Embed Size (px)
Citation preview

COGNITIVE NEUROSCIENCE NEUROREPORT
0959-4965 & Lippincott Williams & Wilkins Vol 12 No 18 21 December 2001 4093
Activation of brain mechanisms of attentionswitching as a function of auditory frequency
change
Elena Yago, MarõÂa Jose Corral and Carles EsceraCA
Neurodynamics Laboratory, Department of Psychiatry and Clinical Psychobiology, University of Barcelona, P. Vall d'Hebron 171,08035-Barcelona, Catalonia, Spain
CACorresponding Author
Received 3 October 2001; accepted 30 October 2001
The activation of the cerebral network underlying involuntaryattention switching was studied as a function of the magnitudeof auditory change. Event-related brain potentials (ERPs) wererecorded during the performance of a visual discrimination taskin which task-irrelevant auditory frequency changes of sixdifferent levels (5%, 10%, 15%, 20%, 40% and 80%) occurredrandomly within the same stimulus sequence. All the frequencychanges elicited a typical ERP waveform, characterized by
MMN, P3a and RON, their respective amplitudes increasinglinearly as a function of the magnitude of change. The resultsindicate that attentional processes in the brain may follow alinear function of activation, contrasting with the well-estab-lished logarithmic functions underlying perceptual and psycho-physical processes. NeuroReport 12:4093±4097 & 2001Lippincott Williams & Wilkins.
Key words: Attention shifting; Distraction; Event-related brain potentials; Mismatch negativity; Orienting response; P3a; RON
INTRODUCTIONEvent-related brain potentials (ERPs) can be used to trackfrom the human scalp the activation of the cerebral net-work engaged by the unexpected occurrence of auditorystimulus change. At early stages of processing (�150 ms)after the occurrence of such changes, the mismatch nega-tivity (MMN) [1] is generated in the auditory cortex totrigger a switch of attention towards that potentiallyrelevant stimulus. Successful attention switches lead to aneffective orienting response, which is apparently signaledover the scalp at �300 ms by the subsequent P3a waveform(for reviews see [2,3]).
The P3a has been proposed to re¯ect the activation of alarge-scale cerebral network involved in the orientingresponse towards novelty and change in the acousticenvironment [4±8]. P3a is larger in amplitude to moreattention-catching events, such as novel sounds, whichsupports its role in the orienting of attention. Cerebralregions involved in P3a generation include the supratem-poral auditory cortex, the anterior and posterior associationcortices, and the posterior hypocampus (for reviews see[2,3]). In agreement with the involvement of this large-scalecerebral network in P3a generation, recent studies havesuggested that, rather than a unitary process, the P3a iscomposed by two consecutive phases with different func-tional roles and different brain areas involved in theirgeneration [8,9].
More recently, a late ERP response involved in thesequence of brain events underlying auditory change and
novelty, processing has been described. This response,called the reorienting negativity (RON) [10], is generatedover frontal areas [11] when irrelevant attention-catchingevents occur during the performance of a given task. In theoriginal study [10], RON was elicited when subjects had toreorient attention to relevant aspects of the auditorystimulation (i.e. duration) after distraction caused by un-expected frequency changes in the same auditory modality.Subsequent studies have also shown that RON can beelicited during visual distraction of visual performance [12]and during auditory distraction of visual performance [9].
It has been proposed that RON re¯ects the reorienting ofattention towards original task performance after momen-tary distraction, as supported by its larger amplitude tomore task-disrupting events [9,11,12] and by the fact ofbeing time-locked to the events of the primary task [9].
There is evidence suggesting that the sequence of brainevents underlying involuntary attention may activate in-creasingly as a function of increasing the magnitude ofauditory change. Indeed, Tiitinen et al. [13] have shownthat the peak amplitude of MMN increases linearly as afunction of the logarithm of the frequency deviance, andAmenedo and Escera [14] have reported a similar logarith-mic relationship between MMN amplitude and devianceby using changes in stimulus duration (increments ordecrements). According to this parametric increase ofMMN amplitude to increasing the magnitude of deviance,one should expect a similar behavior of the subsequentprocessing stages in the cerebral network of involuntary

attention, as indexed by P3a and RON. None of the abovementioned studies, however, analyzed these ERP re-sponses, and therefore the present study aimed at investi-gating their activation as a function of increasing frequencychange and their relationship to behavioral distraction.
MATERIALS AND METHODSEleven healthy human subjects (18±35 years; four males;two left-handed), with normal hearing and vision, partici-pated in the study after informed written consent. Subjectswere presented with 12 blocks of 375 stimulus pairs eachdelivered at a rate of one pair every 1.2 s. Each pairconsisted of an auditory stimulus followed after 300 ms(onset-to-onset) by a visual stimulus. Auditory stimuliwere a 600 Hz standard tone (p� 0.82) and six differentdeviant tones (p� 0.03 each), which, differed in frequencyfrom the standard tone at 5% (630 Hz), 10% (660 Hz), 15%(690 Hz), 20% (720 Hz), 40% (840 Hz) and 80% (1080 Hz),occurring randomly within the stimulus blocks. All audi-tory stimuli were delivered binaurally through headphoneswith a duration of 150 ms (including 10 ms rise and falltimes) and an intensity of 75 dB SPL. Pairs with a devianttone were preceded by a pair in which the tone wasstandard. Visual stimuli were either an even (2, 4, 6, 8) oran odd (3, 5, 7, 9) digit, presented equiprobably on acomputer screen for 200 ms with a vertical visual angle of1.78 and a horizontal angle of 1.18 (150 cm from thesubject's eyes).
Subjects were instructed to use their dominant hand topress one response button with the index ®nger to evennumbers, and another response button with the middle®nger to odd numbers, and to ignore the auditory stimula-tion. Both speed and accuracy were emphasized. Beforethe experimental session, subjects received one practiceblock in which the sounds were turned off, all subjectsreaching a hit rate level of > 85%. A correct button presswithin 1100 ms after visual-stimulus onset was regarded asa hit, the mean reaction time being computed only for thehit trials. Hit rate and reaction time in standard pairs wascompared by means of two-tailed t-tests with performancein each of the deviant pairs.
The EEG (bandpass 0±100 Hz) was continuously re-corded (sampling rate 500 Hz) from the 19 scalp electrodesof the 10-20 system (Fp1, Fp2, F7, F3, Fz, F4, F8, T3, C3, Cz,C4, T4, T5, P3, Pz, P4, T6, O1, O2), and from the left (LM)and right (RM) mastoids. Horizontal and vertical EOGwere recorded with electrodes attached to the canthus and
below the right eye, respectively. The common referenceelectrode was placed on the tip of the nose. ERPs wereaveraged off-line for each auditory stimulus type, for anepoch of 1200 ms including a preauditory stimulus periodof 100 ms. Epochs in which the EEG or EOG exceeded�100 ìV, as well as the ®ve ®rst epochs of each block, wereautomatically excluded from averaging. The standard-tonepairs immediately following a deviant-tone pair were alsoexcluded from the averages. Frequencies . 30 Hz weredigitally ®ltered out from individual ERPs.
The mean amplitude of the relevant ERP componentswas calculated in the difference waves obtained by sub-tracting the ERPs to the standard pairs from those to eachof the six different types of deviant pairs. The meanamplitude of MMN was calculated in the 100±200 ms timewindow after stimulus onset. P3a was identi®ed as thelargest positive peak following MMN, and its mean ampli-tude was calculated in the 225±350 ms time window.Following the P3a, a late negativity was identi®ed in thedifference waves as the RON response, its mean amplitudebeing computed in a 100 ms time window around itsmaximum peak (450±550 ms). The mean amplitude of eachERP component at Fz for each deviant condition wascompared against the zero level by means of t-test com-parisons in order to determine whether each componentwas statistically signi®cant. The scalp distribution of thedifferent ERP responses was analyzed by ANOVA forrepeated measures, with deviance (six levels), frontality(three levels from anterior to posterior) and laterality (®velevels from left to right) performed on normalized meanamplitudes at the electrodes F7, F3, Fz, F4, F8, T3, C3, Cz,C4, T4, T5, P3, Pz, P4 and T6. When appropriate, Green-house-Geisser correction of the degrees of freedom wasapplied, the uncorrected degrees of freedom, the correctedp values and epsilon values being reported. A prioricontrasts were run in the ANOVAs in order to comparethe scalp distributions of the ERPs between adjacentdeviant conditions. Regression analyses were applied onthe data in order to estimate whether the mean amplitudeof each of the studied ERP components could be predictedas a function of increasing the magnitude of deviance.
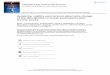
RESULTSStandardÿdeviant difference waves revealed an earlynegative response, peaking between 100 and 200 ms, whichwas identi®ed as the MMN (Fig. 1a). MMN was statisti-cally different from zero for all deviant types (Table 1), its
Table 1. Mean amplitude and values of the t-test comparisons of the mean amplitude at Fz ofMMN (100±200 ms), P3a (225±350 ms) and RON (450±550 ms) against the zero level.
Deviance MMN P3a RON
ìV t values ìV t values ìV t values
5% ÿ0.69 ÿ2.68�� �0.13 0.34 ÿ1.36 ÿ5.73���10% ÿ0.74 ÿ2.83�� �0.67 2.24�� ÿ0.98 ÿ2.06�15% ÿ0.93 ÿ2.91�� �0.58 1.47 ÿ1.23 ÿ3.47���20% ÿ1.21 ÿ5.55��� �0.83 2.54�� ÿ1.32 ÿ4.81���40% ÿ0.72 ÿ2.91�� �1.02 2.76�� ÿ1.62 ÿ4.27���80% ÿ1.37 ÿ3.61��� �1.69 2.66�� ÿ2242 ÿ5.8���
� p ,0.1; �� p ,0.05; ��� p ,0.01
NEUROREPORT E. YAGO, M. J. CORRAL AND C ESCERA
4094 Vol 12 No 18 21 December 2001

latency decreasing progressively with increasing the mag-nitude of the frequency change, as can be seen in Fig. 1a.ANOVA showed no signi®cant interaction of the factordeviance with the factors frontality and laterality, suggest-ing no statistical differences in the scalp distribution ofMMN elicited to the different deviant tones, which had afrontal-central maximum (Fig. 1b). The amplitude of MMN
increased linearly as a function of deviance (slope 0.07 ìVevery 10% change), although the regression model wasonly marginally signi®cant (F(1,4)� 3.25; p� 0.14) probablydue to a drop of MMN amplitude in the 40% deviantcondition (Fig. 2).
Following the MMN, a positive response was identi®edin the difference waves as the P3a (Fig. 1a). The meanamplitude of P3a in the 225±350 ms time window wassigni®cantly different from zero for the 10%, 20%, 40% and80% deviancies, and tended to be signi®cant in the 15%deviant condition (t(10)� 1.47; p� 0.17; Table 1). Differencewaves revealed a different shape of the P3a in the 80%deviant condition, with an evident later phase as comparedwith the other conditions [8] (Fig. 1a). ANOVA showed asigni®cant interaction between the factors deviance andfrontality (F(10,100)� 3.58; p , 0.02; å� 0.37), indicatingdifferent scalp distributions of P3a across deviancies. A
N1/MMN RON
22 µV
12 µV
Fz
(a)
P3a
800 ms
(b)
5%
10%
15%
20%
40%
80%
1/2 1.3 µV 1/2 2 µV 1/2 2.5 µV
1
2
0
Fig. 1. (a) Standardÿdeviant difference waveforms at Fz for the sixtypes of deviant conditions. (b) Scalp distribution maps of MMN, P3a andRON for the six types of deviant conditions.
Y 5 (20.75) 1 (20.007)*Xp 5 0.14
Y 5 0.32 1 0.017*Xp 5 0.04
Y 5 (21.04) 1 (20.015)*Xp 5 0.005
MMN
20 20.2 20.4 20.6 20.8
21 21.2 21.4 21.6 21.8
22
P3a
00.20.40.60.8
11.21.41.61.8
2
20.7 20.9 21.1 21.3 21.5 21.7 21.9 22.1 22.3 22.5
20.5
RON
Mea
n am
plitu
de (
µV)
510
1520
40 80
Deviance in Hz (%)
Fig. 2. Mean amplitudes (ìV) of the MMN (100±200 ms), P3a (225±325 ms) and RON (450±550 ms) responses at the Fz electrode as afunction of frequency change. Regression equations and their statisticalsigni®cance are shown below each regression line.
BRAIN IMAGING AS A FUNCTION OF AUDITORY CHANGE NEUROREPORT
Vol 12 No 18 21 December 2001 4095

priori contrasts revealed no signi®cant differences in scalpdistribution for adjacent deviant conditions, but a signi®-cant different scalp topography between the 40% and the80% deviant conditions (F(1,10)� 26.02; p , 0.001), whichwas due to the more posterior distribution of P3a in thelargest deviant condition, as can be seen in the distributionmaps (Fig. 1b). Regression analyses showed that theamplitude of P3a increased linearly as a function of thefrequency deviance (F(1,4)� 36.2; p , 0.05), with a slope of0.17 ìV every 10% change (Fig. 2).
Subsequent to P3a, a late negative response was alsoobserved in the difference waves (Fig. 1), peaking at about500 ms, which was identi®ed as the reorienting negativityor RON. The mean amplitude of RON was signi®cantlydifferent from the zero level for all types of deviant pairs(Table 1). RON had a frontal distribution which did notvary across deviancies (i.e., no signi®cant interactions be-tween the factors deviance, frontality and laterality werefound in ANOVA; Fig. 1b). The mean amplitude of RONincreased linearly as a function of deviance (F(1,4)� 30.87;p , 0.01), with a slope of 0.15 ìV every 10% change (Fig. 2).
t-test comparisons revealed signi®cant distraction effectsin the 10% deviant condition, as reaction time increasedsigni®cantly by 7.7 ms (t10�ÿ3.24; p , 0.01) when visualstimuli were preceded by a 10% deviant tone in compari-son to those visual stimuli preceded by a standard tone. Incontrast, reaction time decreased by 11.3 ms (t10� 5.21;p , 0.001) in the 5% deviant condition, suggesting a possi-ble facilitatory effect in this condition. No signi®cant be-havioral effects were found in any of the other deviantconditions.
DISCUSSIONThe occurrence of changes in sound frequency elicited athree-phase ERP waveform, in agreement with previousstudies using similar distraction paradigms [8±10,12]. Atearly latencies (� 150 ms), the MMN was elicited to alldeviance levels, its mean amplitude tending to increaselinearly as a function of the magnitude of change. Thesimilar scalp distribution of the MMN in all deviantconditions suggests larger activation of the same brainareas involved in MMN generation, rather than the con-tribution of additional brain regions to large deviancies.The pattern of growth of MMN amplitude to increasingdeviance found here contrasts with the logarithmic func-tions described in previous studies [13,14]. These logarith-mic functions were obtained in passive conditions, in thecontext of pre-attentive perceptual discrimination, andwere similar to those described in animals at neural level,underlying visual [15] or somatosensory [16] perception. Inthe present study, although irrelevant to task performance,the MMN-eliciting sounds could have been attended asthey warned the occurrence of the subsequent visualimperative stimulus [8]. In such a condition, some N1enhancement, shown to be involved in frequency changedetection [17] and contributing to the deviantÿstandarddifference wave to large deviancies [18], may have beenmodulated by attentional demands [19,20]. A previousstudy has also shown a linear increase of the neuralactivity, measured by fMRI, in primary visual cortex toincreasing behavioral accuracy in a visual-cued attentiontask [21]. Together with the linear functions obtained for
the P3a and RON responses, these results suggest thatattentional processes may follow different parametric lawsthan the well-established logarithmic functions underlyingperceptual and psychophysical processes.
Following the MMN, the P3a and RON responses wereelicited, their amplitude increasing linearly as a function ofdeviance, with a slope of growth about double than that ofthe MMN. The scalp distribution of the P3a was moreposteriorly distributed in the 80% deviant condition thanin the smaller deviant conditions, suggesting the engage-ment of different brain regions for the largest frequencychange. This parallels the different shape of the P3a wave-form in this condition, in which a second phase of theresponse appeared evident. In previous studies, two stagesof the P3a elicited to novel sounds have been described,with different functional roles and separate brain areascontributing to their generation as suggested by differentscalp distributions [8,9]. The present results point to theinvolvement of the second phase of the P3a in the proces-sing of stimulus salience, as it was observed only for thelargest deviant condition. At a third stage of activation ofthe cerebral network of involuntary attention, the RONresponse was elicited in all deviant conditions. The scalpdistribution analyses revealed that, similarly to MMN, thesame cerebral regions were engaged during its generationfor all levels of change, these being activated more stronglyto larger deviancies according to a linear function.
Interestingly, the present experiment yielded a lack ofparallel behavior between the electrophysiological dataand task performance, which needs clari®cation in futurestudies. Indeed, the larger activation of the brain networkunderlying involuntary attention with increasing the mag-nitude of change was not paralleled by an attentionalengagement re¯ected in behavioral distraction, i.e. subjectswere only distracted in the 10% deviant condition, whereasa facilitatory effect was observed in the 5% deviant condi-tion. At least two possible explanations may account forthis lack of electrophysiological±behavioral parallel. First,it is plausible that behavioral distraction can only beobserved when an optimal range of cerebral activation isreached, i.e, that obtained for a deviance around 10% [8,9].Once the critical interval of distracting deviance is sur-passed, the larger activation of the brain network under-lying involuntary control of attention may re¯ect acompensatory effect that would prevent behavioral distrac-tion from being manifested. In this context of interpreta-tion, the larger RON response to increasing magnitude ofchange may therefore arise from a compensatory effort toreorient attention effectively towards the original task. Thepresent results also suggest that RON re¯ects the reorient-ing to the originary task not necessarily after actual behav-ioral distraction [9,10,12], but after processing temporarilythe preceding distracting stimulus, a processing which is inturn re¯ected in the preceding P3a response.
The facilitatory effect observed in the 5% deviant condi-tion can also be explained in this context. A small deviantchange may suf®ce to activate change detection, as indexedby MMN, but may fail to trigger successive orienting ofattention, as indexed by the lack of a signi®cant P3a.However, these small changes succeed in causing facilita-tory effects on performance, probably via an enhancementof the processing of the subsequent visual stimuli. Recent
NEUROREPORT E. YAGO, M. J. CORRAL AND C ESCERA
4096 Vol 12 No 18 21 December 2001

evidences have suggested that sounds can facilitate per-formance of a visual task. For example, cross-modal facil-itation has been reported by McDonald et al. [22], whofound better performance to visual targets preceded bywarning tones informing on their location. Similar resultshave been reported by Hackley and Valle-InclaÂn [23] whofound electrophysiological facilitation (i.e. earlier latenciesof the lateralized readiness potential) at early stages ofperceptual processing to visual targets presented simulta-neously with a task-irrelevant sound. The present resultssuggest that, when a visual target is shortly preceded byan auditory change, facilitatory effects can be obtained forthose auditory changes near the discrimination level.
Alternatively, the context of stimulation may have had adeterminant in¯uence in the behavioral results, as inprevious studies distraction effects were obtained withdifferent contextual stimulation. In these studies, subjectswere presented with a similar auditory±visual paradigm inwhich the auditory stimulation intermixed deviant tonesand novel sounds among the repeating standard tones[8,9], whereas in the present study no novel sounds werepresented. It is plausible that the occurrence of novelsounds make the system controlling the orienting of atten-tion more vulnerable to distraction due to a lack of brainresources to cope with novelty, whereas the occurrence ofdifferent deviant tones may not overload the brain re-sources to cope with distraction. Indeed, recent evidence[24] has shown that higher memory load results in greaterinterference effects on behavioral performance from dis-tractor stimuli, as compared to a low memory load condi-tion. These results may also explain why other authors [12]have found distraction effects on performance to smalldeviancies, as the distractors were presented in the samesensory modality than the main task, probably interferingmuch more than in the present study, in which distractorswere presented in a different sensory modality than thestimuli of the primary task. Future studies should thereforeconsider the context of stimulation used to study thecerebral basis of involuntary attention and distraction, as itmay affect the load of memory resources and, conse-quently, the interference of distractors on executive per-formance.
CONCLUSIONWe found that the cerebral network underlying auditorychange processing, as indexed by MMN, P3a and RON,activates linearly as a function of increasing the magnitude
of change in sound frequency. Contrasting with the well-known logarithmic functions describing perceptual andpsychophysical processes, the present results suggest thatattentional processes are best characterized by linear func-tions. Scalp distribution analyses revealed that the samebrain regions contributed to the generation of MMN andRON responses in all deviant conditions, whereas addi-tional brain regions were engaged in salience processing,as indicated by the different scalp distribution and anadditional later phase of the P3a elicited to the largestdeviance. Surprisingly, the linear increase of activation inthis cerebral network was not paralleled by an attentionalengagement re¯ected in behavioral distraction, as subjectswere only distracted in the 10% deviant condition, whereasa facilitatory effect was observed in the 5% deviant condi-tion. This lack of electrophysiological±behavioral agree-ment re¯ecting either low working memory load or theexistence of an optimal range of activation in this cerebralnetwork for interfering executive performance, remains tobe clari®ed in future studies.
REFERENCES1. NaÈaÈtaÈnen R. Attention and Brain Function. Hillsdale: Erlbaum; 1992.
2. Escera C, Alho K, SchroÈger E et al. Audiol Neurootol 5, 151±166 (2000).
3. Friedman D, Cycowicz YM and Gaeta H. Neurosci Behav Rev 25,
355±373 (2001).
4. Squires NK, Squires KC and Hillyard SA. Electroencephalogr Clin
Neurophysiol 38, 387±401 (1975).
5. Knight RT. Electroencephalogr Clin Neurophysiol 59, 9±20 (1984).
6. Grillon C, Courchesne E, Ameli R et al. Arch Gen Psychiatry 47, 171±179
(1990).
7. Woods DL. Electroencephalogr Clin Neurophysiol 84, 456±468 (1992).
8. Escera C, Alho K, Winkler I et al. J Cogn Neurosci 10, 590±604 (1998).
9. Escera C, Yago E and Alho K. Eur J Neurosci 14, 877±883 (2001).
10. SchroÈger E and Wolff C. Neuroreport 9, 3355±3358 (1998).
11. SchroÈger E, Giard MH, Wolff C. Clin Neurophysiol 111,1450±1460 (2000).
12. Berti S and SchroÈger E. Cogn Brain Res 10, 265±273 (2001).
13. Tiitinen H, May P, Reinikainen K et al. Nature 370, 90±92 (1994).
14. Amenedo E and Escera C. Eur J Neurosci 12, 2570±2574 (2000).
15. Seidemann E, Zohary E and Newsome WT. Nature 394, 72±75 (1998).
16. Romo R, HernaÂndez A, Zainos A et al. Neuron 26, 273±278 (2000).
17. Yago E, Escera C, Alho K et al. Neuroreport 12, 2583±2587 (2001).
18. Scherg M, Vajsar J and Picton TW. J Cogn Neurosci 1, 336±355 (1989).
19. Hillyard SA, Hink RF, Schwent VL et al. Science 182, 177±180 (1973).
20. Teder W, Alho K, Reinikainen K et al. Psychophysiology 30, 71±81 (1993).
21. Rees D, Backus BT and Heeger DJ. Nature Neurosci 3, 940±945 (2000).
22. McDonald JJ, Teder-SaÈlejaÈrvi WA and Hillyard SA. Nature 407, 906±908
(2000).
23. Hackley SA and Valle-InclaÂn F. Nature 391, 786±788 (1998).
24. de Fockert JW, Rees G, Frith CD et al. Science 291, 1803±1806 (2001).
Acknowledgements: This research was supported by grant PM99-0167 of the Spanish Ministry of Science and Technology andthe Generalitat de Catalunya (1999FI-003 82-PGUB). We thank Vanessa Carral for her valuable help in the preliminary phases of
the study.
BRAIN IMAGING AS A FUNCTION OF AUDITORY CHANGE NEUROREPORT
Vol 12 No 18 21 December 2001 4097