Embed Size (px)
Citation preview

Acylsugar Acylhydrolases: Carboxylesterase-CatalyzedHydrolysis of Acylsugars in Tomato Trichomes1[OPEN]
Anthony L. Schilmiller2*, Karin Gilgallon3, Banibrata Ghosh, A. Daniel Jones, and Robert L. Last
Department of Biochemistry and Molecular Biology (A.L.S, K.G., B.G., A.D.J., R.L.L.), Department ofChemistry (A.D.J.), and Department of Plant Biology (R.L.L.), Michigan State University, East Lansing,Michigan 48824–1319
ORCID IDs: 0000-0001-7069-7120 (A.L.S.); 0000-0002-7408-6690 (A.D.J.); 0000-0001-6974-9587 (R.L.L).
Glandular trichomes of cultivated tomato (Solanum lycopersicum) and many other species throughout the Solanaceae produce andsecrete mixtures of sugar esters (acylsugars) on the plant aerial surfaces. In wild and cultivated tomato, these metabolites consist ofa sugar backbone, typically glucose or sucrose, and two to five acyl chains esterified to various positions on the sugar core. Thealiphatic acyl chains vary in length and branching and are transferred to the sugar by a series of reactions catalyzed by acylsugaracyltransferases. A phenotypic screen of a set of S. lycopersicum M82 3 Solanum pennellii LA0716 introgression lines identified adominant genetic locus on chromosome 5 from the wild relative that affected total acylsugar levels. Genetic mapping revealed thatthe reduction in acylsugar levels was consistent with the presence and increased expression of two S. pennellii genes(Sopen05g030120 and Sopen05g030130) encoding putative carboxylesterase enzymes of the a/b-hydrolase superfamily. Thesetwo enzymes, named ACYLSUGAR ACYLHYDROLASE1 (ASH1) and ASH2, were shown to remove acyl chains from specificpositions of certain types of acylsugars in vitro. A survey of related genes in M82 and LA0716 identified another trichome-expressed ASH gene on chromosome 9 (M82, Solyc09g075710; LA0716, Sopen09g030520) encoding a protein with similaractivity. Characterization of the in vitro activities of the SpASH enzymes showed reduced activities with acylsugars producedby LA0716, presumably contributing to the high-level production of acylsugars in the presence of highly expressed SpASH genes.
Hundreds of thousands of specialized metabolitesare found in plants, and this large number of structur-ally diverse compounds results from the actions of avariety of enzyme classes, many of which appear toevolve rapidly (Pichersky and Lewinsohn, 2011; Miloand Last, 2012). Acylation of oxygen-containing func-tional groups to form esters is widespread in metabo-lism, particularly in plant specialized metabolism. Theformation of ester-linked derivatives can alter thechemical and physical properties of a molecule, leadingto changes in stability, localization, and solubility, andmany studies have focused on the enzymes that cata-lyze their formation (for review, see D’Auria, 2006;Mugford and Milkowski, 2012). Reverse reactions that
hydrolyze ester linkages in metabolism are also prev-alent, and the responsible enzymes are referred to ascarboxylesterases (CXEs). CXE proteins are classifiedinto three groups based on sequence. Class I and IICXEs contain enzymes belonging to the a/b-hydrolasesuperfamily, while class III proteins belong to the GDSlipase family of enzymes (Gershater and Edwards,2007). Plant CXEs play a role in the modification ofesters of the signaling molecules jasmonic acid, salicylicacid, and auxin (Stuhlfelder et al., 2004; Forouhar et al.,2005; Yang et al., 2008), regulation of metabolite trans-port and localization (Kandzia et al., 1998), deprotectionof functional groups in alkaloid biosynthesis (Ruppertet al., 2005; Winzer et al., 2012), bioactivation of herbi-cides (Gershater et al., 2007), and protection againstpathogen infection (Pontier et al., 1994).
Plant class I CXE proteins are characterized ascontaining Interpro domain IPR013094 (Mitchell et al.,2015), have a catalytic triad of Ser, His, andAsp, and aretypically inhibited by organophosphates. Twenty classICXEgeneswere described fromArabidopsis (Arabidopsisthaliana; Marshall et al., 2003), and a similar number ofgenes were found in apple (Malus domestica) EST col-lections (Souleyre et al., 2011). Only a few plant classI CXEs have been biochemically characterized. SlCXE1from cultivated tomato (Solanum lycopersicum) hydro-lyzes volatile acetate esters in fruit, influencing aro-matic flavor components (Goulet et al., 2012). Similarly,the apple MdCXE1 protein is expressed during fruitripening and is thought to hydrolyze acetate esters
1 This work was supported by the National Science Foundation(grant no. IOS–1025636 to A.D.J. and R.L.L. and Major Research In-strumentation grant no. DBI–0619489).
2 Present address: RTSF Mass Spectrometry and MetabolomicsCore, Michigan State University, East Lansing, MI 48824–1319.
3 Present address: Zoetis, Inc., Lincoln, NE 68521.* Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:Anthony L. Schilmiller ([email protected]).
A.L.S., K.G., and B.G. designed and performed experiments; A.L.S.,K.G., B.G., A.D.J., and R.L.L. analyzed the data; A.L.S. and R.L.L.wrote the article.
[OPEN] Articles can be viewed without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.15.01348
Plant Physiology�, March 2016, Vol. 170, pp. 1331–1344, www.plantphysiol.org � 2016 American Society of Plant Biologists. All Rights Reserved. 1331
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

important for fruit flavor (Souleyre et al., 2011). Apoppy (Papivar somniferum) gene, PsCXE1, is part of acluster of genes encoding enzymes for noscapine bio-synthesis (Winzer et al., 2012); this enzyme is suggestedto catalyze the hydrolysis of an acetyl group frompapaveroxine to form narcotine hemiacetal. Two plantclass I CXE enzymes were shown to catalyze in vitroreactions that differ from the typical hydrolytic cleav-age expected from CXEs: hydroxyisoflavanone dehy-dratase from soybean (Glycine max) and Glycyrrhizaechinata catalyzes the 1,2-elimination of water fromhydroxyisoflavanones (Akashi et al., 2005), while a tu-lip (Tulipa gesneriana) CXE catalyzes an intramoleculartransesterification reaction resulting in the formation ofan antimicrobial lactone product from the Glc esterof 4-hydroxy-2-methylenebutanoate (Nomura et al.,2012). The GA receptor GID1 is a class I CXE that haslost the ability to catalyze hydrolysis reactions and in-stead functions by binding GA, ultimately leading tothe ubiquitination and degradation of transcriptionalrepressors of GA signaling (Ueguchi-Tanaka et al.,2005). Lastly, there are plant class I CXEs associatedwith certain plant responses, but the endogenous sub-strates remain unknown. For example, the ArabidopsisCXE12 protein was demonstrated to convert exoge-nously applied methyl-2,4-dichlorophenoxyacetate tothe active herbicide 2,4-dichlorophenoxyacetic acid(Gershater et al., 2007). Tobacco (Nicotiana tabacum) andpepper (Capsicum annuum) class I CXE genes are in-duced upon pathogen infection and, while the endog-enous substrates are not known, are thought to participatein the hypersensitive response (Baudouin et al., 1997; Koet al., 2005). Although many class I CXEs remain to beanalyzed, all of the enzymatically active class I CXEs fromplants with well-characterized activities function in spe-cialized metabolism.
Many plants throughout the nightshade family (Sola-naceae) have glandular trichomes that produce and se-crete specialized metabolites, including polyacylatedsugar esters known as acylsugars (Schilmiller et al., 2008).Due to their presence on the surface of aerial tissues, theseacylsugars are positioned to provide resistance againstinsect herbivores (Simmons and Gurr, 2005; Weinholdand Baldwin, 2011). Acylsugars and related compoundsalso have commercial and medicinal uses as surfactants(Hill and Rhode, 1999; Dembitsky, 2004), antimicrobialcompounds (Chortyk et al., 1993; Ovenden et al., 2005),and antiinflammatory compounds (Pérez-Castorenaet al., 2010). These varied properties pique interest inunderstanding how acylsugars are produced in tri-chomes and how their levels are regulated.
Within the tomato clade of the genus Solanum,acylsugars consist of a sugar backbone, either Glc orSuc, to which two to five aliphatic acyl chains are es-terified. The acyl chains range in length from two to13 carbons and may be either straight or branched. Thetypes of sugar backbones as well as the lengths andbranching of acyl chains on acylsugars vary widelyamong the species of the Solanaceae and also can dis-play significant variation within accessions of a single
species (Kroumova and Wagner, 2003; Kim et al., 2012;Ning et al., 2015; Schilmiller et al., 2015). We recentlydemonstrated that the biosynthesis of acylsucroses incultivated and wild (Solanum pennellii) tomato involvesBAHD-type acyltransferases that catalyze the transfer ofacyl chains froman acyl-CoAdonor. In theS. lycopersicum,acylsucrose assembly begins with the addition of afive-carbon acyl chain to the pyranose ring of Sucby the enzyme ACYLSUGAR ACYLTRANSFERASE1(ASAT1) followed by the addition of a second pyranosering acyl chain by ASAT2 (Fan et al., 2016). ASAT3 canadd an acyl chain to the furanose ring of diacylsucrosesor add an acyl chain to the pyranose ring of amonoacylsucrose acceptor, depending on the isoform(Schilmiller et al., 2015). ASAT4 adds an acetyl group tothe pyranose ring of a triacylsucrose acceptor (Kimet al., 2012; Schilmiller et al., 2012).
We report the identification of class I CXE familyenzymes from cultivated and wild tomato, which wenamed acylsugar acylhydrolases (ASHs). Introgressionof a dominant locus containing twoASH genes from thewild tomato S. pennellii LA0716 into the S. lycopersicumM82 genetic background results in reduced levels ofacylsugars (Schilmiller et al., 2010a). The dominant in-heritance of the LA0716 locus is due to replacement ofan inactive SlASH2 gene with a highly expressed andfunctional SpASH2 in IL5-3 plants. RNA interference(RNAi)-mediated suppression of the SpASH alleles inthe IL5-3 introgression line caused the restoration ofacylsugar levels, documenting their in planta function.In vitro assays with recombinant ASH proteins showedtheir ability to catalyze the hydrolysis of ester-linkedacyl chains from specific positions of acylsugars.Assays with LA0716 ASH enzymes demonstrated re-duced activity with acylsucroses produced by LA0716compared with high activity against the structurallydifferent acylsucroses produced by M82 trichomes.These results illustrate a mechanism for a negative-effect locus from S. pennellii LA0716 that should beavoided for breeding increased acylsugar levels andsuggest the possibility of a mechanism for the turnoveror remodeling of acylsugar molecules in trichomes.
RESULTS AND DISCUSSION
Identification of an S. pennellii Locus That CausesReduced Acylsugars
Genetic Mapping of the IL5-3 Low-Acylsugar Trait
Previous analysis of trichome-specialized metabo-lites in the S. lycopersicum M82 3 S. pennellii LA0716introgression lines identified a region on chromosome 5that influences total acylsugar levels (Schilmiller et al.,2010a). The parental S. lycopersicum varM82 produces avariety of triacylsucroses and tetraacylsucroses (Fig. 1;Ghosh et al., 2014). In contrast, IL5-3 leaves accumu-lated approximately 30-fold lower levels of acylsugarsthan wild-type M82 plants (Fig. 1, B and D). Theeffect on acylsugar levels was not equivalent for all
1332 Plant Physiol. Vol. 170, 2016
Schilmiller et al.
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

acylsucroses: IL5-3 had a larger reduction in short(C5 or less) acyl chain-containing acylsugars comparedwith acylsugars containing a long (C10 or more) acylchain (Fig. 1E). Nonacylsugar metabolites that areextracted by the leaf-dip method employed, includingrutin and tomatine, were not affected in IL5-3.To understand the genetic basis for reduced acylsugars
in IL5-3 plants, we crossed IL5-3 with the M82 parent.The F1 plants showed the reduced acylsugar phenotype,consistent with the S. pennellii locus being dominant.F2 progeny were generated, and the low:high progeny
showed a ratio of 2.9:1 (x2 = 0.266, n = 1,055), as expectedfor a dominant monogenic trait. The F2 progeny wereused to perform fine-mapping to narrow the locus fromthe approximately 48-Mb IL5-3 span, which waspredicted to encode 1,099 genes (Bombarely et al., 2011;Chitwood et al., 2013), to a region of approximately 129kb (Fig. 2A). The annotation of the S. lycopersicum Heinz1706 genome assembly (Sol Genomics Network [SGN;http://solgenomics.net]; Tomato Genome Consortium,2012) included 17 predicted open reading frames (ORFs)within the mapping interval (Supplemental Table S1).
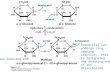
Figure 1. M82 and IL5-3 acylsugarphenotypes. A and B, Base peakintensity (BPI) liquid chromatography-mass spectrometry (LC-MS) chromato-grams for M82 (A) and IL5-3 (B) showreduced levels of acylsugars in IL5-3plants. Levels of rutin (flavonoid gly-coside) and tomatine (glycoalkaloid)are similar for M82 and IL5-3. C,Acylsugar profile of LA0716, whichproduces an abundance of bothacylglucoses (*) and acylsucroses (!).Base peak intensity chromatograms areplots of the abundance of the mostabundant ion in each scan. The number9.00e5 at the top right of each chro-matogram indicates the ion countrepresented by 100% relative abun-dance. IS, Internal standard. D, Totalacylsugar peak areas for [M+formate]–
ions from extracted ion chromatogramsfor LA0716, M82, and IL5-3. Error barsindicate SD from three biological repli-cates. E, Ratio of total peak area foracylsugars containing only short(C5 or less) acyl chains to acylsugarscontaining one long (C10 or more) acylchain. The reduced ratio for IL5-3 in-dicates a stronger reduction in shortchain-containing acylsugars comparedwith acylsugars having a long chain. F,Structures of the most abundantacylsugars of M82 and a generalacylsucrose structure showing positionnumbering for reference. Acylsugarnomenclature, using S3:15 (5,5,5) asan example, is as follows: S indicatesan acylsucrose, 3 indicates three acylchains, 15 indicates 15 total carbons inthe three acyl chains, and in paren-theses are listed the lengths of eachindividual acyl chain.
Plant Physiol. Vol. 170, 2016 1333
Tomato Acylsugar Acylhydrolases
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

Two Genes from This Region Are Expressed in Trichomes
To narrow this list of candidate genes further, welooked for evidence of expression in S. lycopersicum M82and S. pennellii LA0716 trichomes using available tri-chome RNA sequencing (RNAseq) data (Schilmilleret al., 2010b;McDowell et al., 2011;Ning et al., 2015). Fourgenes in the interval showed no evidence of trichomeexpression, and another nine had only a small number ofreads (less than 0.005% of the total reads in the library;reads per kilobase of transcript permillion readsmapped, 10) mapped to each gene, indicating a low level ofexpression in trichomes (Supplemental Table S1). Twogenes in the region, Solyc05g051660 (Sopen05g030120)and Solyc05g051670 (Sopen05g030130; renamed ASH1and ASH2, respectively), had large numbers of reads inthe LA0716 trichome complementary DNA (cDNA) li-brary but low read counts in M82 trichome cDNA(Supplemental Table S1). M82 RNAseq data using iso-lated trichomes from stem tissue and stem tissue afterremoval of trichomes were analyzed to determine
relative levels of expression in trichome versusnontrichome tissue (Ning et al., 2015). The expression ofboth SlASH1 and SlASH2 was enriched in trichome tis-sue, with 10-fold (SlASH1) and 127-fold (SlASH2) higherexpression in trichomes versus stems and petioles afterremoval of the trichomes. Quantitative reverse tran-scription (RT)-PCR using the same cDNA validated thetrichome-enriched expression of SlASH1 and SlASH2(Supplemental Fig. S1). Although SlASH1 expression ishighest in trichome tissue, the expression is not trichomespecific, in contrast to what is seen for the ASAT3 andASAT4 genes, which encode proteins that catalyze thetransfer of acyl chains to form acylsucroses (Schilmilleret al., 2012, 2015). A SlASH1 unigene (SGN-U566465) ispresent in the SGN database, and it consists of nine ESTsfrom various cDNA libraries, including trichomes (one),whole plants (two), developing flower buds (two), sus-pension cell cultures (two), and seeds (two). Flower budsand whole plants contain trichomes and would be ex-pected to show the expression of trichome-specific tran-scripts; however, the ESTs from suspension cells and
Figure 2. Mapping of the IL5-3 locus and relativeexpression of ASH transcripts in trichomes of M82and IL5-3. A, A positional cloning approach using F2recombinants from an M823 IL5-3 cross was used tonarrow the causative locus to a region spanning ap-proximately 129 kb. Markers are shown, with thenumber of recombinants in parentheses. A subset ofthe 129-kb region containing ASH1/2 is shown for theS. lycopersicumHeinz 1706 and S. pennellii LA0716assemblies. The triangle after SpASH2 indicates atransposon insertion in the LA0716 genome at thisposition. B, ASH transcript levels were measured byquantitative RT-PCR in trichomes of M82 and IL5-3.The SpASH2 gene is expressed at higher levels in theIL5-3 trichomes compared with the trichome ex-pression level in M82 for SlASH2, which is predictednot to encode a functional protein. Error bars indicateSD among three biological replicates.
1334 Plant Physiol. Vol. 170, 2016
Schilmiller et al.
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

seeds suggest the possibility of additional non-trichome-localized functions for SlASH1. NoESTs for SlASH2werefound in any tomato cDNA library in the SGN database.The sequences ofASH1 andASH2 inM82 and LA0716
were examined more closely to understand how theycould influence acylsugar levels in IL5-3. A BLASTsearch showed that both SlASH1 and SlASH2 encodeproteins in the carboxylesterase subgroup of thea/b-hydrolase fold superfamily based on the presenceof Interpro domain IPR013094 (Hotelier et al., 2004). TheSlASH1 ORF is 969 bp with no introns and is predictedto encode a protein with 322 amino acids. The currentannotation of the tomato genome (ITAG2.4; http://solgenomics.net) indicates that SlASH2 has two intronsand is predicted to result in an 855-bp ORF encoding aprotein with 284 amino acids. The S. pennellii LA0716genomic sequence shows that both SpASH1 and SpASH2lack introns and are predicted to encode proteins of323 amino acids in length. Alignment of S. lycopersicumand S. pennellii ASH genomic sequences suggests that theSlASH2 gene is likely to be inactive, because it harborsa 31-bp deletion and a 4-bp insertion that disrupts itspredicted ORF (Supplemental Fig. S2). In addition,SlASH2 has 59 single-nucleotide polymorphisms thatare unique to SlASH2 (compared with SpASH2 andboth ASH1 sequences), whereas SlASH1 has only eightunique single-nucleotide polymorphisms, suggestingthat the SlASH2 sequence has been diverging rapidly,as seen commonly for pseudogenes.In addition to the presence of a functional SpASH2
gene in IL5-3, a published RNAseq analysis of gene ex-pression in the S. pennellii introgression lines suggestedthat increased expression of the ASHs in IL5-3 may alsocontribute to the low-acylsugar phenotype. An approx-imately 3- to 4-fold increase was observed for the ex-pression of SpASH1 and SpASH2 in IL5-3 13-d-oldseedlings comparedwith that of other introgression linescontaining the SlASH alleles (Chitwood et al., 2013). Tovalidate these results and test whether this expressiondifference is also observed in trichomes, quantitativePCR (qPCR) was performed using cDNA from isolatedtrichomes collected from stems and petioles of 3-week-old M82 and IL5-3 plants. The SpASH2 transcript levelswere approximately 13-fold higher in IL5-3 trichomeRNA comparedwith SlASH2 inM82 trichomes (Fig. 2B).In contrast, SpASH1 and SlASH1 transcripts accumu-lated to similar levels in trichomes (Fig. 2B). Takentogether, these results led to the hypothesis that the low-acylsugar phenotype in IL5-3 results from introgressionof the more highly expressed SpASH2, replacing themutated SlASH2 gene in the M82 genome. This pro-posed mechanism is consistent with the dominance ofthe S. pennellii locus over the S. lycopersicum M82 locus.
ASH1 and ASH2 Proteins Have AcylsugarAcylhydrolase Activity
Based upon the homology of ASH1 and ASH2 tocarboxylesterases, we tested the hypothesis that the
IL5-3 SpASH enzymes hydrolyze S. lycopersicumacylsucrose acyl chains. The SlASH1, SpASH1, andSpASH2 enzymes were expressed in Escherichia colias 63 His-tagged fusion proteins and assayed usingS. lycopersicum acylsugars as substrates. The triacylsucrosesof S. lycopersicum have acyl chains at the R3, R4, and R39positions, with tetraacylsucroses having an additionalacetyl group at the R2 position; the major acylsugarsin M82 trichome extracts include S3:15 (5,5,5), S4:17(2R2,ai5R3,i5R4,i5R39), S3:22 (ai5R3,i5R4,n12R39), and S4:24(2R2,ai5R3,i5R4,n12R39) (Schilmiller et al., 2010a, 2012;Ghosh et al., 2014). Acylsugar nomenclature is as follows:usingS3:15 (5,5,5) as an example, S indicates anacylsucrose,3 indicates three acyl chains, and 15 indicates 15 totalcarbons in the three acyl chains. The length of each in-dividual acyl chain is in parentheses, with superscriptsindicating the position of the acyl chain, ai = anteisobranched, i = iso branched, and n = normal straightchain (Fig. 1E). Because we observed differences in theimpact of IL5-3 on the accumulation of short and longchain-containing acylsucroses, the M82 acylsugarmixture was separated into two fractions prior to theiruse as substrates. Fraction I contained triacylsucrosesand tetraacylsucroses having only short acyl chains(C5 or less), while fraction II consisted of triacylsucrosesand tetraacylsucroses having a single long acylchain (C10–C12).
ASH1 and ASH2 Hydrolyze the R3 Position of theSix-Membered Ring of Fraction II
Our results demonstrate that the ASH enzymes hy-drolyzed the long chain-containing triacylsucroses andtetraacylsucroses at the R3 position (Fig. 3). Incubationof these two M82 acylsugar fractions with SlASH1,SpASH1, or SpASH2 resulted in the production of new,earlier-eluting peaks, whichwere identified as S2:10 (5,5)and S3:12 (2,5,5) based on fragmentation in negative-ionmode liquid chromatography-time of flight-massspectrometry (Fig. 3; Supplemental Fig. S3). Thesediacylsucrose and triacylsucrose products were not de-tectable in either of the substrate fractions and, thus,result from hydrolysis of acyl chains from thetriacylsucroses and tetraacylsucroses. That the R3 posi-tion acyl chain that was removed was most clearly ob-served in the reactions using the long chain-containingacylsucrose substrate fraction II (Fig. 3, A–D): the S2:10(5,5) and S3:12 (2,5,5) products that were observed couldonly result from hydrolysis of the C12 acyl chain of S3:22(5,5,12) and S4:24 (2,5,5,12), respectively, or from hy-drolysis of C10 from S4:22 (2,5,5,10). No hydrolysisproducts were detected that had a C5 acyl chain re-moved and the long acyl chain remaining in theASH1 orASH2 reaction when fraction II was used as a substrate.
ASH2 Hydrolyzes the R3 Position of the Six-Membered Ringof Fraction I
When fraction I, containing short-chain acylsucrosesfromM82,was tested as a substratewith SpASH2, S2:10
Plant Physiol. Vol. 170, 2016 1335
Tomato Acylsugar Acylhydrolases
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

(5,5) and S3:12 (2,5,5) again were the major productsobserved for each reaction (Fig. 3, E–H). These productsresult from hydrolysis of a single C5 acyl chain fromS3:15 (5,5,5) and S4:17 (2,5,5,5), respectively, or fromhydrolysis of a C4 acyl chain from S4:16 (2,4,5,5). De-termination of the position of hydrolysis is lessstraightforward with these substrates than with long-chain acylsugar fraction II, due to the presence ofmultiple C5 chains, but consideration of additionalLC-MS information helped to narrow the likely position.Fragmentation of these products in positive-ion mode,which results in cleavage of the glycosidic bond be-tween the pyranose (six-membered) and furanose (five-membered) rings of the acylsucrose (Ghosh et al., 2014),showed that each product contained a C5 acyl chain onthe furanose ring (Supplemental Fig. S4). This indicatesthat the hydrolyzed acyl chainwas on the pyranose ring.The S2:10 and S3:12 products produced by the ASH2-fraction I reaction coeluted with S2:10 and S3:12 from
reactions using fraction II, the long chain-containingsubstrate mixture. Taken together, these data are con-sistent with the hypothesis that SpASH2 catalyzed asingle cleavage, at the R3 position of short chain-containing acylsucroses.
ASH1 Cleaves a Single Acyl Chain from the R3 or R4Position of the Six-Membered Ring of Fraction I
Interpretation of the results from the SlASH1 andSpASH1 enzymatic reactions is more complicated. TheS2:10 and S3:12 peaks arising from R3 hydrolysis werealso seen with these enzymes; however, a second, later-eluting S3:12 (2,5,5) peak also was observed fromthe fraction I substrates. Positive-ion mode massspectrometry analysis revealed that this productalso retained the R39 furanose ring C5 acyl chain(Supplemental Fig. S4). The presence of two chro-matographically separable S3:12 (2,5,5) peaks in the
Figure 3. Evidence that ASH1 and ASH2 hydrolyze acyl chains from the R3 or R4 position of M82 acylsucroses. Base peak in-tensity (BPI) LC-MS chromatograms are shown for reactions with the ASH1 and ASH2 enzymes using M82 long chain-containing(fraction II) and short chain-containing (fraction I) acylsucrose substrates. Specific positions of acyl chains are given as superscriptswhen known from NMR-resolved structures (Ghosh et al., 2014); otherwise, only acyl chain lengths are given. IS, Internalstandard. Peak labels for ASH products indicate the acyl chain that was hydrolyzed in the reaction. A, Fraction II substrate. B,SlASH1 + fraction II. C, SpASH1 + fraction II. D, SpASH2 + fraction II. E, Fraction I substrate. F, SlASH1 + fraction I. G, SpASH1 +fraction I. H, SpASH2 + fraction I. Structures of acylsucrose substrates and their reaction products are shown in I. The scale ofchromatogram signals at 100% (representing the most abundant peak) is indicated by the number at the top right corner of eachchromatogram.
1336 Plant Physiol. Vol. 170, 2016
Schilmiller et al.
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

ASH1 samples is most simply explained as the twopositional isomers, (2R2,5R3,5R39) and (2R2,5R4,5R39),resulting from hydrolysis of S4:17 (2R2,5R3,5R4,5R39) atdifferent positions (Fig. 3; Supplemental Fig. S3). Al-ternatively, the two S3:12 peaks could correspond totwo C5 acyl chain isomers, present as either terminallybranched (iso) or subterminally branched (anteiso)chains. However, this scenario is unlikely, because themagnitude of the retention time difference for the twoS3:12 peaks suggests a more significant difference instructure than simply branch point isomers of an acylchain. Clear evidence for R4 hydrolysis by ASH1 comesfrom results shown below for a related hydrolase(ASH3) that cleaves esters at the R4 position. The later-eluting S3:12 product of ASH1 cochromatographs withthe ASH3 S3:12 product. Our results suggest that thelength of the acyl chain at the R3 position influencesASH1 hydrolysis of the R4 position. This is because theASH1 enzymes did not produce S3:19 (2R2,12R3,5R39) orS2:17 (12R3,5R39), which are the expected products of R4hydrolysis of the fraction II substrates. The activity ofASH1 hydrolyzing multiple positions of the shortchain-containing acylsugars is consistent with the ob-served IL5-3 phenotype of substantially lower levels ofthe short chain-containing acylsugars compared withthe long chain-containing acylsugars. Taken together,our data indicate that the ASH1 and ASH2 enzymes arecarboxylesteraseswith hydrolytic activity for the cleavageof acyl chains from specific positions of triacylsucrosesand tetraacylsucroses.
Reducing the Expression of SpASH1/2 in IL5-3 RestoresAcylsugar Levels
RNAi transgenic line phenotypes were analyzed totest the hypothesis that SpASH introgression causedreduced acylsugar levels. Taking advantage of the obser-vation that SpASH1 and SpASH2 are 95% identical at thenucleotide level, we designed an RNAi construct aimed atsilencing both genes (Supplemental Fig. S2). Eight of tenindependent transgenic IL5-3 lines transformed with thisconstruct showed increased acylsugar levels, with twohaving wild-type M82 levels (Fig. 4A). The ratio ofacylsugars having all short chains to acylsugars havinga long acyl chain also was restored to wild-type M82levels in seven of 10 transgenics (Supplemental Fig. S5).The restoration of the short-to-long chain-containingacylsugar ratio and the lack of any new acylsugar in-termediates in the RNAi lines suggest that ASH1/2probably act first followed by other hydrolases thatfurther degrade acylsugars. Nine independent trans-genic lines in the IL5-3 background were recoveredusing an RNAi construct targeting SpASH2 specifically.Of these nine lines, seven showed a small increase inacylsugar levels compared with IL5-3, although thelevels were still statistically significantly lower than theM82 wild-type levels (Fig. 4B). These results suggestthat both S. pennellii ASH genes contribute to reducedacylsugar levels in IL5-3 plants.
Taken together, the introgression of SpASH1 andSpASH2 and the increased expression of SpASH2 canexplain the IL5-3 low-acylsugar phenotype. How-ever, the in vitro assay results suggest that theSlASH1 enzyme in wild-type S. lycopersicum alsoshould be able to hydrolyze acylsugars in the tri-chome, yet the wild-type M82 plants produce easilydetected levels of acylsucroses in the presence ofSlASH1. One hypothesis for explaining the acylsugarphenotype of M82 plants is that the acylsugar levelsreally are quite low, compared with LA0716, and thatthis is due in part to the activity of SlASH1 in thetrichomes. Based on this logic, silencing of ASH genescould potentially lead to increased acylsugar levels.However, silencing of ASH expression in IL5-3 didnot result in acylsugar levels that were significantlygreater than the levels in wild-type M82 plantswhen acylsugars were measured in the primary
Figure 4. RNAi suppression of SpASH1 and SpASH2 restores acylsugarproduction in IL5-3. A, When IL5-3 was transformed with an RNAiconstruct targeting sequence with high identity for both SpASH1 andSpASH2, the primary transformants showed a range of increased totalacylsugar levels, with some showing acylsugar levels comparable to theM82 phenotype. B, An RNAi construct targeting specifically SpASH2did not have a strong impact on acylsugars when transformed into IL5-3plants. IS, Internal standard.
Plant Physiol. Vol. 170, 2016 1337
Tomato Acylsugar Acylhydrolases
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

transformants. This suggests that other modes ofacylsugar regulation exist, possibly at the level ofbiosynthesis or secretion, that result in S. pennelliiLA0716 accumulating acylsugars in amounts muchhigher than S. lycopersicum M82.
Other Tomato CXEs with ASH Activity
The ASH1 and ASH2 enzymes specifically hydro-lyze the R3 and R4 positions of acylsugar substratesfrom S. lycopersicum, yet IL5-3 does not accumulateany detectable diacylsucroses or triacylsucroseslacking acyl chains at these positions. This suggeststhat other ASHs are expressed in trichomes, and thishypothesis was explored in several ways. First, aphylogeny was constructed using the complete setof S. lycopersicum class I carboxylesterase enzymescontaining Interpro domain IPR013094 in the mostrecent annotation (ITAG2.4; Supplemental Fig. S6). Asearch was then conducted to explore for evidence ofexpression of the 33 other predicted tomato CXE genesin the S. lycopersicum and S. pennellii trichome RNAseqdata (Schilmiller et al., 2010a; McDowell et al., 2011;Ning et al., 2015). Two other acylsugar acylhydrolasecandidates, Solyc09g075710 (Sopen09g030520) andSolyc04g005230 (Sopen04g001210), are clustered withASH1 and ASH2 and show trichome-enriched expres-sion (Supplemental Fig. S6).
Assays were performed with recombinant enzymesto test their activities with acylsugar substrates from
M82. The Solyc09g075710 enzyme and correspondingS. pennellii LA0716 isoform, renamed SlASH3 andSpASH3, respectively, showed activity cleaving the C5chain at the R4 position of M82 acylsucroses (Fig. 5).Assays with both ASH3 isoforms using fraction IIsubstrates containing acylsucroses with a long acylchain resulted in three products, S3:17 (2,5,10), S2:17(5,12), and S3:19 (2,5,12), from hydrolysis of a C5 acylchain from S4:22 (2,5,5,10), S3:22 (5,5,12), and S4:24(2,5,5,12), respectively (Fig. 5, A–C; Supplemental Fig.S7). Each of these products retained the R39 C5 acylchain as determined by fragmentation in positive-ionmode LC-MS (Supplemental Fig. S4); therefore,the hydrolyzed chain came from the six-memberedpyranose ring. Because the S3:22 (5,5,12) and S4:24(2,5,5,12) substrates have a single C5 acyl chain on thepyranose ring, we conclude that the cleaved C5 acylchain is located at the R4 position. Similarly, when thefraction I short chain-containing acylsucroses of M82were used as substrate, SlASH3 and SpASH3 pro-duced an S3:12 (2,5,5) peak that results from hydro-lytic removal of a C5 chain from the six-memberedpyranose ring. As mentioned earlier, this productcochromatographs with the later-eluting S3:12 pro-duced in the ASH1 assays that is hypothesized to arisefrom hydrolysis of the R4 position (Fig. 5, D–F). Takentogether, these data show that ASH3 hydrolyzes theC5 chain at the R4 position of triacylsucroses in vitro.In contrast, no activity was detected using theSopen04g001210 enzyme with either M82 acylsugarfraction I or II.
Figure 5. Evidence that SlASH3 and SpASH3 hydrolyze the R4 position ester from M82 acylsucroses. Base peak intensity (BPI)LC-MS chromatograms are shown for reactions with the ASH3 enzymes using M82 long chain-containing (fraction II) and shortchain-containing (fraction I) acylsucrose substrates. IS, Internal standard. A, Fraction II substrate. B, SlASH3 + fraction II. C,SpASH3+ fraction II. D, Fraction I substrate. E, SlASH3 + fraction I. F, SpASH3 + fraction I. Structures of acylsucrose substrates andtheir reaction products are shown in G. The scale of chromatogram signals at 100% (representing the most abundant peak) isindicated by the number at the top right corner of each chromatogram.
1338 Plant Physiol. Vol. 170, 2016
Schilmiller et al.
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

To test whether ASH3 could use the ASH1 productsas substrates, we set up reactions using theM82 fractionI substrate with equal amounts of purified SpASH1 andSpASH3 enzyme. In reactions using only a singleASH enzyme, only one acyl chain was hydrolyzed.When both SpASH1 and SpASH3 were incubatedtogether with M82 fraction I acylsugars, a new andearlier-eluting peak was produced, which corre-sponds to S2:7 (2R2,5R39) (Supplemental Fig. S8). This
result suggests a pathway where multiple ASH en-zymes work sequentially to hydrolyze acylsugars intrichomes.
While introgression of S. pennellii ASH alleles onchromosome 5 resulted in reduced acylsugar levels, noaltered acylsugar phenotype was found for IL9-3, theintrogression region containing the SpASH3 allele(Schilmiller et al., 2010a). This presumably is theresult of several factors. First, the S. lycopersicum and
Figure 6. SpASH enzymes show higher activity with an S. lycopersicum acylsucrose compared with an LA0716 acylsucrose. A, Structures of S3:22fromM82 and S3:19 from LA0716 and the respective hydrolysis products from the SpASH reactions. B to D, Product amounts from assays with a fixedconcentration (33 mM) of substrate and increasing amounts of SpASH1 (B), SpASH2 (C), or SpASH3 (D) enzyme. IS, Internal standard.
Plant Physiol. Vol. 170, 2016 1339
Tomato Acylsugar Acylhydrolases
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

S. pennellii ASH3 isoforms showed the same in vitroactivities, which suggests that they have the same invivo function. Second, the two genes appear to beexpressed similarly: no difference in SpASH3 gene ex-pression was found in IL9-3 compared with other in-trogression lines that have the SlASH3 allele (Chitwoodet al., 2013). The low in vitro activity of ASH3 foracylsugar hydrolysis also suggests that ASH3 has adifferent function in vivo in trichomes.
SpASH Enzymes Display Low or No Activity againstLA0716 Acylsugars
RNAseq data indicated that the SpASH genes arehighly expressed in S. pennellii LA0716 trichomes, yetplants of this accession produce large amounts ofacylsucroses and acylglucoses, estimated to be as highas 20% of leaf dry weight in 17-week-old plants (Fig. 1,C and D; Fobes et al., 1985). To determine how LA0716plants can accumulate such high levels of acylsugarswhile expressing genes encoding acylsugar hydrolasesin trichomes, we compared the relative activities ofS. pennellii ASH enzymes using acylsugars purifiedfrom M82 or LA0716 plants.
LA0716 acylsugars consist of both triacylglucosesand triacylsucroses, with the latter having all threeacylsucrose acyl chains esterified to the pyranose ringof the Suc backbone (Shapiro et al., 1994; Schilmilleret al., 2015). This is in contrast to the triacylsucroses andtetraacylsucroses produced by M82 trichomes, each ofwhich has a single acyl chain (usually C5) present onthe furanose ring of the Suc backbone (Schilmiller et al.,2015). In addition, the M82 tetraacylsucroses have C2chains at the pyranose ring R2 position, whereasLA0716 acylsugars have longer C5 chains. To deter-mine whether either structural difference inhibits ASHactivity, an abundant acylsucrose and an acylglucosewere purified from LA0716 for use in SpASH assays.Structures of the purified acylsugars were character-ized by NMR spectroscopy: the acylsucrose S3:19(ai5R2,i10R3,i4R4) and the acylglucose G3:14 (ai5R2,i5R3,i4R4)from LA0716 both have acyl chains in the R2, R3, and R4positions (Supplemental Tables S2 and S3).
ASH activity assays were first performed using pu-rified acylsucroses with the three recombinant SpASHenzymes. SpASH2 showed the most striking differencein activity: while SpASH2 efficiently hydrolyzed a C5chain from M82 S3:22 (n12R3,i5R4,i5R39), only weak ac-tivity was observed using LA0716 S3:19 (ai5R2,i10R3,i4R4)as substrate, this despite the use of greater than 20-foldmore SpASH2 enzyme thanwas used in assays with theM82 S3:22 substrate (Fig. 6C). Activity assays withSpASH2 using a variety of purified acylsucroses fromSolanum habrochaites with known structures (Ghoshet al., 2014) indicated that the presence of an acyl chainlonger than C2 at the R2 position strongly inhibitedSpASH2 activity (Supplemental Fig. S9). SpASH1 alsoshowed greater activity with the S. lycopersicum S3:22(n12R3,i5R4,i5R39) acylsucrose as substrate compared
with S. pennellii S3:19 (ai5R2,i10R3,i4R4), with the dif-ference seen most clearly at lower SpASH1 enzymeconcentrations (Fig. 6B). Kinetic analysis of SpASH1indicated a similar Km for each substrate (Table I;Supplemental Fig. S10; note that SpASH1 can hydro-lyze two different positions and that Km values werecalculated for each position); however, the maximumvelocity was approximately 2.2-fold higher for hy-drolysis at the R3 position of S3:22 compared withS3:19 and 6.6-fold higher for hydrolysis at the R4 po-sition. SpASH3 activity with either substrate was de-tectable only at very high enzyme concentrations, andthe specific activity was much lower than that ofSpASH1 or SpASH2 (Fig. 6D). These results show thatthe relatively high in vitro activity of SpASH1 andSpASH2 enzymes with the S. lycopersicum S3:22 isconsistent with the observed acylsugar reduction inIL5-3 plants.
S. pennellii LA0716 accumulates even higher amountsof acylglucoses than acylsucroses (Shapiro et al., 1994;Ning et al., 2015). This led us to hypothesize that theseacylglucoses are also poor substrates for SpASHenzymes. As was the case for the LA0716 S3:19acylsucrose (Fig. 6), SpASH2 showed very low activitydetectable only at high enzyme concentrations whenusing G3:14 (ai5R2,i5R3,i4R4) as substrate (SupplementalFig. S11). SpASH1 and SpASH3 both showed higheractivity with G3:14 compared with SpASH2; however,the amount of conversion to product was low even atthe highest enzyme concentrations (Supplemental Fig.S11). Taken together, the low activity of SpASH en-zymes with the S. pennellii acylsucrose and acylglucoseis consistent with the high accumulation of acylsugarsin LA0716.
CONCLUSION
The in vitro activity of SpASH1 and SpASH2 in hy-drolyzing acylsucroses, together with the restorationof acylsugar production following the silencing ofSpASH1/2, indicates that these carboxylesterases con-dition the IL5-3 reduced acylsugar phenotype. Whilethe actual in vivo functions of the ASH enzymes onspecific acylsugar substrates in S. lycopersicum andS. pennellii are less clear, several testable hypothesesseem plausible. One possibility is that the ASH en-zymes play a role in acylsugar turnover in the tri-chomes. The commitment of large amounts of Suc or
Table I. SpASH1 kinetic parameters
Vmax values are not given units because actual product amountscould not be quantified in mmol; instead, they are calculated usingpeak response and are shown relative to Vmax for S3:22 R3 hydrolysis.
Substrate Apparent Km Apparent Vmax
mM
S3:22 R3 hydrolysis 17.4 6 2.5 100S3:19 R3 hydrolysis 24.8 6 10.8 45S3:19 R4 hydrolysis 44.2 6 19.7 15
1340 Plant Physiol. Vol. 170, 2016
Schilmiller et al.
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

Glc or short-chain fatty acids for acylsugar productioncould have a negative effect on apical cell function orplant growth under certain environmental conditions,and the ASHs could return acylsugars back into othermetabolic pathways. Alternatively, the ASH enzymescould play a role in the remodeling of acylsugar struc-tures; remodeling of lipids via acyl editing has beenshown to occur in plant and animal systems (Bateset al., 2007; Robichaud et al., 2013). Similarly, ASHscould remove acyl chains, allowing replacement withan acyl chain of different length or branch structure.Another alternative is that the ASH enzymes couldperform the reverse reaction and catalyze the transfer ofacyl chains to form an ester-linked acylsugar productrather than hydrolyze ester linkages.Analysis of the sequences of the ASH enzymes does
not indicate a subcellular localization other than cyto-solic, which is similar to that of the ASAT BAHD en-zymes that produce acylsucroses. Colocalization ofbiosynthetic enzymes with enzymes capable of hydro-lyzing acylsugars would seemingly create the possi-bility of a futile cycle of synthesis and degradation;however, this does not prevent the accumulation ofacylsugars on S. lycopersicum and S. pennellii leaves.More work is needed to understand whether temporalexpression or localization differences for the ASAT andASH enzymes are important for controlling acylsugarproduction. Labeling experiments similar to that de-scribed by Wang and Jones (2014) using IL5-3 or someof the RNAi lines described in this work may help toreveal the dynamics of acylsugar accumulation and theroles that ASH enzymes play.Due to their role in defense against herbivores,
acylsugar breeding programs are under way to createS. lycopersicum varieties with acylsugar levels andcompositions that increase resistance to insect pests(Alba et al., 2009; Leckie et al., 2012). Quantitative traitlocus (QTL) analyses using populations derived fromS. lycopersicum and S. pennellii LA0716 previouslyidentified a negative-effect QTL on chromosome 5 thatmaps to the same region as ASH1 and ASH2 (Leckieet al., 2012). In fact, one of the markers used to definethe QTL is located only approximately 56 kb away fromthe ASH locus. Our results suggest that SpASH1 andSpASH2 are the causative genes in this negative-effectQTL. The dominance of the S. pennellii alleles is con-sistent with the observed reduction in acylsugar levelswhen the chromosome 5 QTL is present in the hetero-zygous state (Leckie et al., 2012).Our understanding of plant specialized metabolism
is ever increasing as more pathways and enzymes arediscovered (Osbourn and Lanzotti, 2009). Many plantspecialized metabolites are modified by esterificationwith a variety of chemical side chains, including acetyl,aliphatic, phenolic, and organic acid groups. Thesemodifications can modulate the chemical and physio-logical properties of specialized metabolites or serve asblocking or activating groups in metabolic path-way intermediates. Studies focused on the expression(tissue specificity, environmental regulation, etc.) and
activities of carboxylesterases will lead to better un-derstanding of how hydrolysis influences the struc-tures, functions, and amounts of these metabolites.Given the important roles in both the synthesis andturnover of metabolites that the small number of char-acterized CXEs play in plant physiology, a systematicanalysis of this gene family will be important forthe rational engineering of biosynthetic pathways ofvaluable products that are not readily chemicallysynthesized.
MATERIALS AND METHODS
Mapping of the IL5-3 Acylsugar Locus
An F2 mapping population was created by self-pollination of a cultivatedtomato (Solanum lycopersicum) M82 3 Solanum pennellii IL11-3 F1 line kindlyprovided by Dani Zamir (Hebrew University Faculty of Agriculture).Genotyping using PCR-based cleaved-amplified polymorphic sequencemarkers (Supplemental Table S4) and phenotyping by negative-ion modeLC-MS analysis of acylsugars extracted by gentle agitation for 2 min wereperformed as described previously (Schilmiller et al., 2015).
Quantitative RT-PCR
To compare the expression in M82 versus IL5-3 trichomes, total RNA wasextracted from stem and petiole trichomes of 3-week-old M82 and IL5-3 plantsusing the RNeasy Plant Mini Kit (Qiagen). First-strand cDNA was synthesizedfrom 0.5 mg of total trichome RNA using SuperScript II reverse transcriptase(Invitrogen), and the cDNA was diluted 10-fold for qPCR. To determine theexpression levels in M82 trichomes versus the underlying stems and petioletissue, cDNA was prepared from 1 mg of total RNA that was used for RNAseqas described (Ning et al., 2015). ASH transcripts were quantified using thestandard curve method. The qPCR mix consisted of SYBR Select master mix(Life Technologies) with primer concentration of 250 nM and supplementedwith 0.25 mL of Taq polymerase (Invitrogen) per 20-mL reaction. Primer se-quences are listed in Supplemental Table S4. Reactions were analyzed using anABI 7500 Fast qPCR machine (Life Technologies). ASH transcript levels werenormalized to EF1a (Solyc06g009970) transcript levels for comparison acrosstissues and genotypes.
Silencing of ASH1/2 Using RNAi
Constructs for RNAi silencing of SpASH1 and SpASH2 in IL5-3 plants werecreated using the pHELLSGATE-12 binary vector (Eamens and Waterhouse,2011). A 511-bp fragment of SpASH1 was amplified from IL5-3 genomic DNAand cloned into pENTR/D-TOPO (Invitrogen) followed by recombination intopHELLSGATE-12 (primers are listed in Supplemental Table S4). Similarly, a351-bp fragment of SpASH2 from the 39 end of the ORF and untranslated regionwas amplified from IL5-3 genomic DNA, cloned into pENTR/D-TOPO, andrecombined into pHELLSGATE-12. RNAi constructs were transformed intoAgrobacterium tumefaciens strain AGL0, and transformation of IL5-3 tomato wasperformed as described previously (McCormick, 1991).
Phylogenetic Tree Construction
Protein sequences containing Interprodomain IPR013094 from theannotatedtomato genome (ITAG2.4) were aligned together with several other plant classI CXEs using the Muscle algorithm in MEGA software (version 6.06; Tamuraet al., 2013). The phylogeny was constructed using the maximum-likelihoodmethod with the Le and Gascuel substitution model and bootstrapped with1,000 replicates.
Preparation of Acylsugar Substrates
Acylsugar mixtures fromM82were collected by extracting leaves and stemsfrom 3-week-old plants using acetonitrile:isopropanol:water (3:3:2, v/v/v)
Plant Physiol. Vol. 170, 2016 1341
Tomato Acylsugar Acylhydrolases
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

containing 0.1% (v/v) formic acid. The extract was diluted 1:4 with water andthen loadedontoa500-mgC18 solid-phase extraction cartridge (Supelco; catalogno. 57012) equilibrated with 10% (v/v) acetonitrile. Short chain-containingacylsucroses were eluted using 5 mL of 45% (v/v) acetonitrile followed bywashing with 5 mL of 90% (v/v) acetonitrile to elute long chain-containingacylsucroses. Fractions were reduced to dryness under vacuum using a Speed-Vac at room temperature, and the resulting residue was dissolved in 50% (v/v)ethanol for use in assays.
The acylsugars S3:22 (5,5,12) from M82 and S3:19 (4,5,10) and G3:14 (4,5,5)from LA0716were purified as described (Ghosh et al., 2014). NMR spectra wererecorded as described (Schilmiller et al., 2015). The concentrations of purifiedS3:22, S3:19, and S3:14were determined using amodified version of themethoddescribed by Ghosh and Jones (2015). Acyl chains were hydrolyzed by sa-ponification, and the sugar backbone concentration was determined usingstable isotope dilution as follows. A 10-mL aliquot of the purified acylsugarstock solution in acetonitrile was transferred to a 250-mL glass vial, and thesolvent was removed under vacuum in a Speed-Vac at room temperature. Theresulting residue was dissolved in 10 mL of methanol followed by the additionof 10 mL of freshly prepared 1 M sodium hydroxide. After 48 h at room tem-perature (25°C), 10 mL of water plus 70 mL of methanol were added, giving afinal composition of 4:1 (v/v) methanol:water, and samples were further in-cubated at room temperature for another 24 h. Samples were diluted either10- or 100-fold with 4:1 (v/v) methanol:water prior to adding either [13C12]Suc or[13C6]Glc internal standard at a final concentration of 0.5 mM. Saponified sugarswere analyzed using a Waters Quattro Premier triple quadrupole LC-MS deviceinterfaced to a Waters Acquity ultra-performance liquid chromatography ap-paratus. Chromatography was performed using a Waters BEH-amide column(2.1 3 100 mm, 1.7-mm particle size) with column oven set to 40°C. Initialconditions were 10% water + 0.1% formic acid (v/v; solvent A)/90% acetoni-trile (solvent B) at a flow rate of 0.3 mL min21 for 1 min, followed by a lineargradient to 70% A:30% B at 5 min, then to 100% A at 6 min, hold at 100% A to8 min, return to 10% A:90% B at 9 min, and then hold at 10% A:90% B until10 min. Compounds were ionized by electrospray ionization in negative-ionmode, and mass spectra were acquired using multiple reaction monitoring(MRM). The capillary voltage, extractor voltage, and radiofrequency lens set-ting were 3 kV, 5 V, and 4.3, respectively. Cone gas and desolvation gas flowrates were 50 and 600 L h21, and the source and desolvation temperatures were120°C and 350°C. The source cone potentials and collision energies, respec-tively, were as follows: for Suc, 35 and 17 V; for Glc, 16 and 10 V. The precursorand product ion masses used for the MRM transitions were 341 . 179 (Suc),353 . 185 ([13C12]Suc), 179 . 89 (Glc), and 185 . 92 ([13C6]Glc). External stan-dard curves were prepared for Glc and Suc ranging from 0.1 to 125 mM
containing 0.5 mM13C-labeled internal standard, and response factors were
calculated based on the MRM peak areas.
ASH Protein Expression and Enzyme Assays
Recombinant protein for enzyme assays was produced in Escherichia coli asfollows. Full-length ORFs for SlASH1, SpASH1, SpASH2, SlASH3, SpASH3, andSopen04g001210 were amplified using primers (Supplemental Table S4) thatadded restriction enzyme sites to the ends for cloning into pET28b (EMDMillipore). The resulting constructs, which were designed to add an N-terminal63 His tag to the protein for affinity purification, were transformed into BL21Rosetta cells (EMD Millipore). Protein expression was induced in culturesgrowing in Luria-Bertani medium by adding 0.05 mM isopropyl b-D-1-thiogalactopyranoside followed by shaking at 37°C and 200 rpm overnight.Soluble protein was purified by nickel-affinity chromatography. Cells werelysed by sonication in extraction buffer (50 mM sodium phosphate, pH 6.3,150 mM sodium chloride, and 10% [v/v] glycerol), and the cleared lysate wasadded to nickel resin (Qiagen) and incubated for 30 min at 4°C followed by two15-min washes with extraction buffer + 20 mM imidazole. Protein was elutedfrom the resin with extraction buffer + 200 mM imidazole. Protein concentrationwas assayed using the bicinchoninic acid assay (Thermo Scientific).
Acylsugar hydrolase assays using S. lycopersicum M82 acylsucrose frac-tions were performed in a 30-mL volume by incubating 1 mg of purifiedprotein in 50 mM ammonium acetate buffer (pH 6). Reactions were allowed toproceed for 30 min at 30°C followed by termination by the addition of 60 mLof acetonitrile:isopropanol:formic acid (1:1:0.001, v/v/v) containing 10 mM
propyl-4-hydroxybenzoate internal standard and centrifugation at 13,000gfor 10 min. Ten microliters of the supernatant was analyzed by LC-MS asdescribed previously (Schilmiller et al., 2015). Assays for determining theactivity of SpASH enzymes with purified S3:22 (5,5,12), S3:19 (4,5,10), and
G3:14 (4,5,5) were performed by varying the amount of purified enzyme with33 mM final concentration of acylsugar substrate in a 30-mL volume and in-cubating at 30°C for 5 min. Assays for determining SpASH1 kinetic constantswere performed using 10 ng of purified SpASH1 enzyme with varying con-centrations of acylsucrose and incubating at 25°C for 3 min. Five microlitersof the reaction mixture was analyzed by LC-MS to determine velocities (inunits of peak area of hydrolysis product divided by peak area of internalstandard). Apparent Km and Vmax values were determined by nonlinear re-gression using the Michaelis-Menten kinetics model in GraphPad Prism5(GraphPad Software).
Sequence data can be found in the GenBank database under the followingaccession numbers: SlASH1 (KT282356), SlASH2 (KT282357), SlASH3(KT282358), SpASH1 (KT282359), SpASH2 (KT282360), SpASH3 (KT282361),and Sopen04g001210 (KT282362).
Supplemental Data
The following supplemental materials are available.
Supplemental Figure S1. SlASH transcript levels in isolated trichomes.
Supplemental Figure S2. Alignment of S. lycopersicum M82 and S. pennelliiLA0716 ASH1 and ASH2 cDNA sequences.
Supplemental Figure S3. Negative-ion mode mass spectra of ASH1 andASH2 assay products.
Supplemental Figure S4. Positive-ion mode LC-MS analysis of ASH reac-tion products using M82 acylsucrose substrates.
Supplemental Figure S5. Ratio of short-to-long chain-containing acylsugarsin IL5-3 RNAi lines.
Supplemental Figure S6. Phylogeny of tomato CXE proteins containingInterpro domain IPR013094 with other characterized plant CXE proteins.
Supplemental Figure S7. Mass spectra of ASH3 reaction products usingM82 long chain-containing acylsucroses (fraction II) as substrate.
Supplemental Figure S8. Combined SpASH1 and SpASH3 reactions withM82 fraction I substrates.
Supplemental Figure S9. SpASH2 activity with various S. habrochaitesacylsucroses.
Supplemental Figure S10. Nonlinear regression analysis of SpASH1 activ-ity to determine kinetic constants.
Supplemental Figure S11. SpASH enzyme activity with the acylglucoseG3:14 (5,5,4).
Supplemental Table S1. List of genes in the IL5-3 mapping interval.
Supplemental Table S2. NMR chemical shifts for G3:14.
Supplemental Table S3. NMR chemical shifts for S3:19.
Supplemental Table S4. Primer sequences.
ACKNOWLEDGMENTS
We thankAmanda Charbonneau for technical assistance, GauravMoghe forassistance with RNAseq data analysis, Dani Zamir for IL5-3 3 M82 F1 seeds,other trichome project members, and the RTSF Mass Spectrometry andMetabolomics Core Facility at Michigan State University.
Received August 27, 2015; accepted January 23, 2016; published January 25,2016.
LITERATURE CITED
Akashi T, Aoki T, Ayabe S (2005) Molecular and biochemical characterizationof 2-hydroxyisoflavanone dehydratase: involvement of carboxylesterase-like proteins in leguminous isoflavone biosynthesis. Plant Physiol 137:882–891
1342 Plant Physiol. Vol. 170, 2016
Schilmiller et al.
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

Alba JM, Montserrat M, Fernández-Muñoz R (2009) Resistance to the two-spotted spider mite (Tetranychus urticae) by acylsucroses of wild tomato(Solanum pimpinellifolium) trichomes studied in a recombinant inbredline population. Exp Appl Acarol 47: 35–47
Bates PD, Ohlrogge JB, Pollard M (2007) Incorporation of newly synthe-sized fatty acids into cytosolic glycerolipids in pea leaves occurs via acylediting. J Biol Chem 282: 31206–31216
Baudouin E, Charpenteau M, Roby D, Marco Y, Ranjeva R, Ranty B(1997) Functional expression of a tobacco gene related to the serinehydrolase family: esterase activity towards short-chain dinitrophenylacylesters. Eur J Biochem 248: 700–706
Bombarely A, Menda N, Tecle IY, Buels RM, Strickler S, Fischer-York T,Pujar A, Leto J, Gosselin J, Mueller LA (2011) The Sol Genomics Net-work (solgenomics.net): growing tomatoes using Perl. Nucleic Acids Res39: D1149–D1155
Chitwood DH, Kumar R, Headland LR, Ranjan A, Covington MF,Ichihashi Y, Fulop D, Jiménez-Gómez JM, Peng J, Maloof JN, et al(2013) A quantitative genetic basis for leaf morphology in a set of pre-cisely defined tomato introgression lines. Plant Cell 25: 2465–2481
Chortyk OT, Severson RF, Cutler HC, Sisson VA (1993) Antibiotic activ-ities of sugar esters isolated from selected Nicotiana species. BiosciBiotechnol Biochem 57: 1355–1356
D’Auria JC (2006) Acyltransferases in plants: a good time to be BAHD.Curr Opin Plant Biol 9: 331–340
Dembitsky VM (2004) Astonishing diversity of natural surfactants. 1.Glycosides of fatty acids and alcohols. Lipids 39: 933–953
Eamens AL, Waterhouse PM (2011). Vectors and methods for hairpin RNAand artificial microRNA-mediated gene silencing in plants. In J Birchler,ed, Plant Chromosome Engineering: Methods and Protocols. HumanaPress, New York, pp 179–197
Fan P, Miller AM, Schilmiller AL, Liu X, Ofner I, Jones AD, Zamir D,Last RL (2016) In vitro reconstruction and analysis of evolutionaryvariation of the tomato acylsucrose metabolic network. Proc Natl AcadSci USA 113: E239–E248
Fobes JF, Mudd JB, Marsden MP (1985) Epicuticular lipid accumulation onthe leaves of Lycopersicon pennellii (Corr.) D’Arcy and Lycopersiconesculentum Mill. Plant Physiol 77: 567–570
Forouhar F, Yang Y, Kumar D, Chen Y, Fridman E, Park SW, Chiang Y,Acton TB, Montelione GT, Pichersky E, et al (2005) Structural andbiochemical studies identify tobacco SABP2 as a methyl salicylate es-terase and implicate it in plant innate immunity. Proc Natl Acad Sci USA102: 1773–1778
Gershater MC, Cummins I, Edwards R (2007) Role of a carboxylesterase inherbicide bioactivation in Arabidopsis thaliana. J Biol Chem 282: 21460–21466
Gershater MC, Edwards R (2007) Regulating biological activity in plantswith carboxylesterases. Plant Sci 173: 579–588
Ghosh B, Jones AD (2015) Dependence of negative-mode electrosprayionization response factors on mobile phase composition and molecularstructure for newly-authenticated neutral acylsucrose metabolites. An-alyst (Lond) 140: 6522–6531
Ghosh B, Westbrook TC, Jones AD (2014) Comparative structural profil-ing of trichome specialized metabolites in tomato (Solanum lycopersicum)and S. habrochaites: acylsugar profiles revealed by UHPLC/MS andNMR. Metabolomics 10: 496–507
Goulet C, Mageroy MH, Lam NB, Floystad A, Tieman DM, Klee HJ (2012)Role of an esterase in flavor volatile variation within the tomato clade.Proc Natl Acad Sci USA 109: 19009–19014
Hill K, Rhode O (1999) Sugar-based surfactants for consumer products andtechnical applications. Fett Lipid 101: 25–33
Hotelier T, Renault L, Cousin X, Negre V, Marchot P, Chatonnet A (2004)ESTHER, the database of the a/b-hydrolase fold superfamily of pro-teins. Nucleic Acids Res 32: D145–D147
Kandzia R, Grimm R, Eckerskorn C, Lindemann P, Luckner M (1998)Purification and characterization of lanatoside 159-O-acetylesterase fromDigitalis lanata Ehrh. Planta 204: 383–389
Kim J, Kang K, Gonzales-Vigil E, Shi F, Jones AD, Barry CS, Last RL(2012) Striking natural diversity in glandular trichome acylsugar com-position is shaped by variation at the Acyltransferase2 locus in the wildtomato Solanum habrochaites. Plant Physiol 160: 1854–1870
Ko MK, Jeon WB, Kim KS, Lee HH, Seo HH, Kim YS, Oh BJ (2005) AColletotrichum gloeosporioides-induced esterase gene of nonclimactericpepper (Capsicum annuum) fruit during ripening plays a role in resis-tance against fungal infection. Plant Mol Biol 58: 529–541
Kroumova AB, Wagner GJ (2003) Different elongation pathways in thebiosynthesis of acyl groups of trichome exudate sugar esters from var-ious solanaceous plants. Planta 216: 1013–1021
Leckie BM, De Jong DM, Mutschler MA (2012) Quantitative trait lociincreasing acylsugars in tomato breeding lines and their impacts onsilverleaf whiteflies. Mol Breed 30: 1621–1634
Marshall SD, Putterill JJ, Plummer KM, Newcomb RD (2003) Thecarboxylesterase gene family from Arabidopsis thaliana. J Mol Evol 57:487–500
McCormick S (1991) Transformation of tomato with Agrobacteriumtumefaciens. In K Lindsey, ed, Plant Tissue Culture Manual. KluwerAcademic Publishers, Dordrecht, The Netherlands, pp 1–9
McDowell ET, Kapteyn J, Schmidt A, Li C, Kang JH, Descour A, Shi F,Larson M, Schilmiller A, An L, et al (2011) Comparative functionalgenomic analysis of Solanum glandular trichome types. Plant Physiol155: 524–539
Milo R, Last RL (2012) Achieving diversity in the face of constraints: les-sons from metabolism. Science 336: 1663–1667
Mitchell A, Chang HY, Daugherty L, Fraser M, Hunter S, Lopez R,McAnulla C, McMenamin C, Nuka G, Pesseat S, et al (2015) TheInterPro protein families database: the classification resource after15 years. Nucleic Acids Res 43: D213–D221
Mugford ST, Milkowski C (2012) Serine carboxypeptidase-like acyltrans-ferases from plants. Methods Enzymol 516: 279–297
Ning J, Moghe G, Leong B, Kim J, Ofner I, Wang Z, Adams C, Jones AD,Zamir D, Last RL (2015) A feedback insensitive isopropylmalate syn-thase affects acylsugar composition in cultivated and wild tomato. PlantPhysiol 169: 1821–1835
Nomura T, Ogita S, Kato Y (2012) A novel lactone-forming carboxylesterase:molecular identification of a tuliposide A-converting enzyme in tulip. PlantPhysiol 159: 565–578
Osbourn A, Lanzotti V, editors (2009) Plant-Derived Natural Products:Synthesis, Function, and Application. Springer-Verlag, New York
Ovenden SPB, Yu J, Bernays J, Wan SS, Christophidis LJ, Sberna G, TaitRM, Wildman HG, Lebeller D, Lowther J, et al (2005) Physaloside A,an acylated sucrose ester from Physalis viscosa. J Nat Prod 68: 282–284
Pérez-Castorena AL, Martínez M, Maldonado E (2010) Labdanes and su-crose esters from Physalis sordida. J Nat Prod 73: 1271–1276
Pichersky E, Lewinsohn E (2011) Convergent evolution in plant specializedmetabolism. Annu Rev Plant Biol 62: 549–566
Pontier D, Godiard L, Marco Y, Roby D (1994) hsr203J, a tobacco genewhose activation is rapid, highly localized and specific for incompatibleplant/pathogen interactions. Plant J 5: 507–521
Robichaud PP, Boulay K, Munganyiki JÉ, Surette ME (2013) Fatty acidremodeling in cellular glycerophospholipids following the activation ofhuman T cells. J Lipid Res 54: 2665–2677
Ruppert M, Woll J, Giritch A, Genady E, Ma X, Stöckigt J (2005) Func-tional expression of an ajmaline pathway-specific esterase fromRauvolfia in a novel plant-virus expression system. Planta 222: 888–898
Schilmiller A, Shi F, Kim J, Charbonneau AL, Holmes D, Jones AD,Last RL (2010a) Mass spectrometry screening reveals widespread di-versity in trichome specialized metabolites of tomato chromosomalsubstitution lines. Plant J 62: 391–403
Schilmiller AL, Charbonneau AL, Last RL (2012) Identification of a BAHDacetyltransferase that produces protective acyl sugars in tomato tri-chomes. Proc Natl Acad Sci USA 109: 16377–16382
Schilmiller AL, Last RL, Pichersky E (2008) Harnessing plant trichomebiochemistry for the production of useful compounds. Plant J 54:702–711
Schilmiller AL, Miner DP, Larson M, McDowell E, Gang DR, WilkersonC, Last RL (2010b) Studies of a biochemical factory: tomato trichomedeep expressed sequence tag sequencing and proteomics. Plant Physiol153: 1212–1223
Schilmiller AL, Moghe GD, Fan P, Ghosh B, Ning J, Jones AD, Last RL(2015) Functionally divergent alleles and duplicated loci encoding anacyltransferase contribute to acylsugar metabolite diversity in Solanumtrichomes. Plant Cell 27: 1002–1017
Shapiro JA, Steffens JC, Mutschler MA (1994) Acylsugars of the wildtomato Lycopersicon pennellii in relation to geographic distribution of thespecies. Biochem Syst Ecol 22: 545–561
Simmons AT, Gurr GM (2005) Trichomes of Lycopersicon species and theirhybrids: effects on pests and natural enemies. Agric For Entomol 7:265–276
Plant Physiol. Vol. 170, 2016 1343
Tomato Acylsugar Acylhydrolases
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.

Souleyre EJF, Marshall SDG, Oakeshott JG, Russell RJ, Plummer KM,Newcomb RD (2011) Biochemical characterisation of MdCXE1, acarboxylesterase from apple that is expressed during fruit ripening.Phytochemistry 72: 564–571
Stuhlfelder C, Mueller MJ, Warzecha H (2004) Cloning and expression ofa tomato cDNA encoding a methyl jasmonate cleaving esterase.Eur J Biochem 271: 2976–2983
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: Molec-ular Evolutionary Genetics Analysis version 6.0. Mol Biol Evol 30: 2725–2729
Tomato Genome Consortium (2012) The tomato genome sequence pro-vides insights into fleshy fruit evolution. Nature 485: 635–641
Ueguchi-Tanaka M, Ashikari M, Nakajima M, Itoh H, Katoh E,Kobayashi M, Chow TY, Hsing YI, Kitano H, Yamaguchi I, et al (2005)GIBBERELLIN INSENSITIVE DWARF1 encodes a soluble receptor forgibberellin. Nature 437: 693–698
Wang Z, Jones AD (2014) Profiling of stable isotope enrichment inspecialized metabolites using liquid chromatography and multiplexednonselective collision-induced dissociation. Anal Chem 86: 10600–10607
Weinhold A, Baldwin IT (2011) Trichome-derived O-acyl sugars are a firstmeal for caterpillars that tags them for predation. Proc Natl Acad SciUSA 108: 7855–7859
Winzer T, Gazda V, He Z, Kaminski F, Kern M, Larson TR, Li Y, Meade F,Teodor R, Vaistij FE, et al (2012) A Papaver somniferum 10-gene clusterfor synthesis of the anticancer alkaloid noscapine. Science 336:1704–1708
Yang Y, Xu R, Ma CJ, Vlot AC, Klessig DF, Pichersky E (2008) Inactivemethyl indole-3-acetic acid ester can be hydrolyzed and activated byseveral esterases belonging to the AtMES esterase family of Arabidopsis.Plant Physiol 147: 1034–1045
1344 Plant Physiol. Vol. 170, 2016
Schilmiller et al.
https://plantphysiol.orgDownloaded on March 27, 2021. - Published by Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.






![dxdh zekld ly onegzl aey mrp cxe - home vered noam Noam Tchumey Tehara.pdf · [2] mrp cxe 128 inxeb z` mewn meya dpbn epi`e ,zna letih e` dcil zrya oebk ,d`nehd l` daxw e` d`nehd](https://img.pdfslide.net/doc/110x75/5e881e3fa51fed1d3e73248c/dxdh-zekld-ly-onegzl-aey-mrp-cxe-home-vered-noam-tchumey-teharapdf-2-mrp.jpg)

![Pharmacodynamic Impact of Carboxylesterase 1 Gene Variants in … · carboxylesterase 1 (CES1) [10–12]. The activity of CES1 has been associated with marked indi-vidual variability](https://img.pdfslide.net/doc/110x75/60210bd5a855643d9d144092/pharmacodynamic-impact-of-carboxylesterase-1-gene-variants-in-carboxylesterase-1.jpg)



















