decidua and Pinus uncinata was stimulated by FACE treatmentin the first year, the authors concluded that tree growth at high elevationsmust be limited by carbon availability. Not discussed by Hättenschwileret al. was the fact that the FACE treatment also lead to increased foliar accu-mulation of NSCs, evidence that would tend to support the sink limitationhypothesis. However, Körner (2003) argues that foliar NSC concentrationsmay not reliably indicate source-sink relationships in CO2 enrichment studies.Rather, inference must be based on stem tissue NSC concentrations.
Handa, Körner, and Hättenschwiler (2005) reported on years 2 and 3 ofthe FACE experiment described by Hättenschwiler et al. (2002). AlthoughLarix decidua shoot growth was stimulated by enrichment in all 3 years,for Pinus uncinata, there was no enrichment effect in years 2 or 3. Thissuggests that L. decidua may be carbon limited at treeline, whereas P. uncinatagenerally is not; however, a defoliation experiment conducted during the2nd year of enrichment indicated that carbon limitation might periodicallyoccur in response to extreme disturbances at treeline. It is possible thatthese somewhat conflicting results relate to the fact that the present treelinelocation is indicative of past climatic conditions.
CLIMATE CHANGE AND TREELINE
Responses to climate change are expected to be largest at climate-drivenecotones (see Noble, 1993, for a review and critique). There is considerableinterest in the effects of climate change on treeline location: There are clearimplications for both changes in forest structure below the treeline, andbiodiversity of alpine ecosystems above the treeline (Grace et al., 2002).
Analysis of growth ring width along an elevational gradient neartreeline in the Swiss and Austrian Alps indicates a strong relationshipbetween increasing elevation and decreasing ring width for the period1800–1940 (Paulsen, Weber, & Körner, 2000). However, since 1940, Paulsenet al. (2000) found no such trend; the authors suggest that with the pastcentury and a half of warming, minimum temperature requirements forgrowth are now consistently met all the way up to the treeline. Based onhistoric relationships between treeline location and temperature, it is generallyexpected that the present warming trend will cause an upslope migration ofthe treeline. For this to occur, however, new seedlings must become estab-lished above the present treeline location. Smith et al. (2003) identifiedfour forms of microsite facilitation (inanimate, interspecific, intraspecific,and structural) that promote establishment, carbon gain, and ultimately,enhanced growth and survival, of seedlings at treeline (see also Stevens &Fox, 1991).
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

238 A. D. Richardson and A. J. Friedland
The results of Hättenschwiler et al. (2002) would seem to indicate thatrising atmospheric CO2 may be as important as recent temperature increasesfor the health and productivity of trees at the highest elevations, and Handaet al.’s (2005) results indicate that the capacity of alpine trees to respond toCO2 increases is very species-specific. Differential effects of enrichmentcould lead to changes in the competitive balance among species, and mightultimately cause changes in the species composition of treeline forests (seealso Richardson, Denny, Siccama, & Lee, 2003).
However, trends in NSC concentrations reported by Hoch et al.(2002) and Körner (2003) appear to indicate that trees at high elevationsalready take up more CO2 than they can effectively use for growth.Based on Körner’s (1998) impaired sink hypothesis, rapid increases intree growth are predicted in response to climatic warming, as the capacityfor tissue development at low temperatures is very sensitive to temperatureincreases.
SUMMARY AND CONCLUSIONS
In this review, we have described general patterns of treelines around theworld, and compared the physiognomy of treelines in a variety of differenthigh-elevation and high-latitude ecosystems. The variability across differentenvironments suggests that perhaps there is no single theory that can explainthe position of treelines from the equator to the Arctic. Many of the oldertheories to explain treeline, such as excessive light, low carbon dioxide partialpressure, snow depth, desiccation, and wind may help to explain specifictreeline visual patterns, but they fail to provide a globally relevant explanationfor treeline itself. Recent work on factors limiting seedling survival at treeline ishighly relevant, given anticipated changes in treeline location in response toclimate change, and has resulted in interesting hypotheses about the role offacilitation in promoting survival, carbon gain, height growth, and ultimately,treeline advancement. That being said, perhaps the most convincing proposalis Körner’s impaired sink hypothesis, which not only relates treeline tomicroclimatic differences between trees and prostrate vegetation, but alsoprovides a mechanism that, at least in theory, should be effective globallyfrom tropics to Arctic. Furthermore, the recent work demonstrating thattrees at the highest elevations accumulate more non-structural carbohy-drates than those at lower elevations gives strong empirical support for thistheory.
Treelines have fascinated biologists for centuries, and even if theimpaired sink hypothesis does emerge as the consensus theory toexplain treelines around the world, it is certain that forests at the limitsof growth will continue to provide exciting research opportunities inthe future.
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 239
REFERENCES
Anoruo, A. O., & Berlyn, G. P. (1993). Effect of rate of growth and development onneedle photosynthate and phloem transport in Caribbean pine (Pinus caribaeaMor.). Journal of Sustainable Forestry, 1, 127–141.
Baig, M. N., & Tranquillini, W. (1976). Studies on upper timberline: Morphologyand anatomy of Norway spruce (Picea abies) and stone pine (Pinus cembra)needles from various habitat conditions. Canadian Journal of Botany, 54,1622–1632.
Baig, M. N., & Tranquillini, W. (1980). The effects of wind and temperature oncuticular transpiration of Picea abies and Pinus cembra and their significancein desiccation damage at the alpine treeline. Oecologia, 47, 252–256.
Barry, R. G. (1992). Mountain weather and climate (2nd ed.). New York:Routledge.
Beaman, J. H. (1962). The timberlines of Iztaccihuatl and Popocatepetl, Mexico.Ecology, 43, 377–385.
Berlyn, G. P. (1993). Trees. In The new encyclopaedia Britannica (Vol., pp.).Chicago: Encyclopaedia Britannica.
Berlyn, G. P., Anoruo, A. O., Johnson, A. H., Vann, D. R., Strimbeck, G. R., Boyce,R. L., et al. (1993). Effects of filtered air and misting treatment on cuticlesof Red spruce needles on Whiteface Mountain, NY. Journal of SustainableForestry, 1, 25–47.
Billings, W. D. (1973). Arctic and alpine vegetations: Similarities, differences andsusceptibility to disturbance. BioScience, 23, 697–704.
Bliss, L. C. 1956. A comparison of plant development in microenvironments ofArctic and alpine tundras. Ecological Monographs 26: 303–337.
Brink, V. C. (1959). A directional change in the subalpine forest-heath ecotone inGaribaldi Park, British Columbia. Ecology, 40, 10–16.
Chapin, F. S., Schulze, E.- D., & Mooney, H. C. (1990) The ecology and economicsof storage in plants. Annual Review of Ecology and Systematics, 21, 423–447.
Cogbill, C. V., & White, P. S. (1991). The latitude-elevation relationship forspruce-fir forest and treeline along the Appalachian mountain chain. Vege-tatio, 94, 153–175.
Cooper, C. F. (1986). Carbon dioxide enhancement of tree growth at high elevations.Science, 231, 859–859.
Cwynar, L. C., & Spear, R. W. (1991). Reversion of forest to tundra in the centralYukon. Ecology, 72, 202–212.
Daubenmire, R. F. (1943). Vegetational zonation in the Rocky Mountains. BotanicalReview, 9, 325–391.
Daubenmire, R. (1954). Alpine timberlines in the Americas and their interpretation.Butler University Botanical Studies, 11, 119–136.
Davitaja, F. F., & Melnik, J. S. (1962). Radiation heating of the plants active surfaceand the latitudinal and altitudinal limits of forest (in Russian). Meteorol. Gidrol.Moskva, 3–9.
DeLucia, E. H., & Berlyn, G. P. (1984). The effect of increasing elevation on leafcuticle thickness and cuticular transpiration in balsam fir. Canadian Journal ofBotany, 62, 2423–2431.
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

240 A. D. Richardson and A. J. Friedland
Fonda, R. W., & Bliss, L. C. (1969). Forest zonation of the montane and subal-pine zones, Olympic Mountains, Washington. Ecological Monographs, 39,271–301.
Gale, J. (1972). Availability of carbon dioxide for photosynthesis at high altitudes:Theoretical considerations. Ecology, 53, 494–497.
Gale, J. (1986). Carbon dioxide enhancement of tree growth at high elevations.Science, 231, 859–860.
Germino, M. J., Smith, W. K., & Resor, A. C. (2002). Conifer seedling distributionand survival in an alpine-treeline ecotone. Plant Ecology, 162, 157–168.
Grace, J. (1989). Tree lines. Philosophical Transactions of the Royal Society(London) Series B, 324, 233–245.
Grace, J. (1990). Cuticular water loss unlikely to explain tree-line in Scotland.Oecologia, 84, 64–68.
Grace, J., Berninger, F., & Nagy, L. (2002). Impacts of climate change on the treeline. Annals of Botany, 90, 537–544.
Griggs, R. F. (1934). The edge of the forest in Alaska and the reasons for itsposition. Ecology, 15, 80–96.
Griggs, R. F. (1938). Timberlines in the northern Rocky Mountains. Ecology, 19,548–564.
Griggs, R. F. (1946). The timberlines of North America and their interpretation.Ecology, 27, 275–289.
Hadley, J. L., & Smith, W. K. (1983). Influence of wind exposure on needle desicca-tion and mortality for timberline conifers in Wyoming, U.S.A. Arctic and AlpineResearch, 15, 127–135.
Hadley, J. L., & Smith, W. K. (1986). Wind effects of needles on timberline conifers:Seasonal influence on mortality. Ecology, 67, 12–19.
Handa, I. T., Körner, C., & Hättenschwiler, S. (2005). A test of the treeline carbonlimitation hypothesis by in situ CO2 enrichment and defoliation. Ecology, 86,1288–1300.
Hansen-Bristow, K. (1985). Influence of increasing elevation on growth characteristicsat timberline. Canadian Journal of Botany, 64, 2517–2523.
Hättenschwiler, S., Handa, I. T., Egli, L., Asshoff, R., Ammann, W., & Körner, C.(2002). Atmospheric CO2 enrichment of alpine treeline conifers. New Phytologist,156, 363–375.
Hoch, G., & Körner, C. (2003). The carbon charging of pines at the climatic treeline:A global comparison. Oecologia, 135, 10–21.
Hoch, G., Popp, M., & Körner, C. (2002). Altitudinal increase of mobile carbonpools in Pinus cembra suggest sink limitation of growth at the Swiss treeline.Oikos, 98, 361–374.
Johnson, D. M., Germino, M. J., & Smith, W. K. (2004). Abiotic factors limitingphotosynthesis in Abies lasiocarpa and Picea engelmannii seedlings belowand above the alpine timberline. Tree Physiology, 24, 377–386.
Johnson, D. M., & Smith, W. K. (2005). Refugial forests in the southern Appala-chians: Photosynthesis and survival in current-year Abies fraseri seedlings. TreePhysiology, 25, 1379–1387.
Kincaid, D. T., & Lyons, E. E. (1981). Winter water relations of red spruce on MountMonadnock, New Hampshire. Ecology, 62, 1155–1161.
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

Theories to Explain Arctic and Alpine Treelines 241
Körner, C. (1998). A re-assessment of high elevation treeline positions and theirexplanation. Oecologia, 115, 445–459.
Körner, C. (1999). Alpine plant life. Berlin, Germany: Springer-Verlag.Körner, C. (2003). Carbon limitation in trees. Journal of Ecology, 91, 4–17.Körner, C., & Paulsen, J. (2004). A world-wide study of high altitude treeline
temperatures. Journal of Biogeography, 31, 713–732.Lamarche, V. C., Graybill, D. A., Fritts, H. C., & Rose, M. R. (1986). Carbon dioxide
enhancement of tree growth at high elevations. Science, 231, 860–860.Lindsay, J. H. (1971). Annual cycle of leaf water potential in Picea engelmannii
and Abies lasiocarpa at timberline in Wyoming. Arctic and Alpine Research, 3,131–138.
Maher, E. L., & Germino, M. J. (2006). Microsite differentiation among conifer spe-cies during seedling establishment at alpine treeline. Écoscience, 13, 334–341.
Marchand, P. (1996). Life in the cold: An introduction to winter ecology (3rd ed.).Hanover NH: University Press of New England.
Marchand, P. J., & Chabot, B. F. (1978). Winter water relations of tree-line plantspecies on Mt. Washington, New Hampshire. Arctic and Alpine Research, 10,105–116.
Mark, A. F. (1958). The ecology of the Southern Appalachian grass balds. EcologicalMonographs, 28, 293–336.
Michaelis, P. (1934a). Ökologische Studien an der alpinen Baumgrenze. IV. ZurKenntnis der winterlichen Wasserhaushaltes. Jahrb. Wiss. Bot., 80, 169–298.
Michaelis, P. (1934b). Ökologische Studien an der alpinen Baumgrenze. V. OsmotischerWert un Wassergehalt während des Winters in den Verschiedenen Höhenlagen.Jahrb. Wiss. Bot., 80, 337–362.
Noble, I. R. (1993). A model of the responses of ecotones to climate change.Ecological Applications, 3, 396–403.
Paulsen, J., Weber, U. B., & Körner, C. (2000). Tree growth near treeline: Abrupt orgradual reduction with altitude? Arctic, Antarctic and Alpine Research, 32, 14–20.
Perkins, T. D., Adams, G. T., & Klein, R. M. (1991). Desiccation or freezing? Mecha-nisms of winter injury to red spruce foliage. American Journal of Botany, 78,1207–1217.
Platter, W. (1976). Wasserhaushalt, cuticuläres Transpirationsvermögen und Duckeder Cutinschichten einiger Nadelholzarten in verschiedenen Höhenlagen undnach experimenteller Verkürzung der Vegetationsperiode. Doctoral disserta-tion, University of Innsbruck, Innsbruck, Austria.
Richardson, A. D., & Berlyn, G. P. (2002). Spectral reflectance and photosyntheticproperties of Betula papyrifera (Betulaceae) leaves along an elevational gradi-ent on Mt. Mansfield, Vermont, USA. American Journal of Botany, 89, 88–94.
Richardson, A. D., Berlyn, G. P., & Duigan, S. P. (2003a). Reflectance of Alaskanblack spruce and white spruce foliage in relation to elevation and latitude. TreePhysiology, 23, 537–544.
Richardson, A. D., Denny, E. G., Siccama, T. G., & Lee, X. (2003b). Evidence for arising cloud ceiling in eastern North America. Journal of Climate, 16, 2093–2098.
Richardson, A. D., Lee, X., & Friedland, A. J. (2004). Microclimatology of treelinespruce-fir forests in mountains of the northeastern United States. Agriculturaland Forest Meteorology, 125, 53–66.
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4

242 A. D. Richardson and A. J. Friedland
Sakai, A. (1970). Mechanism of desiccation damage of conifers wintering in soil-frozen areas. Ecology, 51, 657–664.
Salisbury, F. B., & Ross, C. W. (1992). Plant physiology. Belmont, CA: Wadsworth.Slatyer, R. O., & Noble, I. R. (1992). Dynamics of montane treelines. In A. J. Hansen
& F. di Castri (Eds.), Landscape boundaries: Consequences for biotic diversityand ecological flows (pp. 346–359). New York: Springer-Verlag.
Smith, A. P., & Young, T. P. (1987). Tropical alpine plant ecology. Annual Reviewof Ecology and Systematics, 18, 137–158.
Smith, W. K., Germino, M. J., Hancock, T. E., & Johnson, D. M. (2003). Anotherperspective on altitudinal limits of alpine timberlines. Tree Physiology, 23,1101–1112.
Sowell, J. B., Koutnik, D. L., & Lansing, A. J. (1982). Cuticular transpiration ofWhitebark pine (Pinus albicaulis) within a Sierra Nevadan timberline ecotone,U.S.A. Arctic and Alpine Research, 14, 97–103.
Sowell, J. B., McNulty, S. P., Schilling, B. K. (1996). The role of stem recharge inreducing winter desiccation of Picea engelmannii (Pinaceae) needles at alpinetimberline. American Journal of Botany, 83, 1351–1355.
Stevens, G. C., & Fox, J. F. (1991). The causes of treeline. Annual Review of Ecologyand Systematics, 22, 177–191.
Terashima, I., Masuzawa, T., Ohba, H., & Yokoi, Y. (1995). Is photosynthesis sup-pressed at higher elevations due to low CO2 pressure? Ecology, 76, 2663–2668.
Tranquillini, W. (1979). Physiological ecology of the alpine timberline. New York:Springer-Verlag.
Troll, C. (1973). The upper timberlines in different climatic zones. Arctic and AlpineResearch, 5, A13–A18.
Virtanen, T., Mikkola, K., Nikula, A., Christensen, J. H., Mazhitova, G. G., Oberman,N. G., et al. (2004). Modeling the location of the forest line in northeast EuropeanRussia with remotely sensed vegetation and GIS-based climate and terrain data.Arctic, Antarctic, and Alpine Research, 36, 314–322.
Vostral, C. B., Boyce, R. L., & Friedland, A. J. (2002). Winter water relations of threeNew England conifers at their upper elevational limits. I. Measurements. TreePhysiology, 22, 793–800.
Wardle, P. (1965). A comparison of alpine timber lines in New Zealand andNorth America. New Zealand Journal of Botany, 3, 113–135.
Wardle, P. (1968). Engelmann spruce (Picea engelmannii Engel.) at its upper limitson the Front Range, Colorado. Ecology, 49, 483–495.
Wardle, P. (1971). An explanation for alpine timberline. New Zealand Journal ofBotany, 9, 371–402.
Wardle, P. (1974). Alpine timberlines. In J. D. Ives & R. G. Barry (Eds.), Arctic andalpine environments (pp. 371–402). London: Methuen.
Wardle, P. (1985). New Zealand timberlines. 3. A synthesis. New Zealand Journal ofBotany, 23, 263–271.
Dow
nloa
ded
by [
Am
s/G
iron
a*ba
rri L
ib]
at 0
8:45
08
Oct
ober
201
4







![In den Baum geschaut - Young Scienceyoung-science-magazin.com › ... › 2016 › 07 › YoungScience_6_IndenB… · [5] Körner, C. (2012) Alpine treelines. Functional ecology of](https://img.pdfslide.net/doc/110x75/5f0deda77e708231d43cc601/in-den-baum-geschaut-young-scienceyoung-science-a-a-2016-a-07-a-youngscience6indenb.jpg)





















