Embed Size (px)
Citation preview

Contents lists available at ScienceDirect
Acta Tropica
journal homepage: www.elsevier.com/locate/actatropica
Aedes aegypti(Linnaeus) larvae from dengue outbreak areas in Selangorshowing resistance to pyrethroids but susceptible to organophosphates
Cherng Shii Leong⁎, Indra Vythilingam, Meng Li Wong, Wan-Yusoff Wan Sulaiman, Yee Ling LauDepartment of Parasitology, Faculty of Medicine, University of Malaya, 50603, Kuala Lumpur, Malaysia
A R T I C L E I N F O
Keywords:Aedes aegyptiInsecticides resistanceSynergistWHO larva bioassays & biochemical assays
A B S T R A C T
The resistance status of Selangor Aedes aegypti (Linnaeus) larvae against four major groups of insecticides (i.e.,organochlorines, carbamates, organophosphates and pyrethroids) was investigated. Aedes aegypti were suscep-tible against temephos (organophosphate), although resistance (RR50=0.21–2.64) may be developing. Theinsecticides susceptibility status of Ae. aegypti larvae were found heterogeneous among the different study sites.Results showed that Ae. aegypti larvae from Klang, Sabak Bernam and Sepang were susceptible against all in-secticides tested. However, other study sites exhibited low to high resistance against all pyrethroids (RR50 =1.19–32.16). Overall, the application of synergists ethacrynic acid, S.S.S.- tributylphosphorotrithioate and pi-peronyl butoxide increased the toxicity of insecticides investigated. However, the application failed to increasethe mortality to susceptible level (> 97%) for certain populations, therefore there are chances of alteration oftarget site resistance involved. Biochemical assays revealed that α-esterase, (Gombak, Kuala Langat, KualaSelangor and Sabak Bernam strains) β-esterase (Klang and Sabak Bernam strains), acetylcholinesterase (KualaSelangor and Sabak Bernam strains), glutathione-S-transferase (Kuala Selangor and Sabak Bernam strains) andmono-oxygenases (Gombak, Hulu Langat, Hulu Selangor and Kuala Langat strains) were elevated. Spearmanrank-order correlation indicated a significant correlation between resistance ratios of: DDT and deltamethrin (r= 0.683, P = 0.042), cyfluthrin and deltamethrin (r = 0.867, P =0.002), cyflyuthrin and lambdacyhalothrin (r= 0.800, P=0.010), cyfluthrin and permethrin (r=0.770, P=0.015) deltamethrin and permethrin (r=0.803,P =0.088), propoxur and malathion (r = 0.867, P = 0.002), malathion and temephos (r= 0.800, P = 0.010),etofenprox and MFO enzyme (r = 0.667, P =0.050). The current study provides baseline information for vectorcontrol programs conducted by local authorities. The susceptibility status of Ae. aegypti should be monitoredsporadically to ensure the effectiveness of current vector control strategy in Selangor.
1. Introduction
Mosquitoes play an important role as vectors of parasites and pa-thogens, due to its blood-sucking characteristics (Benelli and Dunggan;2018). Mosquitoes are threat to humans and animals due to its ability intransmitting destructive parasites and pathogens, including dengue,malaria, yellow fever, Zika virus, chikungunya, filariasis, encephalitis,heartworm and West Nile virus (Benelli, 2015; Mehlhorn, 2016).
The battle against mosquito borne diseases is a huge challenge ofpublic health importance (Benelli and Mehlhorn, 2016). Althoughmalaria cases have declined in recent years, the other arboviruses suchas dengue, Zika virus and chikungunya that are carried by Aedes mos-quitoes have been frequently reported in many parts of the world(Attar, 2016; Benelli and Mehlhorn, 2016). To prevent outbreaks ofmosquito borne diseases, control measures such as application of in-secticides, either as space spraying, indoor residual spraying, longlasting insecticidal nets, bio-control agent and insect growth hormone
were carried out (Benelli and Beier, 2017; Benelli and Romano, 2017Benelli and Mehlhorn, 2016; Mehlhorn, 2016; Benelli, 2015). In-tegrated Vector Management (IVM) has been recommended in makinguse of the full range of vector control tools available to prevent out-breaks (WHO, 2012; Benelli and Beier, 2017). In recent years, OneHealth approach was introduced, a control strategy which emphasizeson cooperation among multiple disciplines to achieve the best healthfor humans, animals and the environment (Benelli and Beier, 2017;Benelli and Duggan, 2018).
Dengue is a mosquito borne viral disease that causes serious publichealth problem in most of the tropical countries. The cases of denguehave increased 30 folds over the past five decades (WHO, 2017). Therewere 4.5 million dengue cases reported in 2016 and 3.9 billion peopleliving in the areas with risk of dengue infection (WHO, 2017). In Ma-laysia, there were 101,357 cases of dengue with 237 deaths (IdengueMOH-Malaysia: unpublished recorded from http://idengue.remotesensing.gov.my/idengue/index.php). Selangor is one of the
https://doi.org/10.1016/j.actatropica.2018.05.008Received 16 January 2018; Received in revised form 2 May 2018; Accepted 6 May 2018
⁎ Corresponding author.
Acta Tropica 185 (2018) 115–126
0001-706X/ © 2018 Published by Elsevier B.V.
T

states in Malaysia which reported the highest number of cases (51,652cases) in the country (MOH, 2016).
Aedes aegypti (Linnaeus) is the vector of dengue and it can transmitall four serotypes as well as other viral diseases like Chikungunya andZika virus (Cheong et al., 1986; Benelli and Romano, 2017). Un-fortunately, there are no drugs to treat dengue and the vaccine is onlypartially effective. Thus, vector control has been the hallmark of thedengue control programme in Malaysia. Application of larvicide liketemephos (Abate 1% Sand Granules) which is an organophosphate hasbeen used for many decades (Chen et al., 2005a) and carried out byhouse owners or during epidemics in which the health personnel willapply the larvicides to containers which cannot be disposed and wherethe water is required by the people. Besides community participation insource reduction, fogging or ultra-low volume (ULV) is carried out bythe authorities when cases of dengue are reported. The insecticides usedfor fogging are mainly organophosphates (malathion & fenitrothion)and pyrethroids.
Insecticides like pyrethroids are not only used in public health butalso in agriculture and most of the aerosol insecticides used by houseowners are also pyrethroids (Yap et al., 2000). The development ofinsecticides resistance has become a serious problem worldwide due tothe excessive use of insecticides and also there is limited insecticides foruse in public health. Aedes aegypti from different localities in the worldhave been documented to be resistant to several classes of insecticides.(Araújo et al., 2013; Garcia et al., 2009; Grisales et al., 2013; Limaet al., 2011; Marcombe et al., 2012; Ocampo et al., 2011;Prapanthadara et al., 2002).
It is not known if the mosquitoes have become resistant to in-secticides and that could be one of the reasons for the increase ofdengue cases. On the other hand, because asymptomatic persons aremore infectious to the virus (Duong et al., 2015), control measuresinstituted after the case has been reported is perhaps too late. Thus, amore proactive surveillance is needed to prevent the increase in thenumber of dengue cases and also the management and use of in-secticides is important.
Although numerous studies have been conducted in Malaysia withregards to insecticide resistance to Ae. aegypti (Chen et al., 2008; Hasanet al., 2016; Lee et al., 2008; Loke et al., 2012; Wan-Norafikah et al.,2010), no robust and comprehensive study has been carried out tocompare the resistance/susceptibility status of the said species to allfour classes of insecticides in dengue outbreak and non-outbreak areasin Selangor state. We therefore examined the susceptibility status of Ae.aegypti larvae to all four classes of insecticides using bioassays andbiochemical methods. The efficacy of synergist on insecticides was alsoexplored.
2. Materials and methods
2.1. Sample collection
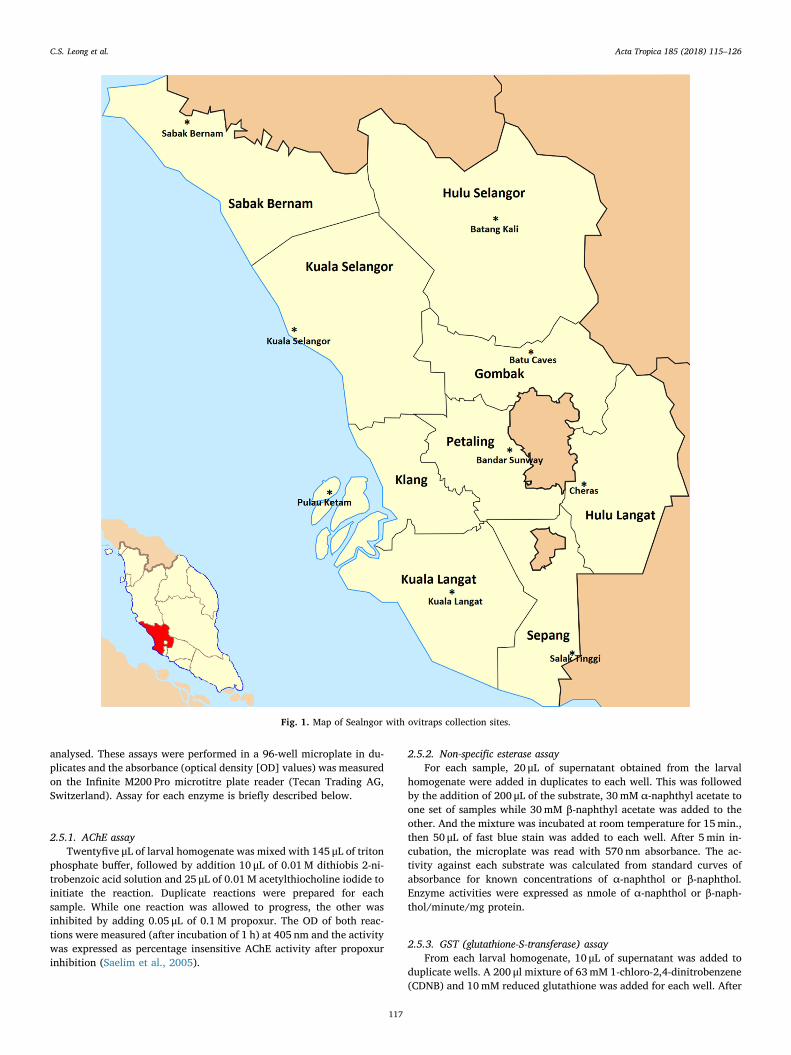
Selangor is the most populated and well-developed state inPeninsular Malaysia. Its central position contributed to rapid in-dustrialization and is the main transportation hub for the country.Selangor contributes 23% of the total GDP of Malaysia (Department ofStatistics, 2014) and comprises of nine districts. Aedes aegypti werecollected from September 2015 to April 2016 using ovitraps and larvalsurveys from all nine districts in Selangor state. Fig. 1 shows the map ofSelangor with ovitraps collection sites. Table 1 shows the districts andsites from where the study was conducted. The sampling sites wereselected based on dengue outbreak and dengue free areas. Eggs col-lected from each site were hatched and maintained as a single colony.All emerged adult mosquitoes were identified to species by morpholo-gical characteristics (WHO, 2003). The Ae. aegypti colonies weremaintained under standard insectary condition with 27 ± 2 °C,75 ± 5% relative humidity, a 10 h : 14 h (light:dark) light cycles andprovided with 10% sucrose solution (vitamin B complex). Blood meals
were provided for five – seven days old adult females Ae. aegypti usinglive white mouse (IACUC no.: 20150407/PARA/R/MBK). Bora-borastrain of Ae. aegypti was used as a reference susceptible strain. The re-ference strains have been reared in the insectary for 134 generationswithout exposure to insecticides.
2.2. Insecticides tested
All four major classes of the neurotoxic insecticides (technicalgrade) were used for the experiments. These were organochlorine: DDT(98%); carbamate: propoxur (99.8%); organophosphates: malathion(98.7%), temephos (97.5%); pyrethroids: cyfluthrin (99.8%), deltame-thrin (99.6%), etofenprox (97.7%), lambdacyhalothrin (97.8%) andpermethrin (98.1%). All insecticides were purchased from SigmaAldrich (Germany).
2.3. Larval bioassays
Larval bioassays were conducted following the larval susceptibilitybioassay procedure (WHO, 1981, 2005). The test was conducted usingdisposable 300ml paper cup containing 249ml of distilled water with1ml of insecticide solution. Twenty to 25 late third to early fourth-instar larvae were randomly selected and transferred to the holding cupfor 10–15min before addition of the insecticide solution. The larvaewere exposed for 24 h after which mortality was recorded. To de-termine the diagnostic dose, Bora-Bora strain was tested with fiveconcentrations for each insecticide in nine replicates to obtain mortalitythat ranges from 5 to 95% to generate LC50 and LC99 values accordingto WHO guidelines (WHO, 1981, 2005). For each test, five controlswere set using 1ml of 10% ethanol. The diagnostic dose, which is de-fined as two times (2X) lethal concentration that kills 99% of the re-ference population tested (LC99 X 2). The mortality rate of F2 genera-tion of field collected mosquito larvae were determined using thediagnostic dosage. Three tests, each performed on three different days,were carried out for each insecticide for all the nine field strains. Si-milarly, five to seven different concentrations for each insecticide wereused to determine lethal concentration of the insecticide that kills 50%(LC50) and 99% (LC99) of test population.
2.4. Synergism tests
All field strains of Ae. aegypti were subjected to synergism tests toevaluate their effectiveness against the detoxification of insecticides.Ethacrynic acid 99% (EA), Piperonyl butoxide 99% (PBO) and S.S.S-tributlyphosphorotrithioate 97.2% (DEF) all from Sigma-Aldrich wereused in this study. Larvae of reference strain were exposed to all threesynergists at different concentrations, and maximum sub lethal con-centration were determined. After a series of trial and error, the sub-lethal doses of larvae synergism test were 0.1, 0.01, and 5mg/L for EA,DEF and PBO, respectively. Larvae synergism tests were conducted al-most similar to larvae bioassays, with an additional step by mixing thesynergist and insecticides in the ratio of 1:1 before exposing to larvae.Each of the synergist was added to all insecticides.
2.5. Biochemical assays
The different enzyme levels in individual larvae were determinedaccording to the WHO procedure, as described by Hemingway andBrogdon (1998). Briefly, fourth-instar larvae were (Bora-Bora and fieldstrains) individually homogenized in 200 μL of distilled water (on ice).Twenty-five microliters of homogenate were used for the AChE (acet-ylcholinesterase) assay. The remaining homogenate was then cen-trifuged at 14,000 rpm at four degrees Celsius for one minute, and thesupernatant was used as an enzyme source for all other biochemicalassays. For each assay, blanks were included (only distilled water in-stead of mosquito homogenate). Ninety-four larvae per strain were
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
116
analysed. These assays were performed in a 96-well microplate in du-plicates and the absorbance (optical density [OD] values) was measuredon the Infinite M200 Pro microtitre plate reader (Tecan Trading AG,Switzerland). Assay for each enzyme is briefly described below.
2.5.1. AChE assayTwentyfive μL of larval homogenate was mixed with 145 μL of triton
phosphate buffer, followed by addition 10 μL of 0.01M dithiobis 2-ni-trobenzoic acid solution and 25 μL of 0.01M acetylthiocholine iodide toinitiate the reaction. Duplicate reactions were prepared for eachsample. While one reaction was allowed to progress, the other wasinhibited by adding 0.05 μL of 0.1 M propoxur. The OD of both reac-tions were measured (after incubation of 1 h) at 405 nm and the activitywas expressed as percentage insensitive AChE activity after propoxurinhibition (Saelim et al., 2005).
2.5.2. Non-specific esterase assayFor each sample, 20 μL of supernatant obtained from the larval
homogenate were added in duplicates to each well. This was followedby the addition of 200 μL of the substrate, 30mM α-naphthyl acetate toone set of samples while 30mM β-naphthyl acetate was added to theother. And the mixture was incubated at room temperature for 15min.,then 50 μL of fast blue stain was added to each well. After 5min in-cubation, the microplate was read with 570 nm absorbance. The ac-tivity against each substrate was calculated from standard curves ofabsorbance for known concentrations of α-naphthol or β-naphthol.Enzyme activities were expressed as nmole of α-naphthol or β-naph-thol/minute/mg protein.
2.5.3. GST (glutathione-S-transferase) assayFrom each larval homogenate, 10 μL of supernatant was added to
duplicate wells. A 200 μl mixture of 63mM 1-chloro-2,4-dinitrobenzene(CDNB) and 10mM reduced glutathione was added for each well. After
Fig. 1. Map of Sealngor with ovitraps collection sites.
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
117
20min of incubation at room temperature, absorbance was determinedat 340 nm. GST activity was calculated based on Beer’s Law (A= €cl)and reported as mmole of CDNB/minute/mg protein. Where A (ODvalue) was transformed to μmole of CDNB conjugates using the € (ex-tinction coefficient) of 4.39mM−1. The path length (the depth of thebuffer solution in the microplate well,) was 0.6 cm.
2.5.4. MFO (Monooxygenases) assayFor MFO activity 20 μL of supernatant was added to duplicate wells.
This was followed by the addition of 80 μL of 0.625M potassiumphosphate buffer (pH 7.2), 200 μL of 3,3,5,5-tetramethylbenzidine(TMBZ) in methanol solution, and 25 μL of 3% hydrogen peroxide. Itwas incubated at room temperature for 2 h before microplate was readat 650 nm. The MFO activity was calculated based on the standardcurve of absorbance for known concentration of cytochrome C(Brogdon and Janet, 1997). Enzyme activity is expressed as equivalentunits of cytochrome P450/min/mg protein.
2.5.5. Protein assayTo account for size variances among individuals, protein con-
centration was used as a standard correction factor for the analysis ofall enzymes activities. A commercialized protein assay (Bio-Rad, USA)was used to obtain the bovine serum albumin standard curve. Proteinconcentration was calculated and transformed based on the bovineserum albumin standard curve. Ten microliters of larval homogenatesolution were mixed with 300 μl of Bio-Rad dye reagent and incubatedfor 5min. and was read at 570 nm.
2.6. Statistical analysis
The mortality rate (%) and resistance ratio (RR50) were used toexpress the Ae. aegypti susceptibility status. Mortality rate was used toevaluate the effectiveness of synergists against insecticides. On theother hand, resistance ratio provides information on the degree of re-sistance of field strains compared to reference strain.
Larval bioassay data within the range 5–95% were subjected toprobit analysis methods of Finney (1947) to obtain the LC50 and LC99
values for each insecticide. Data from bioassays were pooled for ana-lysis. Resistance ratios (RR50) were calculated by dividing values for thefield strain with the reference strain based on the LC50 obtained from
the larval bioassays (Brown, 1958). Levels of resistance were calculatedaccording to WHO (2016) standard where the calculated RR50 va-lues< 1 were expressed as susceptible, RR50 values< 5 were expressedas low resistance, 5–10 were expressed as medium resistance and RR50
values> 10 were expressed as high resistance. If the control mortalitywas> 5%, but< 20% the percentage mortalities was corrected byAbbott’s (1925) formula. The mortality rate 98%–100% indicates sus-ceptible, 80% - 97% suggests the possibility of resistance that need to befurther confirmed and< 80% suggested resistance (WHO, 2016).
Mortality rate and enzymes levels were subjected to Levene’s andKolmogorov-Smirnov tests. Data not normally distributed were trans-formed to arcsine log to stabilize the variance. Two-sample t-test/Mann-Whitney non-parametric test was performed to compared the differencebetween field strains and Bora-bora strain. Statistical significance wasassumed at P<0.05. All data were analysed and interpreted using SPSS(IBM SPSS Statistics 19) software. All graph was generated usingGraphPad Prism V.5.0 (GraphPad Software) and Microsoft Excel ver-sion 2016 (Microsoft Inc.).
3. Results
The Bora-Bora strain of Ae. aegypti exhibited least toxicity to pro-poxur (LC50 0.41755mg/L) and was most susceptible to deltamethrin(0.00006mg/L). The diagnostic dosage of the different larvicides basedon Bora-Bora strain are shown in Table 2. Larvae from all sites showed100% mortality to porpoxur and malathion only (Table 3). For DDT,larvae from all sites showed more than 90% mortality with the excep-tion of Gombak (88.44%) and Hulu Selangor (32.95%). This corrobo-rates well with the RR50 value of these two strains which were 3.84 and5.64 respectively. This shows that the Hulu Selangor strain was mod-erately resistant (Table 4).
Larvae from six of the sites showed 100% mortality towards teme-phos while only the Hulu Langat, Petaling and Kuala Selangor strainsthe mortality ranged from 98.67% to 96.92%. However, the RR50 valueof Petaling strain was 2.64 while all other strains had RR50< 1(Table 4). This shows that temephos was still susceptible to larvae from
Table 1Description of study sites.
District Sitecollection
Coordination Status Landscape
Gombak Batu Caves 3O25’80.90"N101O 64’51.3"E
Dengue-outbreak
Urban
Hulu Langat Cheras 3°01'19.4"N101°45'31.1"E
Dengue-outbreak
Urban
Hulu Selangor Batang Kali 3°27'20.6"N101°39'18.4"E
Non-dengue-outbreak
Sub-urban
Klang Pulau Ketam 3°01'18.5"N101°15'12.4"E
Non dengue-outbreak
Rural
Kuala Langat Banting 2°48'25.9"N101°29'17.8"E
Non-dengue-outbreak
Sub-urban
Kuala Selangor KualaSelangor
3°14'23.8"N101°25'35.2"E
Dengue outbreak Urban
Sabak Bernam SabakBernam
3°45'51.6"N100°59'17.5"E
Non-dengueoutbreak
Rural
Petaling Jaya BandarSunway
3°04'54.3"N101°36'38.6"E
Dengue outbreak Urban
Sepang Salak Tinggi 2°49'42.8"N101°43'34.5"E
Non-dengueoutbreak
Sub-urban
*Dengue-outbreak – Ovitrap was conducted after fogging and frequent denguecases reported in the areas.*Non dengue-outbreak – Ovitrap was conducted with prior survey of no foggingactivity conducted for at least 2 month.
Table 2Diagnostic dose (mg/L) of different larvicides based on Bora-Bora strain.
Insecticides n LC50
(95% CL)(mg/L)
LC99
(95% CL) (mg/L)Diagnostic dose(mg/L)
DDT 1203 0.15482(0.12818-0.18591)
5.60492(3.03397-13.95812)
11.210
Propoxur 1176 0.41755(0.021804-0.68622)
5.82678(2.07143-463.92613)
11.653
Malathion 1211 0.04457(0.03997-0.04926)
0.31488(0.23102-0.49175)
0.629
Temephos 1230 0.00168(0.00142-0.00199)
0.04121(0.02473-0.08533)
0.082
Cyfluthrin 1174 0.00019(0.00017-0.00021)
0.00170(0.00118-0.00289)
0.003
Deltamethrin 1245 0.00006(0.00004-0.00008)
0.00145(0.00054-0.01642)
0.003
Etofenprox 1372 0.00441(0.00113-0.03436)
0.08050(0.06576-2.39374467.5)
0.161
Lambdacyhalothrin 1378 0.00009(0.00007-0.00011)
0.01077(0.00494-0.03337)
0.022
Permethrin 1423 0.00090(0.00079-0.00103)
0.01631(0.01030-0.03148)
0.033
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
118
Table 3Mortality rate of Ae. aegypti larvae 24-hours after exposure to insecticides of diagnostic dosage and synergists.
Insecticides +Synergists
Mean % mortality± SE
Bora-Bora Gombak Hulu Langat Hulu Selangor Klang Kuala Langat Kuala Selangor Petaling SabakBernam
Sepang
Total exposed (n) 1186 1228 1136 1133 1141 1131 1170 1144 1141 1136DDT only 100 ± 0 88.44 ± 1.82* 91.65 ± 2.94* 32.95 ± 3.68*a 100 ± 0 100 ± 0 90.74 ± 0.69 97.33 ± 1.15 100 ± 0 100 ± 0DDT + PBO 100 ± 0 100 ± 0 100 ± 0 89.38 ± 1.88*a 100 ± 0 100 ± 0 100 ± 0 97.81 ± 0.96 100 ± 0 100 ± 0DDT + DEF 100 ± 0 96.89 ± 1.46 100 ± 0 78.72 ± 2.85*a 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0DDT + EA 100 ± 0 100 ± 0 100 ± 0 95.11 ± 2.29a 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0DDT + 3 synergists 100 ± 0 100 ± 0 100 ± 0 80.22 ± 3.08*a 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0Total exposed (n) 1186 1234 1158 1143 1136 1138 1144 1190 1136 1134Propoxur only 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0Propoxur + PBO 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0Propoxur + DEF 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0Propoxur + EA 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0Propoxur + 3
synergists100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0
Total exposed (n) 1235 1233 1146 1142 1135 1131 1145 1143 1135 1128Malathion only 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0Malathion + PBO 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0Malathion + DEF 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0Malathion + EA 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0Malathion + 3
synergists100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0
Total exposed (n) 1213 1178 1141 1147 1136 1130 1143 1155 1136 1129Temephos only 100 ± 0 100 ± 0 98.67 ± 0.94 100 ± 0 100 ± 0 100 ± 0 96.92 ± 1.28 97.33 ± 1.15 100 ± 0 100 ± 0Temephos + PBO 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 97.33 ± 1.49 95.16 ± 1.30 100 ± 0 100 ± 0Temephos + DEF 100 ± 0 100 ± 0 82.67 ± 2.49* 100 ± 0 100 ± 0 100 ± 0 97.33 ± 1.49 89.04 ± 1.93 100 ± 0 100 ± 0Temephos + EA 100 ± 0 100 ± 0 96.04 ± 1.48 100 ± 0 100 ± 0 100 ± 0 96.89 ± 1.60 100 ± 0 100 ± 0 100 ± 0Temephos + 3
synergists100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0 100 ± 0
Total exposed (n) 1156 1211 1138 1146 1131 1129 1133 1178 1131 1127Cyfluthrin only 100 ± 0 64.33 ± 1.59* 80.97 ± 4.35* 11.03 ± 1.61* 100 ± 0 26.86 ± 2.23* 51.69 ± 4.80* 22.22 ± 2.75* 100 ± 0 100 ± 0Cyfluthrin + PBO 100 ± 0 100 ± 0a 92.93 ± 1.61 39.00 ± 2.70*a 100 ± 0 100 ± 0a 54.67 ± 2.83* 86.39 ± 3.12*a 100 ± 0 100 ± 0Cyfluthrin + DEF 100 ± 0 85.42 ± 2.08*a 81.66 ± 2.39* 13.56 ± 2.86* 100 ± 0 53.33 ± 4.42*a 53.28 ± 2.60* 89.92 ± 2.54a 100 ± 0 100 ± 0Cyfluthrin + EA 100 ± 0 79.26 ± 1.34*a 80.61 ± 2.46* 44.44 ± 4.59*a 100 ± 0 75.78 ± 4.33*a 50.37 ± 3.42* 78.18 ± 2.46*a 100 ± 0 100 ± 0Cyfluthrin + 3
synergists100 ± 0 100 ± 0a 83.39 ± 2.38* 34.58 ± 3.81*a 100 ± 0 100 ± 0a 58.58 ± 2.73* 85.29 ± 2.12*a 100 ± 0 100 ± 0
Total exposed (n) 1155 1181 1132 1131 1143 1128 1163 1153 1143 1129Deltamethrin only 100 ± 0 73.01 ± 1.86* 60.35 ± 3.38* 10.64 ± 2.22* 100 ± 0 44.22 ± 2.37*a 26.45 ± 1.63* 24.44 ± 2.35* 100 ± 0 100 ± 0Deltamethrin + PBO 100 ± 0 100 ± 0a 82.42 ± 5.28*a 46.50 ± 3.63*a 100 ± 0 100 ± 0a 29.50 ± 2.08* 85.33 ± 5.89*a 100 ± 0 100 ± 0Deltamethrin + DEF 100 ± 0 77.20 ± 1.48* 65.89 ± 6.76* 12.00 ± 2.58* 100 ± 0 73.33 ± 3.20*a 37.33 ± 2.49* 66.46 ± 2.97*a 100 ± 0 100 ± 0Deltamethrin + EA 100 ± 0 74.34 ± 2.11* 73.68 ± 4.50*a 15.98 ± 3.06* 100 ± 0 62.22 ± 2.12*a 83.92 ± 4.31*a 78.34 ± 3.23*a 100 ± 0 100 ± 0Deltamethrin + 3
synergists100 ± 0 100 ± 0a 74.87 ± 4.11*a 33.45 ± 4.23*a 100 ± 0 100 ± 0a 89.85 ± 2.23a 80.97 ± 1.11*a 100 ± 0 100 ± 0
Total exposed (n) 1150 1168 1137 1134 1139 1131 1140 1143 1139 1128Etofenprox only 100 ± 0 89.78 ± 2.12 94.32 ± 1.13* 61.04 ± 2.91*a 100 ± 0 80.36 ± 1.46* 90.78 ± 1.18 93.33 ± 1.33 100 ± 0 100 ± 0Etofenprox + PBO 100 ± 0 100 ± 0a 96.49 ± 1.04* 91.61 ± 2.26a 100 ± 0 82.32 ± 2.56* 100 ± 0 100 ± 0 100 ± 0 100 ± 0Etofenprox + DEF 100 ± 0 100 ± 0a 89.08 ± 3.62* 83.56 ± 4.39*a 100 ± 0 100 ± 0a 100 ± 0 95.03 ± 1.69 100 ± 0 100 ± 0Etofenprox + EA 100 ± 0 100 ± 0a 100 ± 0 88.96 ± 2.63a 100 ± 0 83.28 ± 2.35* 100 ± 0 96.03 ± 1.33 100 ± 0 100 ± 0Etofenprox + 3
synergists100 ± 0 100 ± 0a 100 ± 0 82.80 ± 2.29*a 100 ± 0 98.67 ± 0.94a 100 ± 0 100 ± 0 100 ± 0 100 ± 0
Total exposed (n) 1151 1913 1141 1133 1143 1129 1137 1186 1143 1127Lambdacyhalothrin
only100 ± 0 75.56 ± 1.41*a 91.91 ± 2.79 29.95 ± 2.65* 100 ± 0 65.30 ± 3.12* 70.36 ± 1.76* 59.56 ± 2.94* 100 ± 0 100 ± 0
Lambdacyhalothrin+ PBO
100 ± 0 100 ± 0a 95.16 ± 1.30 93.42 ± 3.00a 100 ± 0 92.44 ± 1.69*a 72.90 ± 2.74* 97.33 ± 0.94a 100 ± 0 100 ± 0
Lambdacyhalothrin+ DEF
100 ± 0 100 ± 0a 90.91 ± 3.13 43.47 ± 4.60*a 100 ± 0 70.41 ± 3.44* 70.67 ± 1.49 91.26 ± 2.38a 100 ± 0 100 ± 0
Lambdacyhalothrin+ EA
100 ± 0 87.11 ± 1.74*a 87.78 ± 2.97* 96.00 ± 1.86a 100 ± 0 62.22 ± 5.13* 93.33 ± 1.89a 90.88 ± 1.19a 100 ± 0 100 ± 0
Lambdacyhalothrin+ 3 synergists
100 ± 0 100 ± 0a 82.48 ± 2.62* 86.29 ± 2.36*a 100 ± 0 99.56 ± 0.44a 100 ± 0a 90.82 ± 2.59a 100 ± 0 100 ± 0
Total exposed (n) 1204 1192 1148 1134 1143 1131 1139 1165 1143 1131Permethrin only 100 ± 0 100 ± 0 96.18 ± 1.28 91.57 ± 4.02 100 ± 0 90.70 ± 2.50 42.67 ± 3.19* 72.00 ± 2.21* 100 ± 0 100 ± 0Permethrin + PBO 100 ± 0 100 ± 0 100 ± 0 93.81 ± 2.23 100 ± 0 100 ± 0 53.78 ± 4.48* 90.67 ± 2.58a 100 ± 0 100 ± 0Permethrin + DEF 100 ± 0 100 ± 0 91.73 ± 2.37 91.61 ± 2.25 100 ± 0 89.33 ± 1.76 55.66 ± 6.42* 72.85 ± 3.21* 100 ± 0 100 ± 0Permethrin + EA 100 ± 0 100 ± 0 83.88 ± 3.85* 87.32 ± 2.23* 100 ± 0 65.33 ± 2.11* 76.46 ± 2.80*a 95.62 ± 1.37a 100 ± 0 100 ± 0Permethrin + 3
synergists100 ± 0 100 ± 0 92.58 ± 2.36 91.59 ± 2.71 100 ± 0 100 ± 0 88.48 ± 1.42a 90.26 ± 2.32a 100 ± 0 100 ± 0
Mean % mortality followed by asterisk symbol denotes rates that were significantly different when compared with Bora-bora strain (P<0.05, independent T−test).Mean followed by a superscript letter were significant different between synergist(s) threated versus non−synergist(s) threated, P< 0.05, Mann-Whitney U test.
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
119
Table 4Susceptibility of the Ae. aegypti larvae to various insecticides.
Insecticide Strain n LC50
(95% CL) (mg/L)LC99
(95% CL) (mg/L)Slope x2(df) RR50
DDT Bora-Bora 1203 0.15482(0.12818-0.18591)
5.60492(3.03397-13.95812)
9.020 ± 0.135 0.79(3)
–
Gombak 1255 0.59462(0.42305 - 0.80232)
2.90911(1.57590 - 29.21118)
8.354 ± 0.091 11.30 (3) 3.84
Hulu Langat 1146 0.74569*
(0.66852-0.83305)6.31578(4.31662-11.18140)
4.594 ± 0.320 1.04 (3) 4.82
Hulu Selangor 1130 0.87395*
(0.78229 – 0.96309)5.93164(4.30124 - 9.64713)
2.730 ± 0.060 0.71(3)
5.64
Klang 1178 0.01222(0.00595 - 0.02919)
0.72852(0.14527 - 172.54671)
10.014 ± 0.250 10.62 (3) 0.08
Kuala Langat 1130 0.03678(0.01665 - 0.10266)
14.27176(1.75692 - 2982.84299)
8.751 ± 0.147 7.21(3)
0.24
Kuala Selangor 1208 0.33465(0.21873 - 0.46650)
4.66156(2.16930 - 28.09479)
10.373 ± 0.093 6.44(3)
2.16
Petaling 1167 0.15886(0.04137 - 0.60838)
10.37099(1.46911 - 22748.65236)
8.251 ± 0.124 16.78 (3) 1.03
Sabak Bernam 1250 0.07203(0.02274 - 0.32187)
5.52793(0.72965 - 3081529.601)
9.002 ± 1.410 17.40 (3) 0.47
Sepang 1198 0.04019(0.01328 - 0.12832)
9.89097(1.16975 - 11820.30857)
9.939 ± 0.137 11.89 (3) 0.26
Propoxur Bora-Bora 1176 0.41755(0.021804-0.68622)
5.82678(2.07143-463.92613)
8.535 ± 0.771 11.82 (3) –
Gombak 1235 0.68013(0.03969 - 0.11284)
1.33782(0.45234 - 41.37951)
10.383 ± 0.090 9.22 (3) 1.63
Hulu Langat 1134 0.89864(0.67758-1.40572)
4.57604(2.27608-65.72541)
9.637 ± 0.357 11.40 (3) 2.15
Hulu Selangor 1125 0.88679(0.52677 - 1.32921)
4.45991(2.20196 - 186.53030)
10.789 ± 0.322 16.88 (3) 2.12
Klang 1157 0.27382(0.17599 - 0.41134)
2.84803(1.22002 - 39.97898)
10.846 ± 0.330 9.89 (3) 0.66
Kuala Langat 1137 0.10325(0.04743 - 0.23894)
0.88398(0.31580 - 5414.29368)
10.570 ± 0.469 21.71 (3) 0.25
Kuala Selangor 1178 0.02732(0.01622 - 0.04812)
0.64951(0.20942 - 15.21424)
11.163 ± 0.388 9.23 (3) 0.07
Petaling 1154 1.29379(1.18453-1.40352)
6.45977(4.95458 - 9.56722)
9.985 ± 0.296 3.96 (3) 3.10
Sabak Bernam 1229 0.46913(0.26301 - 0.84814)
6.74790(2.28082 - 533.40694)
10.292 ± 0.259 12.73 (3) 1.12
Sepang 1157 1.12195(1.01124 - 1.21150)
4.60518(3.68997 - 6.26600)
2.984 ± 0.064 3.29 (3) 2.69
Malathion Bora-Bora 1211 0.04457(0.03997 - 0.04926)
0.31488(0.23102-0.49175)
10.518 ± 0.352 1.55 (3) –
Gombak 1253 0.03755(0.01837-0.07813)
0.66585(0.20644- 46.84840)
12.334 ± 0.366 16.25 (3) 0.84
Hulu Langat 1243 0.07612*
(0.07054-0.08123)0.24850(0.21014 -0.31523)
11.809 ± 0.865 1.57(3)
1.71
Hulu Selangor 1144 0.03156(0.01545-0.63627)
1.41518(0.34837-112.93681)
11.039 ± 0.319 10.52 (3) 0.71
Klang 1245 0.02275(0.01911- 0.02699)
0.57982(0.35655- 1.13705)
11.543 ± 0.379 1.45 (3) 0.51
Kuala Langat 1131 0.02024(0.01637-0.02532)
1.01909(0.55904-2.29810)
11.699 ± 0.315 2.25 (3) 0.45
Kuala Selangor 1187 0.02883(0.02454-0.03403)
0.62579(0.38428-1.23725)
11.065 ± 0.400 4.22 (3) 0.65
Petaling 1233 0.09891(0.07175-0.15168)
0.38919(0.20978-9.38667)
10.471 ± 0.749 15.87 (3) 2.22
Sabak Bernam 1261 0.02615(0.00836- 0.06870)
1.0313(0.21066-5518.11731)
10.948 ± 0.344 1.57 (3) 0.59
Sepang 1167 0.06025(0.05155-0.07078)
1.14543(0.71852-2.19390)
11.077 ± 0.365 4.45 (3) 1.35
Temephos Bora-Bora 1230 0.00168(0.00142-0.00199)
0.04121(0.02473-0.08533)
10.715 ± 0.434 2.88 (3) –
Gombak 1263 0.00063(0.00027 - 0.00200)
0.26847(0.02795 - 185.59243)
9.772 ± 0.832 8.23 (3) 0.38
Hulu Langat 1170 0.00072(0.00069 - 0.00075)
0.01576(0.01405 - 0.01849)
11.689 ± 1.258 5.17 (3) 0.43
Hulu Selangor 1129 0.00101(0.00082 - 0.00124)
0.04951(0.02551 - 0.12992)
10.021 ± 0.411 4.50 (3) 0.60
Klang 1162 0.00043(0.00037 - 0.00049)
0.00618(0.00401 - 0.01152)
10.472 ± 0.646 3.95 (3) 0.26
Kuala Langat 1135 0.00035(0.00026 - 0.00048)
0.11640(0.04518 - 0.45052)
10.423 ± 0.306 4.77 (3) 0.21
(continued on next page)
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
120
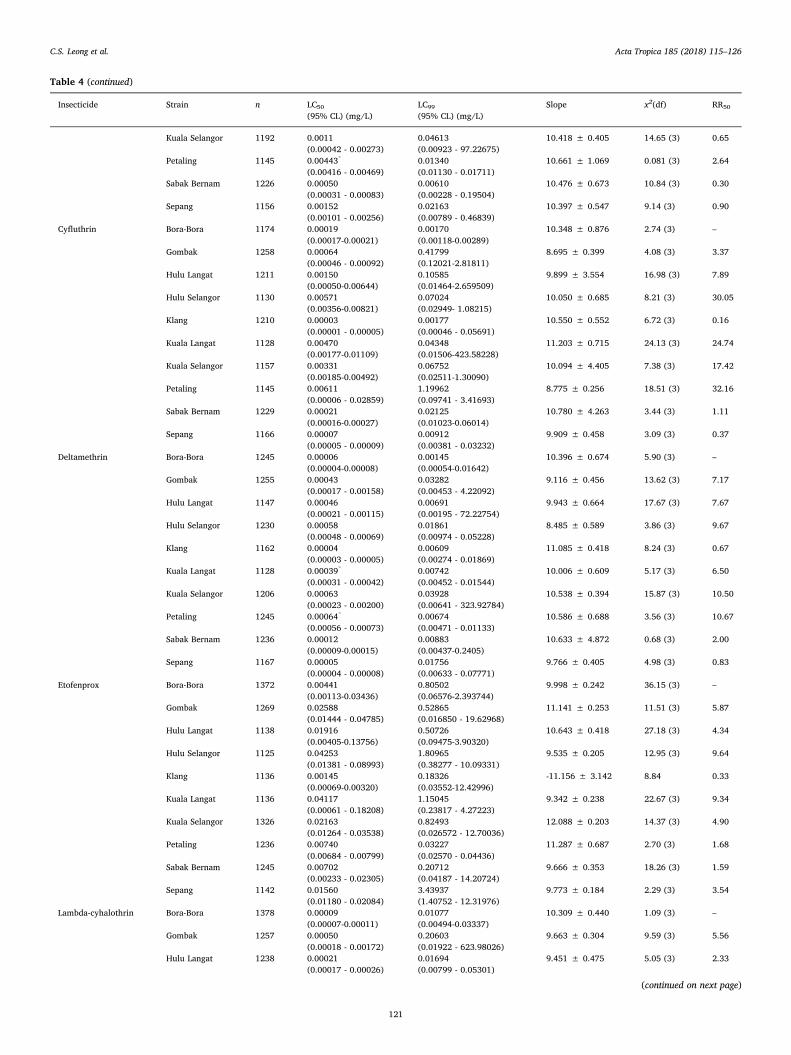
Table 4 (continued)
Insecticide Strain n LC50
(95% CL) (mg/L)LC99
(95% CL) (mg/L)Slope x2(df) RR50
Kuala Selangor 1192 0.0011(0.00042 - 0.00273)
0.04613(0.00923 - 97.22675)
10.418 ± 0.405 14.65 (3) 0.65
Petaling 1145 0.00443*
(0.00416 - 0.00469)0.01340(0.01130 - 0.01711)
10.661 ± 1.069 0.081 (3) 2.64
Sabak Bernam 1226 0.00050(0.00031 - 0.00083)
0.00610(0.00228 - 0.19504)
10.476 ± 0.673 10.84 (3) 0.30
Sepang 1156 0.00152(0.00101 - 0.00256)
0.02163(0.00789 - 0.46839)
10.397 ± 0.547 9.14 (3) 0.90
Cyfluthrin Bora-Bora 1174 0.00019(0.00017-0.00021)
0.00170(0.00118-0.00289)
10.348 ± 0.876 2.74 (3) –
Gombak 1258 0.00064(0.00046 - 0.00092)
0.41799(0.12021-2.81811)
8.695 ± 0.399 4.08 (3) 3.37
Hulu Langat 1211 0.00150(0.00050-0.00644)
0.10585(0.01464-2.659509)
9.899 ± 3.554 16.98 (3) 7.89
Hulu Selangor 1130 0.00571(0.00356-0.00821)
0.07024(0.02949- 1.08215)
10.050 ± 0.685 8.21 (3) 30.05
Klang 1210 0.00003(0.00001 - 0.00005)
0.00177(0.00046 - 0.05691)
10.550 ± 0.552 6.72 (3) 0.16
Kuala Langat 1128 0.00470(0.00177-0.01109)
0.04348(0.01506-423.58228)
11.203 ± 0.715 24.13 (3) 24.74
Kuala Selangor 1157 0.00331(0.00185-0.00492)
0.06752(0.02511-1.30090)
10.094 ± 4.405 7.38 (3) 17.42
Petaling 1145 0.00611(0.00006 - 0.02859)
1.19962(0.09741 - 3.41693)
8.775 ± 0.256 18.51 (3) 32.16
Sabak Bernam 1229 0.00021(0.00016-0.00027)
0.02125(0.01023-0.06014)
10.780 ± 4.263 3.44 (3) 1.11
Sepang 1166 0.00007(0.00005 - 0.00009)
0.00912(0.00381 - 0.03232)
9.909 ± 0.458 3.09 (3) 0.37
Deltamethrin Bora-Bora 1245 0.00006(0.00004-0.00008)
0.00145(0.00054-0.01642)
10.396 ± 0.674 5.90 (3) –
Gombak 1255 0.00043(0.00017 - 0.00158)
0.03282(0.00453 - 4.22092)
9.116 ± 0.456 13.62 (3) 7.17
Hulu Langat 1147 0.00046(0.00021 - 0.00115)
0.00691(0.00195 - 72.22754)
9.943 ± 0.664 17.67 (3) 7.67
Hulu Selangor 1230 0.00058(0.00048 - 0.00069)
0.01861(0.00974 - 0.05228)
8.485 ± 0.589 3.86 (3) 9.67
Klang 1162 0.00004(0.00003 - 0.00005)
0.00609(0.00274 - 0.01869)
11.085 ± 0.418 8.24 (3) 0.67
Kuala Langat 1128 0.00039*
(0.00031 - 0.00042)0.00742(0.00452 - 0.01544)
10.006 ± 0.609 5.17 (3) 6.50
Kuala Selangor 1206 0.00063(0.00023 - 0.00200)
0.03928(0.00641 - 323.92784)
10.538 ± 0.394 15.87 (3) 10.50
Petaling 1245 0.00064*
(0.00056 - 0.00073)0.00674(0.00471 - 0.01133)
10.586 ± 0.688 3.56 (3) 10.67
Sabak Bernam 1236 0.00012(0.00009-0.00015)
0.00883(0.00437-0.2405)
10.633 ± 4.872 0.68 (3) 2.00
Sepang 1167 0.00005(0.00004 - 0.00008)
0.01756(0.00633 - 0.07771)
9.766 ± 0.405 4.98 (3) 0.83
Etofenprox Bora-Bora 1372 0.00441(0.00113-0.03436)
0.80502(0.06576-2.393744)
9.998 ± 0.242 36.15 (3) –
Gombak 1269 0.02588(0.01444 - 0.04785)
0.52865(0.016850 - 19.62968)
11.141 ± 0.253 11.51 (3) 5.87
Hulu Langat 1138 0.01916(0.00405-0.13756)
0.50726(0.09475-3.90320)
10.643 ± 0.418 27.18 (3) 4.34
Hulu Selangor 1125 0.04253(0.01381 - 0.08993)
1.80965(0.38277 - 10.09331)
9.535 ± 0.205 12.95 (3) 9.64
Klang 1136 0.00145(0.00069-0.00320)
0.18326(0.03552-12.42996)
-11.156 ± 3.142 8.84 0.33
Kuala Langat 1136 0.04117(0.00061 - 0.18208)
1.15045(0.23817 - 4.27223)
9.342 ± 0.238 22.67 (3) 9.34
Kuala Selangor 1326 0.02163(0.01264 - 0.03538)
0.82493(0.026572 - 12.70036)
12.088 ± 0.203 14.37 (3) 4.90
Petaling 1236 0.00740(0.00684 - 0.00799)
0.03227(0.02570 - 0.04436)
11.287 ± 0.687 2.70 (3) 1.68
Sabak Bernam 1245 0.00702(0.00233 - 0.02305)
0.20712(0.04187 - 14.20724)
9.666 ± 0.353 18.26 (3) 1.59
Sepang 1142 0.01560(0.01180 - 0.02084)
3.43937(1.40752 - 12.31976)
9.773 ± 0.184 2.29 (3) 3.54
Lambda-cyhalothrin Bora-Bora 1378 0.00009(0.00007-0.00011)
0.01077(0.00494-0.03337)
10.309 ± 0.440 1.09 (3) –
Gombak 1257 0.00050(0.00018 - 0.00172)
0.20603(0.01922 - 623.98026)
9.663 ± 0.304 9.59 (3) 5.56
Hulu Langat 1238 0.00021(0.00017 - 0.00026)
0.01694(0.00799 - 0.05301)
9.451 ± 0.475 5.05 (3) 2.33
(continued on next page)
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
121
all sites.Five insecticides belonging to the pyrethroid group were tested and
of these the Klang, Sabak Bernam and Sepang strains showed 100%mortality to all five insecticides while the Gombak strain had 100%mortality to permethrin (Table 3). All other strains exhibited differentdegrees of resistance to pyrethroids with RR50> 30 for Hulu Selangorstrain against lambdcyhalothrin and cyfluthrin; the Petaling strainagainst cyfluthrin (Table 4). However, Spearman rank-order correlationindicated a significant correlation between resistance ratios of DDT and
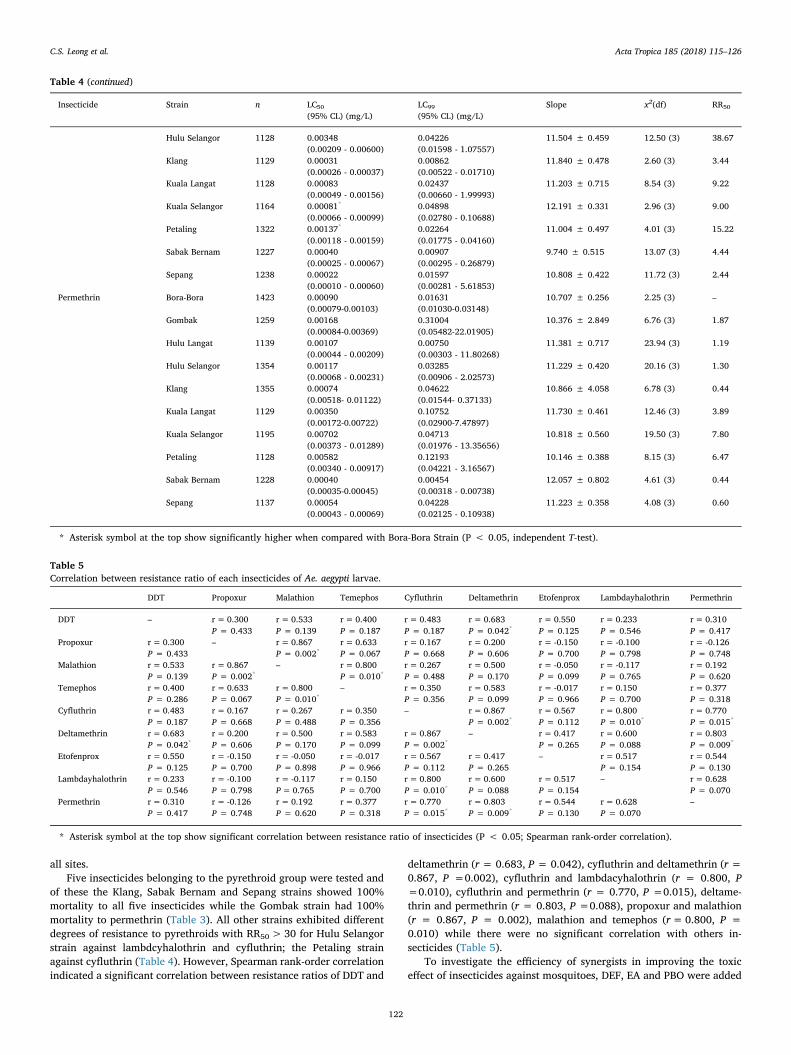
deltamethrin (r = 0.683, P = 0.042), cyfluthrin and deltamethrin (r =0.867, P =0.002), cyfluthrin and lambdacyhalothrin (r = 0.800, P=0.010), cyfluthrin and permethrin (r = 0.770, P =0.015), deltame-thrin and permethrin (r = 0.803, P =0.088), propoxur and malathion(r = 0.867, P = 0.002), malathion and temephos (r= 0.800, P =0.010) while there were no significant correlation with others in-secticides (Table 5).
To investigate the efficiency of synergists in improving the toxiceffect of insecticides against mosquitoes, DEF, EA and PBO were added
Table 4 (continued)
Insecticide Strain n LC50
(95% CL) (mg/L)LC99
(95% CL) (mg/L)Slope x2(df) RR50
Hulu Selangor 1128 0.00348(0.00209 - 0.00600)
0.04226(0.01598 - 1.07557)
11.504 ± 0.459 12.50 (3) 38.67
Klang 1129 0.00031(0.00026 - 0.00037)
0.00862(0.00522 - 0.01710)
11.840 ± 0.478 2.60 (3) 3.44
Kuala Langat 1128 0.00083(0.00049 - 0.00156)
0.02437(0.00660 - 1.99993)
11.203 ± 0.715 8.54 (3) 9.22
Kuala Selangor 1164 0.00081*
(0.00066 - 0.00099)0.04898(0.02780 - 0.10688)
12.191 ± 0.331 2.96 (3) 9.00
Petaling 1322 0.00137*
(0.00118 - 0.00159)0.02264(0.01775 - 0.04160)
11.004 ± 0.497 4.01 (3) 15.22
Sabak Bernam 1227 0.00040(0.00025 - 0.00067)
0.00907(0.00295 - 0.26879)
9.740 ± 0.515 13.07 (3) 4.44
Sepang 1238 0.00022(0.00010 - 0.00060)
0.01597(0.00281 - 5.61853)
10.808 ± 0.422 11.72 (3) 2.44
Permethrin Bora-Bora 1423 0.00090(0.00079-0.00103)
0.01631(0.01030-0.03148)
10.707 ± 0.256 2.25 (3) –
Gombak 1259 0.00168(0.00084-0.00369)
0.31004(0.05482-22.01905)
10.376 ± 2.849 6.76 (3) 1.87
Hulu Langat 1139 0.00107(0.00044 - 0.00209)
0.00750(0.00303 - 11.80268)
11.381 ± 0.717 23.94 (3) 1.19
Hulu Selangor 1354 0.00117(0.00068 - 0.00231)
0.03285(0.00906 - 2.02573)
11.229 ± 0.420 20.16 (3) 1.30
Klang 1355 0.00074(0.00518- 0.01122)
0.04622(0.01544- 0.37133)
10.866 ± 4.058 6.78 (3) 0.44
Kuala Langat 1129 0.00350(0.00172-0.00722)
0.10752(0.02900-7.47897)
11.730 ± 0.461 12.46 (3) 3.89
Kuala Selangor 1195 0.00702(0.00373 - 0.01289)
0.04713(0.01976 - 13.35656)
10.818 ± 0.560 19.50 (3) 7.80
Petaling 1128 0.00582(0.00340 - 0.00917)
0.12193(0.04221 - 3.16567)
10.146 ± 0.388 8.15 (3) 6.47
Sabak Bernam 1228 0.00040(0.00035-0.00045)
0.00454(0.00318 - 0.00738)
12.057 ± 0.802 4.61 (3) 0.44
Sepang 1137 0.00054(0.00043 - 0.00069)
0.04228(0.02125 - 0.10938)
11.223 ± 0.358 4.08 (3) 0.60
* Asterisk symbol at the top show significantly higher when compared with Bora-Bora Strain (P < 0.05, independent T-test).
Table 5Correlation between resistance ratio of each insecticides of Ae. aegypti larvae.
DDT Propoxur Malathion Temephos Cyfluthrin Deltamethrin Etofenprox Lambdayhalothrin Permethrin
DDT – r= 0.300P = 0.433
r=0.533P = 0.139
r= 0.400P = 0.187
r= 0.483P = 0.187
r= 0.683P = 0.042*
r= 0.550P = 0.125
r= 0.233P = 0.546
r= 0.310P = 0.417
Propoxur r= 0.300P = 0.433
– r= 0.867P = 0.002*
r= 0.633P = 0.067
r= 0.167P = 0.668
r= 0.200P = 0.606
r= -0.150P = 0.700
r= -0.100P = 0.798
r= -0.126P = 0.748
Malathion r= 0.533P = 0.139
r=0.867P = 0.002*
– r= 0.800P = 0.010*
r= 0.267P = 0.488
r= 0.500P = 0.170
r= -0.050P = 0.099
r= -0.117P = 0.765
r= 0.192P = 0.620
Temephos r= 0.400P = 0.286
r=0.633P = 0.067
r=0.800P = 0.010*
– r= 0.350P = 0.356
r= 0.583P = 0.099
r= -0.017P = 0.966
r= 0.150P = 0.700
r= 0.377P = 0.318
Cyfluthrin r= 0.483P = 0.187
r=0.167P = 0.668
r=0.267P = 0.488
r= 0.350P = 0.356
– r= 0.867P = 0.002*
r= 0.567P = 0.112
r= 0.800P = 0.010*
r= 0.770P = 0.015*
Deltamethrin r= 0.683P = 0.042*
r= 0.200P = 0.606
r=0.500P = 0.170
r= 0.583P = 0.099
r= 0.867P = 0.002*
– r= 0.417P = 0.265
r= 0.600P = 0.088
r= 0.803P = 0.009*
Etofenprox r= 0.550P = 0.125
r= -0.150P = 0.700
r= -0.050P = 0.898
r= -0.017P = 0.966
r= 0.567P = 0.112
r= 0.417P = 0.265
– r= 0.517P = 0.154
r= 0.544P = 0.130
Lambdayhalothrin r= 0.233P = 0.546
r= -0.100P = 0.798
r= -0.117P=0.765
r= 0.150P = 0.700
r= 0.800P = 0.010*
r= 0.600P = 0.088
r= 0.517P = 0.154
– r= 0.628P = 0.070
Permethrin r= 0.310P = 0.417
r= -0.126P = 0.748
r=0.192P = 0.620
r= 0.377P = 0.318
r= 0.770P = 0.015*
r= 0.803P = 0.009*
r= 0.544P = 0.130
r= 0.628P = 0.070
–
* Asterisk symbol at the top show significant correlation between resistance ratio of insecticides (P < 0.05; Spearman rank-order correlation).
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
122
in combination with each insecticide. The mortality rate of field ob-tained Ae. aegypti larvae against different combination of insecticidesand synergist are shown in Table 3. Generally, the synergists increasedthe mortality rate of different field strains and 100% mortality wasachieved with combination treatment of synergist(s) and insecticide forsome strains. Results show that PBO has significantly increased themortality rate of almost all strains of Ae. aegypti against pyrethroids and100% mortality were also achieved for several insecticides (DDT, te-mephos, cyfluthrin, deltamethrin, etofenprox, lambdacyhalothrin andpermethrin). Results showed that synergist(s) increased the mortality ofall strains of Ae. aegypti against all the tested insecticides. However,from the results, we observed that Hulu Selangor and Kuala Selangorstrains showed low mortality rate (< 80%) against cyfluthrin and del-tamethrin even when synergist(s) was added.
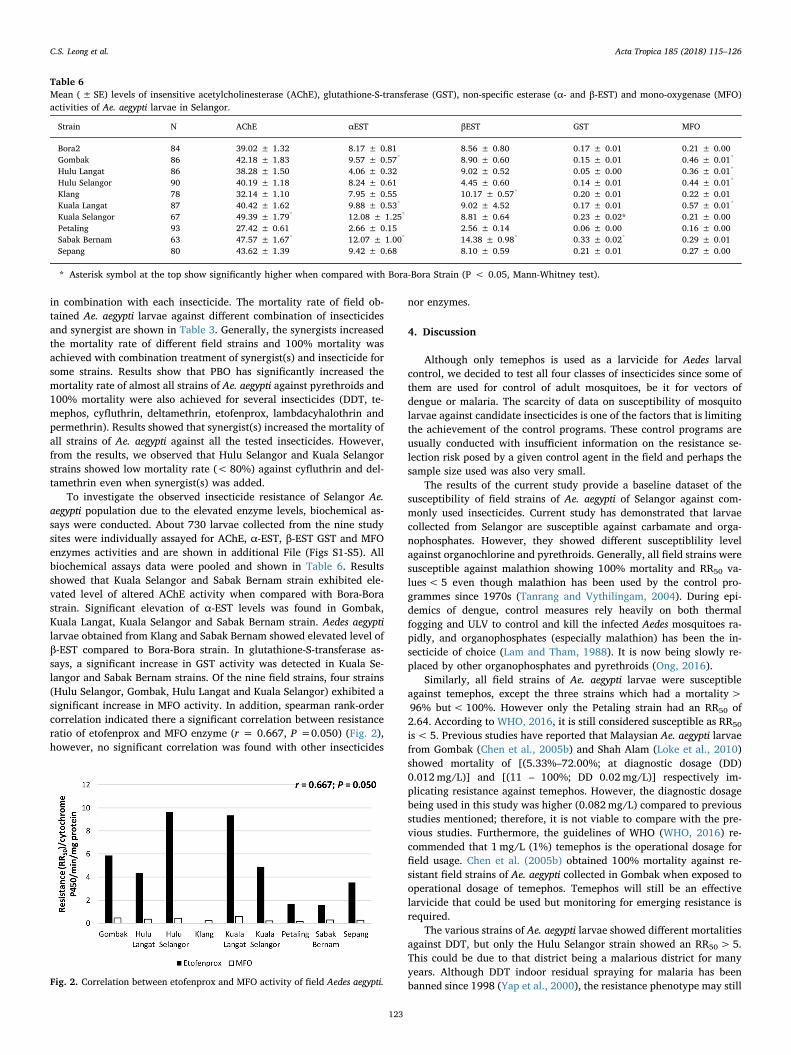
To investigate the observed insecticide resistance of Selangor Ae.aegypti population due to the elevated enzyme levels, biochemical as-says were conducted. About 730 larvae collected from the nine studysites were individually assayed for AChE, α-EST, β-EST GST and MFOenzymes activities and are shown in additional File (Figs S1-S5). Allbiochemical assays data were pooled and shown in Table 6. Resultsshowed that Kuala Selangor and Sabak Bernam strain exhibited ele-vated level of altered AChE activity when compared with Bora-Borastrain. Significant elevation of α-EST levels was found in Gombak,Kuala Langat, Kuala Selangor and Sabak Bernam strain. Aedes aegyptilarvae obtained from Klang and Sabak Bernam showed elevated level ofβ-EST compared to Bora-Bora strain. In glutathione-S-transferase as-says, a significant increase in GST activity was detected in Kuala Se-langor and Sabak Bernam strains. Of the nine field strains, four strains(Hulu Selangor, Gombak, Hulu Langat and Kuala Selangor) exhibited asignificant increase in MFO activity. In addition, spearman rank-ordercorrelation indicated there a significant correlation between resistanceratio of etofenprox and MFO enzyme (r = 0.667, P =0.050) (Fig. 2),however, no significant correlation was found with other insecticides
nor enzymes.
4. Discussion
Although only temephos is used as a larvicide for Aedes larvalcontrol, we decided to test all four classes of insecticides since some ofthem are used for control of adult mosquitoes, be it for vectors ofdengue or malaria. The scarcity of data on susceptibility of mosquitolarvae against candidate insecticides is one of the factors that is limitingthe achievement of the control programs. These control programs areusually conducted with insufficient information on the resistance se-lection risk posed by a given control agent in the field and perhaps thesample size used was also very small.
The results of the current study provide a baseline dataset of thesusceptibility of field strains of Ae. aegypti of Selangor against com-monly used insecticides. Current study has demonstrated that larvaecollected from Selangor are susceptible against carbamate and orga-nophosphates. However, they showed different susceptiblility levelagainst organochlorine and pyrethroids. Generally, all field strains weresusceptible against malathion showing 100% mortality and RR50 va-lues< 5 even though malathion has been used by the control pro-grammes since 1970s (Tanrang and Vythilingam, 2004). During epi-demics of dengue, control measures rely heavily on both thermalfogging and ULV to control and kill the infected Aedes mosquitoes ra-pidly, and organophosphates (especially malathion) has been the in-secticide of choice (Lam and Tham, 1988). It is now being slowly re-placed by other organophosphates and pyrethroids (Ong, 2016).
Similarly, all field strains of Ae. aegypti larvae were susceptibleagainst temephos, except the three strains which had a mortality>96% but< 100%. However only the Petaling strain had an RR50 of2.64. According to WHO, 2016, it is still considered susceptible as RR50
is< 5. Previous studies have reported that Malaysian Ae. aegypti larvaefrom Gombak (Chen et al., 2005b) and Shah Alam (Loke et al., 2010)showed mortality of [(5.33%–72.00%; at diagnostic dosage (DD)0.012mg/L)] and [(11 – 100%; DD 0.02mg/L)] respectively im-plicating resistance against temephos. However, the diagnostic dosagebeing used in this study was higher (0.082mg/L) compared to previousstudies mentioned; therefore, it is not viable to compare with the pre-vious studies. Furthermore, the guidelines of WHO (WHO, 2016) re-commended that 1mg/L (1%) temephos is the operational dosage forfield usage. Chen et al. (2005b) obtained 100% mortality against re-sistant field strains of Ae. aegypti collected in Gombak when exposed tooperational dosage of temephos. Temephos will still be an effectivelarvicide that could be used but monitoring for emerging resistance isrequired.
The various strains of Ae. aegypti larvae showed different mortalitiesagainst DDT, but only the Hulu Selangor strain showed an RR50> 5.This could be due to that district being a malarious district for manyyears. Although DDT indoor residual spraying for malaria has beenbanned since 1998 (Yap et al., 2000), the resistance phenotype may still
Table 6Mean (± SE) levels of insensitive acetylcholinesterase (AChE), glutathione-S-transferase (GST), non-specific esterase (α- and β-EST) and mono-oxygenase (MFO)activities of Ae. aegypti larvae in Selangor.
Strain N AChE αEST βEST GST MFO
Bora2 84 39.02 ± 1.32 8.17 ± 0.81 8.56 ± 0.80 0.17 ± 0.01 0.21 ± 0.00Gombak 86 42.18 ± 1.83 9.57 ± 0.57* 8.90 ± 0.60 0.15 ± 0.01 0.46 ± 0.01*
Hulu Langat 86 38.28 ± 1.50 4.06 ± 0.32 9.02 ± 0.52 0.05 ± 0.00 0.36 ± 0.01*
Hulu Selangor 90 40.19 ± 1.18 8.24 ± 0.61 4.45 ± 0.60 0.14 ± 0.01 0.44 ± 0.01*
Klang 78 32.14 ± 1.10 7.95 ± 0.55 10.17 ± 0.57* 0.20 ± 0.01 0.22 ± 0.01Kuala Langat 87 40.42 ± 1.62 9.88 ± 0.53* 9.02 ± 4.52 0.17 ± 0.01 0.57 ± 0.01*
Kuala Selangor 67 49.39 ± 1.79* 12.08 ± 1.25* 8.81 ± 0.64 0.23 ± 0.02* 0.21 ± 0.00Petaling 93 27.42 ± 0.61 2.66 ± 0.15 2.56 ± 0.14 0.06 ± 0.00 0.16 ± 0.00Sabak Bernam 63 47.57 ± 1.67* 12.07 ± 1.00* 14.38 ± 0.98* 0.33 ± 0.02* 0.29 ± 0.01Sepang 80 43.62 ± 1.39 9.42 ± 0.68 8.10 ± 0.59 0.21 ± 0.01 0.27 ± 0.00
* Asterisk symbol at the top show significantly higher when compared with Bora-Bora Strain (P < 0.05, Mann-Whitney test).
Fig. 2. Correlation between etofenprox and MFO activity of field Aedes aegypti.
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
123

remain in this mosquito population. Nazni et al. (2009) showed that theDDT resistance phenotype still persisted although the laboratory Ae.aegypti strain had been reared in the insectary for 1014 generations.However, it must be noted that DDT was only used in the malariacontrol programme in rural areas and was not used for dengue controlprogrammes. Since DDT and pyrethroids share the same mode of ac-tion/target site, thus, this occurrence of resistance against DDT may bedue to the application of pyrethroids based insecticides since 1996 untilnow (Selvi et al., 2010; Yap et al., 2000). It was shown that An. darlingiin South America was resistant against both DDT and pyrethroid in-secticides, although application of DDT had been stopped for 17 years.Thus, it has been suggested that this cross resistance was due to thecontinuous usage of pyrethroid based insecticides in the study site,since both insecticides shared the same target site – voltage-gated so-dium channels (Fonseca-González et al., 2009).
Among all the insecticides tested in current study, field collected Ae.aegypti was most resistant against pyrethroids (except Klang, SabakBernam and Sepang strains). However, pyrethroids are not used aslarvicides in Malaysia. The observed resistance is likely due to the usageof pyrethroid based adulticides, since pyrethroids is the class of in-secticides used in household insecticides products and also used intreatment of bednets and fogging for control of malaria and denguevectors, respectively (Nazni et al., 1998; Yap et al., 2000). Moreover,pyrethroids are one of the major class of insecticides used by pestcontrol industry. It is suggested that these pyrethroid based insecticidesproducts conferred the pyrethroid resistance detected in the currentstudy. This type of observed resistance among the larval populationssuggests that this phenotype acquired at the adult stage may be dis-played by both mature and immature mosquitoes (Chavasse et al.,1997). However, it should be stressed that this resistance profile inlarvae may not always be consistent in adult mosquitoes, and furtheradulticide bioassays are required for validation (Hemingway et al.,2002; Mebrahtu et al., 1997) and this would be conducted.
Generally, the synergists have increased the mortality of field col-lected Ae. aegypti against insecticides and thus indicates that esterases,monooxygenase and glutathione-S-transferase play important role forDDT and pyrethroid resistance detected in the current study. It wasfound that PBO increased the toxicity of all pyrethroids compared toother synergists. PBO is known as MFO inhibitor, and MFO has beendocumented to play a role in pyrethroid resistance (Hasan et al., 2016;Kasai et al., 2014). Similarly, our study also suggests the involvement ofMFO for the resistance of some field strains towards pyrethroids, wherePBO have increased the mortality of some field collected Ae. aegypti to100%. However, not all field strains have achieved susceptible levelwith the addition of PBO. Furthermore, there were six field strains(Gombak, Hulu Langat, Hulu Selangor, Kuala Langat, Kuala Selangorand Petaling) which showed resistance for two or more different pyr-ethroids and this could be due to the cross-resistance between thepyrethroid groups. Thus, this further suggests that involvement of morethan one mechanism conferring insecticide resistance. Furthermore,cross resistance between propoxur and malathion, malathion and te-mephos, DDT and pyrethroid and within pyrethroids were detected inthe current study. Cross resistance between DDT and pyrethroids hasbeen commonly reported in Thailand (Prapanthadara et al., 2002),Colombia (Fonseca-González et al., 2009), Vietnam (Kawada et al.,2009) and also in Malaysia (Ishak et al., 2015). The mechanisms in-volved in this cross-resistance is mostly related to kdr mutation involtage-gated sodium channel (Hemingway et al., 2002; Hemingwayet al., 2004). Although cross resistance was detected for propoxur-malathion and malathion-temephos, however, all field strain Ae. aegyptilarvae were susceptible against all three insecticides mentioned. Thiscross resistance emerges from the mutation of target site insensitivity ofAChE (Liu et al., 2004).
Biochemical assays have detected elevated levels of ESTs in moststrains. Esterases detoxification is reported as one of the main reasonsof resistance in Ae. aegypti populations due to organophosphates (Bisset
et al., 2003; Nauen, 2007). However, the current study shows the fieldstrains tested were susceptible against temephos and malathion. SinceDEF have significantly increased the mortality of certain field strains ofAe. aegypti larvae when exposed to pyrethroids perhaps elevated es-terases may be involved in pyrethroid resistance.
Insensitive AChE activity of field obtained Ae. aegypti larvae wereminimal with only Sabak Bernam and Kuala Selangor strains showingsignificantly elevated insensitive AChE activity. Cuamba et al. (2010)suspected the hydrolysis of propoxur via others enzymes (such as ESTs)have decreased the level of insensitive AChE in field mosquitoes. Per-haps the concentration of propoxur for AChE inhibition was decreasedas the biochemical assays were conducted using total enzyme extractionon mosquitoes (Koou et al., 2014).
DDT resistance is associated with elevated GST level (Aïzoun et al.,2014; Rodríguez et al., 2007) and this was observed in both the KualaSelangor and Sabak Bernam strains. This elevated GST level maybe dueto the large-scale use of pyrethroids in public health activities as bothDDT and pyrethroids are especially designed to target the voltage-gatedsodium channel of arthropods (Hemingway et al., 2004).
Multiple insecticides resistance implicated that more than one me-chanisms are involved in insecticides resistance. As a matter of fact,evolution/mutation of multiple isolates is not a new phenomenon and isbecoming a serious issue worldwide. Results of the current study do notsupport the hypothesis that enzyme activities are corresponding to in-secticide susceptibility status of Ae. aegypti due to the lack of correlationbetween resistance level and enzyme activities. Yet, there are studiessuggesting that enzyme activities are not axiomatically correlated withtoxicological changes (Montella et al., 2012; Siegfried and Scott, 1992).This suggests that the occurrence of this incidence might be due to theevolution/mutation in voltage-gated sodium channel which is com-monly found in pyrethroids resistant Ae. aegypti (Kasai et al., 2014;Stenhouse et al., 2013; Yanola et al., 2011). Therefore, we propose toconduct molecular work to further uncover the involvement of targetsite insensitivity mechanisms in Ae. aegypti.
In summary, insecticide(s) resistance of Ae. aegypti larvae have beendetected in the current study. Therefore, new strategy for vector controlshould be redesign with consideration of others aspect such as earlierprevention of outbreak (Lau et al., 2017), remodel the methods withdata gather from research and modelling on prediction of outbreak(Benelli et al., 2018). For example, Lau et al. (2017) have reported thatthe data obtained from the combination usages of Gravid OvipositingSticky (GOS) traps and NS1 kit provided earlier detection of dengueoutbreak in a premise in Malaysia. Thus, control measures can takeaction before the dengue outbreak. This control measures fits into OneHealth strategy, where all data collected in small scales can be used andapplied in larger scales where disease outbreak can be preventing(Benelli et al., 2018).
5. Conclusion
Overall, dengue outbreak study sites have shown higher resistancecompared to non-dengue outbreak areas. However, it was found thatSepang strain Ae. aegypti was susceptible against all tested insecticides.Results of current study showed that Ae. aegypti in Selangor are sus-ceptible to organphosphates and carbamate. Temephos remains as aneffective larvicide. It is important to note that Ae. aegypti have devel-oped resistance to pyrethroids and is now one of the main class of in-secticides used in vector control. The choice of alternative insecticidesBacillus thuringiensis israelensis (Bti) or insect growth regulator (IGR)should be recommended for future public health programs. The exactmechanism involved for DDT and pyrethroids resistance in currentstudy remains to be explored to develop alternative measure to manageresistance in mosquitoes. The outcome of current study provides in-formation on resistance and cross resistance pattern occurring inSelangor Ae. aegypti larvae. Thus, control programme should be awareof cross resistance to the same/related active ingredients application on
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
124

the field. At the same time to ensure the success of the control pro-gramme, new developments to overcome resistance is needed and alsonovel strategies should be designed to prevent/minimize the spread andevolution of resistance. Perhaps detection of virus in mosquitoes will bemore proactive strategy as control measures can be instituted beforecases occur.
Acknowledgments
This study was financially supported by the University of MalayaIPP grant (PG004-2015A). This research was regulated by theInstitutional Animal Care and Use Committee (IACUC no.: 20150407/PARA/R/MBK) was obtained before the commencement of the study.
Appendix A. Supplementary data
Supplementary material related to this article can be found, in theonline version, at doi:https://doi.org/10.1016/j.actatropica.2018.05.008.
References
Abbott, W., 1925. A method of computing the effectiveness of an insecticides. J. Econ.Entomol. 18 (2), 265–267.
Aïzoun, N., Aïkpon, R., Gnanguenon, V., Azondekon, R., et al., 2014. Dynamics of in-secticide resistance and effect of synergists piperonyl butoxide (PBO), SSS-tributyl-phosphorotrithioate (DEF) and ethacrynic acid (ETAA or EA) on permethrin, delta-methrin and dichlorodiphenyltrichloroethane (DDT) resistance in two Anophelesgambiae s.l. Populations from Southern Benin, West Africa. J. Parasit. Vector Biol. 6,1–10.
Araújo, A.P., Diniz, D.F.A., Helvecio, E., De Barros, R.A., De Oliveira, C.M.F., Ayres,C.F.J., de Melo-Santos, M.A.V., Regis, L.N., Silva-Filha, M.H.N.L., 2013. The sus-ceptibility of Aedes aegypti populations displaying temephos resistance to Bacillusthuringiensis israelensis: a basis for management. Parasit. Vectors 6, 297.
Attar, N., 2016. ZIKA virus circulates in new regions. Nat. Rev. Microbiol. 14 (62). http://dx.doi.org/10.1038/nrmicro.2015.28.
Benelli, G., 2015. Research in mosquito control: current challenges for a brigther future.Parasitol. Res. 114, 2801–2805.
Benelli, G., Duggan, M.F., 2018. Social and ecological dynamics facing the one healthperspective. Acta Trop. 182, 80–91. http://dx.doi.org/10.1016/j.actatropica.2018.02.015.
Benelli, G., Beier, J., 2017. Current vector control challenges in the fight agaisnt malaria.Acta Trop. 174, 91–96. http://dx.doi.org/10.1016/j.actatropica.2017.06.028.
Benelli, G., Maggi, F., Payela, R., Muruga, K., Govindarajan, M., Vaseeharan, B.,Petrellim, R., Cappellacci, L., Kumar, S., Hofer, A., Youssefi, M.R., Alarfaj, A.A.,Hwang, J.S., Higuchi, A., 2018. Mosquito control with green nanopesticides: towardsthe one health approach? A review of non-target effects. Environ. Sci. Poll. Res.http://dx.doi.org/10.1007/s11356-017-9752-4.
Benelli, G., Romano, D., 2017. Mosquito vectors of Zika virus. Entomol. Gen. 36 (4),309–318. http://dx.doi.org/10.1127/entomologia/2017/0496.
Benelli, G., Mehlhorn, H., 2016. Declining malaria, rising of dengue and Zika virus: in-sights for mosquito vectorl control. Parasitol. Res. 115, 1747–1754.
Bisset, J., Magdalena, R.M., Fernández, D., Pérez, O., 2003. Status of resistance to in-secticides and resistance mechanisms in larvae from Playa municipality collectedduring the intensive campaign against Aedes aegypti in Havana City, 2001-2002.Rev. Cubana. Med. Trop. 56, 61–66.
Brogdon, W., Janet, C., 1997. Heme peroxidase activity measured in single mosquitoesidentifies individuals expressing an elevated oxidase for insecticide resistance. J. Am.Mosq. Control Assoc. 1 (3), 233–237.
Brown, A.W.A., 1958. Insecticide Resistance in Arthropods.World Health OrganizationPublication 244 Columbia University Press, New York.
Chavasse, D.C., Yap, H.H., WHO, 1997. Chemical Methods for the Control of Vectors andPests of Public Health Importance. WHO/CTD/WHOPES/97.2. 129.
Chen, C., Nazni, W., Lee, H., Seleena, B., Sofian-Azirun, M., 2008. Biochemical detectionof temephos resistance in Aedes (Stegomyia) aegypti (Linnaeus) from dengue-en-demic areas of Selangor state, Malaysia. Proc. ASEAN Congr. Trop. Med. Parasitol. 3,6–20.
Chen, C., Nazni, W., Lee, H., Sofian-Azirun, M., 2005a. Susceptibility of Aedes aegypti andAedes albopictus to temephos in four study sites in Kuala Lumpur City center andSelangor State, Malaysia. Trop. Biomed. 22, 207–216.
Chen, C., Nazni, W., Lee, H., Sofian-Azirun, M., 2005b. Weekly variation on susceptibilitystatus of Aedes mosquitoes against temephos in Selangor, Malaysia. Trop. Biomed.22, 195–206.
Cheong, W., Rudnick, A., Lin, T., 1986. The vectors of dengue and dengue hemorrhagicfevers in Malaysia. Dengue fever studies in Malaysia. Inst. Med. Res. Bull. 23,155–167.
Cuamba, N., Morgan, J.C., Irving, H., Steven, A., Wondji, C.S., 2010. High level of pyr-ethroid resistance in an Anopheles funestus population of the Chokwe District inMozambique. PloS One 5, e11010.
Department of Statistics, M., 2014. Malaysia, Monthly manufactruring Statistics. https://www.dosm.gov.my/v1/index.php.
Duong, V., Lambrechts, L., Paul, R.E., Ly, S., Long, K.C., Huy, R., Tarantola, A., Scott,T.W., Sakuntabhai, A., Buchy, P., 2015. Asymptomatic humans transmit dengue virusto mosquitoes. Proc. Natl. Acad. Sci. U. S. A. 112 (47), 14688–14693. http://dx.doi.org/10.1073/pnas.1508114112.
Finney, D.J., 1947. Probit Analysis; a Statistical Treatment of the Sigmoid ResponseCurve. University Press, United State.
Fonseca-González, I., Quiñones, M.L., McAllister, J., Brogdon, W.G., 2009. Mixed-func-tion oxidases and esterases associated with cross-resistance between DDT andlambda-cyhalothrin in Anopheles darlingi root 1926 populations from Colombia.Mem.Inst. Oswaldo. Cruz 104, 18–26.
Garcia, G.P., Flores, A.E., Fernandez-Salas, I., Saavedra-Rodriguez, K., Reyes-Solis, G.,Lozano-Fuentes, S., Guillermo Bond, J., Casas-Martinez, M., Ramsey, J.M., Garcia-Rejon, J., Dominguez-Galera, M., Ranson, H., Hemingway, J., Eisen, L., Black, I.W.,2009. Recent rapid rise of a permethrin knock down resistance allele in Aedes aegyptiin Mexico. PLoS Negl. Trop. Dis. 3, e531.
Grisales, N., Poupardin, R., Gomez, S., Fonseca-Gonzalez, I., Ranson, H., Lenhart, A.,2013. Temephos resistance in Aedes aegypti in Colombia compromises dengue vectorcontrol. PLoS Negl. Trop. Dis. 7, e2438.
Hasan, H.A., Jaal, Z., Ranson, H., McCall, P., 2016. Pyrethroid and organophosphatesusceptibility status of Aedes aegypti (Linnaeus) and Aedes albopictus (Skuse) inPenang Malaysia. Int. J. Entomol. Res. 3, 91–95.
Hemingway, J., Brogdon, W., 1998. Techniques to detect insecticide resistance me-chanisms. WHO/CDS/CPC/MAL/98.6.
Hemingway, J., Field, L., Vontas, J., 2002. An overview of insecticide resistance. Sci. 298,96–97.
Hemingway, J., Hawkes, N.J., McCarroll, L., Ranson, H., 2004. The molecular basis ofinsecticide resistance in mosquitoes. Insect Biochem. Mol. Biol. 34, 653–665.
Ishak, I.H., Jaal, Z., Ranson, H., Wondji, C.S., 2015. Contrasting patterns of insecticideresistance and knockdown resistance (kdr) in the dengue vectors Aedes aegypti andAedes albopictus from Malaysia. Parasit. Vectors 8, 181.
Kasai, S., Komagata, O., Itokawa, K., Shono, T., Ng, L.C., Kobayashi, M., Tomita, T., 2014.Mechanisms of pyrethroid resistance in the dengue mosquito vector, Aedes aegypti:target site insensitivity, penetration, and metabolism. PLoS Negl. Trop. Dis. 8, e2948.
Kawada, H., Higa, Y., Komagata, O., Kasai, S., Tomita, T., Yen, N.T., Loan, L.L., Sánchez,R.A., Takagi, M., 2009. Widespread distribution of a newly found point mutation involtage-gated sodium channel in pyrethroid-resistant Aedes aegypti populations inVietnam. PLoS Negl. Trop. Dis. 3, e527.
Koou, S.Y., Chong, C.S., Vythilingam, I., Ng, L.C., Lee, C.Y., 2014. Pyrethroid resistance inAedes aegypti larvae (Diptera: Culicidae) from Singapore. J. Med. Entomol 51,170–181.
Lam, W., Tham, A., 1988. A field evaluation of the effectiveness of ULV application ofmalathion 96% technical grade and sumithion L-40S against Aedes aegypti(Linnaeus) and Aedes albopictus (Skuse) in Ipoh Municipality, Perak, Malaysia. Trop.Biomed. 5, 81–88.
Lau, S.M., Chua, T.H., Sulaiman, W., Joanne, S., Lim, Y.A., Sekaran, S.D., Chimma, K.,Venugopalan, B., Vythilingam, I., 2017. A new paradigm for Aedes spp. Surveillanceusing gravid ovipositing sticky trap and NS1 antigen test kit. Parast. Vectors 10,151–160. http://dx.doi.org/10.1186/s13071-017-2091-y.
Lee, H., Chen, C., Masri, S.M., Chiang, Y., Chooi, K., Benjamin, S., 2008. Impact of lar-viciding with a Bacillus thuringiensis israelensis formulation, Vectobac WG®, ondengue mosquito vectors in a dengue endemic site in Selangor state, Malaysia.Southeast Asian J. Trop. Med. Public Health 39, 601–609.
Lima, E.P., Paiva, M.H., de Araujo, A.P., da Silva, E.V., da Silva, U.M., de Oliveira, L.N.,Santana, A.E., Barbosa, C.N., de Paiva Neto, C.C., Goulart, M.O., Wilding, C.S., Ayres,C.F., de Melo Santos, M.A., 2011. Insecticide resistance in Aedes aegypti populationsfrom Ceara. Braz. Parasit. Vectors 4, 5.
Liu, H., Cupp, E.W., Micher, K.M., Guo, A., Liu, N., 2004. Insecticide resistance and cross-resistance in Alabama and Florida strains of Culex quinquefasciatus. J. Med. Entomol.41 (3), 408–413.
Loke, S.R., Wei Ann, A., Ahmad, N., Azirun, M., 2012. Insecticide susceptibility status offield-collected Aedes (Stegomyia) aegypti (L.) From a dengue endemic site in ShahAlam, Selangor, Malaysia. Southeast. Asian J. Trop. Med. Public. Health 43, 34.
Loke, S.R., Andy-Tan, W., Benjamin, S., Lee, H., Sofian-Azirun, M., 2010. Susceptibility offield-collected Aedes aegypti (L.)(Diptera: Culicidae) to Bacillus thuringiensis israe-lensis and temephos. Trop. Biomed. 27, 493–503.
Marcombe, S., Mathieu, R.B., Pocquet, N., Riaz, M.A., Poupardin, R., Selior, S., Darriet, F.,Reynaud, S., Yebakima, A., Corbel, V., David, J.P., Chandre, F., 2012. Insecticideresistance in the dengue vector Aedes aegypti from Martinique: distribution, me-chanisms and relations with environmental factors. PLoS One 7, e30989.
Mebrahtu, Y.B., Norem, J., Taylor, M., 1997. Inheritance of larval resistance to perme-thrin in Aedes aegypti and association with sex ratio distortion and life history var-iation. Am. J. Trop. Med. Hyg. 56, 456–465.
Mehlhorn, H., 2016. Nanaoparticles in the Fight Agaisnt Parasites. Parasitol. Res.Monographs 8 Springer, Berlin.
MOH, 2016. Indicators for Monitoring and Evaluation of Strategy health for All. http://www.moh.gov.my/images/gallery/publications/md/hi/hi_2009.pdf (accessed 12.12.17).
Montella, I.R., Schama, R., Valle, D., 2012. The classification of esterases: an importantgene family involved in insecticide resistance-A review. Mem. Inst. Oswaldo. Cruz.107, 437–449.
Nauen, R., 2007. Insecticide resistance in disease vectors of public health importance.Pest Manag. Sci. 63, 628–633.
Nazni, W., Lee, H., Sa’diyah, I., 1998. Rate of resistance development in wild Culexquinquefasciatus (say) selected by malathion and permethrin. Southeast. Asian J.
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
125

Trop. Med. Public. Health 29, 849–855.Nazni, W., Selvi, S., Lee, H., Sadiyah, I., Azahari, H., Derric, N., Vasan, S., 2009.
Susceptibility status of transgenic Aedes aegypti (L.) Against insecticides. DengueBull. 33. 124–129.
Ocampo, C.B., Salazar-Terreros, M.J., Mina, N.J., McAllister, J., Brogdon, W., 2011.Insecticide resistance status of Aedes aegypti in 10 localities in Colombia. Acta Trop.118, 37–44.
Ong, S.Q., 2016. Dengue vector control in Malaysia: a review for current and alternativestrategies. Sains Malays. 45, 777–785.
Prapanthadara, La, Promtet, N., Koottathep, S., Somboon, P., Suwonkerd, W., McCarroll,L., Hemingway, J., 2002. Mechanisms of DDT and permethrin resistance in Aedesaegypti from Chiang Mai, Thailand. Dengue Bull. 26, 185–189.
Rodríguez, M.M., Bisset, J.A., Fernández, D., 2007. Levels of insecticide resistance andresistance mechanisms in Aedes aegypti from some latin American countries. J. Am.Mosq. Control Assoc. 23, 420–429.
Saelim, V., Brogdon, W.G., Rojanapremsuk, J., Suvannadabba, S., 2005. Bottle and bio-chemical assays on temephos resistance in Aedes aegypti in Thailand. Southeast.Asian J. Trop. Med. Public. Health 36, 417.
Selvi, S., Edah, M., Nazni, W., Lee, H., Tyagi, B., Sofian-Azirun, M., Azahari, A., 2010.Insecticide susceptibility and resistance development in malathion selected Aedesalbopictus (Skuse). Trop. Biomed. 27, 534–550.
Siegfried, B.D., Scott, J.G., 1992. Biochemical characterization of hydrolytic and oxida-tive enzymes in insecticide resistant and susceptible strains of the German cockroach(Dictyoptera: Blattellidae). J. Econ. Entomol. 85, 1092–1098.
Stenhouse, S.A., Plernsub, S., Yanola, J., Lumjuan, N., Dantrakool, A., Choochote, W.,Somboon, P., 2013. Detection of the V1016G mutation in the voltage-gated sodiumchannel gene of Aedes aegypti (Diptera: Culicidae) by allele-specific PCR assay, andits distribution and effect on deltamethrin resistance in Thailand. Parasit. Vectors
6 (1).Tanrang, Y., Vythilingam, I., 2004. Field trial to determine the efficacy of pyrethroid
Fendona 10 SC@ application using ultra- low- volume for the control of Aedesmosquitoes. Trop. Biomed. 21.
Wan-Norafikah, O., Nazni, W.A., Lee, H.L., Zainol-Ariffin, P., Sofian-Azirun, M., 2010.Permethrin resistance in Aedes aegypti (Linnaeus) collected from Kuala Lumpur,Malaysia. J. Asia. Pac. Entomol 1 (3), 175–182.
WHO, 1981. Instructions for determining the susceptibility or resistance of mosquitolarvae to insecticides. WHO/VBC/81/807.
WHO, 2003. Guidelines For Dengue Surveillance And Mosquito Control, 2nd Edition.WHO Regional offiece for the Western Pacific, Manila.
WHO, 2005. Guidelines for laboratory and field testing of mosquito larvicides. WHO/CDS/ WHOPES/GCDPP/2005.13.
WHO, 2012. Handbook for Integrated Vector Management. WHO/HTM/NTD/VEM/2012.3.
WHO, 2016. Monitoring and managing insecticide resistance in Aedes mosquito popu-lations: Interim guidance for entomologists. WHO/ZIKV/VC/16.1.
WHO, 2017. Dengue and severe dengue, April 2017 ed. WHO factsheet. http://www.who.int/mediacentre/factsheets/fs117/en/.
Yanola, J., Somboon, P., Walton, C., Nachaiwieng, W., Somwang, P., Prapanthadara, La,2011. High‐throughput assays for detection of the F1534C mutation in the volta-ge‐gated sodium channel gene in permethrin‐resistant Aedes aegypti and the dis-tribution of this mutation throughout Thailand. Trop. Med. Int. Health (16),501–509.
Yap, H.H., Lee, Y., Zairi, J., 2000. Chemical control of mosquitoes. In: Ng, F., Yong, H.(Eds.), Mosquitoes and Mosquito-Borne Diseases: Biology, Surveillance, Control,Personal and Public Protection Measures. Academy of Sciences Malaysia, Malaysia,pp. 197–210.
C.S. Leong et al. Acta Tropica 185 (2018) 115–126
126





























