Embed Size (px)
Citation preview

Acoustic transmission characteristics of the tympanal tracheae of bushcrickets (Tettigoniidae). II: Comparative studies of the tracheae of seven species
Ralf Heinrich, Martin Jatho, and Klaus Kalmring •4G Neurobiologie, FB Biologie-Zoologie, Philipps-Universit•it Marburg, Postfach 1929, D-3550 Marburg, Germany
(Received 4 December 1992; accepted for publication 23 February 1993)
The transmission characteristics of the acoustic tracheae in the forelegs of seven tettigoniid species were investigated by sinusoidal analysis. The species were selected to represent a range of body sizes and leg lengths. Four subfamilies were included, with two species each from three of them; the tracheae in such closely related pairs could be expected to be similar in shape despite their different dimensions. The tracheae were dissected out for morphometric analysis and compared with one another with respect to their overall dimensions and those of typical subsections. The amplitude-versus-frequency response of acoustic transmission in the tracheae was measured at various positions with a probe microphone. The stimuli were continuous sinusoidal signals at an intensity of 100 or 110 dB SPL. The tracheae of all the species studied here (in males and females) are distinguished by a bandpass-limited transmission characteristic. In the frequency range above 5 kHz (at least to 40 kHz) the sound signals are amplified by 10-15 dB during passage through the tracheae. These results are compared with the threshold curves of the auditory organs and the spectra of the conspecific songs. Although in some cases there are considerable differences in the dimensions of the tracheae, the transmission characteristics are very similar; no specific adaptations to the frequency composition of the conspecific song were found.
PACS numbers: 43.80.Lb, 43.80.Cs, 43.80.Jz
INTRODUCTION
It has remained unclear whether acoustic transmission
in the auditory receptor organs in the forelegs of bushcrick- ets is affected by differences in the dimensions of the tra- cheae (Nocke, 1975). In principle, a number of variables could influence the transmission of sound signals from the body surface to the receptor cells of the two auditory or- gans the size of the spiracles, the shape of vesicles, and the diameter and length of the tracheae in the leg region. Fil- tering and amplifying processes in the frequency and time domains could produce a signal fundamentally different from that emitted by the transmitter. Although such alter- ations would limit the range of frequencies audible by the intermediate and tympanal organs, it is also possible that amplification during transmission of the bandpass-filtered signals would improve the bushcricket's heating. In partic- ular, it is conceivable that the transmission characteristics of the tympanal trachea represent adaptations to parame- ters of the conspecific song.
The experiments described here were undertaken to clarify these questions; by means of sinusoidal analysis, the transmission characteristics of the acoustic trachea were
measured in seven species of bushcrickets with legs of dif- ferent dimensions.
I. MATERIALS AND METHODS
The experimental animals included both male and fe- male imagines of the following species: Decticus albifrons (D.a.) and Decticus verrucivorus (D.v.) of the subfamily
Decticinae, Tettigonia w'ridissima (T.v.) and Tettigonia cantans (T.c.) of the subfamily Tettigoniinae, Ephippiger ephippiger (E.e.) and Ephippigerida taeniata (E.t.) of the subfamily Ephippigerinae, and the Australian species My- galopsis marki (M.m.), a conocephaline. The animals were selected according to the degree of kinship and to the size of their forelegs, in order to obtain closely related bush- crickets that differed in the sizes of their spiracles and other tracheal elements. The dimensions of the tracheae
were determined morphometrically and the acoustic trans- mission characteristics were measured at three different
positions along the trachea, with a directly coupled probe microphone. Measurements were carried out on one fore- leg in each of six animals of each species. The other foreleg was used for the morphometric analysis of the acoustic trachea.
A. Morphometric measurement of the acoustic trachea
In order to determine the shape, volume, length, and diameter of the acoustic trachea in the thorax and the
foreleg, the trachea was dissected out of the animal and transferred either to ringer solution or to 70% alcohol. This treatment did not alter the size or shape of the tra- chea. Camera lucida drawings were prepared with a stereo lens system (Wild M3 with drawing mirror), and by mea- suring these the shape and dimensions of the whole trachea as well as its subsections were determined. In comparing the different tracheae, special attention was paid to the size and shape of the spiracle and vesicle in the prothorax, the
3481 J. Acoust. Soc. Am. 93 (6), June 1993 0001-4966/93/063481-09506.00 @ 1993 Acoustical Society of America 3481
Redistribution subject to ASA license or copyright; see http://acousticalsociety.org/content/terms. Download to IP: 132.206.205.6 On: Thu, 04 Dec 2014 10:57:40

D.a. T.c.
1
FIG. 1. Acoustic tracheal systems of the seven species examined. The drawing in the middle of the figure shows a posterior view of the tibial part of the acoustic trachea of D. albifrons, indicating the sites of the acoustic measurements. 1:' distal tibia, 2: proximal tibia, 3: femur, ao: auditory organ, as: air sac, er: colapsed region, tracheal constriction, f: femural part of the trachea, sp: spiracle, v: vesicle.
length and diameter of the trachea in the proximal leg segments as far as the genual joint, the differentiation of the tympanal trachea in the region of the auditory organs, the constriction distal to these organs, and a saclike expansion in the distal region of the tibia (Fig. 1 ). The dimensions of the individual tracheal subsections of males and females of
the various species were tabulated and represented sche- matically for comparison.
B. Acoustic measurement
The measurements were carried out on living bush- crickets. The animal was anesthetized with CO2 and, after removal of the wings, attached to a holder with a wax- colophonium mixture and mounted on a micromanipulator in a reflection-damped chamber. It was positioned in such a way that the spiracle and foreleg on the right side of the body were directly opposite a loudspeaker (Audax TW8 spezial). The distance between speaker and spiracle was 60 cm (free-field conditions). The foreleg was fixed so as to immobilize the knee joint, with the tibia and femur at right angles to one another. Care was taken to keep the slits in the proximal tibial region open. During the measurements a probe microphone was coupled directly to the trachea at three different positions: (1) In the distal tibia, in the re- gion of the "air sac." (2) In the proximal tibia, between the crista acoustica and the intermediate organ (where the trachea is undivided). (3) In the middle region of the femur (see Fig. 1 inset). To enable the measurements, the legs were cut transversely and the distal parts were re- moved. That may have affected the measurements, but was necessary to do such investigations.
114"-microphon
lcm
FIG. 2. Schematic longitudinal section through the probe microphone, incorporating a 1/4-in. microphone capsule.
C. Probe microphone
The probe microphone used to measure transmission in the acoustic trachea consisted of a thick-walled alumi-
num tube 1 cm in diameter and about 6 cm long. A 1/4-in. condenser microphone (Briiel & Kjaer 4135) was fitted into the anterior wall of the tube so that the membrane of
the microphone was parallel to the tube wall. Inserted into the front end of the tube was a hollow cone truncated at
the tip to produce an opening 300/•m in diameter. The back end of the tube was extended by a flexible plastic pipe containing a sound-absorbing material (Fig. 2). This ar- rangement achieved a coupling of microphone to trachea in which interference was largely eliminated. The probe microphone was constructed by Dipl. Ing. Bertram Schapal firm Briiel & Kjaer adapted to the measurement conditions at the tracheae. It was built up on the basis of the probe microphone type 4182 of Briiel & Kjaer con- structed for measurements in very small volumes. The acoustic impedance is very high minimizing reflexions dur- ing the measurements and the load of the source is small. The measurements presented here were carried out with a probe microphone, which offers both advantages and dis- advantages for such experiments. An advantage is that the measurement system can be coupled directly to the trachea by way of the hollow cone. A disadvantage of the system is its low sensitivity, especially in the high-frequency range, so that the highest measurable frequency was 40 kHz.•Fur- thermore, the stimulus intensity had to be very high, 100 or 110 dB SPL, to obtain a usable signal-to-noise ratio.
D. Measurement of the transfer function
Continuous sinusoidal tones were used for the mea-
surements. These were produced with a sine-wave genera- tor (Exact 120 waveform generator) and sent to a loud- speaker (Audax TW8) by way of a power amplifier. The frequency of the tones was monitored with a frequency counter (Dietechnik type 101 ).
The microphone was attached to a sound-pressure- level meter (Briiel & Kjaer 2009) that indicated the peak values of the sound pressure. Frequencies from 2 to 40 kHz were used for the measurement, at intensities of 100-110 dB SPL.
Prior to the experiments the frequency-transmission characteristic of the loudspeaker and the free-field condi- tions in the experimental chamber were measured with a 1/4-in. condenser microphone (Briiel & Kjaer 4135) in the place where the spiracle would be positioned during the
3482 J. Acoust. Soc. Am., Vol. 93, No. 6, June 1993 Heinrich et al.: Bushcricket tracheae, studies of seven species 3482
Redistribution subject to ASA license or copyright; see http://acousticalsociety.org/content/terms. Download to IP: 132.206.205.6 On: Thu, 04 Dec 2014 10:57:40
1,s D.a. 1.4
0,6 • ' ø'tl 0,6
1,0• 1,4
1,8 0 5 10 15 20 25 • •
T.Co E.t.
/
1,8'
1,4: D.v. 1,o: o,6: o,2: (:•
0,2 • o,6:
1,4:
1'8o 5 •o •5'"•6 25 3o 35 0 5 10 15 20 25 30 35
M.m.
length ! mm
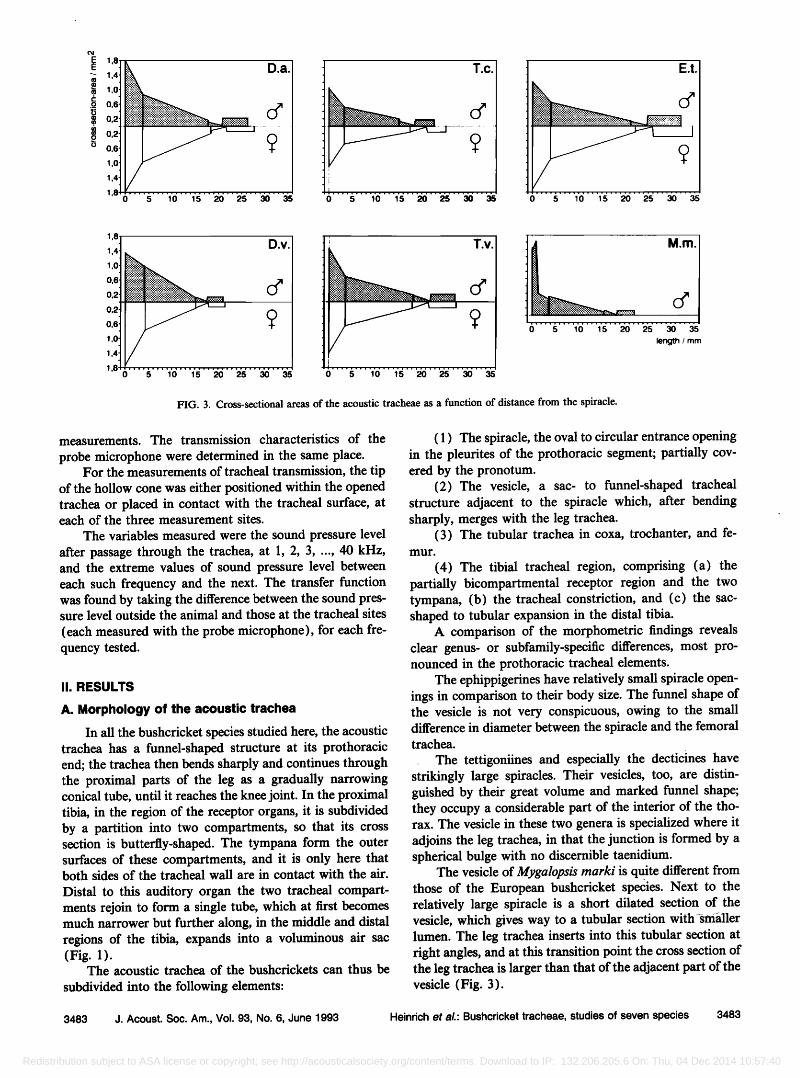
FIG. 3. Cross-sectional areas of the acoustic tracheae as a function of distance from the spiracle.
measurements. The transmission characteristics of the
probe microphone were determined in the same place. For the measurements of tracheal transmission, the tip
of the hollow cone was either positioned within the opened trachea or placed in contact with the tracheal surface, at each of the three measurement sites.
The variables measured were the sound pressure level after passage through the trachea, at 1, 2, 3, ..., 40 kHz, and the extreme values of sound pressure level between each such frequency and the next. The transfer function was found by taking the difference between the sound pres- sure level outside the animal and those at the tracheal sites (each measured with the probe microphone), for each fre- quency tested.
II. RESULTS
A. Morphology of the acoustic trachea
In all the bushcricket species studied here, the acoustic trachea has a funnel-shaped structure at its prothoracic end; the trachea then bends sharply and continues through the proximal parts of the leg as a gradually narrowing conical tube, until it reaches the knee joint. In the proximal tibia, in the region of the receptor organs, it is subdivided by a partition into two compartments, so that its cross section is butterfly-shaped. The tympana form the outer surfaces of these compartments, and it is only here that both sides of the tracheal wall are in contact with the air. Distal to this auditory organ the two tracheal compart- ments rejoin to form a single tube, which at first becomes much narrower but further along, in the middle and distal regions of the tibia, expands into a voluminous air sac (Fig. 1 ).
The acoustic trachea of the bushcrickets can thus be
subdivided into the following elements:
( 1 ) The spiracle, the oval to circular entrance opening in the pleurites of the prothoracic segment; partially cov- ered by the pronotum.
(2) The vesicle, a sac- to funnel-shaped tracheal structure adjacent to the spiracle which, after bending sharply, merges with the leg trachea.
(3) The tubular trachea in coxa, trochanter, and fe- mur.
(4) The tibial tracheal region, comprising (a) the partially bicompartmental receptor region and the two tympana, (b) the tracheal constriction, and (c) the sac- shaped to tubular expansion in the distal tibia.
A comparison of the morphometric findings reveals clear genus- or subfamily-specific differences, most pro- nounced in the prothoracic tracheal elements.
The ephippigerines have relatively small spiracle open- ings in comparison to their body size. The funnel shape of the vesicle is not very conspicuous, owing to the small difference in diameter between the spiracle and the femoral trachea.
The tettigoniines and especially the decticines have strikingly large spiracles. Their vesicles, too, are distin- guished by their great volume and marked funnel shape; they occupy a considerable part of the interior of the tho- rax. The vesicle in these two genera is specialized where it adjoins the leg trachea, in that the junction is formed by a spherical bulge with no discernible taenidium.
The vesicle of Mygalopsis marki is quite different from those of the European bushcricket species. Next to the relatively large spiracle is a short dilated section of the vesicle, which gives way to a tubular section with •srnaller lumen. The leg trachea inserts into this tubular section at fight angles, and at this transition point the cross section of the leg trachea is larger than that of the adjacent part of the vesicle (Fig. 3).
3483 J. Acoust. Soc. Am., Vol. 93, No. 6, June 1993 Heinrich et al.: Bushcricket tracheae, studies of seven species 3483
Redistribution subject to ASA license or copyright; see http://acousticalsociety.org/content/terms. Download to IP: 132.206.205.6 On: Thu, 04 Dec 2014 10:57:40
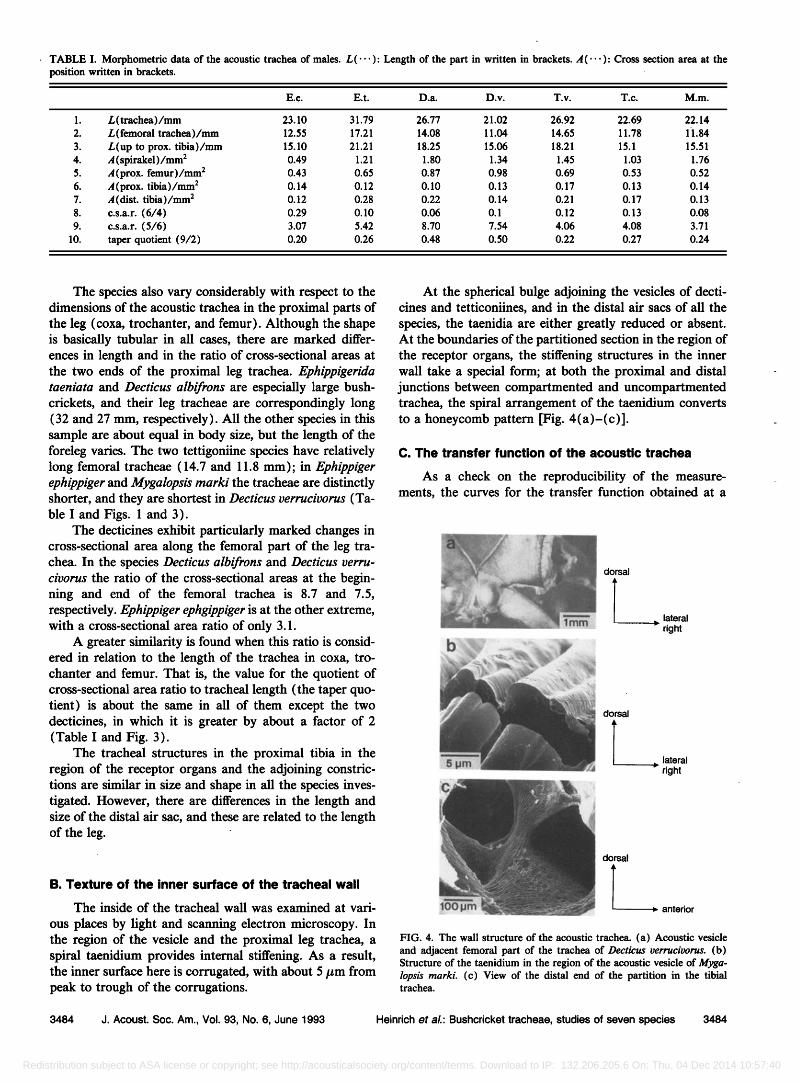
TABLE I. Morphometric data of the acoustic trachea of males. L(--- ): Length of the part in written in brackets. A(.-' ): Cross section area at the position written in brackets.
E.e. E.t. D.a. D.v. T.v. T.c. M.m.
1. L(trachea)/mm 23.10 31.79 26.77 21.02 26.92 22.69 22.14 2. L(femoral trachea)/mm 12.55 17.21 14.08 11.04 14.65 11.78 1'1.84 3. L(up to prox. tibia)/mm 15.10 21.21 18.25 15.06 18.21 15.1 15.51 4. •/(spirakel)/mm 2 0.49 1.21 1.80 1.34 1.45 1.03 1.76 5. ,4 (prox. femur)/mm 2 0.43 0.65 0.87 0.98 0.69 0.53 0.52 6. •/(prox. tibia)/mm 2 0.14 0.12 0.10 0.13 0.17 0.13 0.14 7. ,,/(dist. tibia)/mm • 0.12 0.28 0.22 0.14 0.21 0.17 0.13 8. c.s.a.r. (6/4) 0.29 0.10 0.06 0.1 0.12 0.13 0.08 9. c.s.a.r. (5/6) 3.07 5.42 8.70 7.54 4.06 4.08 3.71
10. taper quotient (9/2) 0.20 0.26 0.48 0.50 0.22 0.27 0.24
The species also vary considerably with respect to the dimensions of the acoustic trachea in the proximal parts of the leg (coxa, trochanter, and femur). Although the shape is basically tubular in all cases, there are marked differ- ences in length and in the ratio of cross-sectional areas at the two ends of the proximal leg trachea. Ephippigerida taeniata and Decticus albifrons are especially large bush- crickets, and their leg tracheae are correspondingly long (32 and 27 ram, respectively). All the other species in this sample are about equal in body size, but the length of the foreleg varies. The two tettigoniine species have relatively long fernoral tracheae (14.7 and 11.8 ram); in Ephippiger ephippiger and Mygalopsis marki the tracheae are distinctly shorter, and they are shortest in Decticus verrucivorus (Ta- ble I and Figs. 1 and 3).
The decticines exhibit particularly marked changes in cross-sectional area along the fernoral part of the leg tra- chea. In the species Decticus albifrons and Decticus verru- civorus the ratio of the cross-sectional areas at the begin- ning and end of the fernoral trachea is 8.7 and 7.5, respectively. Ephippiger ephgippiger is at the other extreme, with a cross-sectional area ratio of only 3.1.
A greater similarity is found when this ratio is consid- ered in relation to the length of the trachea in coxa, tro- chanter and femur. That is, the value for the quotient of cross-sectional area ratio to tracheal length (the taper quo- tient) is about the same in all of them except the two decticines, in which it is greater by about a factor of 2 (Table I and Fig. 3).
The tracheal structures in the proximal tibia in the region of the receptor organs and the adjoining constric- tions are similar in size and shape in all the species inves- tigated. However, there are differences in the length and size of the distal air sac, and these are related to the length of the leg.
B. Texture of the inner surface of the tracheal wall
The inside of the tracheal wall was examined at vari-
ous places by light and scanning electron microscopy. In the region of the vesicle and the proximal leg trachea, a spiral taenidium provides internal stiffening. As a result, the inner surface here is corrugated, with about 5/zm from peak to trough of the corrugations.
At the spherical bulge adjoining the vesicles of decti- cines and tetticoniines, and in the distal air sacs of all the species, the taenidia are either greatly reduced or absent. At the boundaries of the partitioned section in the region of the receptor organs, the stiffening structures in the inner wall take a special form; at both the proximal and distal junctions between compartmented and uncompartmented trachea, the spiral arrangement of the taenidium converts to a honeycomb pattern [Fig. 4 (a)-(c) ].
C. The transfer function of the acoustic trachea
As a check on the reproducibility of the measure- ments, the curves for the transfer function obtained at a
lateral
ß right
•i:.•-•<<•.::•:•$. •!•--:•.•....• ...::•..:..... .... .::.:'":•.:,•!i•.•--•:.:.•:• .....•-•-•-•:•:•---•'•
Z ?::'"' •:::•: .:.•:•: --'-'-::-. ::.:•:. ...•.?:
.:..:: ...... . ........... •?? :•:• -•-.. _ lateral •'?•;• ..... '•:• .... -:: :z::• ' right
dorsal
anterior
FIG. 4. The wall structure of the acoustic trachea. (a) Acoustic vesicle and adjacent femoral part of the trachea of Decticus •erruci•orus. (b) Structure of the taenidium in the region of the acoustic vesicle of Myga- lopsis marki. (c) View of the distal end of the partition in the tibial trachea.
3484 J. Acoust. Soc. Am., Vol. 93, No. 6, June 1993 Heinrich et aL: Bushcricket tracheae, studies of seven species 3484
Redistribution subject to ASA license or copyright; see http://acousticalsociety.org/content/terms. Download to IP: 132.206.205.6 On: Thu, 04 Dec 2014 10:57:40
2ø'T.v.
10
-10;• • • 10 20 30 40 -102 3 5 10 20 30 40 frequency / kHz
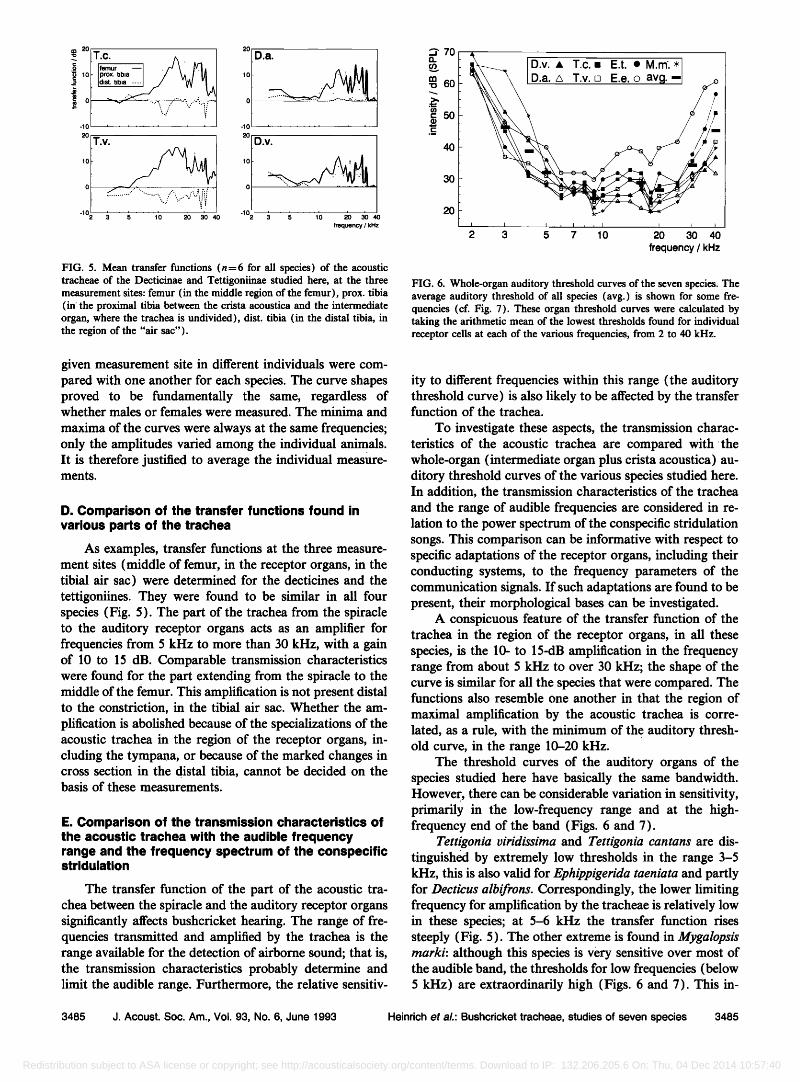
FIG. 5. Mean transfer functions (n=6 for all species) of the acoustic tracheae of the Decticinae and Tettigoniinae studied here, at the three measurement sites: femur (in the middle region of the femur), prox. tibia (in' the proximal tibia between the crista acoustica and the intermediate organ, where the trachea is undivided), dist. tibia (in the distal tibia, in the region of the "air sac").
2 3 5 7 10 2•0 3•0 410 frequency / kHz
FIG. 6. Whole-organ auditory threshold curves of the seven species. The average auditory threshold of all species (avg.) is shown for some fre- quencies (cf. Fig. 7). These organ threshold curves were calculated by taking the arithmetic mean of the lowest thresholds found for individual receptor cells at each of the various frequencies, from 2 to 40 kHz.
given measurement site in different individuals were com- pared with one another for each species. The curve shapes proved to be fundamentally the same, regardless of whether males or females were measured. The minima and
maxima of the curves were always at the same frequencies; only the amplitudes varied among the individual animals. It is therefore justified to average the individual measure- ments.
D. Comparison of the transfer functions found in various parts of the trachea
As examples, transfer functions at the three measure- ment sites (middle of femur, in the receptor organs, in the tibial air sac) were determined for the decticines and the tettigoniines. They were found to be similar in all four species (Fig. 5). The part of the trachea from the spiracle to the auditory receptor organs acts as an amplifier for frequencies from 5 kHz to more than 30 kHz, with a gain of 10 to 15 dB. Comparable transmission characteristics were found for the part extending from the spiracle to the middle of the femur. This amplification is not present distal to the constriction, in the tibial air sac. Whether the am- plification is abolished because of the specializations of the acoustic trachea in the region of the receptor organs, in- cluding the tympana, or because of the marked changes in cross section in the distal tibia, cannot be decided on the basis of these measurements.
E. Comparison of the transmission characteristics of the acoustic trachea with the audible frequency range and the frequency spectrum of the conspecific stridulation
The transfer function of the part of the acoustic tra- chea between the spiracle and the auditory receptor organs significantly affects bushcricket heating. The range of fre- quencies transmitted and amplified by the trachea is the range available for the detection of airborne sound; that is, the transmission characteristics probably determine and limit the audible range. Furthermore, the relative sensitiv-
ity to different frequencies within this range (the auditory threshold curve) is also likely to be affected by the transfer function of the trachea.
To investigate these aspects, the transmission charac- teristics of the acoustic trachea are compared with the whole-organ (intermediate organ plus crista acoustica) au- ditory threshold curves of the various species studied here. In addition, the transmission characteristics of the trachea and the range of audible frequencies are considered in re- lation to the power spectrum of the conspecific stridulation songs. This comparison can be informative with respect to specific adaptations of the receptor organs, including their conducting systems, to the frequency parameters of the communication signals. If such adaptations are found to be present, their morphological bases can be investigated.
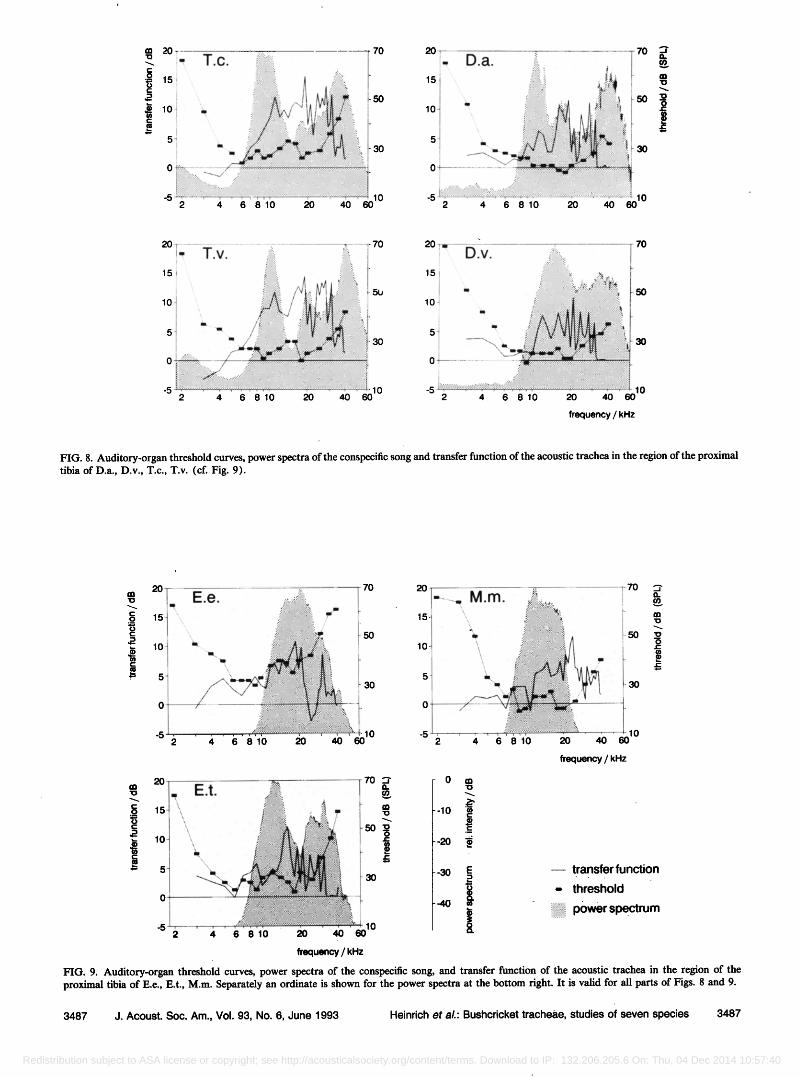
A conspicuous feature of the transfer function of the trachea in the region of the receptor organs, in all these species, is the 10- to 15-dB amplification in the frequency range from about 5 kHz to over 30 kHz; the shape of the curve is similar for all the species that were compared. The functions also resemble one another in that the region of maximal amplification by the acoustic trachea is corre- lated, as a rule, with the minimum of the auditory thresh- old curve, in the range 10-20 kHz.
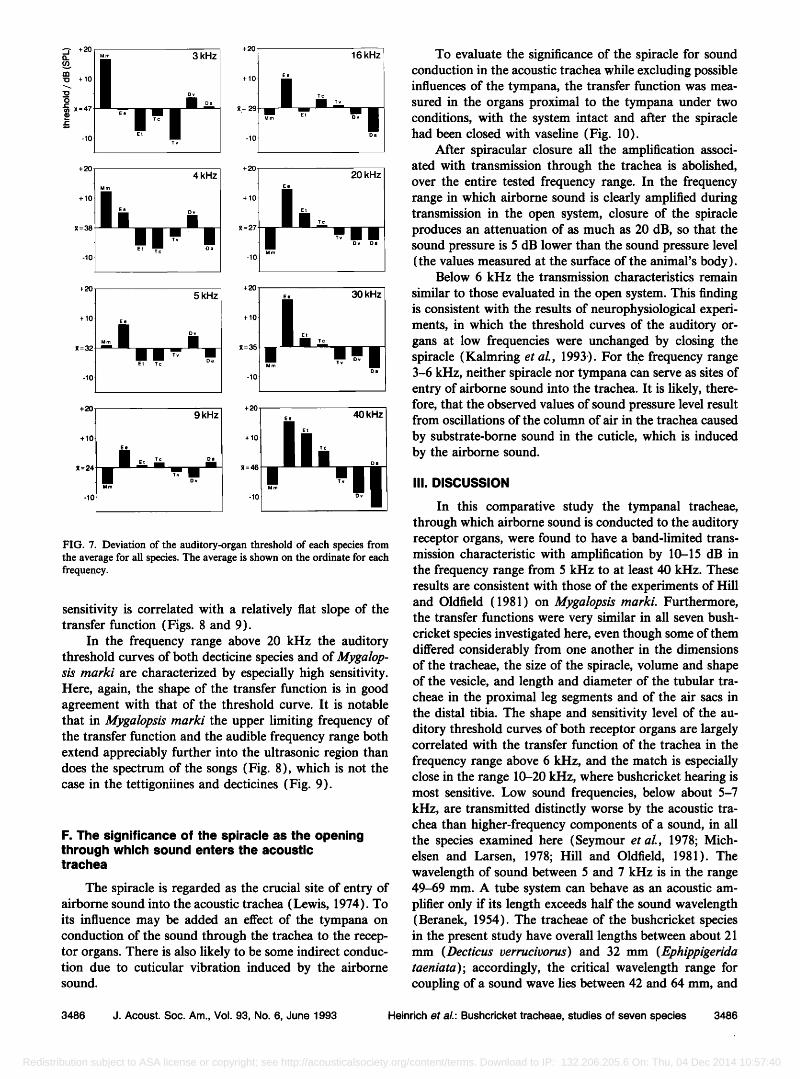
The threshold curves of the auditory organs of the species studied here have basically the same bandwidth. However, there can be considerable variation in sensitivity, primarily in the low-frequency range and at the high- frequency end of the band (Figs. 6 and 7).
Tettigonia viridissima and Tettigonia cantans are dis- tinguished by extremely low thresholds in the range 3-5 kHz, this is also valid for Eœhiœœigerida taeniata and partly for Decticus albifrons. Correspondingly, the lower limiting frequency for amplification by the tracheae is relatively low in these species; at 5-6 kHz the transfer function rises steeply (Fig. 5). The other extreme is found in Mygalopsis marki: although this species is very sensitive over most of the audible band, the thresholds for low frequencies (below 5 kHz) are extraordinarily high (Figs. 6 and 7). This in-
3485 J. Acoust. Soc. Am., Vol. 93, No. 6, June 1993 Heinrich et al.: Bushcricket tracheae, studies of seven species 3485
Redistribution subject to ASA license or copyright; see http://acousticalsociety.org/content/terms. Download to IP: 132.206.205.6 On: Thu, 04 Dec 2014 10:57:40
03 +201 ,,,,,- 3kHz +20 16kHz
• DE e Et• •---29 m __ I• Tv
• = 38 • t• • = 27 • t• I
+20
+101 5 kHz
Ee
v
=32
-10
+ 20.
+10
X=35
-10
i T,_%•
30 kHz
+2o 9 kHz +2ø T •, 40 kHz
+10 +10 œt Tc
FIG. 7. Deviation of the auditory-organ threshold of each species from the average for all species. The average is shown on the ordinate for each frequency.
sensitivity is correlated with a relatively flat slope of the transfer function (Figs. 8 and 9).
In the frequency range above 20 kHz the auditory threshold curves of both decticine species and of Mygalop- sis marki are characterized by especially high sensitivity. Here, again, the shape of the transfer function is in good agreement with that of the threshold curve. It is notable that in Mygalopsis marki the upper limiting frequency of the transfer function and the audible frequency range both extend appreciably further into the ultrasonic region than does the spectrum of the songs (Fig. 8), which is not the case in the tettigoniines and decticines (Fig. 9).
F. The significance of the spiracle as the opening through which sound enters the acoustic trachea
The spiracle is regarded as the crucial site of entry of airborne sound into the acoustic trachea (Lewis, 1974). To its influence may be added an effect of the tympana on conduction of the sound through the trachea to the recep- tor organs. There is also likely to be some indirect conduc- tion due to cuticular vibration induced by the airborne sound.
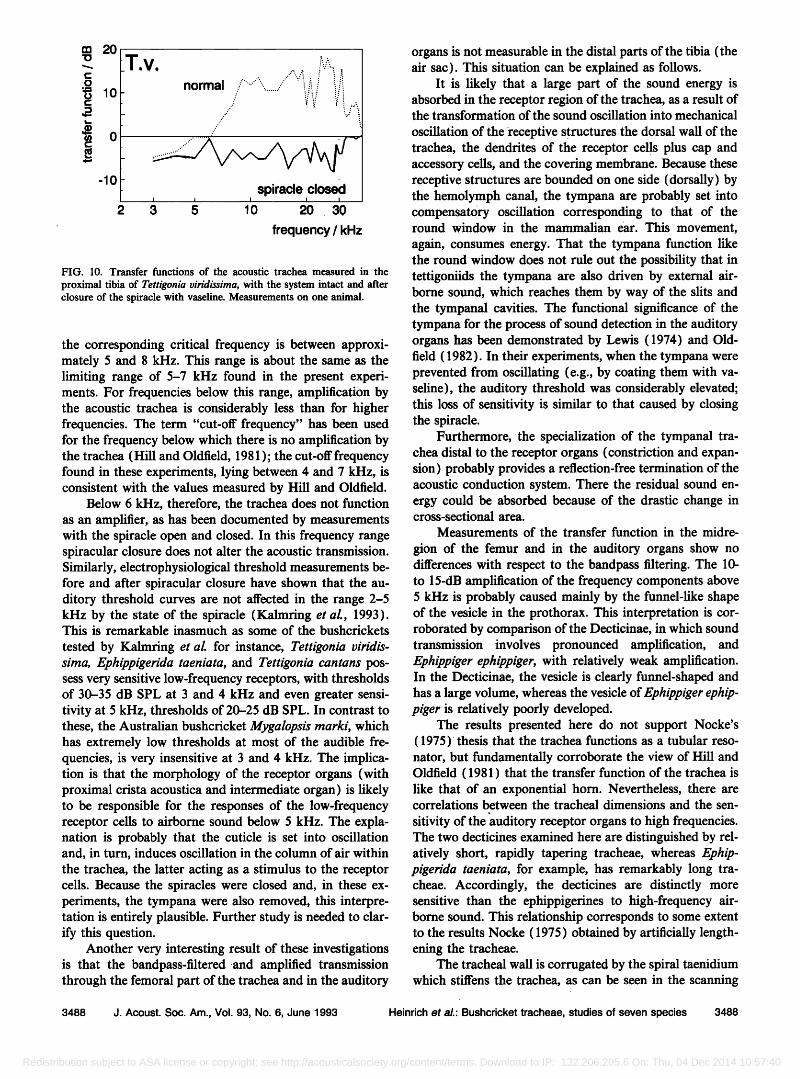
To evaluate the significance of the spiracle for sound conduction in the acoustic trachea while excluding possible influences of the tympana, the transfer function was mea- sured in the organs proximal to the tympana under two conditions, with the system intact and after the spiracle had been closed with vaseline (Fig. 10).
After spiracular closure all the amplification associ- ated with transmission through the trachea is abolished, over the entire tested frequency range. In the frequency range in which airborne sound is clearly amplified during transmission in the open system, closure of the spiracle produces an attenuation of as much as 20 dB, so that the sound pressure is 5 dB lower than the sound pressure level (the values measured at the surface of the animal's body).
Below 6 kHz the transmission characteristics remain
similar to those evaluated in the open system. This finding is consistent with the results of neurophysiological experi- ments, in which the threshold curves of the auditory or- gans at low frequencies were unchanged by closing the spiracle (Kalmring et al., 1993). For the frequency range 3-6 kHz, neither spiracle nor tympana can serve as sites of entry of airborne sound into the trachea. It is likely, there- fore, that the observed values of sound pressure level result from oscillations of the column of air in the trachea caused
by substrate-borne sound in the cuticle, which is induced by the airborne sound.
III. DISCUSSION
In this comparative study the tympanal tracheae, through which airborne sound is conducted to the auditory receptor organs, were found to have a band-limited trans- mission characteristic with amplification by 10-15 dB in the frequency range from 5 kHz to at least 40 kHz. These results are consistent with those of the experiments of Hill and Oldfield ( 1981 ) on Mygalopsis marki. Furthermore, the transfer functions were very similar in all seven bush- cricket species investigated here, even though some of them differed considerably from one another in the dimensions of the tracheae, the size of the spiracle, volume and shape of the vesicle, and length and diameter of the tubular tra- cheae in the proximal leg segments and of the air sacs in the distal tibia. The shape and sensitivity level of the au- ditory threshold curves of both receptor organs are largely correlated with the transfer function of the trachea in the
frequency range above 6 kHz, and the match is especially close in the range 10-20 kHz, where bushcricket hearing is most sensitive. Low sound frequencies, below about 5-7 kHz, are transmitted distinctly worse by the acoustic tra- chea than higher-frequency components of a sound, in all the species examined here (Seymour et al., 1978; Mich- elsen and Larsen, 1978; Hill and Oldfield, 1981). The wavelength of sound between 5 and 7 kHz is in the range 49-69 mm. A tube system can behave as an acoustic am- plifier only if its length exceeds half the sound wavelength (Beranek, 1954). The tracheae of the bushcricket species in the present study have overall lengths between about 21 mm (Decticus verrucivorus) and 32 mm (Ephippigerida taeniata); accordingly, the critical wavelength range for coupling of a sound wave lies between 42 and 64 mm, and
3486 J. Acoust. Soc. Am., Vol. 93, No. 6, June 1993 Heinrich et aL: Bushcricket tracheae, studies of seven species 3486
Redistribution subject to ASA license or copyright; see http://acousticalsociety.org/content/terms. Download to IP: 132.206.205.6 On: Thu, 04 Dec 2014 10:57:40
,.n :)o
.9 15 o
•) 10
70
50
30
20
10
-.... :iiii!!i!!i:i: ,•'--::•i: •!•i..'.'• •,..•::•::•::•..
\ :::::::::::::::::::::::::::::::::: ' i -i•i•::•::!i•:?:•i•i??:i.t•
.............................................................................................`............
........ •i!!!iiii::.:.::i:iiii!i::!!!! ....................... :::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::: ::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::: 10 2 4 6 8 10 20 40 60
70 •'
o
30
20
15
10
5
0
-5 2 4 6 8 10 20 40 60
20 = 70 ": D.v. ............
:::::::::::::•:
ß '. ::::::::::::::::::::::::::::::::::::::::::: . .:'-•.•. ::::::::::::::::::::::::::::::: :... .?.::::::.•.:"!.,• e. !:!:i:!:i:i:i:!:i:i:i:i:i:i:i:!.'.:.,:-!:.•!:. ,:•:i:i:i:i:!:i:•::::..
I i. .;.iiiiiiiii•ii:i:i:i:i:i'::i:i:i 'i: ::::::::::::::::::::::::::::::::::::::::::::
i I .................................................... ........... ............. - '"'":'iiiiiiiiiiiiiii,o 2 4 6 8 10 20 40 60
frequency / kHz
FIG. 8. Auditory-organ threshold curves, power spectra of the conspecific song and transfer function of the acoustic trachea in the region of the proximal tibia of D.a., D.v., T.c., T.v. (cf. Fig. 9).
20 70 20 70
15
10
:iii!iiiiii:::::::::::::::i!iii!i! /
... ::::::::::::::::::::::::::::::::::::::::::::::::::::::::: "•,
....... = ::::::::::::::::::::::::::::::: ::i :::::::::::::::::::::::::::
•.. .'.i: :::::::::::::::::::::: ...
30
15-
10
10 -5 4 6 810 20 40 60 2
ß .... ==================================== •, =================================================
\ :::::::::::::::::::::::::::::::::::::::: •. :.:.:.:.:,:.:.:.:.:.-.- .....
:- .,.'•!•i::ii•:?:•::•: .:•!•! '.• .,.i:!:i:i:i:i•:??
.... ß •... '5::!:
4 6 8 10 20 , 10
' 40 60
frequency / kHz
• 20 / . E.t. .•i•i:•!'::• 7o õ •5
........ ,
5 :.
........... •:J:•:[:?•10 -5 "• • 4 6 810
fr•uency / kHz
-10
-20
-4O
transfer function
..... threshold
i?..-:•i! i p0wer spectrum
FIG. 9. Auditory-organ threshold curves, power spectra of the conspecific song, and transfer function of the acoustic trachea in the region of the proximal tibia of E.e., E.t., M.m. Separately an ordinate is shown for the power spectra at the bottom right. It is valid for all parts of Figs. 8 and 9.
3487 J. Acoust. Soc. Am., Vol. 93, No. 6, June 1993 Heinrich et aL: Bushcricket tracheae, studies of seven species 3487
Redistribution subject to ASA license or copyright; see http://acousticalsociety.org/content/terms. Download to IP: 132.206.205.6 On: Thu, 04 Dec 2014 10:57:40
m 20
o
• 10
o
-lO
T •... ß V., ../.:....:.• ..? ......... ........• normal.... .::?;::
.............. ? .... i! •':' ..... ...' • ¾ ,,.',
.:/" .•..?.....•
..
ß
r
2 3 5 10 20 30
frequency / kHz
FIG. 10. Transfer functions of the acoustic trachea measured in the
proximal tibia of Tettigonia viridissirna, with the system intact and after closure of the spiracle with vaseline. Measurements on one animal.
the corresponding critical frequency is between approxi- mately 5 and 8 kHz. This range is about the same as the limiting range of 5-7 kHz found in the present experi- ments. For frequencies below this range, amplification by the acoustic trachea is considerably less than for higher frequencies. The term "cut-off frequency" has been used for the frequency below which there is no amplification by the trachea (Hill and Oldfield, 1981 ); the cut-off frequency found in these experiments, lying between 4 and 7 kHz, is consistent with the values measured by Hill and Oldfield.
Below 6 kHz, therefore, the trachea does not function as an amplifier, as has been documented by measurements with the spiracle open and closed. In this frequency range spiracular closure does not alter the acoustic transmission. Similarly, electrophysiological threshold measurements be- fore and after spiracular closure have shown that the au- ditory threshold curves are not affected in the range 2-5 kHz by the state of the spiracle (Kalmring et al., 1993). This is remarkable inasmuch as some of the bushcrickets
tested by Kalmring et al. for instance, Tettigonia viridis- sirna, Ephippigerida taeniata, and Tettigonia cantans pos- sess very sensitive low-frequency receptors, with thresholds of 30-35 dB SPL at 3 and 4 kHz and even greater sensi- tivity at 5 kHz, thresholds of 20-25 dB SPL. In contrast to these, the Australian bushcricket Mygalopsis marki, which has extremely low thresholds at most of the audible fre- quencies, is very insensitive at 3 and 4 kHz. The implica- tion is that the morphology of the receptor organs (with proximal crista acoustica and intermediate organ) is likely to be responsible for the responses of the low-frequency receptor cells to airborne sound below 5 kHz. The expla- nation is probably that the cuticle is set into oscillation and, in turn, induces oscillation in the column of air within the trachea, the latter acting as a stimulus to the receptor cells. Because the spiracles were closed and, in these ex- periments, the tympana were also removed, this interpre- tation is entirely plausible. Further study is needed to clar- ify this question.
Another very interesting result of these investigations is that the bandpass-filtered and amplified transmission through the femoral part of the trachea and in the auditory
organs is not measurable in the distal parts of the tibia (the air sac). This situation can be explained as follows.
It is likely that a large part of the sound energy is absorbed in the receptor region of the trachea, as a result of the transformation of the sound oscillation into mechanical
oscillation of the receptive structures the dorsal wall of the trachea, the dendrites of the receptor cells plus cap and accessory cells, and the covering membrane. Because these receptive structures are bounded on one side (dorsally) by the hemolymph canal, the tympana are probably set into compensatory oscillation corresponding to that of the round window in the mammalian ear. This movement,
again, consumes energy. That the tympana function like the round window does not rule out the possibility that in tettigoniids the tympana are also driven by external air- borne sound, which reaches them by way of the slits and the tympanal cavities. The functional significance of the tympana for the process of sound detection in the auditory organs has been demonstrated by Lewis (1974) and Old- field (1982). In their experiments, when the tympana were prevented from oscillating (e.g., by coating them with va- seline), the auditory threshold was considerably elevated; this loss of sensitivity is similar to that caused by closing the spiracle.
Furthermore, the specialization of the tympanal tra- chea distal to the receptor organs (constriction and expan- sion) probably provides a reflection-free termination of the acoustic conduction system. There the residual sound en- ergy could be absorbed because of the drastic change in cross-sectional area.
Measurements of the transfer function in the midre-
gion of the femur and in the auditory organs show no differences with respect to the bandpass filtering. The 10- to 15-dB amplification of the frequency components above 5 kHz is probably caused mainly by the funnel-like shape of the vesicle in the prothorax. This interpretation is cor- roborated by comparison of the Decticinae, in which sound transmission involves pronounced amplification, and Ephippiger ephippiger, with relatively weak amplification. In the Decticinae, the vesicle is clearly funnel-shaped and has a large volume, whereas the vesicle of Ephippiger ephip- piger is relatively poorly developed.
The results presented here do not support Nocke's (1975) thesis that the trachea functions as a tubular reso- nator, but fundamentally corroborate the view of Hill and Oldfield ( 1981 ) that the transfer function of the trachea is like that of an exponential horn. Nevertheless, there are correlations between the tracheal dimensions and the sen-
sitivity of the •tuditory receptor organs to high frequencies. The two decticines examined here are distinguished by rel- atively short, rapidly tapering tracheae, whereas Ephip- pigerida taeniata, for example, has remarkably long tra- cheae. Accordingly, the decticines are distinctly more sensitive than the ephippigerines to high-frequency air- borne sound. This relationship corresponds to some extent to the results Nocke (1975) obtained by artificially length- ening the tracheae.
The tracheal wall is corrugated by the spiral taenidium which stiffens the trachea, as can be seen in the scanning
3488 J. Acoust. Soc. Am., Vol. 93, No. 6, June 1993 Heinrich et al.: Bushcricket tracheae, studies of seven species 3488
Redistribution subject to ASA license or copyright; see http://acousticalsociety.org/content/terms. Download to IP: 132.206.205.6 On: Thu, 04 Dec 2014 10:57:40

electron microscope; the amplitude of the corrugations on the inner surface is 5/zm. The shortest wavelength present in the communication signals of bushcrickets is 4.9 mm (upper limiting frequency 70 kHz). The unevenness of the inner tracheal surface is thus smaller by a factor of 1000 than the shortest wavelengths in the relevant sound signal transmitted through the trachea. It follows that sound con- duction in the trachea is not impaired by the taenidium.
ACKNOWLEDGMENTS
We thank Dipl. Ing. B. Schapal from Briiel & Kjaer for the help to construct the probe microphone that was built by Mr. Thomas and Mr. Muth (Mechanical Work- shop, FB Physik University Marburg). We are also grate- ful to Dr. Ann Thorson and Dr. John Thorson for trans-
lating the German manuscript into English, critically reading the manuscript, and for helpful comments. We thank also Mrs. E. Schaefer for technical help. This work
was supported by the Deutsche Forschungsgemeinschaft, Sonderforschungsbereich 305 (Ecophysiology), part of program A7.
Beranek, L. (1954). Acoustics (Mc Graw-Hill, New York). Hill, K. G., and Oldfield, B. P. (1981). "Auditory function in Tettigoni-
idac (Orthoptera, Ensifera)," J. Comp. Physiol. 142, 169-180. Kalmring, K., R/Sssler, W., Ebendt, R., Ahi., J., and Lakes, R. (1993).
"The auditory receptor organs in the forelegs of bushcrickets," Zool. J. Physiol., 97, 75-94.
Lewis, D. B. (1974). "The physiology of the tettigoniid ear. I-IV," J. Exp. Biol. 60, 821-869.
Michelsen, A., and Larsen, O. N. (1978). "Biophysics of the Ensiferan ear. I. Tympanal vibrations in bushcrickets (Tettigoniidae) studied with laser vibrometry," J. Comp. Physiol. 123, 193-203.
Nocke, H. (1975). "Physical and physiological properties of the tettigo- niid (grasshopper) ear," J. Comp. Physiol. 100, 25-57.
Oldfield, B. P. (1982). "Tonotopic organization of auditory receptors in Tettigoniidae (Orthoptera: Ensifera)," J. Comp. Physiol. 147, 461-469.
Seymour, C., Lewis, B., Larsen O. N., and Michelsen, A. (1978). Bio- physics of the ensiferan ear. II. The steady-state gain of the hearing trumpet in bushcrickets," J. Comp. Physiol. 123, 205-216.
3489 J. Acoust. Soc. Am., Vol. 93, No. 6, June 1993 Heinrich et aL: Bushcricket tracheae, studies of seven species 3489
Redistribution subject to ASA license or copyright; see http://acousticalsociety.org/content/terms. Download to IP: 132.206.205.6 On: Thu, 04 Dec 2014 10:57:40