Embed Size (px)
Citation preview

Actualidades sobre
adaptación, producción,reproducción y mejora genéticaen camélidos
Autores:Edgar Quispe PeñaJoaquin Pablo MuellerJaime Ruiz BejarLeopoldo Alfonso RuizGustavo A. Gutiérrez

Editado por Edgar Carlos Quispe Peña
Tiraje: 500 ejemplares.
Prohibida la reproducción total o parcial de este libro, por cualquier medio, sin autorización por parte de los autores.
Fotografías de portada: Medición del diámetro de fibra en Air Flow, parto y embriones partenogénicos de alpaca.
Fotografías de contraportada: Esquila mecánica para la obtención de fibra, ejemplar ganador de feria, empadre controlado.
Corrección de estilo:Edgar Carlos Quispe Peña
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
Hecho el Deposito Legal en la Biblioteca Nacional del Perú Nº 2008-10814.EDGAR CARLOS QUISPE PEÑADomicilio:Felipe de Zela Nº 796 Dpto. 302 Urb. Los Ficus. Santa Anita. Lima. Perú.UNIVERSIDAD NACIONAL DE HUANCAVELICA. Ciudad Universitaria Paturpampa s/n. Huancavelica. Perú Primera Edición.2008. octubre. Huancayo - Perú.Impreso en Edición Gráfica Industrial E.I.R.L. Huancayo - Perú.

ÍNDICE
PROLOGO
PRESENTACIÓN
INTRODUCCIÓN
ADAPTACIONES HEMATOLÓGICAS DE LOS CAMÉLIDOS SUDAMERICANOS QUE VIVEN EN ZONAS DE ELEVADAS ALTITUDESPor: Quispe, E. C.
CARACTERÍSTICAS DE LA FIBRA DE ALPACA HUACAYA PRODUCIDA EN LA REGIÓN ALTO ANDINA DE HUANCAVELICA, PERÚ
1 2 2 3 1Por: Montes, M. ; Quicaño, I. ; Quispe, R. ; Quispe E. C. ; Alfonso, L.
AVANCES EN BIOTECNOLOGIA REPRODUCTIVA APLICADA EN LA HEMBRA DE LOS CAMELIDOS SUDAMERICANOSPor: Ruiz, J. A.
REVISIÓN DE LA ESTIMACIÓN DE LOS PARÁMETROS GENÉTICOS EN ALPACASPor: Gutiérrez, G. A.
BASES PARA ESTABLECER UN PROGRAMA DE MEJORA DE ALPACAS EN LA REGIÓN ALTOANDINA DE HUANCAVELICA-PERU
1 2 3 1Por: Quispe, E. C. ; Alfonso, L. ; Flores, A. ; Guillén, H.
ESTRATEGIAS PARA EL MEJORAMIENTO DE CAMÉLIDOS SUDAMERICANOSMueller, J. P.
5
5
9
11
37
49
83
93
113
Pag.


PRÓLOGO
La información científica o técnica relacionado con los camélidos sudamericanos y en especial a la Alpaca fue muy escasa hasta mediados de los 70's del siglo pasado comenzando cuando la Universidad Nacional Agraria La Molina inició a dictar el curso de Camélidos Sudamericanos. Un fuerte impulso se obtuvo a fines de los 80's y a principios de los 90's del siglo pasado, en especial cuando se crea el Consejo Nacional de Camélidos Sudamericanos en 1992. Todo ello debido a la fuerte demanda de fibra fina animal que es propia de los camélidos sudamericanos frente a la muy baja cotización de la lana de los ovinos lo que hizo cambiar a Nueva Zelandia y Australia principalmente su explotación ovejera por la alpacuna. Por ello en el libro se muestra la rica y variada bibliografía de las alpacas, relacionadas con la adaptación al medio que se explotan los efectos del medio ambiente y del sexo sobre la característica de la fibra de alpaca Huacaya, los avances en biotecnología reproductiva aplicada en las hembras de los camélidos sudamericanos, la estimación de los parámetros genéticos en alpacas, las bases para establecer un programa de mejora de alpacas en Huancavelica y estrategias para el mejoramiento de camélidos sudamericanos, que nos muestra el progreso de las investigaciones ya sea por su adaptación fisiológica, morfológica así como el efecto de la preadaptación y en especial un mejor aprovechamiento del oxigeno a nivel tisular lo cual impide la hipoxia en las zonas altoandinas.
Se reafirma el efecto del sexo, edad y localidad en el diámetro de la fibra y se desarrolla una completa revisión de las investigación de la biología aplicada en los camélidos sudamericanos desde la recuperación de complejos ovocito – cúmulos hasta la transferencia de embriones pasando por la superovulación, señalando las ventajas y desventajas de los diferentes protocolos, basado en una extensa y fundamental bibliografía, es sin duda un texto para los investigadores en reproducción muy bien compendiada. Se presenta una revisión de los parámetros genéticos de índice de herencia de los métodos para obtenerle de principales características de peso vivo, peso vellón (sucio y limpio), diámetro de fibra y longitud de mecha señalando finalmente que los parámetros genéticos reportados requieren de mayor precisión. Las bases para establecer un programa de mejora de alpaca, se encuentra bien planteado y de la respuesta de la selección esperada. Se demuestra que en la Región Huancavelica existe un buen potencial genético de las dos características más importante en la explotación de la alpaca: finura de la fibra y peso del vellón sucio. Estas características deben ser estudiadas en las diferentes localidades (17) pero explotadas en pasturas excelentes, con buen manejo sanitario y reproductivo que contribuyan a elevar el estándar de vida de los productores alpaqueros.
Finalmente en las estrategias para una mejora genética, se recoge las experiencias identificando los elementos del proyecto y un análisis de los efectuados en el Perú para terminar en estrategias generales en la que se contempla la generación y consolidación de una estructura genética, la importancia del análisis de la fibra, los servicios de evaluación genética e incidiendo finalmente en la participación de los productores en la planificación y ejecución de los proyectos y la formación de recursos humanos.

En resumen el libro es una fuente de información actualizada y valiosa que conlleva a ser un libro necesario para todos los que desean contribuir a la mejora de la explotación de la alpaca en cualquier parte del mundo y en especial en le Perú.
Deseo felicitar a la Universidad Nacional de Huancavelica que viene realizando un esfuerzo sostenido en la mejora de nuestra especie símbolo nacional y al Ing. M. Sc Edgar Quispe Peña el principal promotor y difusor de los avances científicos y tecnológicos.
Alfonso Flores Ph. D Profesor Emerito UNALM

PRESENTACIÓN
Es para mi muy satisfactorio referirme al libro: “Actualidades sobre adaptación, producción reproducción y mejora genética en camélidos”, para señalar que dentro del ámbito del conocimiento de estos temas en nuestro país falta profundizar y sobre todo falta tener referencias bibliográficas que ayuden a formular planes de desarrollo en camélidos sudamericanos en lugares donde aun la tecnología no ha tenido un papel importante en desarrollo de esta especie tan legendaria y tan peruana; destaca especialmente las revisiones concienzudas que se han hecho de aspectos de biotecnología reproductiva que se han desarrollado tanto en nuestro país así como en otros ámbitos a nivel mundial, donde la investigación fundamentalmente ha tenido mucho que ver en el avance de las tecnologías de avanzada que se usan en animales, tales como la inseminación artificial, transferencia de embriones, fertilización in Vitro y otras mas avanzadas que se están ensayando con la intención de lograr protocolos que sean de difusión mayor y que puedan adecuarse una vez que hayan sido validadas y comprobadas. La reproducción va muy de la mano con la mejora genética, para una vez ubicados los mejores animales a través de los estudios de parámetros productivos, puedan estos ser replicados intensivamente, considerando que el período de vida productiva es corto y también lo difícil que es reproducirlos por métodos naturales.
En segundo lugar quiero referirme a los autores del libro quienes son profesionales destacados y jóvenes, entre ellos peruanos de reconocida trayectoria en el campo de la investigación en las áreas de biotecnología reproductiva y mejoramiento genético de los camélidos sudamericanos; ellos con seguridad seguirán avanzando en estas ciencias pecuarias para en el futuro ponerlas a disposición de los ganaderos alpaqueros que como conocemos es parte de la población necesitada de recursos del país; no quiero dejar de mencionar a los autores extranjeros que por su experiencia y dedicación están poniendo la cuota de calidad en investigaciones científicas, dando un nivel adecuado a esta publicación. A todos ellos les deseo éxitos en la producción de futuros trabajos bibliográficos.
Importante igualmente es recomendar la lectura de esta obra que permitirá a los lectores empaparse de información valiosa en el ámbito de la crianza de los camélidos sudamericanos.
Ing. Próspero Cabrera Villanueva Jefe del Banco Nacional de Semen Jefe del Programa de mejora animal - UNALM


INTRODUCCIÓN
Teniendo Perú la mayor cantidad de camélidos sudamericanos (especialmente en lo que se refiere a alpacas y vicuñas) a nivel mundial, no resulta raro que la producción de alpacas, vicuñas y llamas se encuentren considerados como especies importantes de los ejes de desarrollo pecuario, los cuales se vislumbran en los diferentes planes de desarrollos (locales, regionales y nacionales). Por tanto diversas instituciones públicas y privadas, vienen realizando enormes esfuerzos a través de la investigación, con la cual ya se viene logrando la obtención de una tecnología apropiada que permita mejorar la producción de carne, fibra, piel, trabajo, entre otros, de nuestros camélidos. Estos resultados también se deben difundirlos a fin que puedan ser adaptadas a los diferentes sistemas de producción de estas especies animales. La crianza de los camélidos sudamericanos en la Región Huancavelica constituye una actividad económica importante del poblador alto andino, la cual se desarrolla entre los 3,000 a 5,000 m.s.n.m.; las familias están organizadas en comunidades de pastores, bajo un sistema de crianza de Rebaño Mixto Familiar (alpacas, llamas, ovinos). Estos rebaños se caracterizan por carecer de sistemas de crianza adecuados, con escasos criterios de mejoramiento genético, basados exclusivamente como fuente alimenticia en pastizales alto andinos de baja soportabilidad y con falta de cuidados sanitarios. Se estima la existencia de más de 60 comunidades alpaqueras, que agrupan a 3,300 familias aproximadamente, que se encuentran distribuidas en cuatro provincias: Angaraes, Huaytará, Castrovirreyna y Huancavelica.
En Huancavelica e Ica existen alrededor de 330,500 alpacas (23,500 Suri y 307,000 Huacaya), los cuales constituyen el 11.4% de la población nacional. Los vellones obtenidos de las alpacas en las comunidades alpaqueras de Huancavelica son de baja producción y calidad. A nivel de comunidades se estima una producción promedio bianual de 2.2 kg mientras que bajo una crianza medianamente tecnificada se obtiene una producción anual de 2.2 a 2.3 kg. Con respecto a la finura a nivel nacional, el 50% de la producción está dado por fibra gruesa (34 micras), el 40% por fibra de tipo Fleece (26.5 micras) y sólo el 8% está conformado por fibra fina (Baby, de 22.5 micras), lo cual vislumbra el enorme déficit en calidad; también muchos de los vellones son canosos, pintados y canosos-pintados, y en muchos vellones se encuentran gran heterogeneidad en la estructura, pues muchas fibras que lo conforman son de tipo medulado de forma continua o discontinua, lo cual desmerece la calidad del vellón.
En función a las consideraciones anteriores se puede apreciar que existe una problemática en la producción del sector de Camélidos Sudamericanos Domésticos, siendo principalmente tres las causas directas que generan este problema. a) Deterioro de la calidad genética, la cual tiene como causas indirectas, la falta de identificación adecuada de reproductores, inadecuados sistema de reproducción, inexistencia de parámetros genéticos y fenotípicos, desconocimiento del mérito genético e inadecuados registros genealógicos y productivos; b) Inadecuado manejo medioambiental, que tiene como causas indirectas la alta incidencia de enfermedades, deficiente alimentación y

baja tecnología en la producción alpaquera; y c) Unidades productivas desintegradas, debido a una débil organización social y empresarial. Estas tres causas tienen en común una débil y escasa capacitación técnica, empresarial y social de los productores alpaqueros, técnicos e incluso profesionales, los cuales se sitúan como causas indirectas. Las consecuencias que tiene el problema situacional anteriormente identificado se reflejan en la baja productividad de los camélidos, baja competitividad de los productos obtenidos, los cuales redundan en la obtención de bajos ingresos económicos, con una producción de subsistencia, afectando con ello la calidad de vida de los productores.
Los sistemas de producción que se desarrollan en diversas comunidades, relacionados con la crianza de camélidos son generalmente tradicionales, por lo tanto, se hace imprescindible la aplicación de tecnologías apropiadas y conveniente a fin de obtener el cambio a un sistema semitecnificado, que se vería reflejado en los centros de producción que asimilen las adopciones tecnológicas, lo cual también tendrá repercusión en los centros de producción de su entorno.
Unido a la problemática productiva del sector de Camélidos Sudamericanos Domésticos se encuentra también la focalización institucional expresada en la duplicación de acciones y competencias operativas en desmedro de nuestros limitados recursos humanos y financieros. Por un lado al no tener definido el ámbito, naturaleza y alcances de cada institución, y por la falta de una real y efectiva concertación con los actores involucrados en este quehacer, se pierde la gran oportunidad de socializar estas valiosas experiencias. Estos son factores que inciden mayormente en la negativa desarticulación entre el sector productivo y el sector industrial, componentes estratégicos para una real definición en la cadena productiva hacia el mercado, es decir si deseamos la optimización del valor agregado y la generación de un mayor campo ocupacional, este es un aspecto que se espera superar a través de la concertación institucional y organizaciones de productores.
Los aportes en conocimientos, experiencias, recursos humanos y económicos, todos ellos, unidos a un trabajo en donde todos los sectores en su conjunto participen efectiva y coordinadamente, bajo una responsabilidad institucional que represente un reto compartido, quienes al margen de su propia identidad, conjuntamente estén en condiciones de afrontar esta responsabilidad histórica bajo una única concepción estratégica, permitirán conducir a la soluciones de la problemática enmarcada.
En ese sentido la UNH, a través del Programa de Camélidos Sudamericanos (PROCASUD), perteneciente a la Escuela Profesional de Zootecnia de la Facultad de Ciencias de Ingeniería, pone a su alcance este texto universitario: “Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos sudamericanos” en donde diversos especialistas de cada área, de talla nacional e internacional, enfocan temas a nivel del estado de arte actual de nuestros camélidos sudamericanos, a fin de difundir los avances científicos que se vienen logrando en temas que constituyen puntos críticos, pero que se vislumbran prontas soluciones.
Edgar Quispe Peña Autor

ADAPTACIONES HEMATOLÓGICAS DE LOS CAMÉLIDOS SUDAMERICANOS QUE VIVEN EN ZONAS
DE ELEVADAS ALTITUDES
Quispe, E. C.
PROCASUD - Universidad Nacional de HuancavelicaE-mail: [email protected]
ABSTRACT
The South American camelidaes (SAC) not always lived in high altitude areas, because diverse researches have demonstrated that at the pre-Columbian epoch existed great amount of camelidaes that lived in the coast. At the present time the raising of alpacas, llamas and vicunas is mainly carried out at geographic land with high altitude, associated to the diminution of concentration of oxygen, temperature and humidity, as well as to the increase of cosmic radiations, to which the SAC have adapted using biological, morphologic and behavior mechanisms, to which they have a great homeostatic amount of mechanism which they allow his life in that unfavorable atmosphere. This review mainly focus in the hypobaric hypoxia like stress factor and the morphologic and physiological adaptations (haematological and circulatory) that the organism of the SAC has had to modify to confront the diminished oxygen concentration. In relation to the morphologic aspects the adaptability is in the small size and discharge amount of the red blood cells (RBC), that next to the smaller haemoglobin (Hg) level, among others, cause that the SAC have the blood with the low viscosity producing at level of vessels a smaller constrictor effect and greater relaxation. When the kinetic aspect is considered, is very important the greater affinity of the Hg to O2 which together with the elevated density of capillaries (with shorter distances between the capillaries and the cells) would elicit the interchange of gases at alveolar and capillary level. In SAC is not observed pulmonary arterial hypertension due probably to high concentrations of P Substance, which would behave like a powerful vasodilator, which jointly with the increase of the activity of nitric oxide, the presor effect of the hipóxica condition would diminish.
KEY WORDS: SAC, adaptability, hypoxia, red blood cells, haemoglobin, oxygen.
INTRODUCCIÓN
La familia Camelidae, que engloba a los camélidos del viejo y el nuevo mundo (tribus Camelini y Lamini), se originaron en América del Norte durante el Pliocenio, aproximadamente 40 a 50 millones de años atrás. Hace unos 3 millones de años, la tribu Camelini migra al Asia y los Lamini hacia América del Sur. Los representantes más antiguos de la tribu Lamini son los fósiles del género Pliauchenia encontrados en los llanos de América del Norte y con una antigüedad de 9 a 11 millones de años (Wheeler, 1991). El género Pliauchenia dio origen a los Hemiauchenia, los cuales se desplazaron hacia América del Sur al final del Plioceno o comienzos del Pleistoceno, hace unos 3 millones de años. Se estima que los Hemiauchenia dieron origen a los géneros Lama y Vicugna un millón de años más tarde, desapareciendo del continente norteamericano hace unos 10 mil años, quedando sólo las especies Lama y Vicugna, como únicos sobrevivientes de la tribu Lamini.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
11

La denominación de Camélidos sudamericanos (CS), Familia Camelidae, tribu Lamini, géneros Lama y Vicugna, engloba a dos especies domésticas la alpaca y la llama, y dos silvestres la vicuña y el guanaco.
La crianza de alpacas y llamas se constituye en una de las pocas actividades ganaderas que se pueden llevar a cabo en terrenos geográficos ubicados en las grandes alturas lo que por sí sólo constituye un motivo más que justificado para estudiarlas, independientemente del conocimiento requerido por el progresivo desarrollo de esta ganadería en otras regiones de Perú e incluso de la incursión, en diferentes regiones, de cada vez más ejemplares como animales de compañía (Raggi, 2000).
Por otro lado, en Sudamérica la diversidad de mamíferos silvestres está representada principalmente por roedores y murciélagos, siendo los camélidos silvestres, vicuñas (Vicugna vicugna) y guanacos (Lama guanicoe), los ungulados más importantes de las extensas estepas subtropicales del continente. A diferencia de los guanacos que utilizan una gran variedad de ambientes desde el nivel del mar hasta los 4.000 metros de altura, las vicuñas están restringidas a los sistemas puneños y altoandinos entre los 3.000 y 5.000 metros de altura. Los guanacos tienen estructuras sociales más plásticas y variables como consecuencia de sus adaptaciones a diversos hábitats mientras que las vicuñas tienen una organización social relativamente constante. (Vilá, 1999)
La población actual de camélidos en Sudamérica entre alpacas llamas, vicuñas y guanacos se estiman en aproximadamente 8 millones y su distribución por especie se observa en el cuadro 1. En cuanto se refiere a la población de estos animales en Perú, CONACS reporta estimados de: 1'154,848 llamas, 3'036,181 alpacas, 143,526 vicuñas y 3,810 guanacos, haciendo un total de 4'338,365 animales.
Cuadro Nº 1. Población sudamericana de camélidos sudamericanos
Tomado de: CONACS (1999)
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
12
Especie Poblaciónestimada
Alpaca 3 483 200Llama 3 768 670Vicuña 230 585Guanaco 607 000TOTAL 8 089 455

Los llamados ecosistemas montañosos de Sudamérica, - lugar donde se desarrollan la cría y explotación de los camélidos sudamericanos - tienen como característica común la elevación sobre el nivel del mar. Su principal variable climática es la disminución de la concentración de oxígeno atmosférico pero, además, como la temperatura disminuye en función de la altura, aparece el frío como otras de sus características. Asimismo, las radiaciones cósmicas, ultravioleta y otras aumentan con la altura, y la humedad atmosférica disminuye. Es importante recordar entonces, que cuando se habla de adaptación a la altura debe tenerse presente que los humanos y animales deben desarrollar mecanismos adaptativos a múltiples variables climáticas. Contra el frío y las radiaciones, es posible poner en juego, además de mecanismos biológicos, la adaptación por comportamiento que se adquiere culturalmente en humanos e instintivamente en animales. Así es posible desarrollar el vestido adecuado y protegerse del sol, o acurrucarse lo más posible para evitar la pérdida de temperatura. En el caso de la variable oxígeno, ésta adaptación es básicamente biológica aunque pueda darse también por comportamiento, adecuando las tareas físicas a las necesidades de conservar energía. Aparte de la altura, juega un papel importante la latitud del ecosistema montañoso. Los Andes habitados pueden ser considerados como montañas tropicales por su cercanía con la línea del Ecuador, lo que produce altas temperaturas diurnas, quedando la noche expuesta al enfriamiento natural producido por la mayor distancia del centro del globo terrestre y a las escasas propiedades térmicas de una atmósfera más diáfana. (Raggi, 2000).
Sólo una visión ecológica integral permite interpretar los mecanismos biológicos y culturales que condicionan la aclimatación de la flora, la fauna y el ser humano a la altura (Monge y Salinas, 1986).
A medida que se asciende, el aire se hace mucho más seco y las radiaciones solares más intensas, por lo que los habitantes de altura deben ser capaces de resistir la desecación y la fuerte insolación. Como si lo anterior no fuese ya una fuerte limitante hay que agregar el hecho de que en las capas más altas de la atmósfera hay una menor cantidad de oxígeno, de forma que un animal no adaptado a estas condiciones se encuentra frente a un desequilibrio fisiológico difícil de enfrentar. En general las especies adaptadas a la vida en las grandes alturas disponen de una gran cantidad de mecanismos homeostáticos que permiten su vida en este ambiente en extremo desfavorable. Las dificultades anteriormente señaladas son la principal causa de que el número de especies alto andinas sea relativamente reducido, pero esta condición, sumada a su asombrosa adaptación las hace particularmente interesantes y muy importantes desde el punto de vista de la biodiversidad (Raggi, 2000). Lo descrito anteriormente constituye pues el hábitat de los camélidos sudamericanos que en término generales está constituido principalmente por las
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
13

formaciones ecológicas de Puna y Altos Andes. Su altitud oscila entre los 4,800 y 500 metros. Su temperatura promedio es de entre 6ºC y 8ºC . Y su nivel de precipitación es de entre 400mm y 700 mm. En general, los camélidos pueden vivir en zonas más altas de los 5,000 metros. La alpaca prefiere vivir alrededor de las zonas húmedas o bofedales (humedales o wetlands ), La vicuña, en cambio, prefiere las praderas altas. Y la llama habita en todos los niveles, aunque prefiere los lugares secos. (Brenes, 2001). Aunque está muy difundida la creencia de que los camélidos son solamente animales de altura, en realidad, diversos estudios mencionados por Bonavia (1996) demuestran que en la época precolombina existían grandes hatos de camélidos aclimatados en la costa.
Las elevadas altitudes donde se desarrollan los camélidos (especialmente llama, vicuña y alpaca) tienen múltiples factores estresantes medioambientales, pero la hipoxia es el más importante componente. A través de muchos años de evolución, los animales se han movilizado desde un hábitat acuático marino a un hábito de aire atmosférico, conllevando con ello a enfrentar riesgos de baja concentración de O2 en el medio ambiente; es más, la invasión a zonas de elevadas altitudes se ha realizado en un periodo reciente, el cual no ha sido fácil para los seres vivos debido a que la concentración de O2 se encuentra en una función inversa a altitud, por lo que ello ha impuesto una gran reto a la vida animal (Monge y León-Velarde, 1991), debiendo por tanto los camélidos sudamericanos enfrentar ello mediante diversos mecanismo adaptativos fisiológicos.
Si bien el frío está íntimamente asociado con la hipoxia en todos los animales, pero con específicas implicaciones para los animales endotérmicos, esta revisión se concentrará principalmente en la hipoxia hipobárica como factor estresante, así como las adaptaciones morfológicas y fisiológicas hematológicas y circulatorias que el organismo de los camélidos ha tenido que modificar para afrontar ello, por tanto no se tratará aspectos de hipobaria, humedad atmosférica, incremento de radiación atmosférica y otros componentes medioambientales atmosféricos de elevadas altitudes que son más importante para los ectodermos que para los mamíferos y las aves (Monge y León-Velarde, 1991). Consecuentemente la intención de esta revisión es considerar aspectos de transporte de O2 durante la vida de los CS fenotípica y genotípicamente adaptados a elevadas altitudes para un mejor entendimiento de las capacidades y limitaciones que tienen estos animales para vivir en zonas de elevada altitud, para lo cual ha sido necesario examinar algunas investigaciones realizadas sobre fisiología adaptativa no solo en los animales en cuestión, sino también en otras especies animales (incluido el hombre) con finalidades comparativas.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
14

DEFINICIÓN DE TÉRMINOS
Adaptación fisiológica
El término adaptación fisiológica encuentra problemas de demarcación dentro del mismo campo de la fisiología, pues muchas veces se superponen con términos de fisiología comparativa, fisiología evolutiva, genética poblacional y otros campos relacionados a la biología.
Dentro de las adaptaciones también es posible vislumbrar las referidas las de tipo genotípico y las de tipo fenotípico. Mientras que las adaptaciones fenotípicas pueden ocurrir dentro de un periodo de vida del organismo del animal y se suprimen cuando las condiciones estresantes no existen y por lo tanto no son transmisibles a la prole; las adaptaciones genotípicas fijan cambios heredables, pudiendo ser estos cambios de forma y función.
Una convergencia que hay que tener en cuenta es la noción de beneficio que trae consigo las adaptaciones, por ello el concepto debe considerar implícitamente que las modificaciones que se operan en los organismos en presencia de circunstancias o entornos difíciles sirven para favorecer de alguna forma la supervivencia de los seres en cuestión. Los entornos naturales no son constantes, por lo tanto aquellos individuos con mayores capacidades para adaptarse podrían tener ventajas selectivas en la supervivencia y la reproducción.
Para la presente revisión, se tendrá en cuenta el concepto de Jhonson y Blig (citado por Monge y León-Velarde, 1991), que define a las adaptaciones como funciones que reducen el esfuerzo fisiológico producidos por un componente estresante del medioambiente. En el caso de adaptaciones genotípicas es más conveniente hablar de funciones más que de cambios debido a que los organismos adaptados han incorporado la función alterada como parte de su fisiología normal. Obviamente el concepto de beneficio para el individuo y eventualmente para las especies se encuentra implícito dentro de ésta definición.
Adaptación morfológica
En los mamíferos, las adaptaciones de estructura física representan algunos de los mas obvios, casi todos los medios de locomoción, provienen de adaptaciones estructurales de los apéndices, por ejemplo, las aletas en los mamíferos marinos, las piernas largas de los ungulados de las llanuras, las alas membranosas de los murciélagos, con frecuencia esta adaptación del miembro es tan especializada que hace que el mamífero se vuelva peligrosamente lento o torpe fuera de su medio. Las focas por ejemplo están adaptados a la vida acuática, pero en tierra no
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
15

se mueven con mucha rapidez; muchos murciélagos especializados para vivir en el aire apenas se pueden colgar patas arriba cuando aterrizan.
La velocidad depende de las adaptaciones estructurales de las patas y brazos de los mamíferos, los osos relativamente lentos y algunos primates con grandes pies que también son planos deben descansarlos en tierra en cada paso se les llama plantígrados porque apoyan con firmeza la planta en el piso, en cambio los gatos y perros que son mas rápidos, son digitígrados porque se apoyan y caminan sobre dedos, una tercera categoría de mamíferos que incluye caballos, reses y antílopes, son ungulados y se mueven sobre los extremos de las uñas de sus dedos, que están protegidas por las grandes uñas que llamamos pezuñas.
No todas las adaptaciones estructurales están relacionadas con la locomoción, las diversas formas y tamaños de las quijadas y dientes de los mamíferos son adaptaciones estructurales de la alimentación, por ejemplo los agudos caninos del carnívoro lobo se utilizan para acuchillar y desgarrar carne, los premolares y molares han sido adaptados para cortar, en el ciervo rumiante, predominan los dientes moledores, los caninos e incisivos de la mandíbula superior han sido sustituidos por una placa cornea, que sirve para cortar la vegetación (http://www.geocities.com/ludguimx/mamifadap.html).
Los camélidos también tienen presentan adaptaciones estructurales no solo de tipo macro, sino también micro. Las vicuñas tienen los miembros anteriores mucho mas pequeños que los posteriores, los cuales les permiten trepar con mucha rapidez las agrestes montañas donde habitan, y en general los CS han desarrollado una gran capacidad para producir fibras finas que les permiten vivir en condiciones de temperatura bastante bajas. Asimismo existen adaptaciones micro estructurales como el caso de la forma y tamaño de glóbulos rojos que les permiten tener una mejor eficiencia respiratoria – de lo cual se tratará ampliamente en la presente revisión.
Es obvio que dentro de esta terminología no se hace dificultosa la concepción de definición. Es debido a que las adaptaciones morfológicas son de tipo genotípico, vale decir son heredables, y que también son benéficos porque le brinda mayor ventaja que otros organismos para llevar una existencia menos esforzada en contraste con el entorno duro.
También existen adaptaciones conductuales, las cuales involucran modificaciones de comportamiento, sin embargo estas no serán enfocadas en la presente revisión, por lo cual no detallaremos ni en lo mínimo.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
16

Preadaptación.
El término de preadaptación se contrapone al desarrollo de estructuras en anticipación de necesidades futuras. Bock (1959) define el término de preadaptación de la siguiente forma: “una estructura es llamada preadaptada para una nueva función si su actual forma le permite cesar su función original mientras le permite asumir la nueva función siempre que se presente la necesidad de esta función.” Esto obviamente permite ser un paso hacia la evolución, pues así la mayoría – y posiblemente todas- las estructuras nuevas se originaron en base al involucramiento de antiguas estructuras las cuales fueron preadaptados para una función nueva; sin embargo el cambio de función no necesariamente involucra alteración radicación de forma.
Por lo tanto es importante vislumbrar el efecto que ejerce la selección natural en relación a la preadaptación. De este modo los organismos preadaptados estarían poco o casi nada afectados por la selección natural en nuevos entornos comparados con aquellos organismos cuya capacidad de preadaptación a nuevas condiciones medioambientales no están presentes.
Monge (1989) en relación a la adaptación de camélidos y aves a gradientes hipóxicos andinos considera la preadaptación como una capacidad genotípica para invadir un nuevo nicho ecológico como consecuencia de cambios morfológicos y funcionales previamente adquiridos. En el caso de una invasión rápida a nuevos nichos ecológicos, como el sucedido con los camélidos sudamericanos desplazados a los andes por los españoles, es aceptable concebir que la selección natural ha operado a su máximo nivel en éstos animales, quienes sin embargo ya tenían un mínimo nivel de capacidad preadaptativa para un medioambiente hipóxico.
De este modo en concordancia con su historia evolutiva, un animal puede tener diferentes posibilidades en su adaptación para nuevas condiciones medioambientales: Puede estar genotípicamente adaptado como algunos animales nativos a zonas altas, o puede estar fenotípicamente adaptado como muchos animales domésticos introducidos a zonas altas recién hace pocos años atrás, lo cual le permitirían ocupar zonas con altitudes desde el nivel del mar zonas con altitudes bastante elevadas.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
17
ERITROMORFOCINÉTICA EN CAMELIDOS SUDAMERICANOS
Aspectos morfológicos.
a. Sangre periférica.
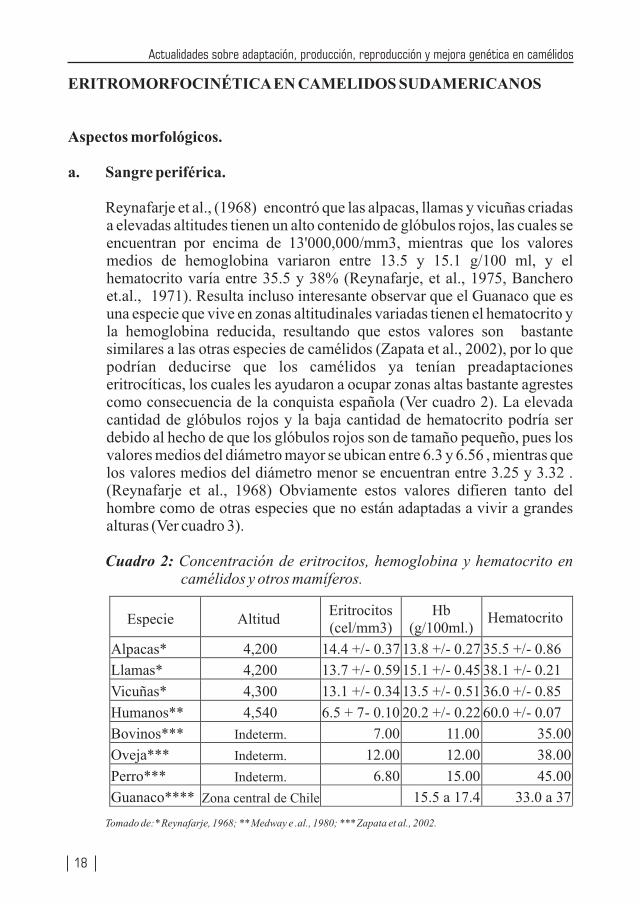
Reynafarje et al., (1968) encontró que las alpacas, llamas y vicuñas criadas a elevadas altitudes tienen un alto contenido de glóbulos rojos, las cuales se encuentran por encima de 13'000,000/mm3, mientras que los valores medios de hemoglobina variaron entre 13.5 y 15.1 g/100 ml, y el hematocrito varía entre 35.5 y 38% (Reynafarje, et al., 1975, Banchero et.al., 1971). Resulta incluso interesante observar que el Guanaco que es una especie que vive en zonas altitudinales variadas tienen el hematocrito y la hemoglobina reducida, resultando que estos valores son bastante similares a las otras especies de camélidos (Zapata et al., 2002), por lo que podrían deducirse que los camélidos ya tenían preadaptaciones eritrocíticas, los cuales les ayudaron a ocupar zonas altas bastante agrestes como consecuencia de la conquista española (Ver cuadro 2). La elevada cantidad de glóbulos rojos y la baja cantidad de hematocrito podría ser debido al hecho de que los glóbulos rojos son de tamaño pequeño, pues los valores medios del diámetro mayor se ubican entre 6.3 y 6.56 , mientras que los valores medios del diámetro menor se encuentran entre 3.25 y 3.32 . (Reynafarje et al., 1968) Obviamente estos valores difieren tanto del hombre como de otras especies que no están adaptadas a vivir a grandes alturas (Ver cuadro 3).
Cuadro 2: Concentración de eritrocitos, hemoglobina y hematocrito en camélidos y otros mamíferos.
Tomado de:* Reynafarje, 1968; ** Medway e .al., 1980; *** Zapata et al., 2002.
Especie AltitudEritrocitos (cel/mm3)
Hb (g/100ml.)
Hematocrito
Alpacas* 4,200 14.4 +/- 0.37 13.8 +/- 0.27 35.5 +/- 0.86
Llamas* 4,200 13.7 +/- 0.59 15.1 +/- 0.45 38.1 +/- 0.21
Vicuñas* 4,300 13.1 +/- 0.34 13.5 +/- 0.51 36.0 +/- 0.85
Humanos** 4,540 6.5 + 7- 0.10 20.2 +/- 0.22 60.0 +/- 0.07
Bovinos*** Indeterm. 7.00 11.00 35.00
Oveja*** Indeterm. 12.00 12.00 38.00
Perro*** Indeterm. 6.80 15.00 45.00
Guanaco**** Zona central de Chile 15.5 a 17.4 33.0 a 37
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
18
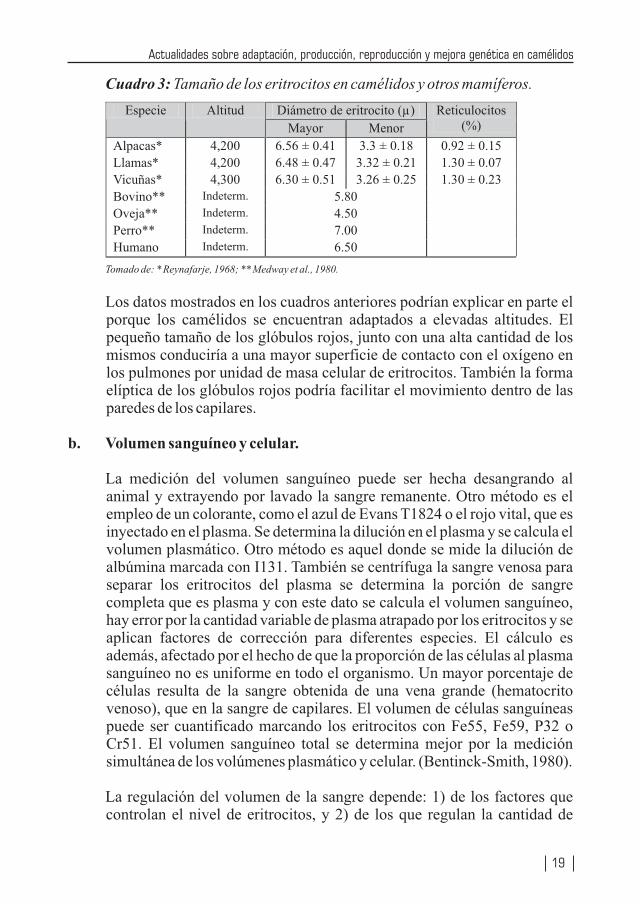
Cuadro 3: Tamaño de los eritrocitos en camélidos y otros mamíferos.
Tomado de: * Reynafarje, 1968; ** Medway et al., 1980.
Los datos mostrados en los cuadros anteriores podrían explicar en parte el porque los camélidos se encuentran adaptados a elevadas altitudes. El pequeño tamaño de los glóbulos rojos, junto con una alta cantidad de los mismos conduciría a una mayor superficie de contacto con el oxígeno en los pulmones por unidad de masa celular de eritrocitos. También la forma elíptica de los glóbulos rojos podría facilitar el movimiento dentro de las paredes de los capilares.
b. Volumen sanguíneo y celular.
La medición del volumen sanguíneo puede ser hecha desangrando al animal y extrayendo por lavado la sangre remanente. Otro método es el empleo de un colorante, como el azul de Evans T1824 o el rojo vital, que es inyectado en el plasma. Se determina la dilución en el plasma y se calcula el volumen plasmático. Otro método es aquel donde se mide la dilución de albúmina marcada con I131. También se centrífuga la sangre venosa para separar los eritrocitos del plasma se determina la porción de sangre completa que es plasma y con este dato se calcula el volumen sanguíneo, hay error por la cantidad variable de plasma atrapado por los eritrocitos y se aplican factores de corrección para diferentes especies. El cálculo es además, afectado por el hecho de que la proporción de las células al plasma sanguíneo no es uniforme en todo el organismo. Un mayor porcentaje de células resulta de la sangre obtenida de una vena grande (hematocrito venoso), que en la sangre de capilares. El volumen de células sanguíneas puede ser cuantificado marcando los eritrocitos con Fe55, Fe59, P32 o Cr51. El volumen sanguíneo total se determina mejor por la medición simultánea de los volúmenes plasmático y celular. (Bentinck-Smith, 1980).
La regulación del volumen de la sangre depende: 1) de los factores que controlan el nivel de eritrocitos, y 2) de los que regulan la cantidad de
Diámetro de eritrocito (µ)Especie Altitud
Mayor Menor
Reticulocitos (%)
Alpacas* 4,200 6.56 ± 0.41 3.3 ± 0.18 0.92 ± 0.15
Llamas* 4,200 6.48 ± 0.47 3.32 ± 0.21 1.30 ± 0.07
Vicuñas* 4,300 6.30 ± 0.51 3.26 ± 0.25 1.30 ± 0.23
Bovino** Indeterm. 5.80
Oveja** Indeterm. 4.50
Perro** Indeterm. 7.00
Humano Indeterm. 6.50
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
19

plasma: presión sanguínea, producción de orina, gasto cardiovascular, sed, ingestión de sodio y acción hormonal de la aldosterona y la hormona antidiurética. El aumento primario en la producción de eritrocitos es raro (policitemia vera), pero en animales en gran altitud puede ocurrir una policitemia secundaria.
Desde que Viault (citado por Monge y León-Velarde, 1991) describió en los andes peruanos incrementos de eritrocitos en humanos visitantes, residentes y animales que vivían en la ciudad de Morococha a 4540 m. de altitud, se despertó un gran interés en el rol adaptativo de la policitemia a elevadas altitudes. Los animales domésticos introducidos a zonas de elevada altitud en periodos históricos recientes, muestran grados variables de policitemia. En contraste a ello los mamíferos genotípicamente adaptados a elevadas altitudes como son los camélidos sudamericanos (alpaca, llama y vicuña) y los roedores andinos (chinchilla, vizcacha), muestran un modesto o ningún incremento en el hematocrito; asimismo la concentración de hemoglobina dentro de los eritrocitos es más alto comparado en la escala de mamíferos. Las llamas, alpacas y vicuñas tienen un tasa de retorno de Fe eritrocítico más alta que los humanos que viven a la misma altitud y la vida media de los eritrocitos es cerca de la mitad que la de los humanos. El volumen eritrocítico que fluctúa alrededor de 25-31 ml/kg es también mucho más bajo. Debido a estas consideraciones la sangre de los camélidos tienen una baja viscosidad pero con una normal concentración de hemoglobina, que dan especial condición para la vida en zonas de elevadas altitudes.
Reynafarge (1968), midió la cantidad de plasma y glóbulos rojos en 12 alpacas, 12 llamas criados en Corpacancha, lugar ubicado a 4,200 m de altitud, utilizando la técnica azul de Evans. También realizó mediciones en vicuñas en la Raya (4300 m de altitud), encontrando valores de masa celular roja de 25, de 24.3 y 31.3 ml/kg de peso corporal en llamas, alpacas y vicuñas. Los valores de plasma resultaron ser dos veces mayor que la masa de células rojas (Cuadro 3).
Cuadro 4: Volumen sanguíneo y celular en camélidos y humanos.
Tomado de: Reynafarje, (1968).
Número Altitudm
Totalml/kg peso corporal
Plasmaml/kg peso corporal
Eritrocitosml/kg peso corporal
10 alpacas 4200 72.0 ± 5.3 45.3 ± 3.4 24.3 ± 2.210 llamas 4200 65.2 ± 4.1 40.2 ± 2.1 25.0 ± 1.410 vicuñas 4300 86.6 ± 2.1 56.0 ± 1.4 31.3 ± 0.910 humanos 4540 88.1 ± 1.5 33.1 ± 0.9 55.0 ± 1.3
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
20

Se hace interesante destacar que en humanos nativos de zonas de elevadas altitudes la masa celular roja es dos veces más grande que la de los camélidos, y que el volumen de plasma es mucho menos en humanos que en camélidos.
De lo anteriormente indicado se hace plausible que tanto los visitantes y nativos a zonas de elevada altitud desarrollan grados variables de policitemia; asimismo un incremento en el hematocrito con la edad de nativos que viven a elevadas altitudes, podría ser interpretada como un indicador de la limitada capacidad de estas poblaciones para adaptarse a éstos entornos, sin embargo pareciera que nuestros camélidos sudamericanos debido a que están genotípicamente adaptados a elevadas altitudes no muestran incrementos de concentración de hemoglobina, ni tampoco muestran policitemia, a pesar de los cambios de altura lo que podría ser consecuencia de una adaptación sin poliglobulia (Raggi, 1998). A dicha adaptación se sumaría el bajo volumen celular el cual hace que la sangre pueda fluir mejor a nivel de los vasos, toda vez que la resistencia de las paredes serían menores.
c. Características de la hemoglobina.
Cuando se analiza la hemoglobina se hace conveniente estudiarla con respecto a su desnaturalización, curva de disociación, electroforesis, fragilidad, entre otros, los cuales nos darían alguna pista sobre los procesos adaptativos involucrados en los camélidos para soportar elevadas altitudes.
La hemoglobina es una proteína conjugada que consta de cuatro grupos “hemos” con un tetraedro. El hemo es una porfirina en cuyo centro está un átomo de hierro en forma ferrosa (Ver figura 1). La porfirina tiene cuatro anillos pirrólicos unidos por cuatro puentes de meteno. El núcleo porfirina tiene substituciones de cuatro metilos: dos virilos y dos grupos de acido propiónico.
Figura 1: Grupo Hemo.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
21

La hemoglobina de la llama difiere de la hemoglobiana humana en que tiene 25 diferentes aminoácidos en la cadena alfa y 24 diferentes aminoácidos en la cadena beta. El hallazgo inicial que la afinidad al oxígeno de la hemoglobina es dependiente de su contenido de 2,3 difosfoglicerato (2,3 DPG), es interpretada como una mutación del 2,3 DPG en posición his â-2 de humanos a la posición Asn Beta 2 en llamas, mientras que uno de los cuatro puntos de contacto se encuentra interrumpido. Esta interrupción daría lugar a una reducción disminuida de la afinidad intrínseca del oxígeno con la molécula de la hemoglobina y explicaría, sobre una base molecular, la afinidad creciente del oxígeno de la hemoglobina de la llama, y por lo tanto, la facilidad de la respiración de las llamas que viven a elevadas altitudes. Por analogía, la afinidad creciente O2 de la hemoglobina fetal humana es interpretada por la inactivación del punto de contacto del fosfoglicerato â-143. (Braunitzer et al., 1978).
La hemoglobina tiene la capacidad de inactivar tanto al factor relajante derivado del endotelio (EDRF) como al óxido nítrico (NO). Se ha demostrado que la hemoglobina puede reaccionar rápidamente con el NO, para formar nitrato a partir de la oxihemoglobina o nitrosol hemoférrico a partir de dioxihemoglobina. En virtud a la indicado las células de la sangre a través de la hemoglobina realiza una limpieza química de NO y puede actuar como vasoconstrictor. En el flujo sanguíneo los glóbulos rojos de la sangre tienden a permanecer en la parte media de los vasos y así evitar las paredes donde se produce NO. Otra de las situaciones es que el empaquetado celular del Hb retarda su reacción con el NO debido a la formación oxidativa de los tionitritos, que mantienen bioactividad cardiovascular en la presencia de Hb, evitando de las reacciones de limpieza de NO, (Singer y Stamler, 2005).
Debido a que la cantidad de hemoglobina en camélidos sudamericanos es levemente diferente (alta comparada con otras especies domésticas, pero baja comparada con humanos) y el hematocrito es baja, el mejor flujo sanguíneo en los vasos podría explicarse debido a que existiría un menor contacto de los glóbulos rojos, y de la hemoglobina con las paredes de los vasos, debido a ello la inactivación del factor relajante derivado del endotelio y del óxido nítrico sería menor, dando mayor oportunidad a éstos para la relajación de los vasos y así disminuir la resistencia del flujo sanguíneo, lo cual evitaría problemas de elevadas presiones vasculares, lo cual dificulta las actividades en humanos y animales no adaptados a zonas altoandinas, conllevando a situaciones extremas de muerte, lo cual no se daría en los camélidos sudamericanos. Esta posibilidad se podría referir como una situación de adaptación morfofisiológica de la llama, alpaca y vicuña que viven en situaciones extremas de altitud.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
22

La resistencia a la desnaturalización por álcali, muestra la persistencia de hemoglobina F, característica de la hemoglobina fetal, la cual tiene alta afinidad por el oxígeno. En la sangre de alpacas adultas se ha encontrado que mas del 50% de la hemoglobina es resistente a la desnaturalización por álcali, el cual es bastante superior al encontrado en humanos adultos que viven en zonas de elevada altitud, donde apenas el 0.7% posee esta resistencia. (Cuadro 5).
Cuadro 5: Desnaturalización alcalina en alpacas y humanos que viven a elevadas altitudes.
Tomado de: Reynafarje, (1975).
Cuando se realiza una corrida electroforética de hemoglobinas de alpacas y humanos, estos últimos tienen una mayor movilidad (Figura 2). Es conocido que la movilidad de la hemoglobina depende de la secuencia de aminoácidos de la cadena polipeptídica, el cual tiene interacción con el grupo hemo, lo cual afecta la afinidad de la hemoglobina por el oxígeno (Reynafarje, 1975), explicando de este modo la mayor afinidad lo cual referido a los camélidos explicaría un mejor mecanismo de adaptación.
Reynafarje, 1975, encontró que en alpacas con respecto a la enzima deshidrogenasa láctica es casi seis veces más alta comparada con humanos (179.7 Vs. 33.0 U/ml de suero), y que los niveles de la deshidrogenasa Glucosa-6-Fosfato es dos veces más alta (327.5 Vs. 162 U/ml de masa globular). Estos hallazgos dan una indicación de la intensidad de la actividad glicolítica en camélidos como compensación de los mecanismos de abastecimiento bajo de oxígeno a elevadas altitudes y condiciones hipóxicas, el cual probablemente sería debido a la mayor tasa de renovación de los eritrocitos en alpacas, como consecuencia de un periodo corto de vida de estos corpúsculos sanguíneos; por tanto el importante rol de la deshidrogenasa láctica en la actividad glicolítica sería otro mecanismo que contribuiría a la adaptación exitosa de los camélidos.
Hb F (%)Alpacas crías 61.8 ± 3.2Alpacas adultas 54.5 ± 8.8Bebes 55.9 ± 3.3Personas adultas 0.7 ± 0.05
Especie
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
23
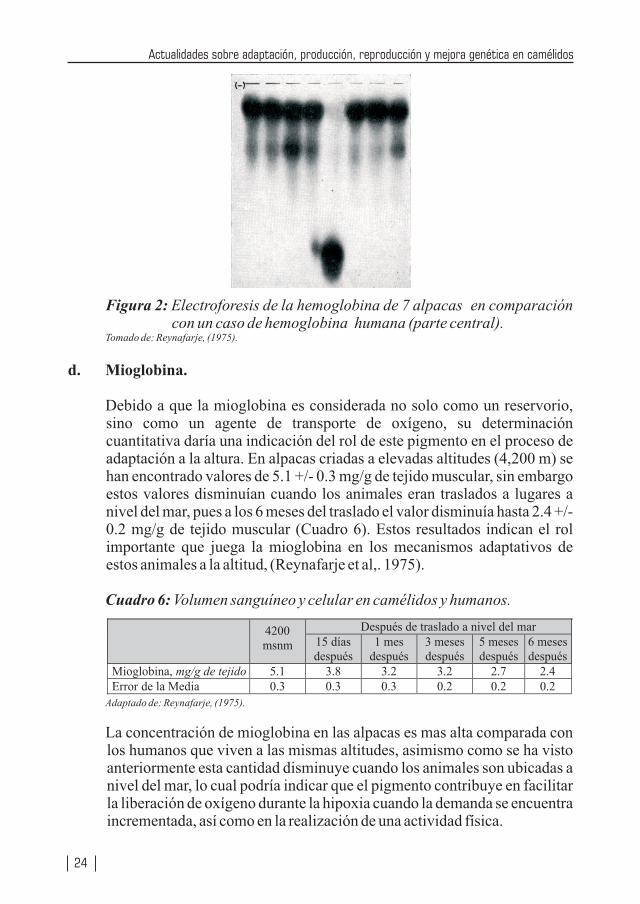
Figura 2: Electroforesis de la hemoglobina de 7 alpacas en comparación con un caso de hemoglobina humana (parte central).
Tomado de: Reynafarje, (1975).
d. Mioglobina.
Debido a que la mioglobina es considerada no solo como un reservorio, sino como un agente de transporte de oxígeno, su determinación cuantitativa daría una indicación del rol de este pigmento en el proceso de adaptación a la altura. En alpacas criadas a elevadas altitudes (4,200 m) se han encontrado valores de 5.1 +/- 0.3 mg/g de tejido muscular, sin embargo estos valores disminuían cuando los animales eran traslados a lugares a nivel del mar, pues a los 6 meses del traslado el valor disminuía hasta 2.4 +/- 0.2 mg/g de tejido muscular (Cuadro 6). Estos resultados indican el rol importante que juega la mioglobina en los mecanismos adaptativos de estos animales a la altitud, (Reynafarje et al,. 1975).
Cuadro 6: Volumen sanguíneo y celular en camélidos y humanos.
Adaptado de: Reynafarje, (1975).
La concentración de mioglobina en las alpacas es mas alta comparada con los humanos que viven a las mismas altitudes, asimismo como se ha visto anteriormente esta cantidad disminuye cuando los animales son ubicadas a nivel del mar, lo cual podría indicar que el pigmento contribuye en facilitar la liberación de oxígeno durante la hipoxia cuando la demanda se encuentra incrementada, así como en la realización de una actividad física.
Después de traslado a nivel del mar4200 msnm 15 días
después1 mes
después3 meses después
5 meses después
6 meses después
Mioglobina, mg/g de tejido 5.1 3.8 3.2 3.2 2.7 2.4Error de la Media 0.3 0.3 0.3 0.2 0.2 0.2
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
24
Aspectos cinéticos.
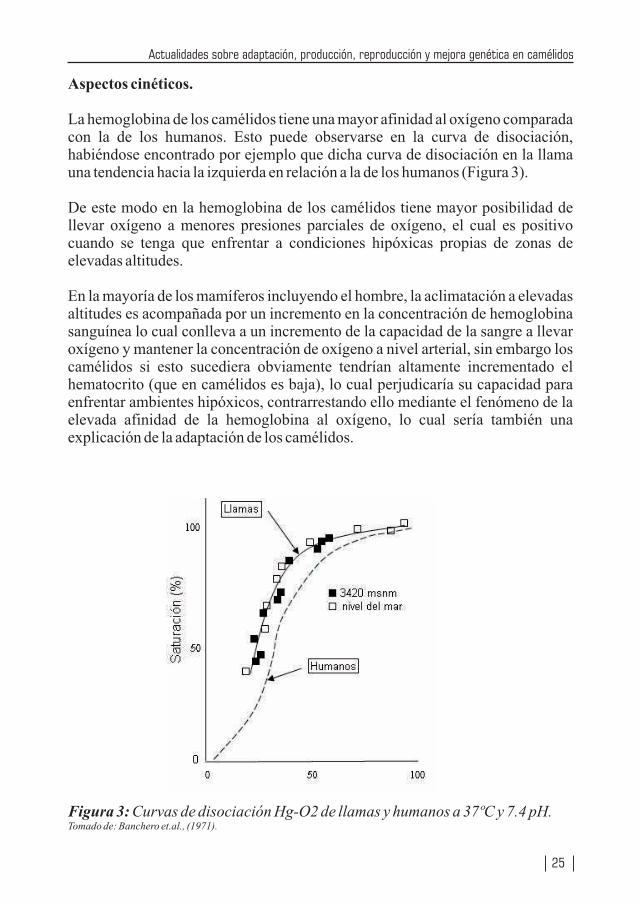
La hemoglobina de los camélidos tiene una mayor afinidad al oxígeno comparada con la de los humanos. Esto puede observarse en la curva de disociación, habiéndose encontrado por ejemplo que dicha curva de disociación en la llama una tendencia hacia la izquierda en relación a la de los humanos (Figura 3).
De este modo en la hemoglobina de los camélidos tiene mayor posibilidad de llevar oxígeno a menores presiones parciales de oxígeno, el cual es positivo cuando se tenga que enfrentar a condiciones hipóxicas propias de zonas de elevadas altitudes.
En la mayoría de los mamíferos incluyendo el hombre, la aclimatación a elevadas altitudes es acompañada por un incremento en la concentración de hemoglobina sanguínea lo cual conlleva a un incremento de la capacidad de la sangre a llevar oxígeno y mantener la concentración de oxígeno a nivel arterial, sin embargo los camélidos si esto sucediera obviamente tendrían altamente incrementado el hematocrito (que en camélidos es baja), lo cual perjudicaría su capacidad para enfrentar ambientes hipóxicos, contrarrestando ello mediante el fenómeno de la elevada afinidad de la hemoglobina al oxígeno, lo cual sería también una explicación de la adaptación de los camélidos.
Figura 3: Curvas de disociación Hg-O2 de llamas y humanos a 37ºC y 7.4 pH.Tomado de: Banchero et.al., (1971).
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
25

La cantidad de oxígeno disponible a nivel de los tejidos celulares depende del ritmo cardiaco y también del contenido de oxígeno de la sangre arterial. En los camélidos como en los humanos, la extracción de oxígeno se encuentra alrededor de 40-45 ml/L. Esto significa que los camélidos en la cual la concentración de hemoglobina es baja y por lo tanto tienen una menor concentración de oxígeno a nivel de sangre arterial, posee mecanismos más eficientes para la extracción de oxígeno. En efecto, se ha probado que el coeficiente de utilización de oxígeno es 50% mas alto que en el hombre, por lo que en llamas se ha encontrado una menor presión parcial y menor saturación de oxígeno a nivel de sangre venosa (Banchero et al., 1971).
Aun no está claro cuales son los mecanismos fisiológicos para esta mejor extracción de oxígeno a nivel de los tejidos, y aunque se podría pensar que debido a una mayor afinidad del oxígeno por la hemoglobina quedaría disminuido la transferencia del oxígeno del nivel arterial al nivel tisular, esto no sucede en los camélidos quienes oxigenan en forma eficiente sus tejidos. Es conocido que el oxígeno pasa a las células por difusión y que sólo un pequeño diferencial de presión es necesario para que a nivel celular se provea de oxígeno a las mitocondrias, que en último caso utilizan dicho insumo para la producción de energía. Un incremento de la densidad de los capilares con distancias mas cortas entre los capilares y las células podrían ciertamente favorecer la difusión desde la sangre a las células favoreciendo la extracción del oxígeno. Este mecanismo ha sido demostrado en cuyes y posiblemente podrían jugar un rol significativo en los camélidos.
TRANSPORTE DE OXIGENO EN CAMELIDOS SUDAMERICANOS
Intercambio de oxígeno a nivel pulmonar.
Después que los alvéolos se han ventilado con aire fresco, el siguiente paso en el proceso respiratorio es la difusión del oxígeno desde los alvéolos hacia la sangre pulmonar y el paso del dióxido de carbono en dirección opuesta: de la sangre pulmonar hacia los alveolos. El proceso de difusión es simple, incluye simplemente el movimiento al azar de las moléculas que entrelazan sus caminos en uno y otro sentido a través de la membrana respiratorio. El oxígeno absorbido continuamente va hacia la sangre de los pulmones, y penetra oxígeno nuevo de modo constante en los alvéolos desde la atmósfera. Cuanto más rápidamente es absorbido el oxígeno, menor resulta su concentración en los alvéolos; por otra parte, cuanto más rápidamente es aportado oxígeno nuevo a los alvéolos desde la atmósfera, mayor va siendo su concentración. Por ello, la concentración de oxígeno en los alveólos, así como su presión parcial dependen, en primer lugar, de la rapidez de absorción de oxígeno hacia la sangre, luego de la rapidez de penetración de oxígeno nuevo en los pulmones gracias al proceso ventilatorio (Guyton, 1988).
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
26

La primera línea de transporte del oxígeno desde el medio ambiente a la sangre es la ventilación de aire por convección. En humanos expuestos a extremas altitudes se ha observado que la ventilación es el parámetro más importante durante el ascenso a elevadas altitudes. La tasa de ventilación por tanto juega un rol importante en la capacidad adaptativa de nativos a elevadas altitudes. Mamíferos tales como llamas, alpacas, ovejas, vacas, cabras expuestas a condiciones hipóxicas bastante largas también muestran respuestas hiperventilatorias, sin embargo resulta interesante ver que de los animales citados tanto la llama y las alpacas con animales nativos adaptados a elevadas altitudes, lo cual muestra en ellas un mecanismo de preadaptación. Sin embargo es de notar que en llamas expuestas a altitudes de 3420 msnm, luego de haber sido criadas a nivel del mar la ventilación alveolar no cambia en contraste con la hiperventilación que normalmente ocurre en humanos. Esta falla de un incremento en la ventilación concuerda al encontrado por diversos investigadores, habiéndose determinado que la respuesta ventilatoria hipóxica ocurre cuando la presión de oxígeno a nivel alveolar cae por debajo de 60 mm mercurio, y la presión de oxígeno arterial cae por debajo de 40 mm Hg. (Monge y León-Velarde, 1991).
Van Nice et al. (1980), en un estudio comparativo de gatos, ratas, llamas, patos, pekines y gansos hizo un intento de correlación entre la presión parcial de oxígeno a nivel arterial de los iniciales incrementos de ventilación con la afinidad de Hb-O, habiendo encontrado que la respuesta ventilatoria resulta ser baja si la afinidad Hb-O es alta. Esto explicaría que los camélidos saltaron la valla de la preadaptación a la valla de adaptación, debido a que su baja respuesta hiperventilatoria estaría subyugado a la elevada afinidad de Hb-O.
La ecuación de difusión de Fick es aplicado a la transferencia de oxígeno entre la sangre y los pulmones:
VO = GLO (PAO -PaO ) ..............................Ecuación 12 2 2 2
y la ecuación de resistencia desarrollada por Roughton y Foster
1/GLO = 1/ GMO + 1/ GBO .......................Ecuación 22 2 2
permiten un análisis de las posibilidades de adaptación de la capacidad de difusión pulmonar [conductancia de difusión (GLO )], sus componentes 2
[conductancia de la membrana (GMO ) y conductancia cinética sanguínea 2
(GBO )], y la diferencia de presión parcial arterial (PAO -PaO ). VO es el 2 2 2 2
consumo de oxígeno corporal.
Desde un punto de vista adaptativo, al analizar la ecuación 1, y teniendo en consideración que (PAO -PaO ), es menor en camélidos y que no cambia el VO , 2 2 2
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
27

entonces lo que estaría incrementado sería GLO m y dependiendo este último de 2
GMO y GBO , sin embargo éste componente sanguíneo no ha sido aún 2 2
debidamente estudiado por lo menos en camélidos. El componente de la membrana pareciera depender del área de la superficie de los alveolos, mas que de la modificación estructural de la membrana propiamente dicha.
En humanos nacidos en zonas de elevada altitud la gradiente (PAO -PaO ) es 2 2
reducida comparada con valores a nivel del mar. Esta gradiente también se encuentra reducida en perros (Monge y León-Velarde, 1991). La falta de respuesta ventilatoria en llamas a valores de PaO entre 51-53 mm Hg, podrían 2
estar relacionados al mantenimiento de una alta saturación de oxígeno arterial (92-93%), mostrándose esto cuando se administran a las llamas una mezcla de gas con bajo nivel de oxígeno, mientras que la PaO cae a 31-33 mm Hg y la 2
saturación disminuye a 72-78% ocurre una moderada hiperventilación, disminuyendo la PaO y por tanto reduciéndose la diferencia PIO -PAO .2 2 2
La forma de la curva de disociación de la Hb-O , la afinidad del O por la Hb, la 2 2
concentración de la Hb en el torrente sanguíneo y la frecuencia cardiaca son factores importantes en el mantenimiento de la presión de oxígeno a nivel de los vasos, por lo tanto es razonable pensar en un elevado nivel de éstos parámetros cuando los mamíferos son expuestos a zonas altas. Varios investigadores recientemente han sugerido que una disminución de la afinidad de la Hb por el O 2
facilita la descarga del O a nivel de los tejidos en humanos en condiciones de 2
hipoxia; sin embargo cuando las llamas ascienden a elevadas altitudes no se encuentra disminución de la afinidad del O por la Hb. 2
Otro aspecto de la aclimatación a elevadas altitudes es el incremento de la concentración de la hemoglobina que conlleva a mejorar la capacidad de transporte de O , sin embargo en llamas la concentración de la hemoglobina 2
sanguínea, así como el hematocrito disminuyen con la altitud (Cuadro 7), sin embargo esta aparente paradoja sería una consecuencia de elevados valores de la concentración de hemoglobina en zonas de baja altitud (Ver Cuadro 2), cuando se comparan con vacunos y ovejas. Esta disminución es una forma clásica de un proceso de aclimatación de los camélidos a zonas bajas, reflejando contrariamente el mecanismo adaptativo a zonas altoandinas.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
28
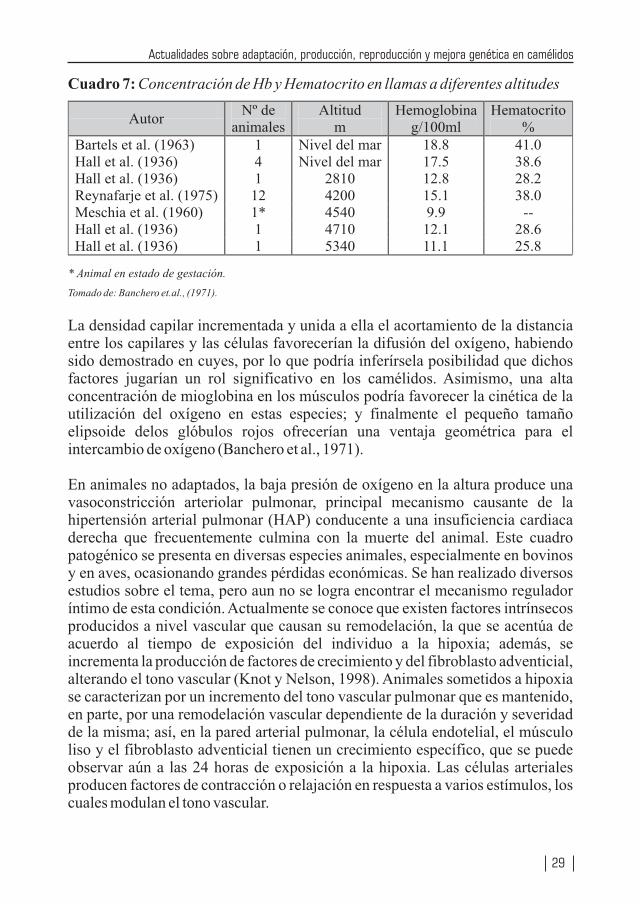
Cuadro 7: Concentración de Hb y Hematocrito en llamas a diferentes altitudes
Tomado de: Banchero et.al., (1971).
La densidad capilar incrementada y unida a ella el acortamiento de la distancia entre los capilares y las células favorecerían la difusión del oxígeno, habiendo sido demostrado en cuyes, por lo que podría inferírsela posibilidad que dichos factores jugarían un rol significativo en los camélidos. Asimismo, una alta concentración de mioglobina en los músculos podría favorecer la cinética de la utilización del oxígeno en estas especies; y finalmente el pequeño tamaño elipsoide delos glóbulos rojos ofrecerían una ventaja geométrica para el intercambio de oxígeno (Banchero et al., 1971).
En animales no adaptados, la baja presión de oxígeno en la altura produce una vasoconstricción arteriolar pulmonar, principal mecanismo causante de la hipertensión arterial pulmonar (HAP) conducente a una insuficiencia cardiaca derecha que frecuentemente culmina con la muerte del animal. Este cuadro patogénico se presenta en diversas especies animales, especialmente en bovinos y en aves, ocasionando grandes pérdidas económicas. Se han realizado diversos estudios sobre el tema, pero aun no se logra encontrar el mecanismo regulador íntimo de esta condición. Actualmente se conoce que existen factores intrínsecos producidos a nivel vascular que causan su remodelación, la que se acentúa de acuerdo al tiempo de exposición del individuo a la hipoxia; además, se incrementa la producción de factores de crecimiento y del fibroblasto adventicial, alterando el tono vascular (Knot y Nelson, 1998). Animales sometidos a hipoxia se caracterizan por un incremento del tono vascular pulmonar que es mantenido, en parte, por una remodelación vascular dependiente de la duración y severidad de la misma; así, en la pared arterial pulmonar, la célula endotelial, el músculo liso y el fibroblasto adventicial tienen un crecimiento específico, que se puede observar aún a las 24 horas de exposición a la hipoxia. Las células arteriales producen factores de contracción o relajación en respuesta a varios estímulos, los cuales modulan el tono vascular.
AutorNº de
animalesAltitud
mHemoglobina
g/100mlHematocrito
%Bartels et al. (1963) 1 Nivel del mar 18.8 41.0Hall et al. (1936) 4 Nivel del mar 17.5 38.6Hall et al. (1936) 1 2810 12.8 28.2Reynafarje et al. (1975) 12 4200 15.1 38.0Meschia et al. (1960) 1* 4540 9.9 --Hall et al. (1936) 1 4710 12.1 28.6Hall et al. (1936) 1 5340 11.1 25.8
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
29
* Animal en estado de gestación.

Investigaciones recientes demuestran la existencia de sustancias vasoactivas que actúan en respuesta a hipoxia, tales como la Sustancia P (SP), un neuromodulador del grupo de neuropéptidos (NPS). Esta sustancia es un potente vasodilatador secretada por los nervios perivasculares y liberada dentro de las células endoteliales (Schaffer et al., 1998).
Se sabe que en camélidos que viven a niveles altos sobre el nivel del mar no desarrollan HAP debido al problema hipóxico, por lo que podría caber la posibilidad de encontrar altas concentraciones de SP lo cual conllevaría a evitar la presencia de HAP. Si bien esto no se ha estudiado en camélidos, sin embargo en pollos Bernilla et al.(2003), pudo encontrar una rápida disminución de la PAPm debido a la aplicación de la SP sugiriendo la posible influencia a nivel endotelial vascular por su acción vasodilatadora, disminuyendo el efecto presor que, a su vez, podría estar relacionado con la hiperpolarización de la membrana o con el aumento de la actividad del óxido nítrico.
Transporte de oxígeno sanguíneo
La ecuación de Fick ,
VO = QB (CaO -CvO ) ..............................Ecuación 32 2 2
donde QB, CaO y CvO representan el flujo sanguíneo por unidad de tiempo, 2 2
contenido de oxígeno a nivel arterial y venoso respectivamente. El producto de QB*CaO representa la cantidad de transporte de oxígeno que puede llevar el 2
torrente sanguíneo, estando QB*CaO en función a la concentración de 2
hemoglobina [Hb] y a la saturación de oxígeno en la sangre arterial SaO .2
CaO = (SaO )*1.3[Hb] ..............................Ecuación 42 2
A elevadas altitudes SaO disminuye, pero en camélidos esta disminución es 2
menor en comparación con otras especies, debido al desplazamiento a la izquierda de la curva de disociación Hb-O lo cual mantiene la capacidad de transporte de oxígeno, y mejorando aún la CaO debido a la elevada cantidad de 2
Hb en la sangre.
Una disminución en QB ha sido descrita en ovinos expuestos a 4,573 m por 12 días. Una redistribución del flujo sanguíneo con un flujo incremento a nivel del corazón y cerebro y una disminución del flujo a nivel de vísceras ha sido observado en ovejas adultas expuestas a hipoxia durante 96 horas. En cambio en llamas criadas a nivel del mar, luego de expuestas por 4 semanas a 4320 m no se encontraron cambios en QB, lo cual conlleva a inferir la capacidad de adaptación de estos animales, pues al no disminuir dicho parámetro no afecta VO .2
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
30

Si la diferencia de CaO -CvO es dividida por CaO , entonces es posible obtener 2 2 2
coeficiente de extracción. Este coeficiente mide la capacidad de los tejidos para extraer O por unidad de nivel de CaO2. Estudios de Banchero et al. (1971), citado 2
por Monge y León-Velarde (1991) encuentra que personas nativas de zonas altoandinas tienen un coeficiente promedio de 0.205, llamas criadas a 3420 m, 0.317, gansos y patos pekines (a 6,100 m por 4 semanas) 0.422 y 0.633 respectivamente. Estos resultados interpretados teniendo en consideración procesos policitémicos en humanos y patos pekines, muestran la complejidad de la fisiología comparativa cuando las analogías son realizadas entre respuestas de animales adaptados fenotípicamente y genotípicamente a elevadas altitudes.
La respuesta eritropoyética a la hipoxia ha sido atribuida a un incremento de la eritropoyetina (EPO). Se podría suponer un mecanismo de retroalimentación negativa que regula la eritropoyesis, el cual sería el siguiente: Una disminución de SaO podría incrementar la producción de EPO a nivel de riñon, 2
produciéndose por tanto una estimulación de la medula roja por la EPO para incrementar la eritropoyesis con un resultado de alta concentración de Hb.
En llamas, vicuñas y chinchilas de la cordillera andina la actividad eritropoyética es mas baja comparada con especies que se crían en zonas bajas y sometidas a las mismas condiciones de hipoxia. En el caso de llamas y alpacas, Reynafarje et al. (1975) encontró una similar concentración de EPO con la de humanos, sin embargo la capacidad de adaptación en los camélidos se reflejaría por la elevada cantidad de eritrocitos producto de una inducción eritropoyética con la misma cantidad o incluso menor de eritropoyetina, sin desmedro de una elevada concentración sanguínea (bajo hematocrito), que permite un adecuado flujo sanguíneo.
Los camélidos sudamericanos tienen una alta afinidad Hb-O , observándose 2
dicha particularidad también en aves, y roedores que viven en zonas altoandinas. Asimismo una similar y una afinidad ligeramente más alta también fue encontradas en otras especies de la familia Camelidae, el camello (tanto Camellus bactrianus y C. dromedraris); por tanto en consideración a ello parece interpretarseque la característica pertenece a la familia Camelidae y no necesariamente a especies que viven en zonas altoandinas, considerándose este proceso como un típico ejemplo de preadaptación.
La elevada afinidad Hb-O en ovejas parece ser determinado por el tipo de Hb, 2
habiéndose encontrado en estas especies dos tipos: A y B, los cuales son transmitidos de acuerdo a una simple herencia mendeliana. La Hb A tiene mayor SaO en condiciones de elevadas altitudes. En camélidos esto aún no ha sido 2
estudiado, y sería interesante determinar la posibilidad de existencia de diferentes tipos de hemoglobina que determinan esta característica preadaptativa.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
31

Resulta conveniente la interpretación de los resultados encontrados acerca de la regulación de la afinidad Hb-O . Al parecer en este sentido la enzima 2,3 2
difosfoglicerasa (2,3DPG) a influye fuertemente en la posición de la curva de equilibrio de oxígeno, debido a que ésta enzima tiene una fuerte tendencia a unirse a la hemoglobina reducida, lo cual disminuye la afinidad de Hb-O , 2
desplazando dicha curva hacia la derecha. Sin embargo al anotar esta característica no se debe dejar de tener en consideración el pH de la sangre, pues la actividad enzimática se ve restringida ante dicho factor. La llama y el guanaco tiene una reacción baja de la 2,3DPG comparadas con otros mamíferos. La llama tiene aspargina en posición BetaNA2, en vez de histidina, uno de los residuos normalmente responsables para la unión de 2,3 DPG. A nivel molecular, este hallazgo explicaría la baja actividad de la 2,3 DPG resultando en una alta afinidad Hb-O en llamas, alpacas y vicuñas. Aunque camélidos sudamericanos y 2
camellos tienen similares afinidades, la regulación de esta propiedad respondería a diferentes mecanismo moleculares. Bauer et al. (citado por Monge y León-Velarde, 1991) mostró que la unión constante de 2,3 DPG de la llama a la deoxihemoglobina es tres veces más pequeña comparada a la del camello, concluyendo que la diferencia se debería a sustitución S55NA2 de la cadena Beta en el sitio de unión a la 2,3 DPG.
Intercambio de oxígeno a nivel tisular.
Podríamos creer que la alta afinidad de Hb-O existente en el torrente sanguíneo 2
de los camélidos podría dificultar la liberación del O a nivel de los tejidos, sin 2
embargo Banchero et al. (1971), encontró que tanto en la llama como en el hombre la extracción de O en condiciones de descanso es alrededor de 40 a 45 2
ml/L. Esto significa que la llama, en la cual la concentración de Hb es más baja y por tanto una CaO también baja, posee un mecanismo mas eficiente para la 2
extracción de oxígeno. De hecho – indica Banchero et al. (1971) –que el coeficiente de utilización es 50% mayor comparado al del hombre. Pero ¿qué mecanismo fisiológico determina esta eficiencia?.
La ecuación de difusión Fick también ayuda a la discusión de la difusión de oxígeno sanguíneo a los tejidos:
VO = GCO (PaO -PTO ) .............................. Ecuación 52 2 2 2
donde GCO es la conductancia de difusión capilar y PTO es la presión parcial 2 2
de oxígeno a nivel tisular. Sin embargo aunque el problema de la medición PTO 2
ha ido mejorando, a través de uso de modelos y análisis teóricos, sugiriendo que la PVO es similar a la PTO esto aun no ha sido utilizado en alpacas ni en llamas, 2 2
por lo que el mecanismo de la eficiencia del descargo de O a nivel de capilares 2
histológicos aun es un punto que falta investigar bastante en camélidos.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
32

Por otro lado, se sabe que la difusión de oxigeno a nivel de tejidos puede ser mejorado mediante el acortamiento de la distancia de difusión, el cual incrementaría el nivel de PTO . Este acortamiento podría ocurrir debido ya sea a 2
un incremento del número de capilares dentro de los tejidos, a la relación células por capilares (C/F) o a la disminución del volumen de las células, manteniendo una constante C/F.
Si bien a nivel de tejido muscular, se han reportado disminución de la distancia de difusión en cuyes, ratas, perros y gatos, considerándose ello como un proceso adaptativo a cambios de hipoxia a elevadas altitudes que podrían proteger de la disminución de PO a nivel de tejidos, si esto sucediera en camélidos también 2
podría ser mejorado dicha adaptación debido a la mayor cantidad de eritrocitos, así como al incremento de superficie de los mismos debido a al pequeño tamaño.
Si bien se han puntualizado aspectos relacionados con la hipoxia, tampoco es menester dejar de mencionar otro factor: La temperatura fría, que también es propio de zonas altoandinas, lo cuales favorecerían o desfavorecería diversos procesos que permiten la adaptabilidad y preadaptabilidad de los camélidos para vivir sin mucho esfuerzo en condiciones donde muchos mamíferos y humanos no la pasan bien digamos, debido al enorme esfuerzo que hacen mediante simples procesos de aclimatación.
Conclusiones.
Es obvio que los camélidos se encuentran adaptados para vivir en zonas bastante elevadas, habiendo sido apoyado por mecanismos preadaptativos antes de invadir el nuevo nicho de la zonas altoandina. Claro que cuando discutimos de zonas altoandinas debiéramos también referirnos a efectos estresantes como la radiación, temperatura, humedad, entro otros, pero sin duda el factor de tensión mas importante resulta ser el efecto hipóxico. Con respecto a este último los camélidos han desarrollado adaptaciones fisiológicas importantes principalmente relacionados al aspecto de abastecimiento de oxígeno a las células tisulares, para lo cual tienen excelentes mecanismos de captación de oxígeno a nivel pulmonar, adecuados mecanismos en el transporte de oxígeno, e intrincados procesos para un mejor aprovechamiento del oxígeno a nivel tisular, lo cual permite no sufrir severos efectos tresores de la hipoxia atmosférica reinante en zonas altoandinas. Los mecanismos a que nos referimos no solo son de tipo morfológico – como la forma de la células o expansión pulmonar – sino tal vez mucho más importante son los mecanismos de tipo fisiológico, siendo el proceso más importante tal vez la elevada afinidad de Hb-O, que permite llevar suficientes cantidades de oxígeno hacia los tejidos a fin de mantener las condiciones homeostáticas para convenientes procesos metabólicos.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
33

Sin duda existen muchos aspectos que aun se mantienen a modo de “caja negra”, siendo tal vez el más importante y urgente de estudio la gran complejidad del proceso de abastecimiento de oxígeno a nivel celular aún cuando el diferencial de presiones a este nivel es baja.
LITERATURA CITADA
Banchero, N., R. Grover y J. Will. 1971. Oxygen transport in the llama (Lama glama). Respiration Physiology. 13:102-115.
Bentinck-Smith, J. 1980. Hematología. Patología Clínica Veterinaria. Editorial UTEHA. Impreso en México. 532 pp.
Bernilla, S., Cueva, S. y M. Ayon, 2003. Efecto de la Sustancia P sobre la presión arterial en pollos expuestos a la altura. Rev Inv Vet. Perú 2003; 14(2): 97-103
Bock, W. J., 1959. Preadaptation and multiple evolutionary pathways. Evolution 13: 194-211.
Bonavia, D. 1996 Los Camélidos Sudamericanos (Una introducción a su estudio). Instituto Francés de Estudios Andinos . Universidad Peruana Cayetano Heredia. Conservation International.
Braunitzer G., B. Schrank, A. Stangl y C. Bauer. 1978. Interaction between phosphate and protein, and the respiration of the llama, the human fetus and the horse. Physiol. Chem. 359(5):547-58.
Brenes,, E., M. Kryssia, F. Pérez y K. Valladares. 2001. El Cluster de los Camélidos en Perú: Diagnóstico Competitivo y Recomendaciones Estratégicas Instituto Centroamericano de Administración de Empresas INCAE.
CONACS. 1999. Población sudamericana de camélidos sudamericanos. Disponible en www.minag.gob.pe/conacs/. Accesado el 25 de Junio de 2006.
Geocities. Adaptación. Disponible enhttp://www.geocities.com/ludguimx/mamifadap.html). Accesado el 23/07/2006.
Guyton, A. 1988. Tratado de Fisiología Médica. Editorial McGraw-Hill. 7ma. Edición. España. 523 p.
Knot, H.J. y M.T. Nelson. 1998. Regulation of arterial diameter and wall Ca2+ in cerebral arteries of rat by membrane potential and intravascular pressure. J. Physiol. 508: 199-209.
Medway, W., Prior, J. y Wilkinson J. 1980. Patología Clínica Veterinaria. Unión Tipográfica Editorial. México. 532 p.
Monge, C. y E. Salinas. 1986. El mal de montaña crónico y la salud de la población andina. Instituto Andino de Estudios en Población y Desarrollo (INANDEP), Cuaderno de Investigación N°5, 24 páginas, Lima, Perú.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
34

Monge, C., 1989. Animal adaptation to low-oxygen Andean gradient. Interciencia 14: 7.
Monge, C. y F. León-Velarde. 1991. Physiological Adaptation to High Altitude: Oxygen Transport in Mammals and Birds. Physiological Reviews. 71(4): 1135-1172
Raggi, L. 1998. Advances in the scientific knowledge of South American Camelids. Avances en Ciencia Veterinarias Vol 13(1): 3-15.
Raggi, L. 2000. El Altiplano chileno: Guía para el turismo de aventura y turismo científico. S.E. Centro Internacional de Estudios Andinos. Universidad de Chile. 20 p.
Reynafarje, C., Faura, J., Paredes, A. y Villavicencio, D. 1968. Erythrokinetics in hith-altitud-adapted animals (llama, alpaca and vicuña). Journal of Applied Physiology. 24(1): 93-97.
Reynafarje, C., Faura, J., Villavicencio, D., Curaca, A., Reynafarje, B., Oyola, L., Contreras, L., Vallenas, E. y Faura, A. 1975. Oxygen transport of hemoglobin in high-altitude animals (Camelidae). Journal of Applied Physiology. 38(5): 806-810.
Schaffer, M., T. Beiter, H.D. Becker, T.K. Hunt. 1998 Neuropeptides: Mediators of inflammation and tissue repair?. Arch. Surg. 133: 1107-1116.
Singer, D. y J. Stamler. 2005. Chemical physiology of blood flow regulation by red blood cells: The Role of Nitric Oxide and S-Nitrosohemoglobin.Annu. Rev. Physiol. 67:99–145.
Vilá, B. 1999. La importancia de la etología en la conservación y manejo de las vicuñas. Etología, 7:63-68.
Wheeler, J.C. 1991. Origen, evolución y status actual. En: Avances y Perspectivas de los Camélidos Sudamericanos. Ed. Saúl Fernández-Baca. Oficina Regional de la FAO para América Latina y el Caribe.
Zapata, B., V. Fuentes, C. Bonacic, B. González, G. Villouta y F. Bas, 2002. Heamatological and clinical biochemistry findings in captive juvenile guanacos (Lama guanicoe Mûller 1776) in central Chile. Small Ruminant Research. 2243:1-7
Van Nice, P.; C. P. Black y S. M. Tenney. 1980. A comparative study of ventilatory responses to hypoxia with reference to hemoglobin O,-affinity in llama, cat, rat, duck and goose. Comp. Biothem. Physiol. A Conzp. Physiol. 66: 347-350, 1980.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
35


CARACTERÍSTICAS DE LA FIBRA DE ALPACA HUACAYA PRODUCIDA EN LA REGIÓN ALTO ANDINA
DE HUANCAVELICA, PERÚ
1 2 2 3 1Montes, M. Quicaño, I. ; Quispe, ; Quispe Alfonso, L.
1 Universidad Pública de Navarra. 31006 Pamplona. Spain2 Desco Huancavelica. Huancavelica. Perú3 Universidad Nacional de Huancavelica. Huancavelica. PerúE:mail: [email protected]
ABSTRACT
The quality of Huacaya alpaca fibre produced in the region of Huancavelica (Peru) is described based on a sample of 203 animals belonging to eight herding communities located between 4,100 and 4,750 m above sea level. The mean fibre diameter, 22.7 ìm (SE 0.2), was lower than values reported for Huacaya alpacas from other areas and varied with sex, age, and community origin (P<0.01). In contrast with results from other studies, males had finer fibre than females, but this may be because they represent selected breeding stock brought from Puno and Cusco. No linear relationship was found between fibre diameter and staple length. Further research is needed to better characterise fibre production traits and quantify their economic values prior to establishing a breeding program to improve fibre production in the region. Although conducting this type of research in the Peruvian Andean Plateau might seem difficult, the active participation of alpaca owners and development promoters made it feasible.
KEYWORDS: Age, diameter, fibre, fibre quality, genetic selection, local development, sex.
INTRODUCCIÓN
Cerca del 90% de la población mundial de alpacas se encuentra en Perú, y el 80% del total de la producción de camélidos sudamericanos se desarrolla en las regiones andinas. Estos animales tienen gran capacidad para adaptarse a elevadas altitudes de esta región, con lugares de crianza entre 3000 y 5000 metros sobre el nivel del mar. La explotación alpaquera constituye una importante actividad económica en los Andes, y se desarrolla bajo un sistema de producción tradicional de pequeñas familias de crianza, llamados alpaqueros.
La calidad de la fibra de la alpaca producida en estas regiones ha disminuido, reduciéndose por lo tanto su valor en los mercados internacionales de fibra. Esta declinación disminución de la calidad podría ser debida a una variedad de factores tales como la presión ejercida por la industria textil con el fin de incrementar la producción de fibra blanca, el insuficiente desarrollo de tecnologías para la reproducción y mejora genética de la alpaca, las inadecuadas
; R. E. C. ;
El original de éste artículo lleva como título: Peruvian Andean Plateau region of Huancavelica., el cual se encuentra publicado en Spanish Journal of Agricultural Research 6(1):33-38.
Quality characteristics of Huacaya Alpaca fibre produced in the
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
37

prácticas de manejo y la pérdida de animales de alta calidad por la exportación de animales a otros países (legal e ilegalmente). Asimismo las alpacas, han sido cruzadas con llamas (Lama glama L.) con el fin de incrementar la cantidad de fibra producidas, a pesar que esta práctica reduce la finura de la fibra (Fairfield, 2006).
Cerca de 220,000 alpacas se localizan en el departamento de Huancavelica, que está situado al sur de la región Sierra del Perú, el cual constituye alrededor de 7% del total nacional. Más del 90% de alpacas en Huancavelica son de la raza Huacaya, caracterizada por tener una fibra compacta, fina, muy rizada y de mechones lacios (Antonini et al., 2004). Cerca del 90% de la población humana de ésta área vive en condiciones de pobreza, mientras que alrededor del 70% viven en situación de extrema pobreza. De hecho, los criadores de alpacas en el Perú se encuentran entre los más pobres de la sociedad peruana. Una de los más serios problemas de los criadores son aquellos relacionados con la comercialización de la fibra (De Los Rios, 2006). El ingreso económico de los productores alpaqueros está basado más en la cantidad que en la calidad de la fibra. Unas pocas plantas de procesamiento dominan la industria, mientras que los intermediarios entre productores y procesadores captan la mayor parte de la cadena de valor, recibiendo los criadores precios muy bajos por su fibra producida (Farfield, 2006). Consecuentemente los criadores no se encuentran motivados para mejorar las técnicas de manejo, lo que ha conllevado a un deterioro genético de la población de alpacas.
El objetivo de este estudio es caracterizar la calidad de la fibra de alpaca producida en la región de Huancavelica a fin de documentar su perfil y proveer información para futuros trabajos de mejora. El efecto del sexo, edad y comunidad sobre el diámetro de la fibra y la relación entre el diámetro de fibra y la longitud de mecha fueron analizados para determinar los factores que deberían ser considerados al diseñar un programa de mejora para la alpaca Huacaya en Huancavelica, Perú.
MATERIAL Y MÉTODOS
Localización y animales
Se tomaron muestras de fibra durante Agosto del 2006 de 203 alpacas de ocho comunidades de la región alto andina de Huancavelica en Perú (Astobamba, Cachimayo, Carhuancho, Choclocoha, Llillinta, Pilpichaca, Pucapampa y Santa Ana), ubicadas entre 4,100 y 4,750 metros sobre el nivel del mar. Los animales pertenecían a Centros de Producción de Reproductores (CPR) (El Peruano, 2005) y manejados por las correspondientes comunidades de alpaqueros
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
38

Los rebaños variaban de tamaño, desde 75 a 200 animales, y la proporción de machos variaban entre el 8 y 13%. Las prácticas de manejo fueron similar en las ocho comunidades, siguiendo las recomendaciones del calendario alpaquero (INFOCIT, 2006). Los animales se alimentaron en base a pastos naturales. Las especies predominantes, las cuales representaban más del 75% del total, pertenecían a las familias Poeacea, Cyperaceae, Astearaceae y Juncaceae.
Cada uno de los animales fueron esquilados una vez al año, usando tijeras (lapiacos), aunque en algunas comunidades también utilizaron esquiladoras mecánicas durante la última campaña del año 2006. Los productores vendían los vellones a los intermediarios, quienes generalmente compraban la fibra basados en el peso más que en la calidad. Más tarde estos agentes, categorizaban la fibra de acuerdo al color y a la calidad para la venta a la industria textil [extra fina (≤23µm); fina (>23 y 26.5 µm); semifina (>26.5 y ≤29 µm); y gruesa (>29 µm)] (NTP, 2004). Los vellones de color se categorizaban en la misma categoría como los vellones de color blanco de la peor calidad.
Las campañas de esquila generalmente ocurrían dos veces al año – en Octubre-Noviembre y en Abril-Mayo – pero los animales podían ser esquilados en diferentes fechas de acuerdo a las necesidades económicas de los alpaqueros. En este estudio los animales fueron esquilados una vez al año en dos de las comunidades (Carhuancho en Abril-Mayo y Santa en Octubre-Noviembre) y dos veces al año en las otras seis, con aproximadamente la mitad de alpacas de cada comunidad esquiladas en cada periodo.
Las muestras fueron obtenidas de machos y hembras de cuatro diferentes grupos de edad (la edad estuvo basada en el desarrollo dental): < 1.5 años (dientes de leche), 1.5 a 3 años (dos dientes), 3 a 4 años (cuatro dientes) y > 4 años (boca llena). La distribución de las muestras se indica en la Tabla 1. Con excepción de la comunidad de Cachimayo, en la cual todas las alpacas eran de color, los animales muestrados fueron de color blanco.
Las muestras de fibra fueron tomada de la zona media a la altura de la décima costilla, debido a considerarse la zona más representativa para la evaluación de la media del diámetro de fibra de alpaca (Aylan-Parker y McGregor, 2002). Utilizando una tijera se toma una muestra entre 6 y 18 gramos de fibra de ambos lados de cada animal. Estas muestras fueron colocadas y selladas en bolsas de plástico para luego ser guardadas para el posterior análisis en el Laboratorio de Fibras Textiles de la Facultad de Zootecnia de la Universidad Nacional Agraria La Molina, ubicada en Lima, Perú.
La longitud de mecha fue medida en 145 alpacas en seis de las ocho comunidades (Astobamba, Carhuancho, Choclococha, Pilpichaca, Pucapampa y Santa Ana).
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
39
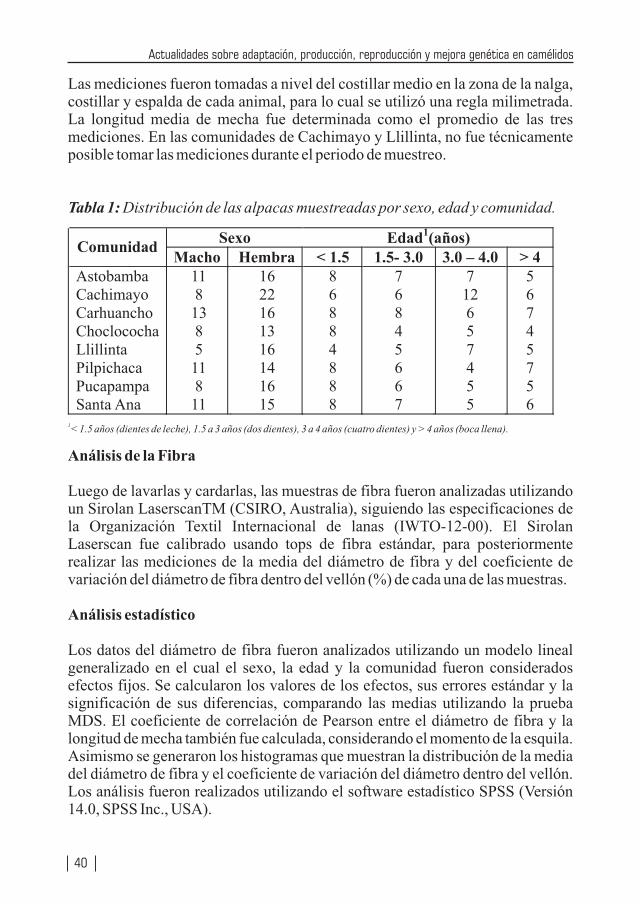
Las mediciones fueron tomadas a nivel del costillar medio en la zona de la nalga, costillar y espalda de cada animal, para lo cual se utilizó una regla milimetrada. La longitud media de mecha fue determinada como el promedio de las tres mediciones. En las comunidades de Cachimayo y Llillinta, no fue técnicamente posible tomar las mediciones durante el periodo de muestreo.
Tabla 1: Distribución de las alpacas muestreadas por sexo, edad y comunidad.
1< 1.5 años (dientes de leche), 1.5 a 3 años (dos dientes), 3 a 4 años (cuatro dientes) y > 4 años (boca llena).
Análisis de la Fibra
Luego de lavarlas y cardarlas, las muestras de fibra fueron analizadas utilizando un Sirolan LaserscanTM (CSIRO, Australia), siguiendo las especificaciones de la Organización Textil Internacional de lanas (IWTO-12-00). El Sirolan Laserscan fue calibrado usando tops de fibra estándar, para posteriormente realizar las mediciones de la media del diámetro de fibra y del coeficiente de variación del diámetro de fibra dentro del vellón (%) de cada una de las muestras.
Análisis estadístico
Los datos del diámetro de fibra fueron analizados utilizando un modelo lineal generalizado en el cual el sexo, la edad y la comunidad fueron considerados efectos fijos. Se calcularon los valores de los efectos, sus errores estándar y la significación de sus diferencias, comparando las medias utilizando la prueba MDS. El coeficiente de correlación de Pearson entre el diámetro de fibra y la longitud de mecha también fue calculada, considerando el momento de la esquila. Asimismo se generaron los histogramas que muestran la distribución de la media del diámetro de fibra y el coeficiente de variación del diámetro dentro del vellón. Los análisis fueron realizados utilizando el software estadístico SPSS (Versión 14.0, SPSS Inc., USA).
Sexo Edad1(años)Comunidad
Macho Hembra < 1.5 1.5- 3.0 3.0 – 4.0 > 4Astobamba 11 16 8 7 7 5Cachimayo 8 22 6 6 12 6Carhuancho 13 16 8 8 6 7Choclococha 8 13 8 4 5 4Llillinta 5 16 4 5 7 5Pilpichaca 11 14 8 6 4 7Pucapampa 8 16 8 6 5 5Santa Ana 11 15 8 7 5 6
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
40
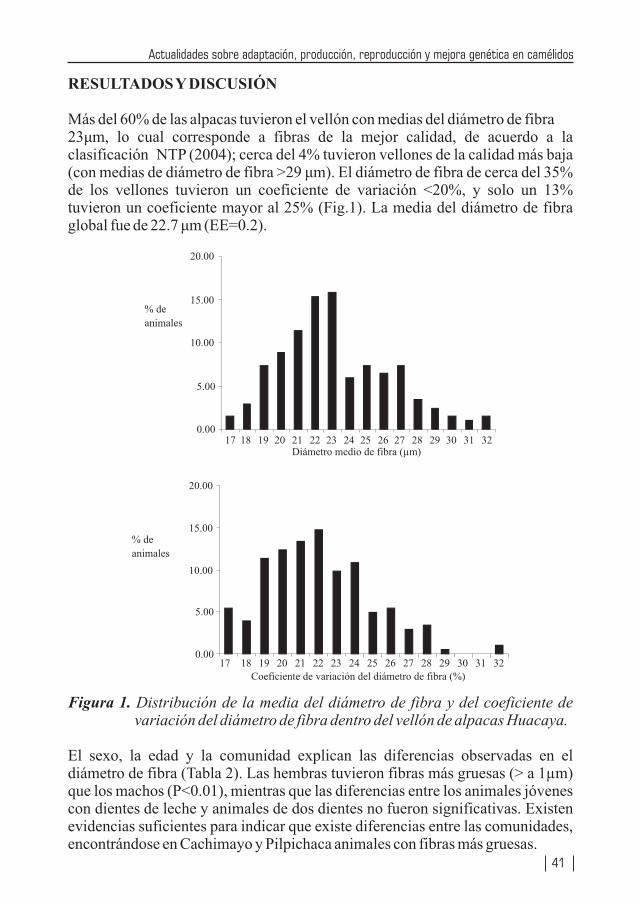
RESULTADOS Y DISCUSIÓN
Más del 60% de las alpacas tuvieron el vellón con medias del diámetro de fibra ≤23ìm, lo cual corresponde a fibras de la mejor calidad, de acuerdo a la clasificación NTP (2004); cerca del 4% tuvieron vellones de la calidad más baja (con medias de diámetro de fibra >29 ìm). El diámetro de fibra de cerca del 35% de los vellones tuvieron un coeficiente de variación <20%, y solo un 13% tuvieron un coeficiente mayor al 25% (Fig.1). La media del diámetro de fibra global fue de 22.7 ìm (EE=0.2).
Figura 1. Distribución de la media del diámetro de fibra y del coeficiente de variación del diámetro de fibra dentro del vellón de alpacas Huacaya.
El sexo, la edad y la comunidad explican las diferencias observadas en el diámetro de fibra (Tabla 2). Las hembras tuvieron fibras más gruesas (> a 1µm) que los machos (P<0.01), mientras que las diferencias entre los animales jóvenes con dientes de leche y animales de dos dientes no fueron significativas. Existen evidencias suficientes para indicar que existe diferencias entre las comunidades, encontrándose en Cachimayo y Pilpichaca animales con fibras más gruesas.
0.00
5.00
10.00
15.00
20.00
17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32Coeficiente de variación del diámetro de fibra (%)
% de
animales
0.00
5.00
10.00
15.00
20.00
17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32Diámetro medio de fibra (µm)
% de
animales
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
41
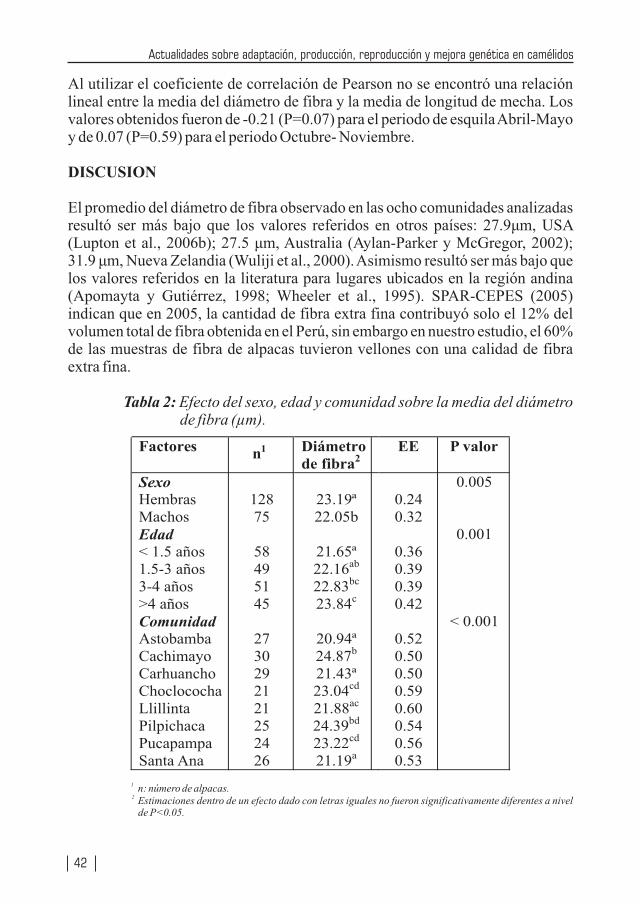
Al utilizar el coeficiente de correlación de Pearson no se encontró una relación lineal entre la media del diámetro de fibra y la media de longitud de mecha. Los valores obtenidos fueron de -0.21 (P=0.07) para el periodo de esquila Abril-Mayo y de 0.07 (P=0.59) para el periodo Octubre- Noviembre.
DISCUSION
El promedio del diámetro de fibra observado en las ocho comunidades analizadas resultó ser más bajo que los valores referidos en otros países: 27.9ìm, USA (Lupton et al., 2006b); 27.5 ìm, Australia (Aylan-Parker y McGregor, 2002); 31.9 ìm, Nueva Zelandia (Wuliji et al., 2000). Asimismo resultó ser más bajo que los valores referidos en la literatura para lugares ubicados en la región andina (Apomayta y Gutiérrez, 1998; Wheeler et al., 1995). SPAR-CEPES (2005) indican que en 2005, la cantidad de fibra extra fina contribuyó solo el 12% del volumen total de fibra obtenida en el Perú, sin embargo en nuestro estudio, el 60% de las muestras de fibra de alpacas tuvieron vellones con una calidad de fibra extra fina.
Tabla 2: Efecto del sexo, edad y comunidad sobre la media del diámetro de fibra (µm).
1 n: número de alpacas. 2 Estimaciones dentro de un efecto dado con letras iguales no fueron significativamente diferentes a nivel
de P<0.05.
Factores n1 Diámetro de fibra2
EE P valor
Sexo 0.005Hembras 128 23.19ª 0.24Machos 75 22.05b 0.32Edad 0.001< 1.5 años 58 21.65ª 0.361.5-3 años 49 22.16ab 0.393-4 años 51 22.83bc 0.39>4 años 45 23.84c 0.42Comunidad < 0.001Astobamba 27 20.94ª 0.52Cachimayo 30 24.87b 0.50Carhuancho 29 21.43ª 0.50Choclococha 21 23.04cd 0.59Llillinta 21 21.88ac 0.60Pilpichaca 25 24.39bd 0.54Pucapampa 24 23.22cd 0.56Santa Ana 26 21.19a 0.53
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
42

Comúnmente, se ha argumentado que la hibridación a gran escala entre llamas y alpacas, que ha sido realizada en el territorio peruano a fin de producir vellones más pesados, ha reducido el número de alpacas genéticamente puras, lo cual afectaría la calidad de la fibra (Kadwell et al., 2001). Como resultado de ello el diámetro de las fibras producidas por las alpacas de hoy está considerado ser significativamente más grueso que el de los animales de la antigüedad. Resultados de los análisis de momias disecadas del periodo Preincaico realizadas por Wheeler et al (1995) sugieren la existencia de dos grupos diferentes de alpacas que estuvieron diferenciados por el diámetro promedio de la fibra: un grupo de fibra extra fina (17.8µm de diámetro) y un grupo de fibra fina (23.6µm de diámetro).
La comparación de los valores de diámetro de fibra de la literatura con los obtenidos en este trabajo deben ser realizados con suma cautela debido a que el diámetro de la fibra es altamente variable. Los valores pueden verse afectados por el muestreo (Aylan-Parker y McGregor, 2002), técnicas de medición (Wiliji et al., 2000), características de los animales (por ejemplo, color; Lupton et al., 2006b) y condiciones de manejo (McGregor, 2002); y estos factores no siempre son comparables. Sin embargo, nuestros resultados indican que la fibra producida en la región de Huancavelica es de alta calidad.
El coeficiente de variación de la media del diámetro de fibra es otra característica importante del vellón porque afecta el rendimiento en el procesamiento de la fibra. En alpacas, el coeficiente de variación varía grandemente de acuerdo a la región corporal del animal (Aylan-Parker y McGregor, 2002), sin embargo diversos estudios descriptivos han mostrado similares valores medios medidos a nivel de la zona media del costillar del animal: 24.3% (DE = 4.0) (Aylan-Parker y McGregor, 2002); 23.5% (DE = 3.6) (Lupton et al., 2006b); y 23.6% (DE = 3.5) (McGregor 2006). Por lo tanto, aunque la comparación con otros resultados puede no ser fiable, la variación de la media del diámetro de fibra de las alpacas Huacaya muestreadas en nuestro estudio parece ser más baja que la obtenida en otros países. Ya que la media del diámetro de fibra (que refleja una fibra extra fina) y el bajo coeficiente de variación sugieren que la calidad de la fibra producida por las alpacas en Huancavelica es bastante buena, se vislumbra la existencia de oportunidades para mejorar los ingresos económicos de los alpaqueros de la zona, si el pago por ellas estuviese basado en la calidad de la fibra en vez de solo tomar en cuenta su peso.
Nuestros resultados indican que el sexo y la edad son dos factores endógenos que afectan significativamente el diámetro de la fibra. Asimismo hemos encontrado que los machos tienen fibras más finas que las hembras, lo cual resulta contradictorio con la idea general de que no existe diferencia entre ningún género en alpacas (Apomayta y Gutiérrez, 1998; Wuliji et al., 2000; McGregor y Butler,
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
43

2004; Frank et al., 2006 y Lupton et al., 2006b) y en otros camélidos (Martínez et al., 1997; Iñiguez et al., 1998). Contrariamente, Aylan-Parker y McGregor (2002) indicaron que existe una relación inversa con respecto al diámetro de la fibra, encontrando que los machos tienen fibras más gruesas que las hembras. La causa más probable de la diferencia de género observada en nuestro estudio es que los machos y las hembras del CPR tuvieron diferentes orígenes genéticos, pues todas las hembras que formaban parte del CPR provenían de las propias comunidades mientras que la tercera parte de los machos tenían orígenes en otras regiones del territorio peruano, y fueron introducidos por la organización no gubernamental Desco a fin de mejorar la calidad de la fibra de alpaca de la zona. Estos machos tenían procedencia principalmente de Puno y Cusco, regiones con mayor población y alta calidad de fibra de alpaca. Los machos se encontraban inscritos en los Libros Genealógicos de la Alpaca Peruana, y habían sido escogidos previamente debido a la calidad de su fibra o habían sido merecedores a premios en algunas ferias ganaderas. Pensando en términos de evaluación genética, bajo un escenario donde el proceso de selección puede ser iniciado, la diferencia entre sexos debería ser cuidadosamente analizada. La confusión entre los efectos del sexo y del origen genético podrían evitarse mediante la consideración de grupos genéticos en un modelo animal (Westell et al., 1988).
Con respecto a la edad, se encontraron diferencias en el diámetro de fibra entre animales más adultos y más jóvenes, lo cual concuerda con la idea generalmente aceptada de que el diámetro de fibra se incrementa con la edad. Sin embargo, McGregor (2002) considera que este concepto es demasiado simplista, ya que debieran tenerse en consideración otras condiciones, tales como la nutrición. Asimismo, McGregor puntualiza que cuando las alpacas pierden peso debido a condiciones nutricionales, también disminuye el crecimiento de la fibra, lo cual está asociado con la disminución del diámetro de la fibra. Lupton et al. (2006a) encontró que alpacas jóvenes que recibían un 20% más de alimento, producían más fibra, la cual tendía a ser significativamente más gruesa. En los Andes peruano, existe una creencia general sobre la relación entre la altitud y la finura de la fibra, aunque esta premisa aún no ha sido concretamente demostrada (Wheeler et al., 1995); sin embargo Braga et al., (2007) recientemente mostró que dentro de un rango altitudinal normal donde se crían las alpacas en los andes peruanos (4,100-4,700 metros), cambios debidos solo a la altitud producirían solo efectos pequeños o nulos sobre las características de la fibra de la alpaca, sin embargo la dieta podría tener una mayor influencia. En base a estas consideraciones, consideramos que la fibra fina observada en las alpacas de la región de Huancavelica podrían deberse a las condiciones pobres de los pastos naturales, más que a sus características genéticas.
Las alpacas de las comunidades Pilpichaca y Cachimayo tienen fibras significativamente más gruesas que las alpacas de las otras seis comunidades.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
44

Estas diferencias son difíciles de explicar, aunque de las ocho comunidades, Pilpichaca se ubica a una altitud mucho menor (4,100 m vs. 4,480-4750 m) pudiendo esperarse que los pastos proveerían a las alpacas mejor aporte nutricional en comparación con las otras siete comunidades. Asimismo, la proporción de machos importados a Pilpichaca de otros lugares fue mucho menor en comparación con las otras comunidades. El hecho de que las alpacas de Cachimayo fueran de color podría ser la explicación del mayor grosor de la fibra encontrado en esta comunidad, sin embargo la literatura sobre la relación entre el color del vellón y la media del diámetro de fibra es variada. Algunos estudios indican que no existen diferencias significativas entre fibras de colores (Wuliji et al., 2000), mientras que otros indican que las fibras de color oscuro son más gruesas que las fibras claras (McGregor y Butler, 2004; Lupton et al., 2006b), y Frank et al., (2006) consideran que los vellones negros son más finos que los blancos.
Aunque el valor económico de los vellones depende principalmente del diámetro de fibra, la longitud de mecha también es una característica de importancia. Con respecto a las alpacas Huacaya de nuestro estudio, no se encontró relación entre la media del diámetro de fibra y la longitud de mecha. Estos resultados no son congruentes con los de la literatura: Lupton et al., (2006b) en un estudio realizado en alpacas en USA, encontraron una correlación negativa entre estos parámetros, mientras que Wuliji et al., (2000) señalan una correlación positiva en alpacas criadas en Nueva Zelanda. De este modo, no parece ser necesario tener en consideración la relación entre diámetro de fibra y longitud de mecha en alpacas criadas con fines de producción de fibra cuando se inicie un programa de mejoramiento genético.
Estudios genéticos en alpacas sugieren que el carácter del diámetro de fibra podría ser mejorado por selección, pues cuenta con heredabilidades estimadas en un rango de 0.18 a 0.73 (Wuliji et all, 2000; León-Velarde y Guerrero, 2001; McGregor y Butler, 2004; Frank et al., 2006).Aunque los factores medioambientales que afectan el diámetro de fibra no son siempre bien entendidos y algunos parecen tener efectos contradictorios, en la región de Huancavelica, la edad y el criador comunal son factores que debieran ser considerados cuando se establezca un programa de mejora genética. Sería conveniente clarificar la relación entre el diámetro de fibra con el sexo y origen genético, por lo que parece necesario realizar más investigaciones para determinar con mayor precisión la relación entre diámetro de fibra y estos y otros factores (por ejemplo, color de vellón). Por otra parte, también parece necesario cuantificar los valores económicos relativos de los atributos de la fibra tal como se ha hecho en otros lugares (McGregor, 2006).
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
45

Aunque la realización de este tipo de trabajos de investigación en la zona alto andina peruana podría parecer dificultosa, este trabajo ha demostrado su factibilidad cuando los criadores y promotores alpaqueros participan, con el convencimiento de que las condiciones actuales pueden ser mejoradas. Pese a todo, el incentivo al precio en la venta de los vellones es con toda probabilidad un requisito esencial en un escenario a medio plazo, previo al inicio de un programa de selección en el que participen de las familias y las comunidades de alpaqueros
AGRADECIMIENTO
Nuestros agradecimientos a los alpaqueros que contribuyeron a éste estudio y a la Universidad Nacional Agraria La Molina (Laboratorio de Fibras Textiles, Facultad de Zootecnia) por los análisis de las muestras de fibra. La Universidad Pública de Navarra y el Gobierno de Navarra brindaron el apoyo financiero para la ejecución de este trabajo.
LITERATURA CITADA
Antonini M., Gonzales M., Valbonesi A., 2004. Relationship between age and postnatal skin follicular development in three types of South American domestic camelids. Livest Prod Sci 90, 241-246.
Apomayta Z.j., Gutiérrez G.a., 1998. Evaluación de las características tecnológicas y productivas de la fibra en alpacas Huacaya esquiladas a los 12 y 17 meses de edad. Anales Científicos UNALM 36, 35-42. [In Spanish].
Aylan-parker J., Mcgregor B. A., 2002. Optimising sampling techniques and estimating sampling variance of fleece quality attributes in alpacas. Small Rumin Res 44, 53-64.
Braga W., Leyva V., Cochran R., 2007. The effect of altitude on alpaca (Lama pacos) fiber production. Small Rumin Res 68, 323-328.
De Los Ríos E., 2006. Producción textil de fibras de camélidos sudamericanos en el área altoandina de Bolivia, Ecuador y Perú. United Nations Industrial Development Organization (UNIDO), Brussels. [In Spanish].
EL PERUANO, 2005. Ley de promoción del mejoramiento genético y conservación de las razas de camélidos sudamericanos domésticos, El Peruano, Normas legales, 022-2005-AG, 5 de mayo de 2005, p. 291992. [In Spanish].
Fairfield T., 2006. The politics of livestock sector policy and the rural poor in Peru. Pro-Poor Livestock Policy Initiative (PPLPI) Working Paper No. 32, FAO, Rome.
Frank E. N., Hick M. V. H., Gauna C. D., Lamas H. E., Renieri C., Antonini M., 2006. Phenotypic and genetic description of fibre traits in South American domestic camelids (llamas and alpacas). Small Rumin Res 61, 113-129.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
46

Infocit, 2006. Calendario de manejo y sanidad de alpacas en la Sierra Central del Perú. Boletín Electrónico CIT Informa, Instituto Nacional de Investigación Agraria, Perú, No. 1/2006 [online]. Available in http://www.inia.gob.pe/boletin/bcit. [May 2007] [In Spanish].
Iñiguez L. C., Alem R., Wauer A., Mueller J., 1998. Fleece types, fiber characteristics and production systems of an outstanding llama population from Southern Bolivia. Small Rumin Res 30, 57-65.
Kadwell M., Fernández M., Stanley H.F., Baldi R., Wheeler J.C., Rosario R., Bruford M.W., 2001. Genetic analysis reveals the wild ancestors of the llama and the alpaca. Proc Biol Sci 268, 2575-2584.
León - Velarde C., Guerrero J., 2001. Improving quantity and quality of Alpaca fiber; using a simulation model for breeding strategies. Proc Third Intnl Symposium on Systems Approaches for Agricultural Development International Potato Center (CIP), Lima, Peru. p. 9. [CDROM].
Lupton C. J., Elvestad R.P., Pfeiffer F. A., Mackinnon K., 2006a. Effects of age, location, and nutrition on body weight, f iber production, and fiber quality characteristics of penned alpaca males. J Anim Sci 84 (Suppl. 1), 58.
Lupton C. J., Mc Coll A., Stobart R. H., 2006b. Fiber characteristics of the Huacaya Alpaca. Small Rumin Res 64, 211-224.
Martínez Z., Iñiguez L. C., Rodríguez T., 1997. Influence of effects on quality traits and relationships between traits of the llama fleece. Small Rumin Res 24, 203-212.
Mc Gregor B. A., 2002. Comparative productivity and grazing behaviour of Huacaya alpacas and Peppin Merino sheep grazed on annual pastures. Small Rumin Res 44, 219-232.
Mc Gregor B. A., 2006. Production, attributes and relative value of alpaca fleeces in southern Australia and implications for industry development. Small Rumin Res 61, 93-111.
Mc Gregor B. A., Butler K.L., 2004. Sources of variation in fibre diameter attributes of Australian alpacas and implications for fleece evaluation and animal selection. Aus J Agric Res 55, 433-442.
NTP, 2004. Peruvian practical standards of the fibre of alpaca. Normas técnicas peruanas 231.300, 231.301. INDECOPI. [In Spanish].
SPAR-CEPES, 2005. Producción y comercialización de fibra de alpaca. Sociedad Peruana de Criadores de Alpacas y Llamas Registradas. Centro Peruano de Estudios Sociales. Miski paqu. Boletín informativo 1, 2-4. [In Spanish].
Westell R. A., Quaas R. L., Van Vleck L. D., 1988. Genetic groups in an animal model. J Dairy Sci 71, 1310-1318.
Wheeler J. C., Russel A. J. F., Redden H., 1995. Llamas and alpacas: pre-conquest breeds and post-conquest hybrids. J Archaeol Sci 22, 833-840.
Wuliji T., Davis G. H., Dodds K. G., Turner P. R., Andrews R. N., Bruce G. D., 2000. Production performance, repeatability and heritability estimates for live weight, fleece weight and fiber characteristics of alpacas in New Zealand. Small Rumin Res 37, 189-201
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
47

Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
48

AVANCES EN BIOTECNOLOGIA REPRODUCTIVA APLICADA EN LA HEMBRA DE LOS CAMELIDOS
SUDAMERICANOS
Ruiz, J. A.
PROCASUD - Universidad Nacional de HuancavelicaE-mail: [email protected]
ABSTRACT
The objective of this review is to describe actual situation of investigation in reproductive biotechnology applied in female of South American camelids. In every chapter I present briefly relevant obtained results for the different researchers who are employed at the topic. In the last years the efforts have been directed to establish protocols to improve efficiency of recovery oocytes in the live females. This oocytes have been used in the in vitro maduration and fertilization, parthenogenesis, intracytoplasmic sperm injection (ICSI) and nuclear transfer (clonation). The evaluation of several protocols of sincronization of follicular wave and the superovulation it is directed to optimizing the embryo transfer programs. All this advances are orientated to obtain a more number of birth of females of high genetics quality and to overcome the physiological limit of a birth by year.
KEY WORDS: Reproductive biotechnology, South American camelids, in vitro embryos production, embryo transfer.
INTRODUCCIÓN
La crianza de alpacas y llamas constituye una actividad económica de vital importancia para un amplio sector de la población altoandina de Perú, Bolivia, Argentina, Chile y Ecuador. Se estima que alrededor de 500 mil familias campesinas de la región andina dependen directamente de la actividad con camélidos sudamericanos (Ruiz 2005), además de otras que se benefician indirectamente de ella aprovechando los subproductos de estas especies como la fibra, carne, piel y estiércol; además la llama por su mayor tamaño y fortaleza es utilizada como animal de transporte (Cárdenas et al 1999).
En la actualidad la explotación de camélidos sudamericanos se lleva a cabo bajo sistemas tradicionales no siempre eficaces agudizando los problemas de morbilidad, mortalidad y baja eficiencia reproductiva (Cárdenas et al 1999); donde los porcentajes de natalidad anual en la mayoría de explotaciones alpaqueras es del orden del 50% (Fernández Baca 1993), con índices de fertilidad (Apaza et al 1998) y de preñez (Apaza et al 2001) que no superan el 65% y 60% respectivamente. Con un reemplazo inadecuado para la reproducción, desconocimiento de la fisiología reproductiva y un inadecuado empleo de reproductores en el apareamiento (Apaza et al 1998). La aplicación de la biotecnología reproductiva puede contribuir a superar estos bajos índices y
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
49

aportar al mejoramiento genético en estas especies, sobretodo en la multiplicación de animales de alto valor productivo mantenidos en núcleos genéticos y estos se convertirían en reproductores para los criadores de camélidos sudamericanos en nuestro país.
A pesar de esta realidad, en la actualidad existen grupos de investigación sobre la fisiología reproductiva de los camélidos sudamericanos en distintos países en el mundo, que están generando conocimientos importantes relacionados con el desarrollo folicular, la ovulación, la función del cuerpo lúteo y sobre el desarrollo embrionario. Además estos conocimientos están permitiendo la investigación en la aplicación de la biotecnología reproductiva, como la sincronización de la dinámica folicular, la inseminación artificial, la transferencia de embriones, la fecundación in vitro y, en los últimos años la producción de embriones partenogenéticos, la inyección intracitoplasmática de espermatozoides y la transferencia nuclear. Todos estos avances permiten cada día entender mejor los procesos involucrados en la función reproductiva de estas especies.
El desarrollo de biotecnologías reproductivas como la inseminación artificial, la transferencia de embriones y el desarrollo de protocolos para la fecundación in vitro, han contribuido a la mejora genética en muchas especies domésticas (Huanca et al 2002). En esta perspectiva la aplicación de biotecnologías reproductivas, podrían rápidamente incrementar el número de machos y hembras genéticamente superiores. De este modo, podría acelerarse la propagación de los núcleos de animales selectos por sus características morfológicas y morfométricas y por la cantidad y la calidad de su fibra (Miragaya et al 2006) y de este modo superar la limitante del número de crías (n ≤ 6) que una hembra puede producir durante toda su vida reproductiva (Franco et al 1999). Sin embargo, la aplicación de la biotecnología reproductiva (Adams y Ratto 2001) se encuentra en un estado formativo en especies domésticas de camélidos sudamericanos (alpaca y llama) y virtualmente no existe para las especies silvestres (vicuña y guanaco).
Por todo lo expuesto, es bastante lo que falta por investigar en la aplicación de biotecnologías reproductivas en camélidos sudamericanos y es necesaria e imprescindible para aplicarlas en beneficio de los criadores de estas especies; para el mejoramiento de la calidad de sus animales, incremento de sus niveles de producción y productividad, que conlleven a un sustancial mejoramiento de sus ingresos.
Recuperación de complejos ovocito-cumulo
Distintas metodologías se han evaluado para la recuperación de COCs (Complejos ovocito-cúmulo) en hembras de camélidos sudamericanos. Ovarios
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
50

de alpacas y llamas beneficiadas son una fuente adecuada para la recuperación de COCs, por la gran disponibilidad de ovocitos para la investigación en la estandarización de protocolos de maduración y fecundación in vitro; y para su utilización como receptores de núcleo de células donadoras en un programa de transferencia nuclear. La primera recuperación de ovocitos para su fecundación in vitro en camélidos sudamericanos se hizo en llamas; en esta investigación se desmenuzó el ovario con ayuda de una hoja de afeitar (Del Campo et al 1994) y se logró recuperar 27 ovocitos por llama descartándose el 26% de los 1324 COCs recuperados. También se han recuperado COCs de llama y alpaca puncionando folículos de ovarios recuperados en el camal con ayuda de una jeringa manual. Con esta metodología Del Campo et al (1992) recuperaron 6,4 ovocitos por llama y Ratto et al (1999a) recuperaron 2,2 COCs/ovario también en llamas con 95% (308/324) seleccionados como aptos para el cultivo en medio de maduración in vitro. Ruiz et al (2007a), utilizando ovarios procedentes del Camal Municipal de Huancavelica aspiraron folículos con agujas 21g y jeringas estériles de 5cc en el laboratorio de Biotecnología de la Universidad Nacional de Huancavelica. Se recuperaron 2,2 COCs/ovario de llama y 3,5 COCs/ovario de alpaca con 66% y 69% COCs de llama y alpaca respectivamente seleccionados como aptos para la maduración in vitro. Esta metodología de trabajo se aplicó por primera vez en el Perú para utilizar los ovocitos recuperados en la producción in vitro de embriones de alpaca y llama.
Figura 1: Lavando los ovarios de alpaca recuperados en el camal Municipal de Huancavelica.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
51
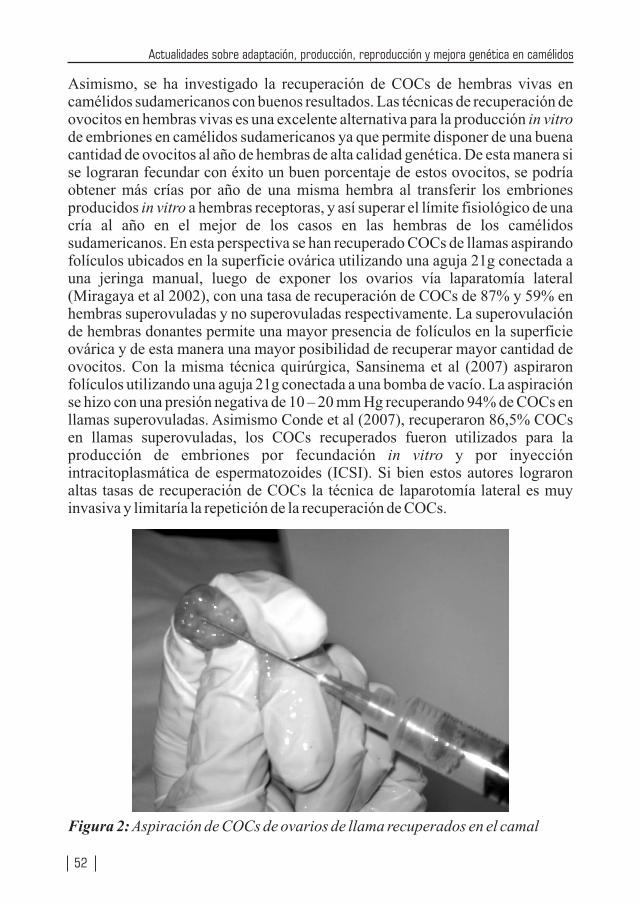
Asimismo, se ha investigado la recuperación de COCs de hembras vivas en camélidos sudamericanos con buenos resultados. Las técnicas de recuperación de ovocitos en hembras vivas es una excelente alternativa para la producción in vitro de embriones en camélidos sudamericanos ya que permite disponer de una buena cantidad de ovocitos al año de hembras de alta calidad genética. De esta manera si se lograran fecundar con éxito un buen porcentaje de estos ovocitos, se podría obtener más crías por año de una misma hembra al transferir los embriones producidos in vitro a hembras receptoras, y así superar el límite fisiológico de una cría al año en el mejor de los casos en las hembras de los camélidos sudamericanos. En esta perspectiva se han recuperado COCs de llamas aspirando folículos ubicados en la superficie ovárica utilizando una aguja 21g conectada a una jeringa manual, luego de exponer los ovarios vía laparatomía lateral (Miragaya et al 2002), con una tasa de recuperación de COCs de 87% y 59% en hembras superovuladas y no superovuladas respectivamente. La superovulación de hembras donantes permite una mayor presencia de folículos en la superficie ovárica y de esta manera una mayor posibilidad de recuperar mayor cantidad de ovocitos. Con la misma técnica quirúrgica, Sansinema et al (2007) aspiraron folículos utilizando una aguja 21g conectada a una bomba de vacío. La aspiración se hizo con una presión negativa de 10 – 20 mm Hg recuperando 94% de COCs en llamas superovuladas. Asimismo Conde et al (2007), recuperaron 86,5% COCs en llamas superovuladas, los COCs recuperados fueron utilizados para la producción de embriones por fecundación in vitro y por inyección intracitoplasmática de espermatozoides (ICSI). Si bien estos autores lograron altas tasas de recuperación de COCs la técnica de laparotomía lateral es muy invasiva y limitaría la repetición de la recuperación de COCs.
Figura 2: Aspiración de COCs de ovarios de llama recuperados en el camal
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
52
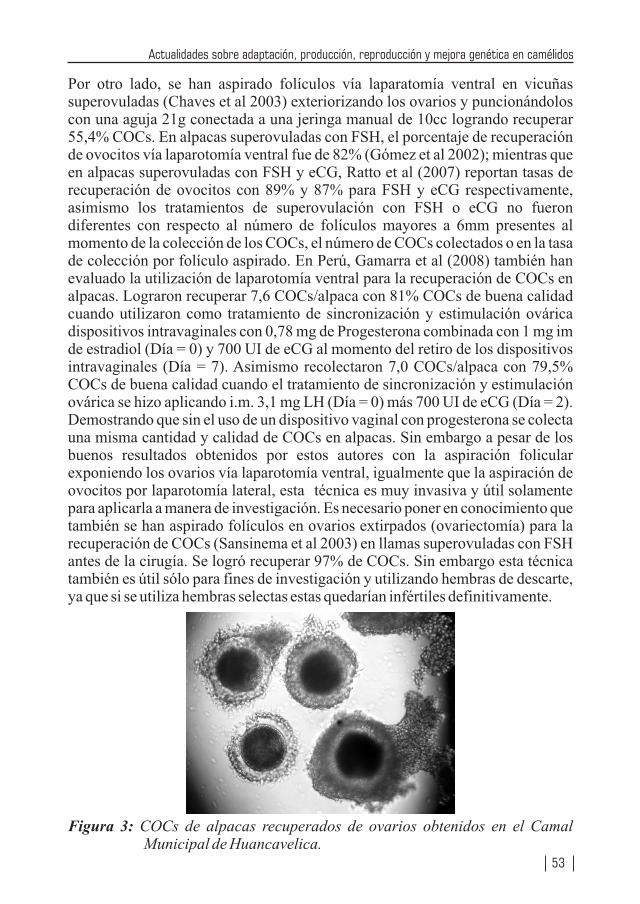
Por otro lado, se han aspirado folículos vía laparatomía ventral en vicuñas superovuladas (Chaves et al 2003) exteriorizando los ovarios y puncionándolos con una aguja 21g conectada a una jeringa manual de 10cc logrando recuperar 55,4% COCs. En alpacas superovuladas con FSH, el porcentaje de recuperación de ovocitos vía laparotomía ventral fue de 82% (Gómez et al 2002); mientras que en alpacas superovuladas con FSH y eCG, Ratto et al (2007) reportan tasas de recuperación de ovocitos con 89% y 87% para FSH y eCG respectivamente, asimismo los tratamientos de superovulación con FSH o eCG no fueron diferentes con respecto al número de folículos mayores a 6mm presentes al momento de la colección de los COCs, el número de COCs colectados o en la tasa de colección por folículo aspirado. En Perú, Gamarra et al (2008) también han evaluado la utilización de laparotomía ventral para la recuperación de COCs en alpacas. Lograron recuperar 7,6 COCs/alpaca con 81% COCs de buena calidad cuando utilizaron como tratamiento de sincronización y estimulación ovárica dispositivos intravaginales con 0,78 mg de Progesterona combinada con 1 mg im de estradiol (Día = 0) y 700 UI de eCG al momento del retiro de los dispositivos intravaginales (Día = 7). Asimismo recolectaron 7,0 COCs/alpaca con 79,5% COCs de buena calidad cuando el tratamiento de sincronización y estimulación ovárica se hizo aplicando i.m. 3,1 mg LH (Día = 0) más 700 UI de eCG (Día = 2). Demostrando que sin el uso de un dispositivo vaginal con progesterona se colecta una misma cantidad y calidad de COCs en alpacas. Sin embargo a pesar de los buenos resultados obtenidos por estos autores con la aspiración folicular exponiendo los ovarios vía laparotomía ventral, igualmente que la aspiración de ovocitos por laparotomía lateral, esta técnica es muy invasiva y útil solamente para aplicarla a manera de investigación. Es necesario poner en conocimiento que también se han aspirado folículos en ovarios extirpados (ovariectomía) para la recuperación de COCs (Sansinema et al 2003) en llamas superovuladas con FSH antes de la cirugía. Se logró recuperar 97% de COCs. Sin embargo esta técnica también es útil sólo para fines de investigación y utilizando hembras de descarte, ya que si se utiliza hembras selectas estas quedarían infértiles definitivamente.
Figura 3: COCs de alpacas recuperados de ovarios obtenidos en el Camal Municipal de Huancavelica.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
53

También se han recuperado con éxito COCs de alpaca y llama con procedimientos menos invasivos y que no requieren de una cirugía, utilizando la aspiración folicular transvaginal y la laparoscopia. Mediante la aspiración folicular transvaginal guiada por ultrasonografía Brogliatti et al (2000) alcanzaron una tasa de colección de COCs de 65% en llamas no estimuladas frente a un 52% en llamas estimuladas hormonalmente para la superovulación. Sin embargo en las llamas superestimuladas se recuperaron más ovocitos (9,0 vs 3,1) y de mayor diámetro (7,7mm vs 4,5mm). Posteriormente estos resultados han sido mejorados por Ratto et al (2005) quienes trabajaron con llamas del Centro de Investigación y Producción Quimsachata de la Estación Experimental ILLPA – INIA – Puno. Lograron recuperar 71% y 74% de COCs con 10,7 ± 2,1 y 11,2 ± 2,3 en promedio en llamas superovuladas con FSH y eCG respectivamente. En otra investigación fuera del país, Sansinema et al (2007) obtuvieron una tasa relativamente menor de recuperación con 61% de COCs en llamas superovuladas con FSH y los COCs recuperados por esta técnica fueron de inferior calidad a los colectados por laparotomía lateral. En el Perú también se ha utilizado la técnica de aspiración folicular guiada por ultrasonido para recuperar COCs en alpacas. Gamarra et al (2008) lograron recuperar en promedio 6 COCs/alpaca cuando el tratamiento para la sincronización y superovulación se hizo con 0,78 mg de progesterona + estradiol (Día = 0) y una dosis de 700 UI de eCG al momento del retiro de las esponjas con progesterona (Día = 7) y recuperaron 6,0 COCs en promedio por alpaca y cuando aplicaron otro tratamiento con LH (Día = 0) y eCG (Día = 2) recuperaron 6,1 COCs/alpaca. Los COCs de buena calidad fueron 7,4% y 64,9% con dispositivo intravaginal y sin este respectivamente. Otro grupo de investigación en el Perú encontraron resultados un tanto diferentes (Huanca et al 2006a) con un promedio de 4,5 folículos ováricos mayores a 6 mm y una tasa de recuperación de 55,5% COCs en alpacas superovuladas con un tratamiento consistente de 200 mg de FSH dividido en una aplicación dos veces/día por 4 días consecutivos realizándose la aspiración de los folículos el sexto día. Por otro lado Ruiz y Correa (2007) recuperaron COCs de alpaca y llama utilizando un laparoscopio con 30° de ángulo de cámara y con fuente de luz tipo 488B. Introdujeron 3 trócares a la cavidad abdominal a unos 6-10 cm craneales a la glándula mamaria. El primero 2 cm a la izquierda de la línea media, el segundo sobre la línea media y el tercero 4 a 6 cm a la derecha de la línea media. A través de la incisión de la izquierda se introdujo un trócar de 7 mm de diámetro a través del cual pasó el laparoscopio conectado a una fuente de luz fría. Ubicado el ovario, se introdujo por la incisión de la derecha un forceps atraumático con el cual se manipuló y sujetó el ovario. Seguidamente se introdujó por el trócar del centro una aguja conectada a una bomba de vacío con 40 mm de Hg de presión negativa con la que se aspiró el fluido folicular con los COCs. Con esta técnica se recuperaron 57% y 60% de COCs en llamas y alpacas respectivamente sin tratamientos para la superovulación.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
54

Figura 4: Aspiración laparoscópica de ovocitos en alpacas
Maduracion in vitro
Los COCs (Complejos ovocito-cúmulo) recuperados requieren de un proceso de maduración artificial en el laboratorio antes de utilizarlos en la producción in vitro de embriones. La maduración in vitro simula el tiempo transcurrido y sobretodo los procesos metabólicos que en forma natural ocurren en el ovocito contenido en el folículo dominante, entre el pico preovulatorio de LH hasta que se produce la ovulación. En este proceso se reactiva la división meiótica desde el diploteno de la profase I de la primera división hasta alcanzar la metafase II de la segunda división meiótica, momento en que el ovocito ovula en forma natural. El ovocito se mantiene en este estado hasta que es fecundado por un espermatozoide o activado artificialmente para producir embriones por partenogénesis, por ICSI (Inyección intracitoplasmática de espermatozoides), por clonación o por transgénesis. Para la maduración in vitro de ovocitos, se utilizan medios de cultivo clasificados por Gordon (1994) en medios simples y medios complejos. Un medio simple es aquel que contiene una solución salina fisiológica bufferada usualmente con bicarbonato y adicionada de piruvato, lactato y glucosa. Un medio de maduración complejo contiene, además de los constituyentes de un medio simple, aminoácidos, vitaminas, purinas y otras sustancias encontradas asociadas al suero. En bovinos el medio tradicional y exitosamente usado es el Tissue Culture Medium 199 (TCM – 199), medio complejo, compuesto por sales Earle`s con HEPES y bicarbonato, como estabilizadores de pH y suplementado con piruvato, lactato, vitaminas, aminoácidos, purinas y proteína como suero fetal bovino o albúmina sérica bovina (Rath, 2001). En camélidos sudamericanos se ha utilizado principalmente medios de maduración similares a los utilizados en bovinos.
Muy pocos estudios han sido conducidos sobre la maduración in vitro de ovocitos (Del Campo et al 1992, 1994; Sansinema et al 2003, 2007; Chaves et al 2003;
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
55

Miragaya et al 2002; Ratto et al 1999a, 2005; Ruiz y Correa 2007) en camélidos sudamericanos, sin embargo existe gran interés de diversos laboratorios en el mundo en investigar en el desarrollo in vitro de embriones de camélidos sudamericanos siendo una de las principales limitantes la disponibilidad de ovocitos: por la distancia de los centros de crianza a laboratorios equipados convenientemente para la maduración y/o fecundación in vitro, por escasos camales de camélidos domésticos al tener los criadores la costumbre del beneficio en el predio y por la prohibición del beneficio de los camélidos silvestres.
Figura 5: COCs de alpaca madurados in vitro. Nótese la expansión de las células del cúmulo.
Ovocitos de llama fueron madurados in vitro satisfactoriamente por primera vez por Del Campo et al (1992), utilizando 36 horas de maduración en un medio de cultivo de tejidos (TCM-199) suplementado con suero fetal bovino mas hormonas y obtuvieron 62% de ovocitos que alcanzaron el estadio de metafase II. En otro experimento el mismo grupo de investigación (1994) obtuvo 30% de ovocitos en metafase II luego de 30 horas de maduración in vitro. La diferencia podría explicarse por los diferentes protocolos de recuperación de ovocitos utilizados en ambos experimentos, en el primer caso se aspiraron los folículos ováricos con ayuda de una jeringa manual mientras que en el segundo caso los ovarios fueron seccionados con ayuda de una hoja de afeitar recuperando una población muy heterogénea de ovocitos desde folículos preantrales y antrales en todos los estados de desarrollo (Hyttel et al 1997). Ratto et al (1999a) utilizando un protocolo similar de maduración que en los casos anteriores mejoraron sustantivamente los resultados anteriores. En su trabajo evaluaron 26, 30 y 36 horas de maduración in vitro de COCs de llama y obtuvieron 58%, 73% y 75% de ovocitos que alcanzaron la metafase II con extrusión del primer corpúsculo polar sin diferencias estadísticas. Miragaya et al (2002) utilizaron un tiempo de 27-30 horas para la maduración in vitro de ovocitos de llama utilizando un medio algo
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
56

diferente que en los casos anteriores. El medio de maduración que utilizaron estaba compuesto por TCM 199 con bicarbonato, 10% de suero fetal bovino y glutamina; obtuvieron 62% y 74% de ovocitos en metafase II en llamas en las cuales se recuperó los COCs sin superovulación y con superovulación respectivamente. En otro estudio, Ratto et al (2005), utilizando un medio de maduración consistente de TCM-199 suplementado con piruvato de Na 0,2 mM, sulfato de gentamicina 25 µg/ml, FSH 0,5 µg/ml, estradiol 17-â 1µg/ml y suero fetal bovino al 10%, encontraron 77,7%, 80,6% y 80,4% de maduración cuando utilizaron 28, 30 y 36 horas para la maduración in vitro de ovocitos de llama no encontrando diferencias estadísticas significativas entre los 3 tiempos evaluados. Por otro lado, Sansinema et al (2003) para la maduración in vitro de ovocitos de llama incrementaron LH en su protocolo de maduración, el medio de maduración que utilizaron estaba compuesto por TCM-199, 5 ìg/ml de FSH, 10 ìg/ml de LH, 1 mg/ml de estradiol 17b y 10% de suero fetal bovino, y obtuvieron 52% de ovocitos maduros. Posteriormente el mismo grupo de investigación (Sansinema et al 2007) utilizaron un medio un tanto diferente para la maduración in vitro de ovocitos de llama consistente de TCM-199, 5 ìg/ml de FSH, 10 ìg/ml of LH, 10 ng/ml of EGF, 10 ng/ml de IGF-1 y 1ìg/ml de estradiol 17b y 10% de suero fetal bovino, mejorando su anterior tasa de maduración con 74% de ovocitos en metafase II luego de 30 horas de maduración in vitro.
También se han madurado con éxito ovocitos de vicuña y alpaca. Chaves et al (2003), reportaron la primera maduración in vitro de ovocitos de vicuña. Utilizaron un tiempo de 27 horas para la maduración en un medio similar al utilizado por Miragaya et al (2002) consistente de TCM 199 con bicarbonato, 10% de suero fetal bovino y glutamina; y obtuvieron 41% de maduración nuclear con extrusión del primer cuerpo polar y la totalidad de los ovocitos presentó maduración citoplasmática finalizado el tiempo de cultivo en medio de maduración. En alpacas, Gómez et al (2002) y Ratto et al (2007) han utilizado 26 horas para la maduración in vitro de ovocitos recuperados por laparatomía ventral. Gómez obtuvo 46% y 40% de ovocitos en metafase II en alpacas donantes superovuladas con FSH y eCG respectivamente. Ratto mejoró los resultados obtenidos por Gómez alcanzando 82% y 64% de ovocitos en metafase II para los mismos tratamientos superovulatorios en las alpacas donantes. Ruiz y Correa (2007) maduraron ovocitos de alpaca y llama durante 27 y 30 horas respectivamente en un medio compuesto por TCM-199 suplementado con piruvato de Na 0,2 mM, sulfato de gentamicina 50 µg/ml, FSH 0,02 unidades/ml, estradiol 17-â 1µg/ml y suero fetal bovino al 10%. Obtuvieron tasas de 75% y 100% de maduración de ovocitos de llama y alpaca respectivamente. Sin embargo es necesario repetir la experiencia con una mayor cantidad de ovocitos de alpaca para confirmar la alta tasa obtenida de ovocitos en metafase II.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
57
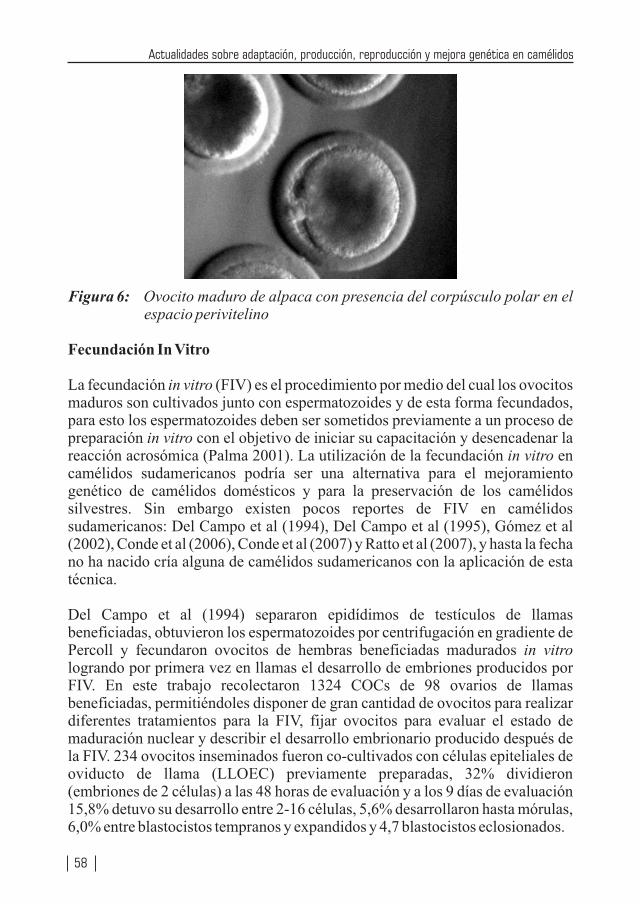
Figura 6: Ovocito maduro de alpaca con presencia del corpúsculo polar en el espacio perivitelino
Fecundación In Vitro
La fecundación in vitro (FIV) es el procedimiento por medio del cual los ovocitos maduros son cultivados junto con espermatozoides y de esta forma fecundados, para esto los espermatozoides deben ser sometidos previamente a un proceso de preparación in vitro con el objetivo de iniciar su capacitación y desencadenar la reacción acrosómica (Palma 2001). La utilización de la fecundación in vitro en camélidos sudamericanos podría ser una alternativa para el mejoramiento genético de camélidos domésticos y para la preservación de los camélidos silvestres. Sin embargo existen pocos reportes de FIV en camélidos sudamericanos: Del Campo et al (1994), Del Campo et al (1995), Gómez et al (2002), Conde et al (2006), Conde et al (2007) y Ratto et al (2007), y hasta la fecha no ha nacido cría alguna de camélidos sudamericanos con la aplicación de esta técnica.
Del Campo et al (1994) separaron epidídimos de testículos de llamas beneficiadas, obtuvieron los espermatozoides por centrifugación en gradiente de Percoll y fecundaron ovocitos de hembras beneficiadas madurados in vitro logrando por primera vez en llamas el desarrollo de embriones producidos por FIV. En este trabajo recolectaron 1324 COCs de 98 ovarios de llamas beneficiadas, permitiéndoles disponer de gran cantidad de ovocitos para realizar diferentes tratamientos para la FIV, fijar ovocitos para evaluar el estado de maduración nuclear y describir el desarrollo embrionario producido después de la FIV. 234 ovocitos inseminados fueron co-cultivados con células epiteliales de oviducto de llama (LLOEC) previamente preparadas, 32% dividieron (embriones de 2 células) a las 48 horas de evaluación y a los 9 días de evaluación 15,8% detuvo su desarrollo entre 2-16 células, 5,6% desarrollaron hasta mórulas, 6,0% entre blastocistos tempranos y expandidos y 4,7 blastocistos eclosionados.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
58

Conde et al (2006) utilizaron un sistema similar para FIV en llamas, la diferencia con Del Campo et al (1994) fue que recuperaron los gametos de animales vivos, ya que es de esta manera en que la producción de embriones por FIV puede contribuir al mejoramiento genético si se logra recuperar gametos de animales de comprobada calidad genética. Los ovocitos fueron recuperados por punción folicular de ovarios expuestos por laparatomía lateral de hembras superovuladas y los espermatozoides fueron recuperados por electroeyaculación de llamas macho. El semen fue tratado con colagenasa para reducir su viscosidad. Las tasas de división (formación de cigotos) a las 48 horas de la fecundación fueron 56% y 50% con y sin agentes capacitantes de espermatozoides (heparina, penicilamina e hipotaurina). Los mismos autores (2007) reportaron 40,8% y 42,2% de división a los dos días y 35% y 47% de blastocistos a los 8 días con y sin agentes capacitantes. Demostrando que la colagenasa no afecta la capacidad fecundante del semen y que es posible producir embriones de llama por FIV con semen fresco.
Gómez et al (2002), fecundaron con semen de epidídimo de llamas beneficiadas ovocitos de alpaca recuperados por punción de los folículos de ovarios expuestos por laparotomía ventral de hembras superovuladas. Los 5 COCs que fueron fecundados y cultivados in vitro desarrollaron hasta mórula a los 6 días de evaluación, ninguno continuó el desarrollo hasta el estadío de blastocisto. Este sería es el primer reporte de la producción de embriones híbridos de alpaca-llama después de una fecundación in vitro heteróloga, experiencia que fue repetida por Ratto et al (2007). Sin embargo, Del Campo et al (1995) indican que fecundaron in vitro ovocitos recuperados de alpacas superovuladas con semen de llama y los embriones producidos desarrollaron hasta el estadío de blastocisto expandido. No existe hasta la fecha reporte científico en el mundo de la producción de un embrión in vitro utilizando los dos gametos en alpacas. Sin embargo en el Perú más de un grupo de investigación viene trabajando en este camino y existe ya el primer reporte periodístico de la obtención de las primeras segmentaciones (embriones de dos células) de alpaca en la Universidad Nacional Agraria La Molina y es muy probable que en un tiempo muy cercano se reporten los primeros blastocistos de alpaca producidos por fecundación in vitro. La importancia de lograr blastocistos radica en que es en este estadio de desarrollo en que los embriones pueden ser transferidos para lograr una preñez exitosa.
Activación química de ovocitos
La activación es un proceso que se inicia normalmente en el ovocito al momento de ser penetrado por el espermatozoide durante la fecundación (Alberio y Zakhartchenko 2001) elevándose en forma breve y periódica los niveles del calcio intracelular (Tanaka 1997). En la mayoría de mamíferos, el ovocito recién liberado del folículo está detenido en Metafase II debido a los altos niveles
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
59

citoplasmáticos del factor promotor de la maduración (FPM). Una vez que se han fusionado el espermatozoide y el ovocito la liberación de calcio intracelular produce entre otros eventos la inactivación del FPM y el consecuente reinicio y termino de la meiosis hasta la extrusión del segundo corpúsculo polar (Williams 2002). La activación química de ovocitos se realiza con el objetivo de imitar en el ovocito los efectos que desencadena la penetración del espermatozoide para estimular el desarrollo de un embrión in vitro. La activación química se utiliza para la producción de embriones partenogenéticos; para activar embriones producidos por inyección intracitoplasmática de espermatozoides (ICSI), por transferencia nuclear (clonación) o tránsgenesis. La activación de ovocitos es una buena herramienta para evaluar la calidad de los ovocitos madurados in vitro o para evaluar la calidad de ovocitos criopreservados luego de la descongelación.
En camélidos sudamericanos existen algunos reportes sobre el uso de la activación química de ovocitos en la producción de embriones in vitro. En llamas, se ha utilizado la Ionomicina de calcio (Sansinema et al 2003, 2007; Conde et al 2006, 2007) como fuente para elevar la concentración de calcio intracelular, combinado con el cicloheximide (Sansinema et al 2003) o 6-DMAP (Conde et al 2006, 2007; y Sansinema et al 2007) como fuentes para inactivar el Factor Promotor de la Maduración (FPM) y reiniciar la meiosis. Existen diferencias en la concentración y el tiempo utilizados para la exposición de los ovocitos a la ionomicina de calcio en los autores citados. Sansinema et al (2003) expusieron los ovocitos enucleados e inyectados con fibroblastos de llama (transferencia nuclear por 5 minutos en Ionomicina de Ca 5 mM a diferencia de Conde et al (2006, 2007) que expusieron ovocitos inyectados con espermatozoides (ICSI) por 10 minutos en 0,5 µM de Ionomicina de Ca. Los embriones producidos por Conde et al desarrollaron hasta el estadío de blastocisto en cambio los embriones producidos por Sansinema et al (2003) solo uno llegó hasta mórula y 10 hasta 8-16 células. Por otro lado Sansinema et al (2007), expusieron embriones producidos por ICSI durante 5 minutos en 7 µM de Ionomicina de calcio y obtuvieron 63% de segmentación y 38% de embriones de 4-8 células. Los diferentes resultados pueden explicarse también por una posible influencia de las diferentes técnicas de micromanipulación utilizadas para la producción de embriones ICSI frente a transferencia nuclear y los distintos medios utilizados para el cultivo de los embriones. Para el caso del DMAP en la activación de ovocitos en diversas especies se prefiere utilizarlo en vez del cicloheximide porque se ha demostrado que acelera la formación de pronúcleos (Guo-Cheng et al 2005), mejora la inactivación del factor promotor de la maduración (FPM) y de un factor citostático (Wang et al 1998) lo que trae como consecuencia el reinicio de la meiosis y la extrusión del segundo cuerpo polar.
En el laboratorio de Biotecnología de la Universidad Nacional de Huancavelica (Ruiz et al 2007a), se utilizó por primera vez en el Perú y en el Mundo la
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
60
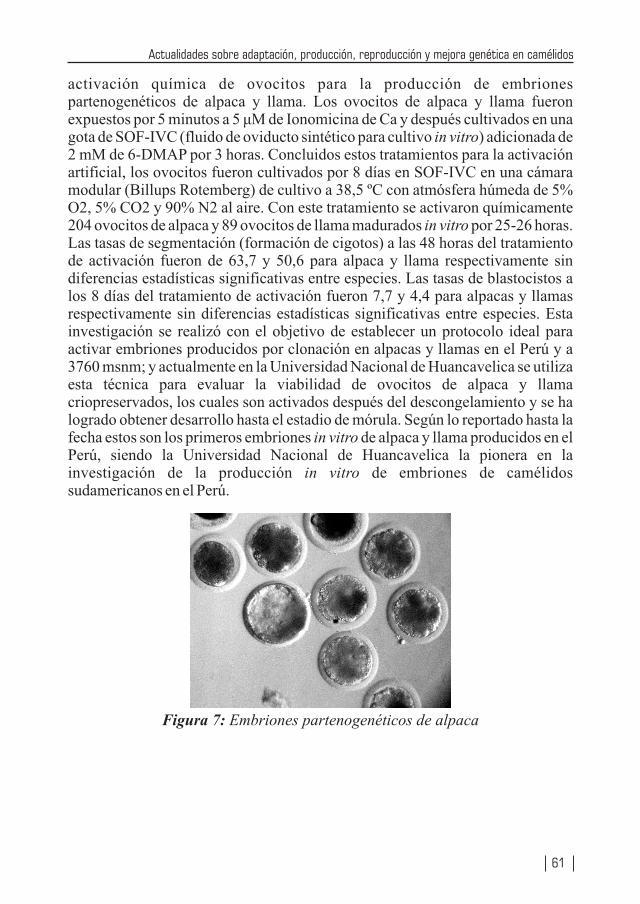
activación química de ovocitos para la producción de embriones partenogenéticos de alpaca y llama. Los ovocitos de alpaca y llama fueron expuestos por 5 minutos a 5 ìM de Ionomicina de Ca y después cultivados en una gota de SOF-IVC (fluido de oviducto sintético para cultivo in vitro) adicionada de 2 mM de 6-DMAP por 3 horas. Concluidos estos tratamientos para la activación artificial, los ovocitos fueron cultivados por 8 días en SOF-IVC en una cámara modular (Billups Rotemberg) de cultivo a 38,5 ºC con atmósfera húmeda de 5% O2, 5% CO2 y 90% N2 al aire. Con este tratamiento se activaron químicamente 204 ovocitos de alpaca y 89 ovocitos de llama madurados in vitro por 25-26 horas. Las tasas de segmentación (formación de cigotos) a las 48 horas del tratamiento de activación fueron de 63,7 y 50,6 para alpaca y llama respectivamente sin diferencias estadísticas significativas entre especies. Las tasas de blastocistos a los 8 días del tratamiento de activación fueron 7,7 y 4,4 para alpacas y llamas respectivamente sin diferencias estadísticas significativas entre especies. Esta investigación se realizó con el objetivo de establecer un protocolo ideal para activar embriones producidos por clonación en alpacas y llamas en el Perú y a 3760 msnm; y actualmente en la Universidad Nacional de Huancavelica se utiliza esta técnica para evaluar la viabilidad de ovocitos de alpaca y llama criopreservados, los cuales son activados después del descongelamiento y se ha logrado obtener desarrollo hasta el estadio de mórula. Según lo reportado hasta la fecha estos son los primeros embriones in vitro de alpaca y llama producidos en el Perú, siendo la Universidad Nacional de Huancavelica la pionera en la investigación de la producción in vitro de embriones de camélidos sudamericanos en el Perú.
Figura 7: Embriones partenogenéticos de alpaca
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
61

Figura8: Embriones partenogenéticos de llama
Inyección intracitoplasmática de espermatozoides
La inyección de espermatozoides intracitoplasmática (IEIC) o ICSI (Intra Cytoplasmic Sperm Injection) también llamada inseminación directa en el citoplasma del ovocito, inseminación intracitoplasmática directa, inseminación intracitoplasmática y microinyección dentro del citoplasma (Vilanova, 1998), permite según Schill et al (1997) la utilización de espermatozoides inmóviles (astenospermia), oligoastenospermia o provenientes del epidídimo por aspiración o del testículo por biopsia en casos de infertilidad humana. También pueden emplearse espermatozoides con graves defectos estructurales (globozoopermia, síndrome de Stump) o funcionales como la reacción del acrosoma defectuosa, baja actividad de la acrosina, fallas en la unión a la zona pelúcida y en la fusión con el ovolema o falta de recondensación nuclear (Schill et al 1997). La técnica de ICSI se basa en la inyección de una cabeza de espermatozoide aislada o el gameto entero directamente dentro del citoplasma del ovocito (Uehara y Yanagimachi 1976) con la asistencia de un micromanipulador. Una de las ventajas de la inyección de espermatozoides es que ha permitido el uso de espermatozoides inmóviles o sólo cabezas de espermatozides, ya que estos no necesitan estar vivos o intactos para llevar a cabo el proceso de fecundación (Goto et al 1990) y por lo tanto permite superar los problemas de capacitación y almacenamiento de semen (Villanueva 1997).
Esta técnica es una buena alternativa para producir embriones en camélidos sudamericanos, sobretodo cuando en la colección de semen se tiene eyaculados de pobre calidad y cuando se tiene baja motilidad después del descongelamiento del semen de alpaca y llama. Igualmente sería muy útil en el caso de tener un reproductor de probada calidad genética con problemas de fertilidad. De igual modo esta técnica así también como la fecundación in vitro (FIV) son buenas alternativas para aumentar más rápidamente los animales de mayor calidad genética, si se sabe que las hembras de los camélidos sudamericanos en el mejor
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
62

de los casos pueden tener una cría al año y menos de 6 en toda su vida reproductiva, ya que con esta técnica se puede tener varias crías al año de la misma hembra aprovechando los buenos resultados de recuperación de ovocitos de hembras vivas obtenidos por diferentes autores en llamas, alpacas y vicuñas (Brogliatti et al 2000, Miragaya et al 2002, Gómez et al 2002, Chaves et al 2003, Ratto et al 2005, Conde et al 2006, Conde et al 2007, Sansinema et al 2007, Ratto et al 2007, Ruiz y Correa 2007, Gamarra et al 2008). En llamas se han producido con diferentes resultados embriones por ICSI (Miragaya et al 2003; Conde et al 2006, 2007 y Sansinema et al 2007) aunque aún no se ha reportado el nacimiento de una cría utilizando esta técnica.
Miragaya et al (2003), utilizaron por primera vez la técnica de ICSI para producir embriones de llama y la única vez que se utilizó semen obtenido con vagina artificial para producir embriones por esta técnica. Evaluaron 2 tratamientos superovulatorios (r-hFSH y eCG) para la recuperación de ovocitos. Con r-hFSH se obtuvo 3/9 y con eCG 6/16 ovocitos que alcanzaron la metafase II mostrando un corpúsculo polar y fueron seleccionados para la técnica de ICSI. Cada ovocito fue inyectado dentro del citoplasma con un solo espermatozoide. Los ovocitos inyectados se cultivaron en medio SOFaas a 38ºC en 5% CO2 y 7% O2 al aire. Se obtuvo con r-hFSH 1/3 y con eCG 3/6 embriones y no se indicó el estadio que alcanzaron. No se utilizó un protocolo de activación química en este experimento.
Conde et al (2006, 2007), compararon el desarrollo de embriones producidos por ICSI con y sin activación química. Los embriones fueron activados con Ionomicina de Ca 0,5 µM por 10 minutos y 6-DMAP 2 mM por 3 horas cuando correspondía. El semen fue colectado por electroeyaculación y fue incubado en 25% (v/v) en una solución de colagenasa a 37 ºC para reducir su viscosidad. Las tasas de división fueron 36% y 7,4% con y sin activación química; y las tasas de blastocistos fueron 44,4% y 50,0% (blastocistos/divididos) y 16% y 3,7% (blastocistos/ovocitos) con y sin activación química. Indican que a pesar haber alcanzado una alta tasa de blastocistos con ICSI sin activación química y sin diferencias estadísticas con el tratamiento de activación química sugieren utilizar una mayor cantidad de ovocitos para determinar si realmente no hay diferencias estadísticas con y sin activación química debido al poco número de ovocitos que no recibieron estimulación química.
Sansinema et al (2007) evaluaron diferentes tratamientos de activación de embriones producidos por ICSI: A) sin activación, B) activados solo con ionomicina y C) activados con Ionomicina y 6-DMAP. Los espermatozoides fueron recuperados de la vagina de llamas hembras colocando un especulo y aspirando el eyaculado con un catéter plástico conectado a una jeringa de 20 ml después de permitir la cópula de un macho de fertilidad comprobada. Las tasas de segmentación a las 36 horas fueron 29%, 33% y 63% para A, B y C con
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
63

diferencias significativas a favor de C. Las tasas de embriones de 4-8 células después de las 48 horas fueron 7%, 13% y 38% para A, B y C con diferencias estadísticas a favor de C. Estos resultados evidencian que los embriones producidos por ICSI en llama requieren de un tratamiento de activación química.
Transferencia nuclear
La transferencia nuclear o clonación, consiste en transferir el núcleo o una célula somática entera en un ovocito enucleado con ayuda de un micromanipulador. El nuevo doblete formado se fusiona con pulsos eléctricos y se somete a un tratamiento de activación química; la nueva célula se comporta como un cigoto, es decir que se divide pasando por el estadio de mórula y puede llegar a formar un blastocisto que si es transferido a una hembra nodriza o receptora puede implantarse y desarrollar un nuevo ser hasta el nacimiento. Sin embargo la eficiencia de la transferencia nuclear en mamíferos, referida como el porcentaje de animales obtenidos respecto a la cantidad de núcleos transferidos, es muy baja aún; Montoliu (2002) indica que es menor al 2%, indicando para ovejas, vacas y ratones, 0,4, 1,4 y 1,3% respectivamente.
Figura 9: Enucleando ovocitos de alpaca con ayuda de un micromanipulador.
La producción de crías clonadas por transferencia nuclear usando células donadoras fetales o adultas se ha logrado satisfactoriamente en ovejas (Wilmut et al 1997), vacas (Cibelli et al 1998, Kato et al 1998), ratones (Wakayama et al 1998), cabras (Baguisi et al 1999), gatos (Skrzyszowska et al 2002, Shin et al 2002), caballos (Galli et al 2003) y con seguridad en otras especies más. En América Latina existen ya reportes de clonación de vacunos en Brasil (Iguma et al 2005), en Argentina el nacimiento de vacunos transgénicos productores de hormona del crecimiento e insulina en la leche (citado por Ruiz et al 2007b) y en
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
64
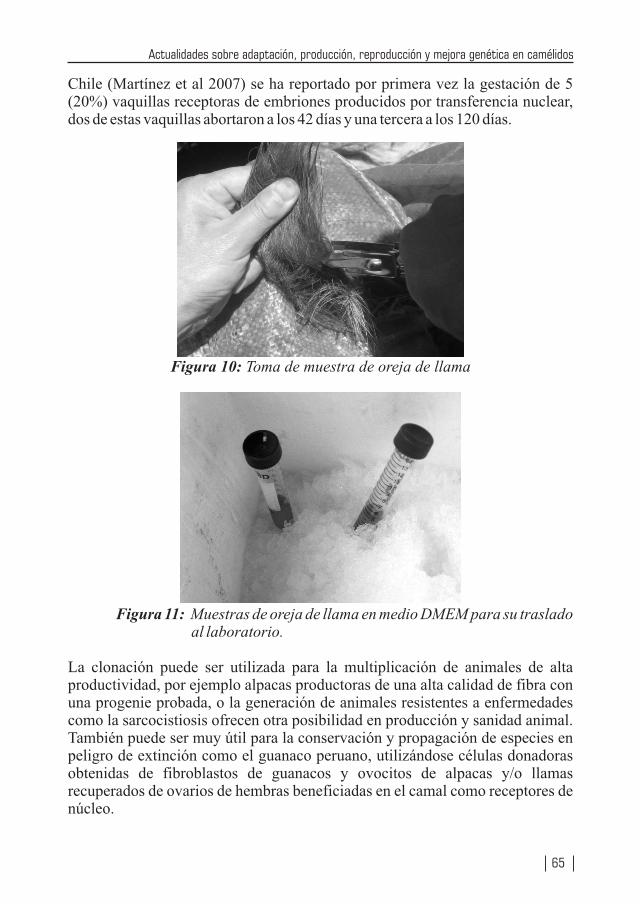
Chile (Martínez et al 2007) se ha reportado por primera vez la gestación de 5 (20%) vaquillas receptoras de embriones producidos por transferencia nuclear, dos de estas vaquillas abortaron a los 42 días y una tercera a los 120 días.
Figura 10: Toma de muestra de oreja de llama
Figura 11: Muestras de oreja de llama en medio DMEM para su traslado al laboratorio.
La clonación puede ser utilizada para la multiplicación de animales de alta productividad, por ejemplo alpacas productoras de una alta calidad de fibra con una progenie probada, o la generación de animales resistentes a enfermedades como la sarcocistiosis ofrecen otra posibilidad en producción y sanidad animal. También puede ser muy útil para la conservación y propagación de especies en peligro de extinción como el guanaco peruano, utilizándose células donadoras obtenidas de fibroblastos de guanacos y ovocitos de alpacas y/o llamas recuperados de ovarios de hembras beneficiadas en el camal como receptores de núcleo.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
65

Figura 12: Fibroblastos de alpaca en cultivo
En camélidos sudamericanos ya se hizo el primer intento de clonación, reportándose el desarrollo in vitro de embriones clonados de llama (Sansinema et al 2003). Las células donadoras se obtuvieron a partir de cultivos primarios de fibroblastos de llama. Además se recuperaron ovocitos con ayuda de una jeringa descartable de 10 ml y una aguja 20g después de la extirpación de los ovarios de llamas superovuladas para utilizarlos como receptores de los fibroblastos cultivados. Se recuperaron 298 ovocitos, de los cuales se seleccionaron 229 para la maduración in vitro y 120 (52%) ovocitos alcanzaron el estadio de metafase II. Un 70% (84/120) de ovocitos fueron enucleados satisfactoriamente con ayuda de un micromanipulador, de los cuales 95% (80/84) fueron reconstruídos con fibroblastos de llamas hembras adultas y 62,5% (50/80) fueron fusionados y activados. Se obtuvo 36% de formación de cigotos (embriones de 2-4 células) a las 48 horas de la activación química y a los 5 días de evaluación 10 embriones de 8-16 células más una mórula los cuales fueron transferidos a 6 hembras receptoras sincronizadas, ninguna hembra mostró signos de preñez a la evaluación ecográfica 14 días después de la siembra de los embriones.
Superovulacion y sincronización de la dinámica folicular
La aplicación de protocolos de ovulación múltiple y transferencia de embriones (MOET) se ha desarrollado con éxito en bovinos y ovinos con aplicación comercial en muchos países del mundo. En camélidos sudamericanos domésticos la respuesta ovárica después de tratamientos para la ovulación múltiple es ampliamente variable (Miragaya et al 2006), con una baja tasa de recuperación de embriones (Bourke et al 1995a). Debido a estos motivos la aplicación de MOET en camélidos sudamericanos todavía se encuentra en una fase de evaluación y estudio. Los tratamientos superovulatorios (Miragaya et al 2006) consisten de la administración de hormonas gonadotróficas: Gonadotropina
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
66

coriónica equina (eCG) u hormona estimulante del folículo (FSH), después de la sincronización de la onda folicular usando una fase luteal natural (inducción de la ovulación) o una fase luteal artificial (progesterona exógena). La superestimulación ovárica o superovulación consiste en inducir el crecimiento, maduración y ovulación de un gran número de folículos simultáneamente (Miragaya et al 2006). Para obtener superovulación es necesario iniciar los tratamientos hormonales en la ausencia de un folículo dominante mayor a 7 mm (Bourke et al 1995b). Iniciar el tratamiento en presencia de un folículo mayor a 5 mm induce el crecimiento de un único folículo (Miragaya et al 2006).
El primer ensayo de superovulación en alpacas donadoras fue reportado por Novoa y Sumar (1968); usaron 1200 UI de PMSG aplicada subcutáneamente en 3 días consecutivos con dosis de 400 UI cada 24 horas, después de la última inyección de PMSG las hembras recibieron vía endovenosa 750 UI de hCG e inmediatamente fueron servidas con un macho fértil. Se colectaron los embriones (3/hembra) a las 72 horas después del servicio.
Novoa et al (1998), evaluaron 2 dosis diferentes de eCG (500 y 750 UI) y dos dosis de hCG (750 y 1000 UI) y su efecto en la superovulación de alpacas. Se utilizaron 22 alpacas que al examen ovárico mediante ultrasonografía mostraron un desarrollo folicular menor de 7 mm de diámetro. Los tratamientos evaluados fueron T1: 750 UI eCG (día 0) y 750 UI hCG (día 5). T2: 500 UI eCG (día 0) y 750 UI de hCG (día 5). T3: 750 UI eCG (día 0) y 1000 UI eCG (día 5). Todas las hembras recibieron el servicio por monta natural con macho entero 2 veces por día el quinto día. Se realizó una laparatomía ventral exponiendo los ovarios para el conteo de los cuerpos lúteos y posterior recuperación de los embriones. Se contaron para T1, T2 y T3: 6,8 ± 2,6; 6,0 ± 2,8 y 12,6 ± 1,8 cuerpos lúteos respectivamente, encontrándose diferencias estadísticas entre T1 y T3, T2 y T3.
Huanca et al (1999), evaluaron la respuesta ovárica a la acción de las gonadotropinas eCG y hCG en alpacas adultas durante la época seca (agosto). Los tratamientos fueron T1: 750 UI hCG (día 0) + 300 UI eCG una vez al día por 3 días (d8, d9 y d10) + PG (d8) + 750 UI hCG (día 11). T2: eCG (300 UI) una vez al día por 3 días (d8, d9, d10) + 750 UI hCG (día 11). T3: Solución salina (d8, d9, d10). En los 3 tratamientos los animales fueron servidos 2 veces por machos fértiles. 7 días después de la monta los animales fueron sometidos a una laparotomía ventral exponiendo los ovarios para la evaluación de la respuesta ovárica. El número de cuerpos lúteos encontrados fueron: T1: 4,1 ± 1,7 y 3,6 ± 2,1; T2: 2,0 ± 2,2 y 1,6 ± 1,7; T3: 0,1 ± 0,4 y 0,1 ± 0,4 para ovario derecho e izquierdo, respectivamente (P< 0.05). El número de folículos mayores de 10 mm encontrado en la superficie ovárica fue T1: 2,3 ± 1,9 y 1,7 ± 1,5; T2: 4,4 ± 2,5 y 3,8 ± 3,1; T3: 0,7 ± 0,3 y 0,1 ± 0,4 (P<0.05). Los resultados obtenidos señalan una mejor respuesta ovulatoria en el T1 con un rango uniforme entre animales y un
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
67

mayor número de folículos mayores de 10 mm en el T2 y sugieren que el uso de hCG puede mejorar la respuesta ovárica en alpacas. Sin embargo, estos resultados difieren de lo señalado por otros autores, en lo referente a la tasa de ovulación, lo cual puede ser atribuido a un posible efecto nutricional y/o estacional.
Agüero et al (1999), compararon diferentes protocolos de superovulación en relación a la respuesta ovárica obtenida y a la cantidad de embriones recuperados. La sincronización de la onda folicular se realizó mediante la aplicación de dispositivos intravaginales (CIDR) durante 5 días. Los tratamientos fueron I (Control): sin tratamiento superovulatorio; II: una única dosis de 500 UI de eCG y III: una dosis de 500 UI de eCG y 50 mg de FSHp. Las hembras recibieron 2 servicios separados por 24 horas con una aplicación de GnRH. La recuperación embrionaria se realizó vía transcervical. La dinámica folicular fue monitoreada mediante palpación rectal y ultrasonografía. El promedio de folículos producidos fue de 2,80 ± 0,83; 5,25 ± 2,50 y 5,94 ± 1,71 para I, II y III respectivamente; el número promedio de embriones obtenidos fue 0,20 ± 0,44; 2,12 ± 2,90 y 1,94 ± 2,74 para I, II y III, respectivamente con diferencias significativas entre I frente a II y III. Concluyeron que el tratamiento II es más sencillo de implementar y económico.
Ratto et al (1999b), evaluaron la respuesta ovárica con 2 tratamientos superestimulatorios en alpacas. Las alpacas fueron tratadas con FSH (200 mg i.m. dividido en 3 días) o eCG (1200 UI im). El tratamiento superestimulatorio se inició cuando se detectó en la superficie ovárica mediante ecografía folículos de 4-5 mm de diámetro; y cuando los folículos alcanzaron un diámetro mayor a 6 mm las alpacas en ambos tratamientos recibieron 1000 UI hCG. Los COCs se colectaron aspirando folículos desde ovarios expuestos durante una laparotomía 18 – 24 horas después del tratamiento con hCG. Se encontraron en promedio por alpaca 31,8 ± 16,9 y 28,8 ± 9,9 folículos mayores a 4 mm para FSH y eCG respectivamente. Se recuperaron en promedio por alpaca 26,3 ± 14,6 y 23,3 ± 9,2 COCs para FSH y eCG respectivamente. La tasa de recuperación de COCs fue 82,7 y 82,3 para FSH y eCG respectivamente. No se encontró diferencias significativas en el número de folículos mayores a 4 mm encontrados ni en el número de COCs recuperados. Estos resultados indican que los tratamientos con FSH y eCG son buenas alternativas para ser usados en programas de superovulación y recuperación de ovocitos por aspiración folicular en alpacas.
Los objetivos para la sincronización de la dinámica folicular en camélidos sudamericanos han sido los siguientes (Miragaya et al 2006): A) Predecir la presencia de un folículo preovulatorio para programar un servicio natural o la inseminación artificial. B) Iniciar tratamientos de superestimulación ovárica en el inicio de una onda folicular en la ausencia de un folículo dominante. C)
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
68

Sincronizar hembras donadoras y receptoras en un programa de transferencia embrionaria.
Ratto et al (2003) realizaron distintos tratamientos para inducir la sincronización de la onda folicular en llamas y predecir el siguiente folículo preovulatorio. Las llamas fueron tratadas con (I) 2 ml de solución salina (control); (II) 1 mg de Estradiol 17-â y 25 mg de Progesterona en 2 ml de aceite; (III) 5 mg im LH; y (IV) Ablación de todos los folículos mayores a 5 mm con ayuda de ultrasonografía transvaginal. La emergencia de una nueva onda folicular desde el día del tratamiento fue de 5,5 ± 1,0, 4,5 ± 0,8, 2,1 ± 0,3, y 2,3 ± 0,3 días para I, II, III y IV. El nuevo folículo dominante alcanzó más de 7 mm a los 8,4 ± 2,0, 7,7 ± 0,5, 5,2 ± 0,5 y 5,0 ± 0,5 días para I, II, III y IV. Siendo más efectivos para inducir la ovulación los tratamientos con LH y la ablación de los folículos (IV) dominantes a través de aspiración folicular guiada por ultrasonido. La emergencia de una nueva onda folicular para predecir del nuevo folículo dominante se ha investigado también con tratamientos a base de progesterona y progestágenos. Ferrer et al (1999), sincronizaron la actividad ovárica con esponjas intravaginales impregnadas con Medroxiprogesterona. Durante 13 días colocaron esponjas intravaginales a llamas, divididas en los siguientes grupos: a) Control sin esponjas (4 llamas), b) Esponjas sin Medroxiprogesterona (4 llamas), c) Esponjas con 120 mg. de Medroxiprogesterona (9 llamas), esponjas con 240 mg. de Medroxiprogesterona (4 llamas) y esponjas con 480 mg de Medroxiprogesterona (8 llamas). Ninguna de las dosis utilizadas inhibió, ni sincronizó el desarrollo folicular. Sin embargo, resultados diferentes encontraron Aba et al (1999) cuando las llamas fueron tratadas por 9 días con esponjas intravaginales impregnadas con 120 mg de acetato de medroxiprogesterona (MAP). Las concentraciones de estradiol 17-â alcanzaron sus niveles más bajos 3-4 días después de la inserción de las esponjas intravaginales y alcanzaron los niveles más altos de 6 días después del retiro de las esponjas lo que se correlacionó con el desarrollo del folículo dominante.
Por otro lado Chaves et al (2002) encontraron un nuevo folículo dominante 5,0 ± 1,0 días después del retiro de dispositivos intravaginales con 0,33 gramos de progesterona (CIDR®). Llamas hembras fueron divididas en 4 tratamientos de acuerdo al tamaño folicular al momento de la inserción del CIDR: I) folículos ≤6 mm; II) folículos entre 6 y 9 mm; III) folículos entre 10 y 14 mm; y IV) folículos en regresión. Los CDIR® con progesterona se colocaron por 8 días. Durante este período se hicieron ecografías ováricas para ver el desarrollo de los folículos. En los grupos II, III y IV se observó un significativo decaimiento en el tamaño folicular después de la inserción de los dispositivos CDIR®. En el grupo I no se observó desarrollo de folículos dominantes hasta el retiro de los CDIR®. En los 4 tratamientos se observó el diámetro folicular más pequeño entre los días 5 y 7 después de la inserción de los CDIR®.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
69

Otro de los objetivos para la sincronización de la dinámica folicular en camélidos sudamericanos es iniciar tratamientos de superestimulación ovárica en el inicio de una onda folicular en la ausencia de un folículo dominante. En esta perspectiva, Bourke et al (1995c) evaluaron 2 tratamientos de superestimulación ovárica en llamas. Un tratamiento (I) aprovechando una fase luteal natural para inducir la ovulación con GnRH y otro tratamiento después de una fase luteal artificial utilizando progesterona exógena con Norgestomet. En (I): las donadoras recibieron 8 ìg im de GnRH (Día 0) para inducir ovulación de un folículo maduro, 1000 UI im eCG (Día 7) y 250 ìg de PGF2á (Día 9). En II: a las donadoras se les colocó un implante subcutáneo (Día 0) con progestágenos (3 mg Norgestomet) por 7 días y 1000 UI im de eCG (Día 5). Cuando la mayoría de los folículos (>65%) en I y II maduraron entre el día 5 al 11 después de la aplicación de eCG, las donadoras recibieron 8 ìg im de GnRH y monta natural. Más folículos (5,4 ± 0,99 y 3,3 ± 0,58) y cuerpos lúteos (5,3 ± 0,89 y 2,4 ± 0,55) se encontraron en el tratamiento II que en I sin embargo recuperaron un número similar de embriones (1,3 ± 0,25 y 1,4 ± 0,45). Aba et al (2005) han evaluado la superestimulación ovárica en vicuñas aplicando eCG luego de inducir una fase luteal artificial con progesterona exógena. TI: Dispositivos intravaginales con 0,33 g de Progesterona se insertaron en la vagina de 2 vicuñas por 5 días y al sexto día se aplicó 750 UI eCG. TII: Se monitoreaba diariamente por ecografia transrectal en 2 vicuñas hembras para confirmar la ausencia de un folículo dominante y en ese momento se aplicó 750 UI de eCG. Los animales de los 2 tratamientos fueron sometidos a cirugía exploratoria 96 horas después de la inyección con eCG, se expusieron los ovarios y se contabilizaron el número de estructuras encontrando 33 (20 folículos > 6 mm, 11 folículos < 6 mm y 2 folículos hemorrágicos) y 58 estructuras (25 folículos > 6 mm, 26 folículos < 6 mm, 3 cuerpos hemorrágicos y 4 folículos hemorrágicos) por hembra en TI y TII respectivamente. En conclusión, la progesterona liberada por el CIDR® ejerce un efecto negativo sobre desarrollo folicular y sugiere que los dispositivos intravaginales podrían ser usados para sincronizar el inicio de la onda folicular durante un tratamiento superestimulatorio. Hubo también una tendencia para un mayor desarrollo folicular cuando los animales fueron previamente tratados con progesterona.
Aller et al (2002) con el objetivo de evaluar la eficacia de eCG para inducir superovulación en vicuñas hembras previamente tratadas con progesterona y estradiol benzoato. Esponjas intravaginales con 60 mg de acetato de medroxiprogesterona fue insertado + 2 mg de estradiol benzoato administrado im (día 0) dentro de 8 vicuñas adultas. Para estimular desarrollo folicular múltiple las hembras fueron tratadas con 750 UI de eCG en el momento del retiro de las esponjas (día 7). En el día 12, las hembras recibieron una inyección intramuscular de 8 ìg GnRH análoga para inducir respuesta ovulatoria. En el día 19 cirugía
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
70

media ventral se realizó para determinar la respuesta ovárica expresada como el número de cuerpos lúteos y folículos ≥ 7 mm. La incidencia de ovulación fue de 50% (4/8) y la respuesta ovárica fue de 87,5% (7/8). En conclusión los resultados sugieren que el tratamiento con progesterona-eCG pueden usarse satisfactoriamente para inducir superovulación en llamas. Número de cuerpos
lúteos promedio 3,9 ± 4,2, número de folículos ≥ 7 mm 2,9 ± 3,3.
Aller et al (2003) evaluaron 3 tratamientos para la sincronización de la onda folicular y estimulación ovárica. TI: dispositivo intravaginal CIDR con 1.9g de progesterona colocado durante 8 días (día 0 = colocación del dispositivo). Al momento de retirar el dispositivo, se inyectó (im) 1200 UI de eCG. TII: esponja intravaginal impregnada con 150 mg de acetato de medroxiprogesterona (MAP) colocada durante 8 días y 5 mg (im) de MAP en vehículo oleoso (día 0). Al día 4, se inyectó 1200 UI de eCG. TIII esponja intravaginal impregnada con 150 mg de acetato de medroxiprogesterona (MAP) colocada durante 8 días y 5 mg de MAP + 2 mg de benzoato de estradiol (BE) en vehículo oleoso (im) (día 0). Al día 6 se inyectó 1200 UI de eCG. El número de folículos encontrado fue 4,3 ± 0,8, 2,5 ± 0,6 y 2,1 ± 0,5 para TI, TII y TIII respectivamente. El número de cuerpos lúteos encontrados fue 2.9 ± 0,5, 8,1 ± 0,9 y 7,3 ± 0,7 para TI, TII y TIII respectivamente. La respuesta ovárica total fue 7,2 ± 0,9, 10,6 ± 1,1 y 9,4 ± 0,9 para TI, TII y TIII respectivamente.
Transferencia de embriones
La transferencia de embriones puede constituir un método valioso para acelerar los programas de mejoramiento genético en camélidos sudamericanos (Franco et al 1999). Sin embargo hasta 1996 solamente 11 crías de camélidos sudamericanos habían nacido en el Mundo por transferencia de embriones (Del Campo, 1997). En el Perú, Sumar y Franco (1974) lograron la primera cría viva de alpacas obtenida por transferencia quirúrgica de embriones. En Estados Unidos, Wiepz y Chapman (1985) lograron el primer nacimiento de una llama por transferencia de embriones no quirúrgica en un ciclo estral natural sin superovulación previa de la donante. Posteriormente, en Perú nacieron 2 crías alpacas (Palomino et al, 1987); 7 crías llamas nacieron en el Reino Unido entre 1992 y 1995 (Bourke et al 1992, Bourke et al 1995b,c); y en Chile, el nacimiento de una cría llama después de 2 transferencia de embriones no quirúrgica (Gatica et al 1994). En los últimos años en criaderos particulares de llamas en Estados Unidos y Chile se ha empleado la transferencia de embriones como herramienta para obtener un mayor número de crías de buena calidad genética por hembra al año y así lograr un progreso genético más rápido (Taylor et al 2000; Von Baer et al 2003a).
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
71

En Estados Unidos durante 10 años de trabajo, Taylor (2003) indica que desde el año 1993 en un criadero particular de llamas, utilizando transferencia de embriones no quirúrgica nacieron 250 crías llamas vivas, mejorando significativamente el último año ya que hasta el año 2002 solamente 40 crías habían nacido vivas. No utilizaban tratamientos de ovulación múltiple en las llamas donantes, lavaban los úteros para la recuperación de los embriones cada 11-12 días continuamente por varios meses, recuperando un sólo embrión en cada lavado presentándose un 5% de recuperación de 2 embriones por lavado. Diariamente monitoreaban el desarrollo folicular en las receptoras mediante el uso de la ultrasonografía. La inducción de la ovulación se realizada indistintamente con LH, GnRH o por monta natural. El lavado uterino de las donantes para la recuperación de los embriones lo realizaban con un catéter Foley de 18-20 Fr. El diagnóstico de preñez lo realizaban con ultrasonografía y/o con un macho.
En Chile Von Baer et al (2003b) evaluaron la factibilidad de realizar transferencias de embriones de alpaca a receptoras llamas. Las alpacas hembras donantes fueron cubiertas naturalmente en presencia de un folículo preovulatorio entre 8 y 13mm de diámetro. Todas las receptoras llamas con un folículo entre 8 y 13 mm de diámetro recibieron una dosis de Conceptal en el mismo momento en que las donantes son cubiertas. Los embriones fueron colectados en forma no quirúrgica 7días ± 2h después de cubiertas las alpacas donantes. Todos los embriones clasificados como blastocistos eclosionados y aparentemente viables fueron transferidos antes de transcurrir las dos horas de ser colectados. La preñez se determinó por ultrasonografía 30 después de la transferencia y evaluada nuevamente al tercer mes. Encontraron tasas similares entre la transferencia de embriones de alpacas a receptoras llamas comparado con transferencias de embriones de llamas a receptoras llamas; la tasa de colección de embriones (56,9% en alpacas y 65,2 en llamas) y la tasa de preñez a los 30 y 90 días (55,2% y 51,7% respectivamente de embriones de alpacas transferidos a llamas; y 53,7% y 50,0% respectivamente de embriones de llamas transferidos a llamas) fueron similares estadísticamente. Los resultados obtenidos indican que es posible realizar transferencias de embriones de alpaca a receptoras llama, obteniéndose las mismas tasas de preñez y de crías nacidas que en el caso de transferencias dentro de la misma especie.
En Argentina, Aller et al (2003) dividieron 64 llamas hembras donantes de embriones en 3 tratamientos superovulatorios, las hembras donantes fueron servidas por monta natural o inseminación artificial, repitiendo el servicio o inseminación artificial a las 24 horas y un tercer servicio a las 12 horas del segundo. La recuperación no quirúrgica de los embriones se realizó 7,5 y 8,5 días después del primer servicio o inseminación artificial. Se obtuvieron 109 embriones luego del lavado uterino de 57 hembras donantes (1,9 ± 0,3) que
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
72

respondieron al tratamiento superovulatorio. Los tratamientos superovulatorios fueron: I) CIDR + eCG (día 8), II) esponja MAP + eCG (día 4) y III) esponja MAP + Benzoato de Estradiol + eCG (día 6) recuperando 2,0 ± 0,5, 2,2 ± 0,6 y 2,2 ± 0,4 embriones por llama y con una tasa de recuperación de embriones de 68%, 27% y 30% para I, II y III respectivamente con una ventaja significativa para el tratamiento I con CIDR + eCG. En las hembras que recibieron doble apareamiento natural (n = 39) se recuperó significativamente un mayor número promedio de embriones por llama con respecto a las hembras que recibieron doble IA (n = 8) (2,1 ± 0,3 y 0,6 ± 0,2, respectivamente). En cambio, en las que recibieron tres apareamientos (n = 10) se recuperaron 2,0 ± 0,7 embriones por llama. No encontraron diferencias significativas entre hembras apareadas dos y tres veces. Por otro lado, el mismo grupo investigador (Aller y col 2002) vitrificaron embriones de llama en una solución de vitrificación compuesta por 20% gricerol + 20% etilenglicol + 0,3M sucrosa + 0,375M glucosa + 3% polietilenglicol, luego de 2 pasajes de 5 minutos en soluciones de equilibrio para acostumbrar a los embriones a las soluciones crioprotectantes. Los embriones fueron descongelados y transferidos a llamas receptoras. Las tasas de gestación fueron 50% (2/4) y 31.3% (5/16) para receptoras que recibieron embriones vitrificados y frescos, respectivamente.
En Perú, Novoa y Sumar (1968) realizaron el primer ensayo de transferencia de embriones en alpacas, los embriones se recuperaron quirúrgicamente 3 días después de la ovulación lavando el oviducto de alpacas donantes superovuladas expuesto por laparotomía abdominal, no consiguieron nacimientos en las 3 hembras receptoras transferidas quirúrgicamente en el cuerno uterino izquierdo. Posteriormente investigadores de nuestro país han obtenido crías por transferencia de embriones en alpacas y llamas. Sumar y Franco (1974) seleccionaron 15 alpacas como donadoras de embriones, utilizaron 44 hembras receptoras de las cuales 4 quedaron preñadas y obtuvieron una cría nacida viva. Palomino et al (1987), usaron 2 alpacas como donadoras de embriones, las 3 hembras receptoras de embriones quedaron preñadas de las cuales 2 crías nacieron vivas. Palomino et al (2000), usaron 8 alpacas donantes de embriones las cuales fueron superovuladas con un tratamiento combinado de FSHp/eCG y los embriones recuperados se transfirieron a 30 alpacas receptoras de las cuales 23 quedaron preñadas y 19 crías nacieron. Huanca et al (2006) obtuvieron por transferencia de embriones transcervical 4 y 23 crías de alpacas y llamas respectivamente.
Huanca et al (2006b), monitorearon ecográficamente 15 alpacas y 16 llamas donadoras de embriones para determinar la presencia de un folículo mayor a 7 mm e iniciar el tratamiento superovulatorio. La recuperación de los embriones se realizó mediante una técnica no quirúrgica utilizando una sonda Foley de dos vías, ubicada en forma alternada en cada uno de los cuernos uterinos. 32 llamas y
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
73

10 alpacas hembras fueron preparadas como receptoras, el mismo día que se realizó la monta a las donantes e inducidas a ovulación con la aplicación de 1 ml de un análogo de GnRH. La transferencia de embriones fue realizada con embriones frescos, colocando un embrión en el cuerno ipsilateral al ovario con presencia de un cuerpo lúteo, determinado previamente por ecografía. Las 42 receptoras utilizadas fueron evaluadas por ecografía para confirmar la ovulación y determinar el lugar de ubicación del cuerpo lúteo. Obtuvieron una tasa de colección de embriones de 1,6 ± 0,1 en alpacas y 4,8 ± 0,9 en llamas, una tasa de preñez al día 90 del 40,0% (4/10) y 71,8 % (23/32) en alpacas y llamas respectivamente y una tasa de natalidad del 40,0% en alpacas y 63,6 % en llamas.
Conclusiones e implicancias
Los avances logrados en la biotecnología reproductiva aplicada en camélidos sudamericanos nos permiten mirar el futuro con optimismo. A pesar que los grupos de investigación en el mundo que trabajan en esta área son escasos comparados con otras especies como bovinos y porcinos, los logros obtenidos son significativos aunque aún es largo el camino que falta por recorrer para obtener crías de alpaca, llama, vicuña y guanaco producidas in vitro. El uso de ovarios y testículos procedentes del camal sigue siendo la fuente principal para obtener ovocitos y espermatozoides para la investigación en la producción in vitro de embriones en estas especies. A pesar de existir diversos reportes sobre la recuperación de ovocitos de hembras vivas por diversas técnicas, no se ha determinado aún cuantas veces al año se puede aspirar a una llama o alpaca donante de ovocitos, no se ha determinado que tiempo debe descansar una donante entre las sesiones de recuperación de ovocitos para no afectar la tasa de recuperación de COCs sobretodo cuando se sabe que la respuesta a la superovulación es variable en las hembras de estas especies. Aunque se entiende que las técnicas de aspiración folicular guiada por ultrasonografía y laparoscopía serían las más indicadas para obtener gametos en hembras de los camélidos sudamericanos de comprobada calidad genética por ser menos invasivas.
A pesar que se ha logrado producir embriones hasta el estadio de blastocisto en llamas por fecundación in vitro, no se logrado obtener preñeces luego de transferir los embriones a las receptoras. El reto en los siguientes años de los investigadores de la fecundación in vitro en camélidos es obtener preñeces y lograr crías de alpacas y/o llamas. Los primeros intentos para la fecundación in vitro se han realizado recuperando gametos de machos y hembras beneficiados en el camal. Sin embargo la producción de embriones recuperando gametos de machos y hembras vivos sería un gran aporte para el mejoramiento genético y también para obtener más crías de una misma hembra de alta calidad genética. No se ha reportado intentos de fecundación in vitro utilizando semen eyaculado de alpaca o llama colectado por vagina artificial este punto es otro gran reto a vencer,
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
74

debido a la dificultad del manejo del semen por la calidad del plasma seminal. La inyección intracitoplasmática de espermatozoides (ICSI) se presenta como una alternativa para fecundar ovocitos debido a que no se requiere una gran calidad de semen reflejado en altas tasas de motilidad y aun se pueden utilizar espermatozoides muertos para introducirlos en el citoplasma de los ovocitos. Con esta técnica se ha logrado también producir embriones de llamas hasta el estadio de blastocisto aunque no se ha reportado el nacimiento de una cría ni se han obtenido preñeces. La gran limitante de esta técnica es la disponibilidad de un micromanipulador y personas capacitadas para su manejo.
La clonación por transferencia nuclear es otro reto para los investigadores en la biotecnología reproductiva aplicada en camélidos sudamericanos. Existe un solo reporte de la producción de embriones en llamas por clonación con el cual no se ha logrado producir crías ni gestaciones. Los embriones producidos por esta técnica lograron un pobre desarrollo. Falta aún caracterizar el desarrollo de las células donadoras en cultivo. En experimentos no publicados realizados por nosotros se observó que los fibroblastos de alpaca y llama desarrollan más lentamente que los fibroblastos de bovinos y necesitan mucho más tiempo para lograr la confluencia. Es posible que ésta sea una respuesta fisiológica de estas especies, sin embargo los medios utilizados para el cultivo de fibroblastos de alpaca y llama fueron los mismos de un protocolo evaluado en bovinos y que estos medios no necesariamente sean los óptimos para el cultivo de fibroblastos de camélidos. Además se debe evaluar el uso de otro tipo de células como donadoras de núcleo, por ejemplo células fetales o células de la granulosa que han sido evaluados con buenos resultados para producir clones en otras especies. Falta aún evaluar el intervalo de tiempo óptimo entre la fusión y la activación química, la intensidad de voltaje para la fusión del doblete formado por un ovocito enucleado y la célula donadora, la metodología para la enucleación de los ovocitos y otros detalles más que podrían contribuir a optimizar el desarrollo de los clones producidos en camélidos. No se debe descartar la producción de embriones transgénicos en camélidos sudamericanos como una alternativa para reproducir animales de alta producción de fibra o fibra de un color determinado, o para producir animales resistente a enfermedades. Si bien aún no se ha logrado aislar algún gen de interés en camélidos se puede evaluar la producción de embriones transgénicos a través de la transfección de alguna proteína en las células donadoras de núcleo. En experimentos no publicados, nosotros logramos transfectar fibroblastos de alpaca con proteína verde fluorescente (GFP) utilizando pEGFP-N1 un plásmido bacteriano que contiene GFP. Las células de alpaca expresaron la GFP. La transfección consiste en introducir algún gen de interés contenido en un plasmidio dentro de una célula cultivada para ser donadora de núcleo.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
75

Se han evaluado diversos protocolos de sincronización de la onda folicular así como de superovulación de donantes con resultados muy variables entre los estudios y entre la respuesta de las donadoras luego de la recuperación de embriones. Esta realidad impide una alta tasa de recuperación embrionaria y hace muy costosa la técnica. Sin embargo el nacimiento de una considerable cantidad de crías por transferencia de embriones convencional en diferentes países del Mundo y en nuestro país permitiría en un futuro cercano lograr crías de embriones producidos in vitro ya sea por fecundación, ICSI, clonación o transgénesis. LITERATURA CITADA
Aba MA, MH Miragaya, MG Chaves, EF Capdevielle, B Rutter, A Agüero. 2005. Effect of exogenous progesterona and eCG treatment on follicular dynamics in vicunas (Vicugna vicugna). Anim. Repro. Sci. 86, 153-161.
Aba MA, MA Quiroga, N Auza, M Forsberg, H Kindahl. 1999. Control of ovarian activity in llamas (Lama glama) with medroxiprogesterone acetate. Reprod. Domest. Anim. 34: 471-476.
Adams G, M Ratto. 2001. Reproductive Biotechnology in South American Camelids. Memorias de la XXIV Reunión Científica Anual de la Asociación Peruana de Producción Animal. Universidad Nacional Mayor de San Marcos. Lima – Perú.
Agüero A, M Chaves, E Capdeville, A Russo. 1999. Superovulación en Llamas: Comparación de 2 tratamientos. II Congreso Mundial sobre Camélidos. Cuzco-Perú.
Alberio R, V Zakhartchenko 2001. Clonación de animales de interés zootécnico. En: Palma G (ed). Biotecnología de la Reproducción. INTA, Buenos Aires, Pp. 353-382.
Aller J, G Rebuffi, K Cancino, RH Alberio. 2003. Factores que afectan la respuesta superovulatoria y la recuperación de embriones en un programa de transferencia embrionaria en llamas (lama glama). III Congreso Mundial de Camélidos. Pp 685-691.
Aller J, G Rebuffi, K Cancino, RH Alberio. 2002. Successful transfer of vitrified llama (Lama glama) embryos. Animal Reproduction Science. 73: 121 – 127.
Apaza N, R Sapana, T Huanca, W Huanca. 2001. Inseminación Artificial en alpacas con semen fresco en comunidades campesinas. Memorias de la XIV Reunión Científica Anual de la Asociación Peruana de Producción Animal. Universidad Nacional Mayor de San Marcos. Lima – Perú.
Apaza N, U Olarte, J Málaga. 1998. Empadre controlado de alpacas Huacaya en el Banco de Germoplasma de la E.E. ILLPA Puno. Memorias de la XXI Reunión Científica Anual de la Asociación Peruana de Producción Animal. Universidad Nacional del Altiplano. Puno-Perú.
Baguishi A, E Behboodi, DT Melican, JS Pollock, MM Destrempes, C Cammuso et al. 1999. Production of goats by somatic cell nuclear transfer. Nat Biotechnol 456-461.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
76

Bourke DA, CE Kyle, TG Mc Evoy, P Young, CL Adam. 1995a. Superovulatory responses to eCG in lamas (lama glama). Theriogenology 44: 255 – 268.
Bourke DA, CE Kyle, TG McEvoy, P Young, CL Adam. 1995b. Advanced reproductive technologies in South American Camelids. In: Proceedings of the Second European Symposium on South American Camelids, pp 235-243.
Bourke DA, CE Kyle, TG McEvoy, P Young, CL Adam. 1995c. Recipient synchronization and embryo transfer in South American Camelids. Theriogenology 43: 171.
Bourke DA, CL Adam, CE Kyle, TG McEvoy, P Young. 1992. Ovulation, superovulation and embryo recovery in llamas. Proc. 12th Int Congr Anim.Reprod. The Hage, 23-27 August 1, 193-195.
Brogliatti GM, AT Palasz, H Rodriguez-Martinez, RJ Mapletoft, GP Adams. 2000. Transvaginal collection and ultrastructure of llama (Lama glama) oocytes. Theriogenology (54) 1269-1279.
Cárdenas O, T Huanca, R Sapana, V Alarcón. 1999. Avances de inseminación artificial de llamas con semen congelado. XXII Reunión Científica Anual de la Asociación Peruana de Producción Animal. Huancavelica – Perú.
Chaves M, M Miragaya, E Capdevielle, B Rutter, S Guliano, A Agüero. 2003. Maduración in vitro de oocitos de vicuña obtenidos por aspiración quirúrgica de folículos de ovarios superestimulados. III Congreso de la Asociación Latinoamericana de Especialistas en Pequeños Rumiantes y Camélidos Sudamericanos. ALEPRYCS. Viña del Mar – Chile.
Chaves MG, M Aba, A Agüero, J Egey, V Berestin, B Rutter. 2002. Ovarian follicular wave pattern and the effect of exogenous progesterone on follicular activity in non-mated llamas. Anim. Reprod. Sci. 69: 37–46.
Cibelli JB, SL Stice, PJ Golueke, JJ Kane, J Jerry, C Blackwell, et al. 1998. Cloned transgenic calves produced from nonquiescent fetal fibroblast. Science 280:1256-1258.
Conde PA, C Herrera, VL Trasorras, S Giuliano, A Director, MH Miragaya, MG Chaves, MI Carchi, D Stivale, C Quintans C, A Agüero, B Rutter, S Pasqualini. 2007. In vitro production of llama (Lama glama) embryos by IVF and ICSI with fresh semen. Animal Reproduction Science. Article in press. doi:10.1016/j.anireprosci.2007.10.004
Conde P, C Herrera, MG Chaves, SM Giuliano, A Director, VL Trasorras, M Pinto, MI Carchi, D Stivale, B Rutter, A Agüero, MH Miragaya, RS Pasqualini. 2006.
In vitro production of llama embryos by IVF or ICSI. Reproduction, Fertility and Development 19 (1) Pages 237 – 238.
Del Campo M. 1997. Reproductive Technologies in South American Camelids. In: Current Therapy in Large Animal. Theriogenology. Ed. Robert S. Younquist. Publisher W.B. Saunders Co. Chapter: 128. pp. 836-839.
Del Campo MR, C.H Del Campo, GP Adams and RJ Mapletoftt. 1995. The application of new reproductive technologies to South American Cameljds. Theriogenology 43: 21-30.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
77

Del Campo M.; C Del Campo C, M Donoso, M Berland, R Mapletoft. 1994. In vitro fertilization and development og Lama glama oocytes using epididymal spermatozoa and oviductal cell co-culture. Theriogenology 41: 1219-1229.
Del Campo MR, MX Donoso, CH Del Campo y col. 1992. In vitro maturation of Llama (Lama glama) oocytes. Proc 12th Int Cong Anim Reprod, 1992; vol 1, p 324.
Fernández Baca S. 1993. Manipulation of reproductive functions in male and female New World camelids. Ani. Reprod. Sci. 33, 307-323.
Ferrer MS, A Agüero, M Flores, B Rutter. 1999. Control de la dinámica folicular en la Llama (Lama glama) utilizando esponjas intravaginales con medroxiprogesterona. II Congreso Mundial sobre Camélidos. Cuzco-Perú.
Franco E, C Ordoñez, E Ampuero. 1999. Retorno del Celo y Fertilidad de Alpacas post-superovuladas utilizando PGF2á. Memorias de la XXIII Reunion Cientifica Anual de la Asociación Peruana de Producción Animal. Universidad Nacional de Huancavelica – Perú.
Gamarra, G., Gallegos, A., Alvarado, E., Asparrin, M. and Vivanco, W. 2008. Techniques for ovum pick-up in gonadotropin-treated alpacas. Reproduction, Fertility and Development 20, 159–160.
Galli C, I Lagutina, G Crotti, S Colleone, P Turini, N Ponderato, R Duchi, G Lazzari. 2003. Pregnancy: a cloned horse born to its daw twin. Nature 424:635.
Gatica R, M Ratto, C Schuler, M Ortiz, J Oltra, JE Correa. 1994. Cría obtenida en una llama (Lama glama) por transferencia de embriones. Agric. Tec. (Chile) 54, 68–71.
Gómez C, MH Ratto, M Berland, M Wolter, GP Adams. 2002. Superstimulatory response and oocyte collection in Alpacas. Theriogenology 57:584 (abstract).
Gordon I. 1994. Laboratory production of cattle embryos. Primera edición, CAB internacional, Cambridge. Inglaterra. 1994.
Goto K, A Kinoshita, Y Takuma, K Ogawa. 1990. Fertilization of bovine oocytes by the injection of inmobilised, killed spermatozoa. Vet. Rec. 127: 517-520.
Guo-Cheng L, H Dong, W Yan-Guanng, H Zheng-Bin, M Suo-Feng, L Xin-Yong, C Chong-Le, T Jing-He. 2005. Effects of duration, concentration, and timing of ionomycin and 6-dimethylaminopurine (6-DMAP) treatment on activation of goat oocytes. Mol Reprod Dev 71, 380-388.
Huanca W, M Ratto, Vásquez M, Cervantes M, Cordero A, Enciso M, Huanca T, Adams G. 2006a. Fertilización In Vitro En Camélidos, I: Recuperación de ovocitos vía transvaginal. Memorias de la XXIX Reunión Científica Anual de la Asociación Peruana de Producción Animal. Universidad Nacional del Centro del Perú. Huancayo.
Huanca W, M Ratto, A Cordero, A Santiani, T Huanca, O Cárdenas, G Adams. 2006b. Respuesta ovárica y transferencia de embriones en llamas y alpacas en la zona altoandina del Perú. Memorias del IV Congreso Mundial de Camélidos. Catamarca-Argentina.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
78

Huanca, W, O Cardenas, T Huanca, A Cordero. 2002. Respuesta Ovárica a la Estimulación Hormonal en Llamas Prepúberes. Memorias de la XXV Reunión Científica Anual de la Asociación Peruana de Producción Animal. Universidad Nacional Pedro Ruiz Gallo. Chiclayo – Perú.
Huanca W, O Cárdenas, A Cordero, T Huanca, R Sapana. 1999. Respuesta ovárica a gonadotropinas (eCG y hCG) en alpacas durante la época seca. II Congreso Mundial sobre Camélidos. Cuzco-Perú.
Hyttel P, T Fair, H Callesen, T Greve. 1997. Oocyte growth, capacitation and final maturation in cattle. Theriogenology 47:23–32.
Iguma LT, SF Lisauskas, EO Melo, MM Franco, I Pivatto, GR Vianna, RV Souza, MA Dode, FJ Aragao, EL Rech, R Rumpf. 2005. Development of bovine embryos reconstructed by nuclear transfer of transfected and non-transfected adult fibroblast cells. Genet Mol Res 31: 55-66.
Kato Y, T Tetsuya, Y Sotomaru, K Kurokawa, J Kato, H Doguchi et al. 1998. Eight calves cloned from somatic cells of a single adult. Science 82: 2095-2096.
Martínez Díaz M, R Gatica, JE Correa, W Eyestone. 2007. Gestaciones producidas con embriones bovinos clonados por transferencia nuclear. Arch. Med. Vet. 39, Nº 1.
Miragaya MH, MG Chaves, A Agüero. 2006. Reproductive biotechnology in South American
camelids. Small Ruminant Research. 61: 299-310.
Miragaya MH, C Herrera, C Quintans, MG Chaves, EF Capdevielle, SM Giuliano, M Pinto, J Egey, B Rutter, S Pascualini, A Agüero. 2003. Producción in vitro de embriones de llama (Lama glama) por la técnica de ICSI: resultados preliminares. Memorias del III Congreso Mundial sobre Camélidos, vol. 1, Potosí, Bolivia, pp. 267–270.
Miragaya MH, MG Chaves, EF Capdevielle, MS Ferrer, M Pinto, B Rutter, DM Neild, A Agüero. 2002. In vitro maturation of llama (Lama glama) oocytes obtained surgically using follicle aspiration. Theriogenology 57 (1): 731.
Montoliu LL. 2002. Organismos modificados genéticamente: clonación en mamíferos. XXIV Curso Internacional de Reproducción Animal. INIA. Madrid.
Novoa C, J Sumar. 1968. Colección de huevos in vivo y ensayo de transferencia en alpaca. In: Boletín Extraordinario IVITA. Univ. Nacional Mayor de San Marcos, Lima (Perú), pp. 31–34.
Novoa C, W Franco, W García, P Danilo. 1998. Dosis de Gonadotropinas (eCG y hCG), superovulación y obtención de Embriones. XXI Reunión Científica Anual de de la Asociación Peruana de Producción Animal. Puno-Perú.
Palma GA. 2001. Producción in vitro de embriones. En Palma GA. Primera edición. Biotecnología de la Reproducción. Pp 225-282. INTA Balcarce.
Palomino H, L Tabacchi, E Avila, O Li. 1987. Ensayo preliminar de transferencia de embriones en Camélidos Sud Americanos. Rev de Camélidos Sud Americanos 5: 10-17.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
79

Palomino Martorell H, 2000. Biotecnología del transplante y micromanipulación de embriones de bovinos y camélidos de los Andes. A.F.A. Editores Importadores, Peru, p. 266 (Chapter 11).
Rath D. 2001. Producción in vitro de embriones porcinos. En Palma GA. Primera edición. Biotecnología de la reproducción. Instituto Nacional de Tecnología Agropecuaria (INTA), Balcarce. Argentina.
Ratto M, C Gómez, M Berland, G Adams. 2007. Effect of ovarian superstimulation on COC collection and maturation in alpacas. Animal Reproduction Science 97: 246-256.
Ratto M, M Berland, W Huanca, J Singh, G Adams. 2005. In vitro and in vivo maturation of llama oocytes. Theriogenology 63: 2445-2457.
Ratto M, J Singh, W Huanca, G Adams. 2003. Ovarian follicular wave synchronization and pregnancy rate alter fixed-time natural mating in llamas. Theriogenology 60, 1645 – 1656.
Ratto M, M Wolter, C Gomez, M Berland, GP Adams. 1999a. In vitro maturation of lama oocytes. Libro de resúmenes. II Congreso Mundial sobre Camélidos. Cuzco-Perú.
Ratto M, C Gómez C, M Wolter, M Berland, GP Adams. 1999b.Superstimulatory response and oocyte collection in alpacas. Libro de resúmenes. II Congreso Mundial sobre Camélidos. Cuzco-Perú.
Ruiz JA 2005. Comercialización de productos y subproductos de la explotación de camélidos sudamericanos. Resumen de conferencia. IV Congreso Latinoamericano de Especialistas en Pequeños Rumiantes y Camélidos Sudamericanos. Curitiba-Brasil.
Ruiz JA, JE Correa, G Ayuque, L Landeo, M Yaranga, A Zacarías. 2007a. Producción in vitro de embriones partenogenéticos de alpaca y llama. I Simposium Internacional de Biotecnología Aplicada en Camélidos Sudamericanos. Universidad Nacional de Huancavelica. Perú.
Ruiz JA, FJ Triviño, F Van Hussel, M Martínez Díaz, JE Correa. 2007b. Posibilidades y perspectivas de la clonación y transgénesis en camélidos sudamericanos. I Simposium Internacional de Biotecnología Aplicada en Camélidos Sudamericanos. Universidad Nacional de Huancavelica. Perú.
Ruiz JA, JE Correa. 2007. Maduración in vitro de ovocitos de alpaca y llama aspirados vía laparoscópica. I Simposium Internacional de Biotecnología Aplicada en Camélidos Sudamericanos. Universidad Nacional de Huancavelica. Perú.
Sansinema M, S Taylos, P Taylor, E Schmidt, R Denniston, R Godke. 2007. In vitro production of llama (Lama glama) embryos by intracytoplasmatic sperm injection: Effect of chemical activation treatments and culture conditions. Animal Reproduction Science 99: 342-353.
Sansinema M, S Taylor, P Taylor, R Denniston, R Godke. 2003. Production of Nuclear Transfer Llama (Lama glama) Embryos From in vitro matured llama oocytes. Cloning Stem cells 5, 191-198.
Shin, T, D Kraemer, J Pryor, L Liu, J Rugila, L Howe et al. 2002. A cat cloned by nuclear transplantation. Nature 415: 859.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
80

Schill W, T Stalf, F Kohn, I Schroeder-Printzen, W Weidner, H Gips. 1997. Intracytoplasmic injection of human spermatozoa. Reprod Dom Anim 32 (1-2).
Skrzyszowska M, L Katska, B Rynska, Z Smorag, G Kania, M Pienkowsky. 2002. In vitro developmental competence of domestic cat embryos after somatic cell cloning: a preliminary report. Theriogenology 58: 1615-1621.
Sumar J, E Franco. 1974. Informe Final IVITA-La Raya. Universidad Nacional Mayor de San Marcos, Lima, Perú.
Tanaka H. 1997. Rol de la activación del oocito en la fecundación y el transplante de núcleos. AGCI e Instituto de Reproducción Animal. Universidad Austral de Chile.
Taylor S, P Taylor, A James, R Godke. 2000. Succesful commercial embryo transfer in the llama (Lama glama). Theriogenology 53: 344.
Taylor, P. 2003. Practical embryo transfer in the South American camelids. III Congreso Mundial sobre Camélidos. Potosí–Bolivia.
Uehara T, R Yanagimachi. 1976. Behavior of nuclei of testicular, caput and cauda epididymal spermatozoa injected into hamster eggs. Biol Reprod 16: 315-321.
Villanueva I. 1997. Fecundación de ovocitos bovinos por inyección de un espermatozoide en el citoplasma. Memoria de titulación. Escuela de Medicina Veterinaria. Universidad Austral de Chile.
Vilanova L. 1998. Inyección espermática intracitoplasmática. Seminario de Investigación. Programa de Doctorado en Ciencias Veterinarias. Universidad Austral de Chile.
Von Baer A, Von Baer L, Donoso M, Poblete P, Miranda H, Del Campo M. 2003a. Efecto de diferentes parámetros en un programa de transferencia de embriones en llamas (Lama glama) y alpaca (Lama pacos). III Congreso Mundial sobre Camélidos. Potosí-Bolivia.
Von Baer A, L Von Baer, X Donoso, M Del Campo. 2003b. Transferencia de Embriones entre Especies de Camélidos Sudamericanos. III Congreso de la Asociación Latinoamericana de Especialistas en Pequeños Rumiantes y Camélidos Sudamericanos. ALEPRYCS. Viña del Mar, Chile.
Wakayama T, ACF Perry, M Zuccotti, KR Johnson, R Yanagimachi. 1998. Full-term development of mice from enucleated oocytes injected with cumulus cell nuclei. Nature 394: 369-374.
Wang W, Z Macháty, L Abeydeera, R Prather, B Day.1998. Parthenogenetic activation of pig oocytes with calcium ionophore and the block to sperm penetration after activation. Biol Reprod 58, 1357-1366.
Wiepz DW, RJ Chapman. 1985. Non-surgical embryo transfer and live birth in a llama. Theriogenology. 24: 251-257.
Williams C. 2002. Signalling mechanism of mammalian oocyte activation. Human Reprod Update. 8, 313-321.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
81

Wilmut I, AE Schnieke, J Mc Whir, AJ Kind, KHS Campbell. 1997. Viable offspring derived from fetal and adult mammalian cells. Nature, 385: 810-813
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
82

REVISIÓN DE LA ESTIMACIÓN DE LOS PARÁMETROS GENÉTICOS EN ALPACAS
Gutiérrez,
Facultad de Zootecnia. Universidad Nacional Agraria La MolinaE-mail: [email protected]
ABSTRACT
The goal of this article is to make a review of genetics parameters estimates for alpaca production traits. The traits studied were live weight, raw and clean fleece weight, mean fiber diameter (FD), coefficient of variation of FD, survival from birth to weaning, and staple length. The heritability estimates ranged from medium to high for all traits but their standard errors were large. The genetics correlation estimates were low and positive but their standard errors were not reported. However, there is abundant genetic variation and hence scope for genetic improvement in fiber production and quality in alpacas population. More accurate estimates of variance components and genetic parameters for economically important traits are required for accurate genetic evaluation and development of breeding plans.
INTRODUCCIÓN
El mejoramiento de la crianza de alpacas a través del uso de la selección es de interés de los criadores y de la industria textil asociada al procesamiento de la fibra de alpaca. El criador busca mejorar sus ingresos vía el incremento de la cantidad de fibra por animal esquilado y disminuyendo el diámetro de fibra de sus vellones, y la industria busca adquirir un mayor volumen de fibra de calidad extrafina y fina, las cuales destinara a la producción de textiles de mayor valor económico en el mercado. En el presente artículo se presenta una revisión de literatura de los valores estimados de heredabilidad y correlaciones genéticas para los principales caracteres productivos de alpacas, con el fin de evaluar su uso en el diseño, implementación y evaluación de los programas, planes o proyectos de mejoramiento genético en esta especie. Los valores estimados de heredabilidad y las correlaciones genéticas son utilizados en las evaluaciones genéticas, en la predicción de la respuesta a la selección y en la elección de las estrategias de mejoramiento genético.
El índice de herencia o heredabilidad de un carácter es la proporción de la variancia fenotípica debida a la varianza aditiva. Si la heredabilidad de un carácter es 0.3 significa que la 30% de la variación fenotípica del carácter es causada por las diferencias en valor de cría entre los individuos de la población. Mayores valores de heredabilidad están asociados a mayor precisión en la evaluación genética y mayor respuesta a la selección por generación. Una reciente revisión del concepto y usos de la heredabilidad se encuentra en Visscher y colaboradores (2008).
G. A.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
83

La correlación genética es la correlación entre los valores de cría o meritos genéticos de dos caracteres medidos del mismo animal (Falconer y Mackay, 1996). Es útil para las evaluaciones genéticas cuando se desea seleccionar por varios caracteres de manera simultánea, por ejemplo en la elaboración de índices de selección, y en la estimación de las respuestas indirectas a la selección para un carácter que no se encuentra incluido en los criterios de selección. Si seleccionamos solo por diámetro de fibra, pero si existe una correlación genética de magnitud importante con el peso de vellón podemos predecir el cambio producida en esta por efecto del esquema de selección propuesto.
INFORMACIÓN Y MÉTODOS PARA LA ESTIMACIÓN DE PARÁMETROS GENÉTICOS
Para estimar los valores de heredabilidad y correlaciones genéticas, se debe partir la varianza fenotípica en sus componentes como la variancia aditiva, variancia ambiental (temporal, permanente o ambos), y las covarianzas aditivas y ambientales (Falconer y Mackay, 1996).
Los métodos de estimación de los parámetros genéticos se basan en la estimación de las covariancias aditivas entre los parientes (Lynch y Walsh, 1996). Si se cuenta con información fenotípica de los padres y las crías se empleara la regresión lineal padre(s)-progenitor, si la información de hermanos es disponible se empleara el análisis de variancia para obtener la correlación intraclase entre hermanos enteros o medios hermanos, si se cuenta con amplia información de genealogía se podrá emplear el modelo animal y usar métodos bayesianos o de máxima verosimilitud restringida (REML) para estimar los componentes de varianza.
La estimación de la heredabilidad por regresión lineal y correlación intraclase han sido descritos en la literatura (Becker, 1986; Cardellino y Rovira, 1987; Falconer y Mackay, 1996; Lynch y Walsh, 1996, Quispe y Alfonso, 2007). Ambos métodos tienen la desventaja de que se requiere ajustar los datos por efectos ambientales y no toman en cuenta el efecto de la selección. Si los factores de ajuste no han sido estimados apropiadamente o si existe apareamiento selectivo en la población, los estimadores de componentes de variancia serán sesgados.
Actualmente el uso del modelo animal y sus variantes como el modelo padre permiten el ajuste simultaneo de los datos por efectos ambientales y la estimación de los componentes de variancia (Mrode, 2005; Thompson y colaboradores, 2005). El modelo animal es un modelo mixto en el cual el valor aditivo de cada animal es considerado como un efecto aleatorio. Este modelo ajusta por efecto de la selección si la información genealógica de los animales involucrados en este proceso es incorporado en la matriz de parentesco.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
84

Existen varios métodos para la estimación de los componentes de variancia mediante el uso del modelo animal, pero los más usados en la ultima década son el método de máxima verosimilitud restringida (REML) y métodos bayesianos basados en el muestreo de Gibbs. Estos métodos han sido incorporados en diferentes programas de computo como el MTDFREML, VCE, DMU, BLUPF90, THRGIBBSF90, WOMBAT y ASREML (Druet y Ducrocq, 2006).
Además, otra ventaja del uso del modelo animal es que se puede incluir en el modelo estadístico otros efectos aleatorios al aditivo, como el efecto genético materno, ambiente permanente directo, ambiente permanente materno. Esto permitiría una mejor descomposición de la varianza fenotípica, siempre y cuando se tenga la información disponible y suficiente para la estimación de sus componentes. También, el modelo animal multicarácter es utilizado para la estimación de las covarianzas genéticas y ambientales entre diferentes caracteres, lo cual nos permite el calculo de sus respectivas correlaciones con mayor precisión.
REPORTES DE HEREDABILIDAD
Peso vivo
Los reportes de estimaciones de heredabilidades para los caracteres de peso vivo a diferentes edades varían de medio a alto (Cuadro 1). En la mayoría de los casos en que se reportan lo errores estándares estos son demasiado altos, lo que implicaría que la información fenotípica no ha sido suficiente como para ganar mayor precisión o que las metodologías empleadas no han sido las apropiadas.
En general el número de observaciones utilizadas para la estimación las heredabilidades para peso vivo son menores a 1000, y se ha utilizado los métodos de regresión padre-progenitor y de correlación intraclase entre medios hermanos para la estimación de las heredabilidades.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
85
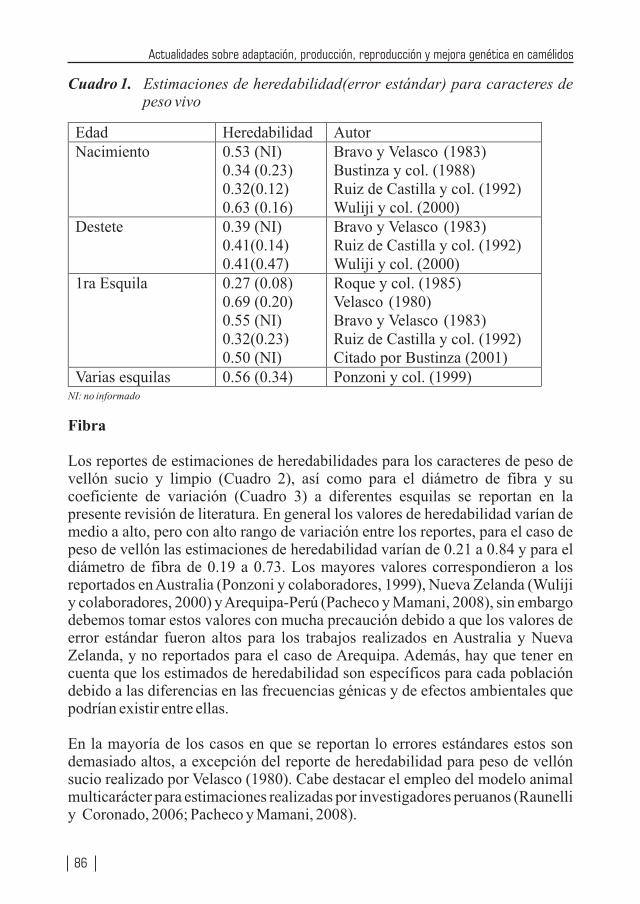
Cuadro 1. Estimaciones de heredabilidad(error estándar) para caracteres de peso vivo
NI: no informado
Fibra
Los reportes de estimaciones de heredabilidades para los caracteres de peso de vellón sucio y limpio (Cuadro 2), así como para el diámetro de fibra y su coeficiente de variación (Cuadro 3) a diferentes esquilas se reportan en la presente revisión de literatura. En general los valores de heredabilidad varían de medio a alto, pero con alto rango de variación entre los reportes, para el caso de peso de vellón las estimaciones de heredabilidad varían de 0.21 a 0.84 y para el diámetro de fibra de 0.19 a 0.73. Los mayores valores correspondieron a los reportados en Australia (Ponzoni y colaboradores, 1999), Nueva Zelanda (Wuliji y colaboradores, 2000) y Arequipa-Perú (Pacheco y Mamani, 2008), sin embargo debemos tomar estos valores con mucha precaución debido a que los valores de error estándar fueron altos para los trabajos realizados en Australia y Nueva Zelanda, y no reportados para el caso de Arequipa. Además, hay que tener en cuenta que los estimados de heredabilidad son específicos para cada población debido a las diferencias en las frecuencias génicas y de efectos ambientales que podrían existir entre ellas.
En la mayoría de los casos en que se reportan lo errores estándares estos son demasiado altos, a excepción del reporte de heredabilidad para peso de vellón sucio realizado por Velasco (1980). Cabe destacar el empleo del modelo animal multicarácter para estimaciones realizadas por investigadores peruanos (Raunelli y Coronado, 2006; Pacheco y Mamani, 2008).
Edad Heredabilidad AutorNacimiento 0.53 (NI)
0.34 (0.23)0.32(0.12)0.63 (0.16)
Bravo y Velasco (1983)Bustinza y col. (1988)Ruiz de Castilla y col. (1992)Wuliji y col. (2000)
Destete 0.39 (NI)0.41(0.14)0.41(0.47)
Bravo y Velasco (1983)Ruiz de Castilla y col. (1992)Wuliji y col. (2000)
1ra Esquila 0.27 (0.08)0.69 (0.20)0.55 (NI)0.32(0.23)0.50 (NI)
Roque y col. (1985)Velasco (1980)Bravo y Velasco (1983)Ruiz de Castilla y col. (1992)Citado por Bustinza (2001)
Varias esquilas 0.56 (0.34) Ponzoni y col. (1999)
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
86
Cuadro 2. Estimaciones de heredabilidad (error estándar) para caracteres de peso vellón sucio y limpio
NI: no informado
Cuadro 3. Estimaciones de heredabilidad (error estándar) para caracteres de diámetro de fibra y coeficiente de variación del diámetro de fibra.
NI: no informado
Sobrevivencia
Existe un reporte de heredabilidad para la sobrevivencia del nacimiento al destete, siendo el valor estimado en 0.10 y su error Standard de 0.17 (Buztinza y colaboradores, 1988).
Carácter Edad Heredabilidad Autor1ra. Esquila 0.35 (0.02)
0.22 (NI)0.21 (0.07)0.38 (0.34)0.31(0.17)0.83 (0.35)0.35 (NI)0.84 (NI)0.45 (NI)
Velasco(1980)Bravo y Velasco (1983)Roque y col. (1985)Mamani (1991)Ruiz de Castilla y col. (1992)Ponzoni y col. (1999)Raunelli y Coronado (2006)Pacheco (2008)Citado por Bustinza (2001)
Peso vellón sucio
Varias esquilas 0.63 (0.22)0.21 (NI)
Wuliji y col. (2000)Raunelli y Coronado (2006)
Peso vellónlimpio
1ra. EsquilaVarias Esquilas
0.79 (0.36)0.68 (0.22)
Ponzoni y col. (1999)Wuliji y col. (2000)
Carácter Edad Heredabilidad Autor1ra. Esquila 0.67 (0.30)
0.19 (NI)0.55 (0.42)0.32 (NI)
Ponzoni y col. (1999)Raunelli y Coronado (2006)Ayala y Chávez (2007)Pacheco y Mamani (2008)
Diámetrode fibra
Varias esquilas 0.73 (0.19)0.53 (NI)
Wuliji y col. (2000)Raunelli y Coronado (2006)
Coeficiente de variación del diámetrode fibra
1ra. EsquilaVarias esquilas
0.46 (NI)0.90 (0.30)
Pacheco y Mamani (2008)Ponzoni y col. (1999)
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
87
REPORTES DE CORRELACIONES GENÉTICAS
Son pocos los estudios que se han realizado para estimar correlaciones genéticas entre los diferentes caracteres de alpacas, en este trabajo se reporta las estimaciones de correlaciones genéticas entre el peso vivo y longitud de mecha y entre varios caracteres de producción y calidad de fibra (Cuadro 4). Los valores reportados de correlación genética son bajos, sin embargo no es posible conocer la precisión en su estimación debido a que no se ha reportado el error estándar de los valores estimados.
La mayoría de criadores de alpacas incluyen en sus objetivos de selección el incremento del peso de vellón y la disminución del diámetro de fibra y su coeficiente de variación, por lo que las correlaciones entre el peso de vellón y estos dos caracteres podría considerarse desfavorable para dicho objetivo, sin embargo la magnitud de la correlación genética es baja. La correlación genética entre el diámetro de fibra y su coeficiente de variación es favorable para el objetivo planteado. Cuadro 4. Estimaciones de correlaciones genéticas (error estándar) para
caracteres de peso vivo, producción y calidad de fibra.
NI: no informado
CONCLUSIONES
Existen reportes en la literatura de heredabilidad y correlaciones genéticas para las principales características productivas de la alpaca. Existen un mayor numero de reportes para valores de heredabilidad para el peso vivo (n=13) y peso de vellón sucio (n=11), pero pocos para caracteres de calidad de fibra como el
Características Heredabilidad AutorPeso vivo- Peso de vellón- Longitud de mecha
0.33 (NI)0.25 (NI)
Citado por Bustinza (2001)
Peso vellón 1ra. Esquila- Diámetro de fibra- Coeficiente de variación del DF
0.23 (NI)0.38 (NI)
Pacheco (2008)
Peso vellón- Longitud de mecha
0.35 (NI) Citado por Bustinza (2001)
Diámetro de fibra- Coeficiente de variación del DF
0.32 (NI) Pacheco (2008)
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
88

diámetro de fibra (n=6), coeficiente de variación del diámetro de fibra (n=2) y peso de vellón limpio (n=2) y sobrevivencia del nacimiento al destete (n=1). No se han hallado reportes de heredabilidad para otros caracteres relacionados a la fertilidad, resistencia a parásitos, susceptibilidad a enfermedades infecciosas, comportamiento, consumo de alimento, rendimiento y calidad de la carne.
Los métodos utilizados para la estimación de heredabilidad han sido el de regresión padre-progenitor y correlación intraclase entre medios hermanos para las estimaciones de heredabilidad del peso vivo y peso de vellón sucio. En cambio para los caracteres de calidad de fibra se han utilizado el modelo animal, quizás debido a la mayor disponibilidad de equipos y programas de cómputo especializados para el análisis del modelo animal y su difusión en los ambientes académicos en últimos 10 años.
En el contexto actual del mercado de fibra de alpaca se busca incrementar la frecuencia de vellones pesados, finos y uniformes en finura, siendo algunas correlaciones genéticas desfavorables al logro de este objetivo mencionado. La correlación genética positiva entre el peso de vellón y diámetro de fibra es desfavorable pero de baja magnitud, lo cual a través del uso de índices de selección podemos identificar y seleccionar a aquellos animales que tengan un mayor merito genético combinado para ambos caracteres.
Todos los estudios hallados en la literatura adolecen de haber contar con un gran numero de información fenotípica y de genealogía para la estimación de la heredabilidad y las correlaciones genéticas, restándole precisión a las estimaciones. Sin embargo los valores de heredabilidad estimados nos indicarían la existencia de una amplia variación genética para la mayoría de los caracteres estudiados, implicando una respuesta favorable a la selección.
Se requiere obtener estimaciones de parámetros genéticos con mayor precisión para los principales caracteres de importancia económica que afectan la producción de alpacas. Ello implicaría la implementación de servicios de colección de información fenotípica y genealógica, y del uso de técnicas estadísticas apropiadas para su estimación, las cuales estarían incluidas o vinculadas a los servicios de evaluación genética de los programas nacionales o planes regionales de mejoramiento genético de alpacas.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
89

LITERATURA CITADA
Ayala, J. y Chávez, J. 2007. Índice de selección genética para el diámetro de fibra de alpacas huacaya tuis y adultas. En: Conferencias Magistrales y Artículos Científicos del I Simposium Internacional de Biotecnología Aplicada en Camélidos Sudamericanos.
Becker, W.A. 1986. Manual de genética cuantitativa. Academic Enterprises.
Bravo, W. y Velasco J. 1983. Índices de herencia de pesos al nacimiento, al destete y primera esquila en alpacas. Compendio de Resúmenes de Proyectos de Investigación realizados por la UNMSM. Tomo III. Lima, Perú.
Bustinza, V. 2001. La Alpaca: conocimiento del gran potencial andino. Universidad Nacional del Altiplano, Puno, Perú.
Bustinza, A. V.; Burfening, P. J. y Blackwell, R. L. 1988. Factors Affecting Survival in Young Alpacas (Lama pacos). J. Anim Sci. 66: 1139-1143
Cardellino, R. y Rovira, J. 1987. Mejoramiento genético animal. Ed. Hemisferio Sur.
Chávez, J. 1991. Mejoramiento genético de alpacas y llamas. En: Avances y perspectivas del conocimiento de los camélidos sudamericanos. Fernández-Baca, S. (ed). p 149-190. FAO. Santiago, Chile.
Druet, T. y Ducrocq, V. 2006. Innovations in software packages in quantitative genetics. En: Proceedings of the 8th World Congress on Genetics Applied to Livestock Producción, Belo Horizonte, Brasil.
Falconer, D.S. y Mackay, T.F.C. 1996. Introduction to quantitative genetics. Fourth Edition. Prentice Hall.
Lynch, M. y Walsh, B. 1997. Genetics and analysis of quantitative traits. Sinauer Associates, Inc.
Mamani, G. 1991. Parámetros genéticos del peso vivo y vellón en alpacas Wacaya de la Raya, Puno. Resúmenes de la VII Convención Internacional de Especialistas en Camélidos Sudamericanos. Jujuy, Argentina.
Mrode, R.A. 2005. Linear models for the prediction of animal breeding values. Second Edition. CABI Publishing.
Pacheco, C. y Mamani, N. 2008. Aplicación práctica del programa de mejoramiento genético en Camélidos Domésticos. Conferencia presentada en el Simposio Internacional “Estrategias de Mejoramiento Genético en Camélidos Sudamericanos Domésticos”, Arequipa, Perú.
Ponzoni, R.W.; Grimson, R.J.; Hill, J.A.; Hubbard, D.J.; McGregor, B.A.; Howse, A.; Carmichael, I. y Judson, G.J. 1999. The inheritance of and association among some production traits in young Australian alpacas. In: Proceedings of the Association for the Advancement of Animal Breeding and Genetics 13:468-471.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
90

Quispe, E. C. y Alfonso, L. 2007. Metodologías para estimar los valores de cría (EVC). Universidad Nacional de Huancavelica.
Raunelli, J. y Coronado J. 2006. Un método de selección aplicable en alpacas. En: Libro de ponencias, resúmenes y posters del II Simposium Internacional de investigación sobre camélidos sudamericanos. Arequipa, Perú.
Roque, J.; Carpio, M. y Blackwell, R. 1985. Transmisión hereditaria de peso vivo y longitud de mecha en alpacas. En resúmenes de la V Convención Internacional sobre Camélidos Sudamericanos. Cuzco, Perú.
Ruiz de Castilla, M.; Alagón, G. y Quirita, C.R. 1992. Estudio de parámetros genéticos en alpacas huacaya. En: Informe de trabajos de investigación en alpacas y llamas de color, 2:1-29.
Thompson, R.; Brotherstone, S. y I.A.M. White. 2005. Estimation of quantitative genetic parameters. Philosophical transactions of the Royal Society B, 360:1469-1477.
Velasco, J. 1980. Heredabilidades y correlaciones de peso corporal y peso de vellón en alpacas. En: Anales de la III Reunión de la Asociación Peruana de Producción Animal. Lima, Perú.
Visscher, P.M.; Hill W.G. y N.R. Wray. 2008. Heritability in the genomics era-concepts and misconceptions. Nature Review Genetics, 19:255-266.
Wuliji T.; Davis G.H.; Dodds K.G.; Turner P.R.; Andrews R.N. y G.D. Bruce. 2000. Production performance, repeatability and heritability estimates for live weight, fleece weight and fiber characteristics of alpacas in New Zealand. Small Ruminant Research, 37:189-201
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
91

Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
92

BASES PARA ESTABLECER UN PROGRAMA DE MEJORA DE ALPACAS EN LA REGIÓN ALTOANDINA DE
HUANCAVELICA-PERU
1 2 3 1Quispe, E. C. ; L. ; A. ; H.
1Departamento Académico de Zootecnia. Facultad de Ciencias de Ingeniería. Universidad Nacional de Huancavelica. Ciudad Universitaria Paturpampa s/n. Huancavelica. Perú.
2 Departamento de Producción Agraria. Universidad Pública de Navarra. Campus de Arrosadía 31006. Pamplona. España.
3 Escuela de Postgrado. Universidad Nacional Agraria. Av. La Molina s/n. La Molina. Lima. Perú.
ABSTRACT
The research was developed in 544 Huacaya alpacas of white color located in 19 centers of production of the Region of Huancavelica, located a rank of altitude between 4,000 and 4.800 meters above sea level. Before the shearing, approximately 20g sample were taken from fleece of the mid side and after the shearing the dirty fleece weight (PVS), sex, age and productive unit was registered of each animal. Fibre diameter (DF), coefficient of variation of fibre diameter (CVDF) and the prickle factor (FP) was determined using the Sirolan-Laserscan. The feeding was based on the pasturing on natural grass, where gramineas species predominate. In function to the collected data the exposition of an improvement scheme was made and on it we considered the answer to the selection under two scenarios. The means±S.E. of PVS, DF, CVDF and FP were of 2300±39g, 21,56±0,12, 22,82±0,12% and 6.33 ± 0.30% respectively. Variation of PVS and DF show significance (p<0,01) for sex, age and productive unit, whereas variations of PF has significance (p<0,01) for age and unit of production and variations of CVFD was only for productive unit (p<0,01). When the relations between the four variables of the fibre were analyzed, only was significance between DF and FP, DF and PVS, CVDF and FP, and CVDF and DF. About these considerations the scheme of improvement leads to a formation of a nucleus elite of reproducers formed by 50 males who could mating to best the 1100 females of the animals with records, that would produce the sufficient amount of males for the replacement of reproducers for this subpopulation. The annual genetic gain prediction, in an individual selection is between 57 and 78g for GFW and -0,17 and -0,23 ìm for FD; whereas, in economic values the annual genetic gain prediccion in a selection by index is between $0,83 and $1,14 depending if a pessimist or optimist scenario is considered. These results would indicate the possibility of making genetic improvement in Andean highland regions, based on the existence of animals with high genetic merit for good quality of fibre.
KEYWORDS: Alpacas, Huacaya, fibre, scheme, genetic gain
INTRODUCCIÓN
En América del Sur se estima que existen más de 7,5 millones de cabezas de camélidos sudamericanos, los cuales son agrupados en cuatros especies, y de ellos dos son silvestres: La vicuña (Vicugna vicugna) y el guanaco (Lama guanicoe); y dos son domésticos: La llama (Lama glama) y la alpaca (Lama
Alfonso, Flores, Guillén,
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
93

pacos). En la actualidad, los camélidos sudamericanos probablemente constituyen el único medio de utilización productiva de las extensas áreas de pastos naturales de las zonas alto andinas donde no es posible la agricultura ni la crianza económica de otras especies de animales domésticos, debido a que convierten, con inusual eficiencia, los pastos pobres de estas alturas en productos de alta calidad como son la fibra y la carne. El Perú tiene el privilegio de ocupar el primer lugar en el mundo en la tenencia de alpacas y vicuñas y el segundo lugar en llamas, después de Bolivia. El aprovechamiento racional de esta ventaja comparativa es el reto que éste país encara como el medio más efectivo de lucha contra la pobreza y la inseguridad alimentaria que afecta a las comunidades campesinas que viven de la crianza de estas especies (Brenes et al., 2001; Caballero y Flores, 2004).
La alpaca es el más importante miembro de los camélidos sudamericanos en cuanto se refiere a la producción de fibra (León-Velarde y Guerrero, 2001; Wuliji et al., 2000; Caballero y Flores, 2004), y en función a ella habría sido seleccionada desde hace más de 3000 años (Wang et al., 2003). La industria textil refiere a la fibra de alpaca como una fibra especial y las prendas que se confeccionan con ellas, están clasificadas como artículos de lujo (Wang et al, 2003). Su población mundial se estima en unos 3,7 millones (FAO, 2005) y el 80% de ellas (aprox. 3 millones) se encuentran principalmente en las zonas alto andinas de Perú, de los que alrededor del 86% son alpacas de color blanco (Brenes et al., 2001); y el remanente se ubica principalmente en Bolivia y Chile, aunque se han introducido también exitosamente en Australia, Canadá, Inglaterra, Francia, Nueva Zelanda y Estados Unidos (Lupton et al., 2006). Perú es el primer productor mundial de fibra de alpaca y en el año 2003 se registró una producción de 6.440 TM por un valor encima de los 82 millones de dólares (De los Ríos, 2006).
Existen dos razas de alpacas: La Huacaya, que se caracteriza por ser compacta, de fibra suave y con presencia de ondulaciones, asimismo el vellón manifiesta una apariencia esponjosa, similar al vellón del ovino de raza Corriedale, lo que le da una apariencia más voluminosa; y la Suri la cual presenta fibras de gran longitud que se organizan en rizos que caen por los costados del cuerpo, similar a lo que se observa en los ovinos de raza Lincoln, dándole al animal una apariencia angulosa (Hoffman y Fowler, 1995; Antonini et al., 2004; FAO, 2005). La raza Huacaya es la que predomina con un 85% mientras que la raza Suri sólo representa el 15% de la población total (Caballero y Flores, 2004).
En Huancavelica - región ubicada en una zona alto andina de la sierra centro de Perú – existen más de 225,000 alpacas (MINAG, 2006), las cuales constituyen alrededor del 7% de la población, y de ellos el 90% son de la raza Huacaya.La crianza de la alpaca constituye una importante actividad económica del poblador
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
94

alto andino, la cual se desarrolla por encima de los 3 700 msnm; las familias están organizadas en comunidades de pastores, bajo un sistema de crianza de Rebaño Mixto Familiar (alpacas, llamas, ovinos). Estos rebaños se caracterizan por carecer de sistemas de crianza adecuados, con escasos criterios de mejoramiento genético, basados exclusivamente como fuente alimenticia en pastizales alto andinos de baja soportabilidad, con escasez de cuidados sanitarios. Se estima en más de 60 las comunidades alpaqueras, que agrupan a 3,300 familias aproximadamente; distribuidas principalmente en cuatro provincias: Angaraes, Huaytará, Castrovirreyna y Huancavelica (Quispe, 2005; Gobierno Regional de Huancavelica, 2006).
Los vellones obtenidos de las alpacas en las comunidades alpaqueras de Huancavelica se consideran de baja producción y calidad. A nivel de comunidades se estima una producción promedio bianual de 2,1 kg, mientras que bajo una crianza medianamente tecnificada se obtiene una producción anual de 2,3 kg (Jáuregui y Bonilla, 1991; Nieto y Alejos, 1999). Con respecto a la finura De Los Ríos (2006), refiere que a nivel nacional, el 20% de la producción está dado por fibra gruesa (mayor de 29), el 46% por fibra semi fina (26.6 a 29 micras), el 22 % por fibra fina (23,1 a 26,5 micras) y sólo el 12% está conformado por fibra extra fina (menor de 23,1 micras), lo cual vislumbra el enorme déficit en calidad; también muchos de los vellones son canosos, pintados y canosos-pintados, y en muchos vellones se encuentran gran heterogeneidad en la estructura, pues muchas fibras que lo conforman son de tipo medulado de forma continua o discontinua, lo cual desmerece la calidad del vellón. En función a estas consideraciones se puede apreciar que existe un deterioro genético de la alpaca principalmente en lo que se refiere a la finura de la fibra, aunque también en lo referente al peso del vellón, lo cual se podría mejorar si se ejecutase un adecuado programa de mejoramiento.
Existen muchos estudios sobre fibra de alpaca, es así que se han realizado caracterizaciones de peso de vellón (Jáuregui y Bonilla, 1991; Mamani, 1991 y Frank et al., 2006); finura, longitud de fibra, medulación y otras características (León, 1998; Hack et al, 1999; Wang et al., 2003; Ayala y Chávez, 2006; Lupton et al., 2006 y Frank et al., 2006). Las relaciones entre peso de vellón, diámetro de fibra y otras características de importancia del vellón también han sido abordados por Hack et al. (1999), Ponzoni, et al. (1999), Wuliji et al. (2000) y Wang et al. (2003). Los efectos tecnológicos y medioambientales fueron parcialmente determinados, en función a peso de vellón, finura y otros (Apomayta y Gutiérrez, 1998; León, 1998; Chávez y Pumayalla, 1988; Hack et al., 1999; McGregor y Butler, 2004; Lupton et al., 2006; Braga et al., 2006). Trabajos con respecto a la estructura interna de fibra, y otras propiedades de importancia para el procesamiento textil han sido reportados por Wang et al. (2003), Wang et al. (2005) y Carpio, (1991); mientras que la determinación de parámetros genéticos
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
95

tales como la repetibilidad y heredabilidad para algunos caracteres de la fibra han sido tratados por Ponzoni et al. (1999), Wuliji et al. (2000) y Ayala y Chávez (2006). Predicciones del progreso genético utilizando índices de selección fueron realizadas por Ponzoni (2000) y León-Velarde y Guerrero (2001), quienes muestran ganancias genéticas referidas a peso de vellón, diámetro de fibra y longitud de mecha, bajo ciertos esquemas de selección. Teniendo en consideración lo planteado en los párrafos anteriores se ha realizado el presente trabajo como línea de base de un programa de mejora de alpacas Huacaya de color Blanco en la Región de Huancavelica-Perú a fin de caracterizar el vellón (con respecto a peso de vellón, diámetro de fibra, coeficiente de variación del diámetro de fibra e índice de confort), identificar los efectos de sexo, edad y localidad sobre las características de la fibra, determinar la correlación fenotípica entre tales características y finalmente en base a dichos resultados estimar el progreso genético que se obtendría bajo diferentes escenarios, tomando en consideración los dos criterios de selección más importantes para la comercialización y la industria textil (el peso de vellón y el diámetro de fibra).
MATERIAL Y MÉTODOS
Localización y animales
El trabajo de campo se desarrolló en 19 centros de producción, ubicados en 17 localidades y 8 comunidades (Pastales Huando, Alto Andino, Pucapampa, Choclococha, Sallcca, Santa Bárbara, Lachocc, Carhuancho) de la Región de Huancavelica (Tabla I), situadas en altitudes que oscilan entre 4.000 y 4,800 metros sobre el nivel del mar, con temperaturas que varían desde -5 °C a 0 °C. por las noches y durante el día entre 14 a 18 °C; y con una precipitación pluvial que alcanza los 752.4 mm/año. Los animales con que se trabajó estaban incluidos dentro del Programa de Mejoramiento Genético de la región de Huancavelica, por lo cual tenían las características raciales fenotípicas de alpacas de la raza Huacaya, eran de color blanco entero y ninguna mostraba defectos genéticos.
Evaluación de las características de la fibra
Los datos de peso de vellón sucio (PVS) fueron tomados de 544 alpacas, previamente identificadas con aretes de plástico sujetos a la oreja derecha, para luego registrarse el sexo y los dientes erupcionados a fin de determinar la edad (<1,5 años = dientes de leche; entre 1,5 a 3 años = dos incisivos permanentes; entre 3 y 4 años = cuatro incisivos permanentes; y > 4 años = incisivos
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
96
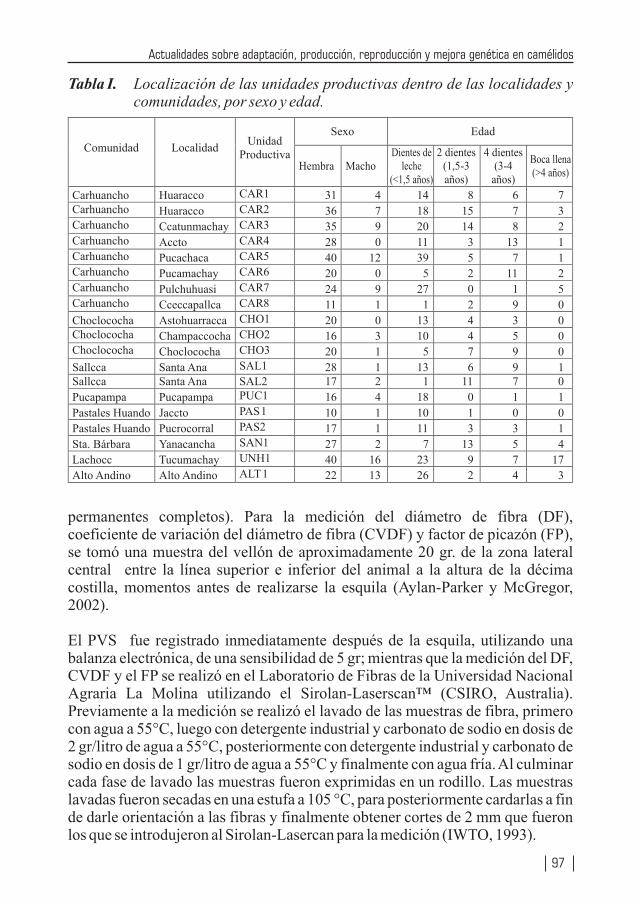
permanentes completos). Para la medición del diámetro de fibra (DF), coeficiente de variación del diámetro de fibra (CVDF) y factor de picazón (FP), se tomó una muestra del vellón de aproximadamente 20 gr. de la zona lateral central entre la línea superior e inferior del animal a la altura de la décima costilla, momentos antes de realizarse la esquila (Aylan-Parker y McGregor, 2002).
El PVS fue registrado inmediatamente después de la esquila, utilizando una balanza electrónica, de una sensibilidad de 5 gr; mientras que la medición del DF, CVDF y el FP se realizó en el Laboratorio de Fibras de la Universidad Nacional Agraria La Molina utilizando el Sirolan-Laserscan™ (CSIRO, Australia). Previamente a la medición se realizó el lavado de las muestras de fibra, primero con agua a 55°C, luego con detergente industrial y carbonato de sodio en dosis de 2 gr/litro de agua a 55°C, posteriormente con detergente industrial y carbonato de sodio en dosis de 1 gr/litro de agua a 55°C y finalmente con agua fría. Al culminar cada fase de lavado las muestras fueron exprimidas en un rodillo. Las muestras lavadas fueron secadas en una estufa a 105 °C, para posteriormente cardarlas a fin de darle orientación a las fibras y finalmente obtener cortes de 2 mm que fueron los que se introdujeron al Sirolan-Lasercan para la medición (IWTO, 1993).
Tabla I. Localización de las unidades productivas dentro de las localidades y comunidades, por sexo y edad.
Comunidad LocalidadUnidad
Productiva
Sexo Edad
Macho
Carhuancho Huaracco CAR1 31 4 14 8 6 7Carhuancho Huaracco CAR2 36 7 18 15 7 3Carhuancho Ccatunmachay CAR3 35 9 20 14 8 2Carhuancho Accto CAR4 28 0 11 3 13 1Carhuancho Pucachaca CAR5 40 12 39 5 7 1Carhuancho Pucamachay CAR6 20 0 5 2 11 2Carhuancho Pulchuhuasi CAR7 24 9 27 0 1 5Carhuancho Cceccapallca CAR8 11 1 1 2 9 0
Choclococha Astohuarracca CHO1 20 0 13 4 3 0Choclococha Champaccocha CHO2 16 3 10 4 5 0Choclococha Choclococha CHO3 20 1 5 7 9 0
Sallcca Santa Ana SAL1 28 1 13 6 9 1Sallcca Santa Ana SAL2 17 2 1 11 7 0
Pucapampa Pucapampa PUC1 16 4 18 0 1 1
Pastales Huando Jaccto PAS1 10 1 10 1 0 0
Pastales Huando Pucrocorral PAS2 17 1 11 3 3 1
Sta. Bárbara Yanacancha SAN1 27 2 7 13 5 4
Lachocc Tucumachay UNH1 40 16 23 9 7 17
Alto Andino Alto Andino ALT1 22 13 26 2 4 3
HembraDientes de
leche(<1,5 años)
2 dientes(1,5-3 años)
4 dientes(3-4
años)
Boca llena(>4 años)
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
97

La esquila se realizó mecánicamente a los animales que tenían una longitud de
mecha ≥ a 9cm, y se adaptaron una 'alzas' a las esquiladoras mecánicas, que permitieron dejar una cubierta de fibra de unos 2 cm, a fin de evitar dejar al animal sin ninguna cubierta de fibra, de modo que puedan tener cierta protección contra las bajas temperaturas de la zona. Las esquilas se realizaron en dos campañas (Noviembre-Diciembre y Marzo-Abril).
Los animales estuvieron sometidos a un sistema de crianza extensiva, pastoreando durante el día y por las noches eran recluidos en unos corrales hechos a base de piedra. La alimentación estuvo basada en pastos naturales, donde predominaban especies pertenecientes a las familias Poaceae, Cyperaceae, Asteraceae y Juncaceae.
Para la evaluación de las características de la fibra se utilizó medias con su respectivo error estándar (E.E.). La determinación del efecto de los factores fijos: edad, sexo y locación sobre el PV, DF, CVDF y FP, se hizo mediante un modelo aditivo lineal de efectos principales; en la comparación de medias se utilizó la prueba de Duncan; y la relación entre las variables de la fibra fue calculada mediante el coeficiente de Pearson. Se utilizó el programa informático SPSS Versión 12.
Propuesta de un esquema de mejora y predicción de la respuesta a la selección (RS)
Para el planteamiento del esquema de mejora se tuvo en cuenta los índices productivos y reproductivos que se muestran en la Tabla II, los cuales fueron encontrados a nivel de la región de Huancavelica por Nieto y Alejos (1999) y a
INDICE PRODUCTIVONº de hembras por macho para empadre% de Machos para reemplazo de reproductores% de problemas en machos de reemplazo% de natalidad% de mortalidad(crias)% de mortalidad(adultos)% de machos de crías nacidas% de animales con registros de la población de alpacas blancas% de animales que aún no producen fibra de la población de alpacas blancasComposición de la población de alpacas blancas con registros (expresado en %)
Crías (machos y hembras)Tuis (< de 3 años)Machos reproductoresHembras reproductoras
VALOR2560124010550
10.030.0
30,028,06,3
35,7
Tabla II. Índices productivos y reproductivos considerados en el planteamiento del esquema de selección.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
98

nivel de Perú por De Los Ríos (2006), FAO (2005) y Brenes et al. (2001). El esquema de mejora a plantearse consideró la formación de un núcleo élite de reproductores machos, con los cuales se posibilitaría el empadre de las mejores reproductoras hembras, para producir crías machos suficientes para el reemplazo del grupo de reproductores machos con registros de producción a fin de empadrar la totalidad de hembras con registros, y de éste modo viabilizar la difusión del material genético deseable a la majada general, mediante el exceso de machos reproductores con registros.
Para la estimación del progreso genético o respuesta a la selección (RS), teniendo como criterio de selección el PVS y DF (caracteres más importantes en la comercialización y la industria de la fibra, de acuerdo a lo indicado por Wang et al., 2003; Frank et al., 2006; McGregor, 2006, Quispe, 2005), se consideraron las desviaciones estándar obtenidas en el presente estudio para PVS y DF respectivamente, mientras que las heredabilidades (h2), presión de selección (Ps) e intervalo generacional (I) fueron obtenidas de revisiones bibliográficas. Se consideraron heredabilidades de 0,30 y 0,25 para PVS y DF respectivamente, en un escenario pesimista y de 0,41 y 0,37 para PVS y DF respectivamente en un escenario optimista .La presión de selección y el intervalo generacional se asumieron bajo dos posibles escenarios: pesimista (Ps = 0,07 y 0,21; I = 5,5 y 6,0; para machos y hembras respectivamente) y optimista (Ps = 0,04 y 0,11; I = 4,5 y 5,5; para machos y hembras respectivamente). La RS se calculó para la selección de cada carácter por separado y combinando ambos en un agregado económico. El cálculo de la RS para cada carácter por separado se predijo según la ecuación: RS = (i*h2*P)/I, considerando i en concordancia con Ps, de acuerdo a la tabla indicada en Bourdon (1997), donde I representa el intervalo generacional e i es la intensidad de selección, ambos valores referidos al promedio de macho y hembra (Bourdon, 1997; Quispe y Alfonso, 2007). Para la predicción de RS en unidades monetarias ($) se consideró un precio de $ 7,00 US por kg/fibra, un costo de producción de $ 3,5 / kg de fibra producida y que por la disminución en 1 µm en un vellón que pesa 2300 g. se obtiene a la venta un adicional de $ 2,38 US. Los pesos económicos utilizados para el cálculo de los índices de selección (con y sin restricción) del PVS (aPVS = 0,0137) y del DF (aDF = -2,375) fueron calculados en base a una derivación parcial de la función de beneficio (Ingresos-Costes), a fin de maximizar ganancias (Quispe y Alfonso, 2007); mientras que la correlación fenotípica entre PVS y DF fue estimada a partir de los datos, y para la correlación genotípica se consideraron diversos valores a fin de asumir múltiples escenarios. Para la predicción del RS anual se utilizó la siguiente ecuación matricial:
Donde “b” es la matriz de coeficientes del índice de selección, “P” es la matriz de varianzas-covarianzas fenotípicas y “G” es la matriz de varianzas-covarianzas genéticas de PVS y DF.
I
i
Gb
bPbRS *
'*
*'*=
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
99

RESULTADOS Y DISCUSIÓN
Evaluación de las características de la fibra.
Las medias ± E.E. obtenidas para PVS, DF, CVDF y FP fueron de 2300±39g, 21,56±0,12ìm, 22,82±0,12% y 6,33±0,30% respectivamente. La media general del PVS resulta ser mayor a los valores reportados por Jáuregui y Bonilla (1991), Castellaro (1998), Wuliji et al. (2000), León-Velarde y Guerrero (2001) y Brenes et al. (2001); similar a lo encontrado por Condorena (1985), Bryant et al. (1989), Nieto y Alejos (1999) y De Los Ríos (2006), aunque resulta ser menor a lo reportado por Ponzoni (1999), Ponzoni (2000), McGregor (2002) y McGregor (2006), pudiendo deberse ésta superioridad a que dichos animales estuvieron bajo condiciones de una mejor alimentación la cual tiene efecto positivo en la producción de la fibra (Bryant et al., 1989), contrario a una nutrición inadecuada lo cual disminuye el crecimiento de la fibra, tal como es discutido por McGregor (2002). Por otro lado el PVS exhibe una regular variabilidad (coeficiente de variación=39,44%), lo que permitiría obtener ganancias genéticas altas si la mejora genética se realizaría por selección teniendo como criterio este carácter (Van Vleck, 1993; Bourdon, 1997), lo que justifica la inclusión de dicho criterio para programas de mejora genética, toda vez que también constituye un factor de importancia al momento de la comercialización del vellón.
El DF es menor a los reportados por Ponzoni et al. (1999), Wuliji et al. (2000), Marti et al. (2000), Wang et al. (2003) y Wang et al. (2005), que conlleva a considerar que un gran porcentaje de vellones que se producen en estos rebaños corresponderían un fibra fina (20,1 a 23 ìm) y superfina (<20ìm), contrario a lo indicado por De Los Ríos (2006) y McGregor (2006), lo cual indicaría la existencia de animales con buena calidad de fibra que podría hacer factible la formación de una élite de reproductores para poder difundir adecuadamente éste deseable potencial genético; sin embargo esta aparente finura podría deberse a las condiciones pobres de alimentación de los animales en estudio (bajo pasturas naturales), lo cual concordaría a lo indicado por McGregor (2002) y Marshall et al. (1981), y corroborado por Wuliji et al. (2000), quienes demostraron que las condiciones nutricionales adversas en alpacas y ovinos reducen el DF, contrario al efecto que causa una buena alimentación, lo cual incrementa el diámetro de fibra.
La media del CVDF obtenida, que resultaría ser bastante buena para las exigencia de la industria textil, es ligeramente inferior a lo obtenido por Hack et al.(1999), Aylan-Parker y McGregor (2002), Wang et al. (2003), McGregor y Butler (2004) y Lupton et al. (2006), quienes reportan valores entre 23,48 y 28,10%. Esta característica tiene alta influencia sobre algunas propiedades requeridas en la industria textil (Lupton et al., 2006), pues conjuntamente con el DF determinan la
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
100

finura para el torcido de la fibra asociado al rendimiento del hilado, propiedad conocida también como “spinning finnes” (SF), cuyo cálculo está basado en el DF con un CVDF de 24%, de modo que valores debajo de ello resultan en un SF más bajo que el DF y valores por encima de ello resultan en un SF más alto (McGregor, 2006), debido a que cuando se tienen fibras de diámetro heterogéneo, se requieren mayor número de ellas para alcanzar igual resistencia y uniformidad (Mueller, 2000). Por otro lado, el FP está asociado a la sensación de picazón (“prickle factor”), por lo que la industria textil requiere de vellones con FP muy bajos. La media ± E.E. del FP encontrada (6,33±0,30%) resulta ser muy inferior a 31,61%, valor que es reportado por Lupton et al. (2006), lo cual se debería al valor alto de DF que encuentra.
El PVS y el DF muestran variaciones altamente significativas (p<0,01) para sexo, edad y unidad productiva, resultando que animales jóvenes tienen menor PVS y DF en contraste con los de mayor edad. Las unidades de producción con menor PVS fueron CHO1, PUC1 y CAR1, y menor DF fueron CHO2 y PAS1, CAR7, SAN1, CHO1 y SAL1; mientras que las que unidades de producción CAR8, CAR4, CAR3, CAR6 y UNH1 tuvieron mayores PVS, y las que tienen mayor DF fueron PUC1, CAR4, UNH1, CAR6, ALT1, CAR8, CAR1, SAL2 y CAR2 (Tabla III).
Las variaciones del PVS por efecto de la edad y el sexo concuerdan a lo reportado por Castellaro et al. (1998), Wuliji et al. (2000), McGregor (2006) y Lupton et al., (2006), de modo que el mayor PVS de los animales adultos se debería al incremento de la superficie corporal (León-Velarde y Guerrero, 2001; Frank et al., 2006), y la influencia del sexo sería debida a que la preñez y la lactación de la hembras reducirían la producción (Newman y Paterson, 1996).
La variación del DF por efecto de la edad concuerda a lo hallado por Wuliji et al. (2000), quien reporta medias de DF de 26,5 y 32,0ìm para crías y adultos respectivamente, lo cual también está acorde a lo encontrado por Lupton et al. (2006). McGregor y Butler (2004), en función a la diferencia de DF que encuentra en alpacas de distintas edades posibilita la existencia del “micron blowout”; término que es utilizado en la industria lanar para describir el incremento del diámetro de la lana con la edad que no es debido a influencias medioambientales temporales. Los ovinos varían en su habilidad para mantener o incrementar levemente el DF a través de todo su periodo de vida o por efecto de una alimentación abundante (Cottle et al., 1995), pudiendo el “micron blowout” ser medido por regresión del DF con la edad o por diferencia entre registros dados a diferentes edades (Hill et al., 1999).
Por otro lado el efecto del sexo sobre el DF encontrado concuerda al de Lupton et al. (2006), aunque difiere a lo reportado por Wuliji et al. (2000), McGregor y
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
101
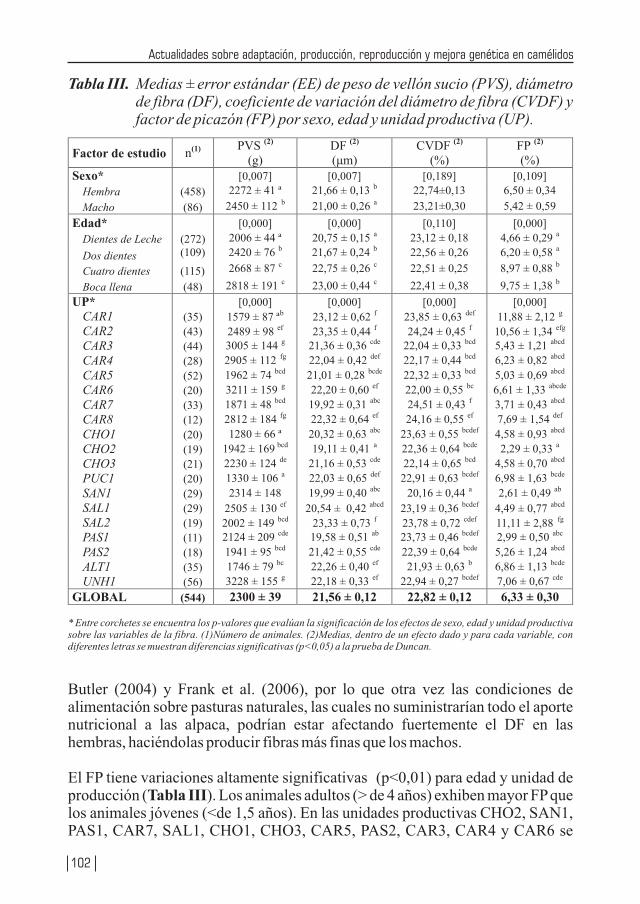
Butler (2004) y Frank et al. (2006), por lo que otra vez las condiciones de alimentación sobre pasturas naturales, las cuales no suministrarían todo el aporte nutricional a las alpaca, podrían estar afectando fuertemente el DF en las hembras, haciéndolas producir fibras más finas que los machos.
El FP tiene variaciones altamente significativas (p<0,01) para edad y unidad de producción (Tabla III). Los animales adultos (> de 4 años) exhiben mayor FP que los animales jóvenes (<de 1,5 años). En las unidades productivas CHO2, SAN1, PAS1, CAR7, SAL1, CHO1, CHO3, CAR5, PAS2, CAR3, CAR4 y CAR6 se
Factor de estudio n(1) PVS (2)
(g)DF (2)
(ìm)CVDF (2)
(%)FP (2)
(%)Sexo* [0,007] [0,007] [0,189] [0,109]
Hembra (458) 2272 ± 41 ª 21,66 ± 0,13 b 22,74±0,13 6,50 ± 0,34
Macho (86) 2450 ± 112 b 21,00 ± 0,26 a 23,21±0,30 5,42 ± 0,59
Edad* [0,000] [0,000] [0,110] [0,000]
Dientes de Leche (272) 2006 ± 44 ª 20,75 ± 0,15 a 23,12 ± 0,18 4,66 ± 0,29 a
Dos dientes (109) 2420 ± 76 b 21,67 ± 0,24 b 22,56 ± 0,26 6,20 ± 0,58 a
Cuatro dientes (115) 2668 ± 87 c 22,75 ± 0,26 c 22,51 ± 0,25 8,97 ± 0,88 b
Boca llena (48) 2818 ± 191 c 23,00 ± 0,44 c 22,41 ± 0,38 9,75 ± 1,38 b
UP* [0,000] [0,000] [0,000] [0,000]
CAR1 (35) 1579 ± 87 ªb 23,12 ± 0,62 f 23,85 ± 0,63 def 11,88 ± 2,12 g
CAR2 (43) 2489 ± 98 ef 23,35 ± 0,44 f 24,24 ± 0,45 f 10,56 ± 1,34 efg
CAR3 (44) 3005 ± 144 g 21,36 ± 0,36 cde 22,04 ± 0,33 bcd 5,43 ± 1,21 abcd
CAR4 (28) 2905 ± 112 fg 22,04 ± 0,42 def 22,17 ± 0,44 bcd 6,23 ± 0,82 abcd
CAR5 (52) 1962 ± 74 bcd 21,01 ± 0,28 bcde 22,32 ± 0,33 bcd 5,03 ± 0,69 abcd
CAR6 (20) 3211 ± 159 g 22,20 ± 0,60 ef 22,00 ± 0,55 bc 6,61 ± 1,33 abcde
CAR7 (33) 1871 ± 48 bcd 19,92 ± 0,31 abc 24,51 ± 0,43 f 3,71 ± 0,43 abcd
CAR8 (12) 2812 ± 184 fg 22,32 ± 0,64 ef 24,16 ± 0,55 ef 7,69 ± 1,54 def
CHO1 (20) 1280 ± 66 ª 20,32 ± 0,63 abc 23,63 ± 0,55 bcdef 4,58 ± 0,93 abcd
CHO2 (19) 1942 ± 169 bcd 19,11 ± 0,41 a 22,36 ± 0,64 bcde 2,29 ± 0,33 a
CHO3 (21) 2230 ± 124 de 21,16 ± 0,53 cde 22,14 ± 0,65 bcd 4,58 ± 0,70 abcd
PUC1 (20) 1330 ± 106 a 22,03 ± 0,65 def 22,91 ± 0,63 bcdef 6,98 ± 1,63 bcde
SAN1 (29) 2314 ± 148 19,99 ± 0,40 abc 20,16 ± 0,44 a 2,61 ± 0,49 ab
SAL1 (29) 2505 ± 130 ef 20,54 ± 0,42 abcd 23,19 ± 0,36 bcdef 4,49 ± 0,77 abcd
SAL2 (19) 2002 ± 149 bcd 23,33 ± 0,73 f 23,78 ± 0,72 cdef 11,11 ± 2,88 fg
PAS1 (11) 2124 ± 209 cde 19,58 ± 0,51 ab 23,73 ± 0,46 bcdef 2,99 ± 0,50 abc
PAS2 (18) 1941 ± 95 bcd 21,42 ± 0,55 cde 22,39 ± 0,64 bcde 5,26 ± 1,24 abcd
ALT1 (35) 1746 ± 79 bc 22,26 ± 0,40 ef 21,93 ± 0,63 b 6,86 ± 1,13 bcde
UNH1 (56) 3228 ± 155 g 22,18 ± 0,33 ef 22,94 ± 0,27 bcdef 7,06 ± 0,67 cde
GLOBAL (544) 2300 ± 39 21,56 ± 0,12 22,82 ± 0,12 6,33 ± 0,30
Tabla III. Medias ± error estándar (EE) de peso de vellón sucio (PVS), diámetro de fibra (DF), coeficiente de variación del diámetro de fibra (CVDF) y factor de picazón (FP) por sexo, edad y unidad productiva (UP).
* Entre corchetes se encuentra los p-valores que evalúan la significación de los efectos de sexo, edad y unidad productiva sobre las variables de la fibra. (1)Número de animales. (2)Medias, dentro de un efecto dado y para cada variable, con diferentes letras se muestran diferencias significativas (p<0,05) a la prueba de Duncan.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
102

encuentras las menores medias del FP, y las mayores medias se ubican en CAR2, SAL2 y CAR1. El CVDF solo muestra variaciones para unidad productiva, con una media inferior encontrada en SAN1, mientras que en PUC1, UNH1, SAL1, CHO1, PAS1, SAL2, CAR1, CAR8, CAR2 y CAR7 (Tabla III).
Por otro lado, la falta de efecto del sexo sobre el CVDF, concuerda con lo reportado por McGregor y Butler (2004), sin embargo la falta de efecto de la edad sobre CVDF, resulta discordante con lo hallado por los investigadores antes citados, quienes encuentran evidencia que el CVDF disminuye rápidamente hasta los 2 a 3 años y para luego incrementar levemente hasta los 10 años. Esta aparente contradicción puede ser explicada debido a que en nuestro estudio la determinación de la edad fue solo considerando la erupción dentaria, la cual constituye sólo una aproximación, mientras que McGregor y Butler (2004), lo hicieron en función a fecha de nacimiento de los animales y evaluándolos cada año.
Nuestros resultados de la variabilidad del FP por efecto de la edad, resulta concordante a lo reportado en alpacas por Lupton et al. (2006), en llamas por Wurzinger et al. (2006) y en ovejas por Notter et al. (2007); asimismo la falta de variabilidad del FP por efecto del sexo encontrada, resulta concordante con Lupton et al. (2006) cuando compara FP entre alpacas machos y hembras, pero discordante con lo encontrado en llamas por Wurzinger et al. (2006), sin embargo estos aspectos merecen una mayor profundidad de estudio debido a la falta de información en alpacas.
Las variaciones de todas las características de la fibra en estudio (PVS, DF, CVDF e FP) debido al efecto de las unidades productivas, resultan justificadas debido a que cada una de ellas pertenecen a diferentes propietarios y se encuentran localizados en diversos lugares de la región (Huancavelica), lo que hace que se encuentren sometidos a diferentes prácticas de manejo (alimentación, sanidad, entre otros) y que también podría estar influyendo el origen genético (Montes et al., 2007), por lo que se convendría el ingreso de este factor fijo dentro de un modelo de evaluación genética.
Las correlaciones entre DF y FP, DF y PVS, y CVDF y FP son altamente significativas (p<0,01), sin embargo sólo la primera resulta ser alta (0,84), mientras que las otras son muy bajas (0.12 y 0.14 respectivamente). La correlación entre CVDF y DF es sólo significativa (<0.05), pero el valor es bajo (-0.088). Finalmente las correlaciones entre PVS con CVDF y FP no son significativas.
Las correlaciones fenotípicas pueden ser buenos indicadores de la magnitud y el signo de las correlaciones genéticas (Lynch y Walsh, 1997), por lo tanto resulta importante la discusión de los hallazgos que pueda realizarse en torno a ellas. La
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
103

existencia de relación encontrada entre PVS y DF es baja frente a correlaciones de 0,25, 0,45 y 0,53 (p<0,01) determinadas en rebaños de Australia, Nueva Zelanda y Norteamérica por Ponzoni et al. (1999), Wuliji et al. (2000) y Lupton et al. (2006) respectivamente; así como en comparación a lo hallado por Safari et al. (2007) en una revisión de 15 trabajos sobre parámetros productivos en ovinos (0,14 y 0,45); sin embargo, esta débil relación encontrado por nosotros podría deberse al amplio rango de intervalo entre esquilas que tuvieron las alpacas en nuestro estudio (los animales fueron esquilado en base a una longitud de mecha ≥ 9 cm), y que también factores como altitud, condición alimenticia, condición corporal, entre otros podrían estar influyendo. La débil de relación entre CVDF con PVS y FD, concuerda con lo reportado por Ponzoni et al. (1999), mientras que la falta de relación encontrada entre PVS y FP, así como la baja relación entre DF y CVDF, no han sido bien estudiadas en esta especie, aunque Safari et al. (2005) al realizar una revisión de 10 investigaciones en ovinos obtiene un coeficiente de correlación entre DF y CVDF de -0,09, el cual resulta casi nula, la que guardaría concordancia con nuestro resultado. Por otro lado, la alta relación encontrada entre DF y FP, se hace evidente, ya que cuando menor sea el DF existirían también menos cantidad de fibras que estén por encima de las 30ìm, como así sucede en ovinos cuando se relaciona el DF con el índice confort (porcentaje de fibras menores a 30µ m, lo cual sería el complemento del FP), para lo cual Notter et al. (2007) encuentra una alta correlación.
Teniendo en consideración que las dos características más importantes que determinan el precio del vellón son el PVS y DF (Ponzoni, 2000; Hack et al., 1999; Leon-Velarde, 2001; Wang et al., 2003; McGregor y Butler, 2004; Quispe, 2005; McGregor, 2006; Quispe y Alfonso, 2007) dentro de los criterios de selección de un programa de mejora en alpacas se hace necesario su inclusión bajo un índice de selección, y debido que actualmente no existe reportes encontrados sobre la correlación genética entre ambos, se haría conveniente asumir relaciones entre nulas a bajas, teniendo en cuenta correlaciones fenotípicas de los reportados por Ponzoni (1999), Wuliji et al. (2000) y Lupton et al. (2006), en estudios realizados en alpacas, y que correlaciones genéticas positivas entre DF y PVS de moderadas a bajas, tales como 0,52, 0,36 y 0,27, también han sido encontradas en ovinos por Clarke et al. (1999), Safari et al. (2005) y Safari et al. (2007).
Esquema de mejora y la respuesta a la selección (RS)
El esquema de mejora planteado (Figura 1) tiene como base una población inicial de alpacas blancas (≈ 193.500) correspondiente a la región de estudio, de la cual se toma una fracción (10%) para el monitoreo de registro de datos de
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
104

producción y genealógicos (Ponzoni et al., 2000, Willis, 1996), y se llega a un núcleo de 50 machos reproductores, que con cubrición natural, podría empadrar a unas 1110 hembras seleccionadas, que bajo las tasas de nacimiento y mortalidad propios de la zona producirán la suficiente cantidad de machos de reemplazo para servir a las hembras en control, de modo que los machos que son reemplazados producirán el flujo de genes hacia la población de animales sin registros (majada).
Este esquema de selección utilizado en un programa de mejoramiento genético de núcleo abierto posibilita su aplicación dentro de un entorno colectivo, lo cual se hace conveniente en la región de estudio, donde los productores tienen hatos conformados por pocos animales (entre 70 a 150 animales), siendo necesario para ello el acuerdo entre los productores (Mueller, 2003), quienes tendrían un rol decisivo para alcanzar los objetivos deseados, de modo que seleccionando animales con alto potencial genético, permitiría la obtención de ganancia genética, lo cual acompañado de mejora de las prácticas de manejo, alimentación y sanidad derivaría en un mejor performance fenotípico.
Los resultados de las predicciones de la RS anual, en magnitudes de los criterios de selección (PVS en g y DF en μm) y en unidades monetarias por animal ($/a), cuando se seleccionan independientemente por producción individual de PVS o DF, así como cuando se seleccionan en conjunto por las producciones de PVS y DF utilizando índices de selección sin restricción encontrados (I = 0,0026*PVS – 0,0020*DF y I = 0,0035*PVS – 0,1100*DF para escenarios pesimista y optimista respectivamente) y con restricción para PVS encontrados (I = 0,0007*PVS – 0,3660*DF y I = 0,0003*PVS – 0,8820*DF para escenarios pesimista y optimista respectivamente), se muestran en la tabla IV, observándose un mejor retorno económico ($ 1,16 US/animal/año) cuando la selección se realizaría utilizando un índice de selección sin restricción en un escenario optimista si la correlación genética fuese baja (r =0,05) y un bajo retorno económica ($ 0,11 G(PVS,DF)
US/animal/año) cuando la selección se realizaría utilizando un índice de selección sin restricción en un escenario pesimista si la correlación genética fuese alta (r =0,65).G(PVS,DF)
Existen muy pocos trabajos referidos a estimaciones de progreso genético en alpacas de color blanco, y prácticamente es nula la información para zonas alto andinas. Nuestras predicciones de RS anual para PVS son cercanas a la predicción realizada por Mueller (2000) en ovinos Merino, sucediendo de forma similar con respecto al DF (-0,26 contra -0,17 y -0,35ìm cuando se realiza selección masal). Por otro lado, nuestra predicción de RS anual (en términos económicos) en una selección por índice resulta ser menor a lo predicho por Ponzoni (2000), en un estudio realizado en Australia, los cual se debería al uso valores económicos, para peso de vellón y diámetro de fibra, más altos que los nuestros ($15,00 y -$3,00 contra $7,00 y -$2,38 respectivamente); pero al
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
105
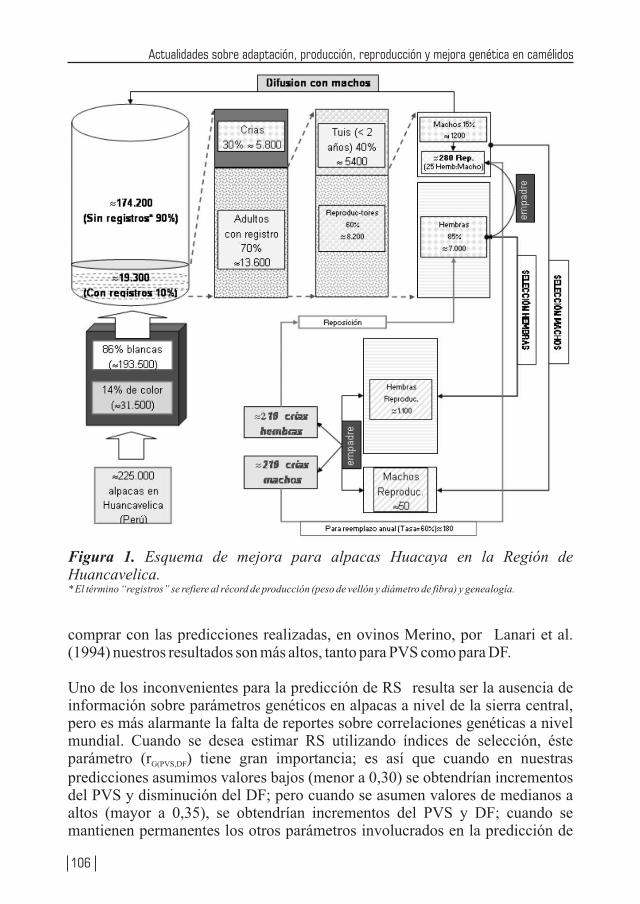
comprar con las predicciones realizadas, en ovinos Merino, por Lanari et al. (1994) nuestros resultados son más altos, tanto para PVS como para DF.
Uno de los inconvenientes para la predicción de RS resulta ser la ausencia de información sobre parámetros genéticos en alpacas a nivel de la sierra central, pero es más alarmante la falta de reportes sobre correlaciones genéticas a nivel mundial. Cuando se desea estimar RS utilizando índices de selección, éste parámetro (r ) tiene gran importancia; es así que cuando en nuestras G(PVS,DF
predicciones asumimos valores bajos (menor a 0,30) se obtendrían incrementos del PVS y disminución del DF; pero cuando se asumen valores de medianos a altos (mayor a 0,35), se obtendrían incrementos del PVS y DF; cuando se mantienen permanentes los otros parámetros involucrados en la predicción de
Figura 1. Esquema de mejora para alpacas Huacaya en la Región de Huancavelica. * El término “registros” se refiere al récord de producción (peso de vellón y diámetro de fibra) y genealogía.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
106
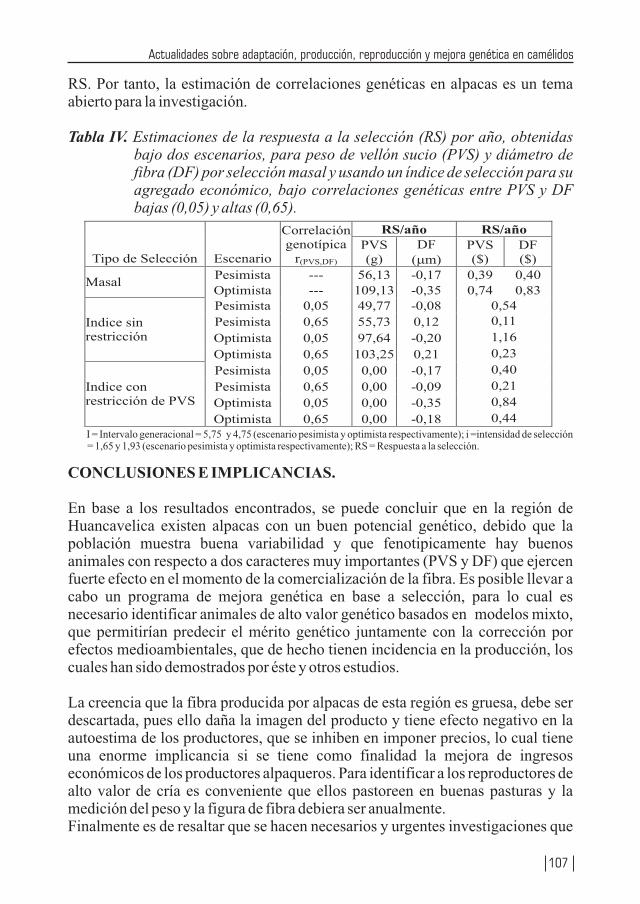
RS. Por tanto, la estimación de correlaciones genéticas en alpacas es un tema abierto para la investigación.
Tabla IV. Estimaciones de la respuesta a la selección (RS) por año, obtenidas bajo dos escenarios, para peso de vellón sucio (PVS) y diámetro de fibra (DF) por selección masal y usando un índice de selección para su agregado económico, bajo correlaciones genéticas entre PVS y DF bajas (0,05) y altas (0,65).
I = Intervalo generacional = 5,75 y 4,75 (escenario pesimista y optimista respectivamente); i =intensidad de selección = 1,65 y 1,93 (escenario pesimista y optimista respectivamente); RS = Respuesta a la selección.
CONCLUSIONES E IMPLICANCIAS.
En base a los resultados encontrados, se puede concluir que en la región de Huancavelica existen alpacas con un buen potencial genético, debido que la población muestra buena variabilidad y que fenotipicamente hay buenos animales con respecto a dos caracteres muy importantes (PVS y DF) que ejercen fuerte efecto en el momento de la comercialización de la fibra. Es posible llevar a cabo un programa de mejora genética en base a selección, para lo cual es necesario identificar animales de alto valor genético basados en modelos mixto, que permitirían predecir el mérito genético juntamente con la corrección por efectos medioambientales, que de hecho tienen incidencia en la producción, los cuales han sido demostrados por éste y otros estudios.
La creencia que la fibra producida por alpacas de esta región es gruesa, debe ser descartada, pues ello daña la imagen del producto y tiene efecto negativo en la autoestima de los productores, que se inhiben en imponer precios, lo cual tiene una enorme implicancia si se tiene como finalidad la mejora de ingresos económicos de los productores alpaqueros. Para identificar a los reproductores de alto valor de cría es conveniente que ellos pastoreen en buenas pasturas y la medición del peso y la figura de fibra debiera ser anualmente.Finalmente es de resaltar que se hacen necesarios y urgentes investigaciones que
RS/año RS/año
Tipo de Selección Escenario
Correlación genotípica
r(PVS,DF)
PVS(g)
DF(mm)
PVS($)
DF($)
Pesimista --- 56,13 -0,17 0,39 0,40Masal
Optimista --- 109,13 -0,35 0,74 0,83
Pesimista 0,05 49,77 -0,08 0,54
Pesimista 0,65 55,73 0,12 0,11
Optimista 0,05 97,64 -0,20 1,16Indice sin restricción
Optimista 0,65 103,25 0,21 0,23
Pesimista 0,05 0,00 -0,17 0,40
Pesimista 0,65 0,00 -0,09 0,21
Optimista 0,05 0,00 -0,35 0,84Indice con restricción de PVS
Optimista 0,65 0,00 -0,18 0,44
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
107

permitan estimar parámetros genéticos (índice de herencia, índice de constancia y correlaciones genéticas) y determinar que factores ambientales son los que ejercen influencia en las características de importancia económica, los cuales influidos en el modelo mixto nos dará estimaciones de los parámetros con mayor precisión y cercano a su verdadero valor.
AGRADECIMIENTO
Agradecemos a todos los propietarios de las diferentes unidades de producción, quienes nos permitieron tomar la información, ejecutar la esquila y obtener muestras de sus rebaños. También apreciamos la asistencia financiera de Dirección Universitaria de Investigación de la Universidad Nacional de Huancavelica, Dirección Universitaria de Economía de la Universidad Nacional Agraria la Molina y de INCAGRO (Contrato Nº 2006-00211), institución dependiente del Ministerio de Agricultura del Gobierno de Perú.
LITERATURA CITADA
Antonini, M., A. Gonzáles y Valbonise. 2004. Relationship between age and postnatal skin follicular development in three types of South American domestic camelids. Livest. Prod. Sci., 90: 241-246.
Apomayta, Z. y G. Gutiérrez. 1998. Evaluación de características tecnológicas y productivas de la fibra en alpacas Huacaya esquiladas a los 12 y 17 meses de edad. Anales Científicos. UNA La Molina. Lima, Perú. 36: 35-42.
Ayala, J. y J.Chávez (2006). Índice de selección genética para características del diámetro de fibra de alpacas de Huacaya Tuis y Adultas en la sierra central del Perú. http://www.infoandina.org/apc-aa-files/237543fdce333f3a56026e59e60 adf7b/ Trabajos Com-pletos.pdf. [28 de Septiembre de 2007].
Aylan-Parker, J. y B.A. McGregor, 2002. Optimising sampling techniques and estimating sampling variance of fleece quality attributes in alpacas. Small Rumin. Res., 44: 53-64.
Bourdon, R.M. 1997. Understanding Animal Breeding. Prentice Hall. New Jersey, USA. 523 pp.
Braga, W., V. Leyva y R. Cochran, 2006. The effect of altitude on alpaca (Lama pacos) fiber production. Technical note. Small Rumin. Res., 68, 323-328.
Brenes, E.R., K. Madrigal, F. Pérez y K. Valladares. 2001. El Cluster de los Camélidos en Perú: Diagnóstico Competitivo y Recomendaciones Estratégicas. Instituto Centroamericano de Administración de Empresas http://www.caf.com/attach/4/default/CamelidosPeru.pdf. [25 de septiembre de 2007].
Bryant, F.C., A. Florez y J. Pfister. 1989. Sheep and alpaca productivity on high andean rangelands in Peru. J. Anim. Sci., 67,3078-3095
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
108

Caballero W. y A. Flores. 2004. La Sierra. Primeria Prioridad para salir del Subdesarrollo Agrario. CONCYTEC
Carpio, M. 1991. Produccion de rumiantes menores: Alpacas. La fibra de camélidos. RERUMEN. 291: 356.
Castellaro, G., J. Garcia-Huidobro y P. Salinas. 1998. Alpaca liveweight variations and fiber production in Mediterranean range of Chile. J. Range Manage., 51, 509-513.
Chávez, J. y A. Pumayalla. 1988. Efectos de localidad, edad y sexo sobre el peso de vellón en alpacas Huacaya de una SAIS de Puno. VI Conv. Int. de Especialistas en Cam., Oruro, Bolivia.
Condorena, N. 1985. Aspectos de un sistema regularizador de la crianza de alpacas. IVITA La Raya. Puno, Perú.
Cottle, D.J., B.C. Russel y K.D. Atkins. 1995. Do Merino hoggets with 'positive micron' wool have a propensity for 'micron blowout'?. Proc. Aust. Assoc. Anim. Breed. Gen., 11: 525-528.
Clarke, J.N., J.L. Dobbie, K.R. Jones, A.L. Wrigglesworth y S.M. Hickey. 1999. Sex and age effects on genetic parameters for wool production and qualities. Proc. N. Z Soc. Anim. Prod., 59: 23-26.
De Los Ríos, E., 2006. Producción textil de fibras de camélidos sudamericanos en el área altoandina de Bolivia, Ecuador y Perú. Organización de las Naciones Unidas para el Desarrollo Industrial (UNIDO). https://www.unido.org/file-storage/download/ ?file_id= 58563. [26 de Septiembre del 2007].
FAO. 2005. Situación Actual de los Camélidos Sudamericanos en el Perú. Organización de las Naciones Unidas para la Agricultura y la Alimentación. Proyecto de Cooperación Técnica en apoyo a la crianza y aprovechamiento de los Camélidos Sudamericanos en la Región Andina TCP/RLA/2914. http://www.fao.org/regional/Lamerica/prior/segalim/animal/ paises/pdf/2914per.pdf. [24 de septiembre de 2007].
Frank E.N., M.V.H. Hick, C.D. Gauna, H.E. Lamas, C. Reniere y M. Antonini. 2006. Phenotypic and genetic description of fibre traits in South American domestic camelids (llamas and alpacas). Small Rumin. Res., 61: 113-129.
Gobierno Regional de Huancavelica. 2006. Plan de Mejoramiento Genético y Medioambiental de Alpacas Huacaya de color blanco a nivel de la región de Huancavelica. Gerencia Regional de Recursos Naturales y Gestión del Medio Ambiente. Proyecto PROALPACA. Huancavelica. Perú.
Hack, W., B. McGregor, R. Ponzoni, G. Judson, I. Carmichael y D. Hubbard. 1999. Australian alpaca fibre. Improving productivity and marketing. http://www.rirdc.gov.au/reports/ RNF/99_140.doc. [1 de octubre de 2007]
Hill, J.A., R.W. Ponzoni y J.W. James. 1999. Micron blowout: heritability and genetic correlations with fibre diameter and secondary follicle diameter. Aust. J. Agric. Res., 50: 1375-1379.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
109

Hoffman, E. y M.E. Fowler. 1995. The Alpaca book. Clay Press Inc., Herald, California.
IWTO. 1993. Measurement of the mean and distribution of fiber diameter using the Sirolan-Lasersacan Fiber Diameter Analyzer. IWTO Test Method, pp. 12-93.
Jaúregui, V. y G. Bonilla. 1991. Productividad de carne, fibra y cuero en alpacas y llamas. XIV Reunión Científica APPA.
Lanari, M.R., E. Tronfi y J.P. Mueller (1994). Revisión de índices de selección PROVINO para la raza Merino en la Argentina. http://www.inta.gov.ar/ bariloche/info/documentos/ animal/genetica/pa244.pdf. [08 de noviembre de 2007].
León, U., 1998. Efecto del ambiente controlado y no controlado sobre el diámetro de lana y fibra de alpaca en verano e invierno. Tesis UNA La Molina. Facultad de Zootecnia. Lima, Perú.
León-Velarde, C.U y J. Guerrero. 2001. Improving quantity and quality of Alpaca fiber; using simulation model for breeding strategies. http://inrm.cip.cgiar.org/ home/publicat/ 01cpb023.pdf. [24 de septiembre de 2007].
Lupton, C.J., A. McColl y R.H. Stobart. 2006. Fiber characteristics of the Huacaya Alpaca. Small Rumin. Res., 64: 211-224.
Lynch, M. y B. Walsh. 1997. Genetic Analysis of Quantitative Traits. Sinauer Assoc. Inc. USA.
Mamani, G. 1991. Parámetros genéticos del peso vivo y vellón en alpacas Huacaya de La Raya, Puno. In: VII Conv. Int. Especialistas en Cam. Sud., Jujuy, Argentina.
Marshall, A.J., V. Bustinza, y T.L. Quispe. 1981. Efecto de la alimentación con alfalfa sobre la producción y reproducción de la alpaca. Resumen de APPA, Ayacucho, Perú.
Marti, S.B., M. Kreuzer, M.R.L. Scheeder. 2000. Analyse von Einflussfaktoren auf die Faserqualit¨at bei Neuweltkameliden mit dem OFDA-Verfahren. Z¨uchtungskunde 72, 389–400.
McGregor, B.A., 2002. Comparative productivity and grazing behaviour of Huacaya alpacas and peppin Merino sheep grazed on annual pastures. Small Rumin. Res., 44: 219-232.
McGregor, B.A. y K.L. Butler. 2004. Sources of variation in fibre diameter attributes of Australian alpacas and implications for fleece evaluation and animal selection. Aust. J. Agric. Res., 55: 433-442.
McGregor, B.A., 2006. Production, attributes and relative value of alpaca fleeces in southern Australia and implications for industry development. Small Rumin. Res., 61: 93-111.
MINAG. 2006. Portal Agrario. Población de alpacas por departamento. http://www. portalagrario.gob.pe/pecuaria/pec_pop001.shtml. [24 de septiembre de 2007].
Montes, M, I. Quicaño, R. Quispe, E.C. Quispe y L. Alfonso. 2007. Quality characteristics of Huacaya Alpaca fibre produced in the Peruvian Andean Plateau region of Huancavelica. Span. J. of Agric. Res. 6(1):33-38.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
110

Mueller, J. (2000). Mejoramiento Genético de la lana. Caracteres de importancia. http://www.inta.gov.ar/bariloche/info/documentos/animal/genetica/pa374.pdf. [01 de octubre de 2007].
Mueller, J. (2003). Estrategías para el mejoramiento genético de pequeños rumiantes. http://www.inta.gov.ar/bariloche/info/documentos/animal/genetica/pa425.pdf. [9 de noviembre de 2007]
Newman, S-A. N., D. J. Paterson (1996).Estimates of environmental effects for liveweight and fleece characteristics of New Zealand cashmere goats. N. Z. J. Agric. Res., 39: 379-386
Nieto, L. e I. Alejos. 1999. Estado económico y productivo del Centro de Producción e Investigación de Camélidos Sudamericanos – Lachocc. XXI Reunión Científica Anual APPA.
Notter, D.R., L.A. Kuehn, R.W. Kott. 2007. Genetic analysis of fibre characteristics in adult
Targhee ewes and their relationship to breeding value estimates derived from yearling fleches. Small Rumin. Res., 67: 164-172.
Ponzoni R.W., R.J. Grimson, J.A. Hill, D.J. Hubbard, B.A. McGregor, A. Howse, I. Carmichael y G.J. Judson. 1999. The inheritance of and association among some production traits in young Australian alpacas. http://www.alpacas.com/AlpacaLibrary/Inheritance Traits.aspx. [26 de septiembre de 2007].
Ponzoni R.W. (2000). Genetic improvement of Australian Alpacas: present state and potencial developments. Proc. Aust. Alpaca Assoc., pp. 71-96.
Quispe, E.C. 2005. Mejoramiento Genético y Medioambiental de Alpacas en la Región de Huancavelica. Proyecto de Inversión Pública a nivel de Perfil. Universidad Nacional de Huancavelica. Perú.
Quispe, E.C. y L. Alfonso. 2007. Metodología para estimar valores de cría: Aplicaciones para el mejoramiento genético de alpacas. Ediciones UNH. Huancavelica. Perú.
Safari, E., N.M. Fogarty, A.R. Gilmour, K.D. Atkins, S.I. Mortimer, A.A. Swan, F.D. Brien, J.C. Greeff y J.H.J. van der Werf. 2007. Genetic correlations among and between wool, growth and reproduction traits in Merino sheep. J. Anim. Breed. Genet., 124: 65-72.
Safari, E., N.M. Fogarty y A.R. Gilmour. 2005. A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep. Livest. Prod. Sci., 92: 271-289.
Van Vleck, L.D. 1993. Selection Index and Introduction to Mixed Model Methods. CRC Press. Florida, USA. 481 pp.
Wang, X., L. Wang y X. Liu. 2003. The Quality and Processing Performance of Alpaca Fibres: Australian Alpaca Fibre Industry and the Fibre properties. http://www.rirdc. gov.au/reports/RNF/03-128.pdf. [25 de septiembre de 2007].
Wang H.M., L. Xin y X. Wang. 2005. Internal Structure and Pigment Granules in Coloured Alpaca Fibers. Fibers and Polymers, 6: 263-268.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
111

Willis, M.B. 1996. Dalton's introduction to practical animal breeding. Editorial Blackewlll Science. Tercera edición. 159 pág.
Wuliji, T., G.H. Davis, K.G. Dodds, P.R. Turner, R.N. Andrews R.N. y G.D. Bruce. 2000. Production performance, repeatability and heritability estimates for live weight, fleece weight and fiber characteristics of alpacas in New Zealand. Small Rumin. Res., 37: 189-201.
Wurzinger M., J. Delgado, M. Nürnberg, A. Valle, A. Stemmer, G. Ugarte y J. Sölkner. (2006). Genetic and non-genetic factors influencing fibre quality of Bolivian llamas. Small Rumin. Res., 61: 131-139
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
112

ESTRATEGIAS PARA EL MEJORAMIENTO DE CAMÉLIDOS SUDAMERICANOS
Mueller,
Instituto Nacional de Tecnología Agropecuaria, Bariloche, Argentina. E-mail: [email protected]
ABSTRACT
All four South American camelid species are present in Peru but the most numerous is the alpaca (Lama pacos) with an estimated population of 3.5 million. Alpacas graze on native grasslands in high altitude and mostly in smallholder flocks. Alpacas provide essential goods and services to its keepers, mainly valuable textile fiber used for local handcrafts and exclusive garments produced in the country. Alpacas with heavy fleeces and fine fibers are preferred by breeders due to its impact on income. Therefore a number of genetic improvement initiatives have been proposed and implemented to improve fleece weight and reduce fiber diameter in Peru. However these efforts have generated little or no genetic improvement for these traits. In this chapter reasons for such failures are analyzed and elements for the formal design and implementation of breeding programs, breeding projects and breeding plans are presented. Intervention strategies are discussed such as the establishment of functional breeding structures. Also the importance of strategic breeding services is highlighted such as the availability of fleece testing laboratories, recording schemes and genetic evaluation expertise. In low input systems an intelligent balance of application of genetic principles and consideration of practical issues is needed. Many solutions to such practical problems may be found in successful programs and, most probably, by active involvement of participating breeders. Most programs need start-up funding but should be designed to be self-sufficient and sustainable in time. That is the key challenge for advisors and field geneticist involved in the organization of genetic improvement programs for camelids in Peruvian conditions.
KEY WORDS: Alpaca, Breeding, Genetics, Peru
INTRODUCCIÓN
Perú concentra aproximadamente 3 millones de las 3.5 millones de alpacas que existen en el mundo. La alpaca en el Perú aporta alimento y fibra a sus criadores además de servir para transporte, aportar estiércol usado como fertilizante, servir de caja de ahorro y cumplir roles relevantes en festividades y rituales. Por estas razones y por su vinculación histórica a los pueblos originarios, la alpaca constituye un acervo cultural fundamental contribuyendo a la identidad peruana. Se calcula que las alpacas del Perú producen anualmente 8.3 mil toneladas de carne que los productores destinan al consumo familiar o venden localmente y 3.5 mil toneladas de fibra que es transformada en artesanías o vendida a empresas textiles locales y extranjeras. Una proporción importante de la fibra de alpaca transformada es exportada como bobinas (tops), telas o prendas terminadas.
J. P.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
113

Con la importancia que la alpaca tiene en el Perú no sorprende que abunden las iniciativas para su mejoramiento genético. Estas iniciativas abarcan desde modestos planes a nivel predial hasta programas de gran escala a nivel nacional. Las metodologías propuestas para el mejoramiento genético también abarcan todo el rango de posibilidades, desde la selección basada en la inspección visual de los animales hasta el uso de información molecular y el empleo de sofisticadas tecnologías reproductivas para la multiplicación de los buenos animales. Sin embargo sorprende que tantas iniciativas no se vean reflejadas en un mejoramiento sensible de la productividad de los animales y en la calidad de la fibra producida (aunque muchas de las iniciativas se han iniciado hace pocos años atrás, por lo que sus resultados no son todavía mensurables). Es posible que los proyectos formales de mejora genética exitosos en realidad no sean tantos y más bien existen problemas en las propuestas y en la adopción de tecnologías de mejoramiento genético.
Aquí haremos un repaso de los elementos a considerar en el diseño del mejoramiento genético de pequeños rumiantes en general y analizaremos motivos por los cuales se dificulta la adopción de tecnologías de mejora genética de alpacas en el Perú. Se revisan algunas estrategias para implementar planes exitosos tomando en cuenta el resultado de experiencias previas con alpacas y otros pequeños rumiantes.
DESARROLLO DEL TEMA
Nivel de tratamiento del mejoramiento genético
Las consideraciones que pretendamos hacer sobre el mejoramiento genético dependen en primer lugar del nivel en el que se pretende intervenir. Por ejemplo podemos considerar el mejoramiento genético de las alpacas de todo el país y en el largo plazo, o podemos analizar las estrategias de mejoramiento de alpacas a nivel de una región o población de alpacas específicas, o también podemos restringirnos al análisis de los procedimientos de selección a nivel de un hato predial o comunal en particular. Estos tres niveles de enfoque suelen abordarse en términos de programas, proyectos y planes de mejora genética, respectivamente (Mueller 1999). En lo que sigue se repasan los elementos básicos a considerar en cada caso.
Elementos para un Programa de Mejora Genética
El programa de mejora genética puede ser interpretado como el marco de referencia que ordena la actividad a gran escala y en el largo plazo. En algunos países este ordenamiento se define y reglamenta en leyes o decretos pero también puede estar contenido en compromisos formales o informales de los
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
114

involucrados. En muchos casos son los organismos oficiales, ministerios o secretarias de agricultura y ganadería, los que tienen la responsabilidad general de la organización del mejoramiento genético en un país. En otros casos son las asociaciones de criadores u otras instituciones públicas o privadas que individualmente o en convenio conducen los programas de mejoramiento. En todo caso el programa debe expresar una finalidad o intención, debe delimitar la población de animales a mejorar y debe aclarar el rol de las organizaciones participantes. Por ejemplo la responsabilidad en los registros genealógicos, registros productivos, servicios de evaluación, etc.
El programa debe ser flexible y susceptible de ser revisado periódicamente para lo cual debería contemplar el establecimiento y la consolidación de un ámbito de discusión permanente. La experiencia indica que en tal ámbito deben participar los responsables directos del mejoramiento genético como así también especialistas en el tema, aparte de los correspondientes funcionarios. El Consejo Nacional de Camélidos Sudamericanos (CONACS) del Perú contemplaba muchos de los elementos mencionados mas arriba, sería deseable que el organismo que lo reemplaza tenga la estructura, continuidad y financiamiento adecuado para ello.
Elementos para un Proyecto de Mejora Genética
Los objetivos del programa deben ser abordados por proyectos específicos. El proyecto de mejora genética es el instrumento para modificar la situación en una región o población de animales determinada. Requiere la planificación de actividades considerando los recursos necesarios e implica básicamente tres tareas: (1) Describir la situación actual analizando la problemática relevante para detectar el cuello de botella por el cual no hay mejora genética. Para ello es necesario analizar las características de la estructura genética vigente (la caracterización de la estructura genética se basa en la clasificación del estrato, rendimiento productivo promedio, existencia de registros de producción y registros genealógicos, origen de los animales, etc.) y los procedimientos de selección utilizados. (2) Elaborar una estrategia de intervención. Esto implica analizar la necesidad de generar o consolidar una estructura genética determinada, seleccionar dentro de la población disponible o introducir animales de otras poblaciones o regiones, etc. (3) Describir la situación final con los resultados esperados.
La planificación del proyecto debe ser el resultado de diagnósticos participativos con las comunidades y productores involucrados. Aspectos críticos en la elaboración de proyectos de mejora genética tienen que ver con: (i) La especificación de la finalidad o consecuencia esperada del proyecto, por ejemplo a quien se pretende beneficiar con el proyecto; al productor, al criador, a
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
115

la comunidad. (ii) La definición de lo que se considera "mejora genética"; al aumento del nivel de progreso genético, al aumento de la tasa de progreso genético, al aumento de la población de animales mejorados, y en este último caso en qué estrato genético. (iii) El listado de insumos necesarios, por ejemplo: Plan de Mejora Genética (ver mas abajo) o plan de capacitación. (iv) Los productos a obtener, por ejemplo número de reproductores mejoradores, o sistema de evaluación y diseminación funcional. (v) Análisis de los supuestos (técnicos, económicos, sociales y políticos), es decir circunstancias que pueden hacer fracasar el proyecto.
Elementos para un Plan de Mejora Genética
El plan de mejora genética es un insumo del proyecto, debe explicitar el trabajo a realizar en el hato. Como cada hato tiene características especiales (infraestructura de manejo, necesidades de reposición, objetivos de mejora específicos, posibilidades de registros de producción, información genealógica, etc.) es posible que cada hato o unidad de mejoramiento requiera un plan específico. El diseño del plan requiere cumplir secuencialmente tres pasos: (1) la determinación del tipo de animal a criar o el objetivo de su mejora; (2) la elección de la información a utilizar o los criterios de selección a aplicar; y (3) el diseño del apareamiento de los animales seleccionados. Aquí no vamos a entrar en detalles sobre estos tres pasos ya que muchos de los aspectos conceptuales y prácticos para el mejoramiento genético de alpacas son tratadas adecuadamente en un libro peruano de reciente edición (Quispe Aragon 2007). Además los aspectos metodológicos específicos para determinar mérito genético también son tratados exhaustivamente en un excelente libro de referencia (Quispe y Alfonso 2007).
Iniciativas de mejora genética de alpacas en el Perú
Varias son las iniciativas para el mejoramiento genético de alpacas en el Perú. Por ejemplo en la Sierra Central, la Universidad Nacional Agraria La Molina (UNALM) conduce un proyecto basado en núcleos multi-comunales formados con las mejoras alpacas de las comunidades campesinas de la región. Así los machos nacidos en los núcleos de Huacahuaganan y de Ayaracra, Pasco se diseminan a las comunidades participantes. El proyecto es financiado en parte por el “Grupo Minero del Centro” y es un buen ejemplo de lo que es posible hacer con pocos recursos (Flores 2007, comunicación personal).
En la Sierra Central de Junín también trabaja organizadamente la SAIS Pachacutec en un plan de mejora de alpacas y llamas diferenciando colores y razas. La SAIS mantiene registros de producción, análisis de fibra y registros genealógicos en sus planteles y disemina machos en su propia población de
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
116

alpacas estimada en 10.000 animales y en todo el Centro del Perú (Terrel 2007, comunicación personal; Ayala y Chávez 2006). Del mismo modo la SAIS Tupac Amaru tiene un plan de mejora genética de sus alpacas.
La Universidad Nacional de Huancavelica (UNH) comenzó un amplio “Plan regional de mejoramiento genético de la alpaca en Huancavelica” que incluye un componente de mejora de la calidad genética con líneas de trabajo en selección, reproducción y difusión genética. El plan se basa en actividades de control de producción y control genealógico y aplicación de tecnologías moleculares y reproductivas de avanzada (Quispe, 2006).
En el Centro de Investigación y Producción (CIP) de Quimsachata, Puno el Instituto Nacional de Investigación y Extensión Agraria (INIA) conduce el proyecto “Mejoramiento genético de camélidos andinos mediante Biotecnología Reproductiva” que beneficiará, de manera directa y en mediano plazo, a 2.482 familias de las provincias de Lampa y Carabaya. La primera etapa del proyecto implica la conservación del germoplasma y biodiversidad genética de los camélidos, así como también, la implementación de un laboratorio de transferencia de embriones y la adquisición de 140 reproductores de alpacas y 28 de llamas de alta calidad genética. Este proyecto tendrá una duración de tres años y tres meses. Durante ese tiempo se prevé conservar 1.500 alpacas de 18 colores y 1.100 llamas de los tipos Q'ara y Ch'aku. El INIA también tiene como meta inseminar a 14.000 alpacas y transferir embriones de alta calidad genética a otras 1.900 alpacas (INIA 2006).
La Sociedad Peruana de Alpacas Registradas (SPAR) de Macusani ha encarado un proyecto de mejora genética basado en la estratificación de alpacas hembras y machos en un núcleo, apareando lo mejor con lo mejor y difundiendo reproductores de fibra fina. A principios de 2006 se establecieron dos núcleos de 150 hembras en Munay Paq'ocha e Itita del SPAR local que proveen reproductores a unos 10 centros de monta ubicados en microcuencas y a los cuales tienen acceso los comuneros. Choquehuanca (2006) resalta cuatro puntos importantes para el éxito del proyecto: productores organizados, concertación interinstitucional, laboratorio de fibras y apoyo de instituciones.
En Puno también hay un trabajo importante de Rural Alianza con sus aproximadas 40.000 alpacas. En un sistema estratificado por peso y calidad de vellón se producen machos para sus miembros. Algunos criadores individuales de la región están utilizando tecnologías genéticas y reproductivas de punta para lograr el mejoramiento de sus alpacas. Tal es el caso de “Pacomarca” del Grupo INCA (Burgos 2007, comunicación personal), “Mallkini” de la empresa Michell (Asperrin 2007, comunicación personal) y “Accoyo” de la familia Barreda.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
117

Con financiamiento del “Proyecto del Centro de Estudios y Promoción del Desarrollo” (DESCO) y en el marco del “Programa de Mejoramiento Genético” (PROMEGE) propuesto para la provincia de Caylloma, Arequipa, Paredes y Renieri (1998) diseñaron un interesante plan de selección para una población total de 18.000 alpacas. El plan se basa en el establecimiento de un núcleo abierto y un estrato de multiplicadores tomando en cuenta colores y razas de alpacas con un detallado plan de toma de datos y apareamientos planificados en cada estrato.
El CONOPA (2007) y el IVITA (Instituto Veterinario de Investigación y Altura) vienen también ofreciendo servicios y ejecutando planes de mejoramiento genético en alpacas, pero no ha sido posible conseguir información precisa de sus actividades en esta área.
Otros proyectos también tienen como finalidad la mejora genética de la alpaca. Por ejemplo un consorcio de instituciones liderado por el Instituto Peruano de Energía Atómica (IPEN) y la Universidad Nacional Cayetano Heredia ejecutan el proyecto “Use of Nuclear Techniques to Improve Alpacas Productive and Reproductive Methods” (Espinoza, 2006) con apoyo económico de la Agencia Internacional de Energía Atómica (IAEA). Por otro lado se formó otro consorcio de instituciones para ejecutar el “Proyecto Internacional Genoma de Alpaca” (Universidad San Martin de Porres, 2007).
Dificultades en la adopción de las propuestas de mejora
A pesar de que muchas de estas iniciativas son exitosas también existen muchas con problemas de implementación, continuidad y resultados concretos. Las causas de los fracasos son múltiples. En parte porque las alpacas está en manos de pequeños productores aislados geográficamente y con limitaciones severas para la adopción de las tecnologías y los conocimientos que exigen los proyectos propuestos. Otra causa de fracasos se debe a que muchos proyectos se organizan en torno a un financiamiento de corto plazo por lo que carecen de la continuidad y el seguimiento necesarios en un proyecto cuyos resultados se evidencian en el largo plazo. Pero la causa mas común es probablemente haber elegido la estrategia de intervención equivocada. Esto sucede cuando el proyecto no se basa en un diagnóstico y planificación con la participación de los propios interesados.
Esto queda claro en la opinión de los propios interesados. Una noticia publicada en Perulactea (2007), dice: “Los campesinos no creen en el mejoramiento genético a través de las técnicas científicas y utilizan la forma práctica y tradicional, mediante la selección de alpacas hembras y machos que tienen fibra de buena calidad. Luego cruzan a estas parejas de alpacas ... para obtener ejemplares que producen fibra más fina, en un proceso natural de selección. Los animales seleccionados y sus crías son inscritos en un registro de control de
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
118

calidad para garantizar la fibra al momento de ser comercializada y demostrar que no está mezclada. Resulta mucho más laborioso y difícil este tipo de selección genética, pero los campesinos de las comunidades criadoras de alpaca se resisten a las técnicas de inseminación artificial. Manifiestan que las técnicas científicas no dan resultado y además son muy caras, por lo que mantienen la tradición de mejoramiento tradicional ... Estos criadores le tienen desconfianza a las entidades que promocionan el mejoramiento e incluso proporcionan ayuda, como el CONACS o la SPAR”.
El dato relevante es que estos productores de las provincias Daniel Alcides Carrión y Pasco, utilizando sus técnicas “tradicionales”, manifiestan haber logrado aumentar la proporción de fibra extrafina producida del 7 al 16%. A los técnicos nos deben preocupar varios aspectos de esta opinión de los campesinos. Es preocupante que las diferentes técnicas de mejoramiento genético sean interpretadas como opuestas o competidoras cuando en realidad solo hay propuestas inteligentes y apropiadas a cada caso. Evidentemente el mensaje técnico no es entendido o no es el adecuado. La elaboración en conjunto de los proyectos puede evitar esos problemas y aumentar considerablemente el compromiso de todos en su ejecución. En ese proceso los técnicos podemos reconocer que las condiciones de los sistemas de producción exigen tecnologías más baratas y sencillas de las pensadas, pero que tienen la ventaja de poder ser operadas por los mismos productores, solo exigiendo capacitación y algún servicio estratégico. Ese tipo de proyectos tiene también una mayor probabilidad de ser sustentable en el tiempo y no depender siempre de financiamiento externo.
Por otro lado las dificultades que experimentan los productores en la aplicación de las técnicas propuestas muchas veces pueden ser resueltas por ellos mismos con conocimientos o estrategias que escapan a los propios técnicos. Estas habilidades y conocimientos pueden hacer la diferencia entre un proyecto viable o no viable, un ejemplo para ello proviene del proyecto de mejora de ovinos de la Sierra Central (Mueller et al. 2002) donde el núcleo se logró formar a sugerencia de los participantes mediante un ingenioso intercambio de ovejas por empadre con reproductores superiores. Experiencias de este tipo de conocimiento local (“local knowledge”) han sido descriptas por Galal et al. (2000) y Mueller (2006a). Obviamente también existen productores y hatos altamente tecnificados y que pueden aprovechar de tecnologías más sofisticadas para continuar su proceso de mejora genética.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
119

ESTRATEGIAS GENERALES
Generación y consolidación de una estructura genética
Aun suponiendo que todas las iniciativas de mejora genética son efectivas es obvio que son insuficientes para cubrir las necesidades de toda la población. Si suponemos que hay 2 millones de alpacas empadradas con un 5% de machos, que a su vez son reemplazados cada 5 años, entonces se necesitan 2.000.000*5%/5=20.000 machos mejoradores nuevos por año, una cifra muy lejana de ser alcanzada por los proyectos actuales e incluso futuros. Esos 20.000 machos podrían ser producidos en un estrato multiplicador. Suponiendo una tasa reproductiva efectiva del 64%, entonces la cantidad de hembras en el estrato multiplicador necesarias para producir esos machos es de al menos 20.000*2/0.64=62.500 y la cantidad de machos necesarios anualmente sería de 62.500*5%/5=625. Suponiendo una presión de selección del 50% las madres necesarias para producir esos 625 reproductores son 625*2*2/0.64=3.906. Estas madres conforman el estrato superior y deberían estar distribuidas en núcleos cubriendo las principales regiones productoras del país. El total de machos candidatos a selección en el estrato superior es de solamente 1250. Los mejores serán padres en el estrato superior y los siguientes en el estrato multiplicador. Estos números serian manejables pero exigen una estructura genética funcional.
Del análisis anterior surge que si se pretende un impacto a nivel de toda la población es estratégico generar y consolidar una estructura genética funcional. Esto no es tarea sencilla ya que implica acuerdo y colaboración interinstitucional. La SPAR puede tener un rol clave para ello, quizá desarrollando la estructura desde la base, primero a nivel de comunidad, luego a nivel regional y finalmente a nivel nacional. A su vez el estado a través de sus organismos de control y regulación debe velar para que los beneficios que genera el funcionamiento de la estructura se distribuyan equitativamente.
Servicios de análisis de muestras de fibras
El objetivo en el estrato superior de la estructura es el de maximizar el progreso genético. Para ello la estimación de mérito genético de los candidatos a selección debe ser lo más precisa posible. Aunque conceptualmente el mérito genético de los animales se puede estimar por simple apreciación visual, la medición permite aumentar considerablemente la precisión y en consecuencia permite aumentar el progreso genético. Es el caso del peso de vellón, peso corporal y fundamentalmente el diámetro de fibras. En ovinos se ha estimado que el uso de mediciones objetivas duplica la tasa de progreso genético obtenida con selección visual. La zona alpaquera tiene una geografía complicada y las comunicaciones son limitadas lo que se justifica la existencia de laboratorios ubicados en lugares
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
120

estratégicos (UNALM, UNH, Munay Paq'ocha, industrias, etc.). El desafío es que estos laboratorios produzcan resultados confiables, a tiempo para tomar decisiones de selección y a un precio acorde a la utilidad del resultado producido. También es importante que los diferentes laboratorios uniformicen los procedimientos de muestro, análisis y presentación de resultados, que permitan usar esta información para un servicio de evaluación genética de carácter regional o nacional.
Servicios de evaluación genética
Dos tipos de servicios de evaluación genética son fundamentales. El servicio de registros genealógicos, y el servicio de análisis de datos para producir estimaciones de merito genético. El primero se refiere a la inspección visual que hasta hace poco hacia personal del CONACS y que entre 2003 y 2005 logró inscribir a unas 8400 alpacas. En otros países este servicio lo suelen ofrecer las Asociaciones de Criadores, aquí podría ser la SPAR con sus delegaciones regionales. El servicio de estimación de mérito genético o “valor de cría” cobra importancia para aquellos núcleos que cuentan con registros de producción y datos productivos. En países desarrollados la evaluación genética es provista por internet. Por ejemplo en Australia y Nueva Zelanda funciona el “Low-cost alpaca across herd genetic evaluation project” (AGE 2007) y en los EEUU el “Genetic Alpaca Improvement Program” (GAIN 2007). Tanto AGE como GAIN ofrecen estimaciones de valores de cría a cambio de un arancel. Ayala (2006) propone un “Servicio de Evaluacion Genética” o EGA para el Perú.
En Argentina funciona el “Programa Nacional de Evaluacion Genética de Ovinos: Provino” (Mueller 2006) que reúne el servicio de análisis de muestras de lana con la provisión de valores de cría e índices de selección. Para ovinos de la raza Merino en Argentina también funciona el “Programa Merino Puro Registrado” (AACM 2007) por el cual animales que superan una inspección visual y superan en producción al promedio de su lote determinado por Provino, son registrados. Los criadores de alpacas del Perú se beneficiarían de un servicio equivalente. Gutierrez y Berger (2006) proponen desarrollar un servicio para alpacas al estilo del “Provino”.
Otros servicios en el futuro
La diferencia genética que se establece entre estratos de una estructura funcional es de aproximadamente 2 generaciones de mejora genética. Es decir que el nivel alcanzado en el núcleo llega al productor comunero con un retraso de 4 generaciones (16 años o más). Para achicar ese retraso y acceder a padres muy superiores puede ser útil la inseminación artificial y en menor medida la transferencia de embriones. En la medida en que se reduzcan los costos y mejore
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
121

la eficiencia de estas técnicas será importante contar con estos servicios. Del mismo modo los servicios para determinación de paternidad, “pureza” y presencia de marcadores genéticos. En algunos casos ya existen algunos de estos servicios en el contexto de proyectos de investigación. En todo caso estos servicios tendrán dimensión e impacto en la medida en que existan hatos organizados con planes de mejora eficientes en ejecución.
CONCLUSIONES E IMPLICANCIAS
El diseño y la implementación de programas, proyectos y planes de mejora para pequeños rumiantes en general deben basarse en el balance inteligente de la aplicación de principios genéticos y la consideración de problemas prácticos. Muchas soluciones a los problemas prácticos pueden ser encontradas en experiencias de otros programas y fundamentalmente pueden surgir de la participación activa de los propios productores participantes. Programas de mejoramiento para pequeños rumiantes en general requieren apoyo económico e intelectual externo importante en su fase inicial pero a mediano plazo deben ser conducidos y sostenidos por los interesados directos. Existen muchos ejemplos de programas que se abandonan una vez que se retira el circunstancial apoyo externo. El desafío para los asesores y genetistas de campo es la organización de programas adecuados a cada situación y sustentables en el tiempo. Para ello es estratégico consolidar una estructura genética funcional, lograr la participación activa de los beneficiarios en la planificación y ejecución de los proyectos, formar recursos humanos correspondientes y disponer de servicios estratégicos.
AGRADECIMIENTOS
A Gustavo Gutiérrez (Universidad Nacional Agraria La Molina, Perú), Karina Cancino, Gustavo Rebuffi y Francisco Rigalt (Instituto Nacional de Tecnología Agropecuaria, Argentina) por la lectura y valiosos aportes a un borrador de este trabajo.
LITERATURA CITADA
AACM 2007. Sitio oficial de la Asociación Argentina Criadores de Merino. http://www.merino.org.ar/index2.htm
AGE. 2007. http://www.alpaca.asn.au/AAA/age/age_intro.shtml
Ayala Paniura JE y Chávez Cossio JF. 2006. Mejoramiento Genético de Fibra de Alpaca Huacaya mediante una estrategia de capacitación y fortalecimiento institucional de productores. Presentación al Technical Workshop to define breeding strategies for South American Camelids. IAEA, 31 mayo a 2 junio, Lima, Perú.
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
122

Ayala Paniura JE. 2006. Estrategia nacional para el desarrollo genético de los camélidos sudamericanos domésticos con énfasis en alpacas. Consultoría dirigida al Programa de Camélidos Domésticos del CONACS, 28 p.
Choquehuanca Z. 2006. SPAR Macusani. Presentación al Technical Workshop to define breeding strategies for South American Camelids. IAEA, 31 mayo a 2 junio, Lima, Perú.
CONOPA. 2007. http://www.conopa.org/proyectos.php#
Espinoza JR. 2006. Genómica de Alpacas: Identificación de Genes Expresados y Marcadores Genéticos Asociados a la Productividad de Fibra Fina en alpacas. Presentación al Technical Workshop to define breeding strategies for South American Camelids. IAEA, 31 mayo a 2 junio, Lima, Perú.
GAIN 2007. http://www.alpacagenetics.com/
Galal S, Boyazoglu J and Hammond K (eds.). 2000. Workshop on developing breeding strategies for lower input animal production environments. ICAR Technical Series No. 3 p. 435-443.
Gutierrez GA y Berger PJ. 2006. Uso de la selección en el mejoramiento de la producción de camélidos sudamericanos. Presentación al Technical Workshop to define breeding strategies for South American Camelids. IAEA, 31 mayo a 2 junio, Lima, Perú.
INIA. 2006. http://www.inia.gob.pe/notas/nota0119/
Mueller JP, Flores ER and Gutierrez G. 2002. Experiences with a large-scale sheep genetic improvement project in the Peruvian highlands. En: Proceedings VII World Congress on Genetics Applied to Livestock Production, Montpellier, France 19-23 August, Communication 25-12.
Mueller JP. 1999. Diseño e implementación de programas de mejoramiento genético de ovinos. Conferencia invitada. Seminario Internacional de Mejoramiento Genético Ovino. Universidad Nacional Agraria La Molina, Lima, Perú 18 de junio. Comunicación Técnica INTA Bariloche Nro PA 356.
Mueller JP. 2006. Avances en el mejoramiento genético de ovinos. Congreso Argentino de Producción Animal, Mar del Plata, Buenos Aires, Argentina 20 de octubre. Comunicación Técnica INTA Bariloche Nro PA 493.
Mueller JP. 2006a. Breeding and conservation programs with local communities. FAO - WAAP Expert Meeting “Sustainable Utilization of Animal Genetic Resources”, Ferentillo, Italy, 2-4 July 2006. Comunicación Técnica INTA Bariloche Nro PA 489.
Paredes MG y Renieri C. 1998. Propuesta de un plan de selección de la población de alpacas en la provincia de Caylloma, Arequipa. En: Frank E, Renieri C y Lauvergne JJ (eds): “Actas del tercer Seminario de Camélidos Sudamericanos Domésticos y Primer Seminario Proyecto SUPREME”, Universidad Católica de Córdoba, Argentina, p. 27-38.
Perulactea. 2007. http://www.perulactea.com/noticias_detalle.php?art_id=2238
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
123

Quispe Aragón A. 2007. Aspectos prácticos para el mejoramiento de alpacas. Sociedad Peruana de Criadores de Alpacas, Macusani, Puno, Perú, 243 p.
Quispe, EC y L. Alfonso. 2007. Metodologías para estimar los valores de Cría (EVC). Aplicaciones para el mejoramiento genético de alpacas. Universidad Nacional de Huancavelica, Perú, 296 p.
Quispe, EC. y L. Alfonso 2006. Propuesta de plan regional de mejoramiento genético de la alpaca en Huancavelica. Presentación al Technical Workshop to define breeding strategies for South American Camelids. IAEA, 31 mayo a 2 junio, Lima, Perú.
Universidad San Martin de Porres. 2007. http://www.universia.edu.pe/noticias/principales/ destacada.php?id=59429
Actualidades sobre adaptación, producción, reproducción y mejora genética en camélidos
124


Esta obra se terminó
de imprimir en Octubre del 2008 en
EDICION Gráfica Industrial E.I.R.L.
Jr. Cusco 421 - Huancayo - Perú
Telf. 211437





























