Embed Size (px)
Citation preview

Adsorption of enzymes at the solid-liquid interface I. Try-psin on polystyrene latex
D. Lewis and T.L. Whaieley Department of Pharmacy, University of Strathclyde, Glasgow G 7 IXW, UK
Presented at Biointeractions ‘87, Cambridge, UK in July 1987
The enzyme, trypsin, has been used to study conformational changes which occur when protein adsorption onto well-characterized, emulsifier-free, polystyrene latex surface takes place. The adsorption isotherm is of the high affinity, langmuirian type with plateau adsorption of trypsin of 2.8 mg m-‘. The enzymic activity of adsorbed trypsin to low molecular weight substrate is found to decrease as the surface coverage decreases indicating that ‘spreading’ or unfolding of the native protein conformation, with consequent loss of enzymic activity, occurs. On the close packed surface such ‘spreading’ is inhibited by steric factors. The view that protein adsorption onto hydrophobic surfaces is dominated by the entropy gain due to protein unfolding to maximize hydrophobic interactions is thus supported.
Keywords: Protein adsorption, trypsin, polystyrene latex. enzyme adsorption, protein conformation
The adsorption of proteins at the solid-liquid interface is of importance in many areas (e.g. biocompatibilityof prostheses and implants; in viva fate of injected foreign colloidal particles used for drug delivery and targeting; immuno- diagnostic methods). Additionally, enzyme adsorption is of interest in immobilized enzyme technology, the use of enzymes as recoverable biocatalysts and in biosensor devices as well as fundamentally in the effects surface and membrane binding have on enzyme activity in vim.
It is particularly difficult to obtain information regarding the conformation and orientation of adsorbed proteins: methods for proteins in solution are not generally applicable to adsorbed material at the solid-liquid interface. Desorbed material may be studied but its relationship to the adsorbed protein is uncertain. The use of an enzyme to study protein adsorption confers advantages in that an additional parameter (i.e. enzyme activity) which is very sensitive to con- formational changes may be monitored in addition to the normal physico-chemical adsorption measurements. In addition to the enzyme activity of adsorbed material to low molecular weight substrates (giving information on gross conformational changes or denaturation) minor conformational changes may be detected by the susceptibility to proteolysis (or, in the case of a proteolytic enzyme, such as trypsin used in this work, to self-digestion or autolysis). This approach has given information on conformational changes involved in the adsorption of trypsin and a-chymotrypsin onto colloidal silica’,’ where the activity of the adsorbed enzyme to low molecular weight substrate is unchanged from the enzyme
.~_ Correspondence to Dr What&y.
in solution, indicating no significant conformational change on adsorption. However, the rate of autolysis of trypsin was dramatically increased in the presence of colloidal silica, indicating a minor conformational change rendering the adsorbed protein molecule more susceptible to proteolysis.
This approach is now extended to the adsorption of trypsin (and the effects on enzyme activity) onto a well- characterized polystyrene latex surface in an attempt to gain further understanding of the conformational changes that occur when proteins adsorb at the solid-liquid interface. The polystyrene latex is an essentially hydrophobic, charge- stabilized system as opposed to the essentially hydrophilic colloidal silica system.
The adsorption of a number of proteins (especially albumin and ribonuclease) onto a range of substrate surfaces, including polystyrene and silica, has been extensively studied by Norde, Lyklema and co-workers3,4. They have concluded that the process of protein adsorption onto a hydrophobic surface is to a large extent entropy-driven. The patterns for adsorption (i.e. saturation plateau coverage, variation with pH) for a wide range of substrates differing in electrical charge with respect to the protein were all similar. Protein adsorption where protein and surface have the same charge proceeds spontaneously due to the loss of con- formational structure on adsorption with consequent gain in entropy. The larger, more readily deformable, proteins adsorbed more strongly than smaller, compact proteins. The use of an enzyme in such studies allows further information to be gained on conformational changes on adsorption and the effect, for instance, of surface coverage on such processes.
Protein adsorption at polymer-water interfaces has
0 1988 Butterworth Et Co (Publwhers) Ltd. 0142-9612/88/010071-05$03.00
Biomaterials 1988, Vol 9 January 71

Tvpsin adsorption: 0. Lewis and TL. Whateley
recently been extensively reviewed by Andrade’ whilst that of polymers at interfaces has been reviewed by Vincent and Whi~ington6.
Sandwick and Schray’ have recently reported on the inactivation of several enzymes (alkaline phosphatase, B- galactosidase, lysozyme, horseradish peroxidase, catalase and 6-phosphate dehydrogenase) on adsorption to a hydro- phobic polystyrene latex surface. In all cases they found that at low solution concentrations, the enzymes lost catalytic activity. However, without parallel adsorption isotherm data, the surface coverage is not known.
In determining the activity of an enzyme at the solid- liquid interface there are complicating factors such as the effect of substrate on enzyme adsorption, increased rates of proteolysis, effects of diffusional restrictions on enzyme activity measurements and the effects of electrostatic interactions on enzyme, substrate and other ions present. We have attempted to take such factors into account and also determined standard adsorption isotherms for trypsin adsorption onto polystyrene latex surface in order to gain further knowledge of the ~onformational changes occurring on protein adsorption.
MATERIALS AND METHODS
Materials
Trypsin (EC 3.4.2 1.4) bovine pancreas, type Ill was obtained from the Sigma Chemical Company as was N, a-benzoyl+- arginine-ethyl ester (BAEE), benzamidine and /V, a-p-tosyl-L- lysine chloromethylketone (TLCK).
latex preparation
A polystyrene latex was prepared according to the method of Goodwin et a/.’ in an emulsifier-free system purged with nitrogen at a temperature of 80°C.
Styrene (70g) was stirred with the aqueous phase (670 ml) and potassium persulphate added as initiator to give a final concentration of 4.4 X 1 Op3 NI on an aqueous phase volume of 720 ml. The total reaction iime was 24 hours.
These conditions were chosen in accordance with the trends noted by Goodwin et a/.’ in order to minimize particle size (i.e. low ionic strength, high initiator concentration, high temperature) and hence maximize the specific surface area of the latex.
The product was dialysed and ion-exchanged twice according to the method of Van den Hul and Vanderhoff’ and the solids content determined after drying samples at 50°C.
Particle size dist~bution
Transmission electron micrographs were obtained using a Phillips EM 400T electron microscope using an accelerating voltage of 60 kV and a dwell time of less than 60 s. The magnification from which size measurements were made was 36 000.
Photon correlation spectroscopy (PCS) was performed using a Malvern Instruments system incorporating a K7027 correlator and data analysis was by the method of cumulants”. The source of coherent light was a Liconix 10 mW helium- cadmium laser (A = 441.6 nm) and the Stokes-Einstein equation was used to calculate the apparent size of the particles from the measured diffusion coefficients. The Pusey polydispersity factor was taken as an indicator of
monodispersity. Measurements were made at 90” to the incident light.
Latex surface charge
Surface charge was determined by conductometric titration under nitrogen at 25”C, with sodium hydroxide (0.025 M)
as titrant. Aliquots (5 ml) of the latex were diluted to 40 ml with doubly distilled water. Additions of titrant were made using an Agla (IO r_ll) microsyringe. Conductivity measure- ments were made using a Wayne-Kerr Autobalance AC conductivity bridge. Time was allowed (2 min) between additions to allow the conductivity to reach a steady value.
Adsorption isotherms
Adsorption isotherms for trypsin and trypsinogen were determined at pH 8.0 and 25°C in the presence of the competitive inhibitor benzamidine (5 mM sodium barbitone, 1 mM benzamide) to prevent autolytic breakdown of the protein. A quantity of latex (e.g. 1 .O ml, 67.8 mg solids) was rapidly added to a solution of the enzyme in a poly~arbonate centrifuge tube and mixed by inversion (twice) before incubation in a water bath at 25°C. The tubes were centrifuged (25 000 g) for 30 min and the supernatants filtered through Durapore hydrophilic filters (pore size 0.45 pm, diameter 13 mm) to remove residual latex particles which in some cases became redispersed from the side of the tube by the falling liquid level during removal. During the filtering of standard protein solutions these filters were shown to absorb less than 0.01 mg of trypsin which was typically 0.5% of the total adsorbed amount in a typical adsorption experiment. Nevertheless the first 2 ml of the filtrate was discarded to ensure a negligible error in the final concentration.
Protein concentrations were determined by the method of Lowry” or that of ltzakhi and Gill” with reference to standard trypsin solutions made up by weight.
No variation in trypsin adsorption to the latex particles was found for contact times before centrifugation of 15 min to 3 h at a final solution protein concentration of 0.1 mg ml-‘. Contact time for both trypsin and ttypsinogen was standardized at 60 min.
Enzyme activity measurements
The activity of trypsin was routinely assayed in the presence and absence of polystyrene latex in a pH-stat vessel (Radiometer Copenhagen) maintained at 25°C + 0.1 “C. Initially this contained the substrate solution (10 ml 5 mM
BAEE, 1 mM sodium barbitone) which was titrated after addition of the enzyme with sodium hydroxide (0.025 M) at pH 8.0. Nitrogen pre-heated to 25°C and saturated with watervapour was blown overthe surface during the titration. BAEE shows negligible self-hydrolysis at this pH.
Whenever enzyme was incubated in the pH-stat for a period before activity determination, a volume of concentrated substrate solution (1 ml, 20 mM BAEE. 1 mM sodium barbitone pH 7.0) was added to the vessel.
Irreversibly-inhibited enzyme
In order to investigate the effect of the substrate on adsorption of the enzyme without its simultaneous hydrolysis a sample of trypsin was prepared, inhibited with TLCK. The preparation showed a very low level of contamination with a- ~hymot~psin and the presence of pseudot~psin which shows no reaction with TLCK. The residual activity was
72 Biomaterials 1988, Vol9 January
Trypsin adsorption: D. Lewis and 7IL. Whareley
about 1% of the starting material and negligible over the time course of the experiments.
RESULTS AND DISCUSSION
Polystyrene latex cheracterization
Table I summarizes the results obtained on characterization of the latex used in the adsorption experiments. The value presented for the particle diameter is a nominal value taken between the values obtained from PCS and TEM (PCS diameter 525 -f 8 nm with a mean polydispersity 0.105, TEM diameter 480 t 10 nm).
A number of authors have reported electron beam damage to polystyrene latex particles resulting in a decreased apparent size by comparison with light scattering and other methods, although with low beam currents good agreement has sometimes been obtained. Effects of variation in ionic strength and counter ion type on PCS measurements have led some authors to conclude that this technique includes either a layer of extended polymer chains at the surface of the particle or a layer of water of hydration.
In the particular case of emulsifier-free latices the presence of low numbers of small aggregates has been inferred from the angular dependence of diffusion coefficients obtained from PCS. The polydispersity factor for our latex (described above) is a mean value and the range of values obtained is quite high (0.006 to 0.135) even after filtering samples through two cellulose nitrate filters (0.12 pm). We take a polydispersity factor of about 0.1 as the criterion for monodispersity.
The hydrodynamic diameter measured by PCS under these conditions is expected to be an overestimate of the ‘true’ diameter but this amounts to a relatively small percentage error for particles of the size used in these experiments. The maximum error is likely to be about 20 nm corresponding to 4% of the value adopted giving an error of 8.2% in the specific surface area calculated from the particle dimensions assuming a density of 1.05 g cme3 for poly- styrene.
The conductometric titrations indicated the presence of both strong (sulphate) and weak (carboxyl) groups. During the period over which the titrations were carried out (2 mths) no decrease in the number of strong acid groups was observed.
The electrophoretic mobility value presented in Table 1 is the mean of the values obtained between pH 6.35 and pH 8.36 in IO-’ M NaCI.
For particles of diameter 0.5,~m suspended in a medium of ionic strength 10e2 M the ratio of the particle radius to the thickness of the diffuse double layer is about 80 and application of the Smoluchowski equation for zeta potential is not strictly correct while the combination of electrophoretic mobilities and ionic strengths lies outside the range of solutions offered by Wiersema et a/.13. The zeta potential presented in the table is therefore calculated from the Henry function.
Table 1 Latex proper?&
Diameter Specific Surface charge density Electrophoretic Zeta nm surface Strong acid Weak acid mobility potential
area charge charge cm2 s-’ V-’ mV cm2 mg-’ PC cm--’
____I-__
500 114 5.0 1.6 5.95 x IOP 78.6
"0 IO 20
Time (mln!
30
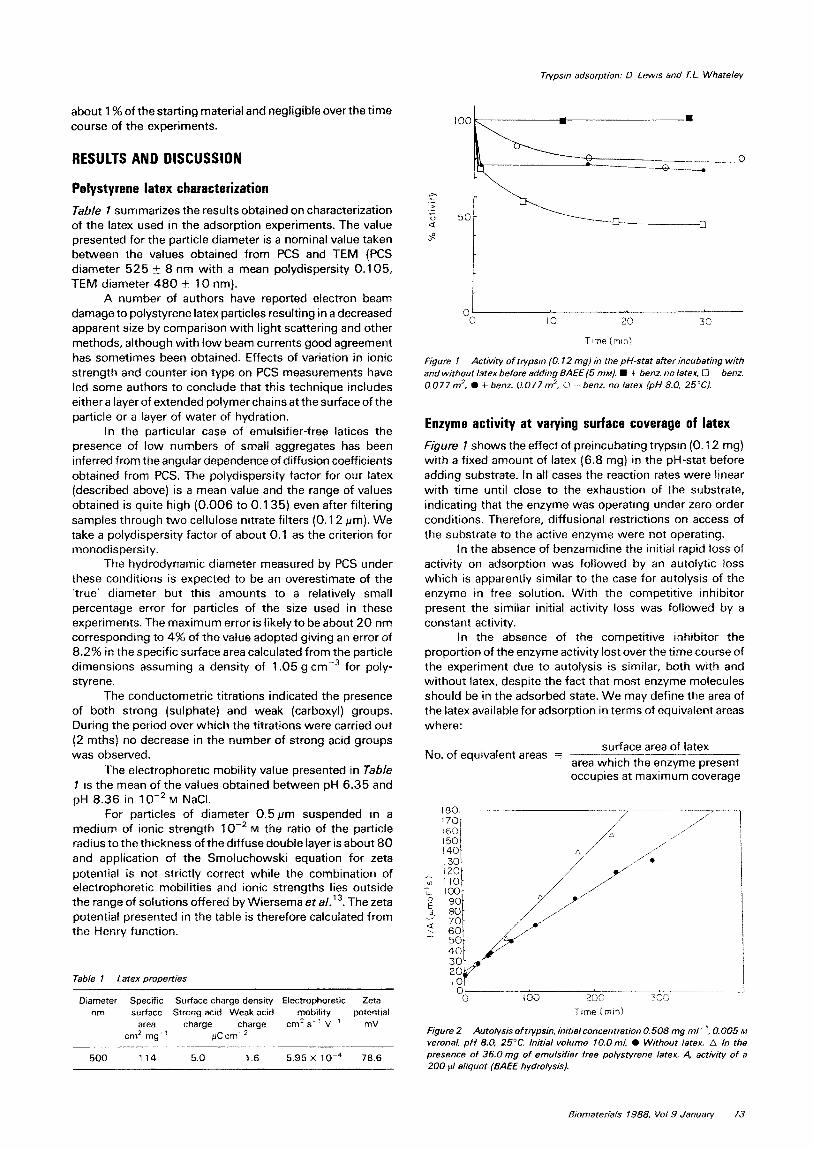
Figure 1 Activjry of try&n (0. I2 rng) in the pH-stat after incubating with and wjthout latex before add;ng 8AfE (5 m&f). S + benz. no latex, Cl - benz. 0.077m’. l f benz. 0.077m’. 0 - benz. no latex {pH 8.0, 25°C).
Enzyme activity at varying surface coverage of latex
Figure 1 shows the effect of preincubating trypsin (0.12 mg) with a fixed amount of latex (6.8 mg) in the pH-stat before adding substrate. In all cases the reaction rates were linear with time until close to the exhaustion of the substrate, indicating that the enzyme was operating under zero order conditions. Therefore, diffusional restrictions on access of the substrate to the active enzyme were not operating.
In the absence of benzamidine the initial rapid loss of activity on adsorption was followed by an autolytic loss which is apparently similar to the case for autolysis of the enzyme in free solution. With the competitive inhibitor present the similar initial activity loss was followed by a constant activity.
In the absence of the competitive inhibitor the propo~ion of the enzyme activity lost over the time course of the experiment due to autolysis is similar, both with and without latex, despite the fact that most enzyme molecules should be in the adsorbed state. We may define the area of the latex available for adsorption in terms of equivalent areas where:
surface area of latex No. of equivalent areas r -----___-....--_____
area which the enzyme present occupies at maximum coverage
Time Imin)
Figure 2 Auto&&s of trypsin. initial concentration 0.508 mg ml- ‘, 0.005 M Verona/, pH 8.0, 25°C. Initial volume lO.Oml. 0 Without latex. A In the presence of 36.0mg of emulsifier free polystyrene latex. 4 activity of a 200 fl aliquot (BAEE hydrolysis).
Biomateriats 7388, Vat 9 Januarv 73

Trypsin adsorption: 0. Lewis and T.L. Whateley
In the case of the data of Figure I the latex presents 1.8 equivalent areas to the protein. It is difficult to explain the high observed rate of autolysis of the adsorbed enzyme by a very low level of enzyme in the free form. Alternatively, enzyme molecules at the interface may have sufficient freedom to move about the surface of the particle to encounter one another and for autolysis to take place. Rates of autolysis in the absence and presence of latex are shown in Figure 2. Compared to the large increase in rate of autolysis of trypsin in the presence of silica surfaces, there is only a small increase in the presence of polystyrene surface.
Figure 3 compares the results obtained by premixing latex and enzyme at a range of equivalent areas to that of adding latex to the enzyme in the presence of BAEE (initially 5 mrvt). The reaction rates in the pH-stat showed a sharp change on adding latex followed by a gradual decrease in activity (as shown by curvature in the pH-stat trace) which was more marked the larger the equivalent area added. The residual activity in this case was taken as that represented by a tangent drawn to the curve 1 min after addition of latex. These values lie to the right of the results obtained by preincubation (i.e. a higher residual activity in the presence of BAEE).
The lower activity found in the case of pre-incubation of latex and enzyme together could have been due to autolysis of the trypsin taking place during this pre-incubation period of 10 min. As shown in Figure 1 (open square symbols) there is a significant loss in activity over a 10 min period. Figure 2 also shows the increased rate of autolysis of trypsin in the presence of the polystyrene latex.
However, pre-incubation of latex and enzyme could result in a higher level of adsorbed enzyme because BAEE
01 i 0 IO 20 30 40 50 6.0
Equlvolent areos
Figure 3 Comparison of the activity due to trvpsin, 0.12 mg, in the pH-stat. 0 Following the addition of enzyme to substrate, BAEE, 0.005 M, veronal. 0.00 1 M, 10 ml and 1.0 min later adding latex. 0 6y pre-incubation of the enzyme with latex veronal, 0.00 1 M, 5.0 ml for 10 min before adding BAEE, 0.02 M, 1.0 ml. (pH 8.0, 25°C).
03 0 0. I 0.2 0.3 0.4 0.5 0.6
Supernatont mm. (mg ml-‘)
Figure 4 Adsorption isotherms onto polystyrene latex in veronal buffer, 0.005 w with benzamidine, 0.00 1 M, pH 8.0,25”C (@ Trypsin, 0 Trypsinogen).
can compete with and reduce the adsorption of trypsin (as detailed later using TLCK-trypsin). With a higher level of adsorbed trypsin, the overall level of measured activity will be lower (open circles, Figure 3).
The effect shown in Figure 3 that as the area of polystyrene available for adsorption increases (i.e. as the surface coverage decreases) the measured enzyme activity decreases, is probably explained by the protein being able to ‘spread’ to a greater extent on the less occupied surface. At high surface coverage (i.e. equivalent area < 1) the close packing of the enzyme on the surface prevents significant ‘spreading’ and loss of conformation. At lower surface coverages hydrophobic interactions between protein and surface can be maximized by the ‘spreading’ of the protein with loss of native conformation and enzyme activity.
The adsorption isotherm for trypsin (that for trypsinogen is also shown) in Figure 4 confirms the high affinity, Langmuirian nature of the adsorption process. Thus, in the results presented in Figure 3, at equivalent areas greater than 1, essentially all enzyme is on the surface.
Trypsin (isoelectric point 10.8) and its zymogen, trypsinogen (isoelectric point 9.3) give the same plateau adsorption value (about 2.8 mg m-‘). This agrees with the conclusions now generally accepted regarding protein adsorption, that for adsorption onto a hydrophobic surface, electrostatic factors are of minor importance, the process being entropically driven, largely by the ‘spreading’ or ‘unfolding’ of the protein at the interface. The results with enzyme activity measurements of trypsin at the polystyrene surface confirm this view3’4,6.
There are a number of potential complicating factors in these experiments. In some situations flocculation of the polystyrene latex occurs on addition of trypsin: this does not occur when a-chymotrypsin is the enzyme and results on
74 Biomaterials 1988, Vol9 January

Table 2 Comparison of the adsorption of trypsin and TLCKinhibited trypsin
to polystyrene latex, the latter in the presence and absence of BAEE (5 mM). Conditions, pH 8.0, 25°C in 5 rnM veronal buffer, contact time 60 min.
Protein BAEE Final solution
concentration mg ml-’
Adsorbed amount mg m-’
-
Trypsin X 0.07 2.72
TLCK-Trypsin X 0.08 2.73
TLCK-Trypsin \ 0.1 1 1.07
this system will be presented later. One can conclude that flocculation in the systems used in this work with trypsin do not affect either measured enzyme activities or adsorption isotherms. The flocculation process has been studied in detail using PCS: rates of particle-particle interaction are on a much longer time-scale than protein-particle and protein- substrate interactions.
There is no evidence that electrostatic interactions between the weakly charged polystyrene latex and substrate, or hydrogen ions, affect the determined enzyme activities. Variation of pH and ionic strength and the determination of enzyme kinetic parameters such as K,,, and V,,,,,, which have been carried out for adsorbed a-chymotrypsin, allow these effects to be eliminated.
However, in the case of trypsin adsorption, the substrate, BAEE, has been found to affect the adsorption properties of trypsin: Table 2 shows the plateau adsorption values for TLCK-trypsin (an irreversibly inhibited form of trypsin, so that BAEE breakdown does not occur during the adsorption experiment). In the presence of BAEE (at the concentration present initially in enzyme assays) the plateau adsorption of trypsin is significantly reduced. This effect is difficult to relate to measured activities of adsorbed trypsin because the BAEE concentration is changing throughout the course of an assay. It may be that the surface coverages
estimated in Figure 3 are lower than indicated due to displacement by substrate.
REFERENCES
1
2
3
4
5
6
7
8
9
10
11
12
13
Trypsin adsorption: 0. Lews and T L. Whateley
Johnson, P. and Whateley. T.L., The effect of glass and silica surfaces on trypsin and a-chymotrypsin kinetics, Biochem. Biophys. Acta. 1972,276.323-327. Johnson, P. and Whateley, T.L., The effects of surface and macro- molecular interactions on the krnetrcs of inactrvation of trypsin and a-chymotrypsin, Biochem. J. 1981, 193, 285-294 Koutsoukos, P.E.. Mumme-Young, CA., Norde, W. and Lyklema, J., Effect of the nature of the substrate on the adsorption of human plasma albumin, Colloids Surfaces 1982, 5. 93-104 Norde, W., MacRitchie, F.. Nowicka, G. and Lyklema, J., Protein adsorption at solid-liquid interfaces: reversibrlity and conformation aspects, J. Co/l. lnterf. Sci. 1986, 112. 447-456 Andrade, J.D. (Ed.), Protein Adsorption at Polymer-Water Interfaces
1985, Plenum Press, New York Vincent, 6. and Whittington, S.G., Polymers at interfaces and in
disperse systems, Surface and Colloid Science (Ed. Matilevic, E.), 1982, Plenum Press, New York, 12. 1- 1 1 7 Sandwick, R.K. and Schray, K.J., The inactrvatton of enzymes upon interactron with a hydrophobrc latex surface,J. Co//. Interf So. 1987, 115, 130-l 38
Goodwin, J.W., Hearn, J., Ho, C.C. and Ottewrll. R.H.. Studies on the preparation and characterisation of monodisperse polystyrene latices, Colloid Poly. Sci. 1974. 252, 464-471. Van den Hul, H.J. and Vanderhoff, J.W., The characterisation of latex particle surfaces by ion exchange and conductometric trtration, J. Electroanal. Chem. lnterf Electrochem. 1972, 37, 161-l 82
Koppel. DE.. Analysis of macromolecular polydispersrty rn intensity correlation spectroscopy: the method of cumulants. J. Chem. Phys. 1972, 57,4814-4820 Lowry. O.H.. Rosebrough. N.J.. Farr. A.L. and Randall, R.J., Protern measurement with the folrn phenol reagent, J. 8iol. Chem. 1951, 193. 265-275
Itzhaki. R.F. and Grll, D.M., A micro-bruret method for estrmatrng proterns, Anal. Biochem. 1964, 9. 40 l-41 0
Wiersma. P.H., Loeb. A.L. and Overbeek, J. Th. G., Calculation of the electrophoretrc mobrltty of a spherical collord partrcle, J. Co//. lnterf SC. 1966, 22. 78-99
Biomaterials 1988, Vol9 January 75





























