Embed Size (px)
Citation preview

Cell, Vol. 44, 337-344, January 31, 1966, Copyright 0 1966 by Cell Press
Abelson Virus Drives the Differentiation of Harvey Virus-Infected Erythroid Cells
Gerald L. Waneck,* Linda Keyes, and Naomi Rosenberg Departments of Pathology, Microbiology, and Molecular Biology Cancer Research Center Immunology Graduate Program Tufts University School of Medicine Boston, Massachusetts 02111
Summary
Abelson murine leukemia virus (A-MuLV) and Harvey murine sarcoma virus (Ha-MSV) are retroviruses carry- ing unrelated one genes. However, both of these viruses are capable of stimulating the growth and differentiation of erythroid precursor cells; the target cells for both appear at the same time during fetal de- velopment and follow a similar pattern throughout on- togeny. In addition, the colonies induced by each vi- rus are morphologically similar and synthesize the adult form of hemoglobin. However, A-MuLV-infected cells are Epo-independent, whereas Ha-MSV-infected cells are Epo-dependent. Superinfection of Ha-MSV- infected cells with AMuLV overrides their Epo-de- pendency. Thus, the consequences of the infection are determined by the interaction of the different one gene products with identical or similar erythroid cells.
Introduction
Several murine retroviruses are capable of inducing erythroid colony formation in vitro, including those carry- ing the one genes ras, abl, src, and mos (Hankins and Scolnick, 1981; Waneck and Rosenberg, 1981; Ratnofsky and Rosenberg, unpublished data; Hankins et al., 1982). In most cases, direct evidence demonstrates that the abil- ity to induce the colonies is a function of the transforming protein of the virus. Because the transforming proteins en- coded by these viruses are structurally distinct and fall into at least two functional classes (reviewed in Weiss et al., 1982), the mechanisms involved in stimulating erythroid colony formation may not be identical for each virus.
Abelson murine leukemia virus (A-MuLV) containing the one gene abl and Harvey murine sarcoma virus (Ha-MSV) containing the one gene ras exemplify the differences among viruses that induce erythroid colonies. The v-abl sequences are expressed as a 180,000 MW protein that is a tyrosine-specific protein kinase (Witte et al., 1978; Reynolds et al., 1978; Rosenberg and Witte, 1980; Witte et al., 1980; Wang et al., 1982). The v-Ha-ras sequences encode a 21,000 MW protein that binds guanine nucleo- tides and is associated with a threonine-specific au-
* Present address: Biogen Incorporated. Cambridge, Massachu- setts 02139.
tophosphorylating activity (Scolnick et al., 1979). Because differences in GTP binding are found when various v-ras or mutated eras proteins are compared to normal c-ras proteins, this binding is believed to be important in trans- formation (Gibbs et al., 1984; McGrath et al., 1984; Sweet et al., 1984).
In vitro, A-MuLV and Ha-MSV transform a variety of cell types including NIHMT3 fibroblasts (Scher and Siegler, 1975; Jainchill et al., 1989) and lymphoblastoid cells pres- ent in mouse bone marrow or late gestation fetal liver (Rosenberg et al., 1975; Rosenberg and Baltimore, 1978; Pierce and Aaronson, 1982). In addition, Ha-MSV induces erythroid colonies after infection of bone marrow cells from mice with drug-induced anemia (Hankins and Scol- nick, 1981) and A-MuLV induces similar colonies when certain fetal tissues are infected (Waneck and Rosenberg, 1981). These initial studies did not address the relation- ship of the two viral systems. Different cells and plating conditions were used and small amounts of the hormone erythropoietin (Epo), a normal mediator of erythrocyte de- velopment, were required for terminal differentiation of the Ha-MSV-infected colonies, while no Epo was used in the A-MuLV system.
The dependence of Ha-MSV-infected cells on Epo for differentiation and the apparent Epo-independence of the A-MuLV-induced colonies suggested that one of two hy- potheses would explain the differences observed in the two experimental systems: the target cell hypothesis that the viruses infect distinct sets of erythroid precursors, or the instructive hypothesis that the expression of the dis- tinct viral one gene products affects growth and differenti- ation of similar erythroid cells in different ways. We have investigated these two hypotheses using an ontogenic ap- proach to examine the target cells and by dissecting the role of virus, feeder cells, and external hormonal signals in colony formation. These experiments show that the tar- get cells for A-MuLV and Ha-MSV are similar in tissue dis- tribution and that the cells in the colonies are morphologi- cally indistinguishable. However, the A-MuLV-infected colony forming cells are relatively Epo-independent while the Ha-MSV-infected cells are strongly dependent on Epo, both for growth and for differentiation. Because A-MuLV infection of erythroid cells initially stimulated to grow by Ha-MSV overrides the Epo-dependency of the Ha-MSV- infected erythroid cells, A-MuLV acts to replace the need for the hormonal signals in this system. Thus, A-MuLV and Ha-MSV seem to infect the same population of erythroid cells, but Ha-MSV is only able to initiate colony formation, while A-MuLV is able both to stimulate growth and to drive differentiation.
Results
Tissue Distribution and Erythropoietin Sensitivity of Colonies Induced by A-MuLV and Ha-MSV We compared the frequency of target cells for A-MuLV and Ha-MSV present in fetal cell populations when the in-
Cell 338
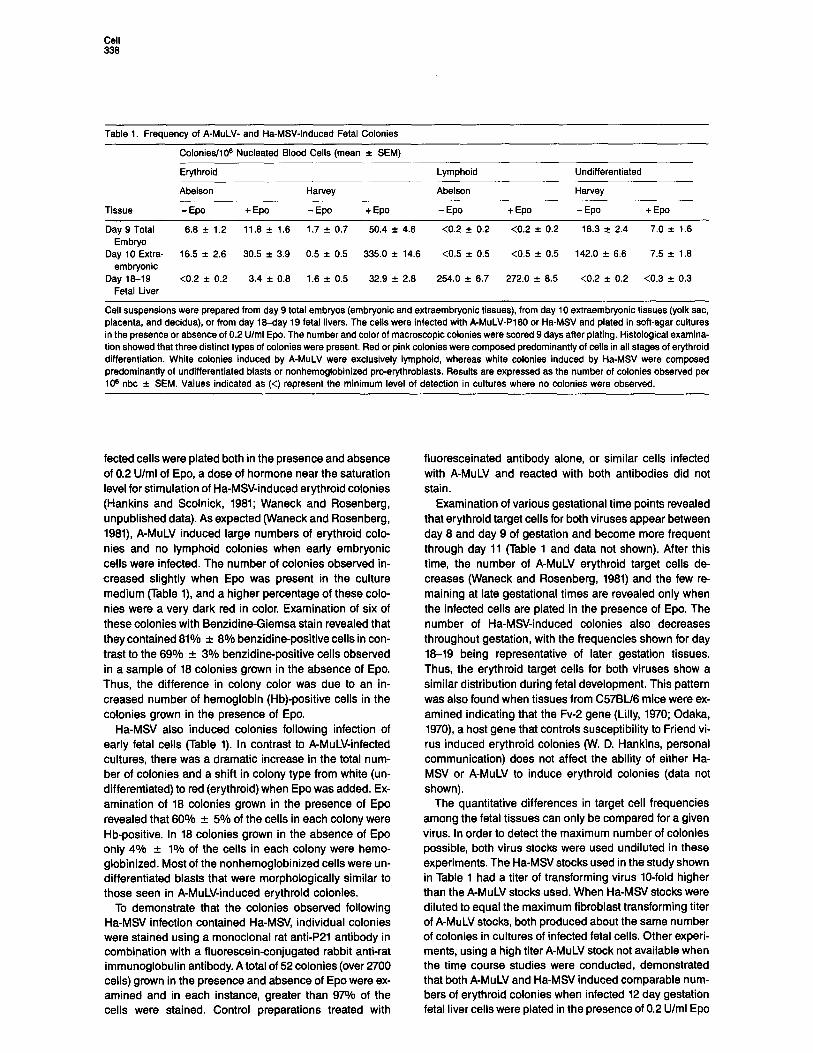
Table 1. Frequency of A-MuLV- and Ha-MSV-Induced Fetal Colonies
Colonies/106 Nucleated Blood Cells (mean f SEM)
Tissue
Erythroid
Abelson
- Epo + Epo
Harvey
-EP + Epo
Lymphoid
Abelson
- Epo + Epo
Undifferentiated
Harvey
- Epo +Epo
Day 9 Total 6.8 f 1.2 11.8 f 1.8 1.7 f 0.7 50.4 f 4.6 <0.2 f 0.2 <0.2 f 0.2 16.3 k 2.4 7.0 f 1.6 Embryo
Day 10 Extra- 16.5 f 2.6 30.5 f 3.9 0.5 f 0.5 335.0 f 14.6 <0.5 f 0.5 <0.5 -c 0.5 142.0 + 6.6 7.5 f 1.8 embryonic
Day 18-19 <0.2 f 0.2 3.4 f 0.8 1.6 f 0.5 32.9 f 2.8 254.0 f 6.7 272.0 f 8.5 <0.2 f 0.2 <0.3 f 0.3 Fetal Liver
Cell suspensions were prepared from day 9 total embryos (embryonic and extraembryonic tissues), from day 10 extraembryonic tissues (yolk sac, placenta, and decidua), or from day I8-day 19 fetal livers. The cells were infected with A-MuLV-P160 or Ha-MS/ and plated in soft-agar cultures in the presence or absence of 0.2 U/ml Epo. The number and color of macroscopic colonies were scored 9 days after plating. Histological examina- tion showed that three distinct types of colonies were present. Red or pink colonies were composed predominantly of cells in all stages of erythroid differentiation. White colonies induced by A-MuLV were exclusively lymphoid, whereas white colonies induced by Ha-MSV were composed predominantly of undifferentiated blasts or nonhemoglobinized pro-erythroblasts. Results are expressed as the number of colonies observed per 10s nbc f SEM. Values indicated as (<) represent the minimum level of detection in cultures where no colonies were observed.
fected cells were plated both in the presence and absence of 0.2 U/ml of Epo, a dose of hormone near the saturation level for stimulation of Ha-MSV-induced erythroid colonies (Hankins and Scolnick, 1981; Waneck and Rosenberg, unpublished data). As expected (Waneck and Rosenberg, 1981) A-MuLV induced large numbers of erythroid colo- nies and no lymphoid colonies when early embryonic cells were infected. The number of colonies observed in- creased slightly when Epo was present in the culture medium (Table l), and a higher percentage of these colo- nies were a very dark red in color. Examination of six of these colonies with Benzidine-Giemsa stain revealed that they contained 81% f 8% benzidine-positive cells in con- trast to the 69% + 3% benzidine-positive cells observed in a sample of 18 colonies grown in the absence of Epo. Thus, the difference in colony color was due to an in- creased number of hemoglobin (Hb)-positive cells in the colonies grown in the presence of Epo.
Ha-MN also induced colonies following infection of early fetal cells (Table 1). In contrast to A-MuLV-infected cultures, there was a dramatic increase in the total num- ber of colonies and a shift in colony type from white (un- differentiated) to red (erythroid) when Epo was added. Ex- amination of 18 colonies grown in the presence of Epo revealed that 60% f 5% of the cells in each colony were Hb-positive. In 18 colonies grown in the absence of Epo only 4% + 1% of the ceils in each colony were hemo- globinized. Most of the nonhemoglobinized cells were un- differentiated blasts that were morphologically similar to those seen in A-MuLV-induced erythroid colonies.
To demonstrate that the colonies observed following Ha-MSV infection contained Ha-MSV, individual colonies were stained using a monoclonal rat anti-P21 antibody in combination with a fluorescein-conjugated rabbit anti-rat immunoglobulin antibody. A total of 52 colonies (over 2700 cells) grown in the presence and absence of Epo were ex- amined and in each instance, greater than 97% of the cells were stained. Control preparations treated with
fluoresceinated antibody alone, or similar cells infected with A-MuLV and reacted with both antibodies did not stain.
Examination of various gestational time points revealed that erythroid target cells for both viruses appear between day 8 and day 9 of gestation and become more frequent through day 11 (Table 1 and data not shown). After this time, the number of A-MuLV erythroid target cells de- creases (Waneck and Rosenberg, 1981) and the few re- maining at late gestational times are revealed only when the infected cells are plated in the presence of Epo. The number of Ha-MSV-induced colonies also decreases throughout gestation, with the frequencies shown for day 18-19 being representative of later gestation tissues. Thus, the erythroid target cells for both viruses show a similar distribution during fetal development. This pattern was also found when tissues from C57BU6 mice were ex- amined indicating that the Fv-2 gene (Lilly, 1970; Odaka, 1970) a host gene that controls susceptibility to Friend vi- rus induced erythroid colonies (W. D. Hankins, personal communication) does not affect the ability of either Ha- MSV or A-MuLV to induce erythroid colonies (data not shown).
The quantitative differences in target cell frequencies among the fetal tissues can only be compared for a given virus. In order to detect the maximum number of colonies possible, both virus stocks were used undiluted in these experiments. The Ha-MSV stocks used in the study shown in Table 1 had a titer of transforming virus lo-fold higher than the A-MuLV stocks used. When Ha-MSV stocks were diluted to equal the maximum fibroblast transforming titer of A-MuLV stocks, both produced about the same number of colonies in cultures of infected fetal cells. Other experi- ments, using a high titer A-MuLV stock not available when the time course studies were conducted, demonstrated that both A-MuLV and Ha-MSV induced comparable num- bers of erythroid colonies when infected 12 day gestation fetal liver cells were plated in the presence of 0.2 U/ml Epo
Abelson Virus-Induced Differentiation 339
E A I 2 3 4 5 6 7 8 9 IO II 12
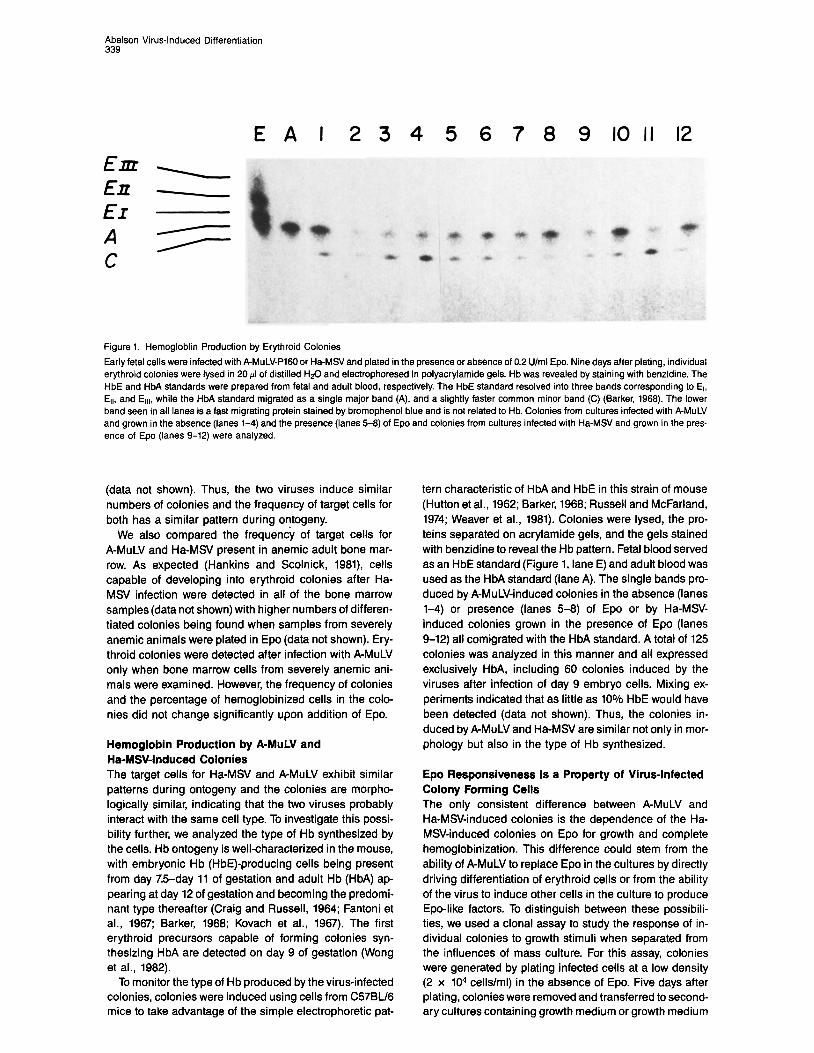
Figure 1. Hemogloblin Production by Erythroid Colonies
Early fetal cells were infected with A-MuLV-PI60 or Ha-MSV and plated in the presence or absence of 0.2 U/ml Epo. Nine days after plating, individual erythroid colonies were lysed in 20 ~1 of distilled HsO and electrophoresed in polyacrylamide gels. Hb was revealed by staining with benzidine. The HbE and HbA standards were prepared from fetal and adult blood, respectively. The HbE standard resolved into three bands corresponding to El, Err, and El,,, while the HbA standard migrated as a single major band (A), and a slightly faster common minor band (C) (Barker, 1966). The lower band seen in all lanes is a fast migrating protein stained by bromophenol blue and is not related to Hb. Colonies from cultures infected with A-MuLV and grown in the absence (lanes 1-4) and the presence (lanes 5-6) of Epo and colonies from cultures infected with Ha-MSV and grown in the pres- ence of Epo (lanes 9-12) were analyzed
(data not shown). Thus, the two viruses induce similar numbers of colonies and the frequency of target cells for both has a similar pattern during ontogeny.
We also compared the frequency of target cells for A-MuLV and Ha-M!3 present in anemic adult bone mar- row. As expected (Hankins and Scolnick, 1981), cells capable of developing into erythroid colonies after Ha- MS/ infection were detected in all of the bone marrow samples (data not shown) with higher numbers of differen- tiated colonies being found when samples from severely anemic animals were plated in Epo (data not shown). Ery- throid colonies were detected after infection with A-MuLV only when bone marrow cells from severely anemic ani- mals were examined. However, the frequency of colonies and the percentage of hemoglobinized cells in the colo- nies did not change significantly upon addition of Epo.
Hemoglobin Production by RMuLV and Ha-MS/-Induced Colonies The target cells for Ha-MSV and A-MuLV exhibit similar patterns during ontogeny and the colonies are morpho- logically similar, indicating that the two viruses probably interact with the same cell type. To investigate this possi- bility further, we analyzed the type of Hb synthesized by the cells. Hb ontogeny is well-characterized in the mouse, with embryonic Hb (HbE)-producing cells being present from day 7.5-day 11 of gestation and adult Hb (HM) ap- pearing at day 12 of gestation and becoming the predomi- nant type thereafter (Craig and Russell, 1964; Fantoni et al., 1967; Barker, 1968; Kovach et al., 1967). The first erythroid precursors capable of forming colonies syn- thesizing HbA are detected on day 9 of gestation (Wong et al., 1982).
To monitor the type of Hb produced by the virus-infected colonies, colonies were induced using cells from C57BU6 mice to take advantage of the simple electrophoretic pat-
tern characteristic of HbA and HbE in this strain of mouse (Hutton et al., 1962; Barker, 1968; Russell and McFarland, 1974; Weaver et al., 1981). Colonies were lysed, the pro- teins separated on acrylamide gels, and the gels stained with benzidine to reveal the Hb pattern. Fetal blood served as an HbE standard (Figure 1, lane E) and adult blood was used as the HbA standard (lane A). The single bands pro- duced by A-MuLV-induced colonies in the absence (lanes l-4) or presence (lanes 5-8) of Epo or by Ha-MSV- induced colonies grown in the presence of Epo (lanes 9-12) all comigrated with the HbA standard. A total of 125 colonies was analyzed in this manner and all expressed exclusively HM, including 60 colonies induced by the viruses after infection of day 9 embryo cells. Mixing ex- periments indicated that as little as 10% HbE would have been detected (data not shown). Thus, the colonies in- duced by A-MuLV and Ha-MSV are similar not only in mor- phology but also in the type of Hb synthesized.
Epo Responsiveness Is a Property of Virus-Infected Colony Forming Cells The only consistent difference between A-MuLV and Ha-MSV-induced colonies is the dependence of the Ha- MSV-induced colonies on Epo for growth and complete hemoglobinization. This difference could stem from the ability of A-MuLV to replace Epo in the cultures by directly driving differentiation of erythroid cells or from the ability of the virus to induce other cells in the culture to produce Epo-like factors. To distinguish between these possibili- ties, we used a clonal assay to study the response of in- dividual colonies to growth stimuli when separated from the influences of mass culture. For this assay, colonies were generated by plating infected cells at a low density (2 x lo4 cells/ml) in the absence of Epo. Five days after plating, colonies were removed and transferred to second- ary cultures containing growth medium or growth medium

Cell 340
100,
90-
SO-
TO-
SO-
50-
40-
30-
20-
IO-
OI
Virus A-M&V A-MuLV l-lo-f&V Ho-MSV
Condttion - to - -tot - to- - tot
Figure 2. Effect of Erythropoietin on Hemoglobinization of Virus- Induced Colonies
Cell suspensions from day 11-12 fetal liver were infected with A-MuLV- P160 or Ha-MSV and plated in the absence of Epo. Five days later, in- dividual colonies were transferred to secondary liquid cultures in the presence (- to +) or absence (- to -) of 0.4 U/ml Epo. Three days later the colonies were dispersed and centrifuged onto microscope slides. The cells were stained with Benzidine-Giemsa and evaluated under oil immersion at 1000x magnification. Each symbol represents the percentage of total benzidine-positive cells per colony. Open cir- cles, Balb/c-derived colonies, at least 100 cells evaluated; solid circles, C57BU6-derived colonies, at least 100 cells evaluated; open squares, Balbk-derived colonies, 20-99 cells evaluated; solid squares, C57BU6 colonies, 20-99 cells evaluated.
supplemented to contain 0.4 U/ml Epo. The colonies were allowed to grow for three more days and then stained with Benzidine-Giemsa and evaluated for cellularity and the frequency of cells synthesizing Hb.
When transferred A-MuLV-induced colonies were exam- ined by Benzidine-Giemsa staining, a high percentage of cells in the colonies synthesized Hb regardless of whether Epo was present after transfer (Figure 2). In contrast, very few cells in Ha-MSV-induced colonies synthesized Hb in the absence of Epo. However, when transferred into Epo- containing medium, the frequency of hemoglobinized cells was similar to that observed for A-MuLV-induced cells. These data indicate that once colonies are initiated, the Epo-independence of A-MuLV-induced colonies is probably due to a direct effect of the virus on the erythroid cell.
The slides prepared from the colonies were also scored for cellularity, as a measure of their growth and survival (Table 2). Most (54%-66%) of the preparations made from A-MuLV-induced colonies showed a high number of intact cells regardless of whether Epo was present. A similar percentage (56%~67%) of Ha-MSV-induced colonies con- tained high numbers of intact cells only if they were trans-
Table 2. Effect of Erythropoeitin on Colony Growth
Percent of Colonies Examined
Balblc c57BU6
Virus Cellularity - Epo + Epo - Epo + Epo
A-MuLV Dead (<lo) 24.0 24.0 25.0 29.1 Low (10-100) 12.0 8.0 16.7 16.7 High (>lOO) 64.0 68.0 58.3 54.2
Ha-WV Dead (<IO) 51.9 11.1 33.3 29.2 LOW (W-100) 29.6 22.2 58.4 12.5 High (>l 00) 18.5 86.7 8.3 58.3
Colonies that were scored for percentage of benzidine-positive cells (Figure 2) were also scored for the number of intact cells in the total preparation. Colonies were scored as dead if <lO cells were observed, low if 10-I 00 cells were observed and high if >lOO cells were observed. ln most cases, slides scored as high contained several hundred to several thousand cells. Results are expressed as the percent of colo- nies examined that fit into each category. Between 12 and 14 colo- nies were examined for each mouse strain and each culture condition. This method of quantitating colony growth and viability correlated close ly with observations of colony development made by visual inspection or using a phase contrast inverted microscope,
ferred into medium plus Epo; when transferred into medium minus Epo, few (8%-19%) of the colonies had large numbers of viable cells by day 8 after infection. Im- munofluorescent staining showed P21 in colonies trans- ferred from the cultures at day5 eliminating the possibility that many of these colonies were uninfected. Thus, it ap- pears that Ha-MS/ provides only an initial signal for colony growth and that unless a hormonal signal is pres- ent, many of the colonies die or do not proliferate.
Epo Is Not Produced in Mass Cultures of AMuLV-Infected Cells The colonies used in the transfer assay were allowed to develop for 5 days in mass cultures to insure that they were large enough to be distinguished from the normal micro- scopic colonies that were present in the cultures. Thus, factors produced either by virus-infected feeder cells or the colony-forming cells themselves during the first 5 days might have important effects on the colonies that were transferred.
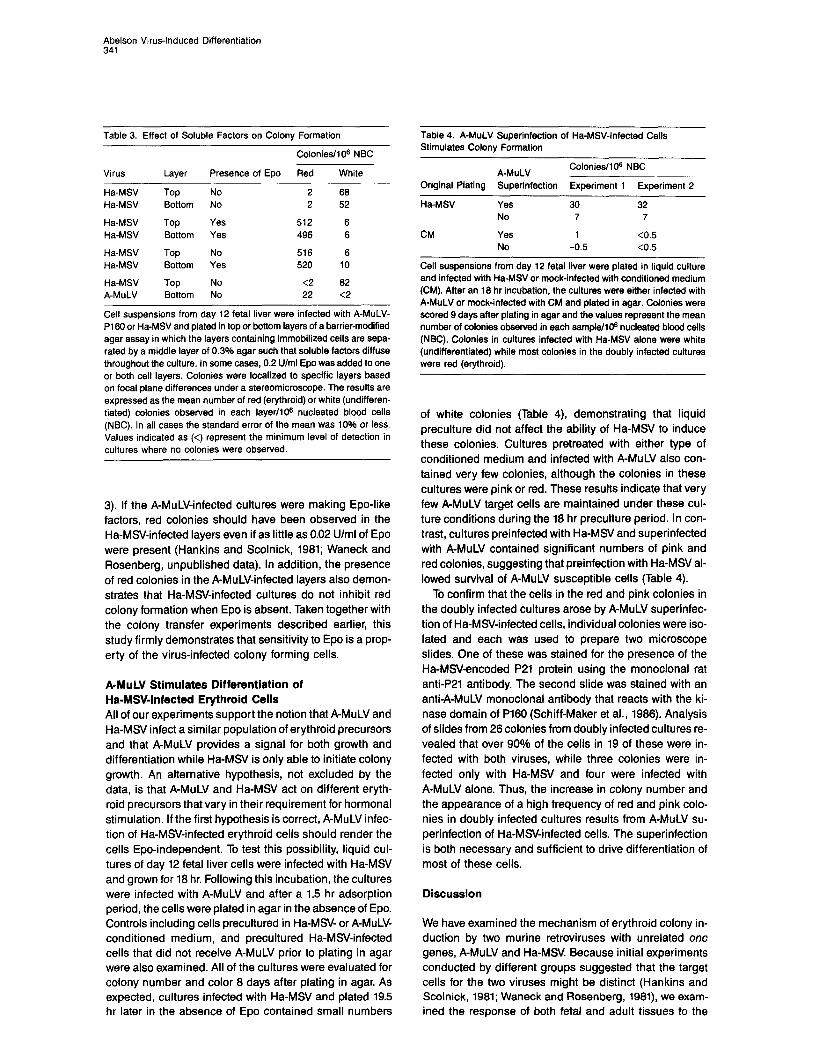
To test for soluble factors in A-MuLV-infected cultures, we developed a barrier-modified agar assay in which top and bottom layers of infected cells are separated by a mid- dle layer of agar medium alone, such that the cells are im- mobilized but soluble factors can diffuse throughout the culture. When this system is used with Ha-MSV-infected fetal cells, addition of Epo to only one cell layer is as effec- tive as addition to both layers in stimulating growth and differentiation of erythroid cells (Table 3).
To determine whether the apparent Epo-independence of A-MuLV-induced colonies was a result of feeder effects, we evaluated cultures that had Ha-MSV-infected cells in the top layer and A-MuLV-infected cells in the bottom layer. No exogenous Epo was added to these cultures. Under these conditions, no red colonies were observed in the Ha-MSV-infected layers while the expected number of red colonies developed in the A-MuLV-infected layers (Table
Abelson Virus-Induced Differentiation 341
Table 3. Effect of Soluble Factors on Colony Formation
Colonies/IO6 NBC
Virus Layer Presence of Epo Red White
Ha-MSV Ha-MSV
Ha-MSV Ha-MSV
Ha-MSV Ha-MSV
Ha-MSV A-MuLV
Top Bottom
Top Bottom
Top Bottom
Top Bottom
No No
Yes Yes
No Yes
No No
2 68 2 52
512 6 496 6
516 6 520 10
<2 62 22 <2
Cell suspensions from day 12 fetal liver were infected with A-MuLV- Pi 60 or Ha-MSV and plated in top or bottom layers of a barrier-modified agar assay in which the layers containing immobilized ceils are sepa- rated by a middle layer of 0.3% agar such that soluble factors diffuse throughout the culture. In some cases, 0.2 U/ml Epo was added to one or both cell layers. Colonies were localized to specific layers based on focal plane differences under a stereomicroscope. The results are expressed as the mean number of red (erythroid) or white (undifferen- tiated) colonies observed in each layer/IO@ nucleated blood cells (NBC). In all cases the standard error of the mean was 10% or less. Values indicated as (<) represent the minimum level of detection in cultures where no colonies were observed.
3). If the A-MuLV-infected cultures were making Epo-like factors, red colonies should have been observed in the Ha-MS/-infected layers even if as little as 0.02 U/ml of Epo were present (Hankins and Scolnick, 1981; Waneck and Rosenberg, unpublished data). In addition, the presence of red colonies in the A-MuLV-infected layers also demon- strates that Ha-MS/-infected cultures do not inhibit red colony formation when Epo is absent. Taken together with the colony transfer experiments described earlier, this study firmly demonstrates that sensitivity to Epo is a prop- erty of the virus-infected colony forming cells.
RMuLV Stimulates Differentiation of Ha-MSV-Infected Erythroid Cells All of our experiments support the notion that A-MuLV and Ha-MSV infect a similar population of erythroid precursors and that A-MuLV provides a signal for both growth and differentiation while Ha-MSV is only able to initiate colony growth. An alternative hypothesis, not excluded by the data, is that A-MuLV and Ha-MSV act on different eryth- roid precursors that vary in their requirement for hormonal stimulation. If the first hypothesis is correct, A-MuLV infec- tion of Ha-MSV-infected erythroid cells should render the cells Epo-independent. To test this possibility, liquid cul- tures of day 12 fetal liver cells were infected with Ha-MSV and grown for 18 hr. Following this incubation, the cultures were infected with A-MuLV and after a 1.5 hr adsorption period, the cells were plated in agar in the absence of Epo. Controls including cells precultured in Ha-MSV- or A-MuLV- conditioned medium, and precultured Ha-MSV-infected cells that did not receive A-MuLV prior to plating in agar were also examined. All of the cultures were evaluated for colony number and color 8 days after plating in agar. As expected, cultures infected with Ha-MSV and plated 19.5 hr later in the absence of Epo contained small numbers
Table 4. A-MuLV Superinfection of Ha-MSV-Infected Cells Stimulates Colony Formation
A-MuLV Colonies/106 NBC
Original Plating Superinfection Experiment 1 Experiment 2
Ha-MSV
CM
Yes No
Yes No
30 32 7 7
1 <0.5 -0.5 <0.5
Cell suspensions from day 12 fetal liver were plated in liquid culture and infected with Ha-MSV or mock-infected with conditioned medium (CM). After an 18 hr incubation, the cultures were either infected with A-MuLV or mock-infected with CM and plated in agar. Colonies were scored 9 days after plating in agar and the values represent the mean number of colonies observed in each sampte/lOa nucleated blood cells (NBC). Colonies in cultures infected with Ha-MSV alone were white (undifferentiated) while most colonies in the doubly infected cultures were red (erythroid).
of white colonies (Table 4), demonstrating that liquid preculture did not affect the ability of Ha-MSV to induce these colonies. Cultures pretreated with either type of conditioned medium and infected with A-MuLV also con- tained very few colonies, although the colonies in these cultures were pink or red. These results indicate that very few A-MuLV target cells are maintained under these cul- ture conditions during the 18 hr preculture period. In con- trast, cultures preinfected with Ha-MSV and superinfected with A-MuLV contained significant numbers of pink and red colonies, suggesting that preinfection with Ha-MSV al- lowed survival of A-MuLV susceptible cells (Table 4).
To confirm that the cells in the red and pink colonies in the doubly infected cultures arose by A-MuLV superinfec- tion of Ha-MSV-infected cells, individual colonies were iso- lated and each was used to prepare two microscope slides. One of these was stained for the presence of the Ha-MSV-encoded P21 protein using the monoclonal rat anti-P21 antibody. The second slide was stained with an anti-A-MuLV monoclonal antibody that reacts with the ki- nase domain of P180 (Schiff-Maker et al., 1988). Analysis of slides from 28 colonies from doubly infected cultures re- vealed that over 90% of the cells in 19 of these were in- fected with both viruses, while three colonies were in- fected only with Ha-MSV and four were infected with A-MuLV alone. Thus, the increase in colony number and the appearance of a high frequency of red and pink colo- nies in doubly infected cultures results from A-MuLV su- perinfection of Ha-MSV-infected cells. The superinfection is both necessary and sufficient to drive differentiation of most of these cells.
Discussion
We have examined the mechanism of erythroid colony in- duction by two murine retroviruses with unrelated one genes, A-MuLV and Ha-MSV. Because initial experiments conducted by different groups suggested that the target cells for the two viruses might be distinct (Hankins and Scolnick, 1981; Waneck and Rosenberg, 1981), we exam- ined the response of both fetal and adult tissues to the

Cell 342
viruses under identical conditions. This study revealed that cells susceptible to both viruses appear between day 8 and day 9 of gestation and show a similar frequency pat- tern throughout gestation. The absence of target cells prior to this time, even though yolk-sac-derived erythroid precursors are abundant early on day 8 (Russell and Bernstein, 1966) indicates that neither virus can stimulate growth of these cells under our culture conditions. This is consistent with the finding that colonies induced by both viruses synthesize only HM. In fact, target cells for both viruses appear at the same point in gestation as erythroid precursors that give rise to normal HbA synthesizing erythroid cells, suggesting a relationship between these two types of cells (Wong et al., 1982).
Both the similar target cell distributions and an inability to separate the target cells for each virus using two differ- ent gradient separation techniques (Keyes and Rosen- berg, unpublished data) argue that the two viruses stimu- late similar, if not identical cell types. This interpretation is also consistent with the observation that the colonies in- duced by A-MuLV in the presence or absence of Epo and those induced by Ha-MSV in the presence of Epo are strik- ingly similar. Not only is the gross morphology of the colo- nies similar, but all contain about l-2 x lo5 cells and mature between days 7-10 of culture. Also, all of the colo- nies synthesize HM. Histological examination of the cells reveals a population of cells in all stages of terminal erythroid differentiation. Normal erythroid colony forming cells are characterized according to these parameters with colony morphology, size, and time of maturation be- ing key criteria to identify similar precursors. Using these parameters, the colonies induced by both of these viruses resemble normal late BFU-E (Eaves et al., 1979), suggest- ing that the target cell for the viruses may be related to this normal cell type.
Epo-sensitivity is the only parameter that distinguishes the A-MuLV- and Ha-MSV-induced colonies. Addition of Epo to Ha-MSV-infected cells enhances both colony num- ber and the frequency of differentiated cells in the colony. In contrast, a slight increase in A-MuLV-induced colonies is observed with little change in the frequency of benzi- dine-positive cells. The transfer experiments identify the Epo-dependency of Ha-MSV-infected cells as a property of the virus-infected cells, and the barrier plating experi- ments eliminate the possibility that either A-MuLV-infected erythroid cells or other cells in the culture produce Epo or Epo-like factors that might function early after colony initi- ation. Thus, A-MuLV-induced colonies are Epo-indepen- dent while Ha-MSV-induced colonies require the hormone for both growth and differentiation.
Although the Epo used in this study is a typical commer- cially available preparation, it is only partially purified. Many of these preparations also contain burst promoting activity (BPA), a potent stimulator of erythroid precursors (Iscove, 1978; lscove et al., 1980); whether this material is present in our Epo lots and may be responsible for some of the effects observed requires further study. The fact that Epo-containing cultures support slightly higher num- bers of lymphoid colonies (unpublished data) also indi-
cates that the Epo preparation may contain several growth factors.
Although all of the evidence discussed so far supports the notion that the one gene products encoded by the two viruses influence the growth and differentiation of eryth- roid cell in different ways, direct evidence for this is pre- sented by the superinfection experiments. These experi- ments show that A-MuLV is capable of stimulating the growth and differentiation of Ha-MSV-initiated colonies in the absence of Epo. Thus, A-MuLV is able to provide a sig- nal that either mimics that of Epo or overrides the cell’s re- quirement for the normal hormonal stimulation. Other groups have recently shown that A-MuLV infection of other types of growth-factor-dependent cells removes the re- quirement for growth factor (Cook et al., 1985; Pierce et al., 1985; Oliff et al., 1985). In addition, in the one instance tested, Ha-MSV was not able to substitute for growth factor (Oliff et al., 1985). Whether A-MuLV is acting in a similar fashion in all of these instances remains to be determined. However, since interleukin 3 is the growth factor as- sociated with each of these responses and this material is the same as BPA (Iscove et al., 1982), a factor often present in commercial Epo preparations similar to those used in these experiments, it is possible that a common pathway is involved.
The role of Ha-MSV in initiating the colonies is not com- pletely clear. The superinfection experiments, as well as the ObSeNatiOn that Epo addition can be delayed for at least 5 days after infection (unpublished data) indicates that Ha-MSV is capable of initiating colony development. These data suggest that at least two signals are required for complete colony maturation and that Epo is only re- quired as a second signal. Thus, the action of theras gene product is sufficient to stimulate transient growth, but it can not provide functions needed to sustain growth or drive differentiation while the abl gene product is able to carry out both of these functions. Because no cell lines have been derived from colonies infected with either virus and even large numbers of cells fail to form tumors in mice (unpublished data), neither virus appears to be capable of immortalizing these cells. Thus, the signal to grow provided by Ha-MSV is not sufficient to stimulate continu- ous growth. With A-MuLV, the signal to grow may be too weak to overcome the signal to differentiate. Further ex- amination of the virus-cell interactions in the erythroid cell system should shed light on the interplay of one gene products and normal signals for growth and differenti- ation.
Experimental Procedures
Cells and Viruses The conditions for growing cells and preparing A-MuLV stocks have been described previously (Rosenberg and Baltimore, 1978; Rosen- berg and Witte, 1980). Ha-MSV stocks were prepared from a producer cell line, derived bysuperinfection of the Ha-S21 clonallyderived trans- formed nonproducer cell line with M-MuLV, clone 2 (Fan and Paskind, 1974; Chang et al., 1980; Hankins and Scolnick, 1981). These cells were a gift of Dr. W. D. Hankins.
Conditioned media (CM) were prepared as described elsewhere (Waneck and Rosenberg, 1981). The Ha-MSV nonproducer cell line

Abelson Virus-Induced Differentiation 343
used for this purpose was prepared by infecting NIH13T3 ceils at a mul- tiplicity of 0.3 Ha-MSV focus forming units and cloning the cells in agar immediately after virus adsorption. Nonproducer colonies were identi- fied by the absence of infectious M-MuLV and Ha-MSV in the culture fluid. The presence of Ha-MSV in the transformed clones was con- firmed by rescuing transforming virus with M-MuLV
Tissue Suspensions and Colony Assays Balblc and C57BU6 mice from our colony at the Tufts Cancer Center were used in all experiments. Fetal and adult cell suspensions were prepared and infected as described (Rosenberg and Baltimore, 1976; Waneck and Rosenberg, 1961). Anemic adult marrow cells were pre- pared from mice that had received intravenous injections of phenyl- hydrazine HCI (0.02, 0.04, or 0.06 mg/g body weight) at 64, 46, and 40 hr prior to use. In some samples, sheep Epo Step Ill (Connaught Laboratories) were added to the plating mixture. Cultures were scored by counting macroscopic colonies 9 days after infection.
In all experiments, erythroid colony formation required the presence of infectious transforming virus. Erythroid colonies could not be in- duced by M-MuLV even when the cells were plated in 1.6 U/ml Epo. Conditioned medium from either A-MuLV or Ha-MSV nonproducer cell lines also did not induce erythroid colonies even when Epo was added to the culture. All assays were linear with respect to virus dose and cell number over the range of 5 x 103-2 x lo5 cells/ml plated in both the presence and absence of Epo (data not shown).
For some experiments, modified agar assays were used. For the barrier modified agar assay, cells were plated in 0.5 ml top and bottom agar layers containing the standard agar plating medium (Rosenberg and Baltimore, 1976) separated by a 0.5 ml middle layer of 03% agar adjusted to contain the same concentration of medium and serum as the top and bottom layers. Colonies were localized to specific layers based on the focal plane differences observed using an American Op- tical stereomicroscope. For colony transfer assays, individual colonies were removed from agar 5 days after plating 2 x lo4 cells/ml in 35 mm plates. The colonies were identified using a dissecting micro- scope and transferred into U-bottom microtiter plates containing RPM1 1640 medium supplemented to contain 20% fetal calf serum and 5 x IO-5 M 2-mercapto-ethanol. Some of the wells were supplemented to contain 0.4 U/ml Epo. The colonies were harvested 3 days after transfer.
fiistologic and lmmunofluorescence Studies For Benzidine-Giemsa staining, colonies that had been centrifuged onto microscope slides were air-dried at least 4 hr, fixed in absolute methanol, stained with 22% benzidine, and counterstained with stan- dard Giemsa solution according to the procedure of Borsook et al. (1969).
For immunofluorescent studies with anti-P21 antibody, the proce- dure of Willingham et al. (1960) was used. The cells were reacted with an anti-P21 monoclonal antibody (a gift of Drs. M. Furth and E. Scol- nick) followed by fluorescein-conjugated rabbit anti-rat immunoglobu- lin (Cappel Laboratories). For staining of the P160 v-abl protein, cells were fixed in 3.7% formaldehyde in phosphate buffered saline for 20 min at room temperature and reacted with an anti-PI60 monoclonal an- tibody (Schiff-Maker et al., 1986) followed by a fluorescein-conjugated rabbit anti-mouse immunoglobulin antibody (Cappel Laboratories). All samples were examined under oil immersion using a Nikon Labophot microscope with ‘an epifluorescent light source.
Hemogloblin Separation Hb was analyzed by polyacrylamide gel electrophoresis as described by Brotherton et al. (1979). HbE and HbA standards were prepared by hemolysis of peripheral blood from C57BU6 12 day fetuses and adults respectively. Individual colonies were prepared for analysis by lysis in distilled water followed by freezing and thawing. Hemolysates were mixed 1:l with sample buffer and electrophoresed in slab gels at 30-35 mA for 3.5 hr at 4OC. Hb bands were detected by staining with benzidine reagent according to the procedure of Clarke (1979).
Acknowledgments
This work was supported by grants from the National Cancer Insti-
tute. N. R. is the recipient of a Research Career Development Award from the National Cancer Institute.
The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received September 11, 1985; revised October 23, 1985
References
Barker, J. E. (1968). Development of the mouse hematopoietic system. I. Types of hemoglobin produced in embryonic yolk sac and liver. Dev. Biol. 18, 14-29.
Borsook, H., Ratner, K., and Tattrie, B. (1969). Studies on erythropoie- sis. II. A method of segregating immature from mature adult rabbit erythroblasts. Blood 34, 32-41.
Brotherton, T W., Chui, D. H. K., Gauldie, J., and Patterson, M. (1979). Hemoglobin ontogeny during normal mouse fetal development. Proc. Natl. Acad. Sci. USA 76, 2853-2857.
Chang, E. H., Maryak, J. M., Wei, C. M., Shih, T. Y., Shober, R., Cheung, H. L., Ellis, R. W., Hager, G. L., Scolnick, E. M., and Lowy, D. R. (1980). Functional organization of the Harvey murine sarcoma vi- rus genome. J. Virol. 35, 76-92.
Clarke, B. J., Nathan, D. G., Alter, 13. P., Forget, B. G., Hillman, D., and Housman, D. (1979). Hemoglobin synthesis in human BFU-E and CFU- E-derived erythroid colonies. Blood 54, 805-817.
Cook, W. D., Metcalf, D., Nicola, N. A., Burgess, A. W., and Walker, F. (1985). Malignant transformation of a growth factor-dependent myeloid by Abelson virus without evidence of an autocrine mechanism. Cell 47, 677-683.
Craig, M. L., and Russell, E. S. (1964). A developmental change in hemoglobins correlated with an embryonic red cell population in the mouse. Dev. Biol. 76, 331-340.
Eaves, C. J., Humphries, R. K., and Eaves, A. C. (1979). In vitro charac- terization of erythroid precursor cells and the erythropoietic differentia- tion process. In Cellular and Molecular Regulation of Hemoglobin Switching, G. Stamatoyannopoulos and A. W. Nienhuis, eds. (New York: Grune and Stratton), pp. 251-278.
Fan, H., and Paskind, M. P (1974). Measurement of the sequence com- plexity of cloned Maloney murine leukemia virus 60 to 70s RNA: evi- dence for a haploid genome. J. Virol. 74, 421-429.
Fantoni, A., Bank, A., and Marks, P A. (1967). Globin composition and synthesis of hemoglobins in developing fetal mice erythroid cells. Science 757, 1327-1329.
Gibbs, J. B., Sigal, I. S., Poe, M., and Scolnick, E. M. (1984). Intrinsic GTPase activity distinguishes normal and oncogenic ras P21 molecules. Proc. Natl. Acad. Sci. USA 87, 5704-5708.
Hankins, W. D., and Scolnick, E. M. (1981). Harvey and Kirsten sar- coma viruses promote the growth and differentiation of erythroid cells in vitro. Cell 26, 91-97.
Hankins, W. D., Kost, T. A., and Pragnell, I. 6. (1982). The myeloprolifer- ative sarcoma virus causes transformation of erythroid progenitor cells in vitro. Mol. Cell. Biol. 2, 138-146.
Hutton, J. J., Bishop, J., Schweet, R., and Russell, E. S. (1962). Hemo- globin inheritance in inbred mouse strains. I. Structural differences. Proc. Natl. Acad. Sci. USA 48, 1505-1513.
Iscove, N. N. (1978). Erythropoietin-independent stimulation of early erythropoiesis in adult marrow cultures by conditioned media from lectin-stimulated mouse spleen cells. In Hematopoietic Cell Differenti- ation, D. W. Golde, M. J. Cline, D. Metcalf, and C. F Fox, eds. (New York: Academic Press), pp. 37-52.
Iscove, N. N., Guilbert, L. J.. and Weyman, C. (1980). Complete replacement of serum in primary cultures of erythropoietin-dependent red cell precursors (CFU-E) by albumin, transferrin, iron, unsaturated fatty acid, lecithin and cholesterol. Exp. Cell Res. 726, 121-126.
Iscove, N. N., Roitsch, C. A., Williams, N., and Guilbert, L. J. (1982). Molecules stimulating early red cell, granulocyte, macrophage and

Cell 344
megakaryocyte precursors in culture: similarity in size, hydrophobicity, and charge. J. Cell. Physiol. Suppl. 7, 85-78.
Jainchill, J. L., Aaronson, S. A., and Todaro, G. J. (1989). Murine sar- coma a@ leukemia viruses: assay using clonal lines of contact- inhibited mouse cells. J. Virol. 4, 549-553.
Kovach, J. S., Marks, P A., Russell, E. S., and Epler, l-l. (1967). Erythroid cell development in fetal mice: ultrastructural characteristics and hemoglobin synthesis. J. Mol. Biol. 25, 131142.
Lilly, F (1970). Fv-2: identification and location of a second gene gov- erning the spleen focus response to Friend leukemia virus in mice. J. Natl. Cancer Inst. 45, 183-189.
McGrath, J. P., Capon, D. J., Goeddel, D. V., and Levinson, A. D. (1984). Comparative biochemical properties of normal and activated human ras P21 proteins. Nature 370, 644650.
Cdaka, T. (1970). Inheritance of susceptibility to Friend mouse leuke- mia virus. VII. Establishment of a resistent strain. Int. J. Cancer 6, 18-23.
Oliff, A., Agranovsky, O., McKinney, M. D., Murty, V. V. V. S., and Bauch- witz, R. (1985). Friend murine leukemia virus-immortalized myeloid cells are converted into tumorigenic cell lines by Abelson leukemia vi- rus. Proc. Nag. Acad. Sci. USA 82, 3308-3310.
Pierce, J. H., and Aaronson, S. A. (1982). Balb and Harvey murine sar- coma virus transformation of a novel lymphoid progenitor cell. J. Exp. Med. 156, 073-087
Pierce, J. H., Di Fiore, l? t?, Aaronson, S. A., Potter, M., Pumphrey, J., Scott, A., and Ihle, J. N. (1985). Neoplastic transformation of mast cells by Abelson-MuLV: abrogation of IL-3 dependence by a nonautocrine mechanism. Cell 47, 885-893.
Reynolds, F H. Jr., Sachs, T S., Doebagkar, D. M., and Stephenson, J. R. (1978). Cells productively transformed by Abelson murine leuke- mia virus express a high molecular weight polyprotein containing structural and nonstructural components. Proc. Natl. Acad. Sci. USA 75, 2874-2978.
Rosenberg, N., and Baltimore, D. (1976). A quantitative assay for trans- formation of bone marrow cells by Abelson murine leukemia virus. J. Exp. Med. 143, 1453-1483.
Rosenberg, N., and Baltimore, D. (1978). The effect of helper virus on Abelson virus-induced transformation of lymphoid cells. J. Exp. Med. 747, 1128-l 141.
Rosenberg, N., and Witte, 0. N. (1980). Abelson murine leukemiavirus mutants with alterations in the virus-specific P120 molecule. J. Virol. 33, 340-348.
Rosenberg, N., Baltimore, D., and Scher, C. D. (1975). In vitro transfor- mation of lymphoid cells by Abelson murine leukemiavirus. Proc. Natl. Acad. Sci. USA 72, 1932-1936.
Russell, E. S., and Bernstein, S. E. (1988). Blood and blood formation. In The Biology of the Laboratory Mouse, Second Edition, E. L. Green, ed. (New York: McGraw-Hill), pp. 351-372.
Russell, E. S., and McFarland, A. (1974). Genetics of mouse hemo- globins. Ann. N. Y. Acad. Sci. 247, 25-38.
Scher, C. D., and Siegler, R. (1975). Direct transformation of 3T3 cells by Abelson murine leukemia virus. Nature 253, 729-731.
Schiff-Maker, L., Burns, M. C., Konopka, J. B., Clark, S., Witte, 0. N., and Rosenberg, N. (1988). Monoclonal antibodiesspecificfor v-abland cab/ encoded molecules. J. Viral., in press.
Scolnick, E. M., Papageorge, A. G., and Shih, T. Y. (1979). Guanine nucleotide binding activity as an assay for the src protein of rat-derived murine sarcoma viruses. Proc. Natl. Acad. Sci. USA 76, 5355-5359.
Sweet, R. W., Yokoyama, S., Kamata, T., Feramisco, J. R., Rosenberg, M., and Gross, M. (1984). The product of fas is a GTPase and the T24 oncogenic mutant is deficient in this activity. Nature 377, 273-276.
Waneck, G. L., and Rosenberg, N. (1981). Abelson leukemia virus in- duces lymphoid and erythroid colonies in infected fetal cell cultures. Cell 26, 79-89.
Wang, J. Y J., Queen, C., and Baltimore, D. (1982). Expression of an Abelson murine leukemia virus-encoded protein in Escherichia co/i causes extensive phosphorylation of tyrosine residues. J. Biol. Chem. 257; 13,181-13,184.
Weaver, S., Comer, M. B., Jahn, C. L., Hutchinson, C. A. Ill, and Edgell, M. H. (1981). The adult pglobin genes of the “single” type mouse C57BI. Cell 24, 403-411.
Weiss, R. A., Teich, N., Varmus, H., and Coffin, J. (1982). RNA Tumor Viruses. (Cold Spring Harbor, New York: Cold Spring Harbor Labo- ratory).
Willingham, M. C., Pastan, I., Shih, T. Y., and Scolnick, E. M. (1980). Localization of the src gene product of the Harvey strain of MSV to the plasma membrane of transformed cells by electron microscopic im- munocytochemistry. Cell 19, 1005-1014.
Witte, 0. N., Rosenberg, N., Paskind, M., Shields, A., and Baltimore, D. (1978). Identification of an Abelson murine leukemia virus-encoded protein present in transformed fibroblast and lymphoid cells. Proc. Natl. Acad. Sci. USA 75, 2486-2492.
Witte, 0. N., Dasgupta, A., and Baltimore, D. (1960). Abelson murine leukeamia virus protein is phosphorylated in vitro to form phosphotyro- sine. Nature 283, 828-831.
Wong, F! M. C., Clarke, B. J., Carr, D. H., and Chui, D. H. K. (1982). Adult hemoglobins are synthesized in erythroid colonies in vitro derived from murine circulating hemopoeitic progenitor cells during embryonic de- velopment. Proc. Natl. Acad. Sci. USA 79, 2952-2956.