Embed Size (px)
Citation preview

Adrenergic Receptor Control Mechanism
for Growth Hormone Secretion
Wn-LwMG. BLACKARDand SYLVIA A. HEIDINGSFELDER
From the Department of Medicine, Louisiana State University School of Medicine,New Orleans, Louisiana 70112
A B S T R A C T The influence of catecholamines ongrowth hormone secretion has been difficult toestablish previously, possibly because of the sup-pressive effect of the induced hyperglycemia ongrowth hormone concentrations. In this study, anadrenergic receptor control mechanism for humangrowth hormone (HGH) secretion was uncoveredby studying the effects of alpha and beta receptorblockade on insulin-induced growth hormone ele-vations in volunteer subjects.
Alpha adrenergic blockade with phentolamineduring insulin hypoglycemia, 0.1 U/kg, inhibitedgrowth hormon elevations to 30-50% of values inthe same subjects during insulin hypoglycemiawithout adrenergic blockade. More complete inhi-bition by phentolamine could not be demonstratedat a lower dose of insulin (0.05 U/kg). Betaadrenergic blockade with propranolol during insu-lin hypoglycemia significantly enhanced HGHcon-centrations in paired experiments. The inhibitingeffect of alpha adrenergic receptor blockade onHGHconcentrations could not be attributed todifferences in blood glucose or free fatty acidvalues; however, more prolonged hypoglycemiaand lower plasma free fatty acid values may havebeen a factor in the greater HGHconcentrationsobserved during beta blockade. In the absence ofinsulin induced hypoglycemia, neither alpha norbeta adrenergic receptor blockade had a detectable
This paper was presented at the Southern Society ofClinical Investigation meetings 27 January 1968. Clin.Res. 1968. 16: 74. (Abstr.)
Address requests for reprints to Dr. William G. Black-ard, Department of Medicine, Louisiana State UniversitySchool of Medicine, New Orleans, La. 70112.
Received for publication 22 December 1967 and in re-vised form 15 February 1968.
effect on HGHconcentrations. Theophylline, aninhibitor of cyclic 3'5'-AMP phosphodiesterase ac-tivity, also failed to alter plasma HGHconcen-trations.
These studies demonstrate a stimulatory effectof alpha receptors and a possible inhibitory effectof beta receptors on growth hormone secretion.
INTRODUCTION
Many of the stimuli for growth hormone secretionsuch as physical and surgical stress (1, 2), psychicstress (2), exercise (2), 2-deoxyglucose (1),hypoglycemia (1), histamine (3), vasopressin (4),and pyrogen (5) result in increased catecholamineconcentrations in blood and tissues (2, 6-10). Thecontrol of pituitary growth hormone secretion bythe hypothalamus is now widely accepted andrecent evidence has shown that at least one of theabove mentioned stimuli caused a discharge ofgrowth hormone-releasing factor from the hypo-thalamus (11). As the catecholamine content ofthe hypothalamus is higher than that of any othercentral nervous system structure (12), the possi-bility that the effect of these stimuli on growthhormone secretion may be mediated by catechola-mines must be considered.
The role of epinephrine in the control of growthhormone secretion has been controversial. Twogroups of investigators (13, 14) have detected astimulatory effect of epinephrine on growth hor-mone secretion while other investigators havebeen unable to demonstrate elevated growth hor-mone concentrations after epinephrine administra-tion (2, 6, 15, 16). Perhaps epinephrine-inducedhyperglycemia masked a stimulatory effect of thehormone on growth hormone secretion in the
The Journal of Clinical Investigation Volume 47 1968 1407

negative studies since hyperglycemia suppressesplasma growth hormone concentrations (1).
The present investigation was performed (a) todetermine the effect of catecholamines on growthhormone secretion under conditions in whichhyperglycemia could not mask a stimulatory effect,(b) to characterize any observed effect in termsof alpha and beta adrenergic receptors, and (c) toassess the role of endogenous catecholamines onthe plasma growth hormone response to insulinhypoglycemia. Administration of alpha and betaadrenergic blocking agents during insulin hypo-glycemia to determine their effect on plasmahuman growth hormone (HGH) elevations proveda suitable experimental design for these purposes.
METHODS21 apparently healthy young male volunteers, 18-35 yrold, were selected for this study. Volunteers were ex-cluded if they were obese (10%o or more above idealweight) or if they had a family history of diabetes. Twointravenous insulin tolerance tests were performed on themajority of the subjects: a control test in which glu-cagon-free insulin was given alone and another test inwhich insulin was given while the patient was receivingeither an alpha or a beta adrenergic blocking agent.Several subjects had three insulin tolerance tests, a con-trol test and one with each of the adrenergic blockingagents, and some subjects received the adrenergic block-ing agents without insulin. The series of tests on eachsubject was performed in random order and was com-pleted within a 10 day period. With few exceptions, atleast 1 day intervened between tests. A total of 52 studieswas performed on these 21 individuals.
The subjects were instructed to abstain from smokingand to take nothing by mouth except water at 10 p.m. thenight before the study. Each subject reported to the roomused for clinical testing at 7 a.m. After the subject hadbeen weighed, he was put to bed and a slow intravenousinfusion of 0.85% NaCl was begun through a 20 gaugeneedle in an antecubital vein. The indwelling needle wasused for withdrawal of blood samples as well as for in-jections of insulin and drugs. The needle was kept openby the saline infusion and no anticoagulants were given tothe subject or were present in the sampling syringes. Asphygmomanometer was placed on the opposite arm tomonitor blood pressure. Blood pressure and pulse ratewere taken every 15 min throughout the experiment.
After a 30 min base line period, glucagon-free insulin(0.05 or 0.1 U/kg) was injected. With the exception ofthe first 15 min interval after insulin injection, bloodsamples were taken every 15 min during the control pe-riod and for 90 min after the insulin had been given.In the alpha adrenergic blockade experiments, phentola-mine was infused at a rate of 0.5 mg/min by a Sage con-stant infusion pump beginning immediately before insulin
administration and continuing throughout the remaining90 min of the experiment. In the beta adrenergic blockadecxperiments, a "stat" dose of propranolol, 3 mg, was givenintravenously and a constant infusion of propranolol at arate of 0.08 mg/min was started immediately before insu-lin administration and was continued until the end of theexperiment. Propranolol was not given to any subj ectwith a resting pulse below 60 beats/min. On this basis,three subjects for whompropranolol experiments had beenplanned were rejected for that part of the study. In 11studies (4 with alpha blockade and 7 with beta blockade)adrenergic blocking agents were given as describedabove, but without insulin. In four subjects, 500 mg oftheophylline ethylenediamine was infused at a constantrate for 15 min at the end of the base line period. Bloodsamples were taken in all studies at the same intervalsas in the experiments in which insulin was given.
Blood samples were collected in chilled heparinizedtubes. Protein-free filtrates for blood glucose were madefrom 0.2 ml of blood immediately after the blood hadbeen obtained. Blood glucose was determined by a glu-cose oxidase method (17). Plasma free fatty acid (FFA)was measured by a modification of the Dole procedureon 1-ml aliquots of plasma (18).
Plasma HGHwas determined by a modification of theradioimmunoassay method of Schalch and Parker (19)utilizing 'I-labeled growth hormone and separating freefrom bound hormone by the double antibody technique.Tracer and standards were prepared from highly puri-fied HGH (Lot HS 968C) supplied to us by Dr. A. E.Wilhelmi. HGHantiserum was generously supplied bythe National Institute of Arthritis and Metabolic Diseasesand the National Pituitary Agency.
Propranolol (Inderal) was kindly supplied by AyerstLaboratories, New York. Phentolamine (Regitine) andtheophylline ethylenediamine (Aminophylline) were pur-chased from CIBA Pharmaceutical Products, Inc., Sum-
60-
N 50-
40-
t30
Z 20-
10-
- Insulin alone.......Insulin+ Phentolamine
5 subjects (paired data)
Insulin (0.1 U/Kg )
h ...I....
-30 -15 0 15 30Minutes
45 60 75 90
FIGURE 1 Effect of alpha adrenergic blockade on plasmahuman growth hormone (HGH) concentrations duringinsulin-induced hypoglycemia, 0.1 U/kg. Alpha adrenergicblockade was produced by phentolamine infusion (0.5mg/min) begun at the time of insulin administration andcontinued throughout the remainder of the experiment.Means ± sEM are shown.
1408 W. G. Blackard and S. A. Heidingsfelder

mit, N. J. and the G. D. Searle & Co., Skokie, Ill., re-spectively.
RESULTS
Alpha adre'nergic blockade during insulin hypo-glycemia. The data shown in Fig. 1 demonstratean inhibitory effect of alpha adrenergic blockadeon plasma HGH concentrations during insulinhypoglycemia. Base line plasma HGHconcentra-tions during the 30 min base line period wereslightly higher in these and subsequent studies(3.1 + .2 m ug/ml) than those usually reportedfor overnight fasting male subjects (1). Our sub-jects were ambulatory before testing which maypartially account for the higher plasma HGHcon-centrations. Glucagon-free insulin administration,0.1 U/kg, resulted in peak plasma HGHconcen-trations at 45-75 min after injection. In the samesubjects during a phentolamine infusion, 0.5 mg/min, plasma HGHresponse to hypoglycemia wasonly one-third to one-half as great as in the ex-periment with insulin alone. Plasma HGHvalueswere significantly different at the 45 and 60 minintervals with P values < 0.01 and < 0,02, respec-tively (paired t test). Blood glucose and plasmaFFA data for these experiments are expressed asper cent of pretest values (Fig. 2). Hypoglycemia
20- Insulin alone (0.1 U/Kg)....... Insulin+Phentolamine
5 subjects (paired data)
0 30 60 900 30 60 90Minutes After Insulin
FIGURE 2 Effect of alpha adrenergic blockade on bloodglucose and plasma FFA during insulin-induced hypo-glycemia, 0.1 U/kg. Mean pretest blood glucose valueswere 74 + 5.3 mg/100 ml and 77 ± 1.5 mg/100 ml in theinsulin alone and alpha adrenergic blockade experiments,respectively. Mean pretest plasma FFA values were 690 +50 AuEq/liter and 780 ± 60 AEq/liter in the insulin aloneand alpha adrenergic blockade experiments, respectively.Means ± SEMare shown.
60-
50-
z 40-
t 30-
t,,
Z@ 20-
10*
- Insulin alone...... Insulin + Phentolamine
5 subjects (paired data)
Insulin (0.05 U/Kg)
P'is .
-30 -15 0Minutes
30 45 60 75 90
FIGURE 3 Effect of alpha adrenergic blockade on plasmaHGH concentrations during insulin-induced hypoglyce-mia, 0.05 U/kg. Alpha adrenergic blockade produced asstated in Fig. 1. Means + SEM are shown.
was slightly more prolonged in the experimentswith insulin alone but analysis of individual sub-ject responses indicated that the greater plasmaHGHelevation with insulin alone could not beattributed to this small difference. The plasmaFFA difference 45 min after insulin injection cannot be explained. As phentolamine alone wasshown to lower plasma FFA in three of foursubjects (Fig. 9) some enhancement of insulin-
20- - Insulin alone (0.05 U/Kg)*------- Insulin+ Phentolamine
5 subjects (paired data)
0 30 60 90 0 30 60 90Minutes After Insulin
FIGURE 4 Effect of alpha adrenergic blockade on bloodglucose and plasma FFA during insulin-induced hypogly-cemia, 0.05 U/kg. Mean pretest blood glucose values were
79 ± 1.7 mg/1O0 ml and 77 ± 2.6 mg/100 ml in the insu-lin alone and alpha adrenergic blockade experiments, re-
spectively. Mean pretest plasma FFA values were 607 +
60 /LEq/liter and 632 + 32 ;LEq/liter in the insulin aloneand alpha adrenergic blockade experiments, respectively.Means ± SEM are shown.
Adrenergic Receptor Control Mechanism for Growth Hormone Secretion 1409

80-
70-
60-
50*
(40-
t3.
": 30-a.,
20-
10-
- Insulin alone........ Insulin + Propranolol
6 subjects (paired data)
Insulin (0.05 U/Kg)
jl2 ............IL .....................
-30 -15 0Minutes
FIGuRE 5 Effect of beta adrenHGH concentrations during itmia, 0.05 U/kg. Beta adrenergiby a stat injection of 3 mg prcfore insulin administration folkpranolol infusion of 0.08 mg/ithe experiment. Means ± SEMar
induced FFA depressionhave occurred.
Similar alpha adrenergicwere performed on five addhypoglycemia produced byinsulin (0.05 U/kg) to deter
1201 Blood Glucose
inhibition of the HGHresponse could be achieved.The smaller dose of insulin resulted in a lesserplasma HGH response (Fig. 3) and althoughcomplete inhibition was not achieved in any sub-ject, phentolamine again depressed the growthhormone response. Plasma HGHvalues were sig-nificantly different at 45 and 60 min with P values< 0.05 (paired t test). Blood glucose and plasma
I ...+L'FFAvalues for these experiments are shown inFig. 4. As blood glucose concentrations with andwithout phentolamine were nearly identical, thepossibility that differences in blood glucose couldaccount for the inhibition of HGHresponse byphentolamine was eliminated. Plasma FFA in
30 45 60 75 90 experiments with insulin alone and those withinsulin and phentolamine reached the same nadir
ergic blockd. onlasma lbut a more rapid and higher rebound in plasmaisulin-induced hypoglyce-.ic blockade was produced FFA occurred in the latter experiments. Because)pranolol immediately be- of the failure of the lower dose of insulin (0.05owed by a constant pro- U/kg) to elicit a HGHresponse in 3 of 10 volun-min until termination of teers, the effect of alpha adrenergic blockade on-e shown. HGHresponse at an even lower insulin dose was
not studied.Beta adrenergic blockade during insulin hypo-
glycemia. Paired experiments in which six vol-.blocad .ex rimn unteers received insulin, 0.05 U/kg, with andial subjects dourngofwithout beta adrenergic blockade showed moderate
enhancement of plasma HGHconcentrations dur-rmine if more complete ing beta adrenergic blockade with propranolol
Plasma FFA (Fig. 5). The sum of plasma HGHconcentrationsat 60, 75, and 90 min after insulin injection was
:80 ...
60 *...I> 40-
. 20- Insulin alone (0.05 U/Kg)*- Insulin + Propranolol
6 subjects (paired data)
0 30 60 90 0 30 60 90Minutes After Insulin
FIGURE 6 Effect of beta adrenergic blockade on bloodglucose and plasma FFA during insulin-induced hypo-glycemia, 0.05 U/kg. Mean pretest blood glucose valueswere 84 +.7 mg/100 ml and 80 + 1.8 mg/100 ml in theinsulin alone and alpha adrenergic blockade experiments,respectively. Mean pretest plasma FFA values were670 ± 88 4Eq/liter and 557 + 64 ,tEq/liter in the insulinalone and alpha adrenergic blockade experiments. Means+ SEMare shown.
q 300-
,! 250-
XtIZ
200-;::% 150-
.,Ct
%100-
' 50-
_0.q0
CZ
T5 Insulin (0.10 U /Kg)+PhentolamineEl Insulin (0.05 U /Kg)+ Phentolomine* Insulin(0.05 U/Kg)+Propranol~ol
30 45 60 75 90Minutes After Insulin Injection
105
FIGURE 7 Effect of alpha and beta adrenergic blockadeon plasma HGH response to insulin hypoglycemia.Means ± SEMare shown.
1410 W. G. Blackard and S. A. Heidingsfelder
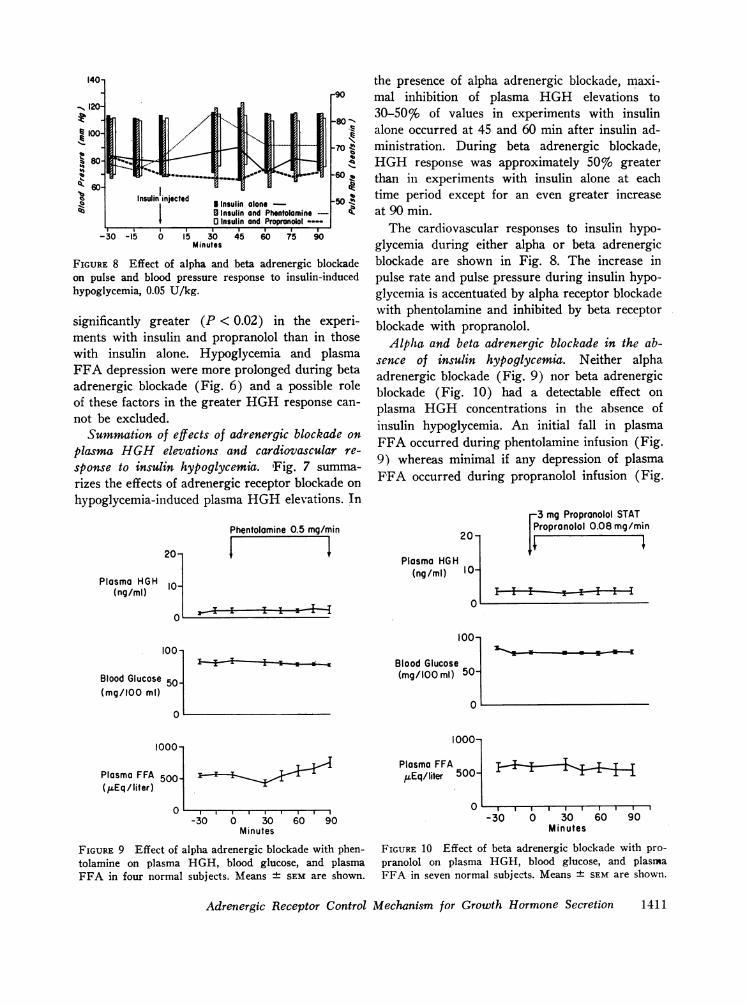
FIGURE 8 Effect of alpha and beta adrenergic blockadeon pulse and blood pressure response to insulin-inducedhypoglycemia, 0.05 U/kg.
significantly greater (P < 0.02) in the experi-ments with insulin and propranolol than in thosewith insulin alone. Hypoglycemia and plasmaFFA depression were more prolonged during betaadrenergic blockade (Fig. 6) and a possible roleof these factors in the greater HGHresponse can-not be excluded.
Summation of effects of adrenergic blockade onplasma HGHelevations and cardiovascular re-sponse to insulin hypoglycemia. Fig. 7 summa-rizes the effects of adrenergic receptor blockade onhypoglycemia-induced plasma HGHelevations. In
Phentolamine 0.5 mg/min
l r lI
20-
Plasma HGH 10-(ng/ml)
the presence of alpha adrenergic blockade, maxi-mal inhibition of plasma HGH elevations to30-50% of values in experiments with insulinalone occurred at 45 and 60 min after insulin ad-ministration. During beta adrenergic blockade,HGHresponse was approximately 50% greaterthan in experiments with insulin alone at eachtime period except for an even greater increaseat 90 min.
The cardiovascular responses to insulin hypo-glycemia during either alpha or beta adrenergicblockade are shown in Fig. 8. The increase inpulse rate and pulse pressure during insulin hypo-glycemia is accentuated by alpha receptor blockadewith phentolamine and inhibited by beta receptorblockade with propranolol.
Alpha and beta adrenergic blockade in the ab-sence of insulin hypoglycemia. Neither alphaadrenergic blockade (Fig. 9) nor beta adrenergicblockade (Fig. 10) had a detectable effect onplasma HGH concentrations in the absence ofinsulin hypoglycemia. An initial fall in plasmaFFA occurred during phentolamine infusion (Fig.9) whereas minimal if any depression of plasmaFFA occurred during propranolol infusion (Fig.
-3 mg Propronolol STATPropronolol 0.08 mg/min
.I; fIPlasma HGH
100-
Blood Glucose 50-(mg/100 ml)
10001
Plasma FFA 500b(kLEq / liter)
0
II iii
100-
Blood Glucose(mg/100 ml) 50-
0
1000-
-30 0 30 60 90Minutes
FIGURE 9 Effect of alpha adrenergic blockade with phen-tolamine on plasma HGH, blood glucose, and plasmaFFA in four normal subjects. Means ± SEM are shown.
Plasma FFA IrI 1 1,uEq/liter 5000i 1
500-30 0 30 60 90
Minutes
FIGURE 10 Effect of beta adrenergic blockade with pro-pranolol on plasma HGH, blood glucose, and plasmaFFA in seven normal subjects. Means + SEM are shown.
Adrenergic Receptor Control Mechanism for Growth Hormone Secretion 1411
20-
Plasma HGHH(ng/mI)
0
100-
Blood Glucose(mg/100 ml)
0
1000-
Plasma FFA 500-(pEq/liter)
OL-30
500 mg Theby constantn
0 30Minutes
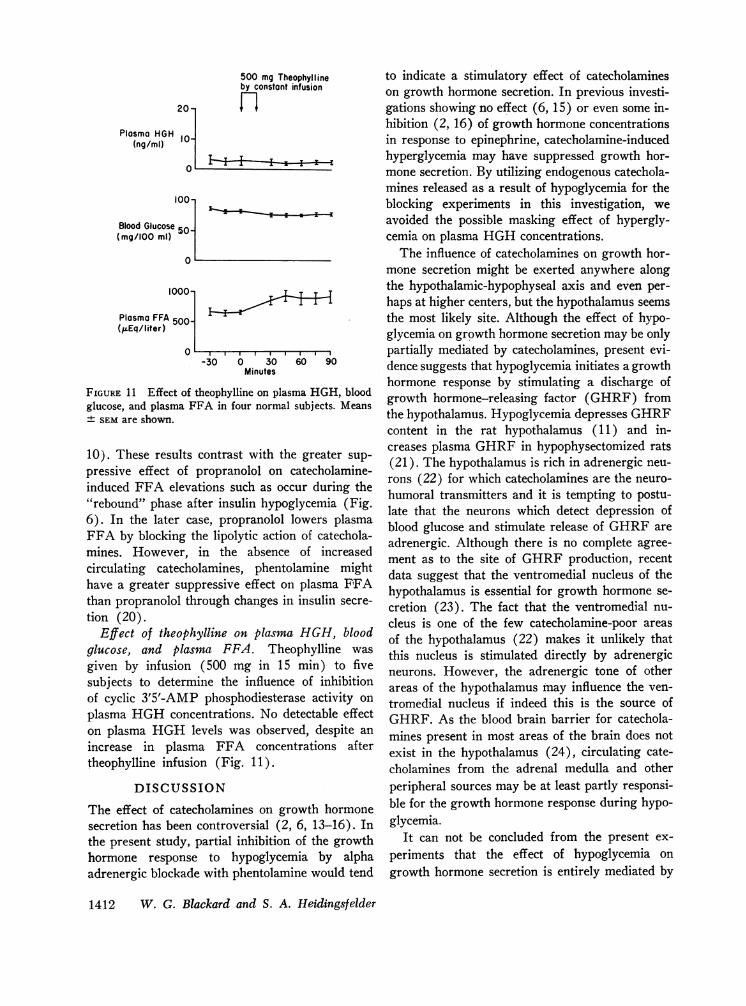
FIGURE 11 Effect of theophylline on plasglucose, and plasma FFA in four normal± SEMare shown.
10). These results contrast with thepressive effect of propranolol on (
induced FFA elevations such as oc("rebound" phase after insulin hypog6). In the later case, propranololFFA by blocking the lipolytic actioimines. However, in the absencecirculating catecholamines, phentohave a greater suppressive effect orthan propranolol through changes intion (20).
Effect of theophylline on plasmaglucose, and plasma FFA. The(given by infusion (500 mg in 15subjects to determine the influenceof cyclic 3'5'-AMP phosphodiesteraplasma HGHconcentrations. No doon plasma HGHlevels was observincrease in plasma FFA concentheophylline infusion (Fig. 11).
DISCUSSION
The effect of catecholamines on grsecretion has been controversial (2,the present study, partial inhibitionhormone response to hypoglyceradrenergic blockade with phentolam
?ophylline to indicate a stimulatory effect of catecholaminesinfusion on growth hormone secretion. In previous investi-
gations showing no effect (6, 15) or even some in-hibition (2, 16) of growth hormone concentrationsin response to epinephrine, catecholamine-inducedhyperglycemia may have suppressed growth hor-mone secretion. By utilizing endogenous catechola-mines released as a result of hypoglycemia for theblocking experiments in this investigation, weavoided the possible masking effect of hypergly-cemia on plasma HGHconcentrations.
The influence of catecholamines on growth hor-mone secretion might be exerted anywhere alongthe hypothalamic-hypophyseal axis and even per-haps at higher centers, but the hypothalamus seemsthe most likely site. Although the effect of hypo-glycemia on growth hormone secretion may be only
,O , , partially mediated by catecholamines, present evi-60 90 dence suggests that hypoglycemia initiates a growth
hormone response by stimulating a discharge ofma HGH, blood growth hormone-releasing factor (GHRF) fromsubjects. Means
the hypothalamus. Hypoglycemia depresses GHRFcontent in the rat hypothalamus (11) and in-creases plasma GHRFin hypophysectomized rats
catecholamine- (21). The hypothalamus is rich in adrenergic neu-
cure during te rons (22) for which catecholamines are the neuro-ur during thie humoral transmitters and it is tempting to postu-
glycemia (Fig. late that the neurons which detect depression oflowers plasma blood glucose and stimulate release of GHRFaren of catechola-
increased adrenergic. Although there is no complete agree-of ight ment as to the site of GHRFproduction, recent
lamne miFFht data suggest that the ventromedial nucleus of the
pinsulansmaFFA hypothalamus is essential for growth hormone se-insulin secre- cretion (23). The fact that the ventromedial nu-
HGH blood cleus is one of the few catecholamine-poor areas
Dphyllin was of the hypothalamus (22) makes it unlikely thatophyllmne was this nucleus is stimulated directly by adrenergic
min) to five neurons. However, the adrenergic tone of othere of inhibition areas of the hypothalamus may influence the ven-Ese activity on tromedial nucleus if indeed this is the source ofetectable effect GHRF. As the blood brain barrier for catechola-ed, despite an mines present in most areas of the brain does notitrations after exist in the hypothalamus (24), circulating cate-
cholamines from the adrenal medulla and otherperipheral sources may be at least partly responsi-
owth hormone ble for the growth hormone response during hypo-6, 13-16). In glycemia.of the growth It can not be concluded from the present ex-
nia by alpha periments that the effect of hypoglycemia on
ine would tend growth hormone secretion is entirely mediated by
1412 W. G. Blackard and S. A. Heidingsfelder
r

catecholamines since complete inhibition of HGHresponse could not be achieved with phentolamine.The growth hormone secretory mechanism is ap-parently sensitive to changes in blood glucose andnot just to catecholamines released by hypoglyce-mia since elevations of blood glucose suppressHGHsecretion (1). Catecholamines may merelymodify the stimulatory effect of hypoglycemia onHGHrelease. Investigations by Sutin have shownan effect of insulin hypoglycemia on the evokedelectrical potential recorded from a probe in theventromedial nucleus of the hypothalamus afterstimulation of the amygdala and septal nuclei (25).These changes in electrical potential from the ven-tromedial nucleus could be reproduced by injectionof norepinephrine into the nucleus (26). Norepi-nephrine modified the amplitude of electrical re-sponse in the ventromedial nucleus to stimuli fromother nuclei but did not initiate a response.
The stimulatory effect of catecholamines ongrowth hormone secretion has important impli-cations as practically all of the recognized stimulifor growth hormone secretion, with the exceptionof amino acid infusion and estrogens, invoke a cate-cholamine response. Even small decreases in bloodglucose (less than 10 mg/100 ml) which havebeen reported to elevate plasma HGHare accom-panied by increased urinary catecholamines (6).In addition, the sympathetic tone in the hypothala-mus may be important in the secretory regulationof other pituitary hormones. Hypothalamic cate-cholamine-depleting agents such as reserpine havebeen shown to influence gonadotropin secretion(27) as well as to inhibit the growth hormone re-sponse to hypoglycemia (28). Vasopressin secre-tion also is probably modified by adrenergic tonein the hypothalamus (29).
The present study characterized the growthhormone response to catecholamines in terms ofalpha and beta adrenergic receptors. During in-sulin hypoglycemia the inhibitory effect of alphablockade on growth hormone secretion was pro-nounced while the stimulatory effect of beta block-ade was less impressive and was associated withmore prolonged hypoglycemia and lower plasmaFFA concentrations. More prolonged hypogly-cemia probably was not responsible for the en-hanced HGHresponse during beta blockade asstudies in monkeys during insulin hypoglycemiahave shown that growth hormone hypersecretion
occurs while blood glucose concentrations are fall-ing but return to normal even though plasma glu-cose concentrations remain depressed (30). Lim-ited data on the effect of plasma FFA on growthhormone levels are available. Schalch and Kipniswere unable to demonstrate an effect of plasmaFFA elevations on plasma HGHconcentrationsin three subjects given a fat meal followed byheparin (31). However, preliminary data fromDr. K. Shizume in Tokyo (personal communica-tion) have suggested that elevations of plasmagrowth hormone concentrations by nicotinic acidmay be related to the fall in plasma FFA. Databy Abramson, Arky, and Woeber, which showeda modest increase in hypoglycemia-induced plasmaHGHelevations during beta adrenergic blockade,support our findings (32).
The present studies demonstrate a definite stim-ulatory effect of alpha adrenergic receptors and apossible inhibitory effect of beta adrenergic re-ceptors on growth hormone secretion. This patternis the opposite of the adrenergic control mechanismfor insulin secretion where it has been shown thatalpha receptors inhibit (33) and beta receptorsstimulate (34) insulin release. Since the predomi-nant effect of catecholamines on hormonal regu-lation appear to be through alpha receptors,another remarkable homeostatic mechanism tomaintain the constancy of the internal milieu is ob-served. During hypoglycemia, catecholamines in-hibit insulin secretion and assist in promoting arise in plasma HGHwhich antagonizes the actionof insulin and also increases plasma FFA whichmay be used for energy requirements.
The lack of effect of adrenergic blockade ortheophylline, an inhibitor of cyclic 3'5'-AMP phos-phodiesterase activity, on plasma HGHconcen-trations in the absence of hypoglycemia probablynegates a significant role of the adrenergic nervoussystem in control of resting plasma HGHlevels.
ACKNOWLEDGMENTSWe would like to acknowledge the capable technical as-sistance of Miss Thelma Cameron.
This paper was supported by grant AM-10151-01 fromthe National Institutes of Health.
REFERENCES
1. Roth, J., S. M. Glick, P. Cuatrecasas, and C. S.Hollander. 1967. Acromegaly and other disorders ofgrowth hormone secretion. Ann. Internal Med. 66:760.
Adrenergic Receptor Control Mechanism for Growth Hormone Secretion 1413

2. Schalch, D. S. 1967. The influence of physical stressand exercise on growth hormone and insulin secretionin man. J. Lab. Clin. Med. 69: 256.
3. Meyer, V., and E. Knobil. 1966. Stimulation of growthhormone secretion by vasopressin in the Rhesusmonkey. Endocrinology. 79: 1016.
4. Gagliardino, J. J., J. D. Bailey, and J. M. Martin.1967. Effect of vasopressin on serum-levels of humangrowth hormone. Lancet. 1: 1357.
5. Kohler, P. O., B. W. O'Malley, P. L. Rayford, M. B.Lipsett, and W. D. Odell. 1967. Effect of pyrogen onblood levels of pituitary trophic hormones. Observa-tions on the usefulness of the growth hormone re-sponse in the detection of pituitary disease. J. Clin.Endocrinol. Metab. 27: 219.
6. Luft, R., E. Cerasi, L. L. Madison, U. S. von Euler,L. D. Casa, and A. Roovete. 1966. Effect of a smalldecrease in blood-glucose on plasma-growth-hormoneand urinary excretion of catecholamines in man.Lancet. 2: 254.
7. Statt, W. H., and M. B. Chenoweth. 1966. Release ofcatecholamines by vasopressin into the cavernoussinus blood of the dog. Proc. Soc. Exptl. Biol. Med.123: 785.
8. Sanford, J. P., J. A. Barnett, and C. Gott. 1960. Amechanism of the glycogenolytic action of bacterialendotoxin. J. Exptl. Med. 112: 97.
9. Laszlo, J., W. R. Harlan, R. F. Klein, N. Kirshner,E. H. Estes, Jr., and M. D. Bogdonoff. 1961. Theeffect of 2-deoxy-D-glucose infusions on lipid andcarbohydrate metabolism in man. J. Clin. Invest. 40:171.
10. Kellaway, C. H., and S. J. Cowell. 1922. On theconcentration of the blood and the effects of hista-mine in adrenal insufficiency. J. Physiol. 57: 82.
11. Katz, S. H., A. P. S. Dhariwal, and S. M. McCann.1967. Effect of hypoglycemia on the content of pitui-tary growth hormone (GH) and hypothalamic growthhormone-releasing factor (GHRF) in the rat. Endo-crinology. 81: 333.
12. Carlsson, A. 1959. The occurrence, distribution, andphysiological role of catecholamines in the nervoussystem. Pharmacol. Rev. 11: 490.
13. Gagliardino, J. J., and J. M. Martin. 1967. Mechanismof growth hormone secretion. In Program of the 49thMeeting. The Endocrine Society. 74. (Abstr.)
14. Meyer, V., and E. Knobil. 1966. The acute elevationof plasma growth hormone concentration in responseto various stimuli. Federation Proc. 25: 379. (Abstr.)
15. Roth, J., S. M. Glick, R. S. Yalow, and S. A. Berson.1963. Hypoglycemia: a potent stimulus to secretion ofgrowth hormone. Science. 140: 987.
16. Rabinowitz, D., T. J. Merimee, J. A. Burgess, andL. Riggs. 1966. Growth hormone and insulin releaseafter arginine: indifference to hyperglycemia andepinephrine. J. Clin. Endocrinol. Metab. 26: 1170.
17. Saifer, A., and S. Gerstenfeld. 1958. The photo-metric microdetermination of blood glucose withglucose oxidase. J. Lab. Clin. Med. 51: 448.
18. Trout, D. L., E. H. Estes, Jr., and S. J. Friedberg.1960. Titration of free fatty acids of plasma: a studyof current methods and a new modification. J. LipidRes. 1: 199.
19. Schalch, D. S., and M. L. Parker. 1964. A sensitivedouble antibody immunoassay for human growth hor-mone in plasma. Nature. 203: 1141.
20. Frohman, L. A., E. Z. Ezdinli, and R. Javid. 1967.Effect of vagotomy and vagal stimulation on insulinsecretion. Diabetes. 16: 443.
21. Krulich, L., and S. M. McCann. 1966. Evidence forthe presence of growth hormone-releasing factor inblood of hypoglycemic hypophysectomized rats. Proc.Soc. Exptl. Biol. Med. 122: 668.
22. Shute, C. C. D., and P. R. Lewis. 1966. Cholinergicand monoaminergic pathways in the hypothalamus.Brit. Med. Bull. 22: 221.
23. Bernardis, L. L., and L. A. Frohman. 1967. Plasmaand pituitary growth hormone levels in rats withhypothalamic lesions. Program of the 49th Meeting.The Endocrine Society. 86. (Abstr.)
24. Weil-Malherbe, H. 1960. The passage of catecholamines through the blood-brain barrier. Adrenergicmechanisms. CIBA Foundation Symposium. J. R.Vane, G. E. W. Wolstenholme, and M. O'Connor,editors. Little, Brown and Co., Boston. 421.
25. Sutin, J. 1963. An electrophysiological study of thehypothalamic ventromedial nucleus in the cat. Electro-encephalog. Clin. Neurophysiol. 15: 786.
26. van Orden, L. S., and J. Sutin. 1963. Differentialeffects of norepinephrine on responses evoked in thehypothalamic ventromedial nucleus. Electroencephalog.Clin. Neurophysiol. 15: 796.
27. Lippmann, W., R. Leonardi, J. Ball, and J. A. Cop-pola. 1967. Relationship between hypothalamic cate-cholamines and gonadotrophin secretion in rats. J.Pharmacol. Exptl. Therap. 156: 258.
28. Muller, E. E., S. Sawano, A. Arimura, and A. V.Schally. 1967. Blockade of release of growth hormoneby brain norepinephrine depletors. Endocrinology. 80:471.
29. Rothballer, A. B. 1959. The effects of catecholamineson the central nervous system. Pharmacol. Rev. 11:494.
30. Abrams, R. L., M. L. Parker, S. Blanco, S. Reichlin,and W. H. Daughaday. 1966. Hypothalamic regulationof growth hormone secretion. Endocrinology. 78: 605.
31. Schalch, D. S., and D. M. Kipnis. 1965. Abnormalitiesin carbohydrate tolerance associated with elevatedplasma nonesterified fatty acids. J. Clin. Invest. 44:2010.
32. Abramson, E. A., R. A. Arky, and K. A. Woeber.1966. Effects of propranolol on the hormonal andmetabolic responses to insulin-induced hypoglycaemia.Lancet. 2: 1386.
33. Porte, D., Jr. 1967. A receptor mechanism for theinhibition of insulin release by epinephrine in man.J. Clin. Invest. 46: 86.
34. Porte, D., Jr. 1967. Beta adrenergic stimulation ofinsulin release in man. Diabetes. 16: 150.
1414 W. G. Blackard and S. A. Heidingsfelder





























