Embed Size (px)
Citation preview
A very terse overview of metabolism
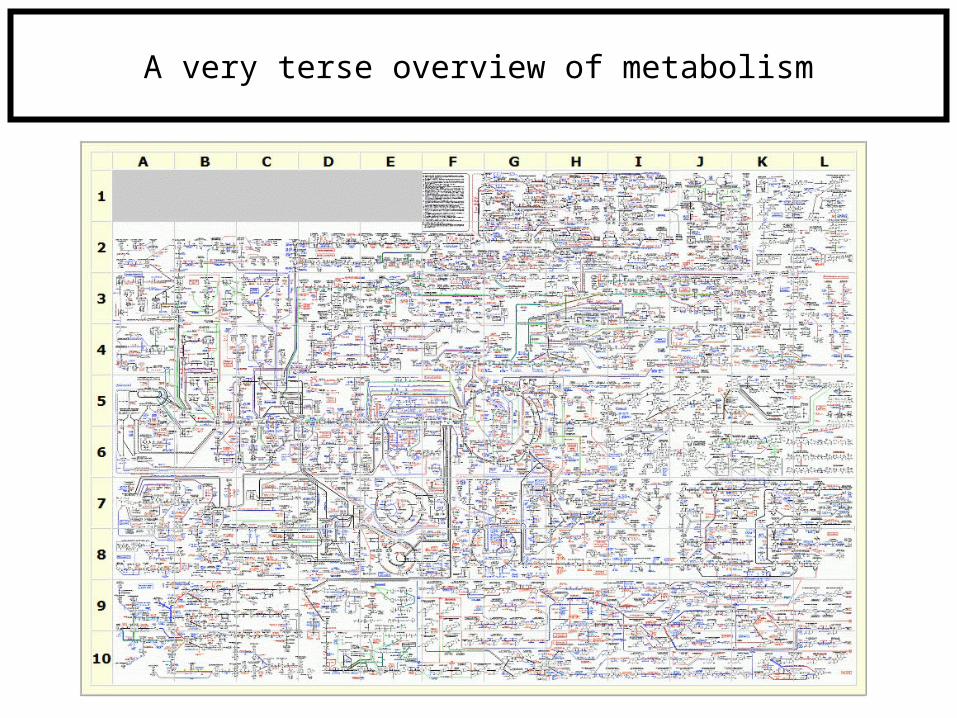
Anabolism vs. Catabolism
“lysis”i.e., glycolysis
“genesis”i.e., gluconeogenesis

Overview of metabolism
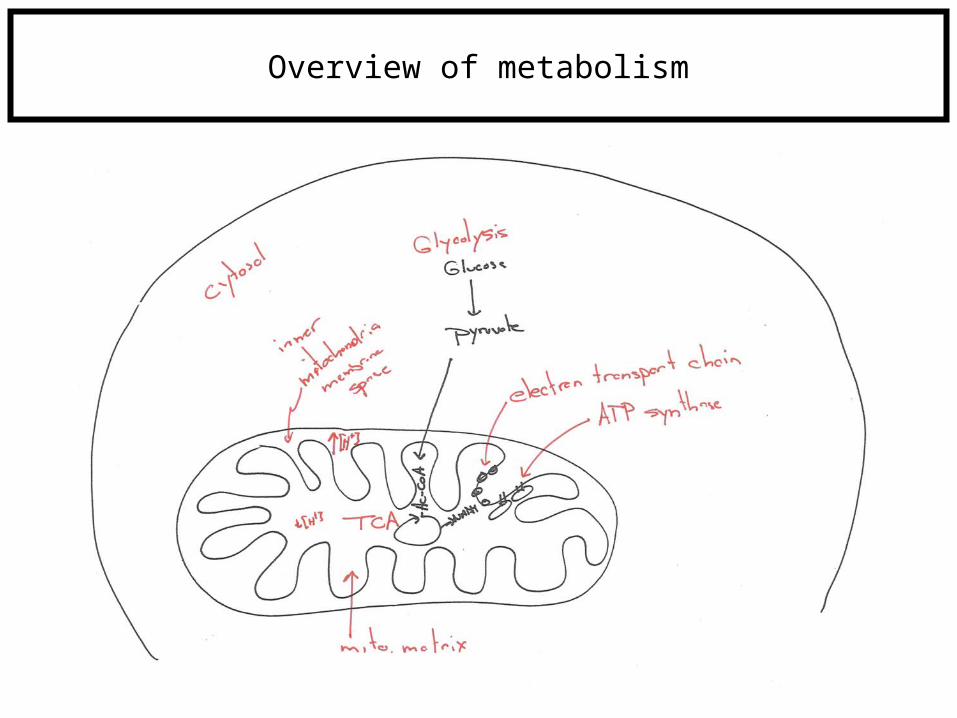
Overview of metabolism
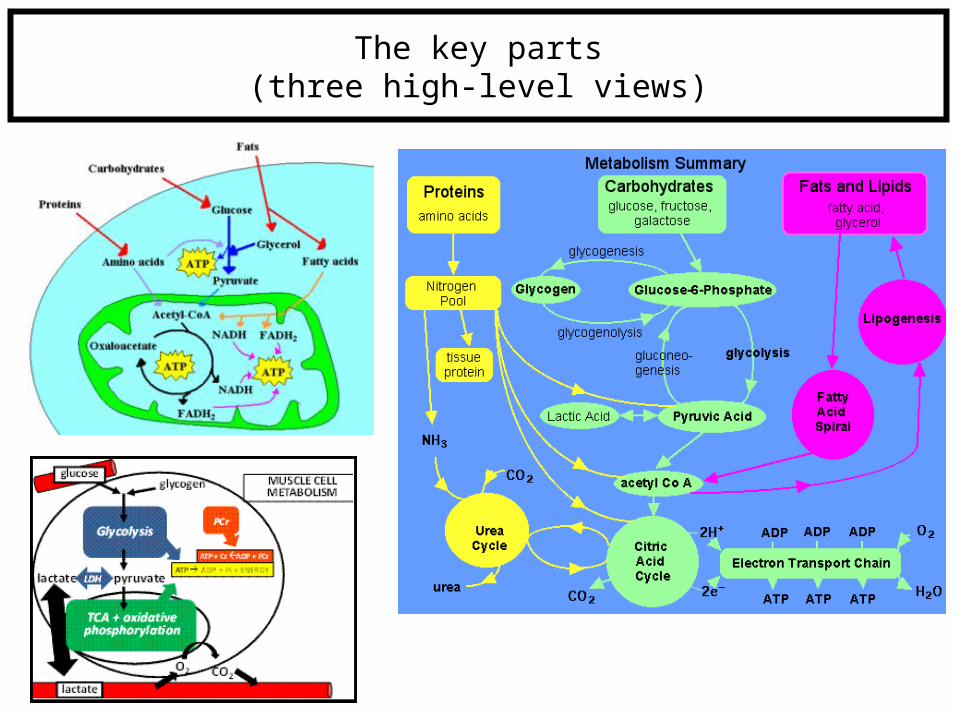
The key parts(three high-level views)
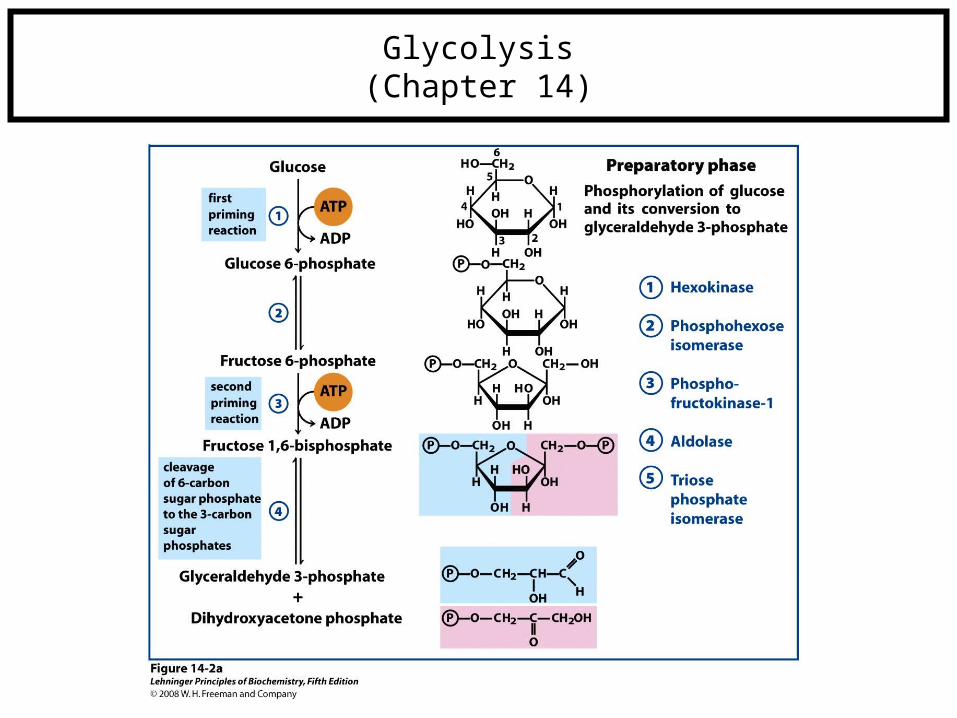
Glycolysis(Chapter 14)
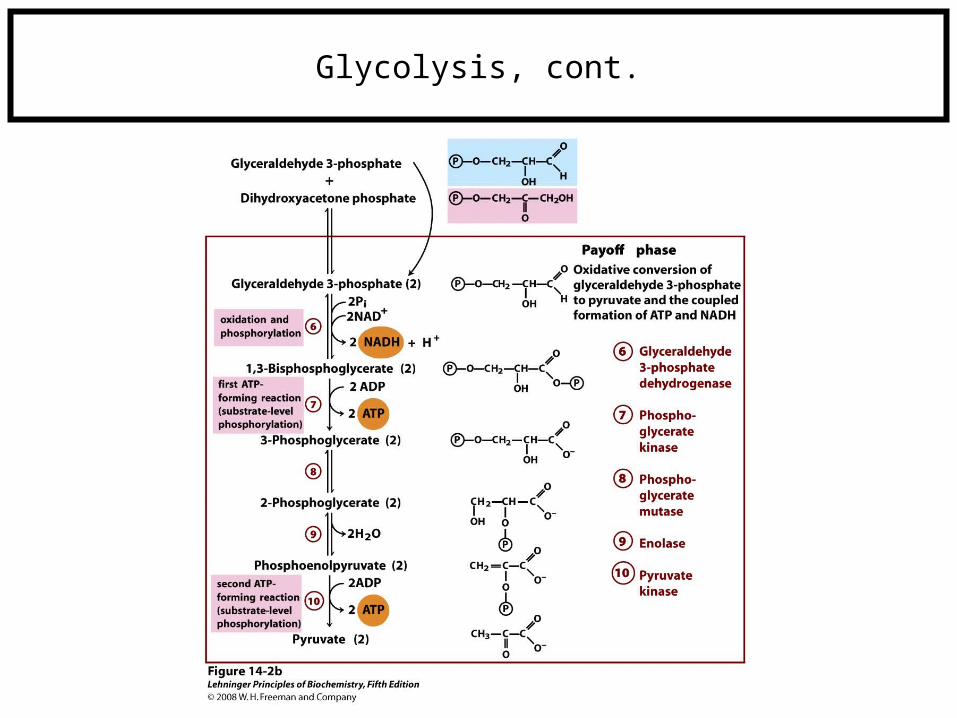
Glycolysis, cont.
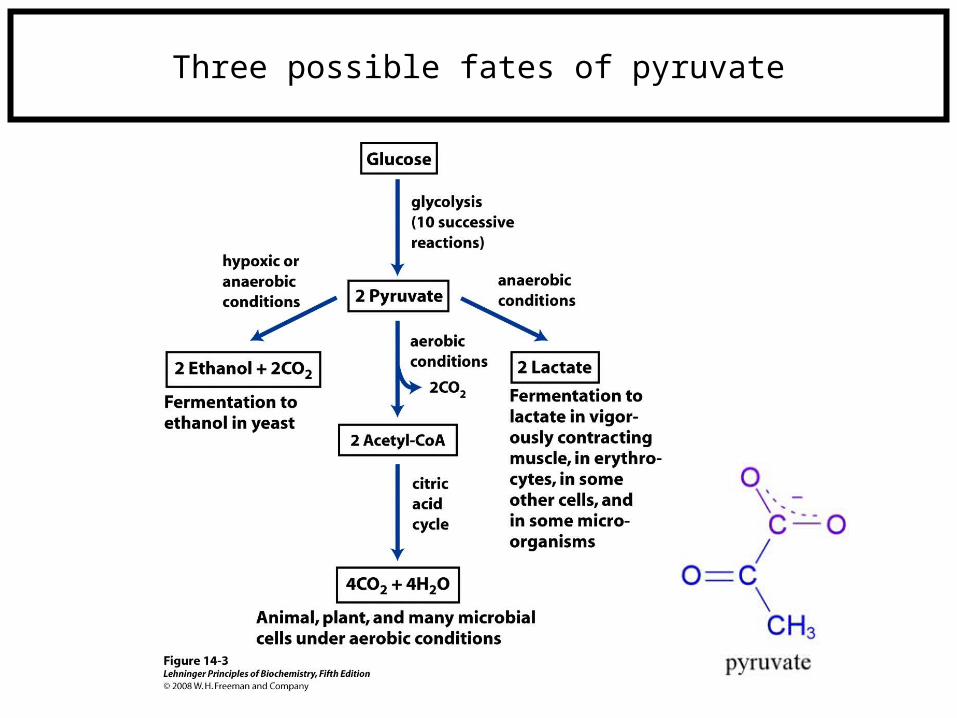
Three possible fates of pyruvate
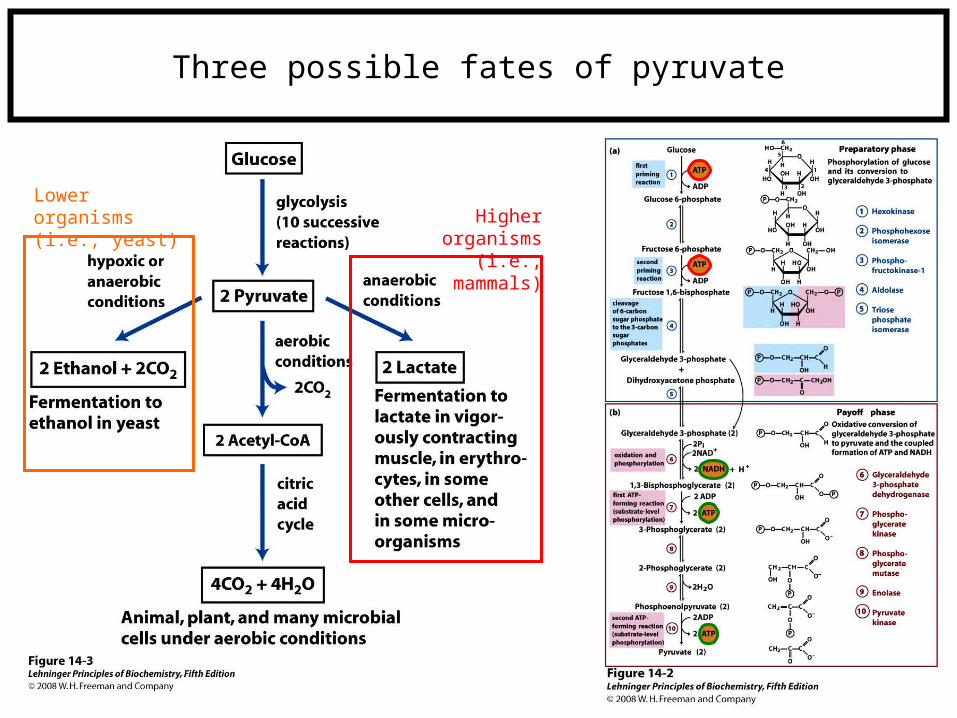
Lower organisms (i.e., yeast) Higher organisms
(i.e., mammals)
Three possible fates of pyruvate
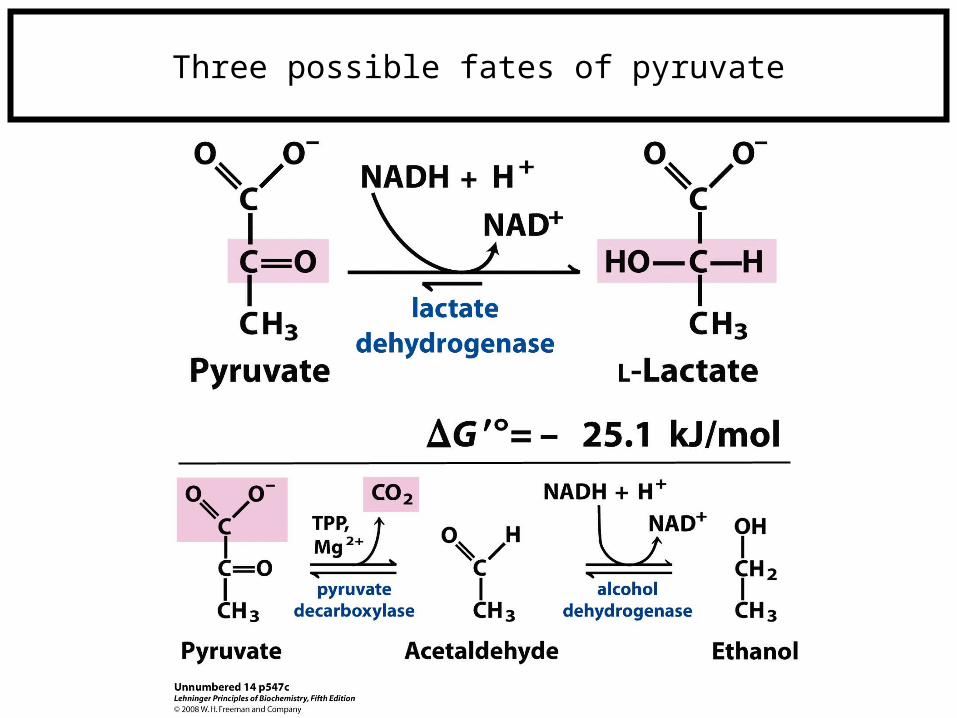
Three possible fates of pyruvate
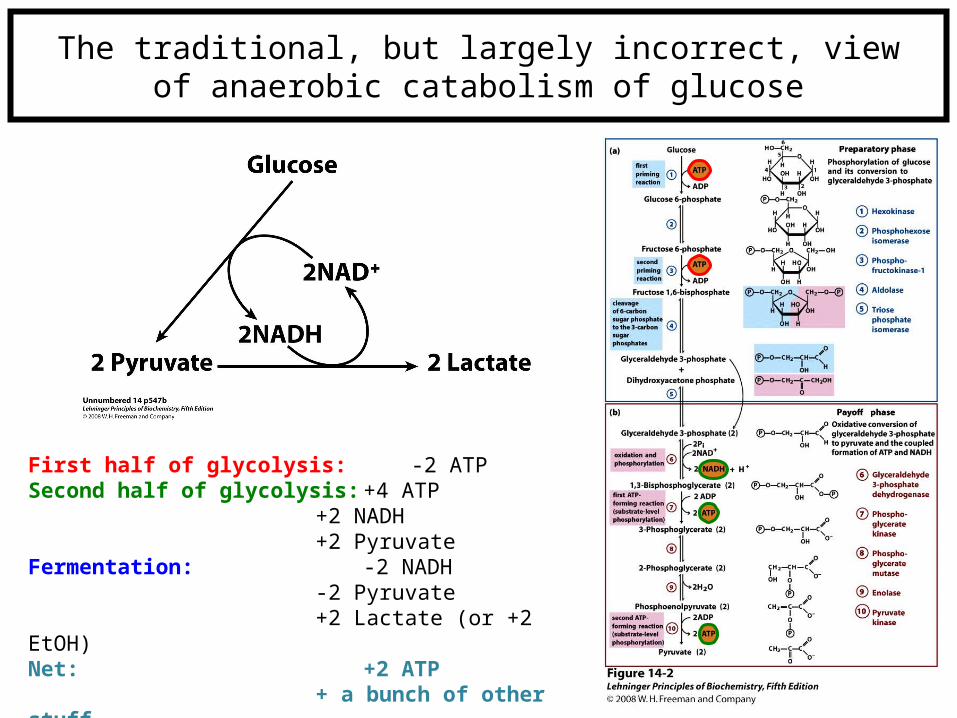
The traditional, but largely incorrect, viewof anaerobic catabolism of glucose
First half of glycolysis: -2 ATPSecond half of glycolysis: +4 ATP
+2 NADH+2 Pyruvate
Fermentation: -2 NADH-2 Pyruvate+2 Lactate (or +2 EtOH)
Net: +2 ATP+ a bunch of other stuff
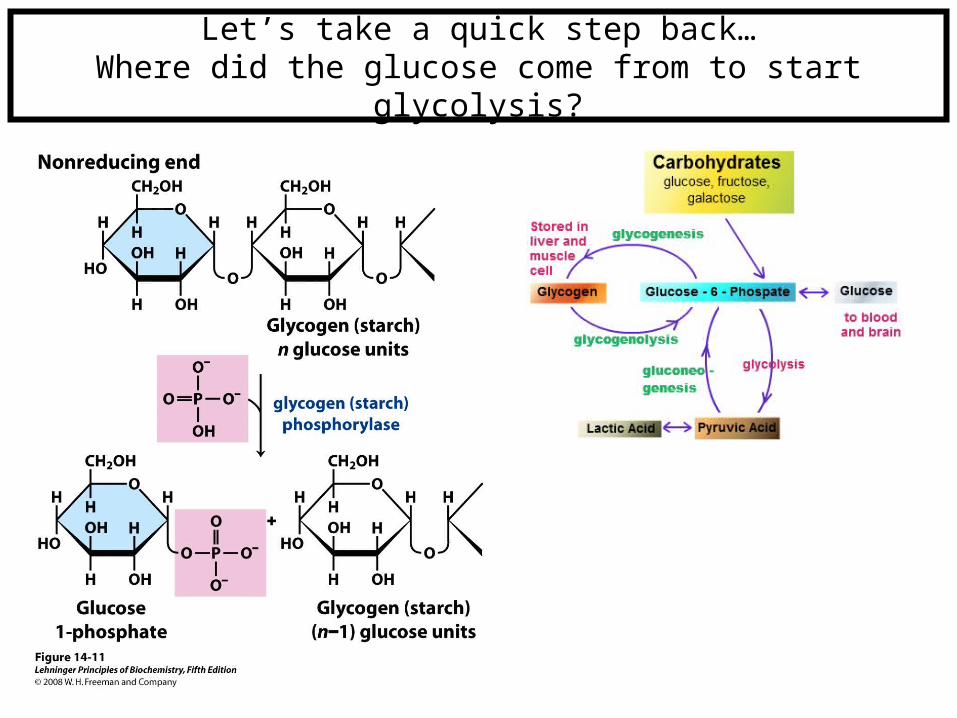
Let’s take a quick step back…Where did the glucose come from to start glycolysis?
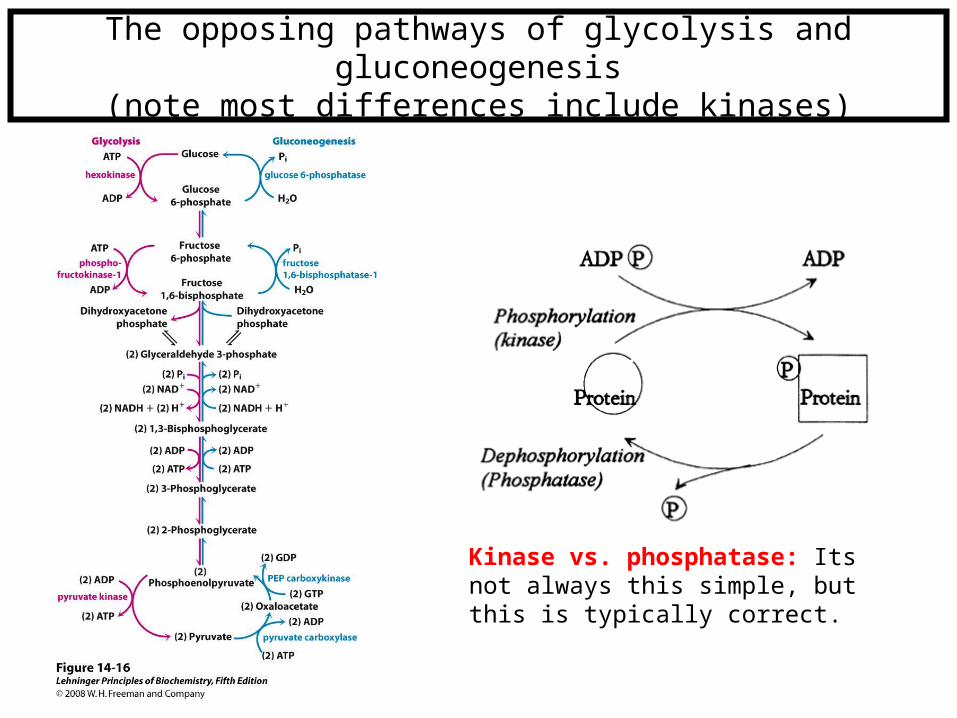
The opposing pathways of glycolysis and gluconeogenesis
(note most differences include kinases)
Kinase vs. phosphatase: Its not always this simple, but this is typically correct.
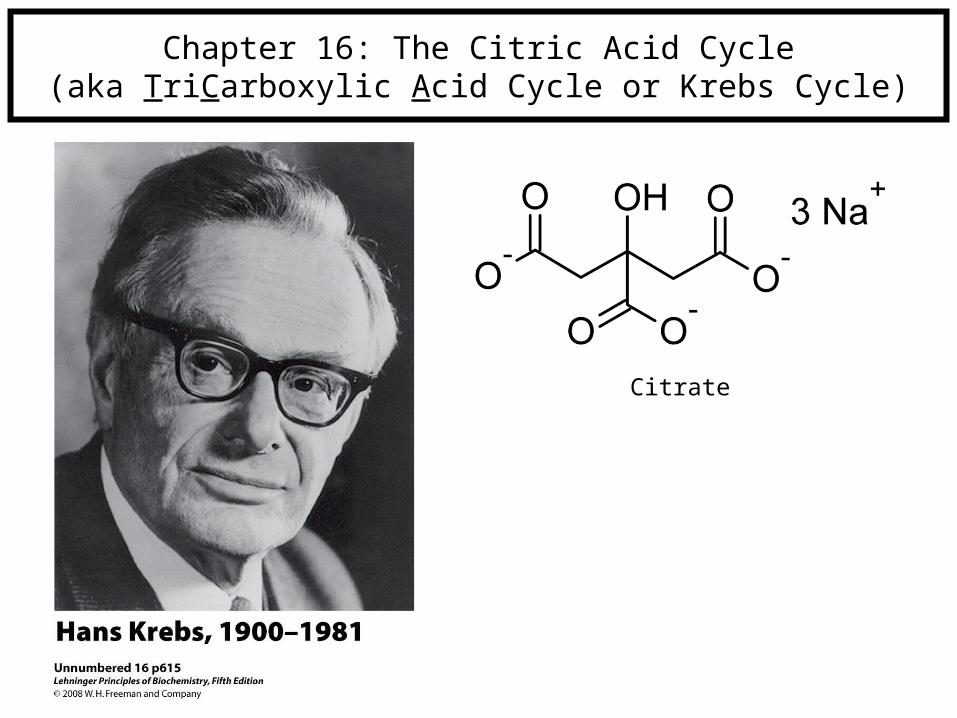
Chapter 16: The Citric Acid Cycle(aka TriCarboxylic Acid Cycle or Krebs Cycle)
Citrate
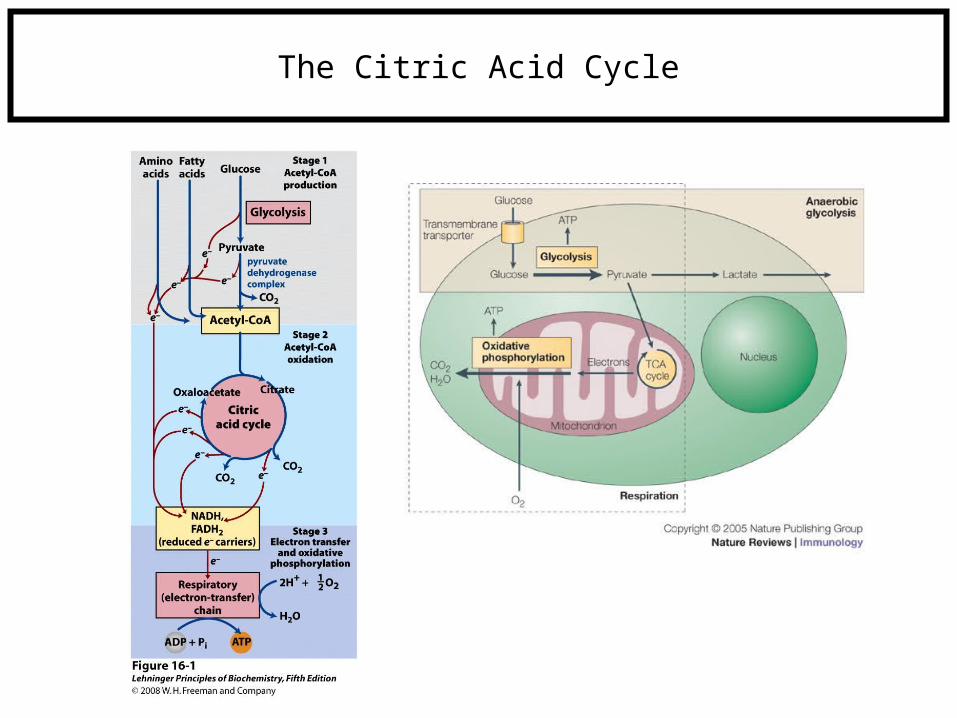
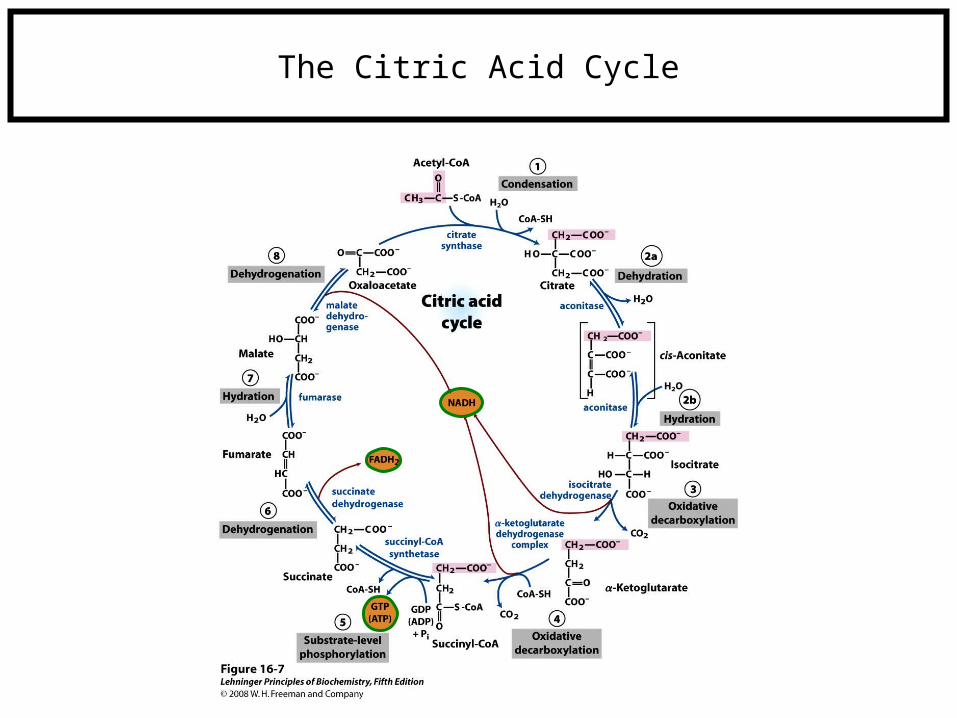
The Citric Acid Cycle
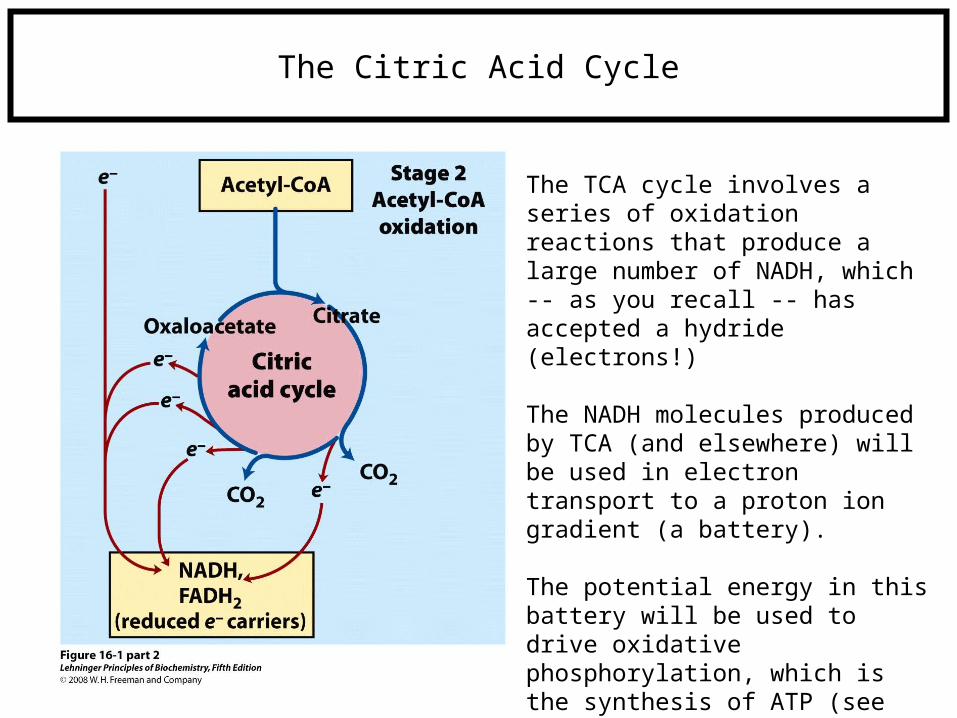
The Citric Acid Cycle
The TCA cycle involves a series of oxidation reactions that produce a large number of NADH, which -- as you recall -- has accepted a hydride (electrons!)
The NADH molecules produced by TCA (and elsewhere) will be used in electron transport to a proton ion gradient (a battery).
The potential energy in this battery will be used to drive oxidative phosphorylation, which is the synthesis of ATP (see purple box on previous slide).
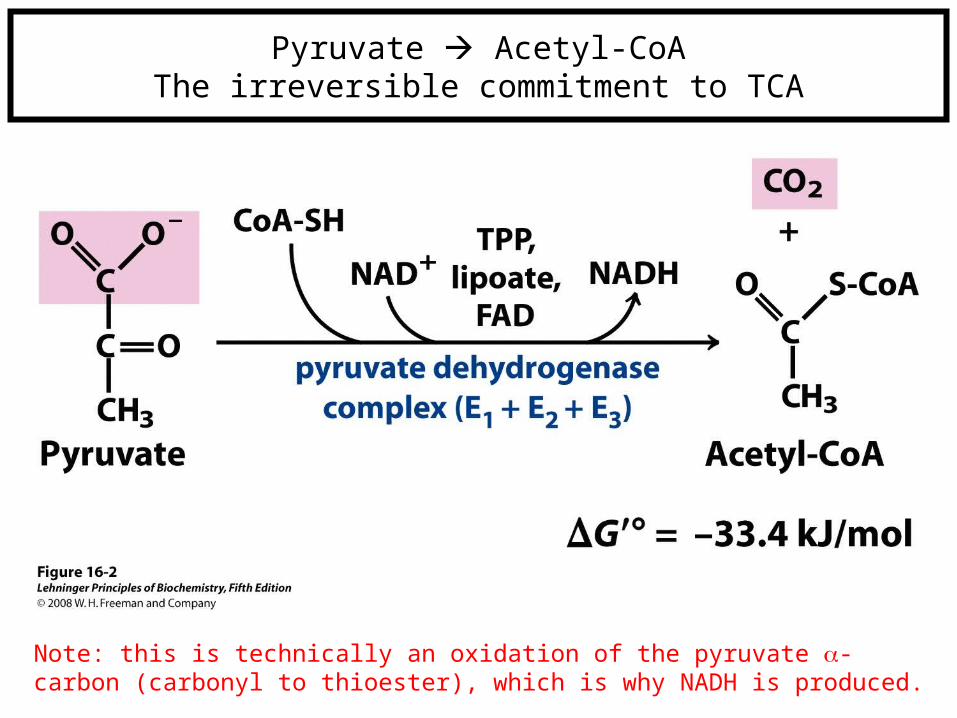
Pyruvate Acetyl-CoAThe irreversible commitment to TCA
Note: this is technically an oxidation of the pyruvate -carbon (carbonyl to thioester), which is why NADH is produced.
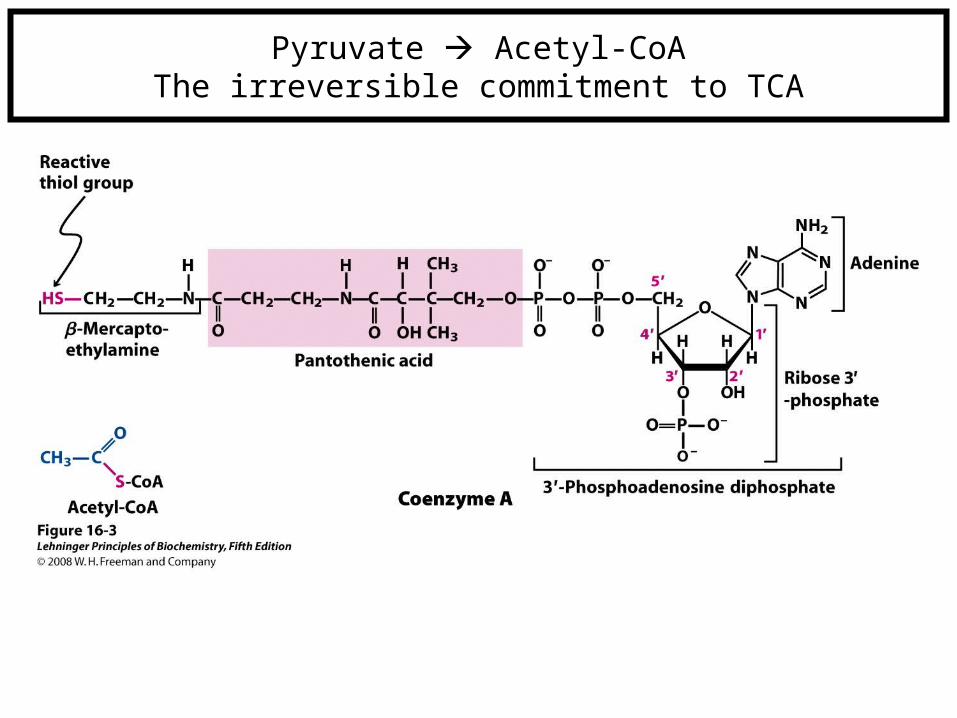
Pyruvate Acetyl-CoAThe irreversible commitment to TCA
The Citric Acid Cycle
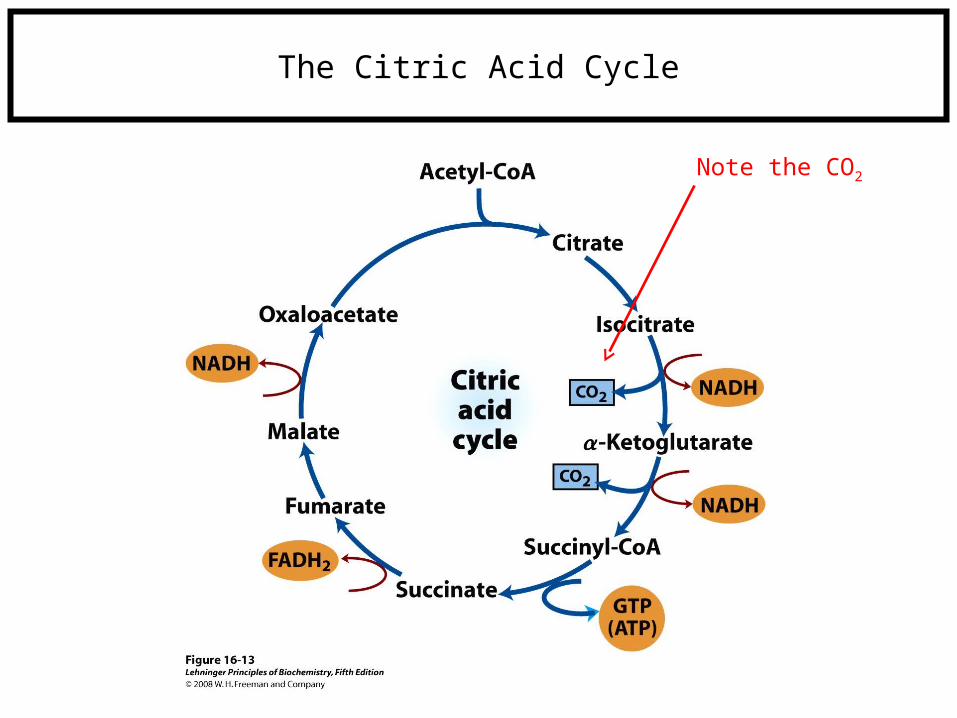
The Citric Acid Cycle
Note the CO2
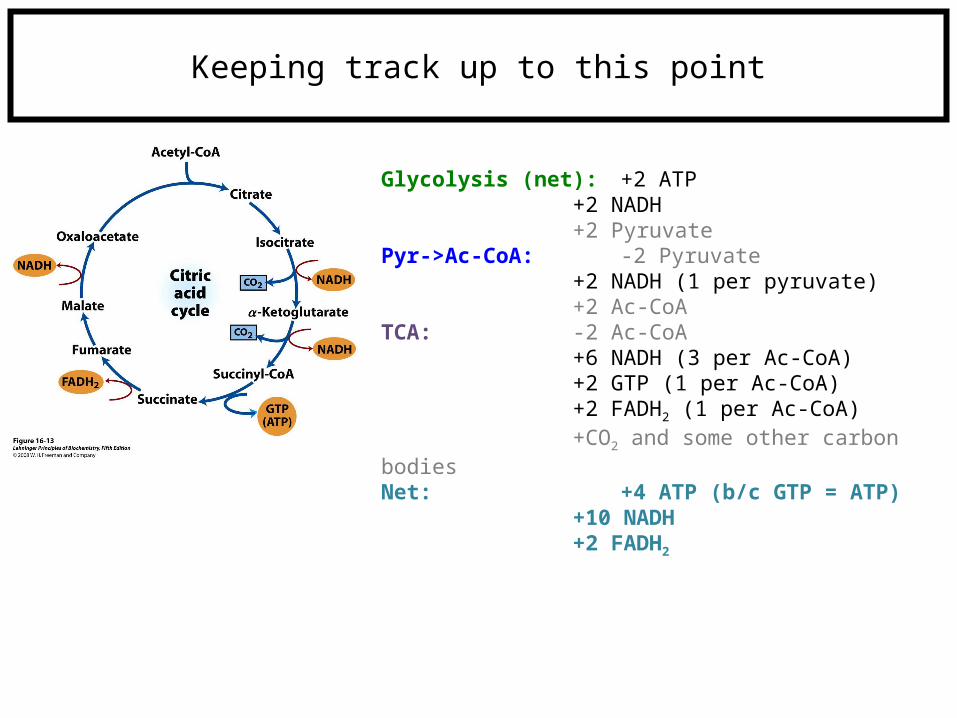
Keeping track up to this point
Glycolysis (net): +2 ATP+2 NADH+2 Pyruvate
Pyr->Ac-CoA: -2 Pyruvate+2 NADH (1 per pyruvate)+2 Ac-CoA
TCA: -2 Ac-CoA+6 NADH (3 per Ac-CoA)+2 GTP (1 per Ac-CoA)+2 FADH2 (1 per Ac-CoA)+CO2 and some other carbon bodies
Net: +4 ATP (b/c GTP = ATP)+10 NADH+2 FADH2
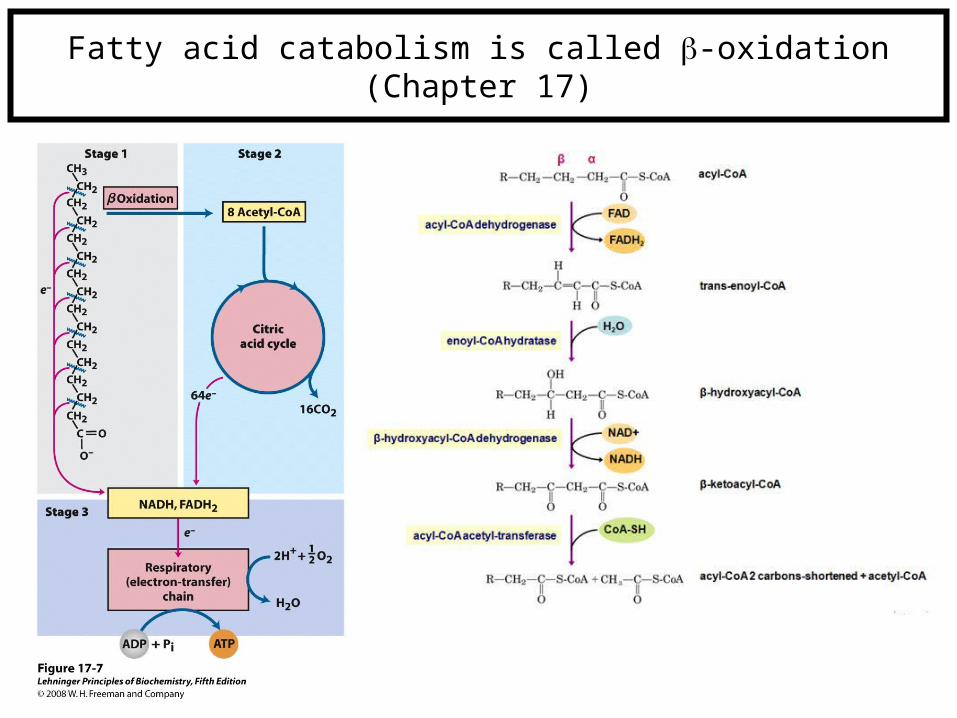
Fatty acid catabolism is called -oxidation(Chapter 17)
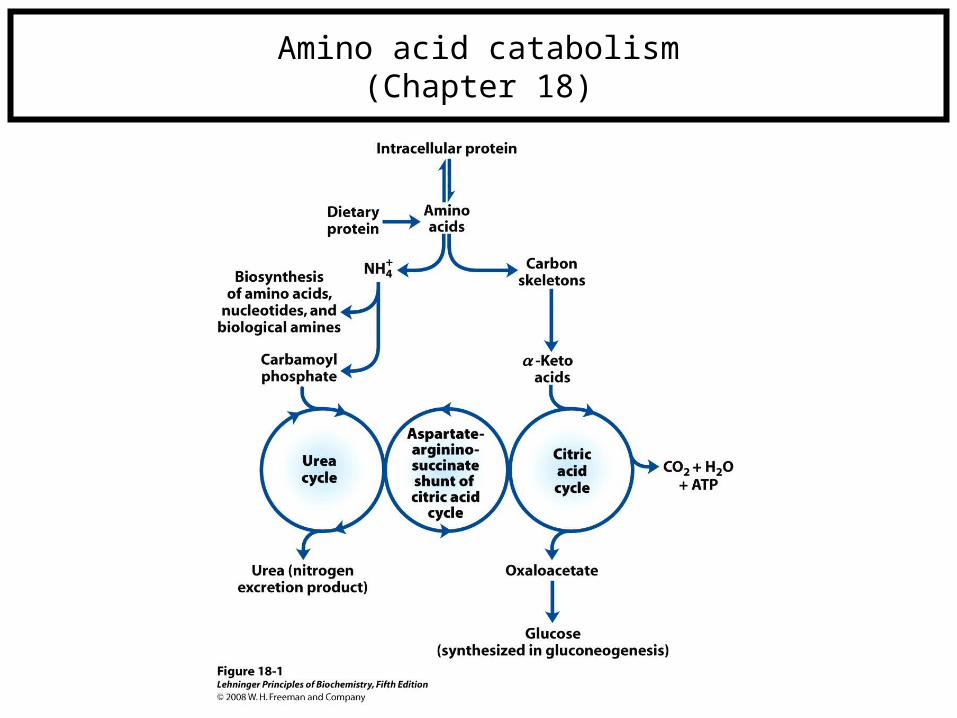
Amino acid catabolism(Chapter 18)

A [H+] gradient is produced using the free energyproduced from electron transport (Chapter 19)
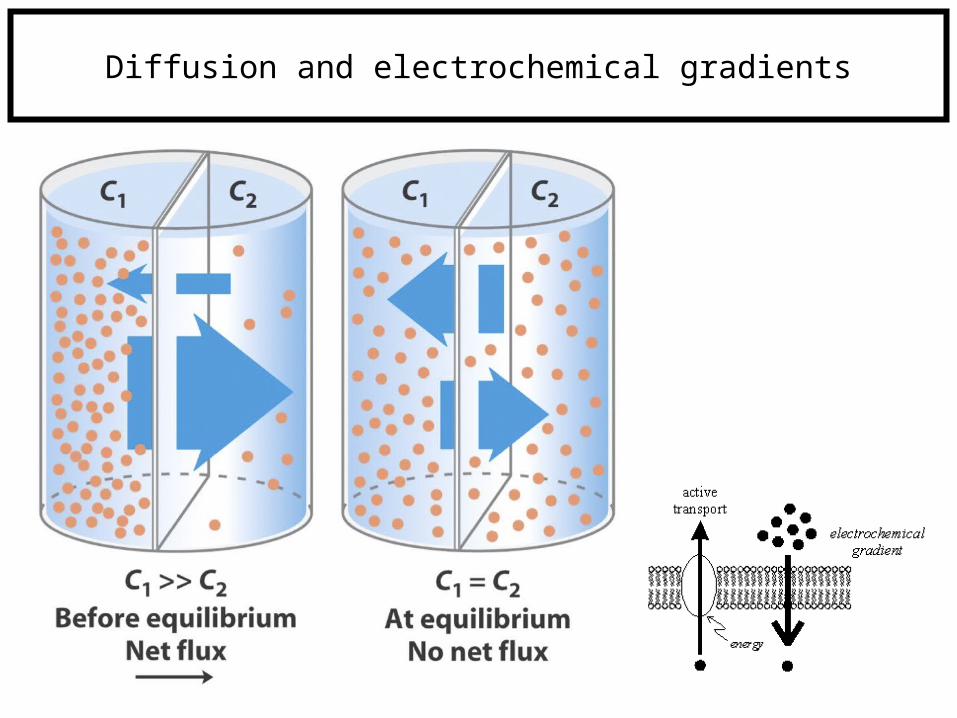
Diffusion and electrochemical gradients
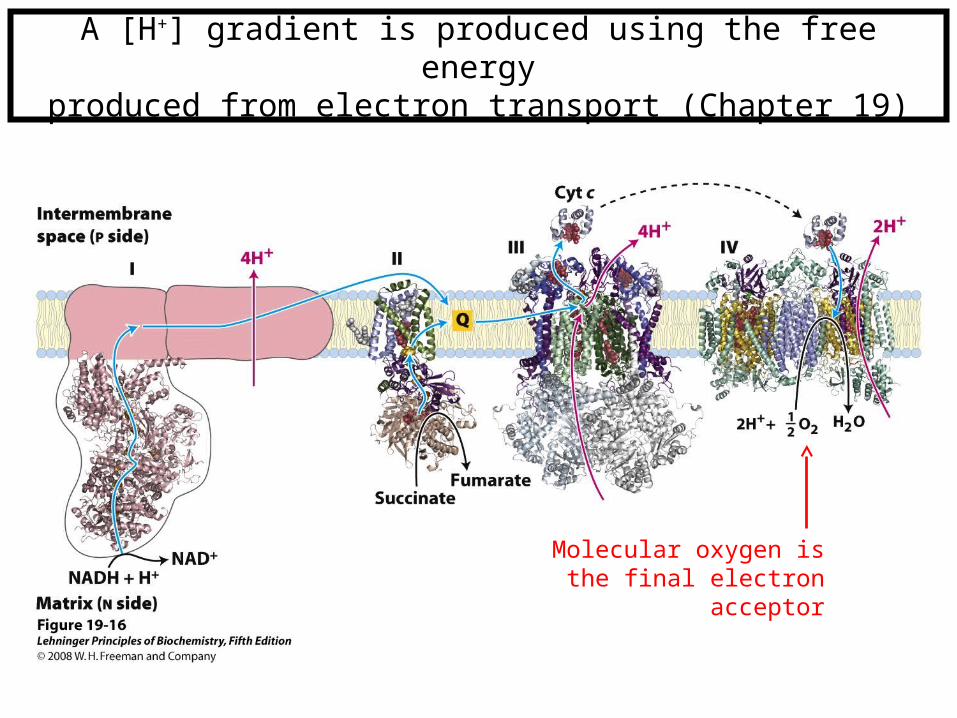
A [H+] gradient is produced using the free energyproduced from electron transport (Chapter 19)
Molecular oxygen is the final electron acceptor
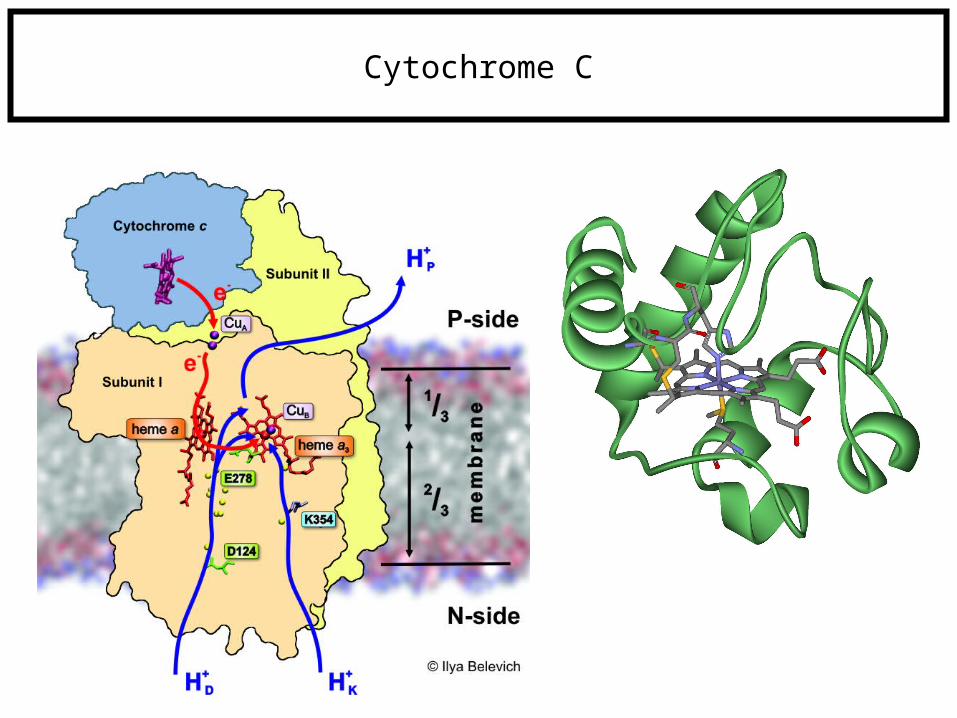
Cytochrome C
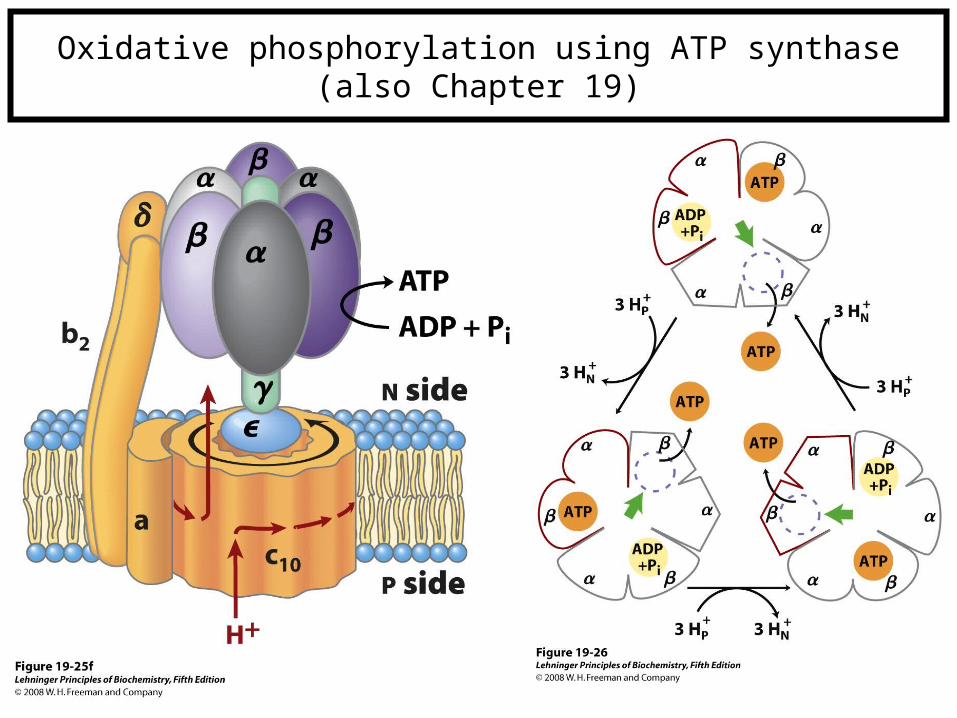
Oxidative phosphorylation using ATP synthase(also Chapter 19)
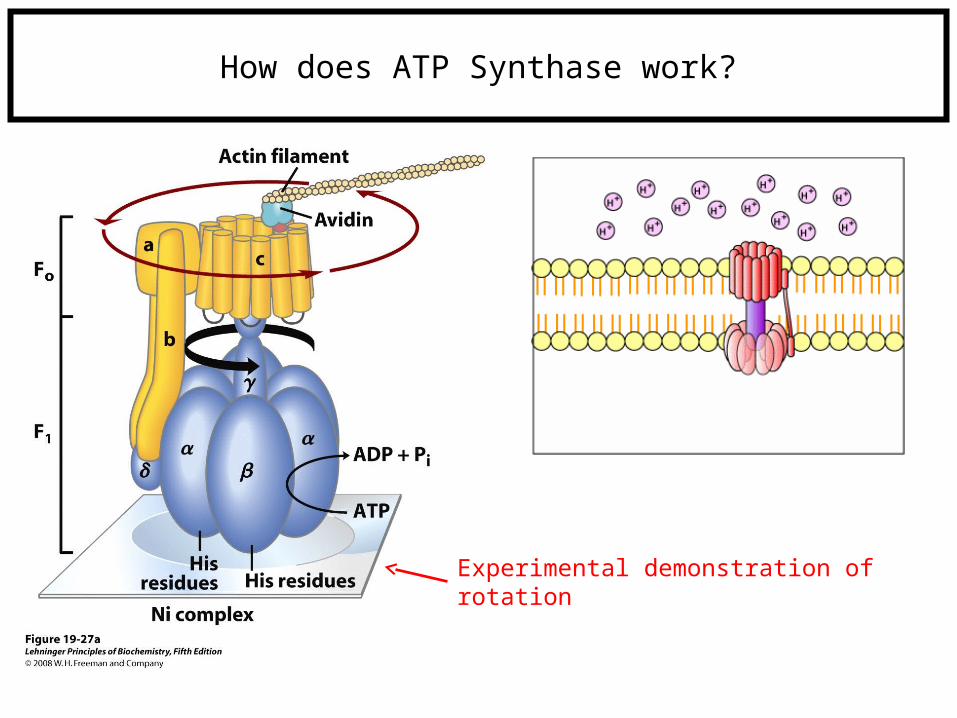
How does ATP Synthase work?
Experimental demonstration of rotation

The final score (sorta)
Glycolysis (net): +2 ATP+2 NADH+2 Pyruvate
Pyr->Ac-CoA: -2 Pyruvate+2 NADH (1 per pyruvate)+2 Ac-CoA
TCA: -2 Ac-CoA+6 NADH (3 per Ac-CoA)+2 GTP (1 per Ac-CoA)+2 FADH2 (1 per Ac-CoA)+CO2 and some other carbon bodies
Net: +4 ATP (b/c GTP = ATP)+10 NADH = +30 ATP (assuming 3 ATP per NADH)+2 FADH2 = +4 ATP (assuming 2 ATP per FADH2)-----------------------------------------------------------------------
+38 ATP (4 from substrate phosphorylation; 34 from oxidative phosphorylation)
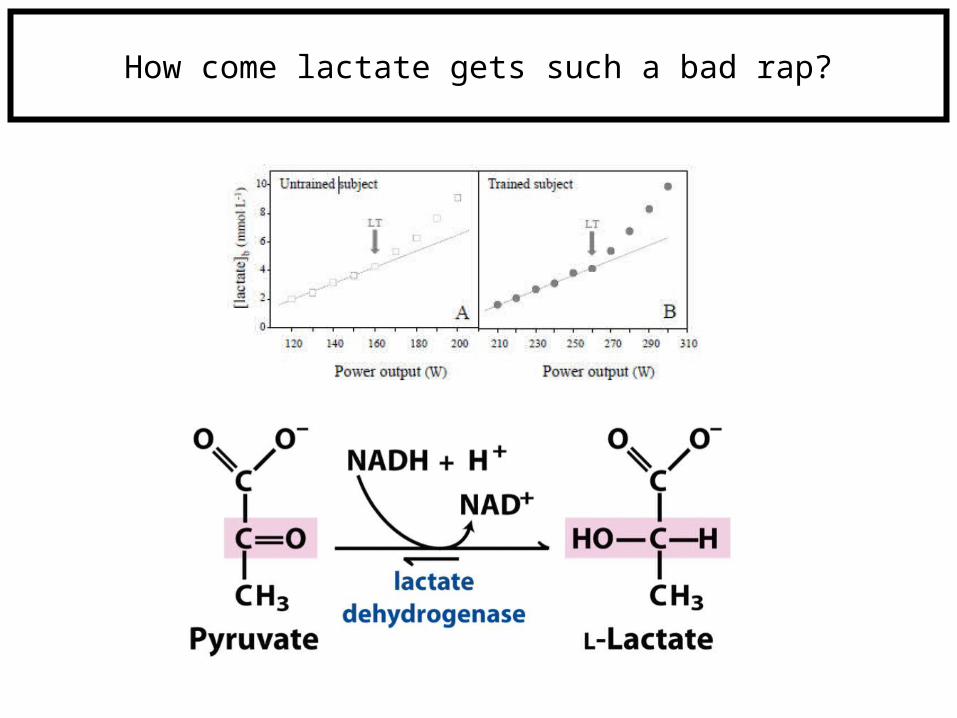
How come lactate gets such a bad rap?
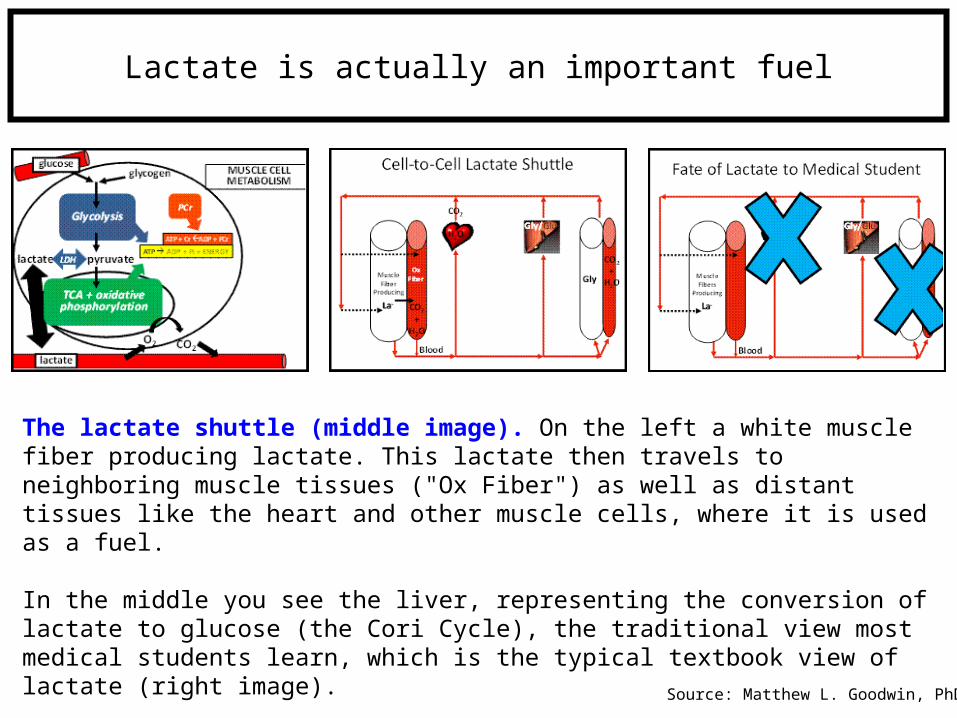
Lactate is actually an important fuel
The lactate shuttle (middle image). On the left a white muscle fiber producing lactate. This lactate then travels to neighboring muscle tissues ("Ox Fiber") as well as distant tissues like the heart and other muscle cells, where it is used as a fuel.
In the middle you see the liver, representing the conversion of lactate to glucose (the Cori Cycle), the traditional view most medical students learn, which is the typical textbook view of lactate (right image).
Source: Matthew L. Goodwin, PhD
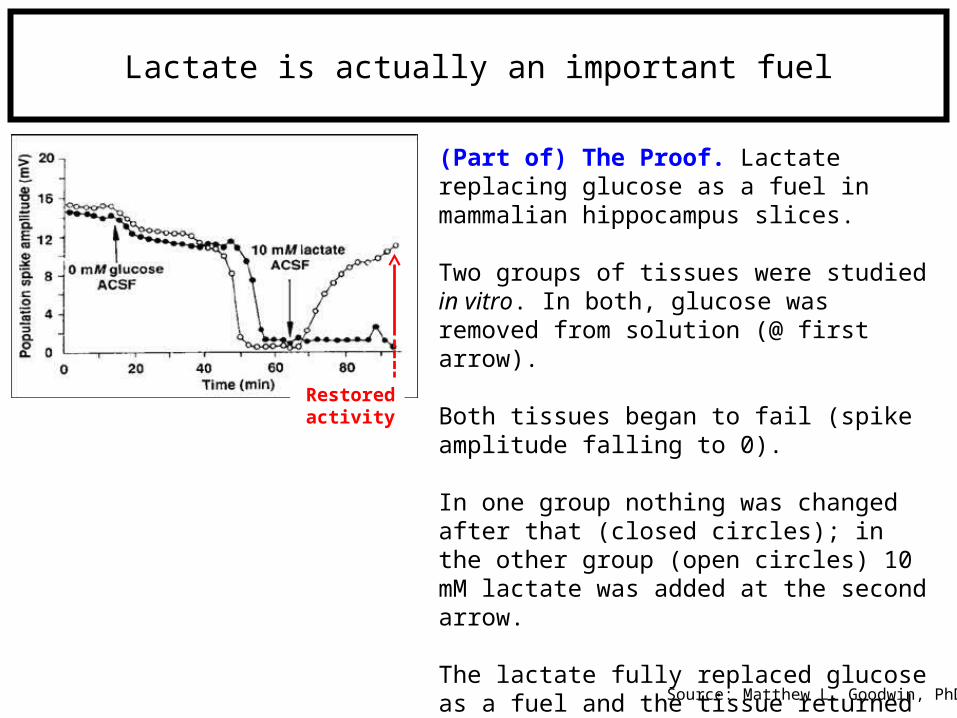
Lactate is actually an important fuel
(Part of) The Proof. Lactate replacing glucose as a fuel in mammalian hippocampus slices.
Two groups of tissues were studied in vitro. In both, glucose was removed from solution (@ first arrow).
Both tissues began to fail (spike amplitude falling to 0).
In one group nothing was changed after that (closed circles); in the other group (open circles) 10 mM lactate was added at the second arrow.
The lactate fully replaced glucose as a fuel and the tissue returned to normal vitality.
Source: Matthew L. Goodwin, PhD
Restored activity
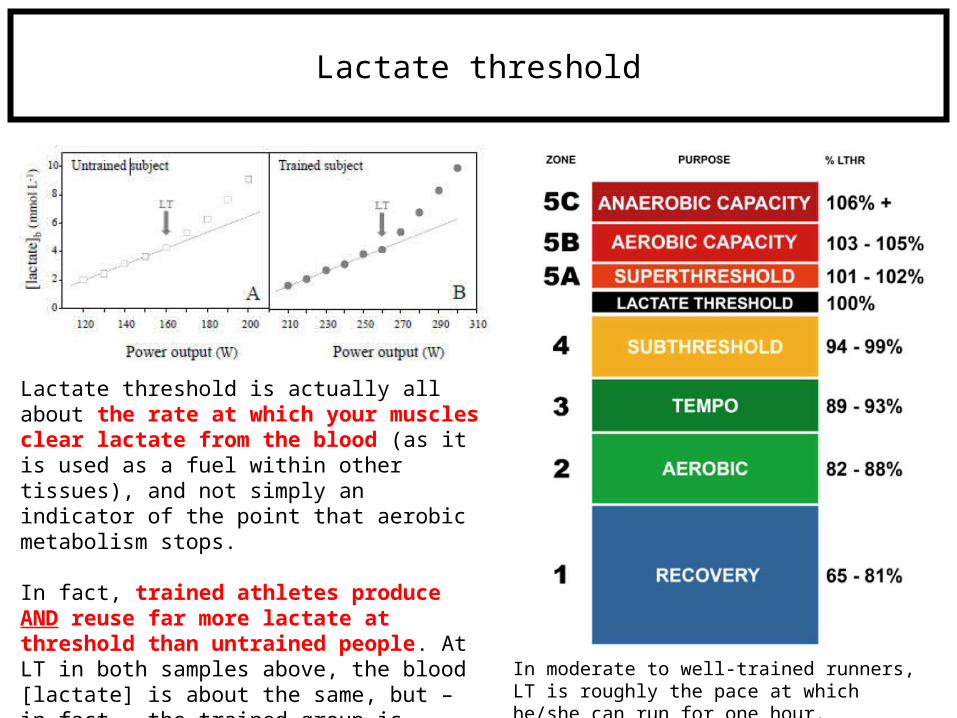
Lactate threshold
Lactate threshold is actually all about the rate at which your muscles clear lactate from the blood (as it is used as a fuel within other tissues), and not simply an indicator of the point that aerobic metabolism stops.
In fact, trained athletes produce AND reuse far more lactate at threshold than untrained people. At LT in both samples above, the blood [lactate] is about the same, but – in fact – the trained group is producing about 60% more lactate, and thus also clearing it 60% faster. In moderate to well-trained runners, LT is roughly
the pace at which he/she can run for one hour.