Embed Size (px)
Citation preview

Biophysical Chemistry 24 (1986) 249-257 Elscvier
249
BPC 01076
ACTIVITY COEFFICIENTS OF SALTS IN HIGHLY CONCENTRATED PROTEIN SOLUTIONS
II. POTASSIUM SALTS IN ISOIONIC BOVINE SERUM ALBUMIN SOLUTIONS
M.D. REBOIRAS *, H. PFISTER and H. PAULY Institut fiir Radicilogir dw Universitiit Erlangen-Niirnherg, D-8520 Erlangen, E:R.G.
Received 13th February 1985 Revised manuscript 24th January 1986 Accepted 5th May 1986
Key words: Protein solution; Bovine serum albumin: Activity coefficient; Ion binding
Mean activity coefficients of different potassium salts KX (X = F-. Cl-. Br-. I-, NO;, SCN- ) have been measured in concentrated isoionic hovine serum albumin (BSA) solutions, by use of the EMF method with ion-exchange membrane electrodes. These solutions may be regarded as simple model systems for the cytoplasm of living cells as far as the influence of the macromolecular component on the activity coefficients of the salts is concerned. Two series of measurements have been carried out: (a) varying the amount of salt from 0.01 to 0.5 molal and maintaining the BSA concentration constant at 20 wt% and (b) varying the protein concentration up to 25 wt% and keeping the salt concentration constant at 0.1 m&l. For all salts studied, the mean activity coefficients in the protein-salt solutions increase as the salt concentration rises, when the protein concentration is maintained constant. In the series of measurements (b) the activity coefficients of all salts change linearly with the protein concentration. Marked qualitative differences, however, were observed depending on the anion species, which could be interpreted in terms of specific ion hinding of X- to the protein molecule. By taking into account BSA-bound ‘non-solvent’ water, the results were analyzed in terms of numbers of anions bound per BSA molecule. Comparison with the results of Scatchard, obtained at low protein concentrations, showed only a very small electrostatic effect of the BSA-(X- ), polyions on the activity coefficient of the salts at higher protein and salt concentrations.
1. Introduction
Proteins and water are the main components of the cytoplasm in living cells, as far as the weight percentages are considered. Electrolytes, in the form of inorganic salts, although present in much smaller weight percentages, play a significant role in the characteristics of the cytoplasm due to their important contribution to fundamental parame- ters such as the colligative properties of the sys- tem.
Since the physicochemical state of the cyto-
plasm components, as measured by their chemical potentials, is not completely known at the present time, it is not possible to give a clear explanation of the cytoplasmatic structure which accounts for the transport of molecules and ions in the cyto- plasm or across the cell membrane.
The activity coefficient of a component in a solution is a measure of the interactions of the component with all the other components in the solution. In the experiments we are reporting here, the assumption is made that the interactions of salts with the protein macromolecules in con- centrated protein-salt solutions are of the same nature as in the cytoplasm, which thus is consid- ered as a macromolecule-salt solution, the ‘solu- tion hypothesis’, in contrast with the ‘association-
* Present address: Departamento de Electroquimica, Facultad de Ciencias. Universidad Autbnoma de Madrid. 28034 Madrid, Spain.
0301.4622/86/$03.50 0 1986 Elsevier Science Publishers B.V. (Biomedical Division)

250 M, D. Rehoiras el al./Artiorty coefficients of salts in protein .wlutims
induction’ hypothesis which regards the cytoplasm as a quasi-solid-state structure.
In a previous study [l] we have determined the activity coefficients of (1,l) salts in bovine serum albumin (BSA) at different protein concentrations (from 0 up to 22 wt%), maintaining the salt con- centration constant at 0.1 molal. The determina- tion was carried out by means of potentiometric
I I 1
Reference saturated KC1 ; KX electrode i solution I
I I
titrations using membrane electrodes. We were then able to show that the activity coefficients of alkaline chlorides in isoionic BSA solutions de- crease very slightly with the protein concentration. This behaviour was explained in terms of the interactions between the components of the solu- tion, the Cl- specifically bound to the protein molecule playing an important role. A significant effect of the cation species was only observed for Li’.
The object of the present work is to study the activity coefficients of different potassium salts KX (X = F-, Cl-, Br-, II, NO,, SCN-) in con- centrated BSA solutions, in order to obtain further information about the interactions of small ions with the protein molecules. Two series of measure- ments were carried out: (a) varying the salt con- centration from 0.01 to 0.5 molal and maintaining the BSA concentration constant at 20 wt%, and (b) varying the macromolecular concentration up to 25 wt%, thus amply covering the ‘average’ protein concentration range corresponding to the cytoplasm of living cells, with the salt concentra- tion being maintained constant at 0.1 molal.
The interactions of low molecular weight ligands with proteins have been studied extensively and for this purpose, there are several experimental methods available [2I. Considerable attention has also been paid to theoretical interpretations [3-61. In most cases, however, the protein concentrations are considerably below those corresponding to an average cytopIasm [7J.
2. Experimental
A cationic or anionic membrane acts as a re- versibie electrode for small cations or anions when they are present in a protein solution. The activity coefficients of the salts in BSA solutions were determined by measuring the EMF of a three- compartment electrochemical cell of the type:
CM denotes a cation-exchange membrane (Nep- ton CR-61) and AM an anion-exchange mem- brane (Nepton AR-1 11) (Ionics, Watertown, MA, U.S.A.).
The EMF of this cell is a function of the mean ionic activities of the salts in solutions I and II according to the relation [8,9]:
(1)
where E denotes the EMF, F the Faraday con- stant, R the universal gas constant, T the absolute temperature and ai and Q+ the mean ionic activ- ity of the salt species in compartment I and II, respectively. t,“” and tzM designate the mean respective coion transference numbers in the an- ion- and cation-exchange membranes, respective- ly *.
Varying the mean activity of KX in compart- ment I, one obtains from eq. 1, for E = 0:
where the right-hand term represents the value of a’+ at E = 0. Considering that, by definition for (1,l) salts
a& = y&m (3)
we obtain for the stoichiometric mean ionic activ- ity coefficient, y+. of the salt KX in the protein
* For derivation and discussion of the validity of eq. 1 for the study presented here, see ref. 8.
M.D. Reboiras et al. /Actimty coefficients of salts in protein solutions
solution the following expression:
y* = (o;>,=, m
where m is the molality of the salt in compart- ment II. Experimentally, for each protein-salt composition, a linear relationship was obtained between E and In a *, which indicates according to eq. 1 that the caion transference number in the membranes does not change significantly within the range of salt concentrations of the reference solutions I used *.
All measurements were carried out at room temperature (23 + 1OC).
The EMF values were obtained by connecting the reference solutions through salt bridges (KC1 saturated in agar-agar) to a vibrating reed elec- trometer (input resistance 10” tin> and recorded with a potentiometric recorder, to an accuracy of +0.15 mV.
Reagent grade dried salts (Merck, p.A.) were used for the preparation of the reference salt solutions with bidistilled deionized water. Special attention was paid to the handling of KI solutions owing to the well-known fact that I- in solution undergoes auto-oxidation-reduction processes when exposed to sunlight. This process is more pronounced at higher concentration and, there- fore, these solutions were prepared immediately prior to use.
The BSA stock solutions were prepared from commercially available samples (Serumalbumin vom Rind, trocken, rein& Behring-Werke, Mar- burg, F.R.G.). Further details of the method, instruments and apparatus used and of prepara- tion of solutions are fully described elsewhere WI.
* No influence was observed from the quantity of protein on the slope of the titration lines. To the contrary, for constant protein concentration the slope of the titration lines de- creases with increasing salt concentration in protein-salt solutions as a result of the increase in the transference numbers of the coions in the membrane.
251
3. Results
3.1. Variation of the activity coefficients of the salts with the protein content in protein-salt solutions
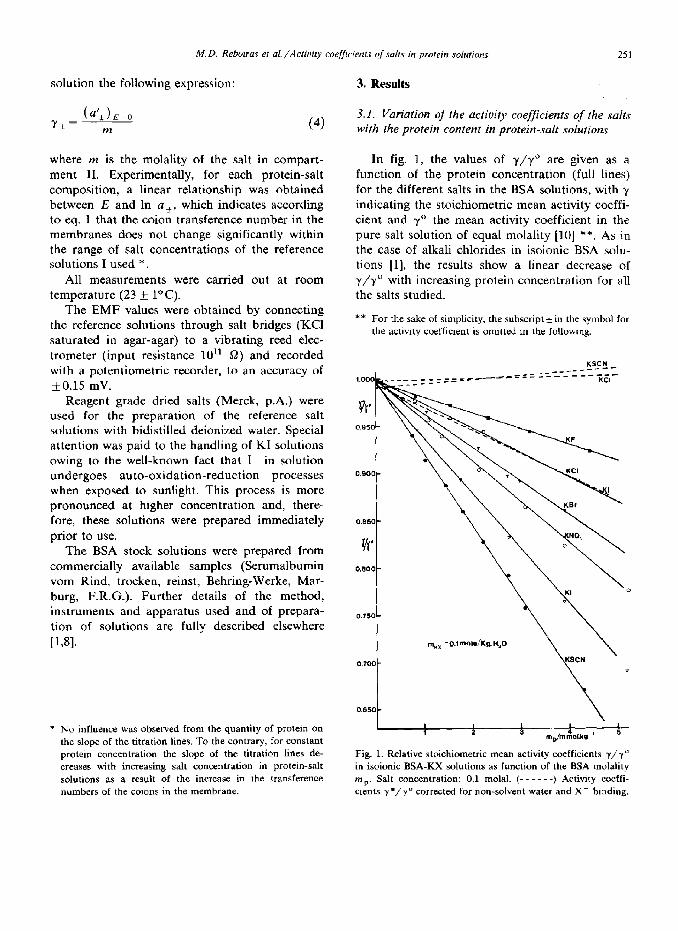
In fig. 1, the values of y/y” are given as a function of the protein concentration (full lines) for the different salts in the BSA solutions, with y indicating the stoichiometric mean activity coeffi- cient and y0 the mean activity coefficient in the pure salt solution of equal molality [lo] **. As in the case of alkali chlorides in isoionic BSA solu- tions [l], the results show a linear decrease of y/y” with increasing protein concentration for all the salts studied.
** For the sake of simplicity. the subscript + in the symbol for the activity coefficient is omitted m the following.
Fig. 1. Relative stoichiometric mean activity coefficients v/y” in isoionic BSA-KX solutions as function of the BSA molality mP. Salt concentration: 0.1 molal. (- - - - - -) Activity coeffi- cients Y*/Y” corrected for non-solvent water and X- binding.
Qualitatively similar results have been obtained for the same salts with hemoglobin [ll].
3.2. Variation of the activity coefficients of the salts with the salt content in protein-salt solutions
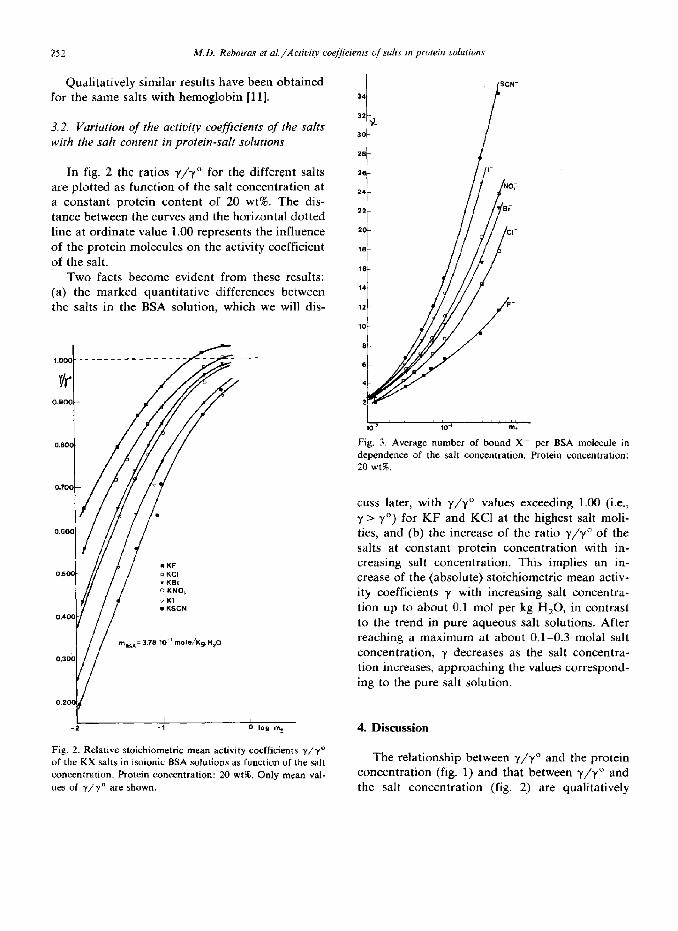
In fig. 2 the ratios y/y” for the different salts are plotted as function of the salt concentration at a constant protein content of 20 wt%. The dis- tance between the curves and the horizontal dotted line at ordinate value 1.00 represents the influence of the protein molecules on the activity coefficient of the salt.
Two facts become evident from these results: (a) the marked quantitative differences between the salts in the BSA solution, which we will dis-
L I I -2 -1 0 log. Ill,
Fig. 2. Relative stoichiometric mean activity coefficients y/y” of the KX salts in isoionic BSA solutions as function of the salt concentration. Protein concentration: 20 wt%. Only mean val- ues of y/y” are shown.
SCN- 34
1 ,1,181l 8,11881 10-2 10-’ m+
Fig. 3. Average number of bound X- per BSA molecule in dependence of the salt concentration. Protein concentration: 20 wt%.
cuss later, with y/y” values exceeding 1.00 (i.e., y > y”) for KF and KC1 at the highest salt moli- ties, and (b) the increase of the ratio y/y” of the salts at constant protein concentration with in- creasing salt concentration. This implies an in- crease of the (absolute) stoichiometric mean activ- ity coefficients y with increasing salt concentra- tion up to about 0.1 mol per kg H,O, in contrast to the trend in pure aqueous salt solutions. After reaching a maximum at about 0.1-0.3 molal salt concentration, y decreases as the salt concentra- tion increases, approaching the values correspond- ing to the pure salt solution.
4. Discussion
The relationship between y/y” and the protein concentration (fig. 1) and that between y/y” and the salt concentration (fig. 2) are qualitatively

M.D. Reborras et al. /Activity coefficients of salts m protein soluiions 253
similar for all the salts studied. The fact that the activity coefficients vary differently for each salt leads us to suppose that there exist specific inter- actions between the components of each salt and the protein molecule.
It is well established [5] that, under the pH conditions of the solutions studied, the BSA mole- cule binds a wide variety of anions, while interac- tions of this type are not observed for alkali ions. In a previous paper [l], we have shown that there are only slight differences in the interactions of the different alkali chlorides with BSA. In the following we assume that no specific interaction exists between BSA and Ki ions of the different salts studied *_
A first approach to the interpretation of the results, therefore, could be to assume that the decrease in the activity of the salts in presence of protein is caused only by the decrease in the concentration of ‘free’ anions due to the specific binding of anions to the protein molecules, as has been done by Scatchard et al. [12] for BSA salt solutions at low concentrations.
According to this assumption, the number of anions X- specifically bound to the BSA mole- cule, vx-, can be calculated [S] from the ratio of the stoichiometric mean activity coefficient in the protein-salt solution to that in pure salt solution,
* See footnote on p. 41 of ref. 1.
y/y’, using the equation
where m and m,, are the molal concentrations of salt and BSA, respectively. The results are given in column 1 of tables 1 and 2. For comparison, in column 3 of the tables the v values for Cl-, I- and SCN- are shown, obtained by use of the equation (see e.g. ref. 2):
v=C n,K,Oy exp( -2wv)(m - vmp)
i 1 -i- Kpy exp( -2wv)( WI - YMp) (6)
with the n, and K,’ values representing the num- ber of different binding sites and their affinity constants, respectively, as determined by Scatchard et al. [13]. m and mp are the salt and protein molal concentrations and y the activity coefficient of the respective ion, assumed here to be equal to that of the pure salt solution of equal salt con- centration. (m - vm,) represents the molality of free anions in solution.
The factor exp( - 2wv) accounts for the electro- static interactions between the sites. The electro- static factor w can be calculated from the electro- static free energy of a single protein molecule. Here, we calculated w according to Scatchard’s theory [13] with the assumptions that the albumin molecule can be represented by a sphere with a radius of b = 30.0 x lo-’ cm, and that the colli-
Table 1
Average number of ions bound to the BSA molecule as function of salt concentration
Conditions:constant protein concentration for each salt (around 20 wt%). The exact BSA concentrations (in mmol per kg H,O) were: 3.77 for KF, 3.78 for KCI, 3.80 for KBr and KI, 3.76 for KNO, and 3.74 for KSCN. (1) P, calculated from the stoichiometric mean activity coefficients. (2) Y, calculated after taking into account the non-solvent water (19.9 kg per mol BSA, corresponding to 0.3 8 per g dry BSA). Each value is the mean of two measurements. (3) Y, calculated from e.q. 6 using the values of n, and KY of Scatchard et al. [13].
m F_ Br- NO, Cl_ I- SCN - ~ ~
1 2 1 2 1 2 1 2 3 1 2 3 1 2 3
0.01 2 2 3 3 2 2 2 2 2 3 3 2 3 3 3 0.03 3 4 5 5 5 6 4 4 4 6 6 6 I 7 7 0.05 3 5 6 7 6 7 4 5 5 8 9 8 Y 10 11 0.07 4 6 7 8 7 9 5 7 7 10 11 10 11 12 13 0.1 3 7 7 10 8 11 6 9 8 11 13 11 13 15 15 0.3 -2 9 6 17 9 19 3 14 12 16 25 17 19 28 21 0.5 - 9 12 4 22 5 24 -1 18 14 22 3X 19 18 34 23
254 M, D. Kebviws et al. /Actiurly coefficiewls of salts in protein solurions
Table 2
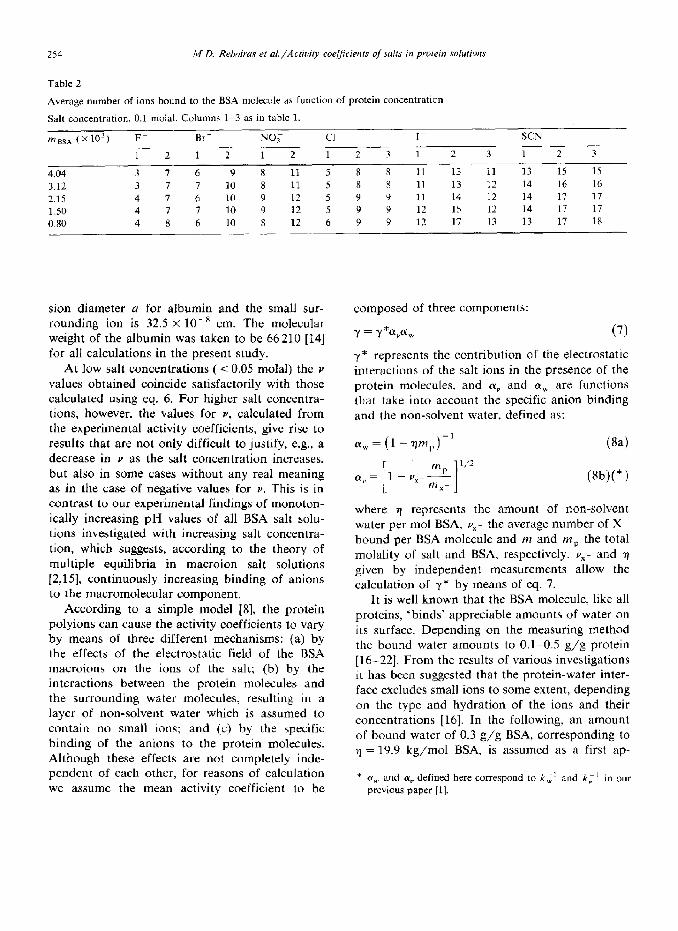
Average numher of ions bound to the BSA molecule as function of protein concentration
Salt concentration, 0.1 molal. Columns 1-3 as in table 1.
mBSA (~10~) F- Br- NO; Cl I_ SCN -
1 2 1 2 1 2 1 2 3 1 2 3 1 2 3
4.04 3 7 6 9 8 11 5 8 8 11 13 11 13 15 15
3.12 3 7 7 IO 8 11 5 8 R 11 13 12 14 16 16
2.15 4 7 6 10 Y 12 5 9 9 11 14 12 14 17 17 1 so 4 7 I 10 9 12 5 9 9 12 I5 12 14 17 17
0.80 4 8 6 10 8 12 6 9 9 12 17 13 13 17 1x
sion diameter a for albumin and the small sur- rounding ion is 32.5 x lO_' cm. The molecular weight of the albumin was taken to be 66210 [14] for all calculations in the present study.
At low salt concentrations ( c 0.05 molal) the Y values obtained coincide satisfactorily with those calculated using eq. 6. For higher salt concentra- tions, however, the values for Y, calculated from the experimental activity coefficients, give rise to results that are not only difficult to justify, e.g., a decrease in v as the salt concentration increases, but also in some cases without any real meaning as in the case of negative values for Y. This is in contrast to our experimental findings of monoton- ically increasing pH values of all BSA salt solu- tions investigated with increasing salt concentra- tion, which suggests, according to the theory of multiple equilibria in macroion salt solutions [2,15], continuously increasing binding of anions to the macromolecular component.
According to a simple model [S], the protein polyions can cause the activity coefficients to vary by means of three different mechanisms: (a) by the effects of the electrostatic field of the RSA macroions on the ions of the salt; (b) by the interactions between the protein molecules and the surrounding water molecules, resulting in a layer of non-solvent water which is assumed to contain no small ions; and (c) by the specific binding of the anions to the protein molecules. Although these effects are not completely inde- pendent of each other, for reasons of calculation we assume the mean activity coefficient to be
composed of three components:
Y = Y *VW (7)
Y* represents the contribution of the electrostatic interactions of the salt ions in the presence of the protein molecules, and (Y, and CY, are functions that take into account the specific anion binding and the non-solvent water, defined as:
WI(*)
where 4 represents the amount of non-solvent water per mol BSA, vxm the average number of XP hound per BSA molecule and m and mp the total molality of salt and BSA, respectively. v,- and q given by independent measurements allow the calculation of y * by means of eq. 7.
It is well known that the BSA molecule, like all proteins, ‘binds’ appreciable amounts of water on its surface. Depending on the measuring method the hound water amounts to O.lLO.5 g/g protein [16-221. From the results of various investigations it has been suggested that the protein-water inter- face excludes small ions to some extent, depending on the type and hydration of the ions and their concentrations [16]. In the following, an amount of hound water of 0.3 g/g BSA, corresponding to 77 = 19.9 kg/mol BSA, is assumed as a first ap-
* a,, and a, defined here correspond to k;' and &,;I in our
previous paper [l].

M.D. Keboiras ef al. /Activity coefficients o/ ml& in protein solutrons 255
proximation to act as non-solvent water for the small ions, independent of the ion type and con- centration.
We have shown in a previous paper [l] that the electrostatic mean activity coefficient y* of alkali chlorides in BSA solutions changes only very little up to 21 wt% BSA, at a constant salt concentra- tion of 0.1 molal. Similarly, the calculation of y*/y’ for Cl-, I- and SCN- carried out by means of eq. 6 shows very little change in the range of BSA concentrations studied, as may be seen in fig. 1 where the ratio y*/y’ is represented by the dotted lines. y*/y” for KC1 and KSCN is practically constant throughout the range of pro- tein concentrations studied, whereas the values for KI decrease slightly with increasing protein con- centration (maximum decrease: 17% at 5 x 1O-3 molal BSA). No evident reason was found for the discrepancy between the results for I- and those for Cll and SCN-, although the most plausible is that of the error in the v values used.
Similar theoretical and experimental results have been obtained [8,23,24] for analogous sys- tems.
Thus, the relatively insignificant influence of the electrostatic interactions with the BSA poly- ions on the chemical potential of those salts, where the amount of specificaly bound anions is known from independent measurements, leads itself to the tentative interpretation of the protein effect on the activity coefficients of the salts in terms of specific anion binding and non-solvent water alone for all the salts investigated. By taking y* = y” in eq. 7 and combining with eq. 8, we arrive at the relation:
In columns 2 of tables 1 and 2 the values calcu- lated from eq. 9 are shown. For all the salts studied, now the number of ions bound to the protein molecule decreases slightly as the con- centration of BSA rises at constant salt concentra- tion. When the BSA concentration is maintained constant, Y increases as the salt concentration increases; the meaningless negative v values (i.e., y/y” > 1 in fig. 2) for F- and Cl- calculated from
eq. 5, have disappeared by the application of the non-solvent water correction. As can be observed, the experimental values for v coincide satisfacto- rily at low electrolyte concentrations with those predicted by Scatchard et al. from eq. 6, within the range of error affecting both groups of results. With increasing salt concentration (> 0.1 molal), the deviations become pronounced. The agree- ment at low concentrations suggests that, with low concentration of salt, the decrease in the activity coefficient must be almost exclusively brought about by the specific binding between the anions and the protein, in accordance with the interpreta- tion made by Scatchard et al. As the salt con- centration increases, other factors such as the elec- trostatic effect of the protein on the ions must come into play as has been shown by RHmisch [24], who indicated a 5% decrease in the activity coefficient because of the electrostatic influence of the protein where the salt concentration exceeds 0.07 molal, for NaCl in BSA solutions. At this concentration 7 Cl- are bound per protein mole- cule. It is around this value of salt concentration that the discrepancies in v between our results and those of Scatchard’s appear. It should be noted, however, that the same effect could be caused either partially or totally by a decrease in non- solvent water with an increased salt concentra- tion, in contrast to the constant value assumed above.
When the values of v calculated by taking into account the constant amount of non-solvent water bound to the protein are plotted as a function of salt concentration (fig. 3), a series of curves is obtained with a similar profile for each salt. Throughout the range of salt concentrations studied the following series can be established for the degree of binding of anions to the protein: F- < Cl- < Br- -z NO; i I- < SCN -. The same sequence is obtained when we consider the in- fluence of BSA concentration on the activity coef- ficients of the salts shown by the different slopes of the straight lines in fig. 1, and similar results have also been obtained in the study of hemo- globin with the same anions [ll].
The v values calculated according to eq. 9 are very sensitive to the amount of non-solvent water assumed. The increase of v due to the term a, in

256 M.D. Reboiros et al./Actiuity coefficients qf salts in protein solutions
eq. 9 is approximately proportional to q under the experimental conditions given here. A variation of the amount of non-solvent water of +0.1 g/g BSA changes by t_ 1.1 and )6.7 for F-, by kO.6 and k5.2 for SCN-, at m = 0.1 and 0.5, respec- tively, and at 20 wt% BSA. At least at the high salt concentrations, therefore, the v values calculated here are subject to ambiguity resulting from the uncertainty of the amount of non-solvent water assumed.
On the basis of the results presented here, it appears that the combination of ion-selective membranes can be used to determine the activity of a large number of inorganic salts in con- centrated protein solutions. It can also be stated that such determinations can be applied to calcu- late the numbers of anions bound to the protein, if the amount of non-solvent water is known from independent measurements with sufficient accu- racy to allow for a precise correction of the non- solvent water which must take into account the influence of both concentration and ion type.
5. Conclusions
Our results show that the stoichiometric mean activity coefficients of the KX salts in the pres- ence of BSA are lower than those in pure salt solutions of equal molality, except for some values for KF and KC1 at high salt concentrations only. This departure depends significantly on the nature of the anion of the salt, whereas there is only a small cation-specific effect in similar BSA salt solutions [l]. Taking into account the effect of the non-solvent water bound to the BSA molecule, it is possible to calculate the number of ions specifi- cally bound to the protein molecule. With increas- ing salt concentration, the activity coefficients of the salt in presence of BSA rise, in contrast to the trend of the activity coefficients of the pure salts in aqueous solutions. This behaviour is similar to that exhibited by salts inside ion-exchanger mem- branes or in poIyelectrolyte solutions, and, purely from a phenomenological point of view, these results are consistent with the concept of using biological ion-exchange models I251 for the inter- pretation of some properties of the cellular cyto-
plasm. One should, however, take into account, when making possible comparisons between the behaviour of salts with synthetic ion-exchangers and in solutions of biological polyions, that the equivalent concentration of fixed ions within the polyelectrolyte matrix, in relation to the amount of water present in the pores, can reach extremely high values (higher than 3 equiv./kg H,O), com- pared to the concentration of the solutions under study. In both cases, however, the activity coeffi- cient decreases as the salt concentration becomes smaller, which indicates that the lower the con- centration the greater is the electrostatic interac- tion between the ions and their macromolecular surroundings.
By and large, the results of our studies pre- sented here and in our previous paper [l] show that the stoichiometric mean activity coefficients of (I,]) salts in isoionic BSA solutions are lowerkd no more than 30-4058 with respect to pure salt solutions at protein and salt concentrations which correspond to those of the cytoplasm of the living cell (about 20 wt% and 0.1 molal, respectively).
Most of this effect is attributed to the specific binding of anions to the protein molecules, to- gether with the assumption of a certain amount of water bound at the surface of the protein mole- cules and acting as non-solvent water which ex- cludes the small ions. Compared to this, the elec- trostatic effect of the protein polyions on the chemical potentials of the salts, as measured by the ‘electrostatic’ mean activity coefficients, seems to be very small (only a few percent).
Acknowledgement
We thank the CAICYT (grant no. 0961/81) for travelling expenses for M.D.R.
References
1 M.D. Reboiras, H. Pfister and H. Pauly, Biophys. Chem. 9 (lY78) 37.
2 J. Steinhard and J.A. Reynolds, Multiple equilibria in pro- teins (Academic Press, New York, 1969).
3 L.W. Nichol and D.J. W&or, Protein-protein interactions, eds. C. Frieden and L.W. Nichol (John Wiley, New York, 1981) p. 1.

M.D. Rcboiras et aI./Actiuity coefficients of .solrs in protein solutinns 257
4 G. Schwartz, Btophys. Chem. 6 (1977) 65. 5 W.E. Briggs, Biophys. Chem. 18 (1983) 67. 6 CL. Ford, D.J. Winzor, L.W. Nichol and M.J. Sculley,
Biophys. Chem. 18 (1983) 1. 7 A.C. Giese, Cell physiology, 4th edn. (W.B. Saunders,
Philadelphia, 1973). 8 H.Pfister, Thesis, Universitlt Erlangen-Niirnberg. F.R.G.
(1971). 9 K. Sollner, Arm. N.Y. Acad. Sci. 148 (1968) 154,
10 R. Parsons, Handbook of electrochemical constants (But- terworths, London, 1959).
11 W. Schmid, Thesis, Universitat Edangen-Niirnberg, F.R.F. (1976).
12 G. Scatchard, I.H. Scheinberg and S.H. Armstrong, J. Am. Chem. Sot. 72 (1950) 535 and 540.
13 G. Scatchard, J.S. Coleman and A.L. Shen, J. Am. Chem. sot. 79 (1957) 12.
14 T. Peters, Jr, Clin. Chem. 23 (1977) 5. 15 C. Tanford, Physical chemistry of macromolecules (John
Wiley, New York, 1961).
16 D. England, in: Water A comprehensive treatise, vol. 4, ed. F. Francks (Plenum Press, New York, 1975) p. 305.
17 GM. Mrevlishvili and P.L. Privalov, in: Water in biological systems, ed. L.P. Kayushin (Consultants Bureau, New York, 1969) p. 63.
18 G. Has1 and H. Pauly, Biophysik 7 (1971) 283. 19 D.J. Cox and V.N. Schumaker, J. Am. Chem. Sot. 83
(1961) 2453. 20 H.B. Bull and K. Breese, Arch. Biochem. Biophys. 128
(1968) 488. 21 G.N. Ling, in: Water and aqueous solutions. Structure,
thermodynamics and transport processes, ed. R.A. Home (Wiley-Interscience, New York, 1972) p. 663.
22 I.D. Kuntz, Jr and W. Kauzman, Adv. Protein Chem 28 (1974) 239.
23 H. Pfister and H. Pauly, J. Polym. Sci. C39 (1972) 179. 24 D. Ram&h, Thesis, UniversitBt Erlangen-Niirnberg, F.R.G.
(1976). 25 R. Damadian, Ann. N.Y. Acad. Sci. 204 (1973) 211.





























