Embed Size (px)
Citation preview

Available online at www.sciencedirect.com
siology 67 (2008) 114–123www.elsevier.com/locate/ijpsycho
International Journal of Psychophy
Affective visual event-related potentials: Arousal, valence, and repetitioneffects for normal and distorted pictures
Bella Rozenkrants a, Jonas K. Olofsson b, John Polich c,⁎
a Department of Psychology, University of California, San Diego, La Jolla, CA, USAb Department of Psychology, Umeå University, Umeå, Sweden
c Molecular and Integrative Neurosciences Department, The Scripps Research Institute, La Jolla, California, USA
Received 2 May 2007; received in revised form 15 October 2007; accepted 23 October 2007
Available online 4 November 2007
Abstract
Event-related potentials (ERPs) were used to assess arousal (low, high), valence (negative, positive), and stimulus repetition effects for normaland distorted images from the International Affective Pictures System (IAPS). Distorted stimuli were constructed by dividing each image intosmall squares and rearranging the segments randomly to produce a “scrambled” picture. Event-related potentials (ERPs) were elicited bypresenting the normal and scrambled images as target stimuli, with a repeated visual pattern used as the standard stimulus. Participants (N=32)were instructed to press a button to the targets and ignore the standard. Stimulus repetition effects were assessed by presenting each stimulus twicein the normal and scrambled condition. High-arousal stimuli yielded larger late positive components for both the normal and scrambled pictures.No overall valence effects were obtained, but arousal and valence influenced component amplitudes for middle-latency ERPs from the scrambledstimuli. For the normal pictures, stimulus repetition was associated with increased component amplitudes for all potentials and decreased RTs ofall affective categories. For the scrambled pictures, no repetition changes were obtained. The findings suggest that stimulus arousal levelcontributes more than valence to affective ERP measures for normal as well as perceptually distorted pictures. Stimulus repetition engagesmemory for previous normal picture items but is not influenced by affective category. Theoretical implications are discussed.© 2007 Elsevier B.V. All rights reserved.
Keywords: International Affective Picture System (IAPS); Event-related potentials (ERPs); P300; Arousal; Valence; Repetition
1. Introduction
The perceptual system utilizes information gained throughprevious exposure to affective events thereby facilitatingsubsequent processing of similar stimuli. As event-relatedpotentials (ERPs) can provide fine-grain temporal resolution ofthe underlying mechanisms, this neuroimaging method hasbecome highly useful for assessing affective neural processing(Bradley et al., 1992; Dolcos and Cabeza, 2002; Ito et al., 1998;
⁎ Corresponding author. Cognitive Electrophysiology Laboratory, Molecularand Integrative Neurosciences Department, The Scripps Research Institute,TPC-10, 10550 North Torrey Pines Road, La Jolla, CA 92037, USA. Tel.: +1858 784 8176; fax: +1 858 784 9293.
E-mail address: [email protected] (J. Polich).
0167-8760/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.ijpsycho.2007.10.010
Lang et al., 1993, 1999). Although affective ERPs can begenerated automatically by motivationally relevant picturemotifs (appetitive, fearful, sexual), emotional stimuli alsomodulate ERPs even if the image content is not readilyperceived (Codispoti et al., 2006; Schupp et al., 2006). Thepresent study employs a stimulus repetition paradigm tocharacterize these issues by comparing affective pictures with“scrambled” image controls.
1.1. ERPs from affective picture stimuli
The International Affective Picture System (IAPS) often isused to provide visual images for assessing emotional outcomes(Lang et al., 1999). These pictures are rated for valence(unpleasant to pleasant) and arousal (calm to exciting) level, andthese dimensions have been related to ERP modulations. The

115B. Rozenkrants et al. / International Journal of Psychophysiology 67 (2008) 114–123
temporal course of valence and arousal effects differs, however,as valence influences are observed for early (100–250 ms)whereas arousal influences occur for later (250–850 ms)components (Codispoti et al., 2007; Olofsson et al., in press).Similar outcomes have been obtained from a wide range oftasks, and as affectively potent stimuli modulate ERPs evenwhen presented subconsciously these effects appear to occurautomatically (Bernat et al., 2001; Cuthbert et al., 2000;Delplanque et al., 2006; Roschmann and Wittling, 1992).
Affective ERP effects have been obtained from passiveviewing, affect discrimination procedures, and for imagespresented as distracting or target stimuli in an oddball task(Delplanque et al., 2004, 2005; Keil et al., 2002; Mini et al.,1996; Schupp et al., 2000). In general, negative affectivevalence produces stronger ERP modulation than positiveaffective valence pictures (Cacioppo et al., 1999; Crawfordand Cacioppo, 2002; Öhman and Mineka, 2001). This“negativity-bias” may reflect rapid amygdala processing ofaversive information (LeDoux, 1989, 1995; Morris et al., 1998;Öhman and Soares, 1998). Moreover, these early affectiveERPs appear related to affective disposition such as trait anxietyand trait fearfulness (Dien, 1999), whereas late affective ERPsare sensitive to individual variation from emotional reappraisaland evaluation strategies (Hajcak and Nieuwenhuis, 2006,Moser et al., 2006).
In addition, when a P300 component from an affectivestimulus is elicited, amplitudes are larger over the parietalcortex and increase with increases in arousal level. Hence,emotional arousal may amplify activity in cortical structuresthat are normally engaged for target processing (Polich, 2007;Sabatinelli et al., 2007). When arousal level across stimuli iswell controlled valence can modulate P300 from frontal sites(Cano et al., in press; Conroy and Polich, 2007; Delplanqueet al., 2005, 2006), although it is unclear whether these effectsconsistently obtain when both factors are manipulated withinthe same session (Cuthbert et al., 2000). Despite evidence forERP modulations from picture valence and arousal (Delplanqueet al., 2004, 2006; Olofsson and Polich, 2007; Schupp et al.,2006), these fundamental affective factors are not typicallyvaried systematically. For example, positive/negative valenceswith generally high-arousal level often are compared withneutral valence stimuli that are low-arousing in a fashion thatconfounds their ERP assessment by differential stimulusprobability.
1.2. Stimulus repetition effects
When a stimulus is presented repeatedly, processing changescan be expected because of variation in stimulus novelty,priming mechanisms, and habituation. Recognition memoryERP studies using neutral and affective stimuli have foundincreased amplitudes for late positive components when astimulus reoccurs within the stimulus series (Bentin et al., 1992;Friedman, 1990; Olofsson and Polich, 2007; Rugg, 1990;Segalowitz et al., 1997; Smith and Halgren, 1989). Severalaffective studies have presented stimuli multiple times butcollapsed over presentations to obtain their ERP averages, so
that repetition effects cannot be assessed directly (Aftanas et al.,2001; Brazdil et al., 2003; Carretie et al., 2004a, 2004b; Itoet al., 1998; Keil et al., 2001; Schupp et al., 2000, 2004).
Olofsson and Polich (2007) found that successive repetitionof affective target stimuli within an oddball sequence producedlarger positive amplitudes for pleasant, unpleasant, and neutralstimuli from around 150 ms as well as evidence of priming withshorter P300 and response latencies across repetition trials. Thisrepetition effect occurred independently of stimulus arousallevel, with larger positivity from around 200 ms for high-compared to low-arousing stimuli. Further, N2 and P3amplitude decreased overall across the experimental session,implying that amplitude habituation was independent ofstimulus arousal level. Taken together, these findings suggestthat arousal, repetition, and habituation appear to be producedindependently of one another and overlap in time, although thetime-course of affective stimulus repetition-induced memoryeffects over longer intervals is unknown.
1.3. Stimulus distortion
Current theories of affective ERP modulation emphasizestimuli with high evolutionary significance as automatictriggers of basic motivational systems whenever the stimuliare perceived (e.g., Schupp et al., 2006). Most ERP studiesusing affective pictures have employed stimulus durations ofN500 ms, enabling a clear perception of picture contents.However, affective information also can be extracted duringconditions of limited perception, such that ERP effects ofarousal are conveyed in a stream of rapidly (120–333 ms)presented pictures (Junghöfer et al., 2001; Schupp et al., 2003,2004, 2006). This method influences perceptual processing ofaffective content and alters higher-level semantic analysis of thepicture content. This interpretation is supported by recentfindings that physical attributes such as featural compositionand complexity modulate affective ERPs perhaps by employingattentional mechanisms not specific to emotion (Bradley et al.,2007; Codispoti et al., 2006, 2007; Cano et al., in press;Delplanque et al., 2007). Thus, if ERP affective modulations arepreserved even when image is distorted the emotionalinformation must be mediated by other perceptual processes.
1.4. Present study
The goals of the present study were to characterize affectivevalence and arousal processing across stimulus repetition ofhighly distinct IAPS stimuli compared to visually distortedcontrols. IAPS images were selected in a way that the stimuliwere either rated as comparatively low or high on arousal level,and relatively strong negative and positive valences selectedwithin each arousal category. No neutral stimuli wereemployed. The chosen stimuli were presented as targets in anoddball paradigm, with normal pictures compared to their“scrambled” counterparts; and each stimulus occurred twice ineach condition.
This approach was devised in order to: (1) assess valence andarousal effects under repetition circumstances within a target
116 B. Rozenkrants et al. / International Journal of Psychophysiology 67 (2008) 114–123
detection task, and (2) temporally track affective ERP effects forthese variables across the waveform to characterize when andwhere they occur. As stimulus repetition is a common procedureand the physical stimulus properties contribute to outcomes, thesefactors were systematically manipulated to characterize theresulting ERP effects relative to previous studies. It washypothesized that arousal would modulate the later ERPcomponents, whereas valence would modulate the early ERPcomponents. Valence effects should be found primarily overfrontal electrodes, whereas arousal effects should be manifestover parietal areas. Stimulus repetition should produce largerlater potentials for normal but not for unrecognizablescrambled pictures, which also should demonstrate an absenceof affective outcomes.
2. Method
2.1. Participants
A total of 32 right-handed undergraduates (16 female) wereemployed as participants. All reported being free of neurolo-
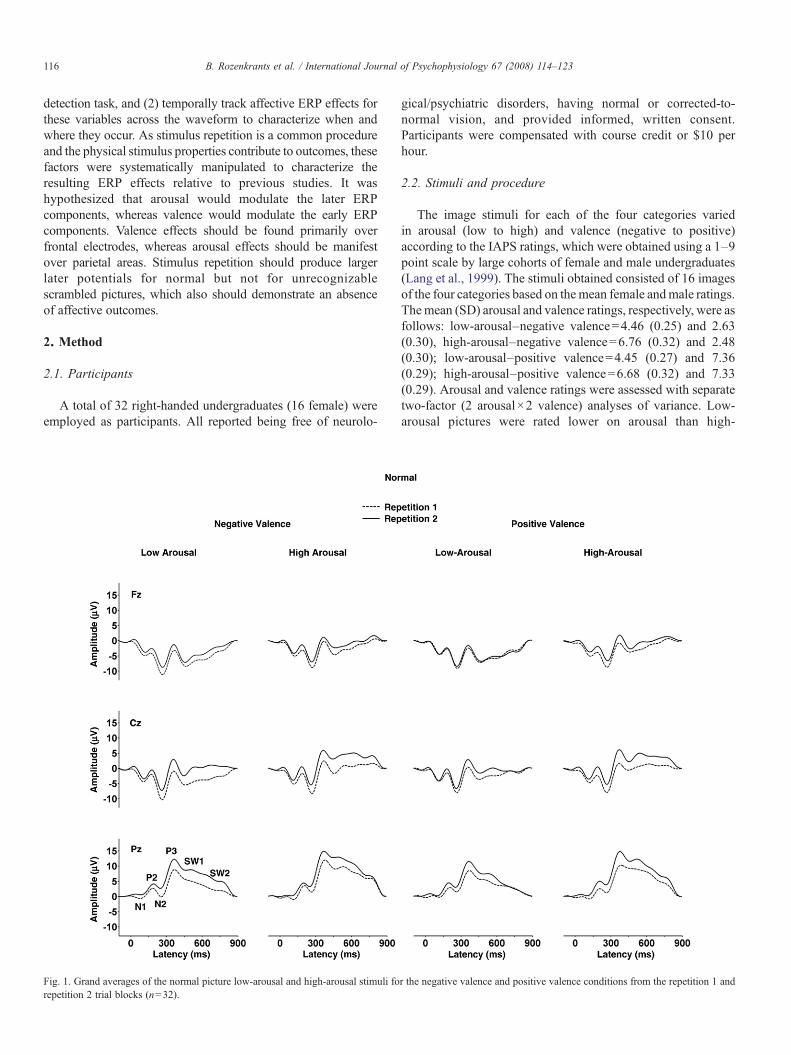
Fig. 1. Grand averages of the normal picture low-arousal and high-arousal stimuli forepetition 2 trial blocks (n=32).
gical/psychiatric disorders, having normal or corrected-to-normal vision, and provided informed, written consent.Participants were compensated with course credit or $10 perhour.
2.2. Stimuli and procedure
The image stimuli for each of the four categories variedin arousal (low to high) and valence (negative to positive)according to the IAPS ratings, which were obtained using a 1–9point scale by large cohorts of female and male undergraduates(Lang et al., 1999). The stimuli obtained consisted of 16 imagesof the four categories based on themean female andmale ratings.Themean (SD) arousal and valence ratings, respectively, were asfollows: low-arousal–negative valence=4.46 (0.25) and 2.63(0.30), high-arousal–negative valence=6.76 (0.32) and 2.48(0.30); low-arousal–positive valence=4.45 (0.27) and 7.36(0.29); high-arousal–positive valence=6.68 (0.32) and 7.33(0.29). Arousal and valence ratings were assessed with separatetwo-factor (2 arousal×2 valence) analyses of variance. Low-arousal pictures were rated lower on arousal than high-
r the negative valence and positive valence conditions from the repetition 1 and
117B. Rozenkrants et al. / International Journal of Psychophysiology 67 (2008) 114–123
arousal pictures, F(1,60)=4242.1, pb0.00001; negative valencepictures were rated more negative than positive valence pictures,F(1,60)=971.8, pb0.00001; no interactions were obtained(pN0.40, both cases).
These pictures were organized into a 9×12 cm format andserved as target stimuli presented to each subject in an oddballtask, with p=0.40 for a picture. A red and white checkedstandard image of equal size and a pattern designed to equatevisual spatial frequency to the target occurred with p=0.60.This stimulus was adapted from previous reports (Delplanqueet al., 2004, 2005, 2006). Additional “scrambled” target imageswere constructed by randomly rearranging 0.75 cm2 areas ofeach image (192 square blocks), which largely maintains thepicture's visual characteristics (color, spatial frequency, inten-sity) but eliminates its content. The Appendix lists the IAPSnumber of each stimulus and presents examples of a normal andscrambled stimulus from each category.
The subjects sat in a chair with 75 cm between the front edgeof the chair and the computer screen. All stimuli were displayedfor a duration of 1000 ms at the center of a computer screen on alight grey background at normal viewing luminance. A 2000 msinter-stimulus interval was used. Participants were instructed torespond with a mouse-click using their dominant right hand
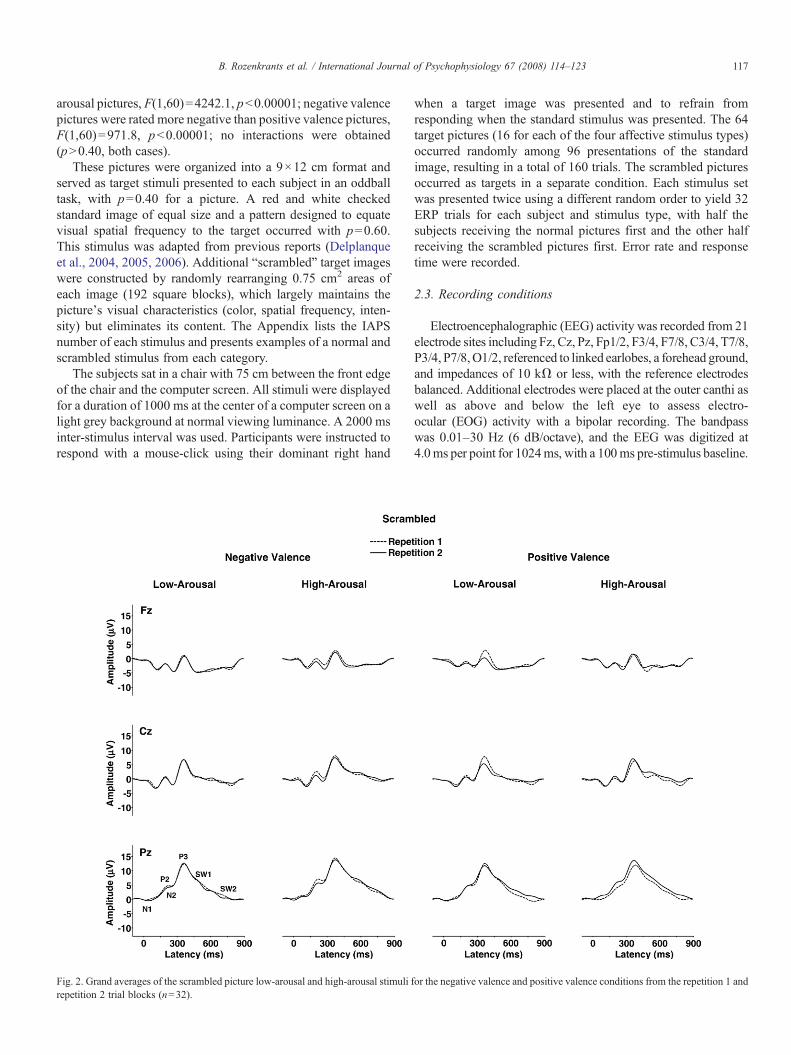
Fig. 2. Grand averages of the scrambled picture low-arousal and high-arousal stimuli frepetition 2 trial blocks (n=32).
when a target image was presented and to refrain fromresponding when the standard stimulus was presented. The 64target pictures (16 for each of the four affective stimulus types)occurred randomly among 96 presentations of the standardimage, resulting in a total of 160 trials. The scrambled picturesoccurred as targets in a separate condition. Each stimulus setwas presented twice using a different random order to yield 32ERP trials for each subject and stimulus type, with half thesubjects receiving the normal pictures first and the other halfreceiving the scrambled pictures first. Error rate and responsetime were recorded.
2.3. Recording conditions
Electroencephalographic (EEG) activity was recorded from 21electrode sites including Fz, Cz, Pz, Fp1/2, F3/4, F7/8, C3/4, T7/8,P3/4, P7/8,O1/2, referenced to linked earlobes, a forehead ground,and impedances of 10 kΩ or less, with the reference electrodesbalanced. Additional electrodes were placed at the outer canthi aswell as above and below the left eye to assess electro-ocular (EOG) activity with a bipolar recording. The bandpasswas 0.01–30 Hz (6 dB/octave), and the EEG was digitized at4.0ms per point for 1024ms, with a 100ms pre-stimulus baseline.
or the negative valence and positive valence conditions from the repetition 1 and

118 B. Rozenkrants et al. / International Journal of Psychophysiology 67 (2008) 114–123
Waveforms were averaged off-line, such that trials on which theEEG or EOG exceeded ±100 μV were rejected. Single-trial dataalso were subjected to an EOG correction procedure to removeany remaining artifact. Rest periods were provided as needed.
3. Results
3.1. Behavioral data
Error rate (ER) was defined as the percent of incorrectresponses, was less than 1%, and is not considered further.Response time (RT) was defined as the time from stimulusonset to the button press response, and the mean RT wascomputed over stimulus trials for each subject from eachcondition across stimulus blocks. RT was assessed with a four-factor (2 arousal×2 valence×2 repetition×2 stimulus type)analysis of variance. Mean RT did not differ amongarousal (pN .70) or valence (pN .30) conditions (low-arousal–negative valence=509, low-arousal–positive valence=502,high-arousal–negative valence=504, high-arousal–positivevalence=502 ms, respectively, pN .15). RT was longer for the
Fig. 3. Normal pictures: Mean amplitude for the N1, P2, N2, P3, early slow wave,negative valence and positive valence stimulus conditions as a function of midline e
first compared to the second repetition, F(1,31) = 12.7,pb0.002, and longer for the normal compared to scrambledstimulus items,F(1,31)=16.2, pb0.001. RTs for normal picturesdemonstrated a significant 45 ms decrease across repetitions,whereas scrambled pictures resulted in a non-significant change(5 ms decrease), yielding a repetition by stimulus typeinteraction, F(1,31)=6.7, pb0.02. Further, RT from the normalstimuli decreased similarly across arousal and valence levels, butthe scrambled stimuli yielded short RT and a stronger decreasefor negative stimuli with longer RT and less of a decrease forpositive stimuli to produce a three-way interaction amongarousal by valence by stimulus type, F(1,31)=4.8, pb0.05. Theorigin of this weak behavioral interaction is unknown.
3.2. ERP analyses
After excluding error and artifact trials, the mean number ofERP averages for each stimulus condition across stimulus type forrepetition block-1 was 14.7, 14.9, 14.7, 14.9, and for repetitionblock-2 was 15.1, 14.8, 14.8, 14.8, trials for the low-arousal–negative, high-arousal–negative, low-arousal–positive, and high-
and later slow wave components from the low-arousal and high-arousal for thelectrode with repetition 1 and repetition 2 trial blocks overlapped.
119B. Rozenkrants et al. / International Journal of Psychophysiology 67 (2008) 114–123
arousal–positive waveforms, respectively. A four-factor (2arousal×2 valence×2 repetition×2 stimulus type) analysis ofvariance performed on the number of trials found no differencesfor arousal (Fb1, pN0.80), valence (Fb1, pN0.50), repetition(Fb1, pN0.15), or stimulus type (F=2.5, pN0.12). Thus, trialnumber did not differ among conditions.
Fig. 1 illustrates the grand averages from the normal pictureimages. Fig. 2 illustrates the grand averages from the scrambledimages for each arousal and valence category. Averages fromeach repetition block overlapped for each of the midlineelectrodes. The ERP components were analyzed by measuringmean waveform amplitude relative to the pre-stimulus baselinewithin component time windows: N1=120–160 ms, P2=160–220, N2=220–300, P3=300–450 ms, early slow wave=550–700, and late slow wave=700–850 ms.
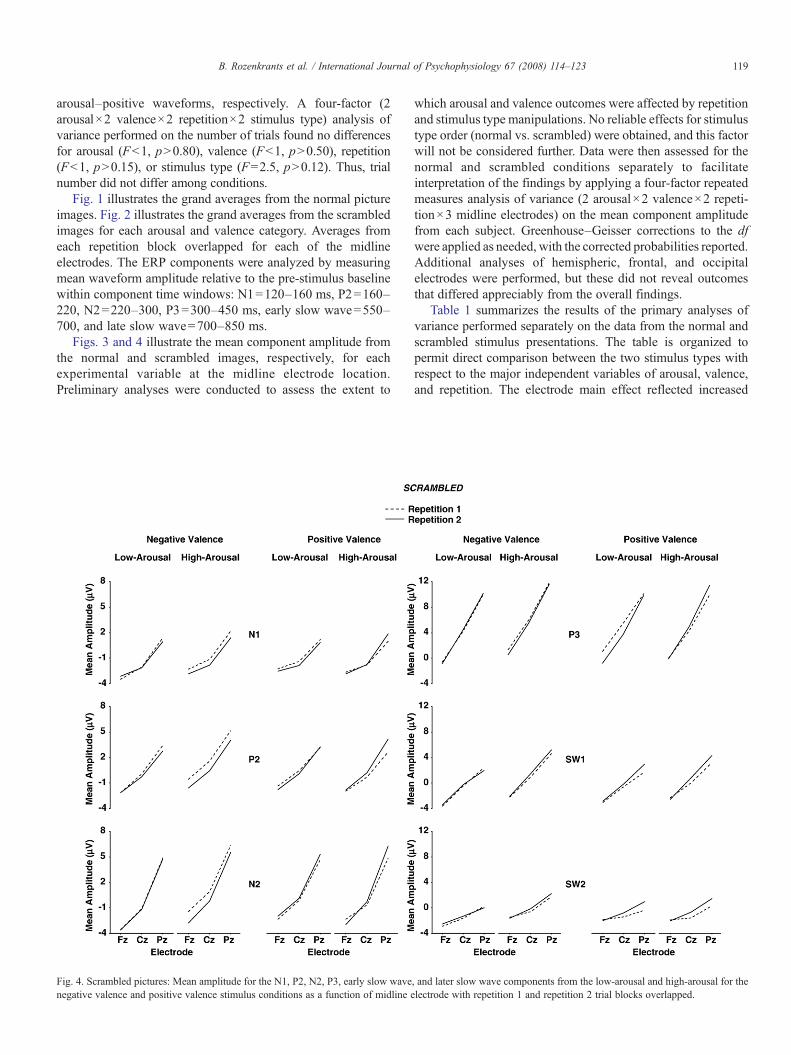
Figs. 3 and 4 illustrate the mean component amplitude fromthe normal and scrambled images, respectively, for eachexperimental variable at the midline electrode location.Preliminary analyses were conducted to assess the extent to
Fig. 4. Scrambled pictures: Mean amplitude for the N1, P2, N2, P3, early slow wavenegative valence and positive valence stimulus conditions as a function of midline e
which arousal and valence outcomes were affected by repetitionand stimulus type manipulations. No reliable effects for stimulustype order (normal vs. scrambled) were obtained, and this factorwill not be considered further. Data were then assessed for thenormal and scrambled conditions separately to facilitateinterpretation of the findings by applying a four-factor repeatedmeasures analysis of variance (2 arousal×2 valence×2 repeti-tion×3 midline electrodes) on the mean component amplitudefrom each subject. Greenhouse–Geisser corrections to the dfwere applied as needed, with the corrected probabilities reported.Additional analyses of hemispheric, frontal, and occipitalelectrodes were performed, but these did not reveal outcomesthat differed appreciably from the overall findings.
Table 1 summarizes the results of the primary analyses ofvariance performed separately on the data from the normal andscrambled stimulus presentations. The table is organized topermit direct comparison between the two stimulus types withrespect to the major independent variables of arousal, valence,and repetition. The electrode main effect reflected increased
, and later slow wave components from the low-arousal and high-arousal for thelectrode with repetition 1 and repetition 2 trial blocks overlapped.
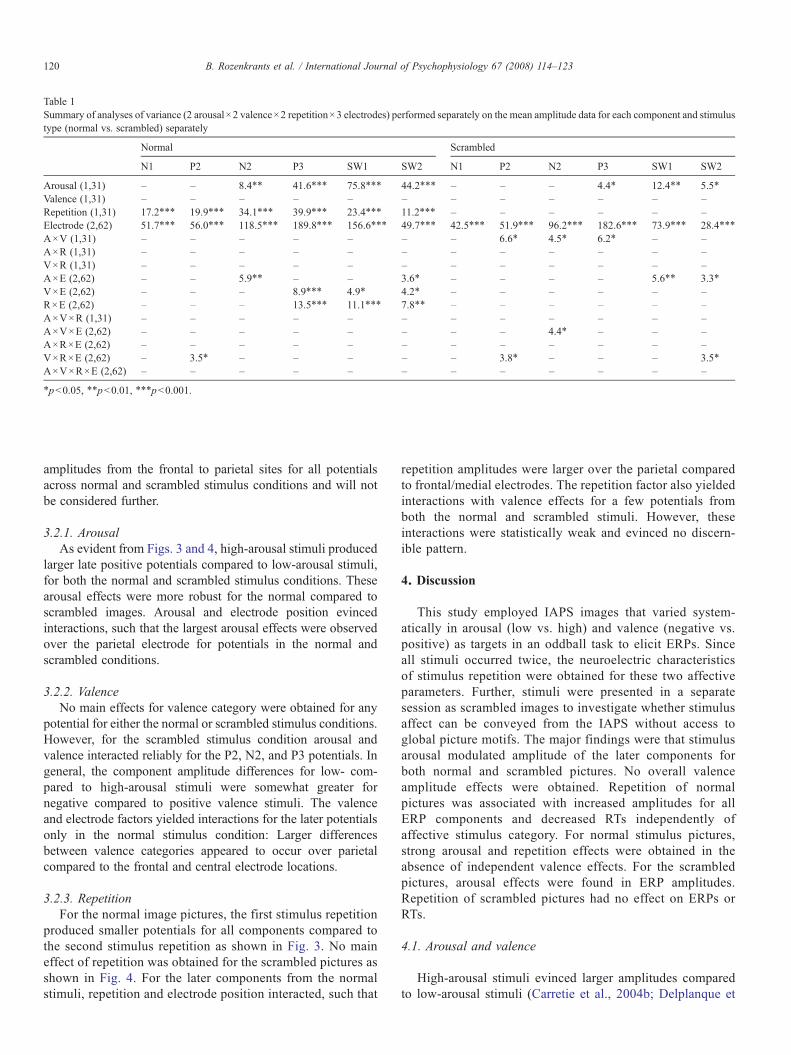
Table 1Summary of analyses of variance (2 arousal×2 valence×2 repetition×3 electrodes) performed separately on the mean amplitude data for each component and stimulustype (normal vs. scrambled) separately
Normal Scrambled
N1 P2 N2 P3 SW1 SW2 N1 P2 N2 P3 SW1 SW2
Arousal (1,31) – – 8.4⁎⁎ 41.6⁎⁎⁎ 75.8⁎⁎⁎ 44.2⁎⁎⁎ – – – 4.4⁎ 12.4⁎⁎ 5.5⁎
Valence (1,31) – – – – – – – – – – – –Repetition (1,31) 17.2⁎⁎⁎ 19.9⁎⁎⁎ 34.1⁎⁎⁎ 39.9⁎⁎⁎ 23.4⁎⁎⁎ 11.2⁎⁎⁎ – – – – – –Electrode (2,62) 51.7⁎⁎⁎ 56.0⁎⁎⁎ 118.5⁎⁎⁎ 189.8⁎⁎⁎ 156.6⁎⁎⁎ 49.7⁎⁎⁎ 42.5⁎⁎⁎ 51.9⁎⁎⁎ 96.2⁎⁎⁎ 182.6⁎⁎⁎ 73.9⁎⁎⁎ 28.4⁎⁎⁎
A×V (1,31) – – – – – – – 6.6⁎ 4.5⁎ 6.2⁎ – –A×R (1,31) – – – – – – – – – – – –V×R (1,31) – – – – – – – – – – – –A×E (2,62) – – 5.9⁎⁎ – – 3.6⁎ – – – – 5.6⁎⁎ 3.3⁎
V×E (2,62) – – – 8.9⁎⁎⁎ 4.9⁎ 4.2⁎ – – – – – –R×E (2,62) – – – 13.5⁎⁎⁎ 11.1⁎⁎⁎ 7.8⁎⁎ – – – – – –A×V×R (1,31) – – – – – – – – – – – –A×V×E (2,62) – – – – – – – – 4.4⁎ – – –A×R×E (2,62) – – – – – – – – – – – –V×R×E (2,62) – 3.5⁎ – – – – – 3.8⁎ – – – 3.5⁎
A×V×R×E (2,62) – – – – – – – – – – – –
⁎pb0.05, ⁎⁎pb0.01, ⁎⁎⁎pb0.001.
120 B. Rozenkrants et al. / International Journal of Psychophysiology 67 (2008) 114–123
amplitudes from the frontal to parietal sites for all potentialsacross normal and scrambled stimulus conditions and will notbe considered further.
3.2.1. ArousalAs evident from Figs. 3 and 4, high-arousal stimuli produced
larger late positive potentials compared to low-arousal stimuli,for both the normal and scrambled stimulus conditions. Thesearousal effects were more robust for the normal compared toscrambled images. Arousal and electrode position evincedinteractions, such that the largest arousal effects were observedover the parietal electrode for potentials in the normal andscrambled conditions.
3.2.2. ValenceNo main effects for valence category were obtained for any
potential for either the normal or scrambled stimulus conditions.However, for the scrambled stimulus condition arousal andvalence interacted reliably for the P2, N2, and P3 potentials. Ingeneral, the component amplitude differences for low- com-pared to high-arousal stimuli were somewhat greater fornegative compared to positive valence stimuli. The valenceand electrode factors yielded interactions for the later potentialsonly in the normal stimulus condition: Larger differencesbetween valence categories appeared to occur over parietalcompared to the frontal and central electrode locations.
3.2.3. RepetitionFor the normal image pictures, the first stimulus repetition
produced smaller potentials for all components compared tothe second stimulus repetition as shown in Fig. 3. No maineffect of repetition was obtained for the scrambled pictures asshown in Fig. 4. For the later components from the normalstimuli, repetition and electrode position interacted, such that
repetition amplitudes were larger over the parietal comparedto frontal/medial electrodes. The repetition factor also yieldedinteractions with valence effects for a few potentials fromboth the normal and scrambled stimuli. However, theseinteractions were statistically weak and evinced no discern-ible pattern.
4. Discussion
This study employed IAPS images that varied system-atically in arousal (low vs. high) and valence (negative vs.positive) as targets in an oddball task to elicit ERPs. Sinceall stimuli occurred twice, the neuroelectric characteristicsof stimulus repetition were obtained for these two affectiveparameters. Further, stimuli were presented in a separatesession as scrambled images to investigate whether stimulusaffect can be conveyed from the IAPS without access toglobal picture motifs. The major findings were that stimulusarousal modulated amplitude of the later components forboth normal and scrambled pictures. No overall valenceamplitude effects were obtained. Repetition of normalpictures was associated with increased amplitudes for allERP components and decreased RTs independently ofaffective stimulus category. For normal stimulus pictures,strong arousal and repetition effects were obtained in theabsence of independent valence effects. For the scrambledpictures, arousal effects were found in ERP amplitudes.Repetition of scrambled pictures had no effect on ERPs orRTs.
4.1. Arousal and valence
High-arousal stimuli evinced larger amplitudes comparedto low-arousal stimuli (Carretie et al., 2004b; Delplanque et

121B. Rozenkrants et al. / International Journal of Psychophysiology 67 (2008) 114–123
al., 2006; Schupp et al., 2000). These effects beganapproximately with the N2, became appreciably stronger forthe P3 and early SW component, and were still robust for thelate SW portion of the waveform as has been previously found(Olofsson and Polich, 2007). Arousal level also increased theamplitude of the later potentials for the scrambled pictures,although the effects were statistically weaker than those fromthe normal pictures. No overall valence effects were obtainedfor either stimulus type, but arousal level and valenceinteracted for several components elicited by the scrambledstimuli.
A novel finding in this study is that highly arousing picturesstill produce an elevated late ERP positivity for the scrambledstimulus images. The distorted pictures were made from thesame IAPS pictures, with each picture segmented into squaresand randomly rearranged. These distorted stimuli are unrecog-nizable and yielded no repetition memory effects for either RTor ERPs components, which were found for the normalpictures. These outcomes are consistent with previous studiesthat altered stimulus perception by quick serial presentation ofaffective pictures, as arousing stimuli appear to cause rapidcortical modulations even when picture content cannot beaccessed due to perceptual limitations (Junghöfer et al., 2001;Schupp et al., 2003, 2006). These and the present results implythat physical stimulus properties (e.g., color, contrast, texture)can produce arousal ERP effects. Although the exact mechan-isms of these influences are unknown, relatively low-levelperceptual features of affective pictures appear to contribute toERP arousal outcomes (Codispoti et al., 2006; Delplanqueet al., 2007).
4.2. Stimulus repetition
Stimulus repetition of the normal pictures producedincreased component amplitudes from the first to the secondtrial block, with no repetition main effects found for scrambledimages. Recognition memory ERP studies have demonstratedincreased amplitudes for late positive components when astimulus is recognized as having been presented previously inthe series (Bentin et al., 1992; Friedman, 1990; Rugg, 1990).Successively repeated emotional stimuli used as targets in anoddball task produced similar amplitude increases from thefirst to the second repetition (Olofsson and Polich, 2007).The current results corroborate previous findings and extendthe repetition effect to non-successive and unpredictablestimulus sequences (Aftanas et al., 2001; Carretie et al.,2001a,b, 2003, 2004a; Ito et al., 1998; Keil et al., 2001;Schupp et al., 2000, 2004). Moreover, the lack of interactionwith the arousal or valence factors indicates that themodulation of rapid brain responses to affective images isstable over repetitions, so that ERP modulations fromreoccurring affective picture stimuli are unlikely to beconfounded by repeated stimulus presentations. That therepetition effects of ERP amplitudes and RTs occurred for thenormal but not the scrambled images suggest that onlynormal images that can be recognized are subject toamplitude increase due to repetition.
4.3. Theoretical perspective
The straightforward nature of the present task with thesame response for each target stimulus in the absence of anyemotive judgments implies that the obtained ERP effectswere produced automatically (Bernat et al., 2001; Olofssonand Polich, 2007; Schupp et al., 2000). Stimulus arousalinfluenced late component amplitudes from both normal andscrambled stimuli and suggest that fundamental perceptualattributes are possible causes of arousal-related processingmodulations, which were statistically weaker for thescrambled stimulus conditions. The variation for componentamplitude between arousal and valence levels observed forthe scrambled stimuli in the middle-latency range also seemto reflect automatic featural processing of affective proper-ties. The exact mechanisms by which arousal-related ERPmodulations are preserved in distorted images are unknown.However, the results from this and other studies suggest thatphysical stimulus characteristics (e.g., color, spatial fre-quency, local/global features) can convey arousing proper-ties (Bradley et al 2007; Cano et al., in press; Codispoti et al2006, 2007). A systematic integration of these findingsshould emerge as additional results accumulate (Olofssonet al., in press).
Although the origins of arousal/valence interaction effects inthe scrambled images remain vague, they may result in partfrom the use of the extreme negative and positive valenceratings coupled with the extreme low- and high-arousal levelsused to select the present stimuli. Perhaps normal picture stimulidid not yield this interaction because these images arenecessarily perceived as a whole percept rather than theindividual components seen in scrambled images. Capturingthe image in its entirety could override basic perceptualinfluences by which valence and arousal interacts (e.g., hue orbrightness). Further, cognitive operations are likely to beutilized more efficiently for normal pictures compared toscrambled images, which would enhance memory performanceand increase P300/slow wave amplitude (Azizian and Polich,2007; Karis et al., 1984; Paller et al., 1988). Such ERPamplitude changes have been obtained when affective memoryformation is assessed (Dolcos and Cabeza, 2002; Palomba et al.,1997). The present results indicate that emotional stimulusproperties also can be conveyed without access to affectivecontent, such that stimulus cues apparently can modulate ERPswhen perceptual quality is limited or altered.
Acknowledgements
The first author received an Undergraduate ResearchFellowship from the University of California, San Diegoand is now in the Neuroscience Graduate Program at theUniversity of Southern California. The second author wassupported by a fellowship from the American–ScandinavianFoundation and Thord-Gray Memorial Fund. This study wassupported by RO1-DA018262 and P50-AA06420. This paperis publication number 18822 from The Scripps ResearchInstitute.

Appendix A. Stimulus category, numbers of selected IAPS pictures, and sample normal and scrambled picture stimuli
122 B. Rozenkrants et al. / International Journal of Psychophysiology 67 (2008) 114–123
The images were presented in color.
References
Aftanas, L.I., Varlamov, A.A., Pavlov, S.V., Makhnev, V.P., Reva, N.V., 2001.Affective picture processing: event-related synchronization within indivi-dually defined human theta band is modulated by valence dimension.Neuroscience Letters 303, 115–118.
Azizian, A., Polich, J., 2007. Evidence for attentional gradient in the serialposition memory curve from ERPs Journal of Cognitive Neuroscience 19,2071–2081.
Bentin, S., Moscovitch, M., Heth, I., 1992. Memory with and without awareness:performance and electrophysiological evidence of savings. Journal ofExperimental Psychology. Learning, Memory, and Cognition 18, 1270–1283.
Bernat, E., Bunce, S., Shevrin, H., 2001. Event-related brain potentials differentiatepositive and negativemood adjectives during both supraliminal and subliminalvisual processing. International Journal of Psychophysiology 42, 11–34.
Bradley, M.M., Greenwald, M.K., Petry, M.C., Lang, P.J., 1992. Rememberingpictures: pleasure and arousal in memory. Journal of ExperimentalPsychology. Learning, Memory, and Cognition 18, 379–390.
Bradley, M.M., Hamby, S., Low, A., Lang, P.J., 2007. Brain potentials in perception:picture complexity and emotional arousal. Psychophysiology 44, 364–373.
Brazdil, M., Roman, R., Daniel, P., Rektor, I., 2003. Intracerebral somatosensoryevent-related potentials: effect of response type (button pressing versusmental counting) on P3-like potentials within the human brain. ClinicalNeurophysiology 114, 1489–1496.
Cacioppo, J.T., Gardner, W.L., Berntson, G.G., 1999. The affect system hasparallel and integrative processing components: form follows function.Journal of Personality and Social Psychology 76, 839–855.
Cano, M.E., Class, Q.A., Polich, J., in press. Affective valence, stimulusattributes, and P300: color vs. black/white and normal vs. scrambled images.International Journal of Psychophysiology.
Conroy, M.A., Polich, J., 2007. Affective valence and P300 when stimulusarousal level is controlled. Cognition and Emotion 21, 891–901.
Carretie, L., Martin-Loeches, M., Hinojosa, J.A., Mercado, F., 2001a. Emotionand attention interaction studied through event-related potentials. Journal ofCognitive Neuroscience 13, 1109–1128.
Carretie, L., Mercado, F., Tapia, M., Hinojosa, J.A., 2001b. Emotion, attention,and the ‘negativity bias’, studied through event-related potentials. Interna-tional Journal of Psychophysiology 41, 75–85.
Carretie, L., Hinojosa, J.A., Mercado, F., 2003. Cerebral patterns of attentionalhabituation to emotional visual stimuli. Psychophysiology 40, 381–388.
Carretie, L., Hinojosa, J.A., Martin-Loeches, M., Mercado, F., Tapia, M., 2004a.Automatic attention to emotional stimuli: neural correlates. Human BrainMapping 22, 290–299.
Carretie, L., Mercado, F., Hinojosa, J.A., Martin-Loeches, M., Sotillo, M.,2004b. Valence-related vigilance biases in anxiety studied through event-related potentials. Journal of Affective Disorders 78, 119–130.
Codispoti, M., Ferrari, V., De Cesarei, A., Cardinale, R., 2006. Implicit and explicitcategorization of natural scenes. Progress in Brain Research 156, 53–65.
Codispoti, M., Ferrari, V., Bradley, M.M., 2007. Repetition and event-relatedpotentials: distinguishing early and late processes in affective pictureperception. Journal of Cognitive Neuroscience 19, 577–586.
Crawford, L.E., Cacioppo, J.T., 2002. Learning where to look for danger:integrating affective and spatial information. Psychological Science 13,449–453.
Cuthbert, B.N., Schupp, H.T., Bradley, M.M., Birbaumer, N., Lang, P.J., 2000.Brain potentials in affective picture processing: covariation with autonomicarousal and affective report. Biological Psychology 52, 95–111.
Delplanque, S., Lavoie, M.E., Hot, P., Silvert, L., Sequeira, H., 2004.Modulation of cognitive processing by emotional valence studied throughevent-related potentials in humans. Neuroscience Letters 356, 1–4.
Delplanque, S., Silvert, L., Hot, P., Sequeira, H., 2005. Event-related P3a andP3b in response to unpredictable emotional stimuli. Biological Psychology68, 107–120.
Delplanque, S., Silvert, L., Hot, P., Rigoulot, S., Sequeira, H., 2006. Arousal andvalence effects on event-related P3a and P3b during emotional categoriza-tion. International Journal of Psychophysiology 60, 315–322.
Delplanque, S., N'diaye, K., Scherer, K., Granjean, D., 2007. Spatial frequenciesor emotional effects? A systematic measure of spatial frequencies for IAPSpictures by a discrete wavelet analysis. Journal of Neuroscience Methods165, 144–150.
Dien, J., 1999. Differential lateralization of trait anxiety and trait fearfulness: evokedpotential correlates. Personality and Individual Differences 26, 333–356.
Dolcos, F., Cabeza, R., 2002. Event-related potentials of emotional memory:encoding pleasant, unpleasant, and neutral pictures. Cognitive, Affectiveand Behavioral Neuroscience 2, 252–263.
Friedman, D., 1990. Cognitive event-related potential components duringcontinuous recognition memory for pictures. Psychophysiology 27, 136–148.
Hajcak, G., Nieuwenhuis, S., 2006. Reappraisal modulates the electrocorticalresponse to unpleasant pictures. Cognitive Affective & BehavioralNeuroscience 6, 291–297.

123B. Rozenkrants et al. / International Journal of Psychophysiology 67 (2008) 114–123
Ito, T.A., Cacioppo, J.T., Lang, P.J., 1998. Eliciting affect using the internationalaffective picture system: trajectories through evaluative space. Personalityand Social Psychology Bulletin 24, 855–879.
Junghöfer, M., Bradley, M.M., Elbert, T.R., Lang, P.J., 2001. Fleeting images: anew look at early emotion discrimination. Psychophysiology 38, 175–178.
Karis, D., Druckman, D., Lissak, R., Donchin, E., 1984. A psychophysiologicalanalysis of bargaining. ERPs and facial expressions. Annals of the NewYorkAcademy of Sciences 425, 230–235.
Keil, A., Muller, M.M., Gruber, T., Wienbruch, C., Stolarova, M., Elbert, T.,2001. Effects of emotional arousal in the cerebral hemispheres: a study ofoscillatory brain activity and event-related potentials. Clinical Neurophy-siology 112, 2057–2068.
Keil, A., Bradley, M.M., Hauk, O., Rockstroh, B., Elbert, T., Lang, P.J., 2002.Large-scale neural correlates of affective picture processing. Psychophy-siology 39, 641–649.
Lang, P.J., Greenwald, M.K., Bradley, M.M., Hamm, A.O., 1993. Looking atpictures: affective, facial, visceral, and behavioral reactions. Psychophysiol-ogy 30, 261–273.
Lang, P.J., Bradley, M.M., Cuthbert, B.N., 1999. International Affective PictureSystem (IAPS): Instruction Manual and Affective Ratings. Gainsville,University of Florida.
LeDoux, J.E., 1989. Cognitive emotional interactions in the brain. Cognitionand Emotion. Special Issue: Development of emotion cognition relations,vol. 3, pp. 267–289.
LeDoux, J.E., 1995. Emotion: clues from the brain. Annual Review ofPsychology 46, 209–235.
Mini, A., Palomba, D., Angrilli, A., Bravi, S., 1996. Emotional informationprocessing and visual evoked brain potentials. Perceptual and Motor Skills83, 143–152.
Morris, J.S., Öhman, A., Dolan, R.J., 1998. Conscious and unconsciousemotional learning in the human amygdala. Nature 393, 467–470.
Moser, E., Hajcac, G., Bukay, E., Simons, R.F., 2006. Intentional modulation ofemotional responding to unpleasant pictures: an ERP study. Psychophysiol-ogy 43, 292–296.
Öhman, A., Soares, J.J., 1998. Emotional conditioning to masked stimuli:expectancies for aversive outcomes following nonrecognized fear-relevantstimuli. Journal of Experimental Psychology. General 127, 69–82.
Öhman, A.,Mineka, S., 2001. Fears, phobias, and preparedness: toward an evolvedmodule of fear and fear learning. Psychological Review 108, 483–522.
Olofsson, J.K., Polich, J., 2007. Affective visual event-related potentials:arousal, repetition, and time-on-task. Biological Psychology 75, 101–108.
Olofsson, J.K., Nordin, S., Sequeira, H., Polich, J., in press. Affective PictureProcessing: an Integrative Review of ERP Findings. Biological Psychology.
Paller, K.A., Mccarthy, G., Wood, C.C., 1988. ERPs predictive of subsequentrecall and recognition performance. Biological Psychology 26, 269–276.
Palomba, D., Angrilli, A., Mini, A., 1997. Visual evoked potentials, heart rateresponses and memory to emotional pictorial stimuli. International Journalof Psychophysiology 27, 55–67.
Polich, J., 2007. Updating P300: an integrative theory of P3a and P3b. ClinicalNeurophysiology 118, 2128–2148.
Roschmann, R., Wittling, W., 1992. Topographic brain mapping of emotion-related hemisphere asymmetries. International Journal of Neuroscience 63,5–16.
Rugg, M.D., 1990. Event-related brain potentials dissociate repetition effects ofhigh- and low-frequency words. Memory & Cognition 18, 367–379.
Sabatinelli, D., Lang, P.J., Keil, A., Bradley, M.M., 2007. Emotional perception:correlation of functional MRI and event-related potentials. Cerebral Cortex17, 1085–1091.
Schupp, H.T., Cuthbert, B.N., Bradley, M.M., Cacioppo, J.T., Ito, T., Lang, P.J.,2000. Affective picture processing: the late positive potential is modulatedby motivational relevance. Psychophysiology 37, 257–261.
Schupp, H.T., Junghöfer, M., Weike, A.I., Hamm, A.O., 2003. Attention andemotion: an ERP analysis of facilitated emotional stimulus processing.Neuroreport 14, 1107–1110.
Schupp, H.T., Junghöfer, M., Weike, A.I., Hamm, A.O., 2004. The selectiveprocessing of briefly presented affective pictures: an ERP analysis.Psychophysiology 41, 441–449.
Schupp, H.T., Flaisch, T., Stockburger, J., Junghöfer, M., 2006. Emotion andattention: event-related brain potential studies. Progress in Brain Research156, 31–51.
Segalowitz, S.J., Van Roon, P., Dywan, J., 1997. The ERP late positivity: agraduated response to stimulus repetition. Neuroreport 8, 757–760.
Smith, M.E., Halgren, E., 1989. Dissociation of recognition memorycomponents following temporal lobe lesions. Journal of ExperimentalPsychology 15, 50–60.













![A Vibrotactile Alarm System for Pleasant Awakening · [23]. The valence level indicates how positive or negative a stimulus is, whereas the arousal level indicates how calming or](https://img.pdfslide.net/doc/110x75/5eaaca730bf65108833e3bde/a-vibrotactile-alarm-system-for-pleasant-awakening-23-the-valence-level-indicates.jpg)