Embed Size (px)
Citation preview

Journal of Physiology (1989). 411, pp. 639-661 639With 12 text-figuresPrinted in Great Britain
SODIUM-CALCIUM EXCHANGE DURING THE ACTION POTENTIAL INGUINEA-PIG VENTRICULAR CELLS
BY T. M. EGAN, D. NOBLE, S. J. NOBLE, T. POWELL, A. J. SPINDLERAND V. W. TWIST
From the University Laboratory of Physiology, Parks Road. Oxford OX] 3PT
(Received 25 May 1988)
SUMMARY
1. Slow inward tail currents attributable to electrogenic sodium-calcium exchangecan be recorded by imposing hyperpolarizing voltage clamp pulses during the normalaction potential of isolated guinea-pig ventricular cells. The hyperpolarizationsreturn the membrane to the resting potential (between -65 and -88 m V) allowingan inward current to be recorded. This current usually has peak amplitude whenrepolarization is imposed during the first 50 ms after the action potential upstroke,but becomes negligible once the final phase of repolarization is reached. The envelopeof peak current tail amplitudes strongly resembles that of the intracellular calciumtransient recorded in other studies.
2. Repetitive stimulation producing normal action potentials at a frequencyof 2 Hz progressively augments the tail current recorded immediately after thestimulus train. Conversely, if each action potential is prematurely terminated at01 Hz, repetitive stimulation produces a tail current much smaller than the controlvalue. The control amplitude of inward current is only maintained if interruptedaction potentials are separated by at least one full 'repriming' action potential.These effects mimic those on cell contraction (Arlock & Wohlfart, 1986) and suggestthat progressive changes in tail current are controlled by variations in the amplitudeand time course of the intracellular calcium transient.
3. When intracellular calcium is buffered sufficiently to abolish contraction, thetail current is abolished. Substitution of calcium with strontium greatly reduces thetail current.
4. The inward tail current can also be recorded at more positive membranepotentials using standard voltage clamp pulse protocols. In this way it was foundthat temperature has a large effect on the tail current, which can change from netinward at 22 °C to net outward at 37 'C. The largest inward currents are usuallyrecorded at about 30 'C. It is shown that this effect is attributable predominantly tothe temperature sensitivity of activation of the delayed potassium current, iK, whosedecay can then mask the slow tail current at high temperatures.
5. Studies of the relationship between the tail current and the membrane calciumcurrent, iCa, have been performed using a method of drug application which iscapable of perturbing ica in a very rapid and highly reversible manner. Partial blockof iCa with cadmium does not initially alter the size of the associated inward current

T. M. EGANAND OTHERS
tail. When iCa is increased by applying isoprenaline, the percentage augmentation ofthe associated tail current is much greater but occurs more slowly. Similarly, the tailcurrent recovers to its initial value more slowly than does ica.
6. These results are interpreted to indicate that the sodium-calcium exchangecurrent flows during the time course of the cardiac action potential and that itsamplitude is more closely related to intracellular calcium release than to the mem-brane calcium current per se. Calculation of the exchange current flowing duringrepolarization suggests that it reaches a peak of about - 150 pA at approximately200 ms. This compares to a net repolarizing current of the order 20 pA. The exchangecurrent should therefore make a significant contribution to the balance of ioniccurrent during the second half of the plateau phase of the action potential. The netcharge transferred would generate a calcium efflux sufficient to balance calciuminflux during the action potential.
INTRODUCTION
Following activation of calcium current (ica) in the heart, repolarization to poten-tials negative to about -35 mV results in slow inward current tails. These wereoriginally attributed to slow deactivation of ica itself. Work with patch clamprecording from isolated single cells shows, however, that ica deactivation occurs veryquickly at this range of potentials. The slow inward current tails that can be recordedin rat (Mitchell, Powell, Terrar & Twist, 1984a; 1987) and in guinea-pig (Fedida,Noble, Shimoni & Spindler, 1987 b) ventricular cells are therefore attributableto another mechanism. Since they can be abolished by buffering intracellularcalcium, by replacing external sodium with lithium, or greatly reduced by replacingcalcium with strontium, they have been attributed to activation of sodium-calciumexchange. Slow inward tail currents in amphibian cardiac cells have also been shownto be carried by the sodium-calcium exchange (Hume & Uehara, 1986; Campbell,Giles, Robinson & Shibata, 1988).Two important questions are raised by these results. When does the process
generating the inward current become activated during normal cardiac electricalactivity and what factors control the current amplitude? Hilgemann & Noble (1987)have shown that, in rabbit atrial muscle, the time course of calcium influx and efflux,inferred by measuring fast extracellular calcium transients (Hilgemann, 1986 a, b), isconsistent with the exchange process being activated during the time course of theplateau phase of the action potential and that it is active enough to generate acalcium efflux that in the steady state balances influx via calcium channels by thetime repolarization is complete. If the system is not in a steady state the magnitudeof the exchange current may be much less than the magnitude of iCa because much ofthe calcium entering the cell is taken up by the sarcoplasmic reticulum. This hypoth-esis, if it is also applicable to ventricular cells (as it seem to be; D. W. Hilgemann,personal communication), leads to the prediction that the slow inward tail currentsattributed to sodium-calcium exchange should be significant only during the timecourse of the action potential and that they should be related more to factorsinfluencing inotropic state, e.g. to the intracellular calcium transient, than to iCaitself. This situation would contrast markedly with that found in amphibian tissue,
640

Na+-Ca2+ EXCHANGE DURING ACTION POTENTIAL
where the exchange process largely pumps calcium out following repolarization andthe current carried is closely related to the time integral of calcium entry via thecalcium current channels (Campbell et al. 1988). We have therefore designed experi-ments to test these predictions. Some of the results described below have beenpreviously communicated at meetings of the Physiological Society (Egan, Noble,Noble, Powell & Twist, 1986a, b).
METHODS
Cell isolationSingle ventricular myocytes were obtained from adult guinea-pigs by enzymic dispersion as
described previously (Powell, Terrar & Twist, 1980: Mitchell, Powell, Terrar & Twist, 1984b:Egan, Noble, Noble, Powell, Twist & Yamaoka, 1988).
Methods and solutionsTwo kinds of perfusion system were used in this study. Both employed small Perspex chambers
mounted on the stage of an inverted microscope (Nikon Diaphot) and superfused with Krebs-Ringer solution at a rate of 1-5 ml/min. The composition of the normal Krebs-Ringer was(in mM): NaCl, 144; KCl, 5; CaCl2, 2-5; MgCl2, 1; NaH2PO4, 03; glucose, 5-6; HEPES, 5, bufferedto pH 7-4 using NaOH.
Perfusion system 1Here the superfusate was heated to 35-37 °C by means of a water-jacket which surrounded the
inflow tube of the perfusion bath. This water-jacket was connected to a circulating water pumphaving a variable temperature control. Thus, the temperature of the incoming superfusate wascontrolled by heat exchange between the water jacket and the solution inflow tube. Temperaturewas monitored continuously by a thermocouple probe placed in the centre of the recordingchamber. While it was easy to increase the bath temperature in a controlled fashion and tomaintain it at a stable value using this system, lowering the temperature within a reasonable timepresented problems because of the large volume of the water-jacket compared with the inflow tube.Thus, to lower the temperature effectively, the circulator had to be switched off and the water inthe water jacket replaced with air. The temperature then dropped fairly quickly but this processwas cumbersome and perfusion system 2 was much more convenient to use when repeated coolingand reheating was required.
Perfusion system 2This bath, like the first, had a thin glass cover-slip set into the base of the chamber to allow for
better adhesion of the cells. The inflow tube was surrounded by an encapsulated wire coil which wasused to heat the inflowing solution from 23 to 37 'C. A thermocouple and feed-back circuit regulatedthe temperature, which was displayed on a digital thermometer. When the power was switched offthe temperature rapidly cooled back to the ambient value. Outflow from the bath was finelycontrolled using a sloping groove-and-wick arrangement. The solution level could be kept constantand at a predetermined value by adjusting the height of the wick; this was a better arrangementthan in perfusion bath 1, where the fluid level tended to fluctuate continuously. Solutions werechanged simply by switching between reservoirs which supplied the baths by gravity feed. Thismethod resulted in application of known concentrations of drug to the myocytes. In addition,isoprenaline and cadmium were often applied in another way when rapid and repetitive fluidexchange was more important than the absolute concentration of drug in the cell vicinity. Amicropipette (tip diameter, 10-20 ,m) filled with a Krebs-isoprenaline (6 /tM) or Krebs-cadmiumchloride (1 mM) mixture was positioned in the superfusate above the cell under investigation andpressure ejected using brief (10-100 ms) pressure pulses (35-140 kPa) which released a few nano-litres of solution (see also Egan et al. 1988). While the absolute concentration of the drug at thesurface membrane is unknown using this method, its upper limit must be set by the concentrationof drug in the pipette (6 uM for isoprenaline or 1 mm for cadmium chloride). By comparing theresults with constant-flow superfusion with the same drugs we estimate that the effective con-
641
21l pH y 411

T. M. EGANAND OTHERS
centration applied to the surface membrane lies between 0-1 and 0 5 of the pipette concentration.Applied in this way, both drugs were completely reversible, wash-out taking a maximum of 1-5min. Chemicals and drugs used in this study included: (-)isoprenaline bitartrate, cadmiumchloride, strontium chloride, caesium chloride, tetrodotoxin, EGTA (Sigma); ryanodine was a giftfrom R. D. Vaughan-Jones.
ElectrophysiologyA single-electrode switch-clamp method (Axoclamp-2, Axon Instruments, Burlingame CA, USA)
was used in many experiments to record membrane current and voltage. The Axoclamp-2 was usedwith conventional microelectrodes filled with either 1-2 M-KCl, 0-5 M-K2SO4 or a mixture of 1 M-KCl and 100 mM-EGTA. When filled with 2 M-KCl these electrodes had DC resistances of 8-12 MQand no differences were noted in the experimental results obtained with these different fillingsolutions, despite the changes in intracellular activities which hypertonic pipette solutions canproduce (Blatt & Slayman, 1983; Desilets & Baumgarten, 1986).
In the remaining experiments, low-resistance suction micropipettes (1-5 MCI) fabricated fromsquare glass were used in conjunction with an Axopatch-IB amplifier (Axon Instruments). Sealresistances were > 1 Gil (cf. Hamill, Marty, Neher, Sakmann & Sigworth, 1981) and, generally,these electrodes needed no fire polishing. The normal internal solution for these micropipettes was(in mM): KCl, 150; NaCl, 5; MgCl2, 1; HEPES, 5, buffered to pH 7*35 with NaOH. In someexperiments KCI was replaced with equimolar CsCl in an attempt to block the delayed potassiumcurrent, iK.
Voltage and current were measured using the Axoclamp-2 switch amplifier at switching fre-quencies of 9-15 kHz (corresponding to a settling time of about 1 ms) and gains of > 2-5 nA/mV, and for the Axopatch-IB with a CV-3 amplifier headstage (gain 0-1 mV/pA) suitable forvoltage clamping large cells. In some experiments using picopump application of drugs (see Fig. 7)the clamp gain was reduced to minimize the risk of oscillation. Records of membrane current andvoltage were digitized and stored on a VCR-recorder (bandwidth DC to 20 kHz) for later analysis.Numerical reconstruction of the results was performed using the computer program OXSOFTHEART (DiFrancesco & Noble, 1985; Hilgemann & Noble, 1987).
RESULTS
Evidence obtained from manipulation of the action potentialUsing isolated guinea-pig myocytes, Fedida et al. (1987) found that voltage
clamp depolarizations from potentials positive enough to inactivate the fast sodiumcurrent, iNa, (i.e. near -40 mV) activated inward current tails which were clearlydistinct from the membrane current, ica. These experiments were performed atroom temperatures of between 18 and 22 °C and utilized conventional square clampprotocols. It seemed reasonable to assume that the depolarization produced by theventricular action potential itself might also switch on this current and we were,moreover, interested to see if we could record it at the more physiological tem-peratures of 35-37 'C. Figure IA and B shows a typical result. Above is the ven-tricular action potential having a duration of 203 ms and a maximum amplitude of105 mV at 35 'C. This action potential was interrupted after 20 ms and the mem-brane potential was clamped back to the resting potential (-65 mV in this case),when a large, inward current was recorded. This current had slow kinetics, takingalmost 100 ms to decay away from a peak amplitude of nearly 2 nA. Figure 1Cand D shows another example of the current, evoked by clamping 30 ms into anaction potential of 234 ms duration back to a resting potential of -76 mV. In thiscase the current changed little in amplitude for about 40 ms, before decaying withkinetics similar to those shown in Fig. 1 B. In different cells the amplitude of thiscurrent varied between as little as 100 pA to about 2 nA and was unaffected by
642
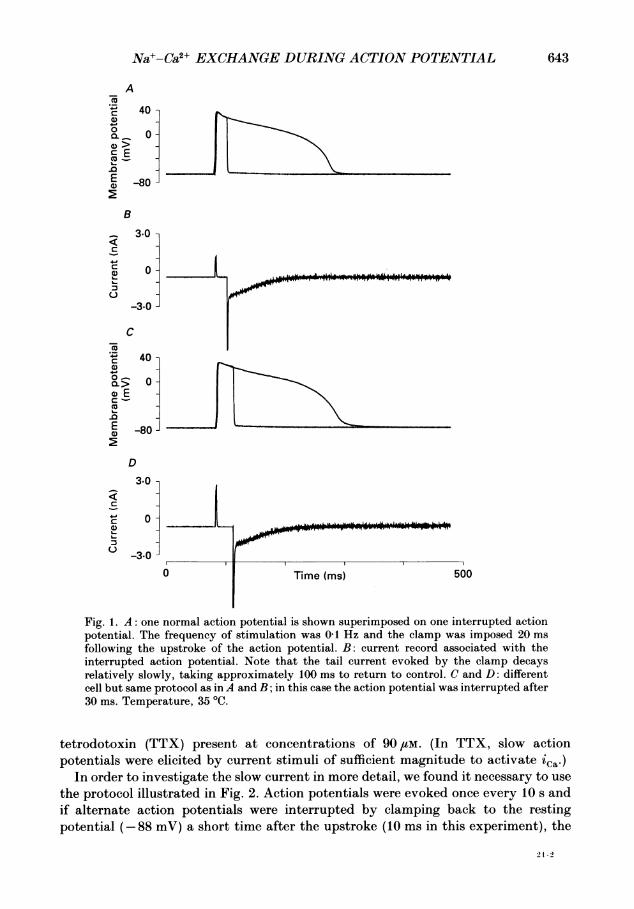
Na+-Ca2+ EXCHANGE DURING ACTION POTENTIAL 643
A
c 40-0-W 0-
-3-0 -
C0E -80 -
B
3.0 -
c~, 0
+-)
-3.0 -
C
+-I
c 40-0)
> 0 -
C
.0
E, -80
D
3.0
c 0-
)-30
0 | Time (ms) 500
Fig. 1. A: one normal action potential is shown superimposed on one interrupted actionpotential. The frequency of stimulation was 0-1 Hz and the clamp was imposed 20 msfollowing the upstroke of the action potential. B: current record associated with theinterrupted action potential. Note that the tail current evoked by the clamp decaysrelatively slowly, taking approximately 100 ms to return to control. C and D: differentcell but same protocol as inA and B; in this case the action potential was interrupted after30 ms. Temperature, 35 'C.
tetrodotoxin (TTX) present at concentrations of 90 /tM. (In TTX, slow actionpotentials were elicited by current stimuli of sufficient magnitude to activate ica-)
In order to investigate the slow current in more detail, we found it necessary to usethe protocol illustrated in Fig. 2. Action potentials were evoked once every 10 s andif alternate action potentials were interrupted by clamping back to the restingpotential (-88 mV) a short time after the upstroke (10 ms in this experiment), the
21-2

T. M. EGANAND OTHERS
current labelled with the filled circle in Fig. 2 was recorded. Interruption of everyaction potential, however, activated the much smaller current indicated by the opensymbol. Further investigation showed that if between 120 and 150 ms (correspondingto the plateau phase) of the action potential was allowed to occur before activatingthe clamp, repriming for the slow current tail would be complete. Action potentials
A
B I
0-
40 mV \ _A[nAi II ._IU
100 ms 100 ms
Fig. 2. A illustrates the experimental protocol. Either alternate action potentials wereinterrupted (@) or every action potential (0), by applying the voltage clamp commandat a predetermined time after the upstroke. B shows an action potential which is inter-rupted after 10 ms (left), and current records for the two sequences illustrated above(right). Note that the current evoked by regular interruption (0) has a time course similarto those shown in Fig. 1, whereas alternate clamping back evokes a much larger current(@) which takes about 30 ms to reach the peak negative level before decaying to zero atabout 120 ms. Frequency of stimulation, 01 Hz. Temperature 35 'C.
continually interrupted after intervals shorter than this could not reprime fully andwere accompanied by current tails of reduced amplitude, thereby suggesting thatsome intracellular process associated with prolonged depolarization must modulatethe response.The time for full repriming appeared to coincide closely with the time after which
no inward tail current could be recorded. Thus, action potentials interrupted after150-200 ms during repolarization were seldom associated with measurable slowcurrent tails. As already mentioned above, we also observed that in a number of cellsrather little inward current could be recorded at any time after the action potentialupstroke when the stimulus frequency was as low as 01 Hz. This is shown to the farleft of Fig. 3, where the duration between upstroke and activation of the clamp was5 ms. Here the current recorded in the control state was only about 300 pA, which
644

Na+-Ca2+ EXCHANGE DURING ACTION POTENTIAL 645
C0CD
0e
~4-Dc Oebo - ;
o.Q
to
4_DO
o
o_4a OQ
ri2
bt0 N4
<I 'C
C3S
Q.' o0,=;"0w;x
Aj,4 0 + 0
ECD
At ~~~~~~~~~~~~~~~I
0
.-0
oCd
0
a
cn
0
CA4+ac13
NI
C4"a0
coCD

T. M. EGANAND OTHERS
appears very small at the gain employed in the experiment shown. However, in suchcases we frequently observed that repetitive stimulation at a higher frequency couldsubstantially alter the amount of slow current available and that this effect washighly cumulative over relatively short bursts of activity. Thus, Fig. 3 also shows theresults of clamping 5 ms into the first action potential triggered 10 s after the cell hadbeen subjected to 60, 120 and 240 suprathreshold stimuli delivered at a frequency of2 Hz. It can be clearly seen that the amount of slow current activated was alreadymuch greater after the first sixty beats (just under 2 nA) and progressively increasedto over 3.5 nA after the longest stimulus train.
A
0_rmV
I 50 mV
'50 ms '
500 pA
Fig. 4. Effect of isoprenaline on action potential duration and slow inward tail currentamplitude. A: action potentials were evoked at 0 1 Hz and alternately interrupted 10 msafter the upstroke (cf. Fig. 2). Isoprenaline was ejected from a pipette placed adjacent tothe cell (see Methods; pipette concentration, 6 ,M) and the action potential increased oversuccessive beats to a maximum (trace marked 0). The isoprenaline was then washed outduring the next six beats and action potential duration returned to the control level (tracemarked 0). B: tail currents evoked during the alternate action potentials also decreasedfrom maximum (@) back to control (0). Temperature 36 'C.
Figure 4 shows the result of an experiment in which we investigated the effect ofisoprenaline on the tail current. In this case, drug was ejected from a pipettecontaining 6 ,UM (see Methods) and the duration of the action potential increased to357 ms (-, Fig. 4A). Over the next six stimuli (pulse interval 10 s), the actionpotential returned to the control duration of 285 ms (0, Fig. 4A). During alternatebeats, when the action potential was interrupted by clamping back to the restingmembrane potential of -86 mV after 10 ms, the evoked slow current decayed inamplitude as the drug was washed out (cf. Fig. 4B).
Influence of duration of depolarizationThe results shown so far involve interrupting the action potential within the first
5-50 ms to reveal the full time course of the tail current. It is important also to know
646
Na+-Ca2+ EXCHANGE DURING ACTION POTENTIAL
how the time of interruption influences the amplitude and time course of the tailcurrent. Thus, if the process that controls the tail current is simply triggered by theaction potential but not itself controlled by it, then the tail currents followinginterruptions at different times should superimpose, whereas if the action potentialitself continues to control the activating process, as is the case for the controlof contraction (Morad & Trautwein, 1968), then the tail currents should not besuperimposable.
A B
1 nA[
1-
_ I, IQ I
O4 -1_
0 200 400Time (ms)
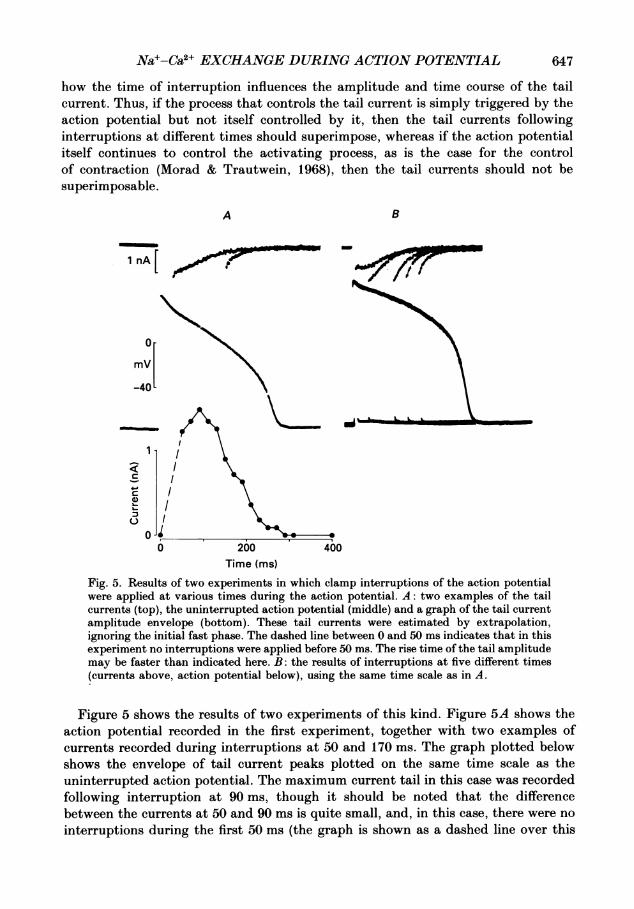
Fig. 5. Results of two experiments in which clamp interruptions of the action potentialwere applied at various times during the action potential. A: two examples of the tailcurrents (top), the uninterrupted action potential (middle) and a graph of the tail currentamplitude envelope (bottom). These tail currents were estimated by extrapolation,ignoring the initial fast phase. The dashed line between 0 and 50 ms indicates that in thisexperiment no interruptions were applied before 50 ms. The rise time of the tail amplitudemay be faster than indicated here. B: the results of interruptions at five different times(currents above, action potential below), using the same time scale as in A.
Figure 5 shows the results of two experiments of this kind. Figure 5A shows theaction potential recorded in the first experiment, together with two examples ofcurrents recorded during interruptions at 50 and 170 ms. The graph plotted belowshows the envelope of tail current peaks plotted on the same time scale as theuninterrupted action potential. The maximum current tail in this case was recordedfollowing interruption at 90 ms, though it should be noted that the differencebetween the currents at 50 and 90 ms is quite small, and, in this case, there were nointerruptions during the first 50 ms (the graph is shown as a dashed line over this
647

T. M. EGAN AND OTHERS
period). Figure5B shows the results of another experiment in which interruptionswere applied at 5, 30, 90, 120 and 150ms. It is clear that the envelope of tail peaksfollows a similar time course to that shown in Fig. 5A. It is also clear that inter-ruptions during the first 100 ms lead to tail currents that do not superimpose.This kind of result was obtained in all cases where the interruptions occurred beforeabout 100-150ms. Later interruptions, though, did give rise to superimposable tailcurrents. This implies that the process controlling the tail current amplitude dependson the maintenance of the action potential plateau for a period between 100 and150 ms and that about 50% of the tail amplitude is independent of the time ofinterruption, i.e. the tail amplitude obtained after the shortest interruptions (5ms)can be as large as 50-80 % of the maximum obtained by interrupting later during theplateau. Thus, the process controlling the tail current amplitude is in great parttriggered in a regenerative manner even by very short depolarizations, while theremaining part is controlled by continued depolarization.
The effect of calcium removal on the tail currentWe found that replacement of extracellular Ca2" with equimolar concentrations of
Sr2+ virtually abolished the slow inward tail current recorded during the actionpotential. Figure 6 shows such a result. In the top panel are displayed a normalaction potential, together with a typical 'strontium' action potential with its muchlonger duration and lower plateau. The second panel shows the slow tail current inthis cell recorded in normal Krebs-Ringer solution by clamping 5 ms into the actionpotential. This is an example of a cell showing a virtually maximal current for thisstimulus frequency (0 1 Hz: alternate action potentials interrupted) of about 2 nA.The third panel, labelled with a star, shows the equivalent steady-state recordobtained in 2-5 mM-strontium-containing Krebs-Ringer solution, where it can beseen that the slow current tail has almost completely disappeared. Finally, thebottom panel illustrates the return to control conditions: the current tail afterstrontium wash-out being only slightly smaller than it was at the beginning of theexperiment.One way to remove intracellular Ca2` is to buffer it with, for example, EGTA.
When using electrodes containing 100 mM-EGTA we found it impossible to recordslow current tails by any technique and this was true in all cell populations wheremyocytes penetrated with normal electrodes showed large inward tail currents. Theeffectiveness of intracellular EGTA diffusion was also assessed visually by observingthe failure of the myocytes to contract in response to suprathreshold stimuli. Otherworkers have previously found that the large tail currents recorded at room tem-perature in guinea-pig ventricular cells or in rat ventricular myocytes at 37 °Cwere abolished in response to intracellular diffusion of EGTA from a second micro-electrode (Fedida et al. 1987 b; Fig. 9: Mitchell et al. 1984 a; 1987).
Finally, we have also used ryanodine to attempt to remove the calcium transientsince this drug has already been successfully used to abolish the inward current tailin rat ventricular myocytes (Mitchell et al. 1984b). In guinea-pig cells this drug hasa more complex action and at 1 ,UM it is ineffective in removing either the contractionor the tail current (Fedida et al. 1987 b). However, we find that in a dose range of8-12 /tm ryanodine abolishes at least 80% of the inward tail current, allowing us to
648
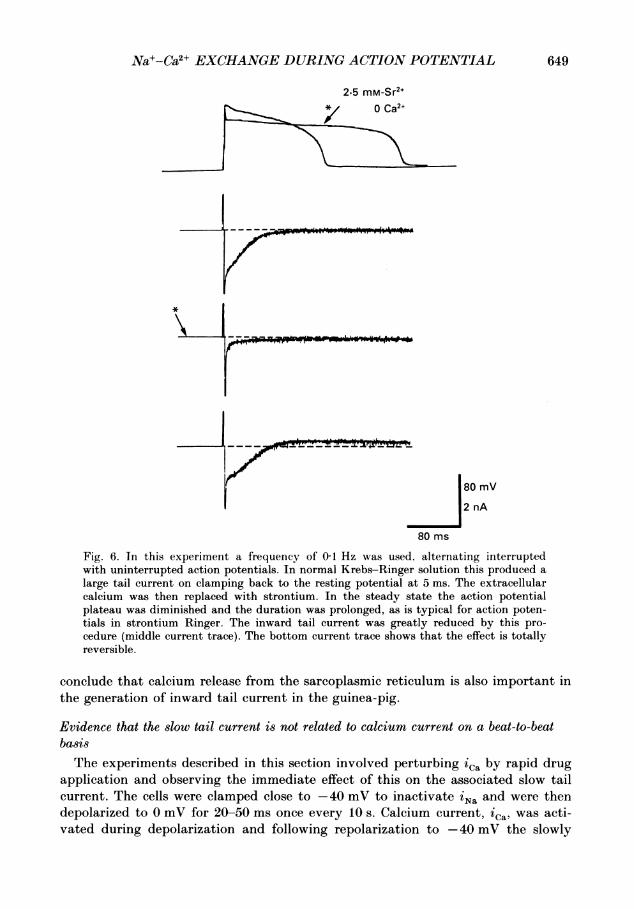
649Na+-Ca2+ EXCHANGE DURING ACTION POTENTIAL
2.5 mM-Sr2+{> ~*/ 0 Ca2+
80 mV
2 nA
80 ms
Fig. 6. In this experiment a frequency of 01 Hz was used, alternating interruptedwith uninterrupted action potentials. In normal Krebs-Ringer solution this produced alarge tail current on clamping back to the resting potential at 5 ms. The extracellularcalcium was then replaced with strontium. In the steady state the action potentialplateau was diminished and the duration was prolonged, as is typical for action poten-tials in strontium Ringer. The inward tail current was greatly reduced by this pro-cedure (middle current trace). The bottom current trace shows that the effect is totallyreversible.
conclude that calcium release from the sarcoplasmic reticulum is also important inthe generation of inward tail current in the guinea-pig.
Evidence that the slow tail current is not related to calcium current on a beat-to-beatbasisThe experiments described in this section involved perturbing 'Ca by rapid drug
application and observing the immediate effect of this on the associated slow tailcurrent. The cells were clamped close to -40 mV to inactivate iNa and were thendepolarized to 0 mV for 20-50 ms once every 10 s. Calcium current, ica, was acti-vated during depolarization and following repolarization to -40 mV the slowly
.1- - -
- --,-
--.- -- --

650
X 40-C
0
o 0 -
0. -~
0 >
c E
E
> -80-
2.0 -
0c
40
0
L-
-6.0 -
cC
-
2.0 -
0 -
-6.0 -
T. M. EGAN AND OTHERS
A
B
c
0 100 200Time (ms)
Fig. 7. Currents (B and C) in response to depolarization from -40 to 0 mV (A) for 50 msat 01 Hz. In each case, after obtaining the control record (0) an ejection from anextracellular electrode containing 1 mM-cadmium chloride was applied using a suddenpressure pulse 2 s before the application of the voltage clamp pulse. In B this was sufficientto produce a 40% block of 'Ca, whereas in C 80% block was produced. In both casesthe slow tail current on return to -40 mV was almost unaffected. Note that in thisexperiment there is a significant voltage sag during the flow of ica itself. This is attribu-table to the low clamp gain required in these experiments to prevent oscillation whenthe capacity is greatly increased by the presence of the picopump electrode (see Methods).There is, however, negligible sag during the flow of tail current.
decaying inward current was recorded. Once stable control records were obtained,pressure ejection of the calcium channel blocker, cadmium chloride, was applied 2 sbefore the next voltage step. We estimate that this produced an almost instan-taneous concentration of between 041 and 1 mm in the vicinity of the cell membrane(see Methods). An example of the effect of this manoeuvre is shown in Fig. 7. As canbe seen from the two examples shown in Fig. 7, cadmium caused a very significant
P*-Poo V -, '104"10.
I

Na+-Ca2+ EXCHANGE DURING ACTION POTENTIAL
block of iCa (0) compared to control (0) without affecting the size or time course ofthe associated slow current tail. Visual inspection of the cell under these conditionsindicated that contraction was still taking place, but at present we have no quanti-tative data on whether cell shortening was modified by the application of cadmium.Responses such as those shown in Fig. 7 were seen in about 50% of the cellsstudied. In the remainder of the experiments both ica and itail were attenuatedon exposure to cadmium. However, it is worth noting in this respect that iNaca(the sodium-calcium exchange current) is also blocked by cadmium in a dose-dependent-manner (Kimura, Miyamae & Noma, 1987; L. Pott, personal communi-cation). The results shown in Fig. 7 can be achieved only by adjusting the cadmiumdose to achieve differential block.A clear advantage of the pressure ejection method of drug application used here
is that it results in almost instantaneous drug exposure. Thus, the time relationbetween associated events can be determined very precisely. This feature is evenmore obvious in the experiment shown in Fig. 8, where the drug in the micropipettewas isoprenaline at a concentration of 6 /LM. The voltage protocol was exactly thesame as in the cadmium experiments and the effect of isoprenaline is shown 12, 22and 32 s post-ejection. It can be seen that after 22 s, the effect of isoprenaline oniCa was maximal and the amplitude of the tail current was also substantiallyincreased. At 32 s, the amplitude of iCa was beginning to fall but the slow currenthad continued to increase. On wash-out the steady decay of ica preceded that of itaii.Notice also that the proportionate change in the tail current is very much larger thanthat of iCaThe time relations for this behaviour are shown graphically in the bottom panel of
Fig. 8. Here the closed symbols represent ica and the open symbols itail, each plottedas a percentage change from control against time from application of the drug. Atthe onset of the experiment the size of the calcium current was 4-7 nA, while theassociated tail current had a magnitude of about 550 pA. There was then an increasein iCa by 41 % to 6-6 nA at 22s, whereas itail increased to 322 % or to around1-8 nA at 32 s. Both currents then decayed, but not completely, back to their controlvalues. In the case of iCa' the slight increase to 5-2 nA measured 92 s after drugapplication appears to represent a real, small change in calcium conductance of themembrane. This increase is probably reflected in the increase in tail size of 790 pAmeasured at 82 s. However, the upswing in the magnitude of the slow tail current to860 pA at 92 s may well indicate the start of a definite change which we frequentlysee in our myocytes as they age: i.e. the tail current can become quite substantiallylarger with time. (When this occurs the general shape of the action potential isusually unaltered.) The main trend is, however, clear from Fig. 8. There was a phaseshift to peak activation of ica compared with itaii of about 10 s (that is, of onestimulus interval), the tail current was augmented much more than the membranecalcium current and returned relatively more slowly to a final, stable value. We haveobtained a similar result in five other cells.
Masking of 'tail by the delayed potassium current, iKWe have observed that the slow tail current can be more difficult to record at
physiological temperatures of 35-37 °C than at room temperature (18-22 °C). The
651

T. M. EGAN AND OTHERS
A12 s _o't
22 s
32 s
Phase shiftbetween peaks (ca. 10 s)
12 22 32 42 52 62Time (s
V1
4 nA
20 ms
0
72 82 92 10,2Fig. 8. A: ionic currents recorded in response to depolarizing pulses from -40 mV to0 mV at a frequency of 0-1 Hz after exposure of the cell to a brief application of 6 ftM-
isoprenaline from an extracellular pressure ejection pipette. The traces show responses at12, 22 and 32 s after drug application. In each case the record is superimposed on thecontrol record. B: the results are plotted as current amplitudes as a percentage of controllevel as a function of time. There is a moderate increase in iCa which is followed by a very
much larger relative increase in tail current. Note that the peak augmentation of iCa
occurs 10 s before the peak augmentation of the tail current.
disappearance of the slow tail at higher temperatures tends to run in cycles so thatwe may fail to see it for prolonged periods of time and then find that it is againpresent and easy to record during an equally long train of experiments. No differencein cell isolation techniques accompany this change, although the fact that it doesoccur suggests that the same enzyme preparations can yield myocytes whose mem-
brane properties vary. One such variable appears to be the extent to which thedelayed potassium current, iK, is activated by short depolarizations to positivepotentials. Since this current is outward at potentials positive to EK, its deactivation
B
0L-
cJ0
+-
C40az
652
)

Nat-Ca2+ EXCHANGE DURING ACTION POTENTIAL 653
at -40 mV could mask the slow inward current tail producing a flat, or evenoutward, record. Figure 9 shows results obtained using a patch clamp electrode in thepresence of05 ,uM-isoprenaline. It was experiments of this kind which first suggestedto us that iK involvement could be relevant to this problem. Under control conditionsat 24 °C (panel A), ica recorded in response to square voltage clamp depolarizationsof 50 ms to + 10 mV from -40 mV was followed by a small, inward current tail.
A B
1 nA
/ 100 ms
C D E
Fig. 9. Membrane currents in response to depolarizing pulses from -40 to 0 mV at 24 °Cbefore (A) and after (B) application of 0-5 /SM-isoprenaline. Note increase in both ic. andtail current. The temperature was then increased to 27 °C (C), 30 °C (D) and 37 °C (E).The inward current tail at 24 °C becomes outward at the higher temperatures.
Both ica and itail became much larger during steady-state perfusion with 0 5 ,UM-isoprenaline at 24 °C: ica increased from 3 nA and itaij from 500 pA to almost 1 nA.The temperature was then increased using perfusion system 2 (see Methods) andthe slow tail gradually reversed in direction until at 37 °C it became markedlyoutward (panels C-E). It is well known that isoprenaline increases activation of boththe delayed rectifier current, iK and the membrane calcium current, ica (Brown &Noble, 1974; Walsh, Begenisich & Kass, 1988). Here, however, it seems clear thatthere must also be a substantial effect of temperature on the level of activation of thetime-dependent potassium current, at least in mammalian ventricular cells, whichallows it to appear in this experiment as an outwardly decaying current despite thelarge inward increase in itai associated with the augmented ica (see also Walsh et al.1988). This same result was later obtained in normal Krebs-Ringer solution butthe tail currents recorded never became net outward following the pulse durationsused in our experiments. Nevertheless, inward tail currents present at about 22 °Cfrequently became flat or virtually flat at 35-37 'C.
Figure 10 shows results from an experiment in which all intracellular and extra-cellular KCl had been replaced by CsCl in an attempt to eliminate the problem

654 T. M. EGAN AND OTHERS
described. Here the protocol used was identical to that described in association withFig. 9. In Fig. 1OA it can be seen that there was a fairly large inward slow tail currentat 26 °C (0-36 nA), which increased to 0-4 nA at 30 °C as shown in B. The kinetics ofdecay were, however, faster at the higher temperature and became most rapid at37 °C as can be seen in C. This result was obtained in all four cells studied in this way.We conclude, therefore, that iK decay tails can significantly mask 'NaCa tails undersome circumstances and that it is then important to eliminate iK before analysingchanges due to iNaCa.
A B |C
0.4 nA
40 ms
Fig. 10. Membrane currents in response to depolarizing pulses from -40 to 0 mV aftersubstituting caesium for potassium ions. The tails are now inward at all temperatures:26 'C (A), 30 'C (B) and 37 'C (C). Note that ic. also increases as the temperature increasesas previously shown by Cavalie, McDonald, Pelzer & Trautwein (1985).
DISCUSSION
Is the tail current activated by [Ca2+]i?Our results show that the slow inward tail current that can be recorded at the
resting potential by interrupting the action potential of guinea-pig ventricular cellsbehaves in a manner that is consistent with it representing the response of thesarcolemmal sodium-calcium exchange process to variations in intracellular freecalcium ([Ca2+]i). Thus, like the tail current following voltage clamp depolarizations(Fedida et al. 1987 b), it is abolished by intracellular calcium buffering and is greatlyreduced by replacing extracellular Ca2+ with Sr2+ ions. In the steady state, it isaugmented by increasing intracellular calcium with isoprenaline (Fig. 4), which notonly increases ica but also the storage and release of calcium from the sarcoplasmicreticulum. Our results also show that maintaining ventricular rhythm is important.Cells that have been quiescent for long periods and which may be hypodynamic canshow very little tail current, and it is then necessary to use a train of repetitivestimulation to enhance the tail current (cf. Fig. 3).
Since we record very large inward current tails by interrupting the action potentialafter only 5-10 ms, it would appear that only a very short depolarization is requiredto activate a large part of the calcium release and that the response of the exchangerto changes in [Ca2+]i is rapid. Recent measurements of [Ca2+]i, using the calcium-sensitive indicator Fura-2, do indeed show a rise to peak of the calcium transient

Na+-Ca2+ EXCHANGE DURING ACTION POTENTIAL
within about 50 ms (Barcenas-Ruiz & Wier, 1987; Wier, Cannell, Berlin, Marban &Lederer, 1987; Cannell, Berlin & Lederer, 1987). This rise time is similar to theduration of depolarization required to activate the maximal slow inward current onrepolarizing to the resting potential (about 50 ms in most cells). Nevertheless, it isdifficult to ascribe a time when the slow inward current tail is at its peak immedi-ately following depolarizations. As shown in Figs 1-5, it characteristically requires10-20 ms more to reach its maximal amplitude or hovers around the same value forthis period of time before starting to decay. This does not seem to be an artifact, sinceclamps at later times into the action potential do not give this type of record, the tailcurrent then declining in a single-exponential manner. The implication is that [Ca2+]iis still rising at the time of repolarization at 5 ms and that, once triggered, therelease continues even after repolarization. This interpretation is supported byMorad & Trautwein's (1968) study of the relation between duration of the actionpotential and the time course and amplitude of contraction. They found that a verylarge fraction of the total contraction could be obtained even when the actionpotential was interrupted only a few tens of milliseconds after the upstroke andconcluded that 'the development of tension can go on after the membrane isrepolarized to the resting state'.Our results are, therefore, consistent with the view that the peak tail current is
activated at about the same time as the peak value of [Ca2+]i and that any delaybetween the increase in [Ca2+]i and the onset of iNaca is in the order of millisecondsonly. The time course of the envelope of tails also fits this interpretation. Thus, tailcurrents are very small or negligible after 150-200 ms as is the calcium transient(Wier et al. 1987; Cannell et al. 1987), Note, however, that the time course is notindependent of the interruption. A tail current can still be obtained by interruptingthe action potential after the time at which the tail current following rapid inter-ruptions has decayed to zero. Moreover, the tail currents following interruptionsat different times do not superimpose (see Fig. 5). This implies that the calciumtransient is shortened by interruption and it fits with Morad & Trautwein's (1968)finding that the duration of contraction increases with the length of the actionpotential.The simplest interpretation of our results is, therefore, that the tail current is
carried by iNaCa which responds to the time course of the [Ca2+]i transient. Thishypothesis is strongly supported by the results of Barcenas-Ruiz, Beuckelmann &Wier (1987) but of course we cannot exclude the involvement of other calcium-activated channels such as the non-specific cation channel, particularly since Ehara,Noma & Ono (1988) have shown that intracellular calcium concentrations in theregion of 15/tM can significantly activate non-specific current flow. However, otherevidence suggests that the sodium-calcium exchange current is activated at lowerintracellular calcium concentrations than that required to open cation channels. Thismight be attributable to the fact that iNaCa is nearly linearly dependent on [Ca2+]iwhereas the channels show activation with a Hill coefficient of 3. Thus, the transientinward current in guinea-pig ventricular cells displays no reversal potential nearzero membrane potential, which is the behaviour expected of the sodium-calciumexchange mechanism (see Fedida, Noble, Rankin & Spindler, 1987 a; Fig. 7), yetthe tail currents recorded here are no larger than the transient inward current.
655

T. M. EGAN AND OTHERS
Moreover, in atrial cells, Mechmann & Pott (1986) have shown that the inwardcurrent first activated by a rise in [Ca2+]i shows no channel noise. Channels onlybecome evident when the non-channel current is already strongly activated.
The time course ofiNaca during the action potentialAt its peak, the tail current amplitude is most usually in the range -0 5 to -1 nA
at -80 mV, although larger currents can sometimes be recorded (see e.g. Fig. 6).It can even reach- 35 nA after long trains of action potentials at 2 Hz. This is a verysubstantial current indeed for cells which generate a peak ica of up to -5 nA, but itis not the amplitude of tail current which would flow during the plateau of the actionpotential. To estimate that amplitude we can use the current-voltage relations foriNaCa recently obtained by Kimura et al. (1987) and by Lipp & Pott (1988). These fitthe equation foriNac. used in the DiFrancesco-Noble (1985) model (see Noble (1986)for an explanation of this equation). Roughly, the inward mode of the exchangeshows an exponential dependence on voltage with a 10-fold change in current overabout 120 mV. From this we can estimate scaling factors for the inward mode of thecurrent at +40 and 0 mV given the current that is recorded at -80 mV. These are01 for +40 mV and 0 3 for 0 mV. Thus, if the inward mode of the exchange carriesa peak current of - 0-5 to -1 0 nA at -80 mV, as in the majority of our experi-mental results, than it would be expected to carry -50 to -100 pA at + 40 mV and-150 to - 300 pA at 0 mV. Part of this current will be offset by the outward modeof the exchange. To calculate both modes and therefore the net current, we also needestimates of [Ca2+]i
In Fig. 11 we have taken this approach a stage further by using the experimentaldata of Fig. 5A to compute the time course of the exchange current and its equi-librium potential during the action potential. First, we converted the tail currentamplitudes to estimates of [Ca2+]i using a scaling factor of 1 nA/,UM for the relationbetween [Ca2+]i and tail current at the resting potential. This scaling factor is basedon the measurements of Kimura et al. (1987) showing that 430 nM [Ca2+]i generatesabout -0-5 nA of exchange current at -80 mV. We then used the DiFrancesco-Noble equation for the exchange current to calculate the current at each timeduring the plateau, using the experimental value of the membrane potential, theestimate of [Ca2+]i and with [Ca2+]o = 2 mm and [Na+]. = 140 mm. Values of [Na2+]ibetween 5 and 15 mm were assumed. The results plotted in Fig. 11 were based onusing [Na+]i = 7 mm. Finally, when the tail current amplitude was negligible, weassumed a basal level of [Ca2+]i to be 20 nm. This is in fact the resting level in theHilgemann-Noble (1987) model and is much lower than the values estimated experi-mentally. It would not, however, alter the main conclusions concerning the ampli-tude and time course of iNaCa during the action potential to choose higher valuesfor the resting [Ca2+]i; this would merely displace the resting level of ENaCa toless negative values and increase the resting level of inward current.
It is striking that, considering the very basic asumptions used in converting thetail current data, the results plotted in Fig. 11 show a remarkable similarity to theexperimental measurements of [Ca2+]i in that the peak value is 1-2 /IM and the timecourse is similar to that monitored using fluorescent probes (Wier et al. 1987; Cannellet al. 1987). The results in Fig. 11 also show that, with these values, the equilibrium
656

Na+-Ca2+ EXCHANGE DURING ACTION POTENTIAL
A
>E0)
o
B0-
CL.-i
-100-C-
-200 r-
657
11
Jo
iNaCa
u Time (ms) 400
Fig. 11. The time course of the equilibrium potential for the sodium-calcium exchange(ENaCa) and of the exchange current ('NaCa) was calculated from the experimental resultsof Fig. 5A. [Ca2+]i was estimated from the envelope of current tail amplitudes assuminga conversion factor of 1 ,jM/nA (Kimura et al. 1987). The equilibrium potential andexchange current were then calculated using the DiFrancesco-Noble (1985) equation with[Na+]i set to 7 mm and the basal (minimum) level of [Ca2+]i set to 20 nm. Other valuesof [Na+]i were also used (see text). The dashed line is used to show that the precise timecourse is uncertain between 0 and 50 ms since no interruptions were applied during thisinterval. This uncertainty does not however greatly influence the exchange currentestimate, which is very low at the beginning of the plateau, despite the high level of[Ca2+]1. The computed exchange current greatly increases as repolarization occurs. Thiseffect reflects the steepness of the current-voltage diagram for the exchange current. Notethat it is responsible for greatly displacing the time of the peak value of current comparedto the peak value of [Ca2+]i.
PHY 41122

T. M. EGANAND OTHERS
potential for the exchange would be positive to the plateau potential and that the netexchange current would therefore be inward during most of the plateau, reaching apeak of about - 150 pA at approximately 200 ms. When [Na+]i was assumed to be5 mm, the exchange current had the same time course but increased to - 200 pA. At[Na+]i = 11 mm, the calculated exchange current was close to zero. To determinethe possible significance of these estimates, it is important to compare them withestimates of the net ionic current and of individual ionic currents during the plateauphase of the action potential. For the net ionic current (ii) we can use the equation:
-i = CdV/dt, (1)
which, with a cell capacitance (C) of about 200 pF and a rate of repolarization(dV/dt) of 0-1 mV/ms, gives a net inward current of only 20 pA. The first conclusionto draw, therefore, is that the peak value of the inward mode of iNac. could liebetween 2 and 10 times the net repolarizing current. This is certainly not negligible.Thus, it is possible that iNaca could contribute a significant current to the balance ofcurrents that produces a net ionic current during the plateau of 20 pA. However, thisfact alone does not answer the question of how much difference iNaca might make byitself to repolarization. The reason for this is that the precise amplitudes of otherplateau currents are also uncertain. Unlike the rat action potential, which dis-plays a late, low plateau attributable almost entirely to sodium-calcium exchange(Mitchell et al. 1984a-c; 1987), the guinea-pig action potential does not normallyshow separate phases of repolarization. Moreover, as Hilgemann & Noble (1987) haveshown, it is not possible to estimate the direct contribution of iNaca to the actionpotential by comparing responses with large and small calcium transients, as forexample during paired-pulse stimulation. The reason for this is that the changes ininactivation of ica in response to variations in [Ca2+]i are much larger than thoseof the exchange current at positive potentials. The early phase of repolarizationtherefore slows when the [Ca2+]i transient is small, which is in the opposite directionto that expected from the change in iNaCa. This fact is sometimes used to argueagainst the role of iNaCa being significant, but the argument is invalid withoutknowing what the change in ica would have produced in the complete absence ofiNaCa. Further experiments are therefore required to determine the precise roleof iNaCa compared to iCa5 iK and other current components. All we can say withreasonable certainty is that the exchange current would not be negligible during theplateau and that it must therefore delay repolarization to some degree, althoughprobably not as dramatically as in rat ventricular cells, where the exchange currentis virtually entirely responsible for maintaining the late plateau.We can, however, be more certain about the fluxes that would be generated.
By integrating the time course of iNaca, we obtain a net charge transfer of 20 pC(an average of 100 pA flows for 200 ms). This would correspond to a net exit ofcalcium carrying 40 pC of charge and would be quite sufficient to balance a net entryof calcium via the calcium channels, assuming that these carry an average current ofabout 500 pA of current for about 80 ms on clamping to the level of the plateau(between 20 and 40 mV). These estimates are, of course, only to be treated as ordersof magnitude, but they are consistent with the conclusion of Hilgemann & Noble
658

Na+-Ca2+EXCHANGE DURING ACTION POTENTIAL 659
(1987) that the sodium-calcium exchange activity during repolarization could besufficient to restore calcium balance before the end of the action potential.
Relation between iCa and iNaCaThe question of the relations between iCa, Ca2+ release, contraction and calcium
efflux via the exchange process is fundamental to a full understanding of exci-tation-contraction coupling in the heart. If the exchange is the main process byA 10
0
flC>, O-
00CL
)C-100
70-
C 05-
10 50 Time (is) 100
Fig. 12. Effect of block or augmentation of ica on voltage clamp currents computed usingHilgemann-Noble (1987) equations for intact rabbit atrium. A: voltage. B: currentsduring normal, 80% block and 100% increase of iCa' C: variations of intracellularfree calcium concentration. It is assumed that the block or enhancement of iCa occursimmediately before each pulse and the computation lasts in each case for only one cycle.Maintained block or enhancement would alter the calcium stored in the model andeventually this would be reflected in reduced or enhanced tail currents. Note that in thismodel the scaling factor for activation of iN8ca by [Ca2"], is close to that measuredexperimentally and used in the calculations shown in Fig. 11.
which calcium efflux occurs then, in the steady state, there will be a strict relationbetween iCa and iNaCa (Hilgemann & Noble, 1987). Since each calcium ion entering viathe channels carries two units of charge, while each one leaving via the exchangerwill generate a net inward current equivalent to one charge, the steady-state timeintegrals of ica and iNaca would be in the ratio 2: 1. However, in non-steady-stateconditions this relation may not hold. Hilgemann & Noble (1987) have shown thatit does break down during paired-pulse stimulation and at the beginning of high-frequency trains, when the calcium current can temporarily generate influx verymuch greater than efflux via the exchanger. This in turn leads to the cell loading withcalcium. Our results show that the opposite departure from balance is also possiblesince, during rapid block of ica, iNaca is initially unaltered (Fig. 7). Arlock & Noble

T. M. EGAN AND OTHERS
(1985) have described a similar result in ferret ventricular muscle. Similarly, rapidenhancement ofiCa only increasesiNaca after a time delay (cf. Fig. 8).
It might be thought that these results are inconsistent with the calcium-inducedcalcium-release hypothesis. However, this is not necessarily the case. First, itshould be noted that such a mechanism must of necessity be non-linear, since thereis the possibility of positive feed-back between [Ca2+]i and calcium release. It maythen require only a relatively smalliCa to initiate the process. Second, we cannotexclude the possibility that calcium may act by catalysing a voltage-dependentprocess, as represented in the Hilgemann-Noble (1987) model. In this connection itis interesting that this model gives precisely the result we obtain experimentallywhen the magnitude of ica is suddenly changed. This is shown in Fig. 12, where theeffect of blocking iCa by 80% or increasing it by 100% on the predicted calciumrelease and the inward tail generated byiNaca can be seen. This degree of pertur-bationofiCa alters the peak calcium transient by only 5%, while the resultingexchange tails almost superimpose, as they do in our experimental results (Figs 7and 8). The insensitivity of the tail current and [Ca2+]i to the amplitudeoficareflects the fact that with the known intracellular buffering power in cardiac cells(e.g. by troponin and calmodulin), the change in [Ca2+]i attributable to calcium entrycan be very small. What then matters is the magnitude of the calcium release fromthe reticulum. This appears to depend more on how much calcium is already storedthan on the calcium triggering its release.
This work was supported by grants from the British Heart Foundation, Medical ResearchCouncil, National Institutes of Health (Heart and Lung Institute) and the Wellcome Trust. Thecomputations done in this study were carried out on a SUN Microsystems Computer purchased bya Wellcome Trust Major Equipment Grant.
REFERENCES
ARLOCK, P. & NOBLE, D. (1985). Two components of 'second inward current' in ferret papillarymuscle. Journal of Physiology 369, 88P.
ARLOCK, P. & WOHLFART, B. (1986). Effect of voltage-clamp duration on force production ofsuccessive twitches in isolated ferret and guinea-pig papillary muscle. Journal of Physiology 381,95P
BARCENAS-RUIZ, L., BEUCKELMANN, D. J. & WIER, W. G. (1987). Sodium-calcium exchange inheart: Membrane currents and changes in [Ca2+]1. Science 238, 1720-1722.
BARCENAS-RUIZ, L. & WIER, W. G. (1987). Voltage dependence of intracellular [Ca2+]1 transientsin guinea-pig ventricular myocytes. Circulation Research 61, 148-154.
BLATT, M. R. & SLAYMANN, C. L. (1983). KCl leakage from microelectrodes and its impact on themembrane parameters of a non-excitable cell. Journal of Membrane Biology 72, 223-234.
BROWN, H. F. & NOBLE, S. J. (1974). Effects of adrenaline on membrane currents underlyingpacemaker activity in frog atrial muscle. Journal of Physiology 238, 51-53P.
CAMPBELL, D. L., GILES, W. R., RoBINSON, K. & SHIBATA, E. F. (1988). Studies of the sodium-calcium exchanger in bull-frog atrial myocytes. Journal of Physiology 403, 317-340.
CANNELL, M. B., BERLIN, J. R. & LEDERER W. J. (1987). Effects of membrane potential changeson the calcium transient in single rat cardiac muscle cells. Science 238, 1419-1423.
CAVALIE', A., MCDONALD, T. F., PELZER, D. & TRAUTWEIN, W. (1985). Temperature-inducedtransitory and steady-state changes in the calcium current of guinea pig ventricular myocytes.Pflugers Archiv 405, 294-296.
DESILETS, M. & BAUMGARTEN, C. M. (1986). K+, Na+ and Cl activities in ventricular myocytesisolated from rabbit heart. American Journal of Physiology 251, C197-208.
DIFRANCESCO, D. & NOBLE, D. (1985). A model of cardiac electrical activity incorporating ionicpumps and concentration changes. Philosophical Transactions of the Royal Society B 307, 353-398.
660

Na+-Ca2+ EXCHANGE DURING ACTION POTENTIAL
EGAN. T. M1. NOBLE., )., NOBLE. S. J.. POWELL, T. & TWIST, V. WV. (1986a). Separation of calciumand [Ca]i-activated current in guinea-pig and rabbit ventricular cells. Journal of Physiology 381,94P.
EGAN, T. M., NOBLE, D., NOBLE, S. J., POWELL, T. & TWIST, V. W. (1986b). The use of very rapiddrug application to single cardiac cells to separate components of ionic current. Journal ofPhysiology 382, 19P.
EGAN, T. M., NOBLE, D., NOBLE, S. J., POWELL, T., TWIST, V. W. & YAMAOKA, K. (1988). On themechanism of isoprenaline- and forskolin-induced depolarization of single guinea-pig ventricularmyocytes. Journal of Physiology 400, 299-320.
EHARA, T., NOMA, A. & ONO, K. (1988). Calcium-activated non-selective cation channel invenitricular cells isolated from adult guinea-pig hearts. Journal of Physiology 403, 117-133.
FEDIDA, D., NOBLE, D., RANKIN, A. C. & SPINDLER, A. J. (1987a). The arrhythmogenic transientinward current iTI and related contraction in isolated guinea-pig ventricular myocytes. Journalof Physiology 392, 523-542.
FEDIDA, D., NOBLE, D., SHIMONI, Y. & SPINDLER, A. J. (1987b). Inward currents and contractionin guinea-pig ventricular myocytes. Journal of Physiology 385, 565-589.
HAMIL, 0. P., MARTY, A., NEHER, E., SAKMANN, B. & SIGWORTH, F. J. (1981). Improved patch-clamp techniques for high-resolution current recording from cells and cell-free patches. PfiugersArchiv 391, 85-100.
HILGEMANN, D. W. (1986a). Extracellular calcium transients and action potential configurationchanges related to post-stimulatory potentiation in rabbit atrium. Journal of General Physiology87, 675-706.
HILGEMANN, D. W. (1986b). Extracellular calcium transients at single excitations in rabbit atrium.Journal of General Physiology 87, 707-735.
HILGEMANN, D. XV. & NOBLE, 1). (1987). Excitation-contraction coupling and intracellular calciumtransients in rabbit atrium: reconstruction of basic cellular mechanisms. Proceedings of the RoyalSociety B 230. 163-205.
HUME, J. R. & UEHARA, A. (1986). Creep currents in single frog atrial cells may be generated byelectrogenic Na/Ca exchange. Journal of General Physiology 87, 857-884.
K[umtRAA J., MIYAMAE, S. & NOMA. A. (1987). Idenitificationi of sodium-calcium exchanige currenitin single ventricular cells of guinea-pig. Journal of Physiology 384, 199-222.
LIPP, P. & POTT, L. (1988). Transient inward current in guinea-pig atrial myocytes reflects achange of sodium-calcium exchange current. Journal of Physiology 397, 601-630.
MECHMANN, S. & POTT, L. (1986). Identification of Na-Ca exchange current in single cardiacmyocytes. Nature 319, 597-599.
MITCHELL, M. R., POWELL, T., TERRAR, D. A. & TWIST, V. W. (1984a). Possible associationbetween an inward eurrent and the late plateau of action potentials in ventricular cells isolatedfrom rat heart. Journal of Physiology 351, 40P.
MITCHELL, M. R., POWELL, T., TERRAR, D. A. & TWIST, V. W. (1984b). The effects of ryanodine,EGTA and low-sodium on action potentials in rat and guinea-pig ventricular myocytes: evidencefor two inward currents during the plateau. British Journal of Pharmacology 81, 543-550.
MITCHELL, M. R., POWELL, T., TERRAR, D. A. & TWIST, V. W. (1984c). Strontium, nifedipine and4-aminopyridine modify the time-course of the action potential in cells from rat ventricularmuscle. British Journal of Pharmacology 81, 551-556.
MI1TC11ELL, I. R., POWELL, T., TERRAR, D. A. & TWIST, V. XV. (1987). Calcium-activated currenitanid contractioni in rat and guiniea-pig ventricular myocytes. Journal of Physiology 391, 545-560.
MORAD, M. & TRAUTWEIN, W. (1968). The effect of duration of the action potential on contractionin mammalian heart muscle. Pftugers Archiv 299, 66-82.
NOBLE, D. (1986). Sodium-calcium exchange and its role in generating electric current. InExcitation and Regulation of Contraction, ed. NATHAN R., pp. 171-199. New York: AcademicPress.
POWELL, T., TERRAR, D. A. & TWIST, V. W. (1980). Electrical properties of individual cells isolatedfrom rat ventricular mvocardium. Journal of Physiology 302, 131-153.
WALSH, K. B., BEGENISICH, T. B. & KASS, R. S. (1988). /-Adrenergic modulation in the heart.Plfugers Archiv 411, 232-234.
WIER, W. G., CANNELL, M. B., BERLIN, J. R., MARBAN, E. & LEDERER, W. J. (1987). Cellular andsubcellular heterogeneity of [Ca]i in single heart cells revealed by fura-2. Science 235, 325-328.
661





























