Embed Size (px)
Citation preview

Insect Biochem. Vol. 15, No. 1, pp. 123-128, 1985 0020-1790/85 $3.00+0.00 Printed in Great Britain. All rights reserved Copyright © 1985 Pergamon Press Ltd
ACTIVATION MECHANISM OF INSECT FAT BODY PHOSPHORYLASE BY COLD
P H O S P H O R Y L A S E K I N A S E , P H O S P H A T A S E A N D A T P LEVEL
YOICHI HAYAKAWA Biochemical Laboratory, Institute of Low Temperature Science, Hokkaido University, Sapporo, Japan
(Received 6 March 1983)
Abstract--Fat body glycogen phosphorylase of overwintering insects is considerably activated by cold; two enzymes, phosphorylase kinase and phosphatase, may be involved in this activation. In this paper, we demonstrate the mechanism by which phosphorylase is activated by cold. Initially, a latent phosphorylase phosphatase was stimulated by freeze-thawing with 67~ saturated ammonium sulphate and mercaptoethanol, enabling partial purification and characterization of this enzyme. Unlike the phosphorylase kinase studied previously, the phosphatase did not function at 0°C. There was no appreciable activation of phosphorylase at 25°C if the phosphatase was present, whereas the phos- phorylase was greatly activated at 0°C even in the presence of phosphatase. It is suggested that ATP regulates phosphorylase activity through its effect on kinase and phosphatase. The activation of fat body phosphorylase that naturally occurs in overwintering insects is also discussed.
Key Word Index: Diapausing insect, Philosamia cynthia, fat body, phosphorylase kinase, phosphorylase phosphatase, ATP, cold-activation
INTRODUCTION
The accumulation of sugar alcohols (glycerol and/or sorbitol) (Chino, 1957; Chino, 1958; Wyatt and Meyer, 1959) or trehalose (Asahina and Tanno, 1964; Hayakawa and Chino, 1981) occurs in many species of over-wintering insects. In certain insects, e.g. di- apausing pupae of Hyalophora cecropia and Philos- amia cynthia silkworms, the production of glycerol or trehalose from glycogen depends on exposure to cold (Hayakawa and Chino, 1981; Wyatt, 1967). The production of such compounds is the result of cold- activation of fat body glycogen phosphorylase (Zi- egler and Wyatt, 1975; Ziegler et al., 1979; Hayakawa and Chino, 1982). Recently, we proposed an hypoth- esis to explain the mechanism of phosphorylase acti- vation by cold (Hayakawa and Chino, 1983). The major phosphorylase kinase of fat body is bound to glycogen-rich particles and demonstrates low but persistent activity at 0°C, whereas phosphorylase phosphatase does not function at 0°C. Thus, temperature-dependent properties of the two en- zymes may result in slow but continuous formation of the active form of phosphorylase. This proposal was based on indirect experimental evidence (Hay- akawa and Chino, 1983) and, to provide definitive proof, it is necessary to study the cumulative actions of the two enzymes, kinase and phosphatase, under various conditions. In this paper, we report the partial purification and characterization of fat body phosphorylase phosphatase as well as the kinase which has been studied previously (Hayakawa and Chino, 1983). On the basis of studies on the two partially purified enzymes, we present further evi- dence for the mechanism by which fat body phos- phorylase is activated at low temperature.
MATERIALS AND METHODS
Animals Pupae of the silkworm, Philosamia cynthia, were collected
outdoors in late summer. The pupae entered diapause almost immediately after collection and were kept at 18-20°C until use.
Chemicals AMP, NADP +, dithiothreitol (DTT), phenylmethylsul-
phonyl fluoride, glucose-l,6-diphosphate, glycerol-3- phosphate, phosphoglucomutase, luciferin-luciferase, and bovine serum albumin (BSA) were purchased from Sigma (USA). ATP and mercaptoethanol were obtained from Wako Chemical Co. (Japan) and oyster glycogen from Nakarai Chemical Co. (Japan). DEAE-cellulose was pur- chased from Whatman (Sweden) and Sephadex G-50, Se- pharose CL-4B from Pharmaeia (Sweden). The glycogen was treated with charcoal before use, to remove traces of AMP (Stevenson and Wyatt, 1964). Rabbit skeletal muscle phosphorylase a and phosphorylase b were obtained from Boehringer-Mannheim (Germany) and the contaminating AMP was removed by dialysis against 1 mM Tris-HC1 buffer (pH 7.5), followed by treatment with charcoal as above. All other chemicals were of analytical grade.
Phosphorylase kinase
The partial purification of glycogen-bound phosphorylase kinase from fat body was performed essentially according to the method described previously (Hayakawa and Chino, 1983). Dissected fat body was homogenized in buffer (50ram Tris-HCl, 10raM glycerol-3-phosphate, 5raM DTT, 2raM phenylmethylsulphonyl fluoride, pH7.5, 2.5 ml/g tissue) and the homogenate centrifuged at 10,000 g for 10 rain at 4°C. The infranatant between the top fat layer and pellet was collected and centrifuged at 100,000g for 90 rain at 4°C. The resulting glycogen-rich pellet containing approx. 60% of the total phosphorylase kinase activity was resuspended in the buffer and subjected to precipitation by
123
124 YOICH! HAYAKAWA
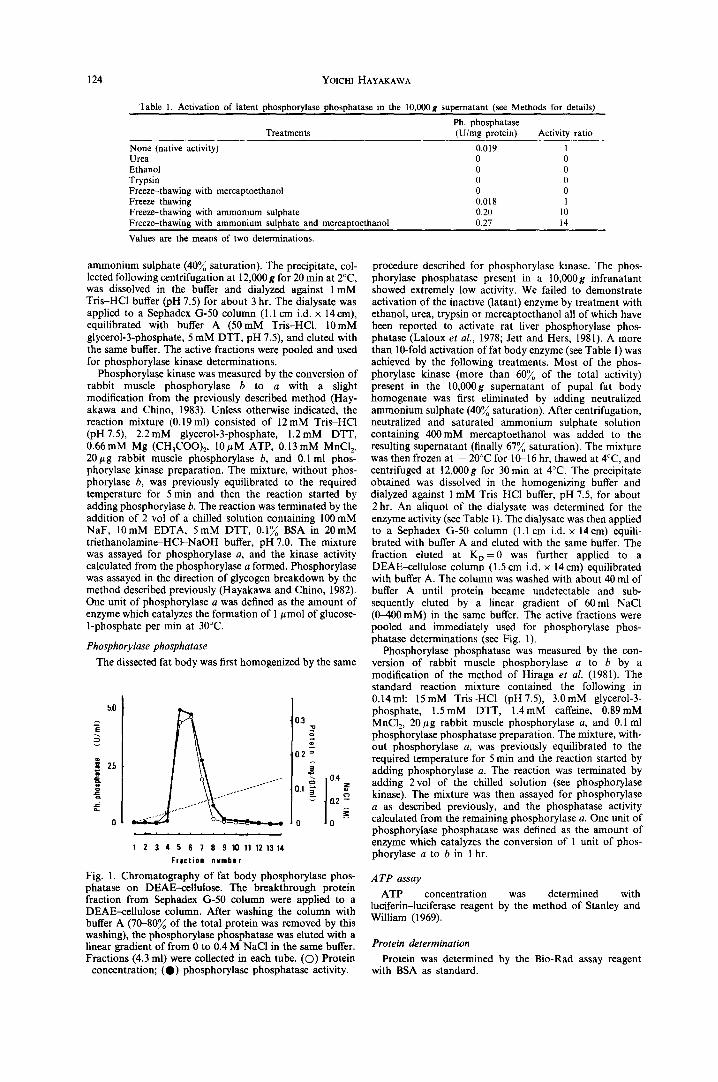
Table 1. Activation of latent phosphorylase phosphatase in the 10,000g supernatant (see Methods for details)
Ph. phosphatase Treatments (U/mg protein) Activity ratio
None (native activity) 0.019 1 Urea 0 0 Ethanol 0 0 Trypsin 0 0 Freeze-thawing with mercaptoethanol 0 0 Freeze-thawing 0.018 1 Freeze-thawing with ammonium sulphate 0.20 10 Freeze--thawing with ammonium sulphate and mercaptoethanol 0.27 14
Values are the means of two determinations.
ammonium sulphate (40~o saturation). The precipitate, col- lected following centrifugation at 12,000 g for 20 min at 2°C, was dissolved in the buffer and dialyzed against 1 mM Tris-HC1 buffer (pH 7.5) for about 3 hr. The dialysate was applied to a Sephadex G-50 column (1.1 cm i.d. x 14cm), equilibrated with buffer A (50mM Tris-HC1, 10mM glycerol-3-phosphate, 5 mM DTT, pH 7.5), and eluted with the same buffer. The active fractions were pooled and used for phosphorylase kinase determinations.
Phosphorylase kinase was measured by the conversion of rabbit muscle phosphorylase b to a with a slight modification from the previously described method (Hay- akawa and Chino, 1983). Unless otherwise indicated, the reaction mixture (0.19ml) consisted of 12mM Tris-HCl (pH7.5), 2.2mM glycerol-3-phosphate, 1.2raM DTT, 0.66mM Mg (CH3COO)2 , 10#M ATP, 0.13mM MnCI2, 20#g rabbit muscle phosphorylase b, and 0.1 ml phos- phorylase kinase preparation. The mixture, without phos- phorylase b, was previously equilibrated to the required temperature for 5 min and then the reaction started by adding phosphorylase b. The reaction was terminated by the addition of 2 vol of a chilled solution containing 100 mM NaF, 10mM EDTA, 5mM DTT, 0.1~ BSA in 20mM triethanolamine-HCl-NaOH buffer, pH 7.0. The mixture was assayed for phosphorylase a, and the kinase activity calculated from the phosphorylase a formed. Phosphorylase was assayed in the direction of glycogen breakdown by the method described previously (Hayakawa and Chino, 1982). One unit of phosphorylase a was defined as the amount of enzyme which catalyzes the formation of 1 #mol of glucose- 1-phosphate per min at 30°C.
Phosphorylase phosphatase
The dissected fat body was first homogenized by the same
~o
E
,~ 2.s
o
0.3
m
0.2 =
1.1
o
0.4
0.2
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Fraction number
Fig. 1. Chromatography of fat body phosphorylase phos- phatase on DEAE-cetlulose. The breakthrough protein fraction from Sephadex G-50 column were applied to a DEAE--cellulose column. After washing the column with buffer A (70-80~ of the total protein was removed by this washing), the phosphorylase phosphatase was eluted with a linear gradient of from 0 to 0.4 M NaC1 in the same buffer. Fractions (4.3 ml) were collected in each tube. (©) Protein
concentration; (O) phosphorylase phosphatase activity.
procedure described for phosphorylase kinase. The phos- phorylase phosphatase present in a 10,000g infranatant showed extremely low activity. We failed to demonstrate activation of the inactive (latant) enzyme by treatment with ethanol, urea, trypsin or mercaptoethanol all of which have been reported to activate rat liver phosphorylase phos- phatase (Laloux et al., 1978; Jett and Hers, 1981). A more than 10-fold activation of fat body enzyme (see Table 1) was achieved by the following treatments. Most of the phos- phorylase kinase (more than 60~o of the total activity) present in the 10,000g supernatant of pupal fat body homogenate was first eliminated by adding neutralized ammonium sulphate (40~o saturation). After centrifugation, neutralized and saturated ammonium sulphate solution containing 400mM mercaptoethanol was added to the resulting supernatant (finally 67~/o saturation). The mixture was then frozen at - 20°C for 10016 hr, thawed at 4°C, and centrifuged at 12,000g for 30 min at 4°C. The precipitate obtained was dissolved in the homogenizing buffer and dialyzed against 1 mM Tris-HC1 buffer, pH 7.5, for about 2 hr. An aliquot of the dialysate was determined for the enzyme activity (see Table 1). The dialysate was then applied to a Sephadex G-50 column (1.1 cm i.d. x 14cm) equili- brated with buffer A and eluted with the same buffer. The fraction eluted at K o = 0 was further applied to a DEAE--cellulose column (1.5 crn i.d. × 14 cm) equilibrated with buffer A. The column was washed with about 40 ml of buffer A until protein became undetectable and sub- sequently eluted by a linear gradient of 60ml NaC1 (0-400 mM) in the same buffer. The active fractions were pooled and immediately used for phosphorylase phos- phatase determinations (see Fig. 1).
Phosphorylase phosphatase was measured by the con- version of rabbit muscle phosphorylase a to b by a modification of the method of Hiraga et al. (1981). The standard reaction mixture contained the following in 0.14ml: 15mM Tris-HC1 (pH7.5), 3.0mM glycerol-3- phosphate, 1.5 mM DTT, 1.4 mM caffeine, 0.89 mM MnCI v 20#g rabbit muscle phosphorylase a, and 0.1 ml phosphorylase phosphatase preparation. The mixture, with- out phosphorylase a, was previously equilibrated to the required temperature for 5 min and the reaction started by adding phosphorylase a. The reaction was terminated by adding 2vol of the chilled solution (see phosphorylase kinase). The mixture was then assayed for phosphorylase a as described previously, and the phosphatase activity calculated from the remaining phosphorylase a. One unit of phosphorylase phosphatase was defined as the amount of enzyme which catalyzes the conversion of 1 unit of phos- phorylase a to b in 1 hr.
A T P assay
ATP concentration was determined with luciferin-luciferase reagent by the method of Stanley and William (1969).
Protein determination
Protein was determined by the Bio-Rad assay reagent with BSA as standard.
Phosphorylase activation by cold
RESULTS 1,o
Activation and partial purification of phosphorylase phosphatase
As indicated in Table 1, phosphorylase phos- phatase present in a 10,000 g infranatant of pupal fat body homogenate exhibits only negligible activity 0 and the various treatments that are reported to stimulate rat liver phosphorylase phosphatase have > no effect for the fat body enzyme. However, the -= enzyme was greatly stimulated by freeze-thawing in the presence of ammonium sulphate (67% saturation) -10 and mercaptoethanol. The activated enzyme solution obtained after dialysis was submitted to further purification (see Method).
An elution profile on DEAE-cellulose column that is the final step of partial purification is illustrated in -2.0 Fig. 1 and demonstrates that the enzyme is eluted by a linear gradient of NaC1 (0-0.4 M) as a single peak. Ten-fold purification (against the original activated enzyme) was achieved at this step.
Properties of phosphorylase phosphatase Time course and temperature-dependence of phos-
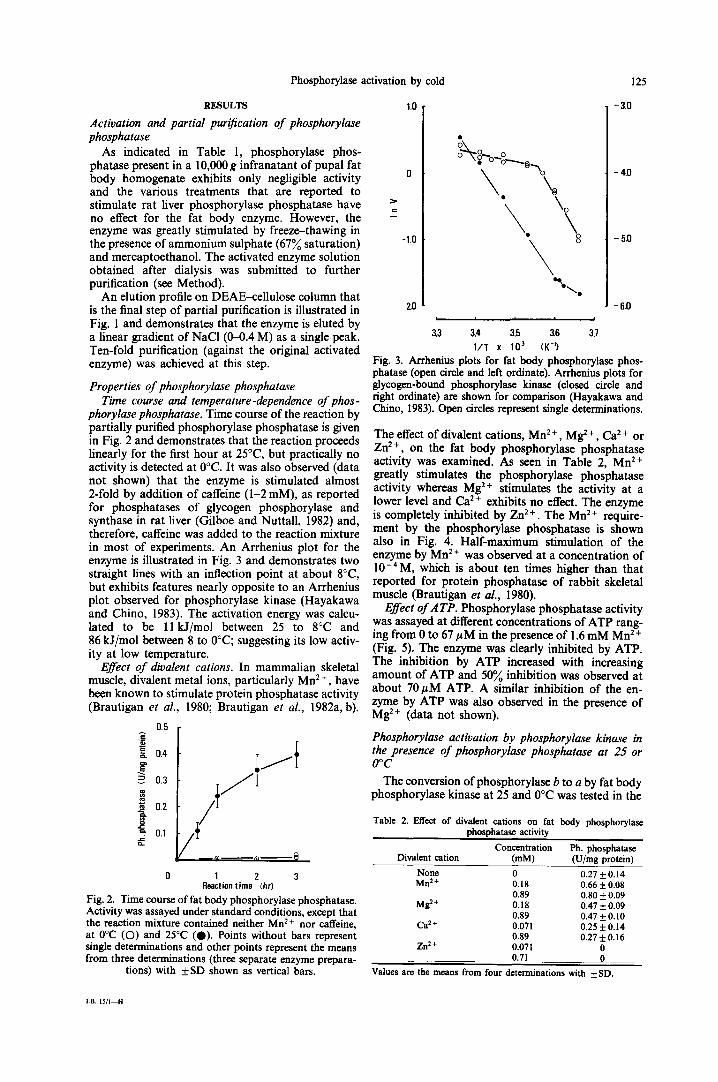
phorylase phosphatase. Time course of the reaction by partially purified phosphorylase phosphatase is given in Fig. 2 and demonstrates that the reaction proceeds linearly for the first hour at 25°C, but practically no activity is detected at 0°C. It was also observed (data not shown) that the enzyme is stimulated almost 2-fold by addition of caffeine (1-2 mM), as reported for phosphatases of glycogen phosphorylase and synthase in rat liver (Gilboe and Nuttall, 1982) and, therefore, caffeine was added to the reaction mixture in most of experiments. An Arrhenius plot for the enzyme is illustrated in Fig. 3 and demonstrates two straight lines with an inflection point at about 8°C, but exhibits features nearly opposite to an Arrhenius plot observed for phosphorylase kinase (Hayakawa and Chino, 1983). The activation energy was calcu- lated to be 11 kJ/mol between 25 to 8°C and 86 kJ/mol between 8 to 0°C; suggesting its low activ- ity at low temperature.
Effect of divalent cations. In mammalian skeletal muscle, divalent metal ions, particularly Mn 2 ÷, have been known to stimulate protein phosphatase activity
1980; Brautigan et al., 1982a, b). (Brautigan et al.,
0.5
0.4
~ 0.3 g ~ 0.2
~'. 0.1
0 I 2 3 Reaction time (hr)
Fig. 2. Time course of fat body phosphorylase phosphatase. Activity was assayed under standard conditions, except that the reaction mixture contained neither Mn 2+ nor caffeine, at 0°C (O) and 25°C (0). Points without bars represent single determinations and other points represent the means from three determinations (three separate enzyme prepara-
tions) with _+SD shown as vertical bars.
125
-3 .0
o o~.. o • v - " ~ , ~
\ . \ \o \
\
- 4 . 0
- 5 . 0
- 6 . 0
I
3.7 i I i I
3.3 3.4 3.5 3.6 I/T x 103 (K -1)
Fig. 3. Arrhenius plots for fat body phosphorylase phos- phatase (open circle and left ordinate). Arrhenius plots for glycogen-bound phosphorylase kinase (closed circle and fight ordinate) are shown for comparison (Hayakawa and Chino, 1983). Open circles represent single determinations.
The effect of divalent cations, Mn 2 +, Mg: +, Ca 2 + or Zn 2+, on the fat body phosphorylase phosphatase activity was examined. As seen in Table 2, Mn 2+ greatly stimulates the phosphorylase phosphatase activity whereas Mg 2+ stimulates the activity at a lower level and Ca 2 + exhibits no effect. The enzyme is completely inhibited by Zn 2 +. The Mn 2 + require- ment by the phosphorylase phosphatase is shown also in Fig. 4. Half-maximum stimulation of the enzyme by Mn 2 + was observed at a concentration of 10-4M, which is about ten times higher than that reported for protein phosphatase of rabbit skeletal muscle (Brautigan et al., 1980).
Effect ofATP. Phosphorylase phosphatase activity was assayed at different concentrations of ATP rang- ing from 0 to 67 #M in the presence of 1.6 mM Mn 2+ (Fig. 5). The enzyme was clearly inhibited by ATP. The inhibition by ATP increased with increasing amount of ATP and 50% inhibition was observed at about 70 # M ATP. A similar inhibition of the en- zyme by ATP was also observed in the presence of Mg 2+ (data not shown).
Phosphorylase activation by phosphorylase kinase in the presence of phosphorylase phosphatase at 25 or O°C
The conversion ofphosphorylase b to a by fat body phosphorylase kinase at 25 and 0°C was tested in the
Table 2. Effect of divalent cations on fat body phosphorylase phosphatase activity
Concentration Ph. pbosphatase Divalent cation (mM) (U/rag protein)
None 0 0.27 + 0.14 Mn 2+ 0.18 0.66 __ 0.08
0.89 0.80 -6_ 0.09 Mg 2+ 0.18 0.47 _-6 0.09
0.89 0.47 _ 0.10 Ca 2+ 0.071 0.25 _-6 0.14
0.89 0.27 ± 0.16 Zn :+ 0.071 0
0.71 0 Values are t h e m e a n s from four determinations with ± SD.
L B . 1 5 / I - - H
126 Yoicm HAYAKAWA
~ 1.0
o .
o.5
o
o//O.
o o
o
~ , , , ~ , 1 i i i i i
6 5 4 3 2 p Mn 2+
Fig. 4. Effect of Mn 2+ on fat body phosphorylase phos- phatase activity. The reaction was carried out for I hr at
25°C. Each point represents single determinations.
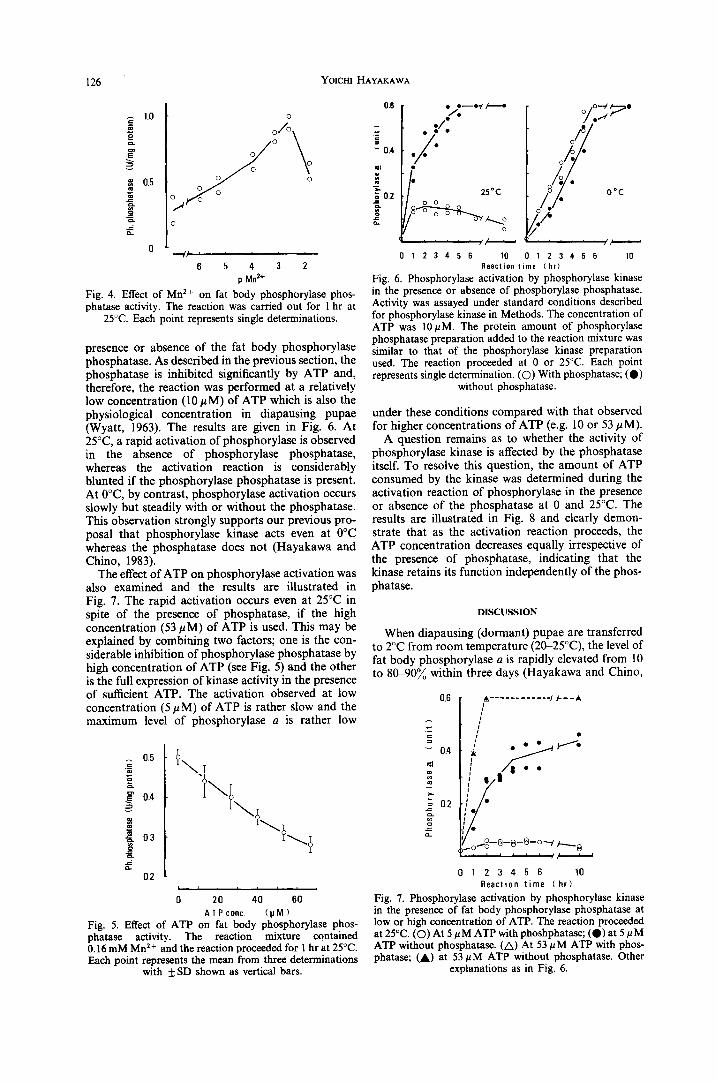
presence or absence of the fat body phosphorylase phosphatase. As described in the previous section, the phosphatase is inhibited significantly by ATP and, therefore, the reaction was performed at a relatively low concentration (10/~M) of ATP which is also the physiological concentration in diapausing pupae (Wyatt, 1963). The results are given in Fig. 6. At 25°C, a rapid activation of phosphorylase is observed in the absence of phosphorylase phosphatase, whereas the activation reaction is considerably blunted if the phosphorylase phosphatase is present. At 0°C, by contrast, phosphorylase activation occurs slowly but steadily with or without the phosphatase. This observation strongly supports our previous pro- posal that phosphorylase kinase acts even at 0°C whereas the phosphatase does not (Hayakawa and Chino, 1983).
The effect of ATP on phosphorylase activation was also examined and the results are illustrated in Fig. 7. The rapid activation occurs even at 25°C in spite of the presence of phosphatase, if the high concentration (53/~M) of ATP is used. This may be explained by combining two factors; one is the con- siderable inhibition of phosphorylase phosphatase by high concentration of ATP (see Fig. 5) and the other is the full expression of kinase activity in the presence of sufficient ATP. The activation observed at low concentration (5 #M) of ATP is rather slow and the maximum level of phosphorylase a is rather low
I/
g2
0.5
0.4
0.3
0.2
0 20 40 60 ATP c0nc. (pM)
Fig. 5. Effect of ATP on fat body phosphorylase phos- phatase activity. The reaction mixture contained 0.16 mM Mn 2+ and the reaction proceeded for 1 hr at 25°C. Each point represents the mean from three determinations
with + SD shown as vertical bars.
0.6
== 0,4
.!'
~, o2 .,1=
e ~ e ~eee/ lee--e ' /
: 25"C o o o
o
i , i i i i ~ ¢ i J
0 1 2 3 4 5 6 10
o ,o--/,~:~O )/o.~
o Ooc
i i i , i i , r i ,
0 1 2 3 4 5 6 10 Reaction time (hr)
Fig. 6. Phosphorylase activation by phosphorylase kinase in the presence or absence of phosphorylase phosphatase. Activity was assayed under standard conditions described for phosphorylase kinase in Methods. The concentration of ATP was 10/~M. The protein amount of phosphorylase phosphatase preparation added to the reaction mixture was similar to that of the phosphorylase kinase preparation used. The reaction proceeded at 0 or 25°C. Each point represents single determination. (O) With phosphatase; (O)
without phosphatase.
under these conditions compared with that observed for higher concentrations of ATP (e.g. 10 or 53/~M).
A question remains as to whether the activity of phosphorylase kinase is affected by the phosphatase itself. To resolve this question, the amount of ATP consumed by the kinase was determined during the activation reaction of phosphorylase in the presence or absence of the phosphatase at 0 and 25°C. The results are illustrated in Fig. 8 and clearly demon- strate that as the activation reaction proceeds, the ATP concentration decreases equally irrespective of the presence of phosphatase, indicating that the kinase retains its function independently of the phos- phatase.
DISCUSSION
When diapausing (dormant) pupae are transferred to 2°C from room temperature (20-25°C), the level of fat body phosphorylase a is rapidly elevated from 10 to 80-90% within three days (Hayakawa and Chino,
0.6
~ 04
m
= 0.2
_¢= ct_
i
i /
o ~ - 8 - 6 - 9 - ° ~ ~ e
0 1 2 3 4 5 6 10 Reaction t ime (h r )
Fig. 7. Phosphorylase activation by phosphorylase kinase in the presence of fat body phosphorylase phosphatase at low or high concentration of ATP. The reaction proceeded at 25°C. (O) At 5 pM ATP with phoshphatase; (O) at 5 pM ATP without phosphatase. (A) At 53 pM ATP with phos- phatase; (A) at 53 #M ATP without phosphatase. Other
explanations as in Fig. 6.

Phosphorylase activation by cold 127
10 o
o i i t t i i
0 1 2 3 1. 5 Reaction time {hr)
Fig. 8. The change of ATP concentration during the phos- phorylase activation. Circle, 25°C; triangle, 0°C. (©, A) With phosphatase; (O, A) without phosphatase. Other
explanations as in Fig. 6.
1982). Two enzymes, phosphorylase kinase and phos- phorylase phosphatase, are likely to be involved in this mechanism (Hayakawa and Chino, 1982; Sack- tot, 1970; Sacktor et al., 1974; McClure and Steele, 1981; Steele, 1982). The present study has been designed to elucidate the mechanism by which these enzymes effect cold-activation of the phosphorylase.
To provide critical evidence for this mechanism, it is necessary to carry out experiments in vitro using isolated and combined preparations of the two en- zymes. Several reports on phosphorylase kinase in insect fat body have been published (Steele, 1982) and we have previously achieved partial purification of the enzyme from pupal fat body (Hayakawa and Chino, 1983). However, little is known about phos- phorylase phosphatase in insect fat body and, indeed, the activity of this enzyme in fat body homogenate is extremely low and almost negligible (Ashida and Wyatt, 1979). It seems desirable to use native enzyme to obtain valid conclusions for the activation mech- anism of phosphorylase by cold in natural conditions described above. However, the activity of native phosphorylase phosphatase was too low and this prevented further experiments. Therefore, we devel- oped a new procedure by which the latent enzyme was considerably activated; freeze-thawing in the presence of 67% saturated ammonium sulphate and mercaptoethanol activated the enzyme more than ten times (Table 1), and facilitated isolation and charac- terization of the enzyme. The nature of the fat body phosphorylase phosphatase essentially resembles that reported for mammalian muscle phosphorylase phos- phatase (Brautigan et al., 1980; Brautigan et al., 1982a, b); the enzyme is considerably stimulated by Mn 2÷ or Mg 2÷ but is not affected by Ca 2÷ (Table 2). Fat body phosphorylase phosphatase appears to be much more sensitive to Zn ~ ÷ than mammalian phos- phatases (Brautigan et al., 1980; Brautigan et al., 1982a, b; Khanddelwal, 1977; Kobayshi, 1975); the former enzyme is almost completely inhibited by Zn 2 + even at concentrations lower than 100 #M. The sensitivity of the insect fat body phosphorylase phos- phatase to Zn 2 + is almost equivalent to that reported for phosphotyrosine phosphatase of some mam- malian tumor cells (Brautigan et al., 1981; Hrrlein et al., 1982).
In the present study, we found that the insect fat
body phosphorylase phosphatase is inhibited by ATP at concentrations lower than 100/zM, even in the presence of Mg 2+. The sensitivity to inhibition by ATP resembles that reported for mammalian protein phosphatase-2A (Ingebritsen and Cohen, 1983; Inge- britsen et al., 1983).
A most significant observation in this study is the difference in temperature-dependence of the phos- phorylase phosphatase compared to that of the phos- phorylase kinase reported previously (Hayakawa and Chino, 1983) (Figs 2 and 3); thus supporting our proposal that the kinase functions even at 0°C whereas the phosphatase has no appreciable activity at such a temperature. These observations together with experimental results obtained by combining the two enzymes, kinase and phosphatase, under different ATP concentrations (Figs 6 and 7) suggest the following as the most likely mechanism of phos- phorylase activation by cold. A low level of phos- phorylase a is established in the fat body of di- apausing pupae that have been kept at 20-25°C because the activity of phosphorylase kinase is lim- ited owing to the low ATP level (about 10/zM). The low level of ATP maintains high phosphorylase phos- phatase activity. If such pupae with low phos- phorylase a are exposed to low temperature (0°C), phosphorylase phosphatase fails to function whereas phosphorylase kinase still retains its function, and this causes a slow but continuous formation of the active form of the phosphorylase.
Acknowledgements--The author wishes to thank Professor H. Chino of this laboratory for his continuous encour- agement and valuable criticism. Thanks are also due to Dr R. G. H. Downer of University of Waterloo, Canada, for reading the manuscript. This study was partly supported by a research grant from Mitsubishi Foundation.
R E F E R E N C E S
Asahina E. and Tanno K. (1964) A large amount of trehalc, oe in a frost resistant insect. Nature 204, 1222.
Ashida M. and Wyatt G. R. (1979) Properties and activa- tion of phosphorylase kinase from silkmoth fat body. Insect Biochem. 9, 403-409.
Brautigan D. L., Bornstein P. and Gallis B. (1981) Phosphotyrosyl-protein phosphatase. Specific inhibition b y Z n 2+ . J. biol. Chem. 256, 6519-6522.
Brautigan D. L., Ballou L. M. and Fischer E. H. (1982a) Activation of skeletal muscle phosphorylase phosphatase. Effects of proteolysis and divalent cations. Biochemistry 21, 1977-1982.
Brautigan D. L., Khatra B. S., Soderling T. R. and Fischer E. H. (1982b) Dephosphorylation of skeletal muscle phosphorylase, glycogen synthase, and phosphorylase kinase fl-subunit by a Mn2+-activated protein phos- phatase. Archs Biochem. Biophys. 219, 228-235.
Brautigan D. L., Picton C. and Fischer E. H. (1980) Phosphor~lase phosphatase complex from skeletal mus- cle. Activation of one of two catalytic subunits by man- ganese ions. Biochemistry 19, 5787-5794.
Chino H. (1957) Conversion of glycogen to sorbitol and glycerol in the diapausing egg of the Bombyx silkworm. Nature 180, 606-607.
Chino H. (1958) Carbohydrate metabolism in diapause eggs of the silkworm, Bombyx mori. II. Conversion of glycogen into sorbitol and glycerol. J. Insect Physiol. 2, 1-12.
Gilboe D. P. and Nuttall F. Q. (1982) Stimulation of liver glycogen particle synthase D phosphatase activity by

128 Yotcm HAYAKAWA
caffeine, AMP, and glucose 6-phosphate. Archs Biochem. Biophys. 219, 179-185.
Hayakawa Y. and Chino H. (1981) Temperature-dependent interconversion between glycogen and trehalose in di- apausing pupae of Philosamia cynthia ricini and pryeri. Insect Biochem. 11, 43-47.
Hayakawa Y. and Chino H. (1982) Temperature-dependent activation or inactivation of glycogen phosphorylase and synthase of fat body of the silkworm Philosamia cynthia: the possible mechanism of the temperature-dependent interconversion between glycogen and trehalose. Insect Biochem. 12, 361-366.
Hayakawa Y. and Chino H. (1983) Insect fat body phos- phorylase kinase is Ca2+-independent and acts even at 0°C. Biochim. biophys. Acta 746, 14-17.
Hiraga A., Kikuchi K., Tamura S. and Tsuiki S. (1981) Purification and characterization of Mg ~ +-dependent gly- cogen synthase phosphatase (phosphoprotein phos- phatase 1A) from rat liver. Eur. J. Biochem. 119, 503-510.
H6rlein D., Byron G., Brautigan D. L. and Bornstein P. (1982) Partial purification and characterization of phosphotyrosyl-protein phosphatase from Ehrlich ascites tumor cells. Biochemistry 21, 5577-5584.
Ingebritsen T. S. and Cohen P. (1983) The protein phos- phatases involved in cellular regulation. 1. Classification and substrate specificities. Eur. J. Biochem. 132, 255-261.
Ingebritsen T. S., Foulkes J. G. and Cohen P. (1983) The protein phosphatases involved in cellular regulation. 2. Glycogen metabolism. Eur. J. Biochem. 132, 263-274.
Jett M. F. and Hers H. G. (1981) Latent phosphorylase phosphatases from rat liver: relationship with the heat- stable inhibitory protein. Fur. J. Biochem. 118, 283-288.
Khandelwal R. L. (1977) Some properties of purified phos- phoprotein phosphatases from rabbit liver. Biochim. bio- phys. Acta 455, 379-390.
Kobayashi M., Kato K. and Sato S. (1975) Multiple molecular forms of phosphoprotein phosphatase. III. Phosphorylase phosphatase and phosphohistone phos- phatase of rabbit liver. Biochim. biophys. Acta 377, 343-355.
Laloux M., Stalmans W. and Hers H. G. (1978) Native and latent forms of liver phosphorylase phosphatase: the non-identity of native phosphorylase phosphatase and synthase phosphatase. Fur. J. Biochem. 92, 15-24.
McClure J. B. and Steele J. E. (1981) The role of extra- cellular calcium in hormonal activation of glycogen phos- phorylase in cockroach fat body. Insect Biochem. 11, 605-613.
Sacktor B. (1970) Regulation of intermediary metabolism, with special reference to the control mechanisms in insect flight muscle. Adv. Insect Physiol. 7, 267-347.
Sacktor B., Wu N.-C., Lescure O. and Reed W. D. (1974) Regulation of muscle phosphorylase b kinase activity by inorganic phosphate and calcium ions. Biochem. J. 137, 535-542.
Stanley P. E. and William S. G. (1969) Use of the liquid scintillation spectrometer for determihing adenosine triphosphate by the luciferase enzyme. Analyt. Biochem. 29, 381-392.
Steele J. E. (1982) Glycogen phosphorylase in insects. Insect Biochem. 12, 131-147.
Stevenson E. and Wyatt G. R. (1964) Glycogen phos- phorylase and its activation in silkmoth fat body. Archs Biochem. Biophys. 108, 420-429.
Wyatt G. R. (1963) Biochemistry of diapause, development, and injury in silkworm pupae. In Insect Physiology (Edi- ted by Brookes V. J.), pp. 23-41. Oregon State Univ, Press, Corvallis, Oregon.
Wyatt G. R. (1967) The biochemistry of sugar and poly- saccharides in insects. Adv. Insect Physiol. 4, 287-360.
Wyatt G. R. and Meyer W. L. (1959) The chemistry of insect hemolymph. III. Glycerol. J. gen. Physiol. 42, 1005-1011.
Ziegler R., Ashida M., Fallon A. M., Wimer L. T., Wyatt S. S. and Wyatt G. R. (1979) Regulation of glycogen phosphorylase in fat body of Cecropia silkmoth pupae. J. comp. Physiol. 131, 321-332.
Ziegler R. and Wyatt G. R. (1975) Phosphorylase and glycerol production activated by cold in diapausing silk- moth pupae. Nature 254, 622-623.





























