Embed Size (px)
DESCRIPTION
Adaptive variation in the vulnerability of woody plants to xylem cavitation.
Citation preview

2184
Ecology, 85(8), 2004, pp. 2184–2199q 2004 by the Ecological Society of America
ADAPTIVE VARIATION IN THE VULNERABILITYOF WOODY PLANTS TO XYLEM CAVITATION
HAFIZ MAHERALI,1,4 WILLIAM T. POCKMAN,2 AND ROBERT B. JACKSON3
1Department of Botany, University of Guelph, Guelph, Ontario Canada N1G 2W12Department of Biology, University of New Mexico, Albuquerque, New Mexico 87131 USA
3Department of Biology and Nicholas School of the Environment and Earth Sciences, Duke University,Durham, North Carolina 27708 USA
Abstract. The ability of plants to supply water to their leaves is intimately associatedwith survival. Water supply to leaves depends on maintaining an intact water column inthe xylem from the roots to shoots. Because this hydraulic pathway is under tension, it isvulnerable to breakage through the induction of air emboli (cavitation). Although the phys-iological benefit of resistance to water-stress-induced xylem cavitation for desiccation tol-erance is clear, there is considerable interspecific variation within and across climates. Tounderstand the adaptive significance of this variation and the potential trade-off with watertransport, we compiled a database of 167 species from 50 seed plant families and examinedrelationships among resistance to xylem cavitation, water transport capacity (as determinedby the specific conductivity of xylem [KS]), and climate. Relationships were evaluated usingstandard cross-species correlations (r). Because inferences about the adaptive significanceof these correlations can be biased by the potential similarity of closely related species,we also analyzed our data using phylogenetically independent contrast correlations (PIC)calculated over a range of alternate seed plant phylogenies. Resistance to cavitation, ex-pressed as the xylem tension at which 50% of hydraulic conductivity was lost (C50), rangedfrom 20.18 to 29.9 MPa for angiosperms and from 21.5 to 214.1 MPa for conifers.Conifers were most resistant to cavitation, with mean C50 80% more negative than angio-sperms. In contrast, KS was 270% higher in angiosperms than conifers. Across all species,cavitation resistance increased with decreasing mean annual precipitation. However, sig-nificant phylogenetically independent contrast correlations between C50 and annual precip-itation were found within the evergreen angiosperms and conifers but not in the deciduousangiosperms. Thus, the adaptive significance of increased resistance to cavitation as amechanism of drought tolerance may be of primary importance in evergreen angiospermsand conifers. In contrast, analysis of independent contrasts indicated that KS increased withdecreasing rainfall in deciduous angiosperms, whereas there was no association betweenKS and water availability for evergreen angiosperms and conifers. These results suggestthat the evolution of increased KS may be a critical adaptation to water limitation in de-ciduous angiosperms. Although there was a significant cross-species correlation betweenC50 and KS, this relationship was not supported by the independent contrast correlation,suggesting that the evolutionary basis for a trade-off between cavitation resistance andwater transport capacity is weak.
Key words: adaptation; climate; correlated evolution; hydraulic conductivity; independent con-trasts; potential evapotranspiration; precipitation; temperature; vegetation; water-stress-induced xy-lem cavitation.
INTRODUCTION
A major cost of acquiring CO2 for photosynthesis isthe evaporation of water from plant leaves. Water sup-ply to leaves depends on maintaining an intact watercolumn in the xylem from the roots to shoots. Becausethis hydraulic pathway is under increasing tension dur-ing transpiration, it is vulnerable to cavitation (Zim-mermann 1983, Carlquist 1988, Tyree and Sperry1989). Cavitation induced by water stress occurs
Manuscript received 29 August 2002; revised 23 October2003; accepted 16 November 2003; final version received 23 De-cember 2003. Corresponding Editor: J. H. Richards.
4 E-mail: [email protected]
through air seeding, when air bubbles are aspirated intowater-filled conduits under tension (Zimmermann1983, Sperry and Tyree 1988, Tyree and Sperry 1989,Jarbeau et al. 1995). The xylem tension required forair seeding to occur is intimately linked to the anatomyof xylem conduits, specifically the structure of the pitmembranes that permit flow between adjacent vesselsor tracheids (Jarbeau et al. 1995). Each time cavitationoccurs, the resulting vapor-filled conduit no longer car-ries water, causing a decrease in xylem hydraulic con-ductivity (Tyree and Ewers 1991, Sperry and Pockman1993). Without responses to limit transpiration, in-cluding stomatal closure (Sperry and Pockman 1993)or leaf shedding (Pataki et al. 1998), decreased hy-

August 2004 2185EVOLUTION OF CAVITATION RESISTANCE
PLATE 1. Catastrophic hydraulic failure and branch die-back in Larrea tridentata following xylem cavitation. Photocredit: W. T. Pockman.
draulic conductivity can drive increased xylem tensionand further cavitation (Tyree and Sperry 1989, Tyreeand Ewers 1991, Meinzer et al. 2001). Ultimately, ifthis positive feedback continues, plants may undergocatastrophic hydraulic failure (see Plate 1). Even with-out hydraulic failure, an immediate consequence of xy-lem cavitation is reduced photosynthesis and growth(Sperry and Pockman 1993, Meinzer et al. 2001). Thedirect link between sustained water transport and plantsurvival and growth suggests that there may be strongselection for the evolution of high resistance to xylemcavitation.
Despite the potential for selection to favor high re-sistance to cavitation, there is considerable interspecificvariation in the resistance of woody plants to water-stress-induced xylem cavitation. Comparative eco-physiological studies indicate that some of this varia-tion is associated with moisture availability (Brodribband Hill 1999, Kolb and Sperry 1999, Pockman andSperry 2000). Variation in cavitation resistance couldalso be driven by evolutionary correlations with otherphysiological traits (Ackerly and Donoghue 1998). Forexample, because of the structural changes in pit mem-branes associated with increased cavitation resistance,there may be a trade-off between resistance to cavi-tation and xylem conductivity (Sperry 2003). If sucha trade-off has an evolutionary basis, low cavitationresistance could be maintained in environments wherehigh conductivity is adaptive. There is evidence forthis trade-off in some taxa (Tyree et al. 1994, Pinoland Sala 2000), but not in others (Sperry et al. 1994).The resistance of xylem to cavitation may also be in-fluenced by shared evolutionary history. If phyloge-netic history is an important adaptive constraint, in-terspecific variation in resistance to cavitation wouldbe conserved, occurring only in groups of closely re-lated taxa. In contrast, evidence of adaptive evolutionwould be provided by trait convergence, with severalindependent origins of high cavitation resistance ob-served (Harvey and Pagel 1991, Ackerly 1999).
A limitation of using cross-species comparisons todetermine the ecological significance of physiologicaltraits is that these correlations may be biased by thepotential similarity of closely related species (Felsen-stein 1985, Harvey and Pagel 1991). This shared evo-lutionary history prevents individual species from be-ing statistically independent data points (Harvey andPagel 1991). As a result, some observed interspecificcorrelations among traits or between traits and ecolog-ical variables may not reflect true functional or adaptiverelationships (Harvey and Pagel 1991, Ackerly andDonoghue 1998). For example, the strong correlationbetween leaf size and leaf life span observed acrossspecies in six biomes was not found when phylogeneticinformation was included (Ackerly and Reich 1999).In this particular case, the interspecific correlation wasdriven largely by the ancestral divergence between co-nifers and angiosperms. It is also equally possible that
the lack of statistical independence and the effects ofunknown historical events can obscure adaptive rela-tionships (Ackerly and Donoghue 1998, Ackerly 1999).Thus, the inclusion of phylogenetic information canreveal significant adaptive associations between traitsthat go undetected by traditional cross-species corre-lation analysis.
Several statistical techniques that incorporate phy-logenetic information have been developed to evaluatethe evolutionary origin of extant trait variation (e.g.,Pagel and Harvey 1991). One of the most powerful isindependent contrasts (Felsenstein 1985), which arecalculated for each speciation event in the phylogeny(e.g., as the difference in trait values between sistertaxa along each branch; Felsenstein 1985, Garland etal. 1992). Trait relationships are then evaluated usingcorrelations among these contrasts. This method iden-tifies adaptive relationships because the strength andsign of the contrast correlation indicates whether evo-lutionary shifts in a trait are associated with changesin another trait or an ecological variable (Pagel 1993,Ackerly and Reich 1999).
In this study, we analyzed the interspecific relation-ships between two major components of plant hydraulicarchitecture (the resistance to water-stress-induced xy-lem cavitation and hydraulic conductivity) and climateusing data from the literature and from global data-bases. Our goals were to determine the adaptive sig-nificance of resistance to xylem cavitation and hy-draulic conductivity in contrasting climates and to eval-uate the strength of these relationships for differentplant functional groups. By using information on phy-logenetic relationships among seed plants, we calcu-lated phylogenetically independent contrasts to test forcorrelated evolutionary change between traits and theenvironment (Felsenstein 1985, Harvey and Pagel1991, Ackerly 2000). We examined character evolutionby mapping traits onto our phylogeny, and using anindex of similarity, we calculated the degree to whichhydraulic traits were conserved within taxa or exhibited
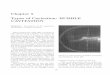
2186 HAFIZ MAHERALI ET AL. Ecology, Vol. 85, No. 8
FIG. 1. The distribution of vulnerability to water-stress-induced cavitation (as determined by xylem tension at which 50%cavitation occurred [C50]) for the species used in this database, ranked by magnitude within five vegetation types. The medianC50 for each vegetation type, along with the sample size for that group, is shown in the inset.
convergent evolution across taxa. We also tested theproposed trade-off between cavitation resistance andwater transport capacity (Tyree and Ewers 1991, Tyreeet al. 1994, Pinol and Sala 2000) by examining cor-related evolution between xylem cavitation resistanceand specific conductivity of xylem (KS).
METHODS
Database of xylem resistance to water-stress-induced cavitation
To examine the relationships between xylem resis-tance to water-stress-induced cavitation, climate, andwater transport, we assembled a database of 60 studiespublished between 1986 and 2000 and our unpublisheddata from field sites in North Carolina and Texas, USA.These studies comprised a total of 230 observationsfor 167 species of woody plants in 50 seed plant fam-ilies (Appendix A). Only studies that reported vulner-ability curves (the response of hydraulic conductivityto changes in xylem tension) of stems or data derivedfrom these curves were used in the analysis. We in-cluded vulnerability curves obtained using methods ofair dehydration (Sperry et al. 1988), air pressurization(Cochard et al. 1992, Sperry and Saliendra 1994), andcentrifugation (Pockman et al. 1995, Alder et al. 1997).We recorded the xylem tension at which 50% cavitationoccurred (C50), a parameter commonly used to char-acterize and interpret interspecific variation in cavita-tion resistance (Tyree and Ewers 1991). To examinethe relationship between water transport efficiency andC50, we also recorded the specific conductivity per unit
xylem area (KS; in kilograms per meter per megapascalper second) only when it was reported in the same studyas C50. Where there were multiple observations for asingle species, mean values were used in the analysis.To be able to examine broad patterns of adaptive in-terspecific variation, we included measurements foradults and saplings, from branches and main stems andfrom field and greenhouse studies. However, we wereunable to assess the influence of intraspecific variationor phenotypic plasticity (Matzner et al. 2001) on ouranalysis; we note that this limitation may compromisethe assumption that trait values were representative forsome species.
Geographic coordinates were recorded and used toobtain climate variables. For greenhouse studies, werecorded the geographic coordinates of the site fromwhich seeds were obtained. When unavailable, thesecoordinates were estimated based on geographic datain the publications. In those cases where geographiccoordinates were unobtainable, our analysis was lim-ited to climatic variables provided by the authors.
Climate data
To examine the adaptive relationships between planthydraulic variables and climate, we gathered data onmean annual precipitation (MAP), temperature (MAT),and potential evapotranspiration (PET) from global da-tabases. We also examined the relationship betweenhydraulic variables and shorter term indicators of cli-mate, such as growing season precipitation. Growingseason precipitation was calculated as the sum of late
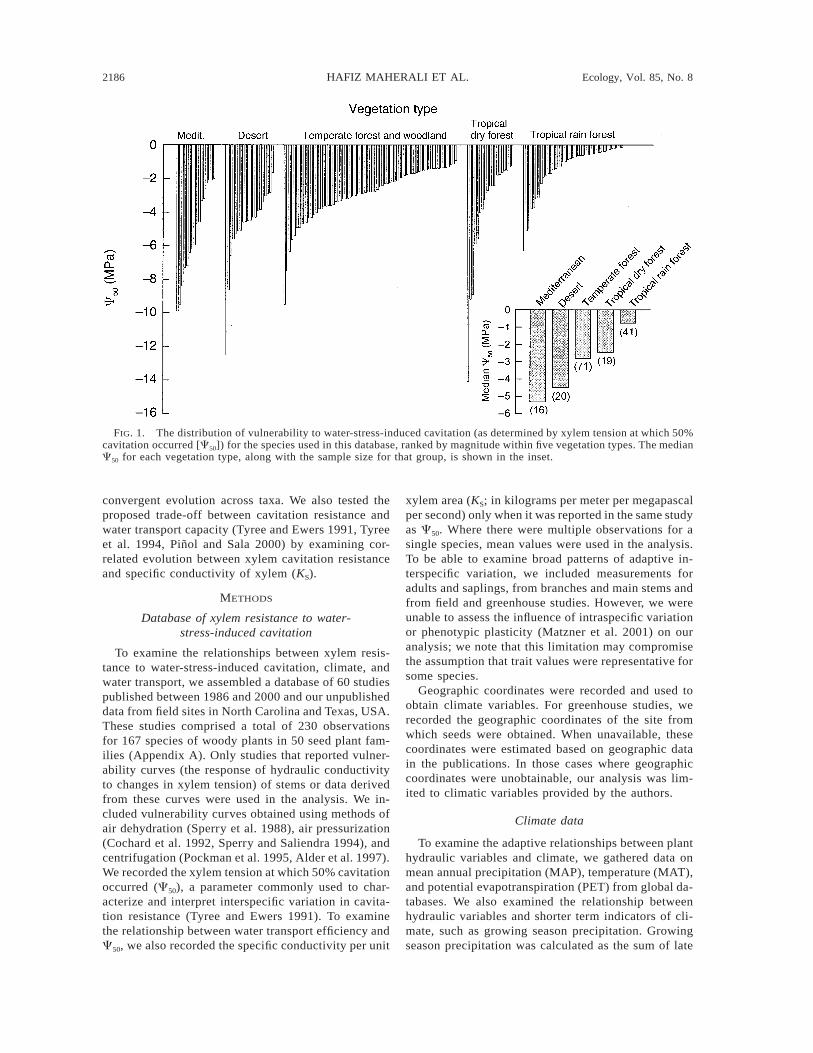
August 2004 2187EVOLUTION OF CAVITATION RESISTANCE
TABLE 1. Summary of data and convergent evolution statistics for vulnerability to xylem cavitation (C50) and specificconductivity (KS).
Comparison
Species values of C50 (MPa)
n Mean 6 1 SE Range QVI‡
Species values of KS (kg·m21·MPa21·s21)†
n Mean 6 1 SE Range QVI‡
All taxaAngiosperms
DeciduousEvergreen
Conifers
167128
626639
23.15 6 0.1922.65 6 0.1822.64 6 0.1822.65 6 0.3124.78 6 0.44
20.18, 214.120.18, 29.920.92, 28.620.18, 29.921.5, 214.1
0.5270.5630.6510.5230.595
8763422124
1.36 6 0.151.70 6 0.191.50 6 0.182.11 6 0.450.46 6 0.05
0.12, 6.70.12, 6.70.16, 4.50.12, 6.70.12, 1.1
0.6730.7540.7390.6740.814
† Specific conductivity is measured per unit xylem area.‡ Quantitative convergence index (scale 0–1, with high values indicating convergent evolution and low values indicating
conserved phylogeny).
TABLE 2. Vulnerability to xylem cavitation (C50, MPa) and specific conductivity (KS; kg·m21·MPa21·s21) for the variousfunctional groups included in this study (means 6 1 SE).
Functional group
Cavitation
C50 MAP (mm) n
Conductivity
KS MAP (mm) n
AngiospermsWinter deciduous shrubsWinter deciduous treesDrought deciduous shrubsDrought deciduous treesEvergreen shrubsEvergreen trees
22.6622.34 6 0.1724.47 6 0.6422.13 6 0.4825.09 6 0.5421.51 6 0.23
140768 6 62196 6 23
1383 6 387761 6 175
2610 6 152
246
95
2144
1.571.58 6 0.221.55 6 0.680.96 6 0.221.75 6 0.742.43 6 0.62
140777 6 85166 6 26
1383 6 387434 6 118
1539 6 247
230
558
12
GymnospermsConifer shrubsConifer trees
28.95 6 1.8124.17 6 0.33
511 6 174984 6 136
534
0.240.48 6 0.05
465817 6 103
222
Notes: Mean (61 SE) annual precipitation (MAP) and sample size (n) for each group and variable combination are includedfor comparison. Standard errors were not calculated for those groups with n , 3.
spring and summer precipitation for each site (May–August for the Northern Hemisphere; November–Feb-ruary for the Southern Hemisphere). Relationships be-tween hydraulic parameters and indices of rainfall werequantitatively similar for both MAP and growing sea-son precipitation across all taxa and within specificfunctional groups (data not shown). However, annualdata were most often included in studies that omittedgeographical coordinates (see Database of xylem re-sistance. . . ). Therefore, we chose annual mean dataover other representations of climate variation becauseit allowed us to keep all species in the analysis. Pre-cipitation and temperature data for each study site wereobtained from the 0.58 gridded global climate databaseof the International Institute of Applied Systems Anal-ysis (IIASA; Laxenburg, Austria; Leemans and Cramer1991). This global database has mean monthly valuesof precipitation and temperature based on .6000weather stations (Leemans and Cramer 1991). Valuesfor PET were obtained from the 0.58 gridded globaldata set of Choudhury (1997) and Choudhury andDiGirolamo (1998) containing mean monthly values ofPET calculated from the Penman-Montieth method(Choudhury 1997). We also examined the relationshipof hydraulic parameters to a derived moisture index,defined as the ratio of MAP to PET (MAP/PET; Schenkand Jackson 2002).
Construction of phylogenetic trees
Because independent contrasts are calculated basedon the difference between pairs of sister taxa at eachnode of a phylogeny, we needed to construct a fullybifurcated tree. To accommodate uncertainties in phy-logenetic hypotheses, we constructed alternate fully bi-furcated phylogenetic trees, using methods modifiedfrom Ackerly and Reich (1999). We started with thestrict consensus phylogenies from published analysesof angiosperms (18S rDNA, rbcL, and atpB gene se-quence data; Soltis et al. 1999) and gymnosperms (28SrRNA sequence data; Stefanovic et al. 1998). Thesetrees were pruned to show relationships among the fam-ilies present in our data set, resulting in a fully bifur-cated family-level phylogeny. Species within each fam-ily were then added, with regions where uncertaintiesarose (i.e., those having more than two genera or spe-cies at a given node) shown as a polytomy. Eleven ofthese 17 species-level polytomies were resolved by ob-taining phylogenetic information on each genus fromsearches of the primary literature (Appendix B). Togenerate a fully bifurcated phylogeny for the calcula-tion of independent contrasts, the six remaining po-lytomies were resolved using the protocol of Donoghueand Ackerly (1996) and Ackerly and Reich (1999). Inbrief, we generated 553 alternate resolved trees (the
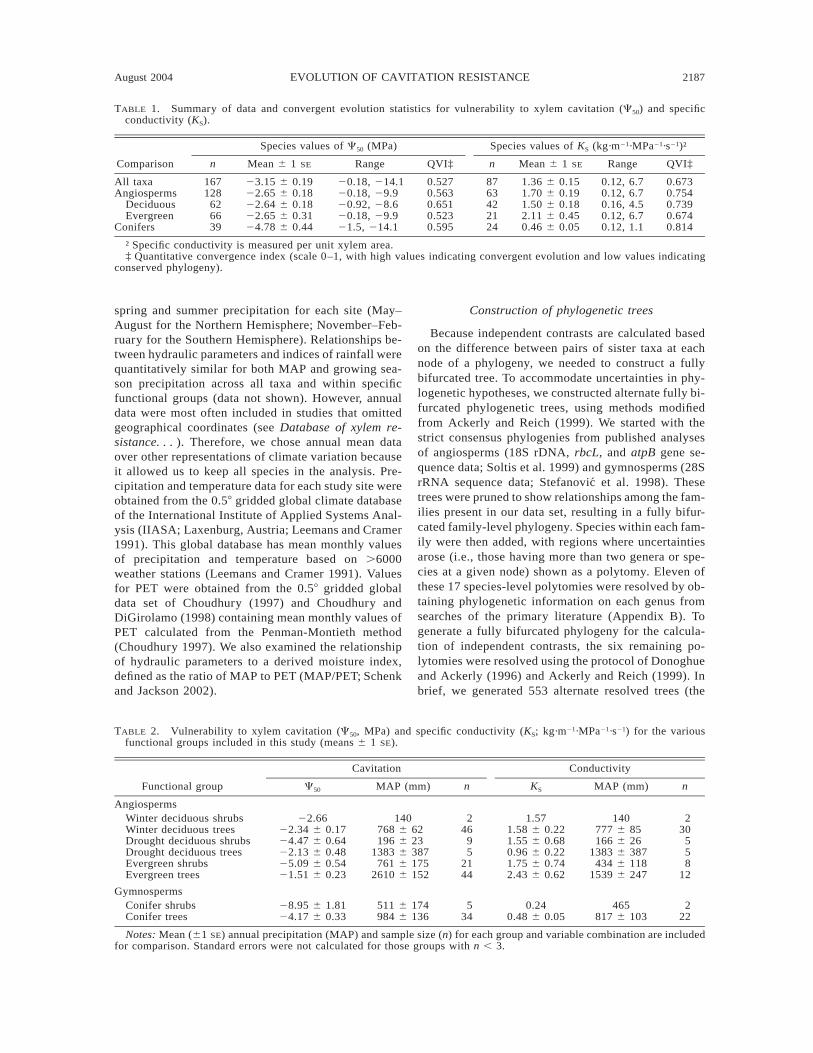
2188 HAFIZ MAHERALI ET AL. Ecology, Vol. 85, No. 8

August 2004 2189EVOLUTION OF CAVITATION RESISTANCE
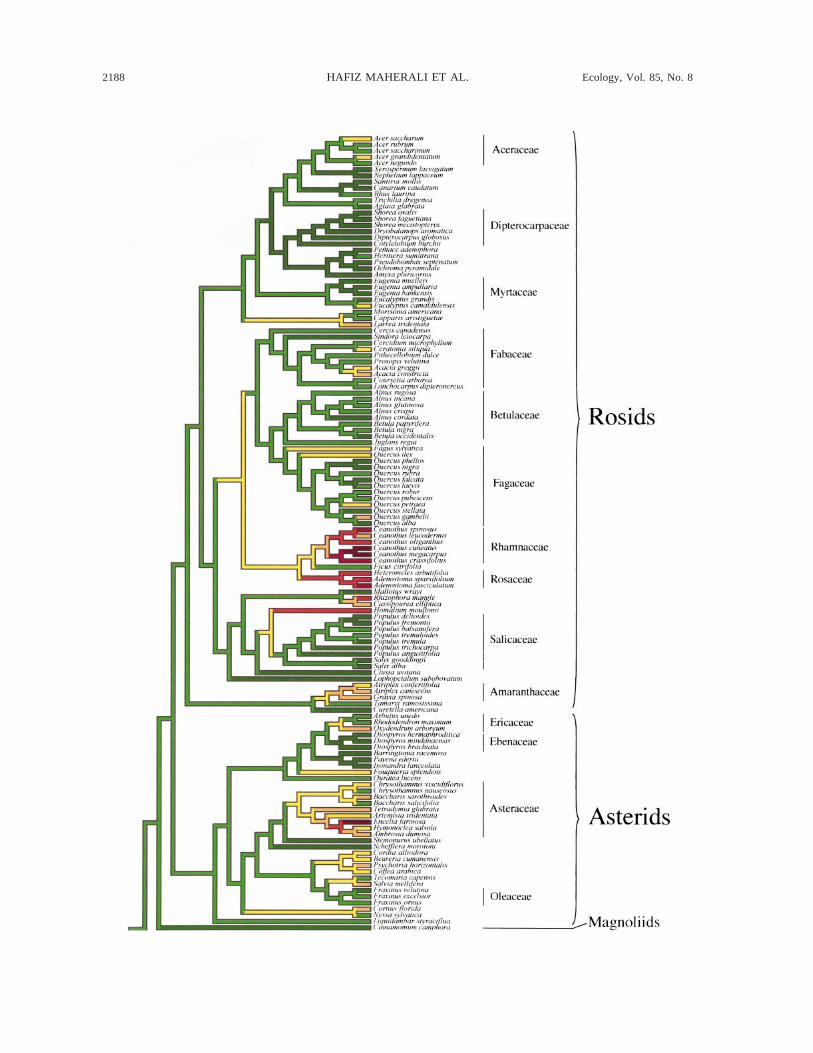
FIG. 2. One of 553 alternate phylogenetic trees showing the evolution of resistance to water-stress-induced xylem cavitationin woody plants (see Methods for details). Overlaying of the continuous characters on the tree was done in MacClade 4.0(Maddison and Maddison 2000). Ancestral state values were determined using squared-change parsimony. However, becausetaxa were nonrandomly sampled across all seed plants, ancestral state values may be biased (Cunningham et al. 1998).Convergence statistics calculated for all 553 trees are shown in Table 2. To facilitate presentation, continuous C50 data weregrouped into 10 discrete color-coded categories of equal size.
result of 1000 randomly resolved trees minus dupli-cates) using the ‘‘randomly resolve current tree’’ optionin MacClade 4.0 (Maddison and Maddison 2000). Thisprocedure resolves polytomies while preserving the re-mainder of the topology.
Comparative statistical analyses
To examine the extent to which interspecific varia-tion in hydraulic traits is the result of convergent evo-lution or constrained by phylogenetic history, we cal-culated the quantitative convergence index (QVI; Ack-erly and Donoghue 1998, Ackerly 1999). QVI valuesrange from 0 to 1, with high values indicating thatdistantly related species are phenotypically similar(convergent evolution) and low values indicating close-ly related species are phenotypically similar (phylo-genetically conserved).
The strength of the cross-species relationships be-tween hydraulic traits and between hydraulic traits andclimate variables without including phylogenetic in-formation were determined using Pearson correlationcoefficients (r; SPSS 10.0, SPSS, Evanston, Illinois,USA). Statistical differences in hydraulic traits be-tween vegetation types and functional groups were test-
ed with analysis of variance (ANOVA). To meet theassumptions of parametric tests, all data were log trans-formed prior to analysis.
To examine the strength of correlated evolutionamong hydraulic traits and between hydraulic traits andclimate parameters, we performed a correlation anal-ysis on calculated phylogenetically independent con-trasts (PICs; Felsenstein 1985, Harvey and Pagel 1991,Garland et al. 1992, Ackerly and Reich 1999) for eachof the 553 fully resolved phylogenies. Independentcontrasts are calculated as the difference in trait valuesbetween sister taxa and standardized by dividing themby the standard deviation of the expected amount ofchange along each branch (Garland et al. 1992). How-ever, because trees from various sources were com-bined to produce our phylogeny, it was impossible toincorporate into our analysis information on branchlengths, which indicate the number of evolutionarychanges along each ancestor–descendant pathway(Harvey and Pagel 1991). In the absence of such in-formation, we assumed that branch lengths are equal,a conservative assumption that minimizes type I errorrate (Purvis et al. 1994, Ackerly 2000).

2190 HAFIZ MAHERALI ET AL. Ecology, Vol. 85, No. 8
Because the sign of an independent contrast is ar-bitrary (depending on which direction is used to cal-culate the difference between sister taxa), correlationanalysis was done through the origin (Garland et al.1992). All comparative analyses were done using CAC-TUS 1.13 (comparative analysis of continuous traitsusing statistics; Schwilk and Ackerly 2001) and thestatistical significance of independent contrast corre-lations was determined using guidelines and statisticaltables for trees with equal branch lengths (Ackerly2000). To facilitate log transformation and analysis, C50
data were converted from negative to positive values.
RESULTS
Variation among species and functional groups
Interspecific variation in C50 spanned nearly two or-ders of magnitude (Fig. 1). The most vulnerable spe-cies, in wet tropical forests, reached 50% cavitation at20.18 MPa, compared with 214.1 MPa for the mostresistant species in dry tropical forests. Cavitation re-sistance also varied substantially within vegetationtypes, ranging from 35-fold in tropical forests to five-fold in the mediterranean zone. This variation not-withstanding, median C50 of species occurring in themost arid vegetation types (e.g., deserts and mediter-ranean zones) were 6–7 times more negative than thoseoccurring in the wettest types (e.g., tropical rain forest,Fig. 1 inset).
On average, conifers were more resistant to water-stress-induced cavitation (C50) than angiosperms; meanC50 was 80% more negative in conifers than angio-sperms (P , 0.0001, Table 1). We note however, thatthe range of C50 values between angiosperms and co-nifers largely overlaps (Table 1). In contrast, mean KS
was 270% higher in angiosperms than conifers (P ,0.0001, Table 1), although minimum values were sim-ilar in both groups (Table 1). Evergreen and deciduousangiosperms had similar mean C50 and KS (P . 0.75,Table 1). Shrubs were more resistant to cavitation thantrees across winter deciduous, drought deciduous, ev-ergreen, and conifer species and also occurred in drierclimates than trees (Table 2). Evergreen angiospermtrees (which were sampled primarily from wet rain for-ests) were the most vulnerable to cavitation, followedby drought deciduous trees, winter deciduous trees andshrubs, conifer trees, drought deciduous shrubs, ever-green shrubs, and conifer shrubs (Table 2). There wereno statistical differences in KS between shrubs and treesfor any group (Table 2).
Quantitative convergence and trait evolution
The impact of variation among the alternate phylog-enies (e.g., from different random solutions to poly-tomies) used in our analysis was quite small. For ex-ample, QVIs and phylogenetically independent contrastcorrelations calculated over 553 alternate phylogenieswere similar (range differed by ,5%). The mean QVI
was always lower for C50 than for KS across all species(Table 1) suggesting that C50 is more conserved thanKS in woody plants. Resistance to cavitation was slight-ly more conserved in angiosperms than conifers (0.563vs. 0.595), although the mean QVI for all taxa was.0.5, indicating greater trait convergence than con-servation (Table 1). Within angiosperms, C50 was moreconserved in evergreen than deciduous species (0.523vs. 0.651). In contrast, KS was highly convergent indeciduous (0.739) and evergreen (0.674) angiospermsand conifer (0.814) species (Table 1).
The extent to which interspecific variation in cavi-tation resistance is the result of convergent evolutioncompared with trait conservation can be illustrated bymapping traits onto a phylogeny (Maddison and Mad-dison 2000). Because the influence of variation in thephylogenies on quantitative parameters was small, wemapped C50 onto one of the 553 phylogenies used inthe analysis (Fig. 2). There was qualitative support forconvergent evolution in cavitation resistance. Withinthe angiosperms, high resistance to cavitation (C50 ,23.15 MPa, the grand mean for all species, Table 1)appeared at least 20 times (Fig. 2). For example, dis-tantly related genera such as Larrea (Zygophyllaceae),Acacia (Fabaceae), Ceanothus (Rhamnaceae), Aden-ostoma (Rosaceae), Atriplex (Amaranthaceae), and En-celia (Asteraceae) were all highly resistant to cavitation(Fig. 2). Within conifers, resistance to cavitation oc-curred in distantly related groups such as the Pinaceae(Pinus edulis and Pseudotsuga menziesii) and Cupres-saceae (Juniperus, Callitris, Actinostrobus, and Wid-dringtonia). In many cases, support for convergent evo-lution on comparatively shorter time scales was alsoprovided by the occurrence of both high and low re-sistance to cavitation within the same genus (e.g., Atri-plex, Baccharis, Pinus, Podocarpus).
Cross-species and independent contrast correlations
Our inclusion of phylogenetic information in thisstudy helped identify the adaptive relationship betweenpairs of traits and between a trait and the environment.We present both cross-species correlations betweenvariables analyzed with Pearson correlation (r) coef-ficients and phylogenetically independent contrasts(PIC). To facilitate log transformation and subsequentcorrelation analysis, C50 data were converted from neg-ative to positive values. We note, however, that forconsistency the data are presented and interpreted withC50 as the usual negative values, such that greater re-sistance to cavitation is indicated by more negative C50
values.Based on interspecific correlations (r), resistance to
water-stress-induced xylem cavitation increased (i.e.,a more negative C50) with decreasing MAP for all taxaanalyzed together (Fig. 3A). These relationships weresupported by correlations using phylogenetically in-dependent contrasts (PICs, Table 3). Similar associa-tions were also found for C50 and an indicator of mois-
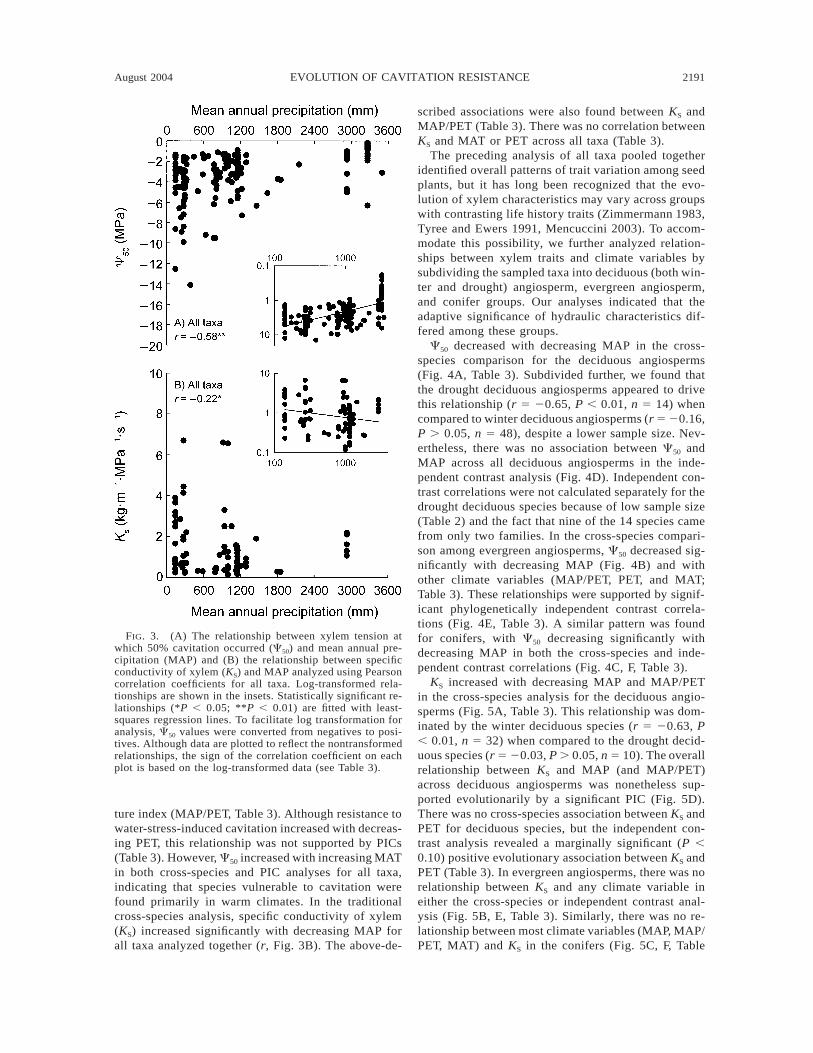
August 2004 2191EVOLUTION OF CAVITATION RESISTANCE
FIG. 3. (A) The relationship between xylem tension atwhich 50% cavitation occurred (C50) and mean annual pre-cipitation (MAP) and (B) the relationship between specificconductivity of xylem (KS) and MAP analyzed using Pearsoncorrelation coefficients for all taxa. Log-transformed rela-tionships are shown in the insets. Statistically significant re-lationships (*P , 0.05; **P , 0.01) are fitted with least-squares regression lines. To facilitate log transformation foranalysis, C50 values were converted from negatives to posi-tives. Although data are plotted to reflect the nontransformedrelationships, the sign of the correlation coefficient on eachplot is based on the log-transformed data (see Table 3).
ture index (MAP/PET, Table 3). Although resistance towater-stress-induced cavitation increased with decreas-ing PET, this relationship was not supported by PICs(Table 3). However, C50 increased with increasing MATin both cross-species and PIC analyses for all taxa,indicating that species vulnerable to cavitation werefound primarily in warm climates. In the traditionalcross-species analysis, specific conductivity of xylem(KS) increased significantly with decreasing MAP forall taxa analyzed together (r, Fig. 3B). The above-de-
scribed associations were also found between KS andMAP/PET (Table 3). There was no correlation betweenKS and MAT or PET across all taxa (Table 3).
The preceding analysis of all taxa pooled togetheridentified overall patterns of trait variation among seedplants, but it has long been recognized that the evo-lution of xylem characteristics may vary across groupswith contrasting life history traits (Zimmermann 1983,Tyree and Ewers 1991, Mencuccini 2003). To accom-modate this possibility, we further analyzed relation-ships between xylem traits and climate variables bysubdividing the sampled taxa into deciduous (both win-ter and drought) angiosperm, evergreen angiosperm,and conifer groups. Our analyses indicated that theadaptive significance of hydraulic characteristics dif-fered among these groups.
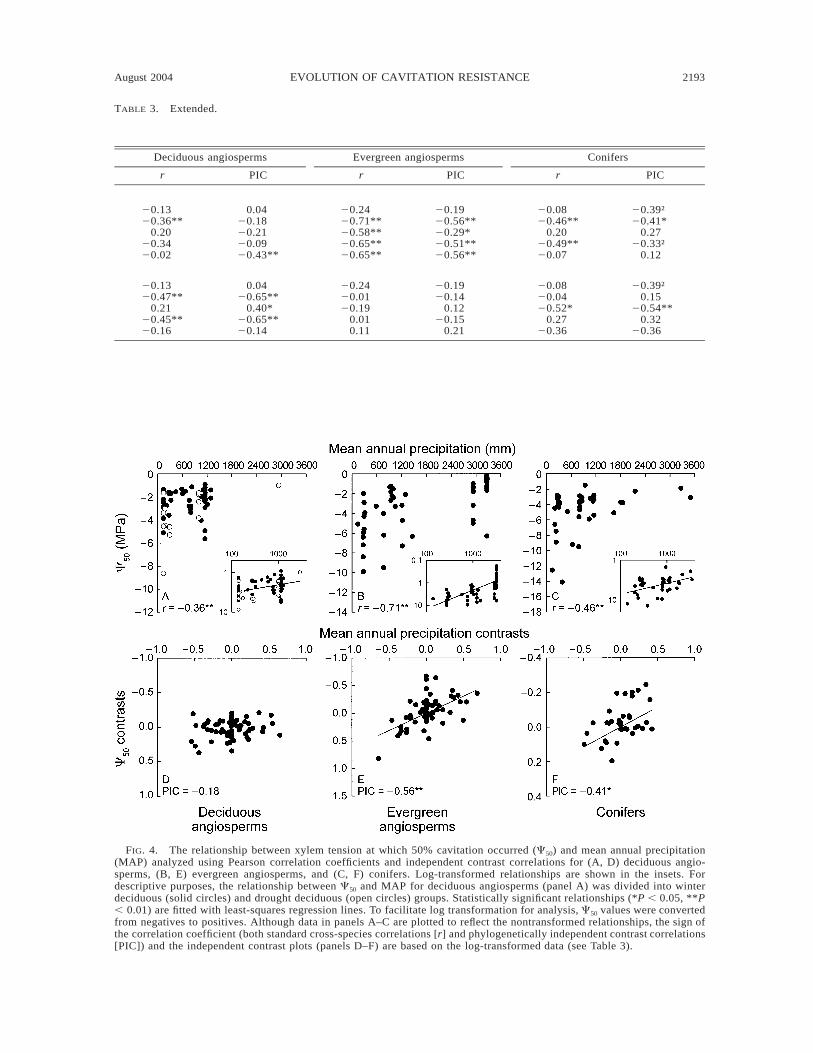
C50 decreased with decreasing MAP in the cross-species comparison for the deciduous angiosperms(Fig. 4A, Table 3). Subdivided further, we found thatthe drought deciduous angiosperms appeared to drivethis relationship (r 5 20.65, P , 0.01, n 5 14) whencompared to winter deciduous angiosperms (r 5 20.16,P . 0.05, n 5 48), despite a lower sample size. Nev-ertheless, there was no association between C50 andMAP across all deciduous angiosperms in the inde-pendent contrast analysis (Fig. 4D). Independent con-trast correlations were not calculated separately for thedrought deciduous species because of low sample size(Table 2) and the fact that nine of the 14 species camefrom only two families. In the cross-species compari-son among evergreen angiosperms, C50 decreased sig-nificantly with decreasing MAP (Fig. 4B) and withother climate variables (MAP/PET, PET, and MAT;Table 3). These relationships were supported by signif-icant phylogenetically independent contrast correla-tions (Fig. 4E, Table 3). A similar pattern was foundfor conifers, with C50 decreasing significantly withdecreasing MAP in both the cross-species and inde-pendent contrast correlations (Fig. 4C, F, Table 3).
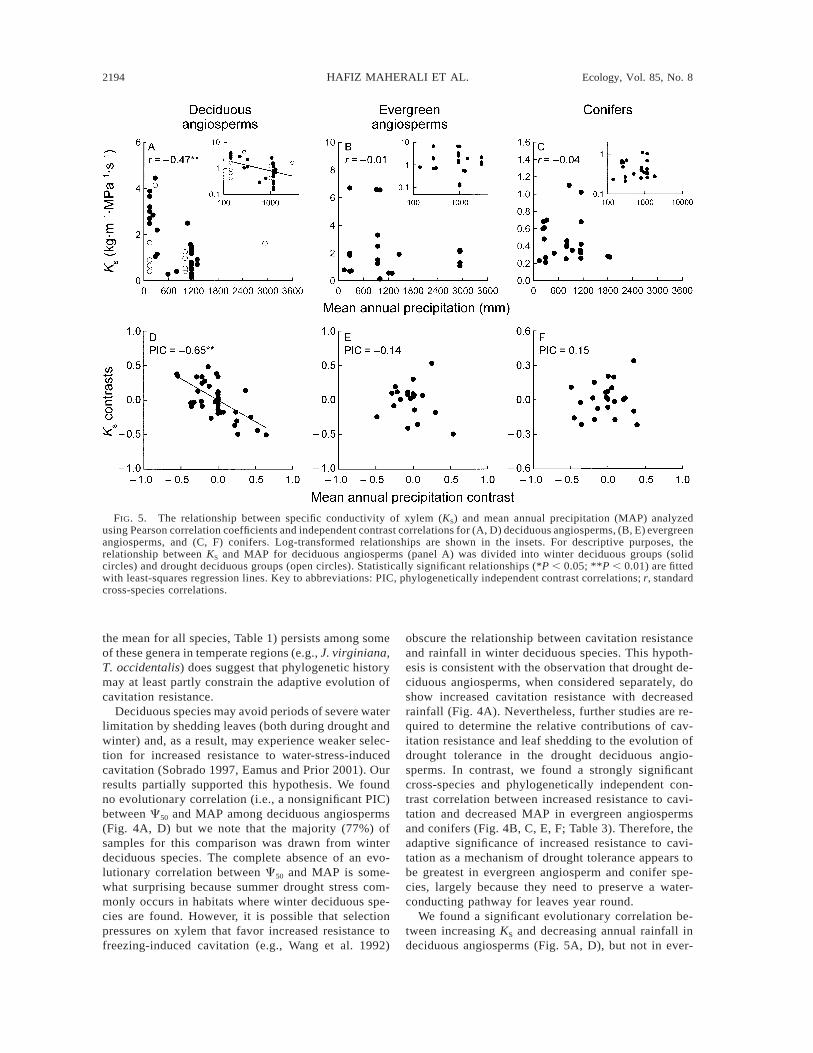
KS increased with decreasing MAP and MAP/PETin the cross-species analysis for the deciduous angio-sperms (Fig. 5A, Table 3). This relationship was dom-inated by the winter deciduous species (r 5 20.63, P, 0.01, n 5 32) when compared to the drought decid-uous species (r 5 20.03, P . 0.05, n 5 10). The overallrelationship between KS and MAP (and MAP/PET)across deciduous angiosperms was nonetheless sup-ported evolutionarily by a significant PIC (Fig. 5D).There was no cross-species association between KS andPET for deciduous species, but the independent con-trast analysis revealed a marginally significant (P ,0.10) positive evolutionary association between KS andPET (Table 3). In evergreen angiosperms, there was norelationship between KS and any climate variable ineither the cross-species or independent contrast anal-ysis (Fig. 5B, E, Table 3). Similarly, there was no re-lationship between most climate variables (MAP, MAP/PET, MAT) and KS in the conifers (Fig. 5C, F, Table
2192 HAFIZ MAHERALI ET AL. Ecology, Vol. 85, No. 8
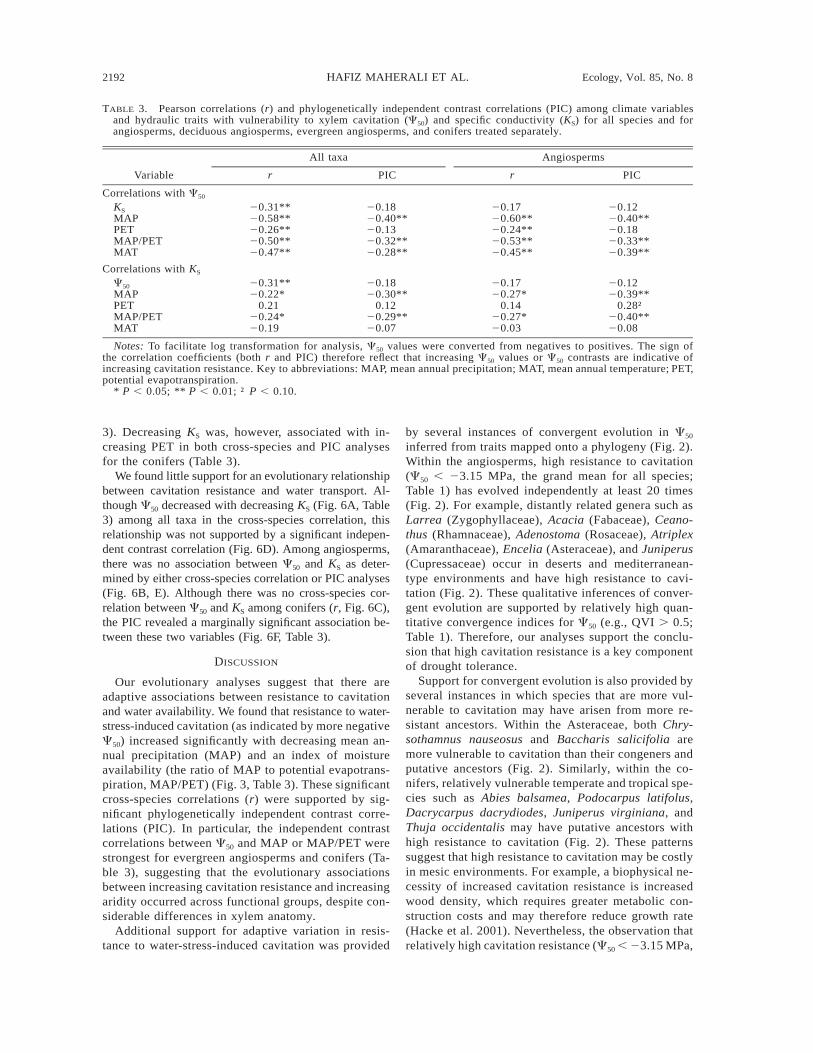
TABLE 3. Pearson correlations (r) and phylogenetically independent contrast correlations (PIC) among climate variablesand hydraulic traits with vulnerability to xylem cavitation (C50) and specific conductivity (KS) for all species and forangiosperms, deciduous angiosperms, evergreen angiosperms, and conifers treated separately.
Variable
All taxa
r PIC
Angiosperms
r PIC
Correlations with C50
KS
MAPPETMAP/PETMAT
20.31**20.58**20.26**20.50**20.47**
20.1820.40**20.1320.32**20.28**
20.1720.60**20.24**20.53**20.45**
20.1220.40**20.1820.33**20.39**
Correlations with KS
C50
MAPPETMAP/PETMAT
20.31**20.22*
0.2120.24*20.19
20.1820.30**
0.1220.29**20.07
20.1720.27*
0.1420.27*20.03
20.1220.39**
0.28†20.40**20.08
Notes: To facilitate log transformation for analysis, C50 values were converted from negatives to positives. The sign ofthe correlation coefficients (both r and PIC) therefore reflect that increasing C50 values or C50 contrasts are indicative ofincreasing cavitation resistance. Key to abbreviations: MAP, mean annual precipitation; MAT, mean annual temperature; PET,potential evapotranspiration.
* P , 0.05; ** P , 0.01; † P , 0.10.
3). Decreasing KS was, however, associated with in-creasing PET in both cross-species and PIC analysesfor the conifers (Table 3).
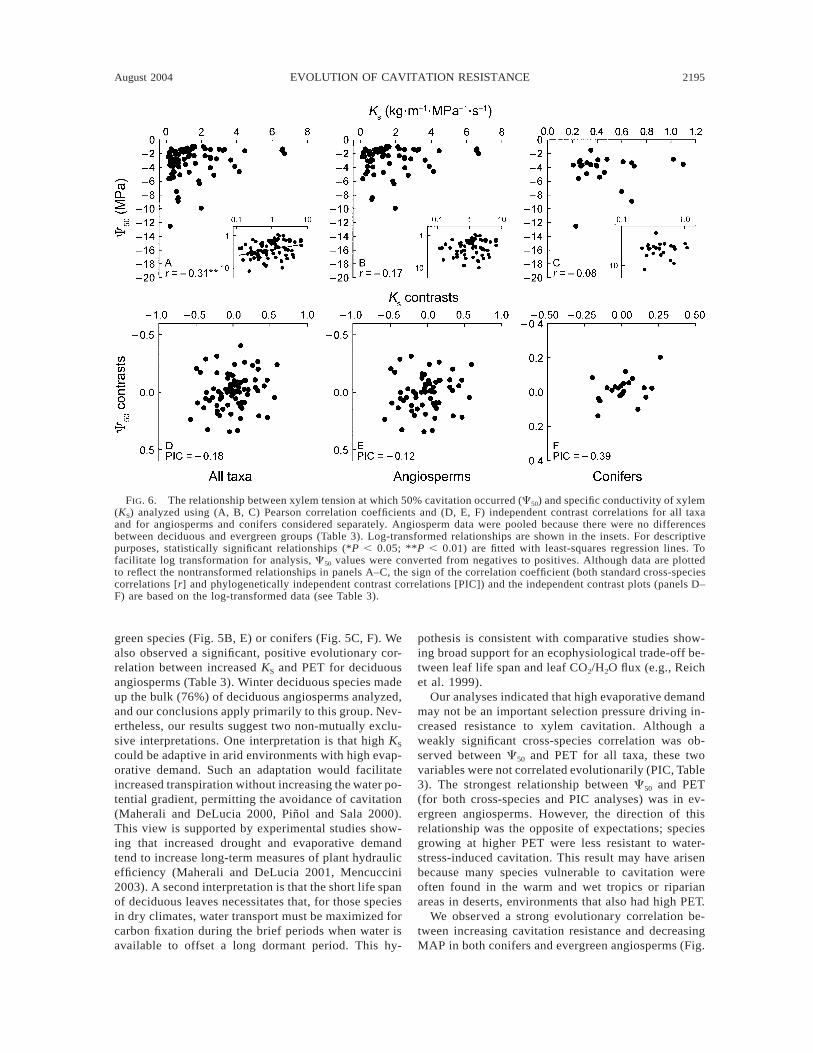
We found little support for an evolutionary relationshipbetween cavitation resistance and water transport. Al-though C50 decreased with decreasing KS (Fig. 6A, Table3) among all taxa in the cross-species correlation, thisrelationship was not supported by a significant indepen-dent contrast correlation (Fig. 6D). Among angiosperms,there was no association between C50 and KS as deter-mined by either cross-species correlation or PIC analyses(Fig. 6B, E). Although there was no cross-species cor-relation between C50 and KS among conifers (r, Fig. 6C),the PIC revealed a marginally significant association be-tween these two variables (Fig. 6F, Table 3).
DISCUSSION
Our evolutionary analyses suggest that there areadaptive associations between resistance to cavitationand water availability. We found that resistance to water-stress-induced cavitation (as indicated by more negativeC50) increased significantly with decreasing mean an-nual precipitation (MAP) and an index of moistureavailability (the ratio of MAP to potential evapotrans-piration, MAP/PET) (Fig. 3, Table 3). These significantcross-species correlations (r) were supported by sig-nificant phylogenetically independent contrast corre-lations (PIC). In particular, the independent contrastcorrelations between C50 and MAP or MAP/PET werestrongest for evergreen angiosperms and conifers (Ta-ble 3), suggesting that the evolutionary associationsbetween increasing cavitation resistance and increasingaridity occurred across functional groups, despite con-siderable differences in xylem anatomy.
Additional support for adaptive variation in resis-tance to water-stress-induced cavitation was provided
by several instances of convergent evolution in C50
inferred from traits mapped onto a phylogeny (Fig. 2).Within the angiosperms, high resistance to cavitation(C50 , 23.15 MPa, the grand mean for all species;Table 1) has evolved independently at least 20 times(Fig. 2). For example, distantly related genera such asLarrea (Zygophyllaceae), Acacia (Fabaceae), Ceano-thus (Rhamnaceae), Adenostoma (Rosaceae), Atriplex(Amaranthaceae), Encelia (Asteraceae), and Juniperus(Cupressaceae) occur in deserts and mediterranean-type environments and have high resistance to cavi-tation (Fig. 2). These qualitative inferences of conver-gent evolution are supported by relatively high quan-titative convergence indices for C50 (e.g., QVI . 0.5;Table 1). Therefore, our analyses support the conclu-sion that high cavitation resistance is a key componentof drought tolerance.
Support for convergent evolution is also provided byseveral instances in which species that are more vul-nerable to cavitation may have arisen from more re-sistant ancestors. Within the Asteraceae, both Chry-sothamnus nauseosus and Baccharis salicifolia aremore vulnerable to cavitation than their congeners andputative ancestors (Fig. 2). Similarly, within the co-nifers, relatively vulnerable temperate and tropical spe-cies such as Abies balsamea, Podocarpus latifolus,Dacrycarpus dacrydiodes, Juniperus virginiana, andThuja occidentalis may have putative ancestors withhigh resistance to cavitation (Fig. 2). These patternssuggest that high resistance to cavitation may be costlyin mesic environments. For example, a biophysical ne-cessity of increased cavitation resistance is increasedwood density, which requires greater metabolic con-struction costs and may therefore reduce growth rate(Hacke et al. 2001). Nevertheless, the observation thatrelatively high cavitation resistance (C50 , 23.15 MPa,
August 2004 2193EVOLUTION OF CAVITATION RESISTANCE
TABLE 3. Extended.
Deciduous angiosperms
r PIC
Evergreen angiosperms
r PIC
Conifers
r PIC
20.1320.36**
0.2020.3420.02
0.0420.1820.2120.0920.43**
20.2420.71**20.58**20.65**20.65**
20.1920.56**20.29*20.51**20.56**
20.0820.46**
0.2020.49**20.07
20.39†20.41*
0.2720.33†
0.12
20.1320.47**
0.2120.45**20.16
0.0420.65**
0.40*20.65**20.14
20.2420.0120.19
0.010.11
20.1920.14
0.1220.15
0.21
20.0820.0420.52*
0.2720.36
20.39†0.15
20.54**0.32
20.36
FIG. 4. The relationship between xylem tension at which 50% cavitation occurred (C50) and mean annual precipitation(MAP) analyzed using Pearson correlation coefficients and independent contrast correlations for (A, D) deciduous angio-sperms, (B, E) evergreen angiosperms, and (C, F) conifers. Log-transformed relationships are shown in the insets. Fordescriptive purposes, the relationship between C50 and MAP for deciduous angiosperms (panel A) was divided into winterdeciduous (solid circles) and drought deciduous (open circles) groups. Statistically significant relationships (*P , 0.05, **P, 0.01) are fitted with least-squares regression lines. To facilitate log transformation for analysis, C50 values were convertedfrom negatives to positives. Although data in panels A–C are plotted to reflect the nontransformed relationships, the sign ofthe correlation coefficient (both standard cross-species correlations [r] and phylogenetically independent contrast correlations[PIC]) and the independent contrast plots (panels D–F) are based on the log-transformed data (see Table 3).
2194 HAFIZ MAHERALI ET AL. Ecology, Vol. 85, No. 8
FIG. 5. The relationship between specific conductivity of xylem (KS) and mean annual precipitation (MAP) analyzedusing Pearson correlation coefficients and independent contrast correlations for (A, D) deciduous angiosperms, (B, E) evergreenangiosperms, and (C, F) conifers. Log-transformed relationships are shown in the insets. For descriptive purposes, therelationship between KS and MAP for deciduous angiosperms (panel A) was divided into winter deciduous groups (solidcircles) and drought deciduous groups (open circles). Statistically significant relationships (*P , 0.05; **P , 0.01) are fittedwith least-squares regression lines. Key to abbreviations: PIC, phylogenetically independent contrast correlations; r, standardcross-species correlations.
the mean for all species, Table 1) persists among someof these genera in temperate regions (e.g., J. virginiana,T. occidentalis) does suggest that phylogenetic historymay at least partly constrain the adaptive evolution ofcavitation resistance.
Deciduous species may avoid periods of severe waterlimitation by shedding leaves (both during drought andwinter) and, as a result, may experience weaker selec-tion for increased resistance to water-stress-inducedcavitation (Sobrado 1997, Eamus and Prior 2001). Ourresults partially supported this hypothesis. We foundno evolutionary correlation (i.e., a nonsignificant PIC)between C50 and MAP among deciduous angiosperms(Fig. 4A, D) but we note that the majority (77%) ofsamples for this comparison was drawn from winterdeciduous species. The complete absence of an evo-lutionary correlation between C50 and MAP is some-what surprising because summer drought stress com-monly occurs in habitats where winter deciduous spe-cies are found. However, it is possible that selectionpressures on xylem that favor increased resistance tofreezing-induced cavitation (e.g., Wang et al. 1992)
obscure the relationship between cavitation resistanceand rainfall in winter deciduous species. This hypoth-esis is consistent with the observation that drought de-ciduous angiosperms, when considered separately, doshow increased cavitation resistance with decreasedrainfall (Fig. 4A). Nevertheless, further studies are re-quired to determine the relative contributions of cav-itation resistance and leaf shedding to the evolution ofdrought tolerance in the drought deciduous angio-sperms. In contrast, we found a strongly significantcross-species and phylogenetically independent con-trast correlation between increased resistance to cavi-tation and decreased MAP in evergreen angiospermsand conifers (Fig. 4B, C, E, F; Table 3). Therefore, theadaptive significance of increased resistance to cavi-tation as a mechanism of drought tolerance appears tobe greatest in evergreen angiosperm and conifer spe-cies, largely because they need to preserve a water-conducting pathway for leaves year round.
We found a significant evolutionary correlation be-tween increasing KS and decreasing annual rainfall indeciduous angiosperms (Fig. 5A, D), but not in ever-
August 2004 2195EVOLUTION OF CAVITATION RESISTANCE
FIG. 6. The relationship between xylem tension at which 50% cavitation occurred (C50) and specific conductivity of xylem(KS) analyzed using (A, B, C) Pearson correlation coefficients and (D, E, F) independent contrast correlations for all taxaand for angiosperms and conifers considered separately. Angiosperm data were pooled because there were no differencesbetween deciduous and evergreen groups (Table 3). Log-transformed relationships are shown in the insets. For descriptivepurposes, statistically significant relationships (*P , 0.05; **P , 0.01) are fitted with least-squares regression lines. Tofacilitate log transformation for analysis, C50 values were converted from negatives to positives. Although data are plottedto reflect the nontransformed relationships in panels A–C, the sign of the correlation coefficient (both standard cross-speciescorrelations [r] and phylogenetically independent contrast correlations [PIC]) and the independent contrast plots (panels D–F) are based on the log-transformed data (see Table 3).
green species (Fig. 5B, E) or conifers (Fig. 5C, F). Wealso observed a significant, positive evolutionary cor-relation between increased KS and PET for deciduousangiosperms (Table 3). Winter deciduous species madeup the bulk (76%) of deciduous angiosperms analyzed,and our conclusions apply primarily to this group. Nev-ertheless, our results suggest two non-mutually exclu-sive interpretations. One interpretation is that high KS
could be adaptive in arid environments with high evap-orative demand. Such an adaptation would facilitateincreased transpiration without increasing the water po-tential gradient, permitting the avoidance of cavitation(Maherali and DeLucia 2000, Pinol and Sala 2000).This view is supported by experimental studies show-ing that increased drought and evaporative demandtend to increase long-term measures of plant hydraulicefficiency (Maherali and DeLucia 2001, Mencuccini2003). A second interpretation is that the short life spanof deciduous leaves necessitates that, for those speciesin dry climates, water transport must be maximized forcarbon fixation during the brief periods when water isavailable to offset a long dormant period. This hy-
pothesis is consistent with comparative studies show-ing broad support for an ecophysiological trade-off be-tween leaf life span and leaf CO2/H2O flux (e.g., Reichet al. 1999).
Our analyses indicated that high evaporative demandmay not be an important selection pressure driving in-creased resistance to xylem cavitation. Although aweakly significant cross-species correlation was ob-served between C50 and PET for all taxa, these twovariables were not correlated evolutionarily (PIC, Table3). The strongest relationship between C50 and PET(for both cross-species and PIC analyses) was in ev-ergreen angiosperms. However, the direction of thisrelationship was the opposite of expectations; speciesgrowing at higher PET were less resistant to water-stress-induced cavitation. This result may have arisenbecause many species vulnerable to cavitation wereoften found in the warm and wet tropics or riparianareas in deserts, environments that also had high PET.
We observed a strong evolutionary correlation be-tween increasing cavitation resistance and decreasingMAP in both conifers and evergreen angiosperms (Fig.

2196 HAFIZ MAHERALI ET AL. Ecology, Vol. 85, No. 8
4E, F). Despite similar patterns of correlated evolution,and the fact that C50 values largely overlapped (Table1, Fig. 4A, B, C), only 35% of evergreen angiosperms(and 26% of deciduous angiosperms) had C50 valuesmore negative than the grand mean for all species(23.15 MPa), whereas 77% of conifers fell into thiscategory. In consequence, conifers were, on average,80% more resistant to water-stress-induced cavitationthan evergreen angiosperms (Table 1). Although tra-cheids are not considered to be inherently superior tovessels in resisting water-stress-induced cavitation(Sperry 2003), why are a greater proportion of conifersmore resistant to water-stress-induced cavitation thanangiosperms? One explanation may be that there arefewer constraints on the evolution of high cavitationresistance in tracheid vs. vessel-bearing wood. For ex-ample, resistance to water-stress-induced cavitation viaair seeding in a large majority of angiosperms dependson the surface tension of the meniscus in each of thepores of the pit membrane that connects adjacent con-duits (Sperry 2003). As a result, increases in resistanceto cavitation must be accompanied by presumably cost-ly decreases in pore hydraulic conductivity (Zimmer-mann 1983, Sperry and Hacke 2004). In conifers, anet-like perimeter (the margo) surrounds a thickenedimpermeable center (the torus). The large pores of themargo allow high pit hydraulic conductivity but appearto place a weak functional limit on air seeding pressuresbecause the aspiration of the torus is responsible forblocking the spread of air between conduits followingcavitation (Sperry 2003, Hacke et al. 2004). Furtherair seeding depends on the elasticity and mechanicalstrength of the torus/margo complex (Sperry and Tyree1990). Because increased air seeding pressure (and thusincreased cavitation resistance) is not accompanied byincreased pit hydraulic resistance, conifers may moreeasily respond to selection for increased cavitation re-sistance than angiosperms.
Given the apparent trade-off between air seedingpressure and pore conductivity, it has long been sug-gested that increased resistance to water-stress-inducedcavitation should be associated with reduced xylemhydraulic transport capacity in the stem (KS), particu-larly in angiosperms (Zimmermann 1983, Tyree andSperry 1989, Tyree and Ewers 1991). This trade-offhas been observed within an individual (e.g., Tyree andSperry 1989, Sperry and Saliendra 1994), but is moredifficult to detect across individuals (Kavanagh et al.1999, Maherali and DeLucia 2000) or species (Tyreeet al. 1994, Pockman and Sperry 2000). We found asignificant, though weak, cross-species correlation be-tween C50 and KS (Fig. 6A). However, there was noevolutionary correlation between the two traits, sug-gesting that there may not be an evolutionary trade-offbetween resistance to cavitation and specific conduc-tivity in woody plants, at least at the broad taxonomicscale of this study. Because angiosperms are on averageless resistant to cavitation than conifers and have higher
KS, the significant cross-species correlation may becaused by the ancestral divergence between these twogroups (e.g., Ackerly and Reich 1999). The absence ofan evolutionary correlation between C50 and KS in an-giosperms, despite a trade-off between air seeding pres-sure and pit conductivity (Sperry and Hacke 2004),may be associated with changes in other traits. Forexample, increased vessel length and diameter both in-crease hydraulic conductivity and may compensate forhydraulic limitations in the pit membrane (Comstockand Sperry 2000, Hacke et al. 2004, Sperry and Hacke2004). Despite the absence of a cross-species corre-lation between C50 and KS in conifers (Fig. 6C), therewas a marginally significant PIC between these twotraits (Fig. 6F), suggesting that there may be a weakevolutionary trade-off between C50 and KS in conifers.
Our study, like all meta-analyses with broad inter-specific data sets, has limitations. In particular, inter-specific relationships analyzed using both traditionalcross-species analysis and independent contrasts arelimited by the inability to randomly sample taxa (Don-oghue and Ackerly 1996, Ackerly 2000). In addition,although our sample was drawn from a broad taxo-nomic range, several families were represented only byone or two species (e.g., Moraceae, Rhizophoraceae,Zygophyllaceae), whereas others had several species(e.g., Fagaceae, Pinaceae, Cupressaceae). Several eco-logically important genera spanning large climate gra-dients (e.g., Eucalyptus, Nothofagus, Ambrosia) werealso not well represented. Finally, our sample only in-cludes woody plants and the C50 and KS values reportedhere may not necessarily be representative of the dis-tribution of these traits across all seed plants (Ackerly2000). Simulation analyses suggest that this type ofnonrandom sampling could result in the underestima-tion of true PIC correlation coefficients (Ackerly 2000).Our analysis of correlated evolution was therefore con-servative, and the significant PICs we report in thisstudy should be regarded as robust. Nevertheless,weaker relationships may have gone undetected,though this limitation must be balanced against inflatedType I error rates of non-phylogenetically correctedcross-species correlations (Garland et al. 1992, Purviset al. 1994, Ackerly 2000).
Our study suggests several avenues of future re-search. First, the cavitation resistance of roots, like thatof stems, can limit transpiration (Alder et al. 1996,Jackson et al. 2000). Although root data were too lim-ited to include in our analysis, roots are generally morevulnerable to water-stress-induced cavitation thanstems (Jackson et al. 2000), and thus we predict thatinterspecific variation in root cavitation resistance alsohas adaptive consequences. Second, our analysis doesnot specifically incorporate vulnerability to freezing-induced cavitation, which is controlled by the volumeof conducting elements in the xylem (Davis et al.1999). The substantial variation that we observed inKS indicates that there is large variation in conduit di-

August 2004 2197EVOLUTION OF CAVITATION RESISTANCE
ameter (e.g., 10 mm to .500 mm; Tyree et al. 1994),and therefore conduit volume, among species in ourdatabase. However, sufficient data on freezing-inducedcavitation were not available for our analysis. Simul-taneous consideration of water-stress-induced andfreezing-induced cavitation in a phylogenetic frame-work, particularly among closely related species thatspan large gradients in precipitation and temperature,will likely be fruitful. Finally, our analysis assumesthat embolism formation is largely irreversible in theabsence of new growth (Zimmermann 1983) or underconditions of positive root or stem pressure (Ewers etal. 1997, Tyree et al. 1999). However, recent studiessuggest that the refilling of emboli may occur duringtranspiration (McCully 1999, Tyree et al. 1999) causinghydraulic conductivity to vary diurnally (Zwienieckiand Holbrook 1998). The mechanism of cavitation re-pair under tension is not known, nor is there enoughinformation in the literature to determine whether it iscommon in many species. Continued research on theseprocesses (Holbrook et al. 2001, Meinzer et al. 2001)across a range of taxa will improve our understandingof the evolutionary and ecological significance of cav-itation resistance.
Additional variables such as the leaf-specific con-ductivity (KL) and the ratio of leaf area to sapwood area(AL/AS) are important in controlling water supply toleaves and are likely associated with variation in re-sistance to xylem cavitation. These variables were rare-ly reported in the studies we used to construct ourdatabase, preventing us from examining several addi-tional hypotheses that deserve further study. For ex-ample, the observation that KL scales directly with leaftranspiration (Mencuccini 2003) suggests that the hy-pothesized trade-off between cavitation resistance andwater transport could be manifested when leaf levelconductivity is taken into account. Alternatively, be-cause increased KL, like KS, facilitates increased tran-spiration without a rise in the water potential gradient,both high KL and high cavitation resistance (a morenegative C50) could represent strategies for coping withprolonged drought (Tyree and Ewers 1991), obscuringa potential trade-off between KL and cavitation resis-tance. Observations of a decrease in AL/AS, which tendsto increase KL, in response to increasing aridity in manyconifer taxa (DeLucia et al. 2000) are consistent withthis view. This decline in AL/AS with increased droughtalso suggests that partial leaf shedding could also bean evolutionarily important response to aridity in ev-ergreen angiosperms and conifers.
Our findings highlight the importance of examiningfunctional correlations in a phylogenetic context in or-der to understand how interspecific variation in planttraits arose. Among other insights, we observed thatcavitation resistance likely has different ecological andevolutionary consequences for deciduous vs. evergreenangiosperms and conifers. Our wide taxonomic sampleprovides evidence of broad patterns, but further studies
at smaller scales (i.e., within specific families or gen-era) that combine physiological data, molecular phy-logenetic hypotheses, and biogeographic informationwill help provide comprehensive tests of the evolutionof functional changes in cavitation resistance. For ex-ample, we found no trade-off between cavitation re-sistance and specific conductivity in a broad sample,but evolutionary associations between these traits mayoccur within specific groups that have very similar leafand xylem structure (e.g., Pinol and Sala 2000; Fig.6F). Resistance to water-stress-induced cavitation is acritical component controlling canopy transpirationand water uptake (Sperry et al. 2002), and interspecificvariation in this trait can influence hydrological pro-cesses (Jackson et al. 2000, 2001). Further understand-ing of the evolutionary origins and ecological signifi-cance of xylem transport and cavitation resistance islikely to improve predictions of plant and ecosystemwater use on both local and global scales and in re-sponse to future environmental change.
ACKNOWLEDGMENTS
This work was supported by grants from the National Sci-ence Foundation, The Natural Sciences and Engineering Re-search Council, the Andrew W. Mellon Foundation, and theNational Institute for Global Environmental Change throughthe U.S. Department of Energy. We thank P. S. Manos formany helpful insights on the construction of phylogenetictrees for comparative analysis. We also thank C. M. Caruso,C. W. Cook, V. C. Engel, P. S. Manos, A. McElrone, C. F.Moura, K. Ogle, C. D. Reid, J. H. Richards, A. Sala, C. J.Willson, and an anonymous reviewer for providing valuablecritiques of the manuscript.
LITERATURE CITED
Ackerly, D. D. 1999. Phylogeny and the comparative methodin plant functional ecology. Pages 391–413 in M. C. Press,J. D. Scholes, and M. G. Baker, editors. Plant physiologicalecology. Blackwell Scientific Press, Oxford, UK.
Ackerly, D. D. 2000. Taxon sampling, correlated evolution,and independent contrasts. Evolution 54:1480–1492.
Ackerly, D. D., and M. J. Donoghue. 1998. Leaf size, saplingallometry, and Corner’s rules: a phylogenetic study of cor-related evolution in maples (Acer). American Naturalist152:767–791.
Ackerly, D. D., and P. B. Reich. 1999. Convergence andcorrelations among leaf size and function in seed plants: acomparative test using independent contrasts. AmericanJournal of Botany 86:1272–1281.
Alder, N. N., W. T. Pockman, J. S. Sperry, and S. Nuismer.1997. Use of centrifugal force in the study of xylem cav-itation. Journal of Experimental Botany 48:665–674.
Alder, N. N., J. S. Sperry, and W. T. Pockman. 1996. Rootand stem xylem embolism, stomatal conductance and leafturgor in Acer grandidentatum populations along a soilmoisture gradient. Oecologia 105:293–301.
Brodribb, T., and R. S. Hill. 1999. The importance of xylemconstraints in the distribution of conifer species. New Phy-tologist 143:365–372.
Carlquist, S. J. 1988. Comparative wood anatomy: system-atic, ecological, and evolutionary aspects of dicotyledonwood. Springer, New York, New York, USA.
Choudhury, B. J. 1997. Global pattern of potential evapo-ration calculated from the Penman-Monteith equation usingsatellite and assimilated data. Remote Sensing of Environ-ment 61:64–81.

2198 HAFIZ MAHERALI ET AL. Ecology, Vol. 85, No. 8
Choudhury, B. J., and N. E. DiGirolamo. 1998. A biophysicalprocess-based estimate of global land surface evaporationusing satellite and ancillary data—I. Model description andcomparison with observations. Journal of Hydrology 205:164–185.
Cochard, H., P. Cruiziat, and M. T. Tyree. 1992. Use of pos-itive pressures to establish vulnerability curves. Plant Phys-iology 100:205–209.
Comstock, J. P., and J. S. Sperry. 2000. Some theoreticalconsiderations of optimal conduit length for water transportin plants. New Phytologist 148:195–218.
Cunningham, C. W., K. E. Omland, and T. H. Oakley. 1998.Reconstructing ancestral character states: a critical reap-praisal. Trends in Ecology and Evolution 13:361–366.
Davis, S. D., J. S. Sperry, and U. G. Hacke. 1999. The re-lationship between xylem conduit diameter and cavitationcaused by freezing. American Journal of Botany 86:1367–1372.
DeLucia, E. H., H. Maherali, and E. V. Carey. 2000. Climate-driven changes in biomass allocation in pines. GlobalChange Biology 6:587–593.
Donoghue, M. J., and D. D. Ackerly. 1996. Phylogeneticuncertainties and sensitivity analyses in comparative bi-ology. Philosophical Transactions of the Royal Society ofLondon, Series B 351:1241–1249.
Eamus, D., and L. Prior. 2001. Ecophysiology of trees ofseasonally dry tropics: comparisons among phenologies.Advances in Ecological Research 32:113–197.
Ewers, F. W., H. Cochard, and M. T. Tyree. 1997. A surveyof root pressures in vines of a tropical lowland forest. Oec-ologia 110:191–196.
Felsenstein, J. 1985. Phylogenies and the comparative meth-od. American Naturalist 125:1–15.
Garland, T., Jr., P. H. Harvey, and A. R. Ives. 1992. Proce-dures for the analysis of comparative data using phyloge-netically independent contrasts. Systematic Biology 41:18–32.
Hacke, U. G., J. S. Sperry, and J. Pittermann. 2004. Analysisof circular bordered pit function. II. Gymnosperm tracheidswith torus-margo pit membranes. American Journal of Bot-any 91:386–400.
Hacke, U. G., J. S. Sperry, W. T. Pockman, S. D. Davis, andK. A. McCulloch. 2001. Trends in wood density and struc-ture are linked to prevention of xylem implosion by neg-ative pressure. Oecologia 126:457–461.
Harvey, P. H., and M. Pagel. 1991. The comparative methodin evolutionary biology. Oxford University Press, Oxford,UK.
Holbrook, N. M., E. T. Ahrens, M. J. Burns, and M. A. Zwien-iecki. 2001. In vivo observation of cavitation and embo-lism repair using magnetic resonance imaging. Plant Phys-iology 126:27–31.
Jackson, R. B., S. R. Carpenter, C. N. Dahm, D. M. McKnight,R. J. Naiman, S. L. Postel, and S. W. Running. 2001. Waterin a changing world. Ecological Applications 11:1027–1045.
Jackson, R. B., J. S. Sperry, and T. E. Dawson. 2000. Rootwater uptake and transport: using physiological processesin global predictions. Trends in Plant Science 5:482–488.
Jarbeau, J. A., F. W. Ewers, and S. D. Davis. 1995. Themechanism of water stress induced embolism in two speciesof chaparral shrubs. Plant Cell and Environment 18:189–196.
Kavanagh, K. L., B. J. Bond, S. N. Aitken, B. L. Gartner,and S. Knowe. 1999. Shoot and root vulnerability to xylemcavitation in four populations of Douglas-fir seedlings. TreePhysiology 19:31–37.
Kolb, K. J., and J. S. Sperry. 1999. Differences in droughtadaptation between subspecies of sagebrush (Artemisia tri-dentata). Ecology 80:2373–2384.
Leemans, R., and W. Cramer. 1991. The IIASA database formean monthly values of temperature, precipitation andcloudiness of a global terrestrial grid. Report Number RR-91–18. International Institute for Applied Systems Anal-ysis, Laxenburg, Austria.
Maddison, W. P., and D. R. Maddison. 2000. MacClade 4.0:analysis of phylogeny and character evolution. Sinauer,Sunderland, Massachusetts, USA.
Maherali, H., and E. H. DeLucia. 2000. Xylem conductivityand vulnerability to cavitation of ponderosa pine growingin contrasting climates. Tree Physiology 20:859–867.
Maherali, H., and E. H. DeLucia. 2001. Influence of climate-driven shifts in biomass allocation on water transport andstorage in ponderosa pine. Oecologia 129:481–491.
Matzner, S. L., K. J. Rice, and J. H. Richards. 2001. Intra-specific variation in xylem cavitation in interior live oak(Quercus wislizenii). Journal of Experimental Botany 52:783–789.
McCully, M. E. 1999. Root xylem embolisms and refilling.Relation to water potentials of soil, roots, and leaves, andosmotic potentials of root xylem sap. Plant Physiology 119:1001–1008.
Meinzer, F. C., M. J. Clearwater, and G. Goldstein. 2001.Water transport in trees: current perspectives, new insightsand some controversies. Environmental and ExperimentalBotany 45:239–262.
Mencuccini, M. 2003. The ecological significance of long-distance water transport: short term regulation, long termacclimation and the hydraulic costs of stature across plantlife forms. Plant Cell and Environment 26:163–182.
Pagel, M. D. 1993. Seeking the evolutionary regression co-efficient: an analysis of what comparative methods mea-sure. Journal of Theoretical Biology 164:191–205.
Pataki, D. E., R. Oren, and N. Phillips. 1998. Responses ofsap flux and stomatal conductance of Pinus taeda L. treesto stepwise reductions in leaf area. Journal of ExperimentalBotany 49:871–878.
Pinol, J., and A. Sala. 2000. Ecological implications of xylemcavitation for several Pinaceae in the Pacific northernU.S.A. Functional Ecology 14:538–545.
Pockman, W. T., and J. S. Sperry. 2000. Vulnerability toxylem cavitation and the distribution of Sonoran desertvegetation. American Journal of Botany 87:1287–1299.
Pockman, W. T., J. S. Sperry, and J. W. O’Leary. 1995. Sus-tained and significant negative water pressure in xylem.Nature 378:715–716.
Purvis, A., J. L. Gittleman, and H. K. Luh. 1994. Truth orconsequences: effects of phylogenetic accuracy on twocomparative methods. Journal of Theoretical Biology 167:293–300.
Reich, P. B., D. S. Ellsworth, M. B. Walters, J. Vose, C.Gresham, J. Volin, and W. Bowman. 1999. Generality ofleaf traits relationships: a test across six biomes. Ecology80:1955–1969.
Schenk, H. J., and R. B. Jackson. 2002. The global bioge-ography of roots. Ecological Monographs 72:311–328.
Schwilk, D. W., and D. D. Ackerly. 2001. Flammability andserotiny as strategies: correlated evolution in pines. Oikos94:326–336.
Sobrado, M. A. 1997. Embolism vulnerability in drought-deciduous and evergreen species of a tropical dry forest.Acta Œcologica 18:383–391.
Soltis, P. S., D. E. Soltis, and M. W. Chase. 1999. Angiospermphylogeny inferred from multiple genes as a tool for com-parative biology. Nature 402:402–404.
Sperry, J. S. 2003. Evolution of water transport and xylemstructure. International Journal of Plant Science 164:S115–S127.
Sperry, J. S., J. R. Donnelly, and M. T. Tyree. 1988. A methodfor measuring hydraulic conductivity and embolism in xy-lem. Plant Cell and Environment 11:35–40.

August 2004 2199EVOLUTION OF CAVITATION RESISTANCE
Sperry, J. S., and U. G. Hacke. 2004. Analysis of circularbordered pit function. I. Angiosperm vessels with homog-enous pit membranes. American Journal of Botany, 91:369–385.
Sperry, J. S., U. G. Hacke, R. Oren, and J. P. Comstock. 2002.Water deficits and hydraulic limits to leaf water supply.Plant Cell and Environment 25:251–263.
Sperry, J. S., K. L. Nichols, J. E. M. Sullivan, and S. E.Eastlack. 1994. Xylem embolism in ring-porous, diffuse-porous and coniferous trees of northern Utah and interiorAlaska. Ecology 75:1736–1752.
Sperry, J. S., and W. T. Pockman. 1993. Limitation of tran-spiration by hydraulic conductance and xylem cavitationin Betula occidentalis. Plant, Cell and Environment 16:279–287.
Sperry, J. S., and N. Z. Saliendra. 1994. Intra-and inter-plantvariation in xylem cavitation in Betula occidentalis. Plant,Cell and Environment 17:1233–1241.
Sperry, J. S., and M. T. Tyree. 1988. Mechanism of waterstress-induced xylem embolism. Plant Physiology 88:581–587.
Sperry, J. S., and M. T. Tyree. 1990. Water-stress-inducedxylem embolism in three species of conifers. Plant Cell andEnvironment 13:427–436.
Stefanovic, S., M. Jager, J. Deutsch, J. Broutin, and M. Mas-selot. 1998. Phylogenetic relationships of conifers inferred
from partial 28S rRNA gene sequences. American Journalof Botany 85:688–697.
Tyree, M. T., S. D. Davis, and H. Cochard. 1994. Biophysicalperspectives of xylem evolution: Is there a tradeoff of hy-draulic efficiency for vulnerability to dysfunction? Inter-national Association of Wood Anatomists Journal 15:335–360.
Tyree, M. T., and F. W. Ewers. 1991. The hydraulic archi-tecture of trees and other woody plants. New Phytologist119:345–360.
Tyree, M. T., S. Salleo, A. Nardini, M. A. Lo Gullo, and R.Mosca. 1999. Refilling of embolized vessels in youngstems of laurel: Do we need a new paradigm? Plant Phys-iology 120:11–21.
Tyree, M. T., and J. S. Sperry. 1989. Vulnerability of xylemto cavitation and embolism. Annual Review of Plant Phys-iology and Plant Molecular Biology 40:19–38.
Wang, J., N. E. Ives, and M. J. Lechowicz. 1992. The relationof foliar phenology to xylem embolism in trees. FunctionalEcology 6:469–475.
Zimmermann, M. H. 1983. Xylem structure and the ascentof sap. Springer, New York, New York, USA.
Zwieniecki, M. A., and N. M. Holbrook. 1998. Diurnal var-iation in xylem hydraulic conductivity in white ash (Frax-inus americana L.), red maple (Acer rubrum L.) and redspruce (Picea rubens Sarg.). Plant Cell and Environment21:1173–1180.
APPENDIX A
A table presenting species, their geographic locations, and studies used in the database is available in ESA’s ElectronicData Archive: Ecological Archives E085-067-A1.
APPENDIX B
A table presenting sources of phylogenetic information and polytomy resolutions for selected taxa is available in ESA’sElectronic Data Archive: Ecological Archives E085-067-A2.