all about adrenal gland of mammalprosthetic gland, adrenal glands etc.
Adrenal gland in mammalsInmammals, theadrenal glands(also known
assuprarenal glands) areendocrine glandsthat sit at the top of
thekidneys; in humans, the right adrenal gland is triangular
shaped, while the left adrenal gland is semilunar shaped.[1]They
are chiefly responsible for releasinghormonesin response
tostressthrough thesynthesisofcorticosteroidssuch
ascortisolandcatecholaminessuch asadrenaline(epinephrine)
andnoradrenaline. These endocrine glands also produceandrogensin
their innermost cortical layer. The adrenal glands affect kidney
function through the secretion ofaldosterone, and recent data
(1998) suggest that adrenocortical cells underpathologicalas well
as underphysiologicalconditions showneuroendocrineproperties;
within the normal adrenal, this neuroendocrine differentiation
seems to be restricted to cells of thezona glomerulosaand might be
important for anautocrineregulation of adrenocortical
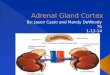
function.[2]Structure[edit]The adrenal glands are located in
theretroperitoneumsuperior to thekidneys, they are quadrilaterial
in shape and are situated bilaterally. The combined weight of the
adrenal glands in an adult human ranges from 7 to 10grams.[3]They
are surrounded by anadipose capsuleandrenal fascia.Each adrenal
gland has two distinct structures, the outeradrenal cortexand the
innermedulla, both of which produce hormones. The cortex mainly
producescortisol,aldosteroneandandrogens, while the medulla chiefly
producesadrenalineandnoradrenaline. In contrast to the direct
innervation of the medulla, the cortex is regulated
byneuroendocrinehormones secreted from thepituitary glandwhich are
under the control of thehypothalamus, as well as by
therenin-angiotensin system.Cortex[edit]Main article:Adrenal
cortexTheadrenal cortexis devoted to production
ofcorticosteroidandandrogenhormones. Specific cortical cells
produce particular hormones includingaldosterone,cortisol,
andandrogenssuch asandrostenedione. Under normal unstressed
conditions, the human adrenal glands produce the equivalent of
3540mg of cortisone acetate per day.[4]The adrenal cortex comprises
three zones, or layers. Thisanatomic zonationcan be appreciated at
the microscopic level, where each zone can be recognized and
distinguished from one another based on structural and anatomic
characteristics.[5]The adrenal cortex exhibitsfunctional zonationas
well: by virtue of the characteristic enzymes present in each zone,
the zones produce and secrete distinct hormones.[5]Zona
glomerulosa[edit]The outermost layer, thezona glomerulosais the
main site for production ofaldosterone, amineralocorticoid, by the
action of the enzymealdosterone synthase(also known
asCYP11B2).[6][7]Aldosterone is largely responsible for the
long-termregulation of blood pressure.[8]The expression of
neuron-specific proteins in the zona glomerulosa cells of human
adrenocortical tissues has been predicted and reported by several
authors[2][9][10]and it was suggested that the expression of
proteins like theneuronal cell adhesion molecule(NCAM) in the cells
of the zona glomerulosa reflects the regenerative feature of these
cells, which would lose NCAM immunoreactivity after moving to
thezona fasciculata.[2][11]However, together with other data on
neuroendocrine properties of zona glomerulosa cells, NCAM
expression may reflect a neuroendocrine differentiation of these
cells.[2]
Paraffin sections ofhuman adrenalsimmunostained forneuronal cell
adhesion molecule(NCAM). Immunohistochemistry was carried out using
4-amino-9-ethylcarbazole(AEC; Dinanova, Hamburg, Germnay) and were
counterstained with hematoxylin. Staining for NCAM was restricted
to thezona glomerulosa(zg) and the adrenal medulla (m); a: x 20; b:
x 200.[2].Zona fasciculata[edit]Situated between the glomerulosa
and reticularis, thezona fasciculatais responsible for
producingglucocorticoids, such
as11-deoxycorticosterone,corticosterone, andcortisolin
humans.[12]Zona reticularis[edit]The inner most cortical layer,
thezona reticularisproducesandrogens,
mainlydehydroepiandrosterone(DHEA),DHEA sulfate(DHEA-S),
andandrostenedione(the precursor totestosterone) in
humans.[12]Medulla[edit]Theadrenal medullais the core of the
adrenal gland, and is surrounded by the adrenal cortex. It secretes
approximately 20% noradrenaline (norepinephrine) and 80% adrenaline
(epinephrine).[12]Thechromaffin cellsof the medulla, named for
their characteristic brown staining withchromic acidsalts, are the
body's main source of the circulatingcatecholaminesadrenaline and
noradrenaline. Catecholamines are derived from the amino
acidtyrosineand these water-soluble hormones are the major hormones
underlying thefight-or-flight response.To carry out its part of
this response, the adrenal medulla receives input from
thesympathetic nervous systemthroughpreganglionic fibersoriginating
in thethoracic spinal cordfrom T5T11.[13]Because it is innervated
bypreganglionic nerve fibers, the adrenal medulla can be considered
as a specializedsympathetic ganglion.[13]Unlike other sympathetic
ganglia, however, the adrenal medulla lacks distinct synapses and
releases its secretions directly into the blood.Cortisol also
promotes adrenaline synthesis in the medulla. Produced in the
cortex, cortisol reaches the adrenal medulla and at high levels,
the hormone can promote the upregulation
ofphenylethanolamineN-methyltransferase(PNMT), thereby increasing
adrenaline synthesis and secretion.[5]
Blood supply[edit]Although variations of the blood supply to the
adrenal glands (and indeed the kidneys themselves) are common,
there are usually three arteries that supply each adrenal gland:
Thesuperior suprarenal arteryis provided by theinferior phrenic
artery Themiddle suprarenal arteryis provided by theabdominal aorta
Theinferior suprarenal arteryis provided by therenal
arteryVenousdrainage of the adrenal glands is achieved via
thesuprarenal veins: Theright suprarenal veindrains into
theinferior vena cava Theleft suprarenal veindrains into the
leftrenal veinor the leftinferior phrenic vein.Thesuprarenal
veinsmay formanastomoseswith theinferior phrenic veins. Since the
right supra-renal vein is short and drains directly into the
inferior vena cava it is likely to injure the latter during removal
of right adrenal for various reasons.The adrenal glands and
thethyroid glandare the organs that have the greatest blood supply
per gram of tissue. Up to 60arteriolesmay enter each adrenal
gland.[14]This may be one of the reasons lung cancer commonly
metastasizes to the adrenals.Function[edit]Aldosterone and
mineralocorticoids[edit]Aldosterone's effects are on thedistal
convoluted tubuleandcollecting duct of the kidneywhere it causes
increased reabsorption of sodium and increased excretion of both
potassium (by principal cells) and hydrogen ions (by intercalated
cells of the collecting duct).[8]Sodium retention is also a
response of the distal colon, and sweat glands to aldosterone
receptor stimulation. Although sustained production of aldosterone
requires persistentcalciumentry through low-voltage
activatedCa2+channels, isolated zona glomerulosa cells are
considered nonexcitable, with recorded membrane voltages that are
too hyperpolarized to permitCa2+channels entry.[15]However, mouse
zona glomerulosa cells within adrenal slices spontaneously generate
membrane potential oscillations of low periodicity; this innate
electrical excitability of zona glomerulosa cells provides a
platform for the production of a recurrent Ca2+channels signal that
can be controlled byangiotensin IIand extracellularpotassium, the 2
major regulators of aldosterone production.[15]Angiotensin II
originates from plasmaticangiotensin Iafter the conversion
ofangiotensinogenbyreninproduced by thejuxtaglomerular cellsof
thekidney.[12]Cortisol and glucocorticoids[edit]Cortisol is the
main glucocorticoid under normal conditions and its actions include
mobilization of fats, proteins, and carbohydrates, but it does not
increase under starvation conditions.[12]Additionally, cortisol
enhances the activity of other hormones including glucagon and
catecholamines. The zona fasciculata secretes a basal level of
cortisol but can also produce bursts of the hormone in response
toadrenocorticotropic hormone(ACTH) from theanterior
pituitary.Androgen production[edit]Adrenaline and
noradrenaline[edit]Clinical significance[edit] Severaladrenal
tumorscause symptoms because they result in the over- or
underproduction of certain hormones by the adrenal gland.
Inhyperaldosteronismthe adrenal glands produce too much
aldosterone. Inpheochromocytomathe adrenal glands secretes
excessive amounts of catecholamines. In endogenousCushing's
syndromethe adrenal glands produce too much cortisol. Adrenal
insufficiencydenotes a group of diseases characterized by
underproduction of cortisol or aldosterone. They can be caused by
problems in the adrenal glands themselves, or by impairment of the
pituitary gland or hypothalamus. TheACTH stimulation testmay assist
in diagnosis. Addison's diseaseis a rare disorder in which the
adrenal glands do not produce sufficient amounts
ofglucocorticoids(mainly cortisol). This can be caused by
anautoimmune reaction, by certain infections or by some other rarer
causes. Congenital adrenal hyperplasiasare genetic defects of
enzymes involved in cortisol production and can affect sex
characteristics of affected patients. WaterhouseFriderichsen
syndromeis adrenal gland failure due to bleeding into the adrenal
glands, caused by severe bacterial infection.
Isolatedhypoaldosteronismcan rarely occur due toaldosterone
synthasedeficiency Absent adrenal gland, rare congenital
condition
Nuclear binding and degradationWhen the RNA polymerase reaches
the end of mRNA coding genes, the nascent transcript is cleaved and
a poly(A) tail is appended to the 3 end of the molecule. A large
complex of proteins, the cleavageand polyadenylation machinery, is
responsible for the recognition of 3-processing signals and for
cleavage, and an associated poly(A) polymerase (Pap1p) synthesizes
the terminal polyadenylate (Buratowski, 2005andZhao etal., 1999).
The presence of a 3 poly(A) tail is both a signal that the mRNA
synthesis is complete and a functional element that will promote
export from the nucleus and translation. Its position close to the
end of the coding region of the mRNA is also a hallmark of quality,
preventing degradation by the quality control system that detects
the occurrence of premature stop codons. The size of the poly(A)
tail (70100 nt inSaccharomyces cerevisiae) is also crucial, as its
shortening in the cytoplasm is the first event committing the
transcript to degradation. Poly(A)-binding proteins are required
for a readout of the tail functions and to influence its synthesis
and length by affecting the processivity of the poly(A) polymerase
(Mangus etal., 2003). InS. cerevisiae, two proteins (Pab1p and
Nab2p) that recognize the poly(A) tail have been described and
extensively studied. Although both proteins shuttle between the
nucleus and the cytoplasm, their main subcellular localization is
cytoplasmic (Pab1p) and nuclear (Nab2p). While a cytoplasmic role
for Pab1p in translation is clearly established, the nuclear role
of the two proteins is still a matter of controversy, and it is not
clear which one is directly involved in the regulation of poly(A)
tail synthesis and its nuclear function(s) (Dunn etal.,
2005andHector etal., 2002).Poly(A) tailing is not only instrumental
toexpress the message carried by an RNA, but also to promote
maturation or degradation. A different polyadenylation pathway,
dependent on an alternative poly(A) polymerase (Trf4p or Trf5p
inS.cerevisiae), exists (Houseley and Tollervey, 2008). This
pathway tags a different class of substrates for faster (or more
efficient) degradation by the nuclear exosome, a protein complex
endowed with both exo- and endonuclease activity (Lebreton and
Seraphin, 2008). In many cases, degradation is halted by stable
ribonucleoprotein complexes, and exonuclease trimming results in
the production of mature and stable RNAs, as in the case of the
small nucleolar RNAs (snoRNAs). It is unclear whether degradative
polyadenylation is coupled to the termination of transcription
and/or to 3 end processing of the primary transcript or if it
occurs posttranscriptionally on substrates that cannot be degraded
efficiently by the exosome, for instance due to secondary
structures. Importantly, it is also unclear whether the
unstructured poly(A) tail is required invivo to promote more
efficient degradation or whether it somehow serves for addressing
the exosome to the substrate.Thus, the poly(A) tail can promote
very different fates, which raises the important question of what
differs in the readout of the same signal. One possibility is that
binding of a poly(A)-binding protein (e.g., Pab1p) after cleavage
and polyadenylation marks the poly(A) tails that have a cytoplasmic
fate while masking at the same time a dangerous degradation signal.
A poly(A) tail that would not associate with poly(A)-binding
proteins during or shortly after synthesis (as could be the case of
a tail added posttranscriptionally by Trf4p) would be exposed to
the degradative activity of the exosome. The paper from Bachand and
colleagues in this issue ofMolecular Cell(Lemay etal., 2010)
challenges to some extent this view and provides a unique
perspective on the function of poly(A)-binding proteins. The
authors show that deletion of the gene encoding a nuclear
poly(A)-binding protein inSchizosaccharomyces pombe, Pab2, leads to
stabilization of extended forms of several snoRNAs. Importantly,
these precursors to the mature snoRNAs are polyadenylated by a
poly(A) polymerase that turns out to be distinct from theS.
pombehomolog of Trf4p. The same extended forms are stabilized by
deletion of Rrp6p (a nuclear exonuclease that is associated with
the exosome), but in a double mutantrrp6pab2 a further
stabilization of the precursors was not observed, suggesting that
Rrp6p and Pab2p work in the same pathway. Importantly, Pab2p binds
these polyadenylated forms invivo and interacts directly with
Rrp6p, both invivo and invitro. This suggests that the binding of
Pab2 to the poly(A) tail is required to target these extended
transcripts to degradation via the direct interaction between Pab2p
and the exosome (Figure1). These findings have two important
implications: first, they provide evidence that the presence of the
poly(A) tail can recruit the exosome (as opposed to favoring its
catalytic activity); second, the mere (and presumably early)
binding of a poly(A)-binding protein is not sufficient to prevent
degradation but actually, in some conditions, promotes it. The
authors suggest that this might reveal the existence of an
alternative pathway to generate a mature form of snoRNAs: either
transcription ends at a proximal site (presumably via the action of
a homolog of theS. cerevisiaeNrd1 complex, involved in termination
of independently transcribed snoRNA genes) or it ends at a
downstream site where the nascent transcript is processed by the
cleavage and polyadenylation machinery and where it interacts with
Pab2b. The presence of Pab2 would lead to the recruitment of the
exosome and subsequent trimming of these transcripts to their
mature length.
However, there is more to the story. In another recent article
(Lemieux and Bachand, 2009), the same group shows that Pab2p also
binds cotranscriptionally to mRNAs and that this occurs even before
the poly(A) tail synthesis (Figure1B). In spite of this, no
stabilization of the tested mRNAs nor an effect on the length of
the poly(A) tail was observed inpab2 cells (Lemay etal., 2010),
suggesting that the presence of Pab2p on the poly(A) tail does not
target these transcripts to the nuclear degradation machinery.
Rather, Pab2p is found in polysomes, indicating that it remains
associated with the mRNA during translation. Therefore, what
determines whether RNAs that share the same tail, possibly
generated by the same apparatus and with the same protein bound to
it, are targeted to the exosome for degradation or to the cytoplasm
for translation remains a hot question. It is obviously possible
that other proteins, aside from Pab2p, bind differentially to mRNAs
and snoRNAs, but it remains unexplained how these species are
distinguished in the act of transcription if they are processed by
the same 3 end apparatus. One possibility is that their fate is
determined kinetically via a take the money and run strategy. If
the RNA is not exported fast enough from the nucleus, it is
targeted to a default exosome pathway for degradation or
processing. If so, the presence of Pab2p on the poly(A) tail might
not have a function in the nucleus other than that of a trigger
that would determine the threshold above which the nuclear
residency time is not compatible with the life of an mRNA.