Embed Size (px)
Citation preview

Journal of the Neurological Sciences 325 (2013) 127–136
Contents lists available at SciVerse ScienceDirect
Journal of the Neurological Sciences
j ourna l homepage: www.e lsev ie r .com/ locate / jns
Acellular spinal cord scaffold seeded with mesenchymal stem cells promoteslong-distance axon regeneration and functional recovery in spinal cord injured rats
Jia Liu a,b,1, Jian Chen a,b,1, Bin Liu c, Cuilan Yang d, Denghui Xie a,b, Xiaochen Zheng a,b, Song Xu a,b,Tianyu Chen a,b, Liang Wang a,b, Zhongmin Zhang a,b, Xiaochun Bai a,b,d,⁎, Dadi Jin a,b,⁎⁎a Department of Orthopedics, the Third Affiliated Hospital of Southern Medical University, Guangzhou, Chinab Orthopedic Research Institute, Southern Medical University, Guangzhou, Chinac Department of Spine Surgery, The Third Affiliated Hospital, Sun Yat-sen University, Guangzhou, Chinad Department of Cell Biology, Southern Medical University, Guangzhou, China
⁎ Correspondence to: X. Bai, Department of Cell BiologyGuangzhou 510515, China. Tel.: +86 20 61648724; fax: +⁎⁎ Correspondence to: D. Jin, Department of Orthopedicof Southern Medical University, Guagnzhou 510665, Chi
E-mail addresses: [email protected] (X. Bai), nyo1 Jia Liu and Jian Chen contributed equally to this stu
0022-510X/$ – see front matter © 2012 Elsevier B.V. Alhttp://dx.doi.org/10.1016/j.jns.2012.11.022
a b s t r a c t
a r t i c l e i n f oArticle history:Received 27 May 2012Received in revised form 23 October 2012Accepted 27 November 2012Available online 11 January 2013
Keywords:Spinal cord injuryAcellular scaffoldsMesenchymal stem cell
The stem cell-based experimental therapies are partially successful for the recovery of spinal cord injury(SCI). Recently, acellular spinal cord (ASC) scaffolds which mimic native extracellular matrix (ECM) havebeen successfully prepared. This study aimed at investigating whether the spinal cord lesion gap could bebridged by implantation of bionic-designed ASC scaffold alone and seeded with human umbilical cordblood-derived mesenchymal stem cells (hUCB-MSCs) respectively, and their effects on functional improve-ment. A laterally hemisected SCI lesion was performed in adult Sprague–Dawley (SD) rats (n=36) andASC scaffolds seeded with or without hUCB-MSCs were implanted into the lesion immediately. All ratswere behaviorally tested using the Basso–Beattie–Bresnahan (BBB) test once a week for 8 weeks. Behavioralanalysis showed that there was significant locomotor recovery improvement in combined treatmentgroup (ASC scaffold and ASC scaffold+hUCB-MSCs) as compared with the SCI only group (pb0.01).5-Bromodeoxyuridine (Brdu)-labeled hUCB-MSCs could also be observed in the implanted ACS scaffoldtwo weeks after implantation. Moreover, host neural cells (mainly oligodendrocytes) were able to migrateinto the graft. Biotin–dextran–amine (BDA) tracing test demonstrated that myelinated axons successfullygrew into the graft and subsequently promoted axonal regeneration at lesion sites. This study provides evi-dence for the first time that ASC scaffold seeded with hUCB-MSCs is able to bridge a spinal cord cavity andpromote long-distance axon regeneration and functional recovery in SCI rats.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
The regeneration capacity of native spinal cord is limited. Spinalcord injury (SCI) often results in severe sensory and motor deficitsbeyond the level of injury because of extensive neuronal loss, acuteaxonal damage, demyelination, apoptosis and functional damage ofperipheral neurons [1,2]. Therapeutic application of stem cell-basedtreatment of SCI seems to be effective in overcoming injury by replac-ing damaged cells, providing a better conducive microenvironmentand establishing new synapses [3–6].
An attractive source of cell transplantation is umbilical cordblood-derived mesenchymal stem cells (UCB-MSCs) [7,8]. The studiesin vitro showed that UCB-MSCs can secrete a number of cytokines
, Southern Medical University,86 20 61648208.s, The Third Affiliated Hospitalna. Tel./fax: +86 20 [email protected] (D. Jin).dy.
l rights reserved.
that could be beneficial to SCI recovery [9]. Moreover, UCB-MSCsmaintain typical biological characteristics associated with stem cells[10], such as self-renewal potential [11], and multi-potential[12–14] including differentiating functional neural cells [15–19].Furthermore, UCB-MSCs have more advantages in comparison withadult sourced mesenchymal stem cells and embryonic stem cells.UCB-MSCs can be easily obtained [20,21], without ethically contro-versial, more primitive [22], higher proliferative capacity [23] andless expensive. Most importantly, UCB-MSCs exhibit strong immuno-suppressive and immunomodulatory properties, and play an impor-tant role in modifying graft-versus-host reactions during allogenictransplantations [24–26]. Recently much attention has been focusedon the therapeutic potential of hUCB-MSCs on the injury of centralnervous system [27,28], which have been xenografted in animalmodels of spinal cord injury and have displayed some positive effects[6,29–31]. Accordingly, owing to their primitive nature and ability todevelop into functional neural cells, hUCB-MSC subpopulation is avaluable tool to facilitate spinal cord repair [7,10].
Approaches of tissue engineering are designed to repair lost ordamaged tissue through the use of cellular transplantation and
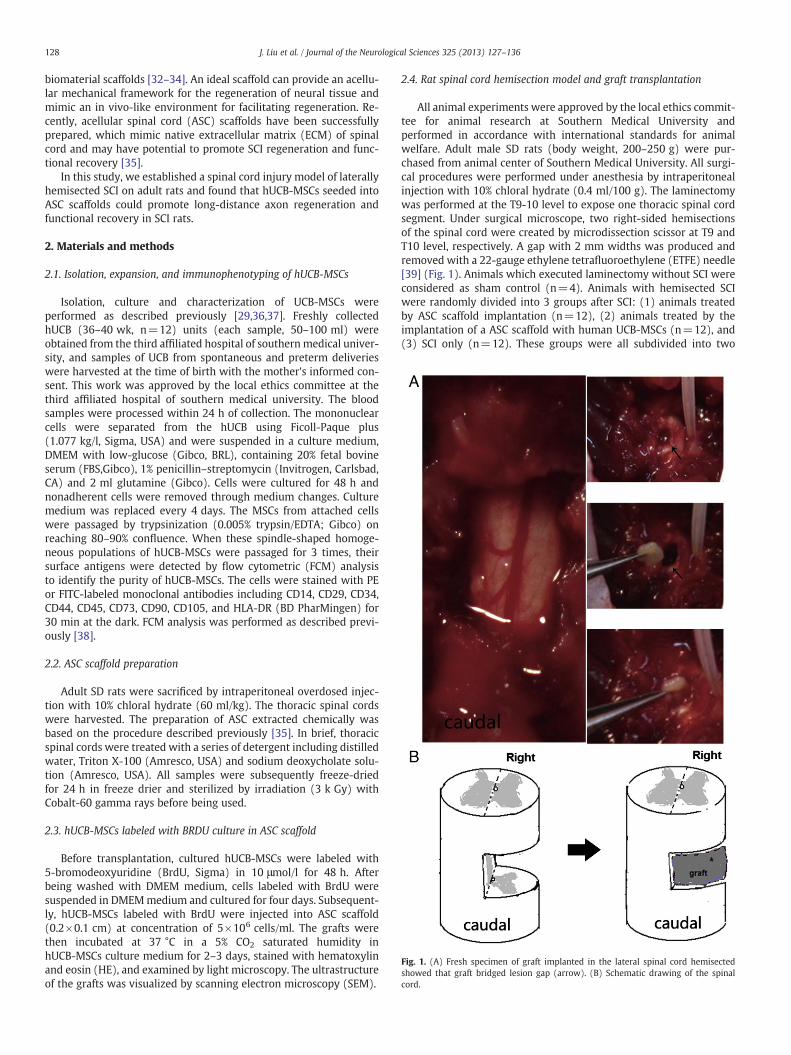
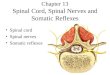
Fig. 1. (A) Fresh specimen of graft implanted in the lateral spinal cord hemisectedshowed that graft bridged lesion gap (arrow). (B) Schematic drawing of the spinalcord.
128 J. Liu et al. / Journal of the Neurological Sciences 325 (2013) 127–136
biomaterial scaffolds [32–34]. An ideal scaffold can provide an acellu-lar mechanical framework for the regeneration of neural tissue andmimic an in vivo-like environment for facilitating regeneration. Re-cently, acellular spinal cord (ASC) scaffolds have been successfullyprepared, which mimic native extracellular matrix (ECM) of spinalcord and may have potential to promote SCI regeneration and func-tional recovery [35].
In this study, we established a spinal cord injury model of laterallyhemisected SCI on adult rats and found that hUCB-MSCs seeded intoASC scaffolds could promote long-distance axon regeneration andfunctional recovery in SCI rats.
2. Materials and methods
2.1. Isolation, expansion, and immunophenotyping of hUCB-MSCs
Isolation, culture and characterization of UCB-MSCs wereperformed as described previously [29,36,37]. Freshly collectedhUCB (36–40 wk, n=12) units (each sample, 50–100 ml) wereobtained from the third affiliated hospital of southernmedical univer-sity, and samples of UCB from spontaneous and preterm deliverieswere harvested at the time of birth with the mother's informed con-sent. This work was approved by the local ethics committee at thethird affiliated hospital of southern medical university. The bloodsamples were processed within 24 h of collection. The mononuclearcells were separated from the hUCB using Ficoll-Paque plus(1.077 kg/l, Sigma, USA) and were suspended in a culture medium,DMEM with low-glucose (Gibco, BRL), containing 20% fetal bovineserum (FBS,Gibco), 1% penicillin–streptomycin (Invitrogen, Carlsbad,CA) and 2 ml glutamine (Gibco). Cells were cultured for 48 h andnonadherent cells were removed through medium changes. Culturemedium was replaced every 4 days. The MSCs from attached cellswere passaged by trypsinization (0.005% trypsin/EDTA; Gibco) onreaching 80–90% confluence. When these spindle-shaped homoge-neous populations of hUCB-MSCs were passaged for 3 times, theirsurface antigens were detected by flow cytometric (FCM) analysisto identify the purity of hUCB-MSCs. The cells were stained with PEor FITC-labeled monoclonal antibodies including CD14, CD29, CD34,CD44, CD45, CD73, CD90, CD105, and HLA-DR (BD PharMingen) for30 min at the dark. FCM analysis was performed as described previ-ously [38].
2.2. ASC scaffold preparation
Adult SD rats were sacrificed by intraperitoneal overdosed injec-tion with 10% chloral hydrate (60 ml/kg). The thoracic spinal cordswere harvested. The preparation of ASC extracted chemically wasbased on the procedure described previously [35]. In brief, thoracicspinal cords were treated with a series of detergent including distilledwater, Triton X-100 (Amresco, USA) and sodium deoxycholate solu-tion (Amresco, USA). All samples were subsequently freeze-driedfor 24 h in freeze drier and sterilized by irradiation (3 k Gy) withCobalt-60 gamma rays before being used.
2.3. hUCB-MSCs labeled with BRDU culture in ASC scaffold
Before transplantation, cultured hUCB-MSCs were labeled with5-bromodeoxyuridine (BrdU, Sigma) in 10 μmol/l for 48 h. Afterbeing washed with DMEM medium, cells labeled with BrdU weresuspended in DMEMmedium and cultured for four days. Subsequent-ly, hUCB-MSCs labeled with BrdU were injected into ASC scaffold(0.2×0.1 cm) at concentration of 5×106 cells/ml. The grafts werethen incubated at 37 °C in a 5% CO2 saturated humidity inhUCB-MSCs culture medium for 2–3 days, stained with hematoxylinand eosin (HE), and examined by light microscopy. The ultrastructureof the grafts was visualized by scanning electron microscopy (SEM).
2.4. Rat spinal cord hemisection model and graft transplantation
All animal experiments were approved by the local ethics commit-tee for animal research at Southern Medical University andperformed in accordance with international standards for animalwelfare. Adult male SD rats (body weight, 200–250 g) were pur-chased from animal center of Southern Medical University. All surgi-cal procedures were performed under anesthesia by intraperitonealinjection with 10% chloral hydrate (0.4 ml/100 g). The laminectomywas performed at the T9-10 level to expose one thoracic spinal cordsegment. Under surgical microscope, two right-sided hemisectionsof the spinal cord were created by microdissection scissor at T9 andT10 level, respectively. A gap with 2 mm widths was produced andremoved with a 22-gauge ethylene tetrafluoroethylene (ETFE) needle[39] (Fig. 1). Animals which executed laminectomy without SCI wereconsidered as sham control (n=4). Animals with hemisected SCIwere randomly divided into 3 groups after SCI: (1) animals treatedby ASC scaffold implantation (n=12), (2) animals treated by theimplantation of a ASC scaffold with human UCB-MSCs (n=12), and(3) SCI only (n=12). These groups were all subdivided into two
129J. Liu et al. / Journal of the Neurological Sciences 325 (2013) 127–136
subgroups each one including six animals and sacrificed on the 2ndand 8th weeks after surgery respectively. Neurologic symptomswere assessed weekly with the BBB locomotor rating scale beforeand after surgery. Histological examinations and immunohistochem-istry of neural cells were performed on all animals. Myelin basic pro-tein (MBP) staining and BDA anterograde tracing were used to detectmyelinated nerve fibers in animals 8 weeks after surgery. The musclelayers and skin were closed with suture. The animals were returnedto their cages with water and food made easily accessible. To preventinfection, rats were treated with ampicillin (100 mg/kg) and genta-micin (12 mg/kg) subcutaneously once a day following surgery for3 days. Manual bladder expression was performed twice a day untilthey regained bladder control, approximately for 3 to 5 days after ini-tial injury.
Fig. 2. (A) Primary cell colonies of hUCB-MSCs. (B) The third passage culture of hUCB-MSCB=100 μm).
2.5. Behavioral assessment after SCI
BBB scoring: A behavioral test was performed to measure thefunctional recovery of rats' right hind limb following the treatments[40], and results were scored in the range from 0 to 21. Individualrats were placed on an open field and observed for 4 min by two ob-servers blind to the treatments. The test was carried out once a weekafter transplantation up to the 8th week.
2.6. BDA anterograde tracing
Seven weeks after injury, the rats were anesthetized again andplaced on a stereotaxic frame. Two pieces of bone approximately3 mm×5 mm on each side of sutura sagittalis were removed using
s. (C) Immunophenotype of hUCB-MSCs (the third passage). (Scale bar: A=200 μm;
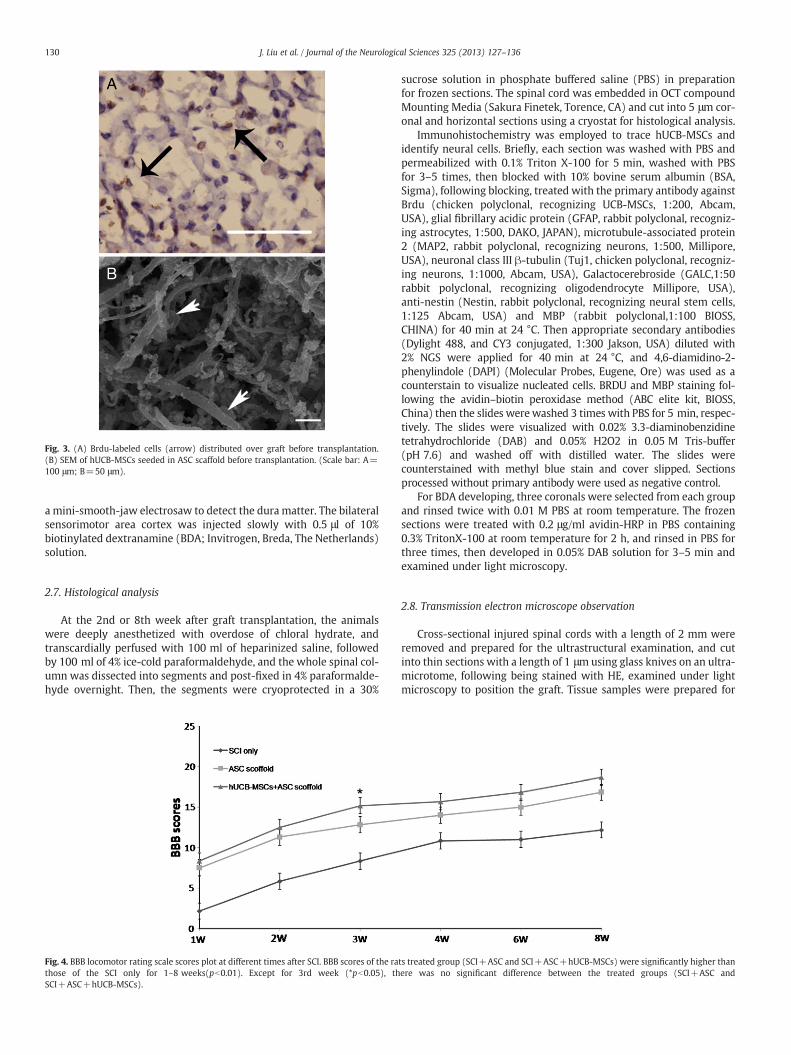
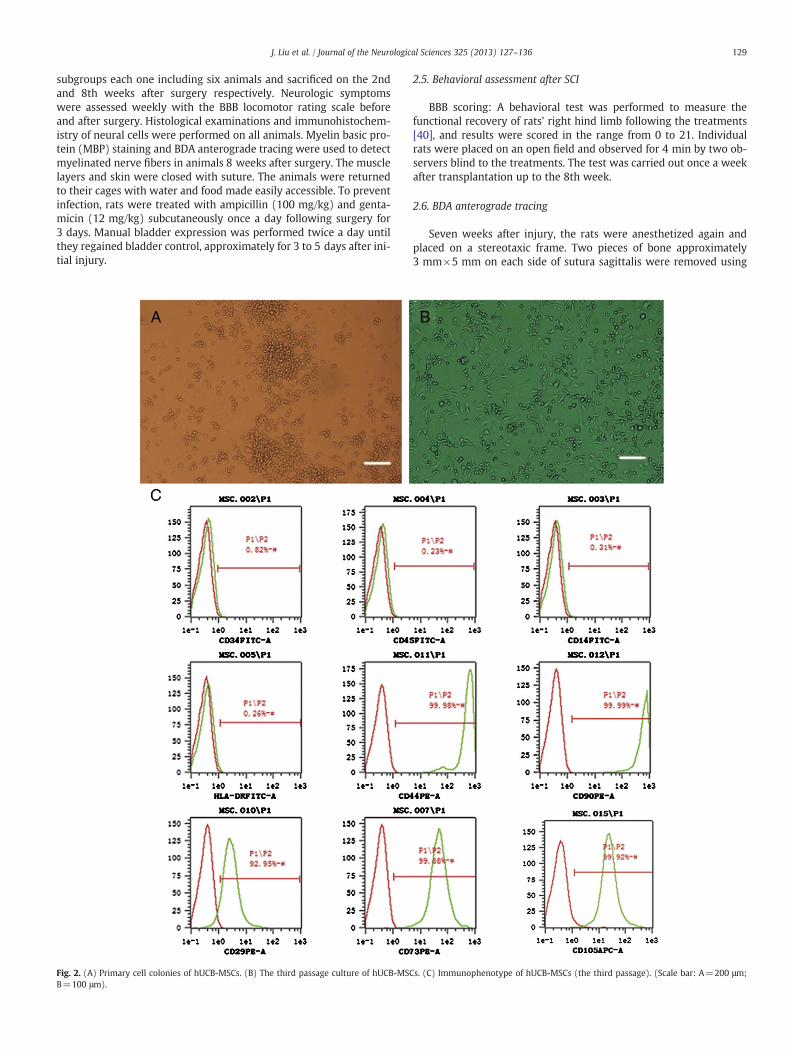
Fig. 3. (A) Brdu-labeled cells (arrow) distributed over graft before transplantation.(B) SEM of hUCB-MSCs seeded in ASC scaffold before transplantation. (Scale bar: A=100 μm; B=50 μm).
130 J. Liu et al. / Journal of the Neurological Sciences 325 (2013) 127–136
a mini-smooth-jaw electrosaw to detect the dura matter. The bilateralsensorimotor area cortex was injected slowly with 0.5 μl of 10%biotinylated dextranamine (BDA; Invitrogen, Breda, The Netherlands)solution.
2.7. Histological analysis
At the 2nd or 8th week after graft transplantation, the animalswere deeply anesthetized with overdose of chloral hydrate, andtranscardially perfused with 100 ml of heparinized saline, followedby 100 ml of 4% ice-cold paraformaldehyde, and the whole spinal col-umn was dissected into segments and post-fixed in 4% paraformalde-hyde overnight. Then, the segments were cryoprotected in a 30%
Fig. 4. BBB locomotor rating scale scores plot at different times after SCI. BBB scores of the rathose of the SCI only for 1–8 weeks(pb0.01). Except for 3rd week (*pb0.05), thSCI+ASC+hUCB-MSCs).
sucrose solution in phosphate buffered saline (PBS) in preparationfor frozen sections. The spinal cord was embedded in OCT compoundMounting Media (Sakura Finetek, Torence, CA) and cut into 5 μm cor-onal and horizontal sections using a cryostat for histological analysis.
Immunohistochemistry was employed to trace hUCB-MSCs andidentify neural cells. Briefly, each section was washed with PBS andpermeabilized with 0.1% Triton X-100 for 5 min, washed with PBSfor 3–5 times, then blocked with 10% bovine serum albumin (BSA,Sigma), following blocking, treated with the primary antibody againstBrdu (chicken polyclonal, recognizing UCB-MSCs, 1:200, Abcam,USA), glial fibrillary acidic protein (GFAP, rabbit polyclonal, recogniz-ing astrocytes, 1:500, DAKO, JAPAN), microtubule-associated protein2 (MAP2, rabbit polyclonal, recognizing neurons, 1:500, Millipore,USA), neuronal class III β-tubulin (Tuj1, chicken polyclonal, recogniz-ing neurons, 1:1000, Abcam, USA), Galactocerebroside (GALC,1:50rabbit polyclonal, recognizing oligodendrocyte Millipore, USA),anti-nestin (Nestin, rabbit polyclonal, recognizing neural stem cells,1:125 Abcam, USA) and MBP (rabbit polyclonal,1:100 BIOSS,CHINA) for 40 min at 24 °C. Then appropriate secondary antibodies(Dylight 488, and CY3 conjugated, 1:300 Jakson, USA) diluted with2% NGS were applied for 40 min at 24 °C, and 4,6-diamidino-2-phenylindole (DAPI) (Molecular Probes, Eugene, Ore) was used as acounterstain to visualize nucleated cells. BRDU and MBP staining fol-lowing the avidin–biotin peroxidase method (ABC elite kit, BIOSS,China) then the slides were washed 3 times with PBS for 5 min, respec-tively. The slides were visualized with 0.02% 3.3-diaminobenzidinetetrahydrochloride (DAB) and 0.05% H2O2 in 0.05 M Tris-buffer(pH 7.6) and washed off with distilled water. The slides werecounterstained with methyl blue stain and cover slipped. Sectionsprocessed without primary antibody were used as negative control.
For BDA developing, three coronals were selected from each groupand rinsed twice with 0.01 M PBS at room temperature. The frozensections were treated with 0.2 μg/ml avidin-HRP in PBS containing0.3% TritonX-100 at room temperature for 2 h, and rinsed in PBS forthree times, then developed in 0.05% DAB solution for 3–5 min andexamined under light microscopy.
2.8. Transmission electron microscope observation
Cross-sectional injured spinal cords with a length of 2 mm wereremoved and prepared for the ultrastructural examination, and cutinto thin sections with a length of 1 μmusing glass knives on an ultra-microtome, following being stained with HE, examined under lightmicroscopy to position the graft. Tissue samples were prepared for
ts treated group (SCI+ASC and SCI+ASC+hUCB-MSCs) were significantly higher thanere was no significant difference between the treated groups (SCI+ASC and
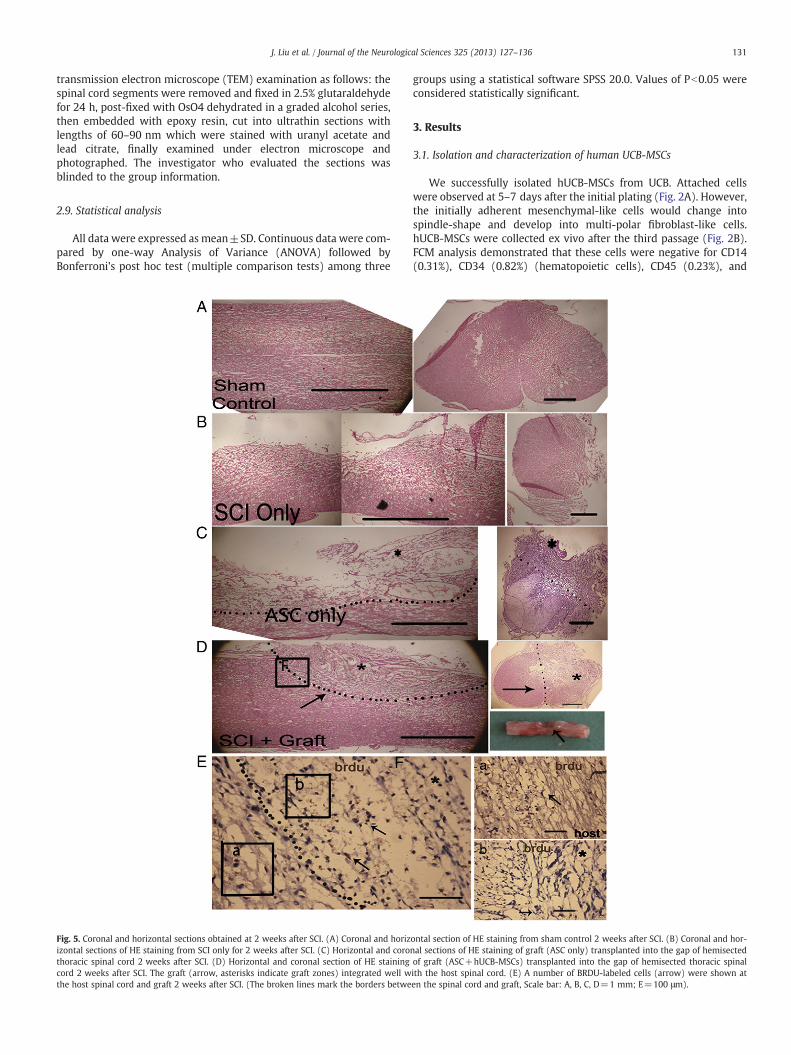
131J. Liu et al. / Journal of the Neurological Sciences 325 (2013) 127–136
transmission electron microscope (TEM) examination as follows: thespinal cord segments were removed and fixed in 2.5% glutaraldehydefor 24 h, post-fixed with OsO4 dehydrated in a graded alcohol series,then embedded with epoxy resin, cut into ultrathin sections withlengths of 60–90 nm which were stained with uranyl acetate andlead citrate, finally examined under electron microscope andphotographed. The investigator who evaluated the sections wasblinded to the group information.
2.9. Statistical analysis
All data were expressed as mean±SD. Continuous data were com-pared by one-way Analysis of Variance (ANOVA) followed byBonferroni's post hoc test (multiple comparison tests) among three
Fig. 5. Coronal and horizontal sections obtained at 2 weeks after SCI. (A) Coronal and horizizontal sections of HE staining from SCI only for 2 weeks after SCI. (C) Horizontal and corothoracic spinal cord 2 weeks after SCI. (D) Horizontal and coronal section of HE stainingcord 2 weeks after SCI. The graft (arrow, asterisks indicate graft zones) integrated well wthe host spinal cord and graft 2 weeks after SCI. (The broken lines mark the borders betwe
groups using a statistical software SPSS 20.0. Values of Pb0.05 wereconsidered statistically significant.
3. Results
3.1. Isolation and characterization of human UCB-MSCs
We successfully isolated hUCB-MSCs from UCB. Attached cellswere observed at 5–7 days after the initial plating (Fig. 2A). However,the initially adherent mesenchymal-like cells would change intospindle-shape and develop into multi-polar fibroblast-like cells.hUCB-MSCs were collected ex vivo after the third passage (Fig. 2B).FCM analysis demonstrated that these cells were negative for CD14(0.31%), CD34 (0.82%) (hematopoietic cells), CD45 (0.23%), and
ontal section of HE staining from sham control 2 weeks after SCI. (B) Coronal and hor-nal sections of HE staining of graft (ASC only) transplanted into the gap of hemisectedof graft (ASC+hUCB-MSCs) transplanted into the gap of hemisected thoracic spinalith the host spinal cord. (E) A number of BRDU-labeled cells (arrow) were shown aten the spinal cord and graft, Scale bar: A, B, C, D=1 mm; E=100 μm).

132 J. Liu et al. / Journal of the Neurological Sciences 325 (2013) 127–136
HLA-DR (0.26%), but positive for CD90 (99.99%), CD29 (92.95%), CD44(99.98%), CD105 (99.92%) and CD73 (99.88%) (Fig. 2C). Interestingly,the immunophenotypes of hUCB-MSCs were inconsistent with that ofMSC reported previously [7,14,41].
3.2. hUCB-MSCs culture in ASC scaffold
hUCB-MSCs were seeded into ASC scaffold and the grafts appearedin white color and displayed soft properties two days later.hUCB-MSC cells with positive Brdu were distributed in the networkof ECM in a cross-section before use (Fig. 3A). Under SEM, ASC scaf-fold was made up of thin fibers forming uniform mesh pores, onwhich was adhered by hUCB-MSC cells (Fig. 3B).
3.3. Locomotory behavior
All animals had normal motor function before SCI (BBB=21±0).After SCI, the right legs of animals were paraplegic (BBB=0.3±0.1).Over 8 weeks, motor function of all animals was gradually improvedso that they were able to move extensively all 3 joints of their hindlimbs. The animals were divided into 3 groups (SCI only, SCI+ASConly, and SCI+ASC+hUCB-MSCs). BBB scores of SCI+ASC andSCI+ASC+hUCB-MSCs group were significantly improved com-pared with SCI only group (pb0.01). In the 3rd week after operation,BBB scores of the animals treated with ASC seeded with hUCB-MSCswere significantly improved, compared with animals treated withASC only (pb0.05) (Fig. 4).
Fig. 6. Neural cells and Brdu positive cells distribution on graft (ASC+hUCB-MSCs) 2 weeks(C) Green-galc, red-BrdU, blue-DAPI. (D) Green-nestin, blue-DAPI. (Scale bar: A, B, C, D=1indicate graft zones).
3.4. Histological evaluation and immunohistochemical analysis
The spinal cord was removed at the 2nd and 8th weeks after trans-plantation in each group. Nonlesion spinal cord was in the sham controlgroup (Fig. 5A). In the SCI only group, HE staining showed that the lesioncompletely disrupted the dorsal and ventral of right lateral funiculus(Fig. 5B). In the SCI+ASC only and SCI+ASC+hUCB-MSCs groupsgross looking and HE staining showed that the graft was completelyfused in lesion gap (Fig. 5 C, D). A certain number of hUCB-MSC cellswith Brdu immunoreactivity survived on the graft and scattered in hostspinal cord in the 2nd week after transplantation (Fig. 5E). The stainedBrdu and neural makers could be detected in SCI+ASC+hUCB-MSCsgroup in the 2nd week after transplantation (Fig. 6).
Oligodendrocytes were the main type of neural cells distributed ongraft in the 8th week (SCI+ASC and SCI+ASC+hUCB-MSCs), whichsuggested that treatment with ASC scaffold promoted the infiltrationof oligodendrocytes into the lesion site (Fig. 7A, E). A few number ofneural stem cells and neurons could be observed on the graft (SCI+ASConly and SCI+ASC+hUCB-MSCs) (Fig. 7B, F). However, only few astro-cytes could be observed on the graft (SCI+ASC and SCI+ASC+hUCB-MSCs), which resulted in an astrocytes removed area (Fig. 7C, G).The remyelination of the sprouting axonswas visualized byMBP positivestaining. These results suggested that some remyelination of ingrownaxons appeared in the grafts (Fig. 7D, H).
3.5. Antergrade labeling of descending axons
Corticospinal tracts (CST) stained positively by BDA should exist inthe right funiculus of whitematter of spinal cord. Our research revealed
after SCI. (A) Green-gfap, red-BrdU, blue-DAPI. (B) Green-MAP2, red-BrdU, blue-DAPI.00 μm. The broken lines mark the borders between the spinal cord and graft, asterisks
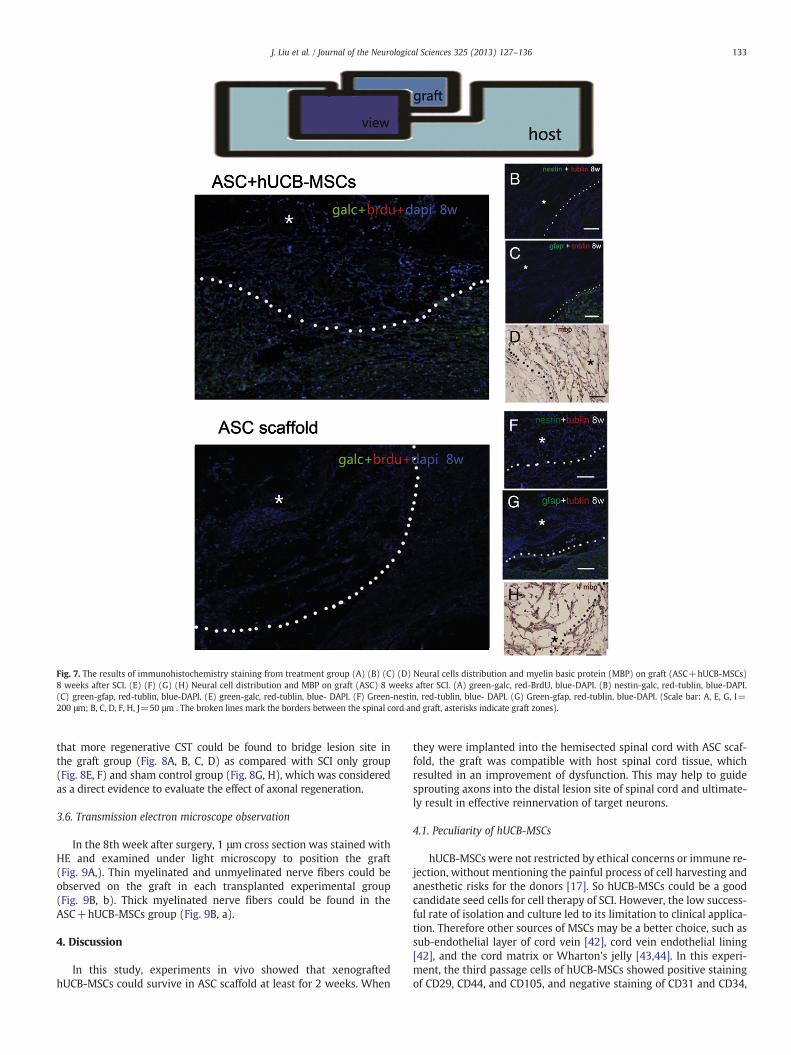
Fig. 7. The results of immunohistochemistry staining from treatment group (A) (B) (C) (D) Neural cells distribution and myelin basic protein (MBP) on graft (ASC+hUCB-MSCs)8 weeks after SCI. (E) (F) (G) (H) Neural cell distribution and MBP on graft (ASC) 8 weeks after SCI. (A) green-galc, red-BrdU, blue-DAPI. (B) nestin-galc, red-tublin, blue-DAPI.(C) green-gfap, red-tublin, blue-DAPI. (E) green-galc, red-tublin, blue- DAPI. (F) Green-nestin, red-tublin, blue- DAPI. (G) Green-gfap, red-tublin, blue-DAPI. (Scale bar: A, E, G, I=200 μm; B, C, D, F, H, J=50 μm . The broken lines mark the borders between the spinal cord and graft, asterisks indicate graft zones).
133J. Liu et al. / Journal of the Neurological Sciences 325 (2013) 127–136
that more regenerative CST could be found to bridge lesion site inthe graft group (Fig. 8A, B, C, D) as compared with SCI only group(Fig. 8E, F) and sham control group (Fig. 8G, H), which was consideredas a direct evidence to evaluate the effect of axonal regeneration.
3.6. Transmission electron microscope observation
In the 8th week after surgery, 1 μm cross section was stained withHE and examined under light microscopy to position the graft(Fig. 9A,). Thin myelinated and unmyelinated nerve fibers could beobserved on the graft in each transplanted experimental group(Fig. 9B, b). Thick myelinated nerve fibers could be found in theASC+hUCB-MSCs group (Fig. 9B, a).
4. Discussion
In this study, experiments in vivo showed that xenograftedhUCB-MSCs could survive in ASC scaffold at least for 2 weeks. When
they were implanted into the hemisected spinal cord with ASC scaf-fold, the graft was compatible with host spinal cord tissue, whichresulted in an improvement of dysfunction. This may help to guidesprouting axons into the distal lesion site of spinal cord and ultimate-ly result in effective reinnervation of target neurons.
4.1. Peculiarity of hUCB-MSCs
hUCB-MSCs were not restricted by ethical concerns or immune re-jection, without mentioning the painful process of cell harvesting andanesthetic risks for the donors [17]. So hUCB-MSCs could be a goodcandidate seed cells for cell therapy of SCI. However, the low success-ful rate of isolation and culture led to its limitation to clinical applica-tion. Therefore other sources of MSCs may be a better choice, such assub-endothelial layer of cord vein [42], cord vein endothelial lining[42], and the cord matrix or Wharton's jelly [43,44]. In this experi-ment, the third passage cells of hUCB-MSCs showed positive stainingof CD29, CD44, and CD105, and negative staining of CD31 and CD34,

Fig. 8. The results of BDA staining (arrow) in coronal sections. (A) (B) (C) (D): BDA staining (arrow) in coronal sections from treated group (ASC+hUCB-MSCs) showed that abun-dant of regenerative axons were found in graft. It seemed that axons choose shortcut way to bridge lesion site. (E) (F) BDA stained axons in ASC only group. (G) (H) BDA stainedaxons in SCI only group. (I) (J) BDA stained axons in sham control spinal cord. (Scale bar: A, E, G=200 μm; B, C, F=100 μm. D, H=40 μm. The asterisk represented the graft.)
134 J. Liu et al. / Journal of the Neurological Sciences 325 (2013) 127–136
which suggested the hUCB-MSCs cultured by our protocol expressedthe markers of MSCs [7,8].
4.2. ASC scaffold
As aforementioned, tissue engineering aimed at providing three-dimensional functional construction to serve as replacement tissue ororgan by combining cells with scaffolds. Much attention recently hasbeen focused on acellular scaffoldswhich retain the structural and func-tional proteins of extracellular matrix (ECM) and original three-dimensional (3D) distribution [32,35]. Accordingly, acellular scaffold isbecoming an ideal candidate for tissue engineering repair strategies.In this study, we showed that ASC scaffold had a beneficial effect onfunction recovery of SCI by providing an environment favorable
to host oligodendrocyte differentiation, proliferation and axonalremyelination, which promoted endogenous regeneration of neuralcells. Our results demonstrated the ASC scaffold in favor of host neuralinduction and conduction.
4.3. Cell fate of xenografted hUCB-MSCs on ASC scaffold in vivo
As compared with SCI only group, ASC scaffold only and hUCB-MSCsseeded into ASC scaffold had a gradually increased functional recoveryafter co-treatment for 8 weeks as measured by BBB scores. However,only a fewxenografted hUCB-MSCs survived for 2 weeks after transplan-tation and we failed to observe any neural differentiation [45]. It wassupposed that hUCB-MSCs might cause immune rejection whenxenogenically transplanted into nonimmune-suppressed host vertebral

Fig. 9. HE and TEM examination in treated group in the 8th week after SCI. (A) HE staining of graft in treated group. (B) Observation of graft in lesion site showed some of theregenerated axons and thin myelin sheath (arrow) distribute on graft. The integrity of myelin sheath (arrow) was better in ASC+hUCB-MSCs (a) than that in ASC scaffold (b).(Scale bar: A=100 μm; B=5 μm).
135J. Liu et al. / Journal of the Neurological Sciences 325 (2013) 127–136
column, but hUCB-MSCs had hypo-immunological features, such as theabsence of HLA class II expression and inhibition of T-cell response[24–26]. Our results showed that hUCB-MSCs migrated widely in thehost spinal cord, but neuronal replacement in quantity was far insuffi-cient to produce any significant therapeutic effect, which suggestedthat hUCB-MSCsmight play neurotrophic protective effects [9,46], possi-bly contributing to SCI recovery through the release of factors that re-duced secondary injury [28,47]. Moreover, treatment with hUCB-MSCsalso changed the cellular components in the lesion microenvironment,whichmay be a source of growth factors in the injured spinal cord. Final-ly, no xenografted hUCB-MSCs could be observed in the 8th week aftertransplantation, and this indicated the proliferation, long term survivaland maturation of xenografted cells in the host milieu were morechallenging.
4.4. Myelinate regenerating axons inside the implant
The improved locomotor function of animals with ASC scaffoldtransplanted with hUCB-MSCs could also be explained with thesemodels by providing a neurotrophic environment, especially mucholigodendrocyte in the astrocytes removed area, which was in favorof migration and proliferation of host neural cells, and significantremyelination was consistently observed. Our BDA anterograde trac-ing test directly proved that the disrupted nerve fibers had bypassingconnection, re-established ascending/descending tracts, and local syn-apses, thus released signals of the right column, which is importantfor retrieval behavior. In addition, interestingly, the re-establishedaxon tracts didn't lose their orientation in the graft but could chooseshortcut way to re-establish axons.
In conclusion, the temporary survival of xenografted hUCB-MSCs,which were seeded into ASC scaffold of injured spinal cord of rats wassufficient to achieve improved locomotor recovery, through providingneurotrophic protective effects and changing lesion microenvironment
to help host neural cells to regenerate. Therefore, this model might be apromisingway for repairing damaged spinal cords. In addition, we needto better understand the molecule mechanisms of reestablishingdisrupted nerve fiber in CNS environment.
Conflict of interest
The authors have no conflicts of interest.
Acknowledgments
This work was supported by National Natural Sciences Foundationof China (30900555, 30973036 and 91029727), and Program forChangjiang Scholars and Innovative Research Team in University(IRT1142).
References
[1] Rossi SL, Keirstead HS. Stem cells and spinal cord regeneration. Curr OpinBiotechnol 2009;20:552–62.
[2] Enzmann GU, Benton RL, Talbott JF, Cao Q, Whittemore SR. Functional consider-ations of stem cell transplantation therapy for spinal cord repair. J Neurotrauma2006;23:479–95.
[3] Ban DX, Kong XH, Feng SQ, Ning GZ, Chen JT, Guo SF. Intraspinal cord graft of au-tologous activated Schwann cells efficiently promotes axonal regeneration andfunctional recovery after rat's spinal cord injury. Brain Res 2009;1256:149–61.
[4] Cho SR, Yang MS, Yim SH, Park JH, Lee JE, Eom YW, et al. Neurally induced umbil-ical cord blood cells modestly repair injured spinal cords. Neuroreport 2008;19:1259–63.
[5] Dobkin BH. Recommendations for publishing case studies of cell transplantationfor spinal cord injury. Neurorehabil Neural Repair 2010;24:687–91.
[6] Kuh SU, Cho YE, Yoon DH, Kim KN, Ha Y. Functional recovery after human umbil-ical cord blood cells transplantation with brain-derived neutrophic factor into thespinal cord injured rat. Acta Neurochir 2005;147:985–92 [discussion 92].
[7] Arien-Zakay H, Lazarovici P, Nagler A. Tissue regeneration potential in humanumbilical cord blood. Best Pract Res Clin Haematol 2010;23:291–303.

136 J. Liu et al. / Journal of the Neurological Sciences 325 (2013) 127–136
[8] Malgieri A, Kantzari E, Patrizi MP, Gambardella S. Bone marrow and umbilicalcord blood human mesenchymal stem cells: state of the art. Int J Clin Exp Med2010;3:248–69.
[9] Neuhoff S, Moers J, Rieks M, Grunwald T, Jensen A, Dermietzel R, et al. Prolifera-tion, differentiation, and cytokine secretion of human umbilical cord blood-derived mononuclear cells in vitro. Exp Hematol 2007;35:1119–31.
[10] Hilfiker A, Kasper C, Hass R, Haverich A. Mesenchymal stem cells and progenitorcells in connective tissue engineering and regenerative medicine: is there a futurefor transplantation? Langenbecks Arch Surg/Dutsche Gesellschaft fur Chirurgie2011;396:489–97.
[11] Kern S, Eichler H, Stoeve J, Kluter H, Bieback K. Comparative analysis of mesenchy-mal stem cells from bone marrow, umbilical cord blood, or adipose tissue. StemCells 2006;24:1294–301.
[12] Lee OK, Kuo TK, Chen WM, Lee KD, Hsieh SL, Chen TH. Isolation of multipotentmesenchymal stem cells from umbilical cord blood. Blood 2004;103:1669–75.
[13] Wagner W, Wein F, Seckinger A, Frankhauser M, Wirkner U, Krause U, et al. Com-parative characteristics of mesenchymal stem cells from human bone marrow,adipose tissue, and umbilical cord blood. Exp Hematol 2005;33:1402–16.
[14] Montesinos JJ, Flores-Figueroa E, Castillo-Medina S, Flores-Guzman P, Hernandez-Estevez E, Fajardo-Orduna G, et al. Human mesenchymal stromal cells from adultandneonatal sources: comparative analysis of theirmorphology, immunophenotype,differentiation patterns and neural protein expression. Cytotherapy2009;11:163–76.
[15] Sun W, Buzanska L, Domanska-Janik K, Salvi RJ, Stachowiak MK. Voltage-sensitiveand ligand-gated channels in differentiating neural stem-like cells derived fromthe nonhematopoietic fraction of human umbilical cord blood. Stem Cells2005;23:931–45.
[16] Jin W, Xing YQ, Yang AH. Epidermal growth factor promotes the differentiation ofstem cells derived from human umbilical cord blood into neuron-like cells viataurine induction in vitro. In Vitro Cell Dev Biol Anim 2009;45:321–7.
[17] Lim JY, Park SI, Oh JH, Kim SM, Jeong CH, Jun JA, et al. Brain-derived neurotrophicfactor stimulates the neural differentiation of human umbilical cord blood-derivedmesenchymal stem cells and survival of differentiated cells through MAPK/ERKand PI3K/Akt-dependent signaling pathways. J Neurosci Res 2008;86:2168–78.
[18] Jeong JA, Gang EJ, Hong SH, Hwang SH, Kim SW, Yang IIH, et al. Rapid neural dif-ferentiation of human cord blood-derived mesenchymal stem cells. Neuroreport2004;15:1731–4.
[19] Lee MW, Moon YJ, Yang MS, Kim SK, Jang IK, Eom YW, et al. Neural differentiationof novel multipotent progenitor cells from cryopreserved human umbilical cordblood. Biochem Biophys Res Commun 2007;358:637–43.
[20] Can A, Karahuseyinoglu S. Concise review: human umbilical cord stroma with re-gard to the source of fetus-derived stem cells. Stem Cells 2007;25:2886–95.
[21] Weiss ML, Troyer DL. Stem cells in the umbilical cord. Stem Cell Rev 2006;2:155–62.
[22] Lu LL, Liu YJ, Yang SG, Zhao QJ, Wang X, Gong W, et al. Isolation and characteriza-tion of human umbilical cord mesenchymal stem cells with hematopoiesis-supportive function and other potentials. Haematologica 2006;91:1017–26.
[23] Moezzi L, Pourfathollah AA, Alimoghaddam K, Soleimani M, Ardjmand AR. The ef-fect of cryopreservation on clonogenic capacity and in vitro expansion potential ofumbilical cord blood progenitor cells. Transplant Proc 2005;37:4500–3.
[24] Glennie S, Soeiro I, Dyson PJ, Lam EW, Dazzi F. Bone marrow mesenchymal stemcells induce division arrest anergy of activated T cells. Blood 2005;105:2821–7.
[25] Kode JA, Mukherjee S, Joglekar MV, Hardikar AA. Mesenchymal stem cells:immunobiology and role in immunomodulation and tissue regeneration. Cytotherapy2009;11:377–91.
[26] Abumaree M, Al Jumah M, Pace RA, Kalionis B. Immunosuppressive Properties ofMesenchymal Stem Cells. Stem Cell Rev 2012;8(2):375–92.
[27] Low CB, Liou YC, Tang BL. Neural differentiation and potential use of stem cellsfrom the human umbilical cord for central nervous system transplantation thera-py. J Neurosci Res 2008;86:1670–9.
[28] Chua SJ, Bielecki R, Yamanaka N, Fehlings MG, Rogers IM, Casper RF. The effect ofumbilical cord blood cells on outcomes after experimental traumatic spinal cordinjury. Spine 2010;35:1520–6.
[29] Dasari VR, Spomar DG, Li L, Gujrati M, Rao JS, Dinh DH. Umbilical cord blood stemcell mediated downregulation of fas improves functional recovery of rats afterspinal cord injury. Neurochem Res 2008;33:134–49.
[30] Veeravalli KK, Dasari VR, Tsung AJ, Dinh DH, Gujrati M, Fassett D, et al. Humanumbilical cord blood stem cells upregulate matrix metalloproteinase-2 in ratsafter spinal cord injury. Neurobiol Dis 2009;36:200–12.
[31] Lee JH, Chung WH, Kang EH, Chung DJ, Choi CB, Chang HS, et al. Schwann cell-likeremyelination following transplantation of human umbilical cord blood(hUCB)-derived mesenchymal stem cells in dogs with acute spinal cord injury. JNeurol Sci 2011;300:86–96.
[32] Zhang XY, Xue H, Liu JM, Chen D. Chemically extracted acellular muscle: a newpotential scaffold for spinal cord injury repair. J Biomed Mater Res A 2012;100:578–87.
[33] Hejcl A, Sedy J, Kapcalova M, Toro DA, Amemori T, Lesny P, et al. HPMA-RGDhydrogels seeded with mesenchymal stem cells improve functional outcome inchronic spinal cord injury. Stem Cells Dev 2010;19:1535–46.
[34] Pritchard CD, Slotkin JR, Yu D, Dai H, Lawrence MS, Bronson RT, et al. Establishinga model spinal cord injury in the African green monkey for the preclinical evalu-ation of biodegradable polymer scaffolds seeded with human neural stem cells. JNeurosci Methods 2010;188:258–69.
[35] Guo SZ, Ren XJ, Wu B, Jiang T. Preparation of the acellular scaffold of the spinalcord and the study of biocompatibility. Spinal Cord 2010;48:576–81.
[36] Newcomb JD, Willing AE, Sanberg PR. Umbilical cord blood cells. Methods MolBiol 2009;549:119–36.
[37] Seo Y, Yang SR, Jee MK, Joo EK, Roh KH, Seo MS, et al. Human umbilical cordblood-derived mesenchymal stem cells protect against neuronal cell death andameliorate motor deficits in Niemann Pick type C1 mice. Cell Transplant2011;20:1033–47.
[38] Gang EJ, Jeong JA, Han S, Yan Q, Jeon CJ, Kim H. In vitro endothelial potential ofhuman UC blood-derived mesenchymal stem cells. Cytotherapy 2006;8:215–27.
[39] Itosaka H, Kuroda S, Shichinohe H, Yasuda H, Yano S, Kamei S, et al. Fibrin matrixprovides a suitable scaffold for bone marrow stromal cells transplanted into in-jured spinal cord: a novel material for CNS tissue engineering. Neuropathol Off JJpn Soc Neuropathol 2009;29:248–57.
[40] Basso DM, Beattie MS, Bresnahan JC. A sensitive and reliable locomotor ratingscale for open field testing in rats. J Neurotrauma 1995;12:1–21.
[41] Kogler G, Sensken S, Wernet P. Comparative generation and characterization ofpluripotent unrestricted somatic stem cells with mesenchymal stem cells fromhuman cord blood. Exp Hematol 2006;34:1589–95.
[42] Romanov YA, Svintsitskaya VA, Smirnov VN. Searching for alternative sources ofpostnatal human mesenchymal stem cells: candidate MSC-like cells from umbili-cal cord. Stem Cells 2003;21:105–10.
[43] Weiss ML, Mitchell KE, Hix JE, Medicetty S, El-Zarkouny SZ, Grieger D, et al. Trans-plantation of porcine umbilical cord matrix cells into the rat brain. Exp Neurol2003;182:288–99.
[44] Wang HS, Hung SC, Peng ST, Huang CC, Wei HM, Guo YJ, et al. Mesenchymal stemcells in the Wharton's jelly of the human umbilical cord. Stem Cells 2004;22:1330–7.
[45] Saporta S, Kim JJ, Willing AE, Fu ES, Davis CD, Sanberg PR. Human umbilical cordblood stem cells infusion in spinal cord injury: engraftment and beneficial influ-ence on behavior. J Hematother Stem Cell Res 2003;12:271–8.
[46] Liu CH, Hwang SM. Cytokine interactions in mesenchymal stem cells from cordblood. Cytokine 2005;32:270–9.
[47] NewmanMB, Willing AE, Manresa JJ, Sanberg CD, Sanberg PR. Cytokines producedby cultured human umbilical cord blood (HUCB) cells: implications for brain re-pair. Exp Neurol 2006;199:201–8.