Embed Size (px)
Citation preview

[ 777 ]
A STUDY OF OVARIAL DEVELOPMENT AND ITSRELATION TO ADULT NUTRITION IN THE BLOWFLY
PROTOPHORMIA TERRAE-NOVAE (R.D.)
BY PHYLLIS M. HARLOW, Ph.D., A.R.C.S.
Imperial College, London
{Received 12 July 1956)
I. INTRODUCTIONIn recent years considerable attention has been directed to the hormonal control ofinsect growth and, in particular, to those hormones which affect ovarial develop-ment. In contrast, although the nutritional requirements for growth of the immaturestages of several insects have been investigated, comparatively little is known of thefood requirements for ovarial growth specifically.
In the blowfly maturation of the ovaries is controlled by an endocrine complexincluding corpus allatum, corpus cardiacum and median neurosecretory cells ofthe brain (Thomsen, 1952). Evidence is also accumulating to show that the corpusallatum produces hormones which influence both protein and carbohydratemetabolism (Thomsen, 1942, 1949, 1952; Pfeiffer, 1945).
On the maintenance diet of sucrose and water the adult female blowfly neverlays eggs; ovarial growth ceases at an early stage, although normal changes frompupal to adult fat body and from ring gland to corpus allatum occur. When meat isadded to the diet of such flies growth of the ovaries, fat body and corpus allatumis resumed immediately and the ovaries mature a few days later. Since both corpusallatum and the fat body undergo further growth on the complete diet, it is probableon these grounds alone that their functions concern reproduction. Fraenkel (1940)showed that the sucrose of the maintenance diet could be replaced by certain othersugars or polyhydric alcohols but not by protein. The additional requirements forovarial maturation have been only partly investigated. Hobson (1938) showed thatat least two meat meals (approx. 30 ml. beef liver juice) were required for eachoviposition and that blood serum + Marmite, but not serum alone, could replacemeat. Dorman, Hale & Hoskins (1938) found that meat could not be replaced bycasein + butter + yeast, by soluble casein, by blood albumen, or by Lemco beefextract, and concluded that both protein and unidentified accessory substanceswere required. Banki (1937) was able to rear both larvae and adults on the followingmixture: agar, sugar, peptone extract, dried yeast extract, olive oil, cholesterol,ascorbic acid and vitamins A and D. He did not differentiate between active andnon-active constituents but found that vitamin E was not required for growth ofeither larva or adult. Rasso & Fraenkel (1954) have undertaken the most recentinvestigation. Using Phormia regina (Meig.) they report that eggs may be producedwith diets containing foods such as casein, casein enzymic hydrolysate, egg

778 PHYLLIS M. HARLOW
albumen, yeast extract, etc. Proteins must be in soluble form; dialysis reduced theiefficacy of sodium caseinate. Better ovarial growth with these diets was obtainedby the addition of a mixture of B vitamins and minerals. Choline, nicotinic acidand potassium phosphate were the most effective of the added substances. Syntheticdiets never gave such good development as liver. The amino-acid requirements forgrowth have been investigated in several other insects, e.g. Tribolium confusum(Lemonde & Bernhard, 1951), Blatta germanica (House, 1949), Drosophila melano-gaster (Schultz, St Lawrence & Newmeyer, 1946) and Apis melUfica (De Groot,1953); nothing, however, is known of the amino-acid requirement of theblowfly.
In the work here reported the normal development of the ovaries of the blowflyhas been studied, and a general investigation made of the food requirements forovarial growth in adults maintained on a diet of sucrose and water. Althoughsterile conditions were not used, the risk of bacterial contamination of food wa3decreased by a strict feeding procedure. It is believed that such bacterial con-tamination and the use of insufficiently controlled and, in some cases solid, foodsmay explain the conflicting accounts of former investigators.
II. MATERIALS AND METHODS
(a) Care of stock. Protophormia terrac-novae were used throughout. Adultswere reared under conditions of continuous light, relative humidity 70-80%, andtemperature 25° C. Each day, newly laid eggs were transferred to fresh meat in asmall plastic dish covered with muslin. The resultant larvae were fed daily withsmall amounts of whale-meat and were kept moist with wet cotton-wool wrapped infine muslin. On the sixth day the larvae and meat were transferred to Petri disheson sand: the migrating prepupae then moved to the clean dry sand below in whichthey pupated. In-breeding was not controlled and the flies emerging on one daycame from only two to four batches of eggs. 200-300 closely related flies wereobtained daily and kept in 7 lb. glass jars until required.
(b) Experimental procedure. Unless otherwise stated, newly emerged femaleflies were used for experiments. They were anaesthetized lightly with ether and thensexed. Random groups of ten females were marked individually with paint andweighed. Each group of ten was kept in a 7 lb. glass jar covered with Tigron(plastic muslin) and carpeted with filter-paper. All flies were supplied with solidsucrose in crucibles.
In each experiment at least three units of ten flies were used. These flies weretreated similarly except in the type of food given which was as follows:
(1) The sucrose control where flies were given a constant supply of water in5 ml. flasks with cotton-wool wicks. They were given no food other than the sucrose.
(2) The whale-meat control where flies were given a liquid preparation1 of whale-meat at specific times, and after the last feed were given water as in (1).
1 Measured quantities of whale-meat and water homogenized in a Waring Blendor and strainedthrough butter muslin. This is referred to in the text as 'whale-meat mixture'.

Ovanal development and relation to adult nutrition in blowfly 779
(3) The experimental jar where flies were given a different type of liquid foodwhich, except where stated, was of a similar protein concentration to the whale-meat control.
A large number of jars with different foods was generally included in this lastsection, each of which could be compared with another. The two control jarschecked the size and activity of the flies and the suitability of the experimentalconditions. Where it was necessary to use more than ten flies the jars wereduplicated.
For test feeding only liquid was given. It was spread from 1 ml. pipettes overthe surface of the muslin cover and protected from evaporation with an invertedPetri dish. Light above the jars attracted the flies to this surface and they fed easilyfrom the film of liquid and the hanging droplets. Spilt food dried quickly on thefilter-paper floor, and after 2 hr. the stale food was removed by exchanging themuslin cover. Stock food was refrigerated. In this way the effects of prolongedbacterial contamination of the food were avoided. Flies were given at least eight2 hr. feeding periods (meals) during the first 7 days. After the last meal flies wereagain anaesthetized and weighed On return to the jar they were given sucrose andwater continuously until dissection. The increased weight of the flies during thefeeding period showed that an adequate amount of food had been ingested by themajority in this time. No adverse after effect of ether was discovered.
All flies of one set were killed for dissection on the same day (this was alwaysbetween 8 and 14 days after emergence) and dissected under Ringer's solutionwithin an hour. The condition of the crop contents, ovaries and fat body of eachfly was recorded and the size of each ovary was measured. A micrometer eyepiecewas used to measure the dimensions of the ovaries which were spread out flat on aslide. Two diameters at right angles, and the length of the largest egg chamber, weremeasured. Owing to the regular compact shape of the ovary, the product of thesethree figures gave a value which, although having no absolute meaning, wasroughly proportional to the volume of the ovary. This value was used for findingthe mean ovarial size of each fly and each group of flies, thus providing a basis forstatistical work. Fat body was classed as pupal or adult, and the latter graded byeye into grades 1-4 according to size, 1 being the smallest.
(c) The effects of experimental conditions. The following were found to be criticaland were kept constant throughout each experiment: number of flies per jar; sizeof jar; light intensity; number of meals and the interval between them; concentra-tion of amino-nitrogen in the food (protein or amino-acid); solubility of food; thetime between last feeding and dissection.
Certain other factors were found to have no effect on ovarial development. Thesewere: the relative humidity, if above 5 5 % R . H . , the initial age of the flies atcommencement of feeding tests if previously maintained on sucrose and water,and if less than 2 weeks; the larval diet; the diet of the preceding six generations oflarvae and adults; the weight of the fly at emergence; the presence of males whichencouraged laying, but not growth, of the eggs.

780 PHYLLIS M. HARLOW
HI. RESULTS(a) Stages in ovarial development
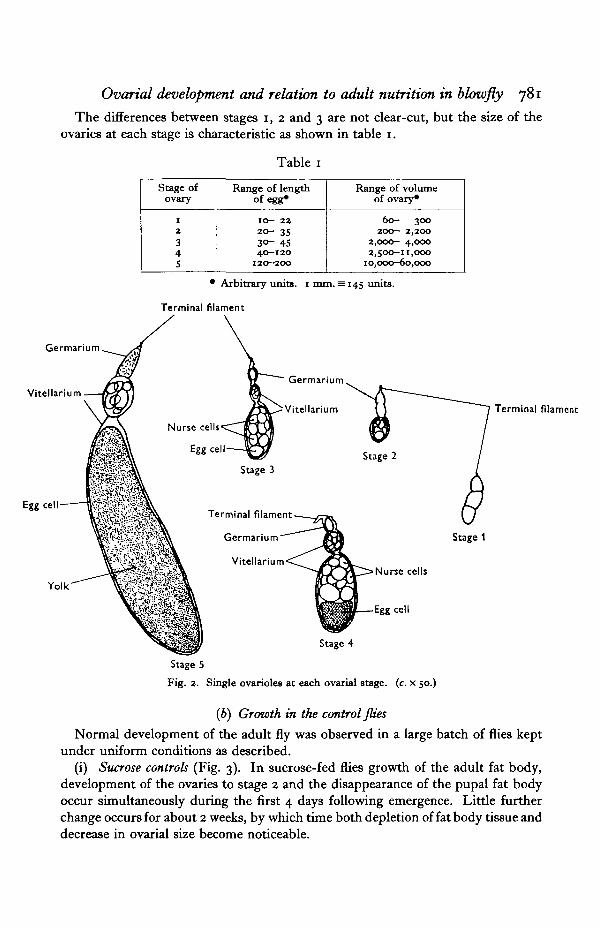
The ovaries in the blowfly are of the polytrophic type containing roughly 110-180ovarioles. Only one egg from each ovariole matures at one time, and all the eggsin one fly are at the same stage of development. Five stages of ovarial developmenthave been established and defined as follows (see also Figs. 1 and 2):
Stage 1. At emergence the ovary is small, white and opaque, the ovarioles adhereclosely, and the primary egg chamber is not clearly differentiated from thegermarium.
Stage 1
Egg cellwith yolk
Stage 3Stage 2
Stage 4Stage 5
Fig. 1. Whole ovaries to show stages in ovarial development ( x 15.)
Stage 2. The ovary is more translucent. Primary egg chambers are clearlydifferentiated from the germarium and nurse cells are visible at high magnification.
Stage 3. The primary egg chambers are enlarged, spherical and adhering lessclosely, giving a bubbly appearance to the ovary.
Stage 4. The primary egg chambers are elliptical, due to growth of both thenurse cells and the egg. The latter contains yolk and is often crescent shaped andgenerally larger than the nurse cells. Secondary egg chambers have reached stage 2.
Stage 5. The mature ovary is very large. Nurse cells have disappeared and thelong curved cylindrical egg fills the primary egg chamber and is fully yolked. Theyolk may be quite opaque (O) or less dense and partly translucent (P).
Ovarial development and relation to adult nutrition in blowfly 781
The differences between stages 1, 2 and 3 are not clear-cut, but the size of theovaries at each stage is characteristic as shown in table 1.
Table 1
Stage ofovary
12345
Range of lengthof egg*
10— 2220- 3530- 4540—120
120-200
Range of volumeof ovary*
60- 300200- 2,200
2,000- 4,0002,500-11,000
10,000-60,000
Germarium
Vltellarium
Egg cell
Yolk
• Arbitrary units, i mm. = 145 units.
Terminal filament
Germarium
Vitellarium
Stage 3
Terminal filament
Germarium
Vitellarium
Terminal filament
Stage 2
Stage 1
Nurse cells
Egg cell
Stage 4
Stage 5
Fig. 2. Single ovarioles at each ovarial stage, (c. x 50.)
(b) Growth in the control flies
Normal development of the adult fly was observed in a large batch of flies keptunder uniform conditions as described.
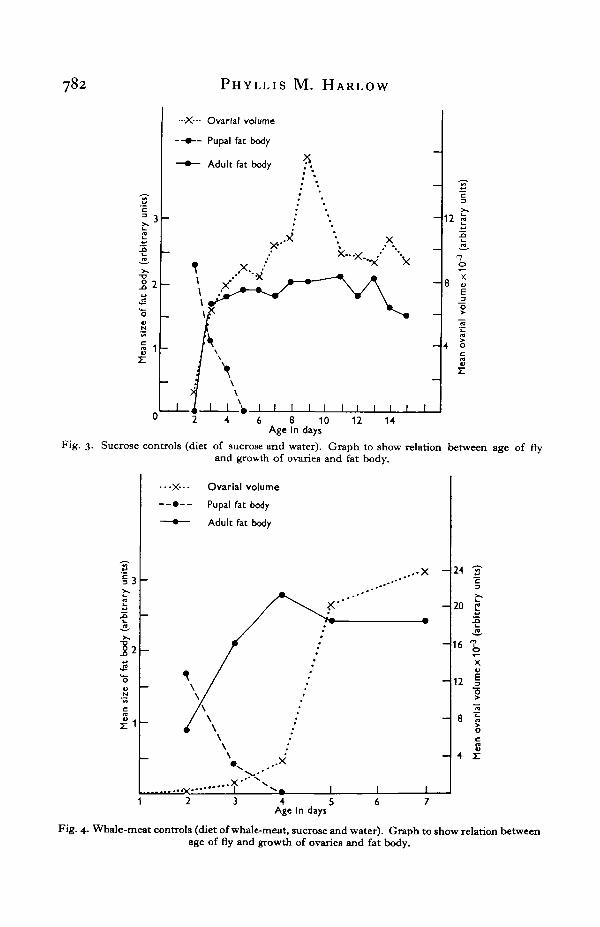
(i) Sucrose controls (Fig. 3). In sucrose-fed flies growth of the adult fat body,development of the ovaries to stage 2 and the disappearance of the pupal fat bodyoccur simultaneously during the first 4 days following emergence. Little furtherchange occurs for about 2 weeks, by which time both depletion of fat body tissue anddecrease in ovarial size become noticeable.
782 PHYLLIS M. HARLOW
•X-" Ovarial volume
• — Pupal fat body
• — Adult fat body
6 8 10Age In days
Fig. 3. Sucrose controls (diet of sucrose and water). Graph to show relation between age of flyand growth of ovaries and fat body.
- - •X- - - Ovar ia l vo lume
- - • - - Pupal fat body
— • — Adul t fat body
42"o
• X - 24
20 | *
16
12
Cz
1 2 3 4 5 6 7Age In days
Fig. 4, Whale-meat controls (diet of whale-meat, sucrose and water). Graph to show relation betweenage of fly and growth of ovaries and fat body.

Ovarial development and relation to adult nutrition in blowfly 783
(ii) Whale-meat controls (Fig. 4). In whale-meat fed flies, similar changes in fatbody and ovaries have taken place by the third day of adult life. Pupal fat bodypersists to the third day only, undoubtedly because the basal metabolic rate hasbeen much increased by ingestion of meat. Both ovaries and fat body then undergovery rapid growth and the complete ovarial maturation is accomplished within5 days. The striking increase in ovarial size accompanying yolk deposition takesplace in one day. The speed of yolk formation indicates that there is a large readilymobilized food reserve built up. This is probably greater than can be providedfrom either the haemolymph or direct absorption of food from the gut. Since thefat body is always large at the onset of yolk formation and has diminished signifi-cantly when yolk formation is complete, it is evident that a transfer of materialfrom the fat body to the ovary occurs during this time.
(c) The effects of food on ovarial development
(i) The type and amount of food taken
Normally adult flies can only ingest solid food in the form of very fine particles,although a few foods, e.g. sucrose, are dissolved by the sahvary fluid ejected duringfeeding. All foods employed in this work were used in the form of clear solution,fine suspension or homogenized mixture such as is produced by the WaringBlendor. Dissection disclosed the presence of food in the crop immediately afterfeeding and up to 7 days later. The presence of coloured food in the crop at autopsyindicated adequate food intake, whilst the presence of congealed meat juice ormilk a week after ingestion showed that extensive digestion of protein did notoccur in the crop which appears to act chiefly as a store for both food and water.Since ingestion of much water may retard digestion by diluting the food, flies werenot supplied with water until experimental feeding was complete.
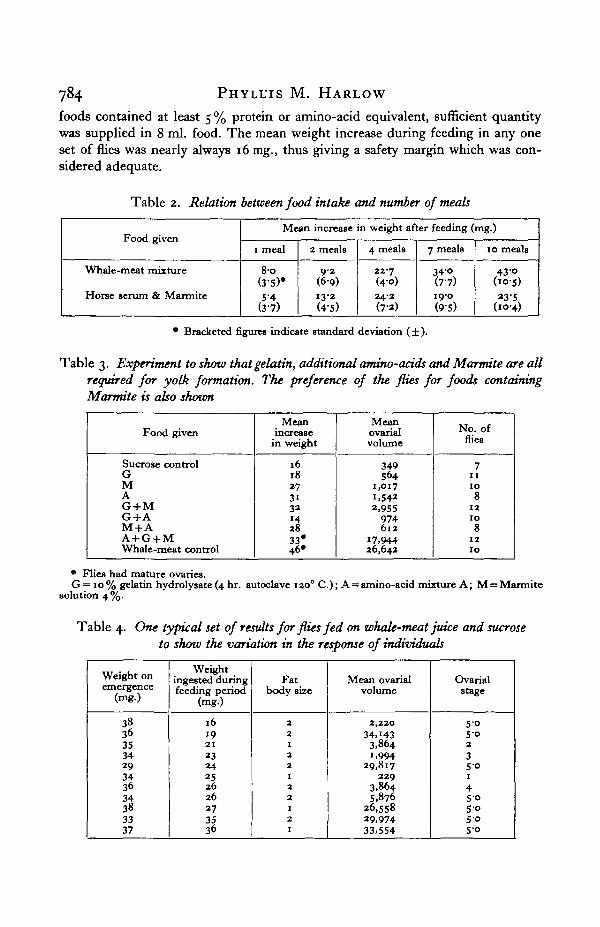
Each fly was weighed before and after the complete feeding period. The differencein weight was used to express its food intake and results showed (Table 2) that thisamount increased with the number of meals. Taste was also important. Table 3shows that Marmite-containing foods were preferred to others, although sufficientamounts of even less acceptable foods were taken. Minor inaccuracies in thisfigure are caused by regurgitation, defaecation, water loss, metabolic activity, andalso variation due to unequal growth of the ovaries and fat body. The variation infood intake and ovarial growth of individual flies, all of which had ingested con-siderable amounts of whale-meat juice, is shown in Table 4. In these cases ovarialmaturation could not be related to food intake since all the flies had taken in suf-ficient food; where no ovarial maturation occurred the cause of non-developmentmust be inherent in the fly.
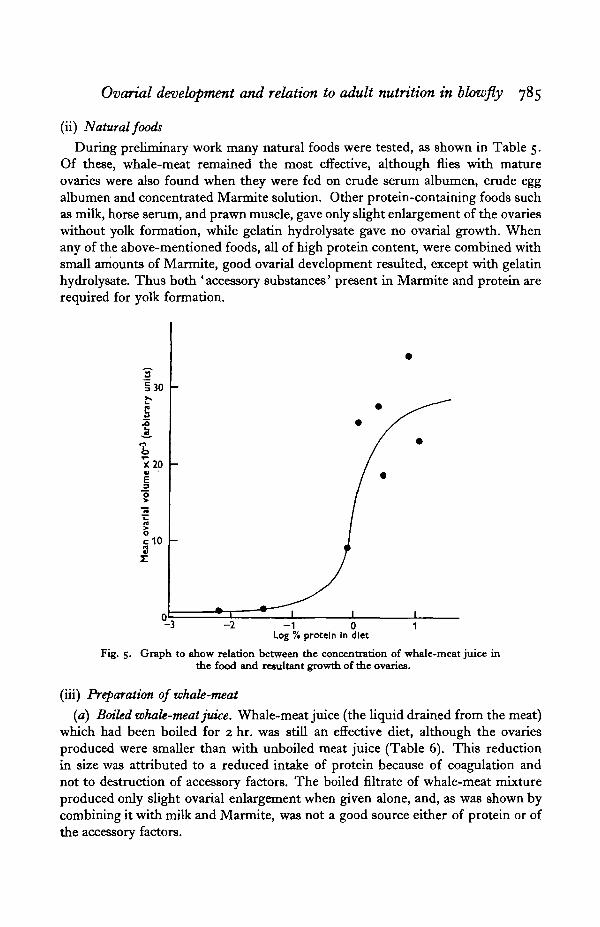
The usual range of weight increase per fly during feeding in any one batch offlies was 16-40 mg. By feeding successive dilutions of a whale-meat juice it wasshown that a solution containing 1 % protein was the lowest to give satisfactoryovarial growth under standard conditions (Fig. 5). One fly therefore requires notmore than 0-4 mg. protein for ovarial maturation. Since most experimental
7 8 4 PHYLLIS M. HARLOW
foods contained at least 5 % protein or amino-acid equivalent, sufficient quantitywas supplied in 8 ml. food. The mean weight increase during feeding in any oneset of flies was nearly always 16 mg., thus giving a safety margin which was con-sidered adequate.
Table 2. Relation between food intake and number of meals
Food given
Whale-meat mixture
Horse serum & Marmite
Mean increase in weight after feeding (mg.)
1 meal
80(3-S)*
< &
2 meals
92(6-9)I3'2
(4-5)
4 meals
22-7(40)24-2(7-2)
7 meals
34O(7-7)19-0(9-S)
10 meals
43-o(io-5)a3-S
(IO-4)
• Bracketed figures indicate standard deviation ( ± ).
Table 3. Experiment to show that gelatin, additional amino-acids and Marmite are all
required for yolk formation. The preference of the flies for foods containing
Marmite is also shown
Food given
Sucrose controlGMAG + MG + AM + AA + G + MWhale-meat control
Meanincrease
in weight
1618273 i3214283 3 *4 6 *
Meanovarialvolume
349S64
1,017
i,5422,955
974612
17,94426,643
No. offlies
71 1
1 0
81 2
1 0
81 2
1 0
• Flies had mature ovaries.G = 10% gelatin hydrolysate (4 hr. autoclave 1200 C) ; A = amino-acid mixture A; M = Marmite
solution 4 %.
Table 4. One typical set of results for flies fed on whale-meat juice and sucrose
to show the variation in the response of individuals
Weight onemergence
(mg.)
3836353429343634383337
Weightingested duringfeeding period
(mg.)
16192 1
2324252626273536
Fatbody size
221
22
1
221
21
Mean ovarialvolume
2,220
34,1433,8641,994
29,817229
3,8645,876
26,55829,97433,554
Ovarialstage
5-0SO2
35 O1
4SOS o5 05-o
Ovarial development and relation to adult nutrition in blowfly 785
(ii) Natural foods
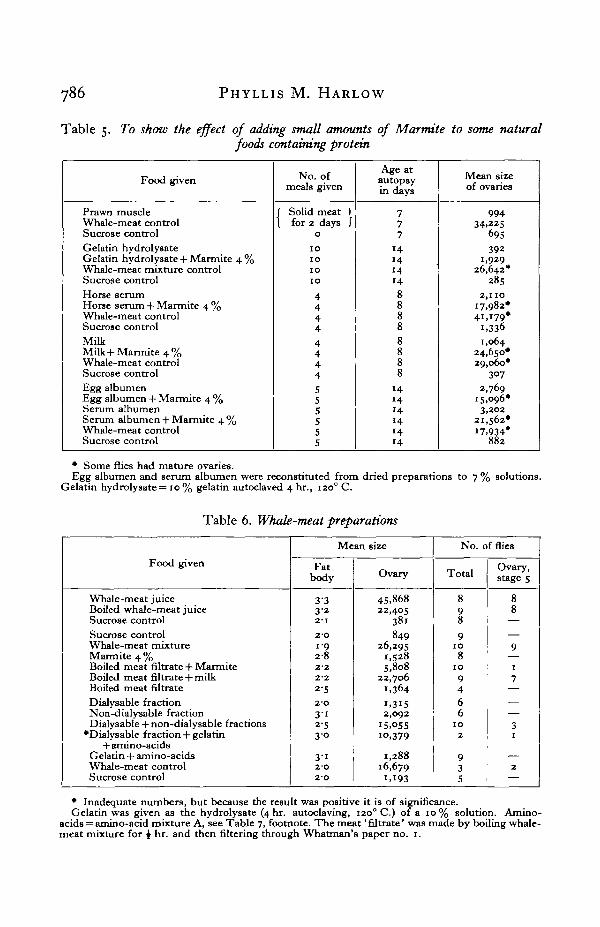
During preliminary work many natural foods were tested, as shown in Table 5.Of these, whale-meat remained the most effective, although flies with matureovaries were also found when they were fed on crude serum albumen, crude eggalbumen and concentrated Marmite solution. Other protein-containing foods suchas milk, horse serum, and prawn muscle, gave only slight enlargement of the ovarieswithout yolk formation, while gelatin hydrolysate gave no ovarial growth. Whenany of the above-mentioned foods, all of high protein content, were combined withsmall amounts of Marmite, good ovarial development resulted, except with gelatinhydrolysate. Thus both ' accessory substances' present in Marmite and protein arerequired for yolk formation.
3§30
ja
X20
oc10a
-3 -2 - 1 0Log % protein In diet
Fig-S- Graph to show relation between the concentration of whale-meat juice inthe food and resultant growth of the ovaries.
(iii) Preparation of whale-meat
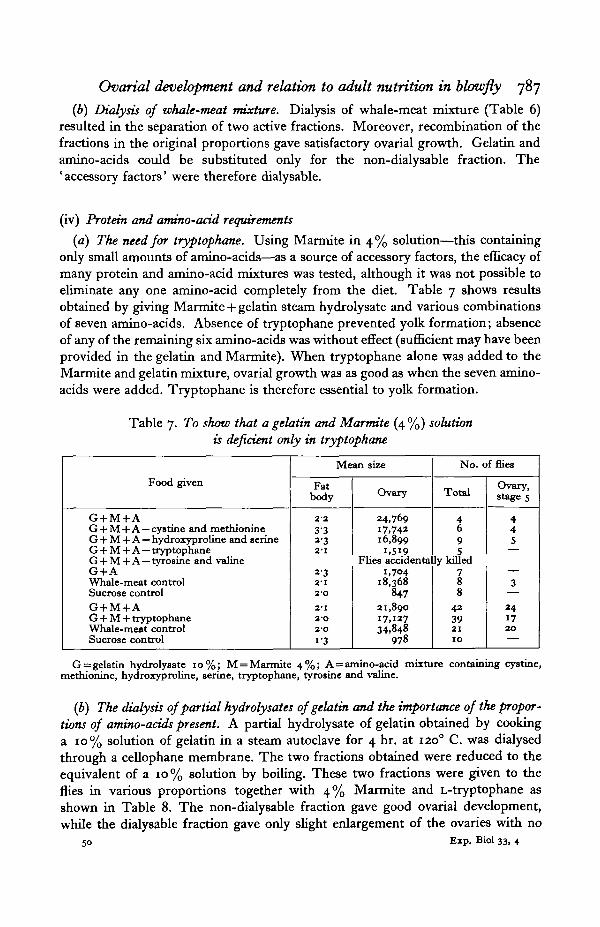
(a) Boiled whale-meat juice. Whale-meat juice (the liquid drained from the meat)which had been boiled for 2 hr. was still an effective diet, although the ovariesproduced were smaller than with unboiled meat juice (Table 6). This reductionin size was attributed to a reduced intake of protein because of coagulation andnot to destruction of accessory factors. The boiled filtrate of whale-meat mixtureproduced only slight ovarial enlargement when given alone, and, as was shown bycombining it with milk and Marmite, was not a good source either of protein or ofthe accessory factors.
786 PHYLLIS M. HARLOW
Table 5. To show the effect of adding small amounts of Marmite to some naturalfoods containing protein
Food given
Prawn muscleWhale-meat controlSucrose controlGelatin hydrolysateGelatin hydrolysate + Marmite 4 %Whale-meat mixture controlSucrose controlHorse serumHorse serum + Marmite 4 %Whale-meat controlSucrose controlMilkMilk + Marmite 4 %Whale-meat controlSucrose controlEgg albumenEgg albumen 4- Marmite 4 %Serum albumenSerum albumen + Marmite 4%Whale-meat controlSucrose control
No. ofmeals given
( Solid meat I( for 2 days I
0
1 01 01 01 0
4444
4444
555555
Age atautopsyin days
777
1414141488888888
141414141414
Mean sizeof ovaries
99434,225
695392
1,92926,642*
285
2,11017,982*4 i , i79 #
1,3361,064
24,650*29,060*
3072,769
15,096*3,202
21,562*17,934*
882
* Some flies had mature ovaries.Egg albumen and serum albumen were reconstituted from dried preparations to 7 % solutions.
Gelatin hydrolysate = 10% gelatin autoclaved 4 hr., 1200 C.
Table 6. Whale-meat preparations
Food given
Whale-meat juiceBoiled whale-meat juiceSucrose controlSucrose controlWhale-meat mixtureMarmite 4%Boiled meat filtrate + MarmiteBoiled meat filtrate + milkBoiled meat filtrate
Dialysable fractionNon-dialysable fractionDialysable + non-dialysable fractions
•Dialysable fraction + gelatin+ amino-acids
Gelatin + amino-acidsWhale-meat controlSucrose control
Mean size
Fatbody
333-22-12O1928222-22-52-O312-53O
31202O
Ovary
45,86822,405
381849
26,2951,5285,808
22,7061,3641,3152,092
I5,O5510,379
1,28816,679
1,193
No. of flies
Total
898
9108
10946
102
935
Ovary,stage 5
8QO
9
1
7
31
• Inadequate numbers, but because the result was positive it is of significance.Gelatin was given as the hydrolysate (4 hr. autoclaving, 1200 C.) of a 10% solution. Amino-
acids = amino-acid mixture A, see Table 7, footnote. The meat' filtrate' was made by boiling whale-meat mixture for J hr. and then filtering through Whatman's paper no. 1.
Ovarial development and relation to adult nutrition in blowfly 787
(b) Dialysis of whale-meat mixture. Dialysis of whale-meat mixture (Table 6)resulted in the separation of two active fractions. Moreover, recombination of thefractions in the original proportions gave satisfactory ovarial growth. Gelatin andamino-acids could be substituted only for the non-dialysable fraction. The' accessory factors' were therefore dialysable.
(iv) Protein and amino-acid requirements
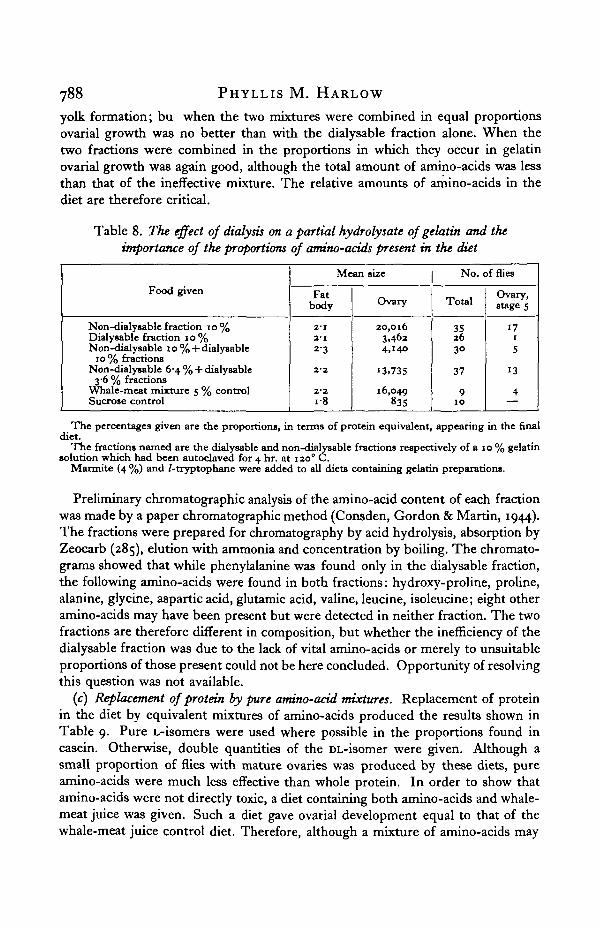
(a) The need for tryptophane. Using Marmite in 4% solution—this containingonly small amounts of amino-acids—as a source of accessory factors, the efficacy ofmany protein and amino-acid mixtures was tested, although it was not possible toeliminate any one amino-acid completely from the diet. Table 7 shows resultsobtained by giving Marmite + gelatin steam hydrolysate and various combinationsof seven amino-acids. Absence of tryptophane prevented yolk formation; absenceof any of the remaining six amino-acids was without effect (sufficient may have beenprovided in the gelatin and Marmite). When tryptophane alone was added to theMarmite and gelatin mixture, ovarial growth was as good as when the seven amino-acids were added. Tryptophane is therefore essential to yolk formation.
Table 7. To show that a gelatin and Marmite (4%) solutionis deficient only in tryptophane
Food given
G + M+AG + M + A — cystine and methionineG + M + A — hydroxyproline and serineG + M+A — tryptophaneG + M + A — tvrosine and valineG + AWhale-meat controlSucrose controlG + M + AG + M + tryptophaneWhale-meat controlSucrose control
Mean size
Fatbody
2'23"32-32-1
232" I2-0
2-12-O2'Oi-3
Ovary
24,76917,74^16,899i,5i9
Flies accidenta1,704
18,368847
21,89017,12734,848
978
No. of flies
Total
69
Jy killed
88
42392110
Ovary,stage s
445
3
241720
G = gelatin hydrolysate 10%; M = Marmite 4 % ; A = amino-acid mixture containing cystine,methionine, hydroxyproline, serine, tryptophane, tyrosine and valine.
(b) The dialysis of partial hydrolysates of gelatin and the importance of the propor-tions of amino-acids present. A partial hydrolysate of gelatin obtained by cookinga 10% solution of gelatin in a steam autoclave for 4 hr. at 1200 C. was dialysedthrough a cellophane membrane. The two fractions obtained were reduced to theequivalent of a 10% solution by boiling. These two fractions were given to theflies in various proportions together with 4% Marmite and L-tryptophane asshown in Table 8. The non-dialysable fraction gave good ovarial development,while the dialysable fraction gave only slight enlargement of the ovaries with no
50 Exp. Biol 33, 4
788 PHYLLIS M. HARLOW
yolk formation; bu when the two mixtures were combined in equal proportionsovarial growth was no better than with the dialysable fraction alone. When thetwo fractions were combined in the proportions in which they occur in gelatinovarial growth was again good, although the total amount of amino-acids was lessthan that of the ineffective mixture. The relative amounts of amino-acids in thediet are therefore critical.
Table 8. The effect of dialysis on a partial hydrolysate of gelatin and theimportance of the proportions of amino-acids present in the diet
Food given
Non-dialysable fraction 10%Dialysable fraction 10%Non-dialysable io% +dialysable
10% fractionsNon-dialysable 6-4% + dialysable
3'6% fractionsWhale-meat mixture 5 % controlSucrose control
Mean size
Fatbody
2-12'J
2'2
2'21-8
Ovary
20,0163.4024,140
13.735
16,049835
No. of flies
Total
3526
3°
379
10
Ovary,stage 5
171
5
13
4
The percentages given are the proportions, in terms of protein equivalent, appearing in the finaldiet.
The fractions named are the dialysable and non-dialysable fractions respectively of a 10 % gelatinsolution which had been autoclaved for 4 hr. at 120° C.
Marmite (4 %) and /-tryptophane were added to all diets containing gelatin preparations.
Preliminary chromatographic analysis of the amino-acid content of each fractionwas made by a paper chromatographic method (Consden, Gordon & Martin, 1944).The fractions were prepared for chromatography by acid hydrolysis, absorption byZeocarb (285), elution with ammonia and concentration by boiling. The chromato-grams showed that while phenylalanine was found only in the dialysable fraction,the following amino-acids were found in both fractions: hydroxy-proline, proline,alanine, glycine, aspartic acid, glutamic acid, valine, leucine, isoleucine; eight otheramino-acids may have been present but were detected in neither fraction. The twofractions are therefore different in composition, but whether the inefficiency of thedialysable fraction was due to the lack of vital amino-acids or merely to unsuitableproportions of those present could not be here concluded. Opportunity of resolvingthis question was not available.
(c) Replacement of protein by pure amino-acid mixtures. Replacement of proteinin the diet by equivalent mixtures of amino-acids produced the results shown inTable 9. Pure L-isomers were used where possible in the proportions found incasein. Otherwise, double quantities of the DL-isomer were given. Although asmall proportion of flies with mature ovaries was produced by these diets, pureamino-acids were much less effective than whole protein. In order to show thatamino-acids were not directly toxic, a diet containing both amino-acids and whale-meat juice was given. Such a diet gave ovarial development equal to that of thewhale-meat juice control diet. Therefore, although a mixture of amino-acids may

Ovarial development and relation to adult nutrition in blowfly 789
be more difficult to metabolize efficiently, its presence in a diet does not affect thenormal metabolic changes. (Considerable difficulty accompanied the feeding of ahighly concentrated amino-acid mixture because of the insolubility of some con-stituents. Treatment with the Waring Blendor provided the most suitable diet,insoluble material being reduced to a suspension which could be ingested by theflies.) Use of a mixture containing DL-serine and L-isomers of the remainingamino-acids did not result in better development than that obtained with mixturesused for most of the above tests. The latter contained about 4 DL-isomers inaddition to DL-serine. Therefore, unless DL-serine was strongly inhibitive (therewas no evidence to favour this) the presence of small amounts of the unnaturalisomer in the diet did not interfere with growth. When, however, a mixture con-taining mainly DL-isomers was tested and compensating double quantities were notgiven, there was no yolk formation in any fly. Since the effective concentrationsof such amino-acids in this particular mixture had been reduced by half this suggeststhat, as with insects other than Protophormia, many D-isomers cannot be metabolized.(L-isomers of all amino-acids have only since become commercially available.)
Table 9. The result of replacing protein in the dietby equivalent mixtures of amino-acids
Food given
Amino-acids 5 % (B) + Mannite 4%Amino-acids 6 % (C) + Marmite 4 %Whale-meat mixtureSucrose controlAmino-acids 5 % (G) + Marmite 4 %Whale-meat mixture 5 %Amino-acids 5 % (G) + whale-meatmixture 5%
Amino-acids 2-5 % (G) + whale-meatmixture 2-5 %
Sucrose controlL-Amino-acids (E)L-Amino-acids (E) + Marmite 4 %DL-Amino-acids (F) + Marmite 4 %Whale-meat mixtureSucrose control
Mean size
Fatbody
24232319232-1
2-O
I'8i-5i-62 3192-oi-5
Ovary
19,30912,26424.460
1,05212,96333,O5528,201
30,865
9681,1468,0181.833
39,125923
No. of flies
Total
28362117363434
38
1915302568
Ovary,stage 5
51214
123123
34
4
6
Bracketed letters refer to the amino-acid mixture used. See Appendix, Table 1.
(d) Combination of protein and amino-acid mixtures. It has already been shownthat protein hydrolysates and pure amino-acid mixtures can only partially replaceprotein in the diet.
Although no record has been found where the existence of a non-specificsubstance present in proteins and not in amino-acid mixtures is proved, thefollowing experiments were made to show whether the presence of small amountsof protein or other undialysable breakdown products of protein significantlyimproved ovarial growth in flies given pure amino-acids. In (a) a small proportion
JO-2
79° PHYLLIS M. HARLOW
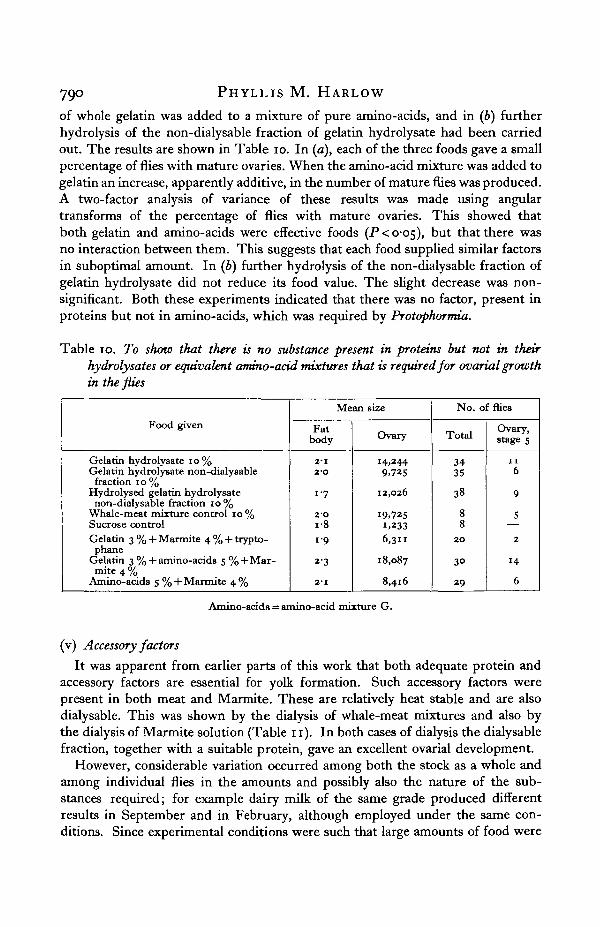
of whole gelatin was added to a mixture of pure amino-acids, and in (b) furtherhydrolysis of the non-dialysable fraction of gelatin hydrolysate had been carriedout. The results are shown in Table 10. In (a), each of the three foods gave a smallpercentage of flies with mature ovaries. When the amino-acid mixture was added togelatin an increase, apparently additive, in the number of mature flies was produced.A two-factor analysis of variance of these results was made using angulartransforms of the percentage of flies with mature ovaries. This showed thatboth gelatin and amino-acids were effective foods (P<o-O5), but that there wasno interaction between them. This suggests that each food supplied similar factorsin suboptimal amount. In (b) further hydrolysis of the non-dialysable fraction ofgelatin hydrolysate did not reduce its food value. The slight decrease was non-significant. Both these experiments indicated that there was no factor, present inproteins but not in amino-acids, which was required by Protophormia.
Table 10. To show that there is no substance present in proteins but not in theirhydrolysates or equivalent amino-acid mixtures that is required for ovarial growthin the flies
Food given
Gelatin hydrolysate 10%Gelatin hydrolysate non-dialysablefraction 10%
Hydrolysed gelatin hydrolysatenon-dialysable fraction 10 %
Whale-meat mixture control 10%Sucrose controlGelatin 3 % + Marmite 4 % + trypto-phane
Gelatin 3 % +amino-acids 5 % +Mar-mite 4 %
Amino-acids S % + Marmite 4 %
Mean size
Fatbody
2-12-O
1-7
2-O1819
23
2-1
Ovary
14.2449,725
12,026
19.7251,2336,311
18,087
8,416
No. of flies
Total
3435
38
88
20
3°
29
Ovary,stage s
116
9
5
2
14
6
Amino-acids = amino-acid mixture G.
(v) Accessory factors
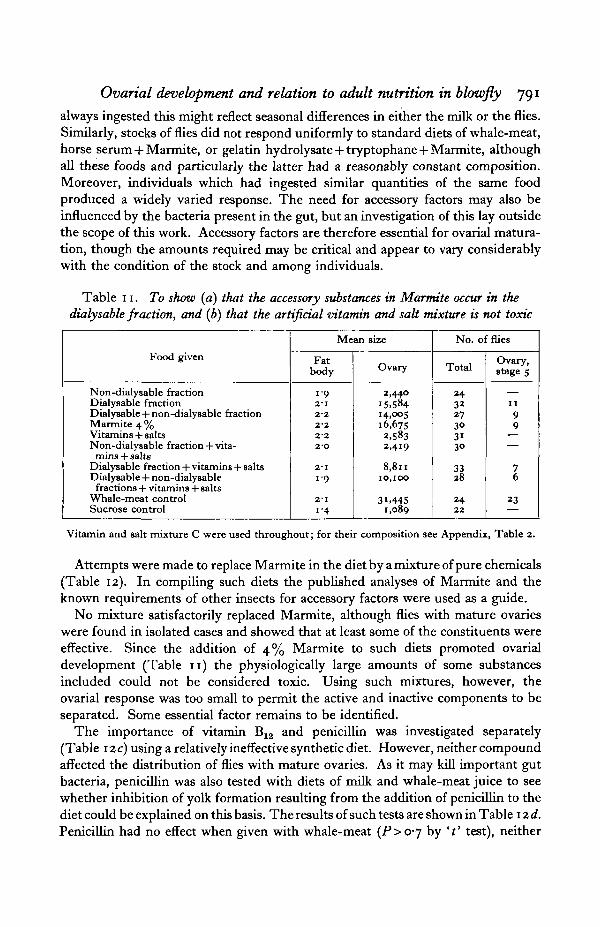
It was apparent from earlier parts of this work that both adequate protein andaccessory factors are essential for yolk formation. Such accessory factors werepresent in both meat and Marmite. These are relatively heat stable and are alsodialysable. This was shown by the dialysis of whale-meat mixtures and also bythe dialysis of Marmite solution (Table 11). In both cases of dialysis the dialysablefraction, together with a suitable protein, gave an excellent ovarial development.
However, considerable variation occurred among both the stock as a whole andamong individual flies in the amounts and possibly also the nature of the sub-stances required; for example dairy milk of the same grade produced differentresults in September and in February, although employed under the same con-ditions. Since experimental conditions were such that large amounts of food were
Ovarial development and relation to adult nutrition in blowfly 791
always ingested this might reflect seasonal differences in either the milk or the flies.Similarly, stocks of flies did not respond uniformly to standard diets of whale-meat,horse serum + Marmite, or gelatin hydrolysate + tryptophane + Marmite, althoughall these foods and particularly the latter had a reasonably constant composition.Moreover, individuals which had ingested similar quantities of the same foodproduced a widely varied response. The need for accessory factors may also beinfluenced by the bacteria present in the gut, but an investigation of this lay outsidethe scope of this work. Accessory factors are therefore essential for ovarial matura-tion, though the amounts required may be critical and appear to vary considerablywith the condition of the stock and among individuals.
Table 11. To show (a) that the accessory substances in Marmite occur in thedialysable fraction, and (b) that the artificial vitamin and salt mixture is not toxic
Food given
Non-dialysable fractionDialysable fractionDialysable +non-dialysable fractionMarmite 4 %Vitamins + saltsNon-dialysable fraction + vita-mins + salts
Dialysable fraction + vitamins + saltsDialysable + non-dialysablefractions + vitamins + salts
Whale-meat controlSucrose control
Mean size
Fatbody
1-92-1222'2222O
2-11 9
211-4
Ovary
2,44015.58414.00516,6752,5832,419
8,81110,100
31.4451,089
No. of flies
Total
243227303i30
3328
2422
Ovary,stage 5
1199
76
23
Vitamin and salt mixture C were used throughout; for their composition see Appendix, Table 2.
Attempts were made to replace Marmite in the diet by a mixture of pure chemicals(Table 12). In compiling such diets the published analyses of Marmite and theknown requirements of other insects for accessory factors were used as a guide.
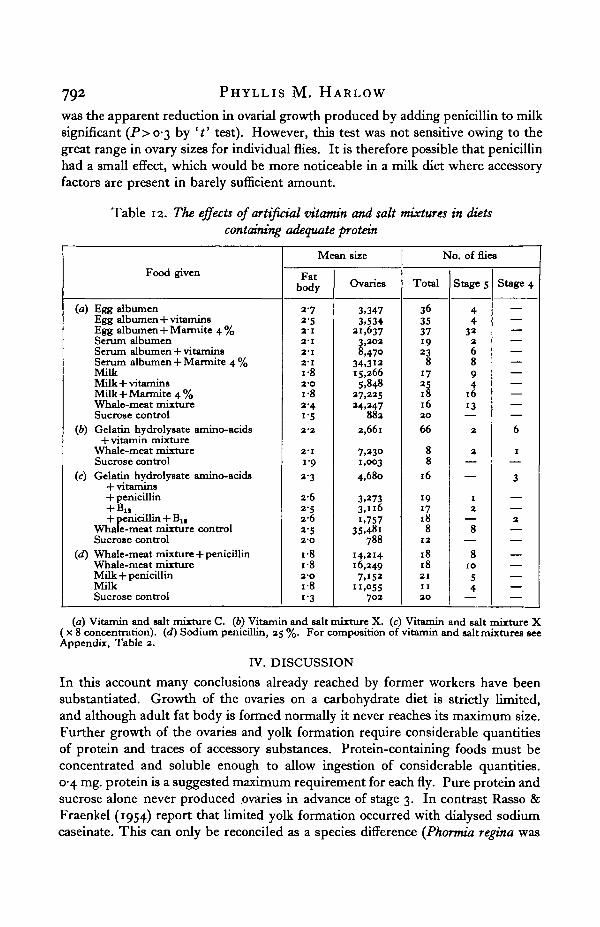
No mixture satisfactorily replaced Marmite, although flies with mature ovarieswere found in isolated cases and showed that at least some of the constituents wereeffective. Since the addition of 4% Marmite to such diets promoted ovarialdevelopment (Table 11) the physiologically large amounts of some substancesincluded could not be considered toxic. Using such mixtures, however, theovarial response was too small to permit the active and inactive components to beseparated. Some essential factor remains to be identified.
The importance of vitamin B12 and penicillin was investigated separately(Table 12 c) using a relatively ineffective synthetic diet. However, neither compoundaffected the distribution of flies with mature ovaries. As it may kill important gutbacteria, penicillin was also tested with diets of milk and whale-meat juice to seewhether inhibition of yolk formation resulting from the addition of penicillin to thediet could be explained on this basis. The results of such tests are shown in Table 12 d.Penicillin had no effect when given with whale-meat (P > 07 by ' t' test), neither
792 PHYLLIS M. HARLOW
was the apparent reduction in ovarial growth produced by adding penicillin to milksignificant (P>o>3 by 't' test). However, this test was not sensitive owing to thegreat range in ovary sizes for individual flies. It is therefore possible that penicillinhad a small effect, which would be more noticeable in a milk diet where accessoryfactors are present in barely sufficient amount.
Table 12. The effects of artificial vitamin and salt mixtures in dietscontaining adequate protein
Food given
(a) Egg albumenEgg albumen + vitaminsEgg albumen + Marmite 4 %Serum albumenSerum albumen + vitaminsSerum albumen + Marmite 4 %MilkMilk + vitaminsMilk + Marmite 4%Whale-meat mixtureSucrose control
(b) Gelatin hydrolysate amino-acids+ vitamin mixture
Whale-meat mixtureSucrose control
(c) Gelatin hydrolysate amino-acids+ vitamins+ penicillin+ Bj,+ penicillin + Blt
Whale-meat mixture controlSucrose control
(d) Whale-meat mixture + penicillinWhale-meat mixtureMilk + penicillinMilkSucrose control
Mean size
Fatbody
2-7252-12-12-12-11-82-Oi-82-4i-5
2-2
2'I
19232-625262-52-0i-8i-82'Oi-813
Ovaries
3,3473,534
21,6373,2028,470
34,31215,2665,848
27,22524,247
882
2,661
7,2301,0034,680
3,2733,"6i,757
35,48i788
14,21416,2497,152
",055702
No. of flies
Total
36353719238
1725
162066
88
16
1917188
12
1818211120
Stage 5
44
322689
:o13
2
2
12
8
81054
Stage 4
6
1
3
2
(a) Vitamin and salt mixture C. (b) Vitamin and salt mixture X. (c) Vitamin and salt mixture X( x 8 concentration), {d) Sodium penicillin, 25 %. For composition of vitamin and salt mixtures seeAppendix, Table 2.
IV. DISCUSSION
In this account many conclusions already reached by former workers have beensubstantiated. Growth of the ovaries on a carbohydrate diet is strictly limited,and although adult fat body is formed normally it never reaches its maximum size.Further growth of the ovaries and yolk formation require considerable quantitiesof protein and traces of accessory substances. Protein-containing foods must beconcentrated and soluble enough to allow ingestion of considerable quantities.0-4 mg. protein is a suggested maximum requirement for each fly. Pure protein andsucrose alone never produced .ovaries in advance of stage 3. In contrast Rasso &Fraenkel (1954) report that limited yolk formation occurred with diah/sed sodiumcaseinate. This can only be reconciled as a species difference (Phormia regina was

Ovarial development and relation to adult nutrition in blowfly 793
used) or, as the former workers have suggested, the precautions against bacterialcontamination of the food were not adequate.
Differentiation of the primary egg chambers and initial enlargement of theovaries occur in sucrose-fed flies in the first 4 days of adult life, when the pupal fatbody is disappearing. These processes have been correlated with the activities ofthe side lobes of the ring gland which persists until the fourth day of adult life(Thomsen, 1952). Since little further growth of either the ovaries or fat bodyoccurs when a protein diet is withheld this early growth must depend on foodreserves formed during larval stages and preserved in the pupal fat body. Pupalfat body, however, disappears more rapidly when a complete diet is given. This maybe correlated with an increased metabolic rate since the corpus allatum, which hasbeen shown by Thomsen (1949) to influence metabolic rate, also increases in sizeduring this time. Moreover, the adult fat body reaches a maximum size duringyolk formation and is smaller in the mature fly. The simultaneous decrease in sizeof fat body and enormous increase in ovarial size which occurs during yolk formationindicate that a transfer of food reserves from the fat body to ovary takes place.Since the gonadotrophic function of the corpus allatum hormone has beenestablished it would be interesting to discover if its effect is to control this transferof food material, as in the grasshopper Melanophis differentiaUs (Pfeiffer, 1945).
The investigation of specific amino-acid requirements has been hindered by thedifficulty in procuring a diet from which individual amino-acids could be eliminated,but it has been possible to show that tryptophane is essential. Pure protein couldnot be entirely replaced by equivalent amino-acid mixtures, ovarial growth beingreduced by at least half. This reduction was not due to toxicity or to the presenceof D-isomers. Growth was, however, greatly restricted when a mixture containingpredominantly DL-isomers of amino-acids was used. This suggests that, as withother insects, D-isomers are not always utilizable. It was found that the relativeamounts of amino-acids in the diet are very critical when unnatural mixtures areused. Unsuitable proportions may have been the reason for the inadequacy of themixtures of pure amino-acids used here.
There was no evidence of the requirement of a specific factor present in proteinsbut not in hydrolysates of proteins or pure amino-acid mixtures. Addition ofamino-acid mixtures to dilute protein diets gave an additive but not synergisticeffect. The importance of amino-acid balance already noted was emphasized whenthe proportions of dialysable to non-dialysable fractions of gelatin hydrolysatewere varied, and it was found that presence of a higher proportion of the inadequatedialysable fraction reduced ovarial growth although no reduction in the concentra-tion of the non-dialysable fraction had been made. This is comparable to the'relative lysine deficiency' found by Fraenkel (1948) in TriboHum tnfestans and the'relative tryptophane deficiency' found by Maddy & Elvehjem in rats (1949).Resolution of this problem in the blowfly will hot be possible until an accurateanalysis of the amino-acid content of the two fractions has been made.
Accessory factors were less satisfactorily investigated owing to the failure toproduce a chemically defined diet. Some essential factor or factors was/were still

794 PHYLLIS M. HARLOW
missing from the vitamin and salt mixtures tested, although almost every substanceknown to be important for growth of other insects had been included. It wouldseem that a new factor is involved. Sodium penicillin had no marked effect on theefficacy of any diet and vitamin B12 did not improve growth on a synthetic diet.The complete vitamin and salt mixture did, however, contain some active con-stituents, but in the absence of essential substances their individual effects were toosmall to make their investigation practicable.
In contrast to Evans's (1936) results with Lucilia, ovarial development at55 % R.H. under optimum feeding conditions was as good as at 95 % R.H., althoughflies were very much more active at the lower humidities. It was also surprising tofind no correlation between the weight of a fly at emergence and its ovarial size atany time. However, ovarial size at emergence may be governed by genetic factorsand not by the larval conditions which are very important in determining bodyweight.
There were considerable variations both in the behaviour of the stock at differenttimes and in the individual flies which had been given the same amount of food.In some an apparent genetic abnormality resulted in the failure of the ovaries togrow beyond stage 1. In one winter stock a tendency to an accumulation of fatbody with failure of ovarial growth on a complete diet was noted. This could beinterpreted as an adult diapause. Such flies responded to much higher concentra-tions of Marmite or whale-meat juice than those normally needed, and apparentlyrequired unusually large amounts of the accessory factors for ovarial development.
V. SUMMARY1. A method has been developed for obtaining 200—300 flies daily from a culture
of Protophormia terrae-novae reared on whale-meat.2. Normal growth changes in adults have been described, and five stages in
ovarial development have been denned.3. Experimental conditions affecting ovarial growth in adults have been
investigated and a satisfactory routine method for examining the effect of adultnutrition on ovarial growth has been devised. Precautions were taken againstbacterial contamination of food although non-sterile conditions were used.
4. Ovarial development was not related to relative humidity, if above 55 % R.H.,nor to the weight of the adults at emergence.
5. Evidence has been given for the conversion of food material from pupal toadult fat body and for the transfer of material from fat body to ovaries at yolkformation.
6. Healthy adult females can be maintained on a diet of sucrose and water.Addition to the diet of protein or amino-acids produces only slight enlargement ofthe ovaries. Yolk formation only occurs if the diet is further supplemented byaccessory factors, or substances required only in small amount. Ovarial maturationrequires considerable amounts of protein, though not more than 0-4 mg. per flyis needed and food solutions must be concentrated. The accessory factors have not

Ovarial development and relation to adult nutrition in blowfly 795
been identified but are known to be resistant to slow boiling and are dialysable.Certain mineral salts and B vitamins are thought to be involved.
7. Tryptophane is an essential part of the amino-acid mixture.8. Protein hydrolysates or equivalent amino-acid mixtures can be substituted
for the whole protein but are less effective. The proportions of amino-acids presentare very critical, particularly when the amino-acids are in a free state. The amino-acid mixtures have no toxic effect and the presence of some D-isomers do not inhibitgrowth, although some D-isomers cannot be used by the flies for growth.
9. A chemically defined diet similar to that used for Drosophila did not promoteovarial growth in Protophormia. A mixture containing ten B vitamins, ribosenucleic acid and mineral salts and cholesterol contained some effective accessorysubstances but lacked some unidentified factor required for yolk formation. Themissing factor was not vitamin B12. Penicillin had no significant effect on ovarialgrowth whether used in minute or in antibiotic concentration.
I wish to thank those of the Zoology Department, Imperial College of Scienceand Technology, London, who have given valuable advice and criticism duringthis work, notably Prof. O. W. Richards, my supervisor, Dr N. Waloff, Mr R. G.Davies and Dr. D. R. P. Murray. Grateful thanks are also due to Dr. R. E. Blackithfor his advice on statistics and Dr J. Russell for his interest in the preparation offoods. I am very grateful to the Agricultural Research Council who supportedthe work with a grant, and to Prof. J. W. Munro for the facilities he allowed me inhis department. Pupae were kindly supplied by Mr McKenny-Hughes, London,Dr A. A. Greene, Slough, and Dr L. Davies, Durham, and samples of sodiumpenicillin and vitamin B12 by Dr W. F. J. Cuthbertson of Messrs Glaxo Ltd.
REFERENCESBANKI, L. (1937). Calliphora erythrocephala. The importance of vitamin E in larval and adult
nutrition. AUart Kirzlem Budapest, 34, 21.CONSDEN, R., GORDON, A. M. & MARTIN, A. J. P. (1944). Qualitative analysis of proteins; a partition
method using paper. Biochem. J. 38, 224.DORMAN, S. C , HALE, W. C. & HOSKINS, W. M. (1938). The laboratory rearing of flesh flies and the
relations between temperature, diet and egg production. J. Earn. Ent. 31, 44.EVANS, A. C. (1936). The physiology of the sheep blowfly, Lurilia tericata (Meig.). Trans. R. Ent.
Soc. Lend. 85, pt. 15, 363.FRAKNKEL, G. (1940). Utilization and digestion of carbohydrates by the adult blowfly. J. Exp. Biol.
17, 18.FRAENKEL, G. (1948). The effect of a relative deficiency of lysine and tryptophane in the diet of an
insect. Biochem. J. 43, no. 1, xiv.DE GROOT, A. P. (1953). Phytiol. Camp. 3, fasc. 2 & 3.HOBSON, R. P. (1938). Sheep blowfly investigations. VII. Observations on the development of the
eggs and oviposition in the sheep blowfly Lucilia sericata. Ann. Appl. Biol. 25, no. 3, 573.HOUSE, H. L. (1949). Nutritional studies with Blatella germanica (~L) reared under aseptic conditions.
III. Five essential amino-acids. Canad. Ent. 81, 105.LEMONDE, A. & BERNHARD, R. (1951). Nutrition of Tribolittm confusum larvae. I. A satisfactory
synthetic basal diet for the larvae. Canad. J. Zool. 291, 80.MADDY, K. H. & ELVEHJEM, C. A. (1949). Studies on growth of mice fed rations containing free
amino-acids. J. Biol. Chem. 177, 577.Marmite Food Extract Co. Ltd., London E.C. 3. Marmite Yeast Extract in Medicine and Dietetics.
796 PHYLLIS M. HARLOW
PFEFFFKR, I. W. (1945). The effect of the corpora allata on the metabolism of the adult femalegrasshopper Mtlanopltu differentiate. J. Exp. Zool. 99, 183.
RASSO, S. C. & FRAENKEL, G. (1954). The food requirements of the adult female blowfly, Phormiaregina (Meig.) in relation to ovarian development. Ann. Ent. Sac. Amer. 47, 636.
SCHULTZ, J. P., S T LAWRENCE, P. & NEWMEYER, D. (1046). A chemically denned medium for thegrowth of Droiopkila melanogatter. Anat. Rec. 96, 44.
THOMSEN, E. (1943). An experimental and anatomical study of the corpus allatum in the blowflyCaUiphora erytkrocephala. Vidensk. Medd. dantk. naturh. Foren. Kbh. 106, 320.
THOMSEN, E. (1949). The influence of the corpus allatum on the oxygen consumption of adultCaUiphora erythrocephala. J. Exp. Biol. 26, no. 2, 137.
THOMSEN, E. (1952). Functional significance of the neurosecretory brain cells and the corpuscardiacum in the female blowfly CaUiphora erythrocephala (Meig.). J. Exp. Biol. 29, no. i, 137.
APPENDIX
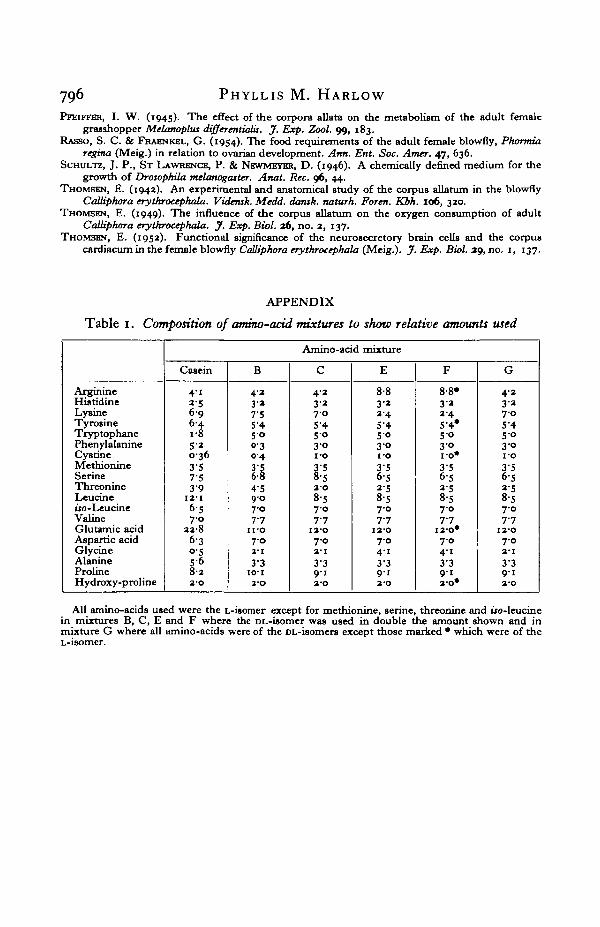
Table 1. Composition of ammo-acid mixtures to show relative amounts used
ArginineHistidineLysineTyrosineTryptophanePhenylalanineCystineMethionineSerineThreonineLeucineiro-LeucineValineGlutamic acidAspartic acidGlycineAlanineProlineHydroxy-proline
Amino-acid mixture
Casein
4 12-56 96-41 85 20-363 57-53 9
12-16-57 0
33-86-3o-55-68 22-O
B
4-23-a7-55'45 00-30 4
6-14'59 07-07-7
1 i-o7-03-1
3 3IO-I
j - o
C
4-23 27 05-45 03°i -o3/58-52-O
8-57-07-7
I2-O7-02-1
3-39 13-O
E
8-83-23 45'45-03 0i -o3 56-52'58-S7-07 7
I2-O7-O4' i3 39'i2'O
F
8-8»3 13 45-4*5 03°1 0 *
3'56-52-58-57-07-7
120*7 04' i3-39 12 O *
G
4-33-37 05'45°3-0i-o3'56 5*"58-57-07-7
I2-O7-02-1
3-39 12-O
All amino-acids used were the L-isomer except for methionine, serine, threonine and iio-leucinein mixtures B, C, E and F where the DL-isomer was used in double the amount shown and inmixture G where all amino-acids were of the DL-isomers except those marked • which were of theL-isomer.
Ovarial development and relation to adult nutrition in blowfly 797
APPENDIX (cont.)
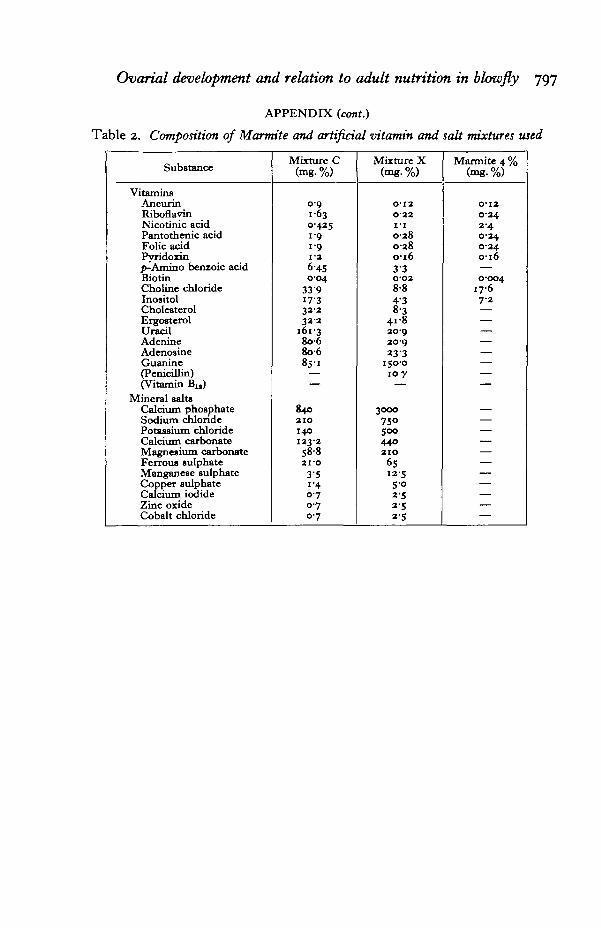
Table 2. Composition of Marmite and artificial vitamin and salt mixtures used
Substance
VitaminsAneurinRiboflavinNicotinic acidNico t in i c acidFolic acidPyridoxinp-Amino benioic acidBiotinCholine chlorideInositolCholesterolErgosterolUracilAdenineAdenosineGuanine(Penicillin)(Vitamin Bu)
Mineral saltsCalcium phosphateSodium chlorideSodium chlorideCalcium carbonateMagnesium carbonateFerrous sulphateManganese sulphateCopper sulphateCalcium iodideZinc oxideCobalt chloride
Mixture C(mg. %)
o-o1 6 30-425i-o1-91-2
6-450-04
33917-33 3 23 3 2
161-38 0 68o-68 5 1
——
8402 1 0140123-2S§-82I-O
3'5i -40-70-70-7
Mixture X(mg. %)
O-I2O'22I - IO-28O-280-163'3O-O28-84'38-3
41-820-920-92 3 3
150-010 y
3000
75°500
44°a i o6512-5S-o2 52 52-5
Marmite 4 %(mg.%)
0-120-242-40-240-240-16—0-004
1 7 67-2
———————
———————————

























![Optical(Coherence(Tomography:(Age(estimation(of(Calliphora ......Introduction(Pupae,)a)sedentary)and)long]lasting)stage)of)the)blowfly)lifecycle,)are)often)reported)at)crime)scenes)](https://img.pdfslide.net/doc/110x75/60b54e2575eeff0dad7cccb6/opticalcoherencetomographyageestimationofcalliphora-introductionpupaeasedentaryandlonglastingstageoftheblowflylifecycleareoftenreportedatcrimescenes.jpg)



