Embed Size (px)
Citation preview

882
Activation of Calpain I and Hydrolysis ofCalpain Substrates (Actin-Binding Protein,
Glycoprotein Ib, and Talin) Are Nota Function of Thrombin-Induced
Platelet Aggregation
June D. Wencel-Drake, Janice R. Okita, Douglas S. Annis, and Thomas J. Kunicki
Calcium-activated neutral proteinase (calpain) has been shown to cleave proteins involved inthe maintenance of cell structure. In human platelets, substrates of calpain include glycopro-tein Ib (GPIb), actin-binding protein (ABP), and talin. GPIb-ABP complexes can be isolatedin detergent extracts and are thought to represent membrane-cytoskeleton attachment sites. Ithas been hypothesized that the hydrolysis of GPIb-ABP by calpain is regulated by the extentof binding of this proteinase to the plasma membrane-cytoskeleton interface with plateletactivation. Recently, another calpain substrate (talin) has been shown to redistribute from thecytoplasm to the plasma membrane-cytoskeleton interface as the result of thrombin stimula-tion. To investigate the intracellular distribution of calpain I, we employed the monoclonalantibody B27D8, specific for the heavy chain (catalytic subunit) of calpain I. Indirectimmunofluorescent staining of resting human platelets revealed undetectable surface antigen.Permeabilization with Triton X-100, however, revealed a diffuse intracellular antigen consistentwith a cytosolic distribution. To determine whether this antigen distribution reflected theproenzyme or the activated form of calpain I and to assess the degree of hydrolysis of ABP,GPIb, and talin, we employed B27D8 and murine monoclonal antibodies against ABP (1B3 and3D1), GPIb (LJIblO), and rabbit polyclonal antibodies against talin (A2 and Bl l ) in aquantitative immunotransblot assay. Examination of resting platelets revealed that calpain Iexisted as the 85 -kd proenzyme form and that ABP, GPIb, and talin existed in their nativeintact forms. When platelets were aggregated with thrombin, autoproteolysis of calpain Ioccurred within the 30 seconds required to completely solubilize platelet aggregates in sodiumdodecyl sulfate—containing buffer and not as a direct result of thrombin-induced activation.When EDTA was added to platelet samples before the addition of sodium dodecyl sulfate lysisbuffer, negligible proteolysis of calpain I, ABP, or talin occurred. When platelets were lysed withsodium dodecyl sulfate in the absence of EDTA, autoproteolysis of calpain I and cleavage ofABP and talin occurred. GPIb is not proteolyzed under either condition. In parallel immuno-fluorescent studies, examination of thrombin-stimulated platelets demonstrated minimalsurface staining. Permeabilization revealed a redistribution of the calpain I epitope to anintracellular circumferential ring pattern. We conclude that with thrombin-induced plateletactivation, calpain I redistributes intracellularly in the absence of activation of the 85-kdproenzyme form or significant hydrolysis of three important calpain substrates, ABP, GPIb,and talin. Failure to chelate divalent cations at the time of platelet lysis in sodium dodecylsulfate enables the redistributed calpain I in thrombin-activated platelets to efficientlyproteolyze ABP and talin. This proteolysis is initiated at the time of solubilization and isunrelated to thrombin aggregation per se. These findings raise further doubts concerning therole of calpain I in the events that initiate and maintain thrombin-induced platelet aggregation.(Arteriosclerosis and Thrombosis 1991;11:882-891)
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Wencel-Drake et al Platelet Calpain I 883
It is well documented that human platelets con-tain calcium-activated neutral proteinase(calpain, EC 3.4.22.17). Calpain activity is evi-
dent in platelet lysates created by sonication1 or bysolubilization in Triton X-1002 (Tx) in the absence ofcalpain inhibitors a*nd in the presence of free calciumions (Ca2+). In such lysates, calpain catalyzes thelimited proteolysis of actin-binding protein (ABP),3-4
talin/P235,1 glycoprotein Ib (GPIb),2 fibrinogen,5 andvon Willebrand factor,6 as evidenced by the require-ment for Ca2+ and inhibition by leupeptin, antipain,Af-ethylmaleimide, or mersalyl.
Fox and coworkers7 reported the hydrolysis ofABP and talin on treatment of human platelets withthrombin, collagen, or the Ca2+ ionophore A23187.Hydrolysis products of the two proteins were identi-fied and appeared to be identical to the cleavageproducts found when platelets are lysed in the ab-sence of calpain inhibitors (EDTA or leupeptin).Thus, it was concluded that on stimulation of plate-lets, calpain is activated and hydrolysis of at least twoof its substrates (ABP and talin) occurs.
Complexes containing GPIb and ABP can be iso-lated in detergent extracts of platelets.78 Such com-plexes are thought to represent attachment sitesbetween the plasma membrane and the cytoskeletonthat may be involved in maintaining the structure ofplatelets and in the shape change that occurs onplatelet activation.78 Because GPIb, ABP, and talinare substrates for calpain and hydrolysis of ABP andtalin has been reported to occur during plateletaggregation, it is thought that calpain hydrolysis ofGPIb-ABP complexes and/or talin may be involvedin platelet activation.7-9 Because thrombin stimula-tion results in the redistribution of talin to the innerface of the plasma membrane,9 one could be led tohypothesize that the proteolysis of calpain substratesis regulated by the degree to which calpain localizesto the plasma membrane-cytoskeleton interface onplatelet activation.
Two calpains requiring either low (calpain I) orhigh (calpain II) concentrations of Ca2+ have beendescribed in numerous tissues, including platelets.10
Muscle calpains I and II undergo limited autoprote-olysis, resulting in a decreased requirement for Ca2+
and an enhanced proteolytic activity.11-12 Recently,
From the Departments of Medical Laboratory Sciences andPharmacology (J.D.W.-D.), University of Illinois at Chicago, Chi-cago, 111., and the Blood Center of Southeastern Wisconsin, Inc.(J.R.O., D.S.A., T.J.K.) and Department of Anatomy and CellBiology (T.J.K.) Medical College of Wisconsin, Milwaukee, Wis.
Supported by National Heart, Lung, and Blood Institute grantHL-38489 (to J.D.W.-D.), American Heart Association of Metro-politan Chicago grant-in-aid (to J.D.W.-D.), National Heart,Lung, and Blood Institute grant HL-33925 (to J.R.O.), AmericanHeart Association of Wisconsin grant-in-aid No. 86-GA-68 (toJ.R.O.), and National Heart, Lung, and Blood Institute grantHL-32279 (to T.J.K.).
Address for correspondence: Dr. June D. Wencel-Drake, De-partment of Medical Laboratory Sciences (M/C 518, 690 CME),University of Illinois at Chicago, 808 South Wood Street, Chicago,IL 60612.
Received December 27,1990; revision accepted March 14,1991.
Samis and coworkers13 have reported that intracellu-lar platelet calpain I is not active until it is convertedto a 76-78-kd form. Autoproteolysis of calpain isthus an indicator of the state of activation of theenzyme. In the present study, we investigatedwhether calpain I undergoes autoproteolysis duringthrombin-induced platelet aggregation, whether thisactivation of the proteinase correlates with its redis-tribution to the site of GPIb-ABP interaction, andwhether such redistribution initiates hydrolysis of thesubstrates ABP, GPIb, and talin.
MethodsChemicals
The following were purchased from Sigma Chem-ical Co., St. Louis, Mo.: bovine serum albumin (re-crystallized once, essentially globulin free) and poly-L-lysine. Paraformaldehyde was obtained fromPolysciences, Warrington, Pa. Sepharose 2B, proteinA-Sepharose, and Sephadex G-100 superfine wereobtained from Pharmacia, Inc., Piscataway, N.J. Txwas purchased from J.T. Baker Chemicals, Phil-lipsburg, N.J. All other chemicals were reagentgrade. a-Thrombin was the generous gift of JohnFenton, New York State Department of Health,Albany, N.Y., and Richard A. Marlar, Blood Centerof Southeastern Wisconsin, Milwaukee, Wis.
Platelet Preparation
Platelet-rich plasma was prepared from acid/cit-rate/dextrose-anticoagulated whole blood obtainedfrom healthy aspirin-free volunteers as described.14
Washed platelets were subsequently prepared bycentrifugation and gel filtration on Sepharose 2B asdescribed.15 In activation studies platelets were stim-ulated with a-thrombin at concentrations of 0.2-1unit/ml at 37°C for 0.5, 1, 2, 3, or 5 minutes, at whichtime the cells were either fixed with 2% paraformal-dehyde and processed for routine immunofluores-cence studies or solubilized in sample buffer (25 mMtris[hydroxymethyl]aminomethane [Tris] HC1, 25%glycerol, 1% sodium dodecyl sulfate [SDS], pH 6.8,with or without 0.25 mM EDTA), vortexed immedi-ately, and processed for immunotransblot studies.
AntibodiesJohn S. Elce (Queen's University, Kingston, Can-
ada) kindly provided the cell line (B27D8 sc El l)that produces a mouse monoclonal immunoglobulin(Ig) Gl (B27D8) specific for the heavy chain (cata-lytic subunit) of calpain I.13 The specificity of B27D8was confirmed by quantitative immunotransblot of aplatelet cytosolic fraction as described.16 AlthoughB27D8 also reacts with calpain II, the cross reactivityis extremely low and negligible in these experi-ments.13 The cell line B27D8 sc El l was used toproduce ascites fluid in BALB/c BYJ mice (JacksonLabs, Bar Harbor, Me.), and IgG was purified bycombining and modifying17 the rivanol and caprylicacid procedures.18-19 For immunofluorescence stud-
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
884 Arteriosclerosis and Thrombosis Vol 11, No 4 July/August 1991
Calpain Actin
x
Ascites Prebleed
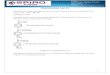
FIGURE 1. Photomicrographs showing im-munofluorescent localization of calpain I inresting platelets. Intact or detergent-permeabil-ized fixed platelets were stained with B27D8 (amonoclonal anti-calpain I antibody) and rab-bit anti-actin and counterstained with rhodam-inated goat anti-mouse immunoglobulin G(IgG) and fluoresceinated goat anti-rabbit IgG.Intact resting cells demonstrated minimal sur-face staining for both calpain (panel a) andactin (panel b). Permeabilization revealed ex-tensive immunofluorescent staining for bothproteins, which occupied the cell interior (pan-els c and d). Cells doubly labeled with irrele-vant ascites control (panel e) and rabbit pre-immune serum (panel f) did not demonstratesignificant labeling. Tx, Triton X-100. xl,200.
ies the IgG fraction was concentrated fivefold by useof a Centricon 30 microconcentrator (Amicon, Dan-vers, Mass.). The murine monoclonal anti-GPIb an-tibody LJIblO was a gift of Zaverio Ruggeri, ScrippsClinic and Research Foundation, La Jolla, Calif.20
Two monoclonal antibodies, 1B3 and 3D1, directedagainst ABP and its major proteolytic fragments at190, 100, and 90 kd were kindly provided by RobertEzzell, Whitehead Institute for Biomedical Re-search, Cambridge, Mass.21 Rabbit polyclonal anti-
bodies A2 and Bll raised against avian smoothmuscle talin and human platelet talin, respectively,were generously provided by Mary Beckerle, Univer-sity of Utah, Salt Lake City, Utah.9
Immunofluorescence Microscopy
Immunofluorescent staining of platelets was per-formed as previously described.16 Briefly, resting orthrombin-stimulated platelets were fixed with 2%paraformaldehyde on ice for 1 hour. Unreacted
Calpain Actin
xh-
FIGURE 2. Photomicrographs show-ing immunofluorescent localization ofcalpain I in thrombin-stimulatedplatelets. Examination of intact cellsthat had been stimulated with 1unit/ml thrombin for 30 seconds re-vealed minimal surface staining (pan-el a). In these experiments, actin la-beling was used as an internal controlto establish that cells were imperme-able to antibody when stained in theabsence of Triton X-100 (Tx) (panelb). Permeabilization, however, re-vealed the apparent redistribution ofthe B27D8 epitope to a peripheralintracellular ring pattern (panel c),which generally colocalized with intra-cellular actin (panel d). y.1,200.
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
Wencel-Drake et al Platelet Calpain I 885
Calpain Actin
cE
cE
CVJ
cE
CO
cE10
FIGURE 3. Photomicrographs showing time course ofredistribution of calpain I. Platelets were stimulated with 1unit/ml thrombin and stained as in Figure 2. Examinationof additional timepoints at 1 (panels a and b), 2 ("panelsc and d), 3 (panels e and i), and 5 (panels g and h)minutes (min.) revealed that redistribution of calpain I to aperipheral ring pattern is maintained over time, x 1,200.
aldehyde was blocked with NH4Cl-Tris-buffered sa-line, pH 7.4, and the cells were permitted to settle onpoly-L-lysine-coated glass coverslips. In some cases,the cells were treated with 0.1% Tx for 3 minutes to
3 4 8 98 5 , .8 1 - 17 8 - '
EDTA
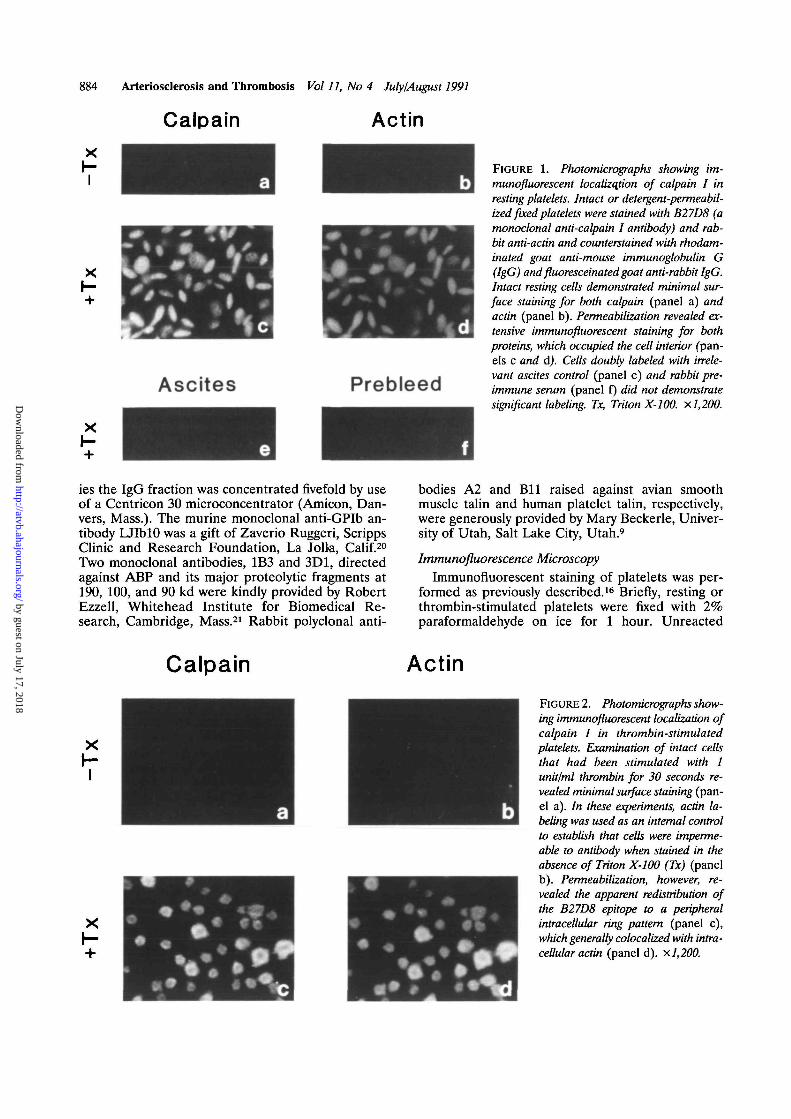
FIGURE 4. Photograph of immunotransblot assay of calpainI in resting platelets. Solubilized platelet proteins were reducedwith 5% fi-mercaptoethanol and resolved by 5% sodiumdodecyl sulfate-potyacrylamide gel electrophoresis. Proteinswere electrophoretically transferred to nitrocellulose, andcalpain I heavy chain was detected with B27D8 followed byiodine-125 -labeled rabbit anti-mouse immunoglobulin. Onlythe portion of the transblot autoradiogram containing the threeB27D8-reactive bands is shown. A standard (lane 1), preparedas described in "Methods," was run on each gel to demon-strate that mobilities of three calpain I bands (85, 81, and 78kd) can be distinguished. Composite transblots of gel-filteredplatelets (lanes 2-5), minus external Ca2+ (lanes 6 and 7),plus external Ca2+ (lanes 8 and 9), and in the absence (lanes2, 4, 6, and 8) or presence (lanes 3, 5, 7, and 9) of 0.25 mMEDTA in solubilization buffer.
render them permeable to antibody before staining.The permeable or intact cells were then rinsed withTris-buffered saline containing 0.1% bovine serumalbumin and incubated for 20 minutes with either theprimary antibody (B27D8), an irrelevant ascites, orpreimmune serum control. After the cells wererinsed they were stained with rhodaminated goatF(ab')2 anti-mouse IgG (Organon-Technika, CappelDivision, Malvern, Pa.). In double-label experiments,cells were additionally stained with rabbit anti-actin(ICN Biomedicals, Inc., Costa Mesa, Calif.) andcounterstained with a fluoresceinated goat anti-rab-bit IgG (Organon-Technika).
The platelets were viewed with a Zeiss standard16 microscope equipped with an HBO 50-W mer-cury lamp, an IVF1 epifluorescence condenser, BP485 and 546 excitation filters, and BP 520-560 andLP 590 barrier filters and were photographed withKodak Tri-X panchromatic film (Eastman Kodak,Rochester, N.Y.)
Quantitative Immunotransblot Assay for Calpain ICalpain I heavy-chain antigen was quantified by
immunotransblot assay as described.22 Briefly, solubi-lized platelet proteins were reduced with 5% /3-mer-
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from

886 Arteriosclerosis and Thrombosis Vol 11, No 4 July/August 1991
81 —
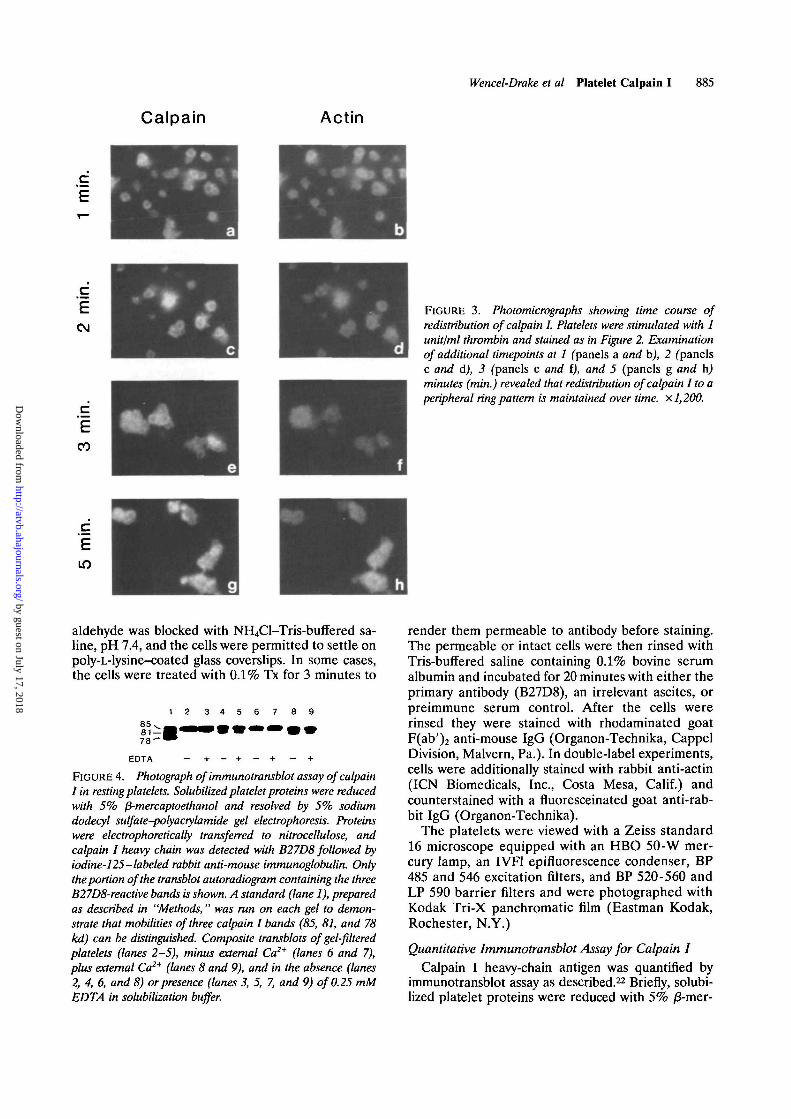
FIGURE 5. Photograph of immunotransblot assay of calpainI in thrombin-stimulated platelets. Standards (lane 1) andcomposite transblots of thrombin-stimulated platelets wereprepared as in Figure 4. Sample treatments were 1 unit/mlthrombin (lane 2), 0.05 unit/ml thrombin (lane 3), and 1unit/ml thrombin, with 0.25 mM EDTA added before lysis(lane 4). Time of stimulation for all samples was 1 minute.
captoethanol and were resolved by 5% SDS-poly-acrylamide gel electrophoresis (PAGE) to allowresolution of 85-, 81-, and 78-kd bands. The proteinswere electrophoretically transferred to nitrocellulose,and calpain I heavy chain was detected with B27D8followed by iodine-125-labeled rabbit anti-mouse Ig.The labeled bands were cut out using the autoradio-graph as a guide, and the amount of 125I was quanti-fied. Additionally, parallel samples were transblottedwith LJIblO (to detect GPIb and its proteolytic prod-uct glycocalicin), with 1B3 and 3D1 (to detect ABPand its major proteolytic fragments at 190,100, and 90kd), and with A2 and Bll (to detect talin).
To verify the separation of calpain I antigen intothree bands on each gel, a standard was prepared asfollows. A platelet cytosolic fraction was incubatedat 4°C for 1-5 minutes in the presence of lowconcentrations of Ca2+ (0.2 mM). Under theseconditions, calpain I heavy chain undergoes limitedautoproteolysis. Aliquots were chosen that con-tained significant amounts of the three forms ofcalpain I (85, 81, and 78 kd). These standardpreparations were run on SDS-PAGE gels alongwith test samples to verify the separation of calpainI into three distinct bands.
ResultsImmunofluorescent Localization of Calpain I inResting and Thrombin-Stimulated Platelets
Intact resting platelets doubly labeled with B27D8and rabbit anti-actin demonstrated minimal surface
staining (Figures la and lb). Cells made permeablewith Tx demonstrated extensive intracellular stainingfor both antigens (Figures lc and Id), while irrele-vant ascites (Figure le) and rabbit preimmune con-trols (Figure If) remained negative. Intact plateletsthat had been stimulated with 1 unit/ml thrombin for30 seconds also demonstrated minimal surface label-ing (Figures 2a and 2b). These cells, subsequentlystained in the permeable state, revealed a centralizedclearing or an apparent redistribution of the B27D8antigen to a peripheral fluorescent ring pattern (Fig-ure 2c). That this pattern represents an internalredistribution is evidenced by the lack of labelingwhen cells were stained in the intact state.
Examination of multiple time points revealedthat the peripheral distribution of calpain I ismaintained while the cells subsequently aggregate(Figure 3). Based on the redistribution of calpain Iantigen seen by immunofluorescence microscopy,we were compelled to evaluate the state of activa-tion of this protease under the same conditions. Todo so, we took advantage of the fact that onceactivated, calpain I undergoes autoproteolysis andthe apparent molecular weight of its catalytic sub-unit decreases from 85 to 81 and 78 kd.
Calpain I Catalytic Subunit in Resting PlateletsBy immunotransblot assay, the majority of resting
gel-filtered platelet preparations contained calpain Ias a single 85-kd band (Figure 4, lanes 2 and 3).However, some preparations demonstrated signifi-cant amounts of the 81-kd band (Figure 4, lanes 4and 5). To determine the effect of external Ca2+ onthis apparent variability, gel-filtered platelets wereincubated at 37°C for 30 minutes in the presence orabsence of 2 mM external Ca2+. In the absence ofexternal Ca2+, no detectable amounts of the 81-kdform of calpain I were observed (Figure 4, lanes 6and 7). In contrast, the 81-kd band was consistentlyobserved in cells incubated in the presence of exter-nal Ca2+ (Figure 4, lanes 8 and 9). The amount of the85- or 81-kd forms of calpain I in resting platelets was
— 205
— GPIb
— 103
-68
1 2 3 4Thrombin — + — +
EDTA - - + +
5 6 7 8 9
FIGURE 6. Photograph of immunotransblot assay ofcalpain I and gtycoprotein lb (GPIb) in resting and throm-bin-stimulated platelets. Resting or thrombin-stimulatedplatelets (1 unit/ml, 5 minutes, 37 °C) were spiked with 0.25mM EDTA or buffer before solubilization. Calpain I trans-blots (lanes 1-4) were prepared as in Figure 4. Parallelsamples blotted with a monoclonal anti-GPIb antibodyLJIblO are shown in lanes 6-9. Molecular weight standardswere run in lane 5. Protein band at a molecular weight of 68kd represents platelet albumin that nonspecifically adsorbsLJIblO antibody in this assay.
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from
Wencel-Drake et al Platelet Calpain I 887
c IOCH
O)
IOOJ
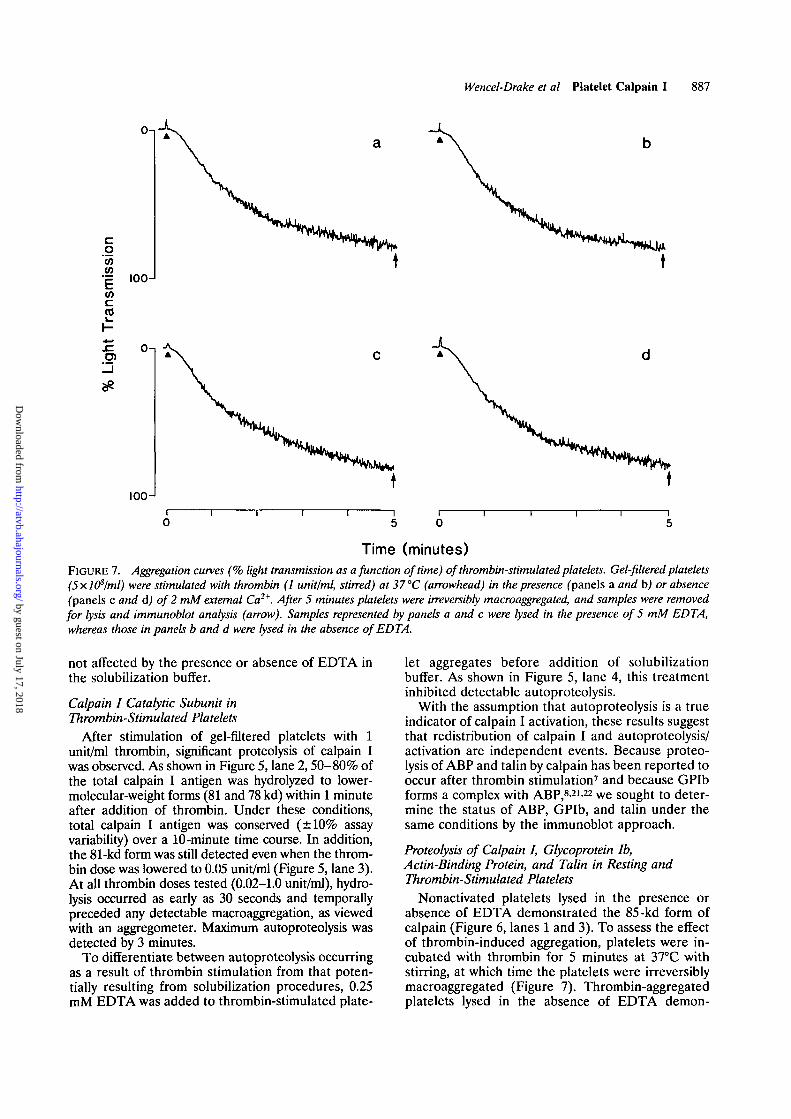
Time (minutes)FIGURE 7. Aggregation curves (% light transmission as a function of time) of thrombin-stimulated platelets. Gel-filtered platelets(5x.l(Plml) were stimulated with thrombin (1 unit/ml, stirred) at 37 °C (arrowhead) in the presence (panels a and b) or absence(panels c and d) of 2 mM external Ca2*. After 5 minutes platelets were irreversibly macroaggregated, and samples were removedfor lysis and immunoblot analysis (arrow). Samples represented by panels a and c were lysed in the presence of 5 mM EDTA,whereas those in panels b and d were lysed in the absence of EDTA.
not affected by the presence or absence of EDTA inthe solubilization buffer.
Calpain I Catalytic Subunit inThrombin-Stimulated Platelets
After stimulation of gel-filtered platelets with 1unit/ml thrombin, significant proteolysis of calpain Iwas observed. As shown in Figure 5, lane 2,50-80% ofthe total calpain I antigen was hydrolyzed to lower-molecular-weight forms (81 and 78 kd) within 1 minuteafter addition of thrombin. Under these conditions,total calpain I antigen was conserved (±10% assayvariability) over a 10-minute time course. In addition,the 81-kd form was still detected even when the throm-bin dose was lowered to 0.05 unit/ml (Figure 5, lane 3).At all thrombin doses tested (0.02-1.0 unit/ml), hydro-lysis occurred as early as 30 seconds and temporallypreceded any detectable macroaggregation, as viewedwith an aggregometer. Maximum autoproteolysis wasdetected by 3 minutes.
To differentiate between autoproteolysis occurringas a result of thrombin stimulation from that poten-tially resulting from solubilization procedures, 0.25mM EDTA was added to thrombin-stimulated plate-
let aggregates before addition of solubilizationbuffer. As shown in Figure 5, lane 4, this treatmentinhibited detectable autoproteolysis.
With the assumption that autoproteolysis is a trueindicator of calpain I activation, these results suggestthat redistribution of calpain I and autoproteolysis/activation are independent events. Because proteo-lysis of ABP and talin by calpain has been reported tooccur after thrombin stimulation7 and because GPIbforms a complex with ABP,821-22 we sought to deter-mine the status of ABP, GPIb, and talin under thesame conditions by the immunoblot approach.
Proteolysis of Calpain I, Glycoprotein Ib,Actin-Binding Protein, and Talin in Resting andThrombin-Stimulated Platelets
Nonactivated platelets lysed in the presence orabsence of EDTA demonstrated the 85-kd form ofcalpain (Figure 6, lanes 1 and 3). To assess the effectof thrombin-induced aggregation, platelets were in-cubated with thrombin for 5 minutes at 37°C withstirring, at which time the platelets were irreversiblymacroaggregated (Figure 7). Thrombin-aggregatedplatelets lysed in the absence of EDTA demon-
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from

888 Arteriosclerosis and Thrombosis Vol 11, No 4 July/August 1991
TABLE 1. Quantification of Thrombin-Induced Proteolysis of Calpain I, Actin-Binding Protein, and Talin
Calpain (B27D8)
85 kd
81 kd
78 kd
Total
ABP (1B3)
Parent
190 kd
Total
ABP (3D1)
Parent
100 kd
90 kd
90 kd
Total
Talin (A2)
223 kd
200 kd
Total
Talin (Bll)
223 kd
200 kd
Total
Without thrombinor EDTA
l,163±163(90±4)
69±26(5±2)
54±26 (4±2)
1,285
1,616±117(98±1)
28±14(2+1)
1,645
1,655±185(99±1)
12±20(l±l)
5±7(0.±l)
5±7(0.±l)
1,672
773±139(90±8)
93±98(10±8)
866
4,825 ±703 (95 ±2)
233±134(4±2)
5,058
With thrombin,without EDTA
1,021+264(67+16)
371 + 149 (24±9)
146+138 (9±8)
1,538
1,856+641 (97±2)
59+72 (3±2)
1,916
1,517+60 (93±2)
80+32 (5 ±2)
80+32 (5 ±2)
38+29 (2±2)
1,635
633+103 (78± 14)
190+150(22+14)
823
4,010+660 (70±7)
l,680±611 (29±7)
5,690
Without thrombin,with EDTA
1,115±147(92±3)
60±26(5±2)
35±17(3±2)
1,210
1,726±133(98±1)
28±14(2±1)
1,754
1,675±165(96±2)
39±30 (2±2)
39±30 (2±2)
36+24 (2±1)
1,750
736 ±49 (93 ±6)
54±53 (7±6)
790
4,775+738 (94 ±4)
239±62(5±2)
5,014
With thrombinand EDTA
1,214±26 (89±4)
96+37 (7±2)
57±23 (4+2)
1,366
l,850±163(98±2)
47±43 (2±2)
1,898
1,717±34(97±1)
37+12 (2±1)
37+12 (2±1)
26±24(1±1)
1,780
741 ±96 (91+4)
78±32 (10±4)
819
4,671 ±675 (91 ±3)
455 + 192(9+3)
5,126
Values are counts per minute obtained from corresponding gel slices and represent mean±SD of four determinations.Percentage of total counts per minute is in parentheses.ABP, actin-binding protein.
strated significant proteolysis with the appearance ofthe 81- and 78-kd forms of calpain (Figure 6, lane 2).Inclusion of EDTA to such aggregates before lysis(Figure 6, lane 4) resulted in no detectable 81- or 78-kd forms. Parallel studies run on the same lysatesblotted with a monoclonal anti-GPIb antibody,LJIblO, indicated that GPIb remained intact underall conditions tested (Figure 6, lanes 6-9).
To evaluate activation of calpain I and hydrolysisof ABP and talin, platelet lysates from four individ-uals were blotted with a panel of monoclonal andpolyclonal antibodies. The results obtained with eachof the four individuals were nearly identical, andmean values are listed in Table 1. Figure 8 depicts atypical set of results from one individual. Lysatesblotted with B27D8 confirmed that autoproteolysis ofcalpain I occurred only when EDTA was omittedfrom thrombin-stimulated samples before lysis (Fig-ure 8, lanes 1-4; and Table 1).
Parallel blots prepared with 3D1 and 1B3, mono-clonal anti-ABP antibodies that recognize the 190-and 90-100-kd proteolytic fragments of ABP, respec-tively, are shown in Figure 8. When 3D1 was used asthe primary antibody, 190-kd fragments of ABP weredetected in platelets aggregated by thrombin (Figure8, lanes 5-8). 1B3 blots revealed detectable 100- and90-kd fragments in identical samples (Figure 8, lanes9-12). The percentage of degraded ABP, estimated
by excising the bands identified by autoradiographyand determining the counts per minute of 125I-labeledsecond antibody, is shown in Table 1.
These data suggest that a low level of proteolysis(3-7%) of ABP may occur as a result of platelet lysisin the absence of EDTA after thrombin activation.However, when EDTA was added to the thrombinaggregates before lysis, the counts per minute asso-ciated with the 90-, 100-, and 190-kd bands wereessentially eliminated with no evidence of ABP di-gestion (Table 1). Blots prepared with A2, specificfor avian smooth muscle talin (Figure 8, lanes 13-16), and Bll, specific for human platelet talin (Fig-ure 8, lanes 17-20), yielded similar results (Table 1).In the case of talin, the 200-kd fragment represents9-10% of the total antigen when thrombin-aggre-gated platelets are lysed in the presence of EDTA.This compares with a value of 5-7% when the sameplatelets are lysed in the presence of EDTA but nottreated first with thrombin. On the other hand, thesame fragment represents 22-29% of antigen whenplatelets are aggregated by thrombin but lysed in theabsence of EDTA.
DiscussionIn the resting platelet, the calpain I catalytic
subunit displays a homogeneous cytosolic distribu-tion. This observation is in agreement with previous
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Wencel-Drake et al Platelet Calpain I 889
_n ARP 19O n ABP 100 /90 a Talin (ASM) a Talin (H)
KBP —
190 —
— 223
— 200
9 10 11 12 13 14 15 161 2 3 4 5 6 7 8
Thrombin - + - + - + - +
EDTA - - + + - - + + - - + + - - + + - - + +
FIGURE 8. Photograph of immunotransblot assay of calpain I, actin-binding protein (ABP), and talin in resting andthrombin-stimulated platelets. Platelet lysates, prepared as in Figure 6, were run in parallel and transblotted with B27D8, amonoclonal antibody directed against calpain I; 1B3 or 3D1, monoclonal antibodies directed against ABP and its proteolyticfragments at 90, 100, and 190 kd; and A2 or Bll, polyclonal rabbit antibodies directed against avian smooth muscle talin andhuman platelet talin, respectively. Lanes 1-4, B27D8 transblots; lanes 5-8, 1B3 transblots; lanes 9-12, 3D1 transblots; lanes13-16, A2 transblots; and lanes 17-20, Bll transblots. A total of four individuals were studied in parallel experiments.Representative blots from a single individual are shown.
subcellular fractionation studies that indicate that95% of the total calpain I antigen as well as morethan 95% of the total calpain proteolytic activity arefound in the cytosolic fraction.116
When platelets were activated by thrombin, weobserved a redistribution of the calpain I antigen intoa circumferential ring. This redistribution occurredwithin 30 seconds after addition of thrombin togel-filtered platelets. Whether this reflects directedmovement of calpain I antigen or is a passive eventresulting from cytoskeletal reorganization and gran-ule centralization remains to be determined. Therequirement for permeabilization before visualiza-tion of the calpain I antigen by immunofluorescencemicroscopy suggests that the epitope defined byB27D8 is not accessible on the platelet surface, eitherin the resting state or after thrombin stimulation. Thepresumptive localization of calpain I to the cytoplas-mic face of the plasma membrane and the fact thatcalpain is thought to exert its proteolytic effect oncomponents of the plasma membrane-cytoskeletoninterface1-4-7 would be consistent with the argumentthat redistribution of calpain I is involved in modu-lation of membrane-cytoskeleton attachment sitesduring platelet activation.
To determine whether redistributed calpain I waspresent in its proenzyme or activated form, we em-
ployed a quantitative immunotransblot assay.16 Rest-ing gel-filtered platelets incubated in the absence ofexternal Ca2+ or in the presence of prostaglandin Ejplus EDTA16 demonstrated the 85-kd form ofcalpain I. In contrast, platelets incubated in thepresence of external Ca2+ exhibited both the 85- andthe 81-kd forms. These results suggest that in theresting cell, calpain I exists as the 85-kd form andthat a variable amount of proteolysis occurs duringthe process of gel filtration. Similarly, when plateletswere stimulated with thrombin, we observed thatautoproteolysis of calpain I occurred within the 30-second time frame required to completely solubilizeplatelet aggregates in SDS-containing buffer. Addi-tion of EDTA to the aggregate before solubilizationcompletely inhibited detectable autoproteolysis.With the assumption that autoproteolysis is a truereflection of calpain activation, these results suggestthat calpain I is not activated as a result of thrombin-induced platelet aggregation, in agreement with therecent reports by Samis et al13 and Fox and Elce.23
Given the magnitude of the previously reportedproteolysis of ABP and talin by calpain after throm-bin stimulation7 and the known association of GPIbwith ABP,82122 we sought to determine the status ofABP, GPIb, and talin under the same conditions withthe immunoblot method. GPIb remained intact un-
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from

890 Arteriosclerosis and Thrombosis Vol 11, No 4 July/August 1991
der all conditions tested. In contrast, the 90-, 100-,and 190-kd proteolytic fragments of ABP were de-tected in platelets aggregated by thrombin. Afterelimination of artifactual proteolysis of ABP associ-ated with lysis in the absence of EDTA, it is apparentthat the degree of degradation of ABP is consider-ably smaller than previous estimates7 and is essen-tially negligible. These results question the signifi-cance of ABP proteolysis during thrombin-inducedaggregation. While low-level degradation of ABPlocalized to specialized regions of the cytoskeletonduring thrombin-induced aggregation may be impor-tant, more precise quantification of that degree ofhydrolysis is required to document this point. Todate, none of the approaches that have been used toquantify ABP hydrolysis have had the necessary de-gree of sensitivity. The quantitative immunoblot assayemployed in this study is more sensitive than previ-ously applied techniques7 yet still fails to detectsignificant hydrolysis of ABP as a result of thrombin-induced aggregation. Similarly, quantitative immu-notransblots with antibodies against talin revealed alack of significant proteolysis occurring as a result ofthrombin stimulation.
Our results seriously question a role for calpain Iin the events that initiate and complete irreversibleplatelet aggregation. In addition, our results indicatethat autoproteolysis of calpain I does not coincidewith and is probably not required for its peripheralredistribution as a result of thrombin activation (ag-gregation). Thus, calpain migrates to the plateletperiphery in an inactivated form. If autoactivation viaproteolysis is then required for proteolysis of ABP ortalin, then this, too, must occur at the point ofsubsequent platelet solubilization. A most intriguingobservation that follows from our data is that plateletaggregation brings platelet calpain and certain of itssubstrates (e.g., talin) in close physical proximity,thereby predisposing them to subsequent proteolysis.The exact nature of this physical proximity and therole, if any, of subsequent proteolysis in the eventsthat follow activation remain interesting questionsthat beg further investigation.
AcknowledgmentsThe authors gratefully acknowledge the generous
gift of the murine hybridoma cell line of B27D8 scEl l from John S. Elce, Queen's University, Kingston,Canada. LJIblO, a monoclonal anti-GPIb antibody,was a gift of Zaverio Ruggeri, Scripps Clinic andResearch Foundation, La Jolla, Calif. Two monoclo-nal antibodies designated 1B3 and 3D1 directedagainst ABP were kindly provided by Robert Ezzell,Whitehead Institute for Biomedical Research, Cam-bridge, Mass. Additionally, rabbit polyclonal anti-bodies A2 and Bll raised against avian smoothmuscle talin and human platelet talin, respectively,were generously provided by Mary Beckerle, Univer-sity of Utah, Salt Lake City, Utah. Finally, we wouldlike to acknowledge the technical expertise and pho-tography of Michael G. Dieter.
References1. Phillips DR, Jakabova M: Ca++-dependent protease in human
platelets: Specific cleavage of platelet polypeptides in thepresence of added Ca++. J Biol Chem 1977;252:5602-5605
2. Solum NO, Olsen TM, Gogstad GO, Hagen I, Brosstad F:Demonstration of a new glycoprotein Ib-related component inplatelet extracts prepared in the presence of leupeptin. Bio-chim Biophys Acta 1983;729:53-61
3. Truglia JA, Stracher A: Purification and characterization of acalcium-dependent sulfhydryl protease from human platelets.Biochem Biophys Res Commun 1981;100:814-822
4. White GC II: Calcium-dependent protease in platelets:Response of calcium-activated protease in normal and throm-basthenic platelets to aggregating agents. Biochim BiophysActa 1980;631:130-138
5. Kunicki TJ, Mosesson MW, Pidard D: Cleavage of fibrinogenby human platelet calcium-activated protease. Thromb Res1984;35:169-182
6. Kunicki TJ, Montgomery RR, Schullek J: Cleavage of humanvon Willebrand factor by platelet calcium-activated protease.Blood 1985;65:352-356
7. Fox JEB, Reynolds CC, Phillips DR: Calcium-dependentproteolysis occurs during platelet aggregation. / Biol Chem1983;258:9973-9981
8. Fox JE: Identification of actin-binding protein as the proteinlinking the membrane skeleton to glycoproteins on plateletplasma membranes. / Biol Chem 1985;260:11970-11977
9. Beckerle MC, Miller DE, Bertagnolli ME, Locke SJ: Activa-tion-dependent redistribution of the adhesion plaque protein,talin in intact human platelets. / Cell Biol 1989;109:3333-3346
10. Sakon M, Kambayashi JI, Ohno H, Kosaki G: Two forms ofCa++-activated neutral protease in platelets. Thromb Res1981;24:207-214
11. Dayton WR: Comparison of low- and high-calcium-requiringforms of the calcium-activated protease with their autocata-lytic breakdown products. Biochim Biophys Acta 1982;709:166-172
12. Hathaway DR, Werth DK, Haaeberle JR: Limited autolysisreduces the Ca++ requirement of a smooth muscle Ca++-activated protease. / Biol Chem 1982;257:9072-9077
13. Samis JA, Zboril G, Elce JS: Calpain I remains intact andintracellular during platelet activation: Immunochemical mea-surements with monoclonal and polyclonal antibodies. Bio-chem J 1987;246:481-488 '
14. Wencel-Drake JD, Plow EF, Zimmerman TS, Painter RG,Ginsberg MH: Immunofluorescent localization of adhesiveglycoproteins in resting and thrombin stimulated platelets. AmJPathol 1984;115:156-164
15. Ginsberg MH, Plow EF: Fibronectin expression on the plate-let surface occurs in concert with secretion. / Supramol Struct1981;17:91-98
16. Okita JR, Frojmovic MM, Kristopeit S, Wong T, Kunicki TJ:Montreal platelet syndrome: A defect in calcium-activatedneutral proteinase (calpain). Blood 1989;74:715-721
17. Audran R, Pejaudier L, Steinbuch M: Preparation of iramu-noglobulin G, A, M (IgGAM) for therapeutic use: Conditionsfor enrichment in IgA or IgM. Rev Fr Transfus Immunohematol1975;18:119-135
18. Franek F: Purification of IgG monoclonal antibodies fromascites fluid based on Rivanol precipitation. Methods Enzymol1986;121:631-638
19. Steinbuch M, Audran R: The isolation of IgG from mamma-lian sera with the aid of caprylic acid. Arch Biochem Biophys1969;134:279-289
20. Handa M, Titani K, Holland LZ, Roberts JR, Ruggeri ZM:The von Willebrand factor-binding domain of platelet mem-brane glycoprotein Ib. J Biol Chem 1986;261:12579-12585
21. Ezzell RM, Kenney DM, Egan S, Stossel TP, Hartwig JH:Localization of the domain of actin-binding protein that bindsto membrane glycoprotein Ib and actin in human platelets. JBiol Chem 1988;263:13303-13309
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Wencel-Drake et al Platelet Calpain I 891
22. Okita JR, Pidard D, Newman PJ, Montgomery RR, Kunicki aggregation in human platelets (abstract). J CellBiol 1989;107:TJ: On the association of glycoprotein Ib and actin-binding 391aprotein in human platelets. / Cell Biol 1985;100:317-321
23. Fox MJ, Elce JS: Calpain I activation is not correlated with KEY WORDS • immunofluorescence • quantitativethrombin, PMA, or A23187-induced serotonin release and immunotransblot assay • calpain I
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from

J D Wencel-Drake, J R Okita, D S Annis and T J Kunickiglycoprotein Ib, and talin) are not a function of thrombin-induced platelet aggregation.
Activation of calpain I and hydrolysis of calpain substrates (actin-binding protein,
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 1991 American Heart Association, Inc. All rights reserved.
Avenue, Dallas, TX 75231is published by the American Heart Association, 7272 GreenvilleArteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.11.4.8821991;11:882-891Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/11/4/882World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document.Permissions and Rights Question and AnswerFurther information about this process is available in theis being requested is located, click Request Permissions in the middle column of the Web page under Services.Clearance Center, not the Editorial Office. Once the online version of the published article for which permission
can be obtained via RightsLink, a service of the CopyrightArteriosclerosis, Thrombosis, and Vascular Biology Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on July 17, 2018http://atvb.ahajournals.org/
Dow
nloaded from