Embed Size (px)
Citation preview

MARINE MAMMAL SCIENCE, 12(1):54-88 (January 1996) 0 1996 by the Society for Marine Mammalogy
AGE DETERMINATION IN MANATEES USING GROWTH-LAYER-GROUP COUNTS IN BONE
MIRIAM MARMONTEL Department of Wildlife and Range Sciences, University of Florida,
Gainesville, Florida 326 11
THOMAS J. O’SHEA HOWARD I. KOCHMAN
U.S. Fish and Wildlife Service National Ecology Research Center, Sirenia Project, 412 NE 16th Ave., Gainesville, Florida 32601
STEPHEN R. HUMPHREY Florida Museum of Natural History, University of Florida,
Gainesville, Florida 32611
Growth layers were observed in histological preparations of bones of known- age, known minimum-age, and tetracycline-marked free-ranging and captive Florida manatees (Trichechus manatus latirostris), substantiating earlier prelim- inary findings of other studies. Detailed analysis of 17 new case histories showed that growth-layer group (GLG) counts in the periotic bone were consistent with known age, or time since tetracycline administration, but were less reliable in other bones. GLG counts were also made in periotic bones of 1,196 Florida manatees of unknown age found dead from 1974 through 1991. These counts were conducted in order to assess variability and to determine relationships among estimated age, size, sex, and degree of bone resorption. Resorption can interfere with accuracy of GLG counts. This effect does not occur until ages greater than about 15 yr and body lengths greater than 300 cm are attained. GLGs were also observed in periotic bones of Antillean manatees (Trichechus manatus manatus) but were not validated against known-age specimens. Use of GLG counts in the periotic bone is suitable for application to studies of population dynamics and other age-related aspects of manatee biology.
Key words: age determination, Florida, growth layers, manatee, Sirenia, Tri- chechus manatus, bone.
Current addresses: 1 Projeto Mamiraua, Av. Brasil, 197, 69470 Tefe AM, Brazil; 2 > National Biological Service, Midcontinent Ecological Science Center, 4512 McMurry Ave., Fort Collins, Colorado 80525-3400, U.S.A.; 3 now with National Biological Service, same address.
54

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 55
Understanding of manatee population biology has been hampered by the lack of anatomical age-determination techniques. This is, in part, because most methods of age determination in mammals cannot be applied to manatees. For example, manatees lack bacula, baleen, earplugs, pelage, horns, and obvious secondary sexual characteristics. Counting of corpora albicantia in ovaries cannot provide age estimates because manatees are polyovular, and corpora albicantia do not remain visible throughout life but are resorbed at an unknown rate (Marmontel et al. 1992). Among marine mammals, annuli in the teeth have been used to determine age in pinnipeds (Laws 1952, Payne 1978), and odon- tocetes (Perrin and Myrick 1980, Hohn et al. 1989). This approach has also been followed in the dugong (Dugong dugon), where the permanent, tusklike incisors possess growth-layer groups (GLGs) that are deposited annually (Mitchell 1978, Marsh 1980). Unlike dugongs, manatees lack incisors, and unlike almost all other mammals, manatees have an indeterminate number of supernumerary molars that are continually replaced throughout life (Domning and Hayek 1984). This renders teeth inappropriate for age-determination studies, except in young individuals.
Periodic growth marks in skeletal tissues of mammals were recognized as criteria for age assessment as early as 1950 (Scheffer 1950). Excellent reviews on this topic have been produced subsequently (e.g., Klevezal’ and Kleinenberg 1969, Morris 1972). GLGs in bone have been validated as annuli in some mammals (Klevezal’ and Kleinenberg 1969, Laws 1960). Quantification of GLGs in bone, therefore, has promise as a technique for age determination in manatees. However, bone is dynamic. Assumptions that growth layers (GL) are annual and that their total number corresponds to true age must be carefully validated by known-age material. The prior absence of known-age specimens and the pachyosclerotic nature of sirenian bones (requiring more elaborate tech- niques for specimen preparation) have heretofore delayed thorough evaluation of bone GLGs as an indicator of age in manatees.
However, preliminary research previously suggested the potential utility of GLGs in bone for manatee age determination. Domning and Myrick (1980) found GLGs in ribs and mandibles of one Amazonian manatee and an unspecified small number of Florida manatees. Best (1982) also reported GLGs in bone of a small sample of Amazonian manatees. Myrick and Kelly (unpublished manu- script) and Myrick (1985) carried out a preliminary study of GLGs in bone of 12 Florida manatees in 1980. Results suggested that the periotic bone provided maximum counts of GLGs in manatees in comparison with teeth, mandibles, and ribs. None of the 12 manatees examined in the latter two studies were of known age; one was treated with tetracycline for therapeutic purposes but died before a full year had elapsed. Tetracycline produces a fluorescent mark in living bone at the time of injection (Frost et al. 1961), allowing calibration of sub- sequent layering. To verify findings of these preliminary studies, we examined skeletons from a series of tetracycline-labeled or known-age Florida manatees. The objective of our study was to use these specimens, as well as a much larger sample of individuals of unknown age, to determine more fully the potential for developing age-determination methods for Florida manatees based on GLGs

56 MARINE MAMMAL SCIENCE. VOL. 12. NO. 1. 1996
in bone. A progress report on this work (Marmontel et ul. 1990) presented findings based on a smaller data set.
MATERIALS AND METHODS
Specimens examined-Bones from 17 known-age, known minimum-age, or tetracycline-marked manatees were used to validate GLG counts. Known-age specimens included two captive-born manatees and individually recognizable wild manatees from intensively studied winter aggregations. Study areas included Blue Spring on the upper St. Johns River, Crystal River on the northern Gulf coast, and Atlantic coast sites as described by O’Shea and Hartley (in press), Rathbun et al. (in press) and Reid et al. (in press). Specimens used for GLG validation also included nine manatees administered tetracycline in the field or at oceanaria. We also examined periotic bones from 1,196 manatees of unknown age and without tetracycline labels. This sample included most carcasses recovered in the southeastern United States between August 1974 and December 1991 and allowed examination of GLGs over a wider range of body sizes. Antillean manatees (Trichechus manatus manatus) from Puerto Rico and Belize were treated separately. In describing case histories we use museum catalog numbers, or field numbers for specimens not accessioned into museums (specimen num- bering is detailed in Marmontel 1993). Body-size measurements are total lengths (Bonde et al. 1983).
We use the terms calf, subadult, and adult to denote relative age. Calves are nursing young; subadults are weaned but reproductively immature; adults are reproductively mature. Photoidentification studies demonstrate that wild calves nurse for l-2 yr and that sexual maturity occurs at ages from 3 to 8 yr (Rathbun et al., in press, O’Shea and Hartley, in press). These terms describe specimens with known histories and those for which necropsy reports indicate reproductive status (Marmontel, in press). Where such information is unavail- able, we u.se size criteria (O’Shea et al. 1985): manatees less than 175-cm total length are defined as calves, manatees greater than 275 cm as adults, and. those of intermediate size as subadults.
Tetracycline marks--Tetracycline binds with calcium and is incorporated into active tissues within 48 h of administration (Frost et al. 1961, Frost 1.969). Under visible light, tetracycline-marked bone exhibits a yellow-brown color, and under ultraviolet light it produces a characteristic yellow-gold fluorescence (Frost 1983). Because of its utility in GLG calibration, the Sirenia Project and co- operators began routinely injecting captive manatees and wild individuals from selected areas with tetracycline in 1981. Sixty-nine manatees were marked through December 1992. Liquamicyn LA-200 oxytetracycline (Pfizer) was administered intramuscularly at a dosage of 30 mg/kg to 19 wild manatees caught by net at Blue Spring. Injection was at multiple sites (up to 15 ml/injection site) in the dorsal peduncle. This pattern of administration, based on recommendations by D. P. Domning (unpublished report) and participating veterinarians at oceanaria, was also followed routinely for captive manatees. Nine manatees were recovered dead between 7 and 13 1 mo after tetracycline administration. One

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 57
specimen (UF 20757) received therapeutic oral and intravenous doses in cap- tivity, and was previously examined by Myrick and Kelly in 1980 (unpublished manuscript).
Sample Preparation -Methods developed for examination of teeth and bone of dolphins (Perrin et a/. 1976, 1977; Hohn 1980; Hohn et al. 1989) at the National Marine Fisheries Service, Southwest Fisheries Science Center (SWFSC) were modified for manatee bone investigations. Details of modifications are provided by Marmontel (1993), and Marmontel et al. (1990). One of us (MM) received initial training at SWFSC by A. Hohn, building on the methods of Myrick and Kelly. Nearly all bones collected prior to 1989 were stored in air. Earbones collected since late 1989 were fixed in 10% neutral buffered formalin and maintained in that medium or transferred to 70% alcohol. Bones were rough-sectioned on a rock saw to provide a holding surface for fine sectioning. A 4-mm section was cut for decalcification and histological preparation. Un- treated thick sections (approximately 140 pm) of tetracycline-labeled bone were mounted directly on a slide for examination under ultraviolet light. Dry bone sections were fixed in formalin overnight prior to decalcification. Specimens were then decalcified in RDO (Apex Engineering Products) and rinsed thoroughly. A freezing microtome produced 40-wrn sections, which were subsequently stained in Mayer’s hematoxylin (modified by Grue) and mounted in glycerin. We calibrated the rate of GLG formation for tetracycline-labeled specimens by comparing the stained section with the thick section of the same bone. The distance from the tetracycline label to the edge of the periosteal layer was measured for the thick section. Subsequently, the number of bone GLGs within that distance in the stained section was compared with the time between tetra- cycline injection and death. A Microcomp morphometry microcomputer program (Southern Micro Instruments) allowed accurate measurement and comparison of GL in stained and fluorescence sections. Multiple measurements (n I 10) were made in the middle third of the dome section of the periotic bone (= pars temporalis or tegmen tympani of Robineau 1969). An IBAS image analysis system was used to measure, enhance, and photograph GLGs. Both fluorescence and stained sections were photomicrographed to allow detailed examination. Information on archiving of slides and skeletal material is available from the Sirenia Project.
Definitions and enumeration of GLGs--A post-hoc method of defining GLGs for age-class estimation was developed based on initial findings. The method is supported by studies of other species. In dentine and cement of other mammals (Klevezal’ and Kleinenberg 1969, Perrin and Myrick 1980) a broad and weakly stained band followed by a narrow and intensely colored line represents an annual layer. The wide bands represent periods of active bone growth, and narrower lines represent reductions in apposition of bone (Weinmann and Sicher 1947; Sissons 1949, 1971). In stained sections of decalcified bone tissue prepared for this study, broad, opaque bands, rich in cells and well vascularized (“zones”), also stained less intensely than the thinner bordering hematoxylinophilic lines (adhesion lines, or Castagnet’s “arrest lines of growth” [Castagnet 1974, 1980]) that separate them. Adhesion lines lack fibrils, consist of an adhesive substance

58 MARINE MAMMAL SCIENCE, VOL. 12, NO. 1, 1996
impregnated with calcium salts, and are more calcified than the collagen of bone, and therefore stain intensely (Weinmann and Sicher 1947, Amprino and Engs- trom 1952). In our study a broad zone (light stain) followed by an adhesion line (dark stain) is considered to be a GLG representing an annual period. The difference in distinctiveness of the first, and sometimes the second, GL in manatee bone is supported by studies of cement, dentine, and bone in other species (Laws 1953, 1958; Mansfield 1958).
GLG counts were carried out on histological sections of the central area of the dome portion of the periotic bone (halfway from its anterior end to its junction with the mastoid portion). GLs were clearly defined in this region, as found by Myrick and Kelly (unpublished manuscript), and counts were most consistent with known age or tetracycline calibration compared with other bones (see Results). Age-class estimation, using microscopic sections of the periotic dome, started with identifying two types of reticular tissue and the subsequent change in structure to compact bone, as noted in the examinations of neonates and young specimens. The next step was location of the region marking ap- proximately 1 yr of bone growth (see Res.dt.r and Fig. 4). This region was variably characterized by a lighter-stained or unstained thin band (“white rim”), sometimes with traces of an adhesion line, or in some cases no thin band. Canals in this region changed from more lengthy and horizontally packed to more shortened and irregularly packed. All other adhesion lines distal to this region were counted as subsequent years. Counts of incomplete, faint traces of lines or individual arms of bifurcated (“double layer” or “split”) lines were omitted, because the significance of these patterns has not been determined. We define the first age class (AC) as age class 0. This age class includes neonates and manatees estimated to be less than 1 yr old.
Microscope filters were used to enhance contrast. Adhesion lines were followed as far as possible in resorptive bone and matched to lines on opposite sides of intruding Haversian systems to obtain minimum counts. Photographic montages were employed when simultaneous viewing of several fields was required to count and match GLGs. Resolution of microstructure of layered, hard tissues of dolphins has been improved with the examination of thin, ground sections under a polarized light microscope (Myrick 1980). We attempted this technique on manatee bones but found no improvement over the decalcification and staining process.
Variability in counts of GLGs--Histological sections of periotic bones from 1,078 manatees were read five times each by one observer (MM) in a series of blind replicates, with the time between each reading ranging from two days to several months. If three of five readings were the same, the mode was used as the age estimate; otherwise a mean was taken. This sample did not include bones with heavy resorption because of greater subjectivity in estimating age. Variance was calculated for the readings for each manatee, and the means of the variances for each age class were computed and plotted.
Determining effects of resorption on GLG counts-Resorption in bones observed microscopically was categorized as none, light, moderate, or heavy. Bones with light resorption had low numbers of Haversian systems and widely scattered

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 59
secondary osteons which did not affect GLG counts. With moderate resorption, small aggregations of Haversian systems were seen sporadically. Closer attention was required to obtain a reading, and the stage had to be moved around these aggregations to count layers. Heavy resorption was characterized by large areas covered by Haversian systems. Counting lines was difficult, and many lines had seemingly disappeared due to remodeling. Portions of the bone looked smooth and devoid of adhesion lines. Recently deposited Haversian systems obliterated earlier ones in a dense network of secondary osteons. In cases undergoing moderate or heavy resorption, a “best estimate” approach was taken by examining bone structure, the extent and degree of resorption, and the distances between visible layers. The “first-year zone” was identified or, if not visible, the first adhesion line. Interrupted lines were followed along the microscope slide and matched to lines on the opposite sides of intruding Haversian systems to obtain minimum counts. When large areas were affected by resorption, the number of lines missed was estimated on the basis of the distance between GLGs, considering that the first 3-6 GLGs are wider than subsequent ones. Minimum and maximum possible ages were assigned, and the midpoint used as the age estimate. The mean of five readings was used as the “approximate age class” (-AC).
The relationship of resorption to body length, age class, and sex was examined by logistic regression analysis (Harrell and Lee 1985). This technique uses maximum likelihood estimation to model a nominal, binary, or ordinal depen- dent variable as a linear function of both continuous and categorical predictors. A SAS statistical procedure, LOGISTIC (SAS Institute Inc. 1990), was used to sequentially add terms for length, age class, and sex to logistic regression models that included the ordered resorption categories as the dependent variable. Values of length and age were log-transformed to correct for curvilinearity detected in arithmetic scale, and sex was recoded to a numeric “dummy” variable (0 = female, 1 = male). Resorption, which consisted of four ordinal levels (O- 3), was regressed against the predictors in a stepwise sequence. Cases where resorption interfered with age determination were excluded from the analysis. At each step, maximum likelihood estimates of regression coefficients were calculated, including standard errors and chi-square test statistics, for association, improvement of fit, and lack of fit. We tested hypotheses that resorption was the same for males and females, after adjusting for differences in length and age. In addition, we tested for relationships of resorption with age class and length, adjusted for sex.
RESULTS
Presence of Growth-Layer Groups in Bones
A small series of bones was examined in several manatees of known or approximately known age (Table 1). GLGs were most consistently found in the dome portion of the periotic bone. Layers were also clearly defined in the stapes and tympanic bone. Numbers of layers in the stapes did not correspond to those in the periotic bone consistently and often were fewer (Table 1). This was

Table I. Florida manatee bones examined for growth-layer groups and corresponding age-class estimates Samples characterized by heavy resorption are indicated with an asterisk (*). See text for the basis of actual-age age is indicated by (->. Abbreviations: St = stapes; Ty = tympanic; Do = dome; Ma = mastoid, Rb UI = ulna; Px = phalanx; Cp = carpal, Pv = pelvic bone; M = mandible; Hy = hyoid.
Identification number Age St TY Do Ma Rb Hm Rd Ul
UF 20758 2.0 0 1 1 2 4 UF 20774 -2.5 4 UF 23993 -3.5 4 ; 1 4 SWFTm-9002B -3.5 2 : 2 UF 20757 34 2 ; 4 3 UCF-9 13 1 >7 7 7 6 KDL-882 7 7.9 7 7 7 7 SWFTm-83 12B >9.0 11 11 8 SWFTm-7918B 12 10 11 8 MNE-9 102 >17 10 -19 10 SWFTm-92 15B >27 -29 7 SWFTm-8646B >21 1 :* -18 12+ 12 11 5 5 UF 20773 old >3* >2* -27 >8* +15

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 61
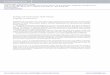
1 CM
Figure 1. Manatee tympano-periotic complex (right side). A, dorsolateral view, anterior at top; B, stapes. 1 = tympanic bone; 2 = periotic dome; 3 = mastoid portion of the periotic bone; 4 = petrous portion of the periotic bone. Dashed lines represent planes of section. The middle dashed line in the center of the complex shows the preferred plane of section for preparation of histological slides for age estimation.
verified in 17 manatees of unknown ages. In this sample the number of layers in the stapes equalled that in the dome in only seven cases. Tetracycline labels were never identified in the stapes (n = 5). GLGs in tympanic bones of young animals (up to age-class 7) were consistent with counts in the dome area in samples of known or approximately known age (Table 1). However, in a sample of 30 manatees of unknown ages, correspondence in number of GLGs between the tympanic and dome occurred in only 17 cases, and in 15 of these GLG counts were 0 or 1. Ribs, mandibles, humeri, radii, ulnae, pelvic bones, pha- langes, carpal bones, and hyoid bones did not reliably show consistent patterns of layering relative to the periotic dome, known age (Table l), or time since tetracycline administration. Although all portions of the tympano-periotic bone sectioned (tympanic, dome, and mastoid; Fig. 1) contained layering, the number of layers was not consistent among these portions in the known or approximately known age sample (Table 1). This was also found in a comparison of GLG counts among these bones in a sample of 56 individuals of unknown age. The highest number of layers, corresponding most closely to actual age of known- age individuals, was found in the dome portion.
Under microscopic examination, the innermost area of the periotic dome consisted of a primary reticular vascular tissue, where primary osteons (vascular canals not surrounded by Haversian lamellae) formed an unorganized, irregular reticulum. Primary osteons had l-3 lamellae and rounded osteocytes. This primary reticular area was observed in the dome and periotic portions and was

6 2 MARINE MAMMAL SCIENCE. VOL. 12. NO. 1. 1996
Figure 2. Microscopic appearance of bone structure in the periotic dome of a near-term manatee fetus (MNW-8906F). PV, primary vascular bone; SV, secondary vascularbone. Original magnification 40×.
followed by a wide band of secondary reticular vascular tissue. These two typesof bone were the only kinds present in periotic domes of neonates.
In some cases GLGs in the periotic dome were variable in definition, withirregular and tortuous deposition. Bifurcated layers were observed. Spacingbetween layers was not equal, but overall there was a tendency for layers to becloser together as they approached the periosteal edge. GLGs were more easilydiscerned in the central third of the microscopic section of the periotic dome.Therefore examination of the dome for age estimation and measurements oflayers were performed in the central third of the section.
Case Histories
GLGs in periotic bones of neonates and calves— In young manatees the perioticdome was primarily composed of reticular vascular tissue. A small fetus (KDL-8738), a near-term fetus (MNW-8906F, Fig. 2), and two perinatal cases (MSE-8914 and MSE-8915) presented both primary and secondary vascular tissue.The primary vascular tissue (PV, Fig. 2) had a porous appearance, with irregularly

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 63
spaced vascular canals and osteocytes. The secondary vascular tissue was denser and contained fewer pores. Small (presumably very young) calves had variable amounts of dense compact bone as well as primary and secondary reticular tissue (Fig. 3). This was also the pattern in a captive-born manatee (“Corky”) that died at 8 d old. Osteocytes were more organized in the compact bone. Vascular canals were oriented primarily parallel to the bone’s surface, giving the tissue a more regular and compact appearance.
The periotic dome of manatees in their first year exhibited a wide band (Fig. 4) of transition from reticular bone with circular lacunae (SV) to organized lamellar bone with flattened lacunae (CL) distal to the secondary vascular tissue. The CL band was usually darkly stained with no obvious line demarcating the outer reticular bone from the inner edge of the compact lamellar tissue. The distal edge of this darker band of compact lamellar tissue seems to represent a region marking about one year’s growth. Canals in this region are shortened and more irregularly packed. A lighter-stained or unstained band (white rim), sometimes with traces of an adhesion line, may occur in this region (Fig. 4).
Characteristics of periotic bones of calves were confirmed by examination of MJAV-8921. First seen at Blue Spring (BS-101) as a very small nursing calf in November 1988, this male was recovered dead on 20 October 1989 at slightly over 1 yr old. In addition to the early reticular material, the dome presented a broad band of compact lamellar bone, without lighter-stained bands or adhesion lines (Fig. 5). This manatee was classified as age-class 1 (between 1 and 2 yr old).
Known-age subadults and adults- A female from Blue Spring (SWFTm- 9002B, BS-87) born during summer 1986 died in January 1990 at between 3 and 4 yr old. The periotic dome was characterized by inner layers of primary and secondary vascular tissue, followed by the broad, dark, compact-bone layer and a change in structure and orientation of vascular canals typical of the region marking the end of the first year. Two thin adhesion lines were present distal to this region, marking the end of the second and third year. The adhesion line marking the third year was continuous and close to the edge of the bone. Based on the structural change marking the end of year 1 plus two adhesion lines, this manatee was assigned to age-class 3 (between 3 and 4 yr old), corresponding to true age.
UF 20774 was an orphaned neonate rescued in August 1980 at a body length of 122 cm. She died in captivity in January 1983, at a length of 186 cm and approximately 2.5-3.0 yr old. Only one tympanic bone was available. The transition between the secondary reticular vascular tissue to lamellar bone was evident. After the transition, three complete GLGs were seen ending in a thin adhesion line. Based on layering in the tympanic bone (not the periotic dome) she was assigned to age-class 3 (between 3 and 4 yr old). The count based on the tympanic bone was not reliable in older animals but may be consistent with true age in young animals (Table 1). There is no clear explanation for the discrepancy of about one year between the GLG count and the animal’s history. A different count may have been obtained had the periotic dome been available. Growth may have been abnormal in this individual. It was a runt,

64 MARINE MAMMAL SCIENCE, VOL. 12. NO. 1, 1996
Figure 3. Thin section of the periotic dome of a young manatee calf (CAS 23285).Note the transition from secondary vascular to compact lamellar bone. Original magni-fication 20×.

Figure 4. Photomicrograph (A) and line drawings (B) of whole section (original magnification 10×) and detail (original magnification 100×)from the central portion of the periotic dome of an age-class-8 Florida manatee. PV, primary vascular bone; SV, secondary vascular bone; CL,compact lamellar bone; WR, white rim. Numerals 1–8 mark the growth-layer groups.

66 MARINE MAMMAL SCIENCE, VOL. 12, NO. 1, 1996
Figure 5. Thin section of the periotic dome of an approximately 1-yr-old, individuallyrecognizable manatee from Blue Spring (MJAV-8921). Note the transition from secondaryvascu1ar (SV) to compact lamellar (CL) bone and the change from regular to irregularlypacked vascular canals (first-year zone, 1) at the top of the figure. Original magnification4 0 × .
based on its total length at death compared to other manatees of similar age(Fig. 6).
KDL-8827 was a Blue Spring female (BS-36) born in May 1980 and founddead 7 yr 11 mo later. This female had successfully given birth at age 5 and

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 67
360
320
290
z 200
g 230
z y 200
2
rr!
170
140
110
350
320
290
E 260
I I- 230
iii 2 200
a I-
e
170
140
110
a0
Blue Spring sample
I f 171 I (4) (7)
1 2 3 4 5 6 7 a 9
YEAR CLASS
Carcass sample
(1031
181,
Ill I 134) (421
‘I371 (491
0 1 2 3 4 5 6 7 a
AGE CLASS
Figure 6. Total body length and age for wild, known-age Florida manatees from Blue Spring (top) measured alive (O’Shea and Reep 1990), and a separate group of dead manatees collected throughout the range (bottom). Age classes of dead manatees were estimated based on growth-layer groups in the periotic dome. Solid lines and squares represent ranges and means, respectively. Numbers next to means are sample sizes.

68 MARINE MAMMAL SCIENCE. VOL. 12. NO. 1, 1996
Figure 7. Photomicrograph of thin section of the periotic dome of a 7.9-yr-old,individually recognizable manatee from Blue Spring (KDL-8827). Note the change relativeto the first-year zone (1). Numerals 2–7 mark growth-layer groups. Original magnification2 0 × .
was pregnant at death. The periotic dome exhibited the region marking the firstyear of growth, with an additional six GLGs with thin adhesion lines. In theperiotic dome section (Fig. 7), the white rim was represented by a thin line notconspicuous throughout the bone. Lines 5 and 6 both presented a bifurcatedpattern of double lines. The outermost tissue consisted of a light band and wasnot a constant feature on the surface. It measured about half (53.3 µ m) of thewidth of the immediately preceding layer (110.8 µ m). The animal was judgedto be age-class 7 (between 7 and 8 yr old), which agrees closely with true age.Evidence of moderate resorption was present, and lower GLG counts wererecorded in other bones (Table 1).
GLGs in known minimum-age manatees— SWFTm-9215B was a large female(350 cm, 1,216 kg) first sighted at Blue Spring as an adult accompanied by acalf in winter 1970–1971 (O’Shea and Hartley, in press). She was lactating atdeath in August 1992. Field observations suggest she was at least 26 yr old(assuming the calf accompanying her in 1970 was her first and she gave birthat age 4). The periotic had undergone heavy resorption, and only an approximateage could be obtained (Fig. 8). The “best estimate” was age-class 29, consistentwith the known minimum age.
M-453 was a female first identified as an independent subadult on the AtlanticCoast in winter 1978–1979. Based on size she was assumed to be l–3 yr oldat the time. The 292-cm carcass was recovered in May 1985, 6.5 yr after the

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 6 9
Figure 8. Thin section of the periotic dome of an individually recognizable femalemanatee from Blue Spring (SWFTm-9215B) that had been known as an adult for aminimum of 22 yr. Original magnification 40×.
first sighting; thus, the animal probably was between 7.5 and 9.5 yr old atdeath. The periotic dome showed a transitional area corresponding to the endof the first year, and six additional complete GLGs. This configuration resultedin a categorization as age class 7 (between 7 and 8 yr old). This estimatecorresponds with the time since initial sighting and its estimated age at thattime, as well as with lengths of known 7–8-yr-olds (Fig. 6).
UF 24971 was a large (387 cm) female from Crystal River first identifiedas adult in winter 1977–1978 and found dead in 1984. Fifteen GLGs wereseen in addition to the first year’s growth. Adhesion lines included double,multiple, and faint lines (Fig. 9). She was classified as age-class 16 (between16 and 17 yr old).
GLGs in tetracycline-marked free-ranging manatees— UF 23993 was firstidentified at Blue Spring (BS 47) in November 1981, when he was the size ofa recent weanling (220 cm). He was assumed to have been born during 1980.In February 1983 he was captured and administered tetracycline. The 265-cmcarcass was recovered 15 mo later. The periotic dome section had early reticulartissue followed by a broad, darkly stained band of dense compact bone endingat a white rim marking the end of the first year. This was followed outwardlyby two complete GLGs. A third GLG was incomplete, with a narrow band oftissue following the second adhesion line (Fig. 10). Based on the GLG count,UF 23993 was age-class 3 (at least 3 but less than 4 yr old), an estimatecompatible with its history. Tetracycline was incorporated into the GL at thetime of injection and was followed the next year by another GLG including an

Figure 9. Drawing (A) and photomicrograph (B) of thin section of the periotic dome of UF 24971, a known minimum-age manatee whereknown age was at least 9, and estimated age was at least 16. Note the change in structure in the first-year zone (1). Numerals 2–16 markgrowth-layer groups. Original magnification 40×.

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 71
Figure 10. Photomicrograph (A) and drawing (B) of thin section of the perioticdome of an approximately 3–4-yr-old, individually recognizable manatee from Blue Spring(UF 23993). Note the change relative to the first-year zone (1). Numerals 2–3 markgrowth-layer groups. Original magnification 20×.

72 MARINE MAMMAL SCIENCE. VOL. 12. NO, 1, 1996
Figure 11. Photomicrograph of thin section of the periotic dome of a 343-cm (totallength) female from Blue Spring (MNE-9102), known alive from 1974 to 1991. Originalmagnification 40×.
adhesion line. Additional tissue corresponding to formation of a new GLG alsowas present. The distance between the label and the outer edge of both bonescorresponded to the location of the second outermost layer in the stained sections.The tetracycline label confirmed that a GLG corresponds to one year in theperiotic dome.
MNE-9102 was a female first seen at Blue Spring in 1974 as an adult. InFebruary 1983 she was administered tetracycline in the field. Eight years andone month later she was found dead at 343 cm total length. Moderate resorptionand a pattern of double and closely appressed GLGs allowed only an approx-imation of age (Fig. 11). The average of five readings resulted in an estimatedage of 19 yr (possible range 17–20). The tetracycline label was narrow, dull,and discontinuous but still recognizable under ultraviolet light. The distancebetween the periosteal edge and the label (306 µ m) corresponded to the areabetween the 7th and 8th adhesion lines inward from the periosteum in thestained section (346 µ m). This corresponds to the elapsed time of eight yearsand one month from tetracycline administration to death.
UF 20773 was a Blue Spring male (BS-6) first observed as an adult in 1970and recovered dead in November 1982 (338 cm total length). He was capturedand administered tetracycline 22 mo prior to death. A thin section of the perioticdome viewed under ultraviolet light showed a tetracycline label in several smallsegments, with an average distance of 26.89 µ m from the periosteal edge.Analysis of the stained section equivalent to the same region produced an average

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 73
distance for the outermost layer from the edge of 12.3 7 pm, and for the second 25.57 pm. Thus, the two outer broad bands represent approximately two years, corresponding to the 22 mo elapsed between tetracycline administration and death. The “best estimate” approach yielded an estimate of AC -28 with a range of 26 to 32 GLGs (Fig. 12). The tissue was undergoing heavy resorption, and some previously existing layers may have been obliterated. This estimate is compatible with the animal’s history.
SWFTm-8646B was a large female from Blue Spring administered tetra- cycline in January 1981 and recovered dead 66 mo later. SWFTm-8646B was first recorded as an adult with a calf in winter 1970-1971. If the calf with her in 1970 was her first and she gave birth at age 4, this manatee could be no younger than 20 yr old at death. A range of 15 to 24 GLGs, with a best esti- mate of 19 (AC - 19) was obtained from the periotic dome. Closely appressed adhesion lines, two double and three multiple, were counted as one GLG each. Tetracycline was observed as a few short labels (varying from 185 to 707 pm in length) very close to the edge. The average distance between the tetracycline label and the outer edge of the dome (66 pm> was smaller than the distance from the edge to the 4th (135 pm> and 5th (186 pm> outermost adhesion lines. If the fluorescence matched the stained section, the label should have fallen in that vicinity. The reduced number of layers relative to minimum age in SWFTm-8646B may be attributed to the presence of heavy resorption. Re- sorption, and perhaps changes in growth, may also account for the presence of tetracycline label in regions inconsistent with GLGs interpreted as annuli.
GLGs in tetracycline-marked captive manatees-UCF-9 13 1 was a subadult- sized male rescued in May 1984 at 211 cm total length. Tetracycline was administered at 81 and 42 mo before its death in August 1991 (275 cm total length). The dome portion of the periotic showed seven complete GLGs, followed by a band of compact bone, and no resorption. Therefore, it was estimated to be between 7 and 8 yr old at death. This is compatible with likely true age. The periotic dome section showed two fluorescent labels, 960 pm and 576 pm from the outer edge. In the stained sections the distance from the periosteal edge to the second label encompassed three complete and one incomplete GLG, in accordance with the three years and six months since tetracycline administra- tion. The distance from the periosteal edge to the first label contained six complete GLGs, parts of the most recent GLG, and a portion of bone prior to the white rim. This also corresponds to the six years nine months since administration.
SWFTm-7918B was a female rescued as a neonate in August 1979 and kept captive until death 12.2 yr later. During 1981 she received intramuscular injections of tetracycline on 9 January and 16 July. This represents the longest labeling record in the sample (i.e., 12 5 and 13 1 mo prior to death). The estimated age obtained through a GLG count was AC 11 (between 11 and 12 yr old). Resorption on the dome was light. Under ultraviolet light the untreated section of the dome revealed two clear tetracycline labels. The innermost label was 1,296 pm inward from the periosteal edge, falling just above the location of the first-year zone in the stained section (1,3 52 pm). The second label was 384 pm away from the first, corresponding to an area below the first conspicuous

Figure 12. Photomicrograph (A) and line drawing (B) of thin section of the periotic dome of an adult, individually recognizable manateefrom Blue Spring (UF 20773) known from 1970 to 1982. Note the Haversian systems (HS) and a large erosion bay (EB) indicating resorption.Numerals 2–26 mark growth-layer groups. Adhesion lines corresponding to growth-layer groups 23–26 appear throughout the section as thinnerlines than the previous, Original magnification 100×.

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 75
Figure 13. Photomicrograph of the thick section of the periotic dome of UF 20758under ultraviolet light. The fluorescent band represents tetracycline deposition in the bonetissue. Original magnification 40×.
adhesion line (i.e., GLG 2) at 891 µ m from the periosteal edge of the stainedsection. This agrees with SWFTm-7918B’s history of tetracychne administration.
SWFTm-8321B was a subadult-sized (231 cm) female rescued in August1983. She received tetracycline in November 1984 and died in captivity inSeptember 1992 (316 cm). Examination under ultraviolet light did not reveala label in the periotic dome. Tetracycline lines in the tympanic and mastoidportions were continuous, but thin and faint. The stained section of the domeshowed 11 GLGs. The first-year zone was followed by a second faint line(corresponding to GLG 2) without a clear adhesion line. Adhesion line 3 wasdouble, and all subsequent lines were single. A considerable amount of bonetissue was deposited after adhesion line 11, and resorption was light. Based onGLGs, this manatee was between 11 and 12 yr old. This estimate is reasonableif she were l–3 yr old when rescued (an age compatible with size).
UF 20758 was a female born in captivity in August 1980. She was admin-istered tetracycline in March 1981 and died in August 1982. Distinctive adhesionlines were not present in the periotic dome, but two adhesion lines (one of whichwas bifurcated) appeared in the stapes, and one in the tympanic bone. In theperiotic dome, the core had reticular vascular bone, followed by a wide, darkband of compact lamellar bone. The “white rim” was followed by a wide bandof tissue, with more irregularly oriented canals. Based on this structure, she wascategorized as age-class 1 (between 1 and 2 yr old), which was an underestimateof true age (2 yr, 25 d). The untreated slide examined under ultraviolet light

76 MARINE MAMMAL SCIENCE, VOL. 12, NO. 1, 1996
showed a strong tetracycline line 1,200 pm inward from the periosteal edge. The white rim area was distal to the tetracycline line (260 gm), confirming the chronology of deposition of this region (Fig. 13).
UF 20757 was a subadult-sized (247 cm) female rescued in September 1979 and treated with tetracycline therapeutically. Death occurred in September l980. The periotic dome section was composed of primary and secondary vascular tissue bordered by a lightly colored narrow band. This band also marked a region where tissue structure changed from lamellar to more compact bone with fewer vascular canals, marking the first year of bone growth. Three subsequent GLGs followed (Fig. 14), with a small amount of tissue present distal to the last adhesion line. Based on these criteria, UF 20757 was assigned to age class 4 (between 4 and 5 yr old). This classification is consistent with the range of ages associated with body lengths of wild manatees similar to that of UF 20757 at capture (Fig. 6) and the additional year in captivity. Repeated therapeutic administration of tetracycline to UF 20757 resulted in distinct labeling of some bones. Only a very short amount of label (48 pm) was seen in the dome portion of the periotic bone, and this was very close to the edge (11 pm). Wide bands of tetracycline were present on the mandible (both longitudinal and transverse sections). Despite high dosage, tetracycline was not evident in the ribs.
GLGs in Relation to Size, Sex, and Resorption
Heavy resorption accompanied by GLG counts lower than known minimum age in a key specimen (SWFTm-8646B) suggests that resorption decreases reliability of GLG counts for age determination in adult manatees. To investigate this problem further, data were analyzed for 1,2 12 manatees (16 known-age and 1,196 additional specimens) classified on the basis of resorption category, age class, total body length, and sex. In most cases resorption was not observed, or was classified as light, in either sex until total body lengths exceeded 300 cm (Fig. 15). Resorption also was not observed, or was classified as light to moderate, in age classes up to about 15 yr (Fig. 16).
Analyzed separately, both age and length were highly significant (P < 0.0001) when added to logistic regression models of resorption, indicating that older and larger animals were more likely to show the highest resorption levels regardless of sex. Adding length to a model that already contained sex and age, however, did not significantly improve the fit (P = 0.0941), but age remained sigmficant (P < 0.0001) (Table 2). This indicated that resorption increases with age independent of length. The apparent relationship between resorption and length, therefore, was largely caused by the correlation of length with age (Fig. 6). There was also a highly significant sex effect (P < O.OOOl>, because females were more likely to have higher resorption levels after adjusting for age and length differences (Table 2). There were no interactions involving sex with age or length (P > 0.45).
To verify relationships between body length and age, we compared age-class assignments based on GLG counts from dead specimens with the range of total- length measurements determined from a separate group of living, known-age

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 77
Figure 14. Photomicrograph (A) and line drawing (B) of thin section of the perioticdome of UF 20757, a manatee of unknown age, estimated to be age-class 4. Note thechange from regularly to irregularly spaced canals in the first-year zone (1). Numerals 2–3 mark growth-layer groups. Original magnification 40×.

78 MARINE MAMMAL SCIENCE, VOL. 12, NO. 1, 1996
200
150
h k$ ioc W
B
50
C -
F
- N I F
120- Wl- 241- 180 240 300
-
M FM FM SEX
> 300 SIZE CLASS (CM)
Figure 15. Frequency of manatees with no, light, moderate, or heavy resorption of bone in the periotic dome, classified by sex and size class.
wild manatees from Blue Spring (Fig. 6). Field data for this comparison were from wild manatees measured each year up to age class 9, with body lengths approaching a maximum of 300 cm (O’Shea and Reep 1990). This comparison also suggests that GLG counts in the periotic dome are accurate estimates of age for manatees up to at least 300 cm total length and through the first 10 yr of life.
Variability in GLG Counts
Best precision was obtained in the lower age classes where GLGs were usually wide and better defined and resorption was minimal. Variability in replicate

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 79
300
250
6
6”
&o
& 100
50
0
FM FM FM FM FM FMSEX O-3 4-7 8-11 12-15 >15 ? AGE
CLASS
RESORPTION I NONE CZZZI LIGHT BZB MODERATE m HEAVY
Figure 16. Frequency of manatees with no, light, moderate, or heavy resorption of bone in the periotic dome, classified by sex and age class.
counts of GLGs increased with age, especially after age class 10 (Fig. 17). However, sample sizes also decreased in the older age categories.
Maximum Age-Class Assignments and Manatee Longevity
Despite resorption-based interference in age estimation through GLGs, some individuals had high counts. For example, 59 GLGs were estimated in UF 15 115, a 362-cm female; -37 in M-229, a 309cm male; and several large animals over 300 cm had GLG counts of over 20. Such GLG counts are consistent with other knowledge of manatee longevity in captivity and from field studies (Table 3).
GLGs in Bones of Antillean Manatees
Periotic domes from 18 specimens of Trichechus manatus manatus were examined ( 17 from Puerto Rico and 1 from Belize). Body lengths ranged from 119 to 33 5 cm. The depositional pattern conformed with that described for Florida manatees. A typical section consisted of reticular tissue, followed by lamellar tissue bordered by a poorly stained band. A pattern of bands and adhesion lines followed, with some cases of double and split lines, especially in

80 MARINE MAMMAL SCIENCE, VOL. 12, NO. 1, 1996
T&e 2. Parameter estimates and test statistics for logistic regression of resorption against age class, length (cm), and sex based on GLG counts in the periotic dome of Florida manatees (n = 991)a.
Variable Parameter Standard estimate error Chi-squarec P
Age dassb -15.1546 1.2257 Lengthb -7.5817 4.5292 Sex 0.9873 0.2529
a Excludes all cases of heavy resorption. b Log transformed. ‘df = 1.
152.8582 <0.0001 2.8021 0.0941 15.2374 <0.0001
larger specimens. Similar to the pattern seen in Florida manatees, resorption appeared to be more intense in older specimens and affected readings of animals greater than AC 20.
DISCUSSION
GLG counts in the dome portion of the periotic bone of Florida manatees provide a reliable technique for age estimation. Age estimates based on this method appear to be especially accurate for individuals ranging up to about 10-l 5 yr old with body lengths up to about 300 cm. The procedure was verified by case histories of 17 manatees that were either known age, known minimum age, or had tetracycline markers. Case studies of three wild manatees from Blue Spring ranging from about 1 to nearly 8 yr old showed close agreement between estimated age and true age. Location of fluorescent marks and subse- quent layering patterns confirmed that GLGs were annual and corresponded well to time since tetracycline administration in three free-ranging and three captive manatees. Double in vivo labeling with tetracycline in SWFTm-7918B and UCF-9131 further confirm the finding of one GLG per year. SWFTm- 7918B also shows that tetracycline marks may persist even 10 yr after admin- istration. Fluorescent marks and GLG counts did not correspond with known minimum age or elapsed time since tetracycline administration for one free- ranging adult female from Blue Spring. However, this specimen exhibited heavy bone resorption. Analysis of other specimens also showed that resorption was likely to interfere with age estimation for larger, older manatees.
Examination of manatees younger than 1 yr revealed that changes in cell types, packing, and shape and orientation of vascular canals were keys to assignment of age categories based on early growth. Although neonatal lines representing the time of birth have been identified in teeth of odontocetes and dugongs (Marsh 1980, Perrin and Myrick 1980), a neonatal line is not present in periotic domes of manatees. Neonatal lines consist of a thin translucent or opaque layer thought to be due to disturbances in nutrition in the immediate postpartum period (Perrin and Myrick 1980). The identification of a neonatal

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 81
40
39 T T
0 2 4 6 8
19 23
II
0
22 24 T T
5
7
IL!
4
10 12 14 16 18 20 22 24 26 28 AGE CLASS
J Figure 17. Variance among five replicate growth-layer-group counts for 1,078 Florida
manatees. Squares indicate the mean variance for each age class, vertical lines show the range and numbers above the lines are sample sizes.
line is usually based on specimens caught soon after birth; our sample contained numerous small calves, but a neonatal line was not detected in any of them.
Unlike some mammals, the first year in manatees is marked by a zone of osteological change with a variable appearance, but it is not typically or consis- tently characterized by an adhesion line. This first-year zone was characterized by a change from lengthy and horizontally packed canals to more shortened and irregularly packed canals, and may occur with or without a lightly stained or unstained band (“white rim”) or traces of an adhesion line.
The first strong adhesion line seems to be formed at the end of the second year, as demonstrated in some known-age animals. For example, MJAV-8921, known from the field to be older than 1 but less than 2 yr old, had a first-year zone in the periotic dome, but no adhesion lines. Manatee SWFTm-9002B was known to be between 3 and 4 yr old, and had her first dark adhesion line 2 GLGs inward from the outer edge of the periotic dome. Similarly, GLG counts in the dome of SWFTm-8646B (the 7.9-yr-old wild female) from the edge of the bone inward showed an innermost (first) adhesion line corresponding to year 2.
For mammals in general, deposition of thin adhesion lines corresponds to a seasonal slowing of growth in winter months (Klevezal’ 1980, Klevezal and

82 MARINE MAMMAL SCIENCE, VOL. 12, NO. 1, 1996
TabZe 3. Examples of minimum ages of living individual Florida manatees recorded in field studies and in captivity. For field studies, CR = Crystal River and BS =: Blue Spring. Year of most recent record for current status is 1994 for captives and 1992 for field studies, except as noted.
Individual
Year of birth/ capture
Captives
Relative age at first sighting
Current status
Elapsed time
Snooty d Romeo 8 Juliet P Rosie Q
Individual
1948 1957 1958 1968
Year of first sighting
born 21 July young adult adult adult
Field Studies Relative age at
first sighting
living 46 yr living 37 yr living 36 yr living 26 yt
Current Elapsed status time
BS-19 $ BS-11 d BS-02 d CR-28 Q CR-60 Q CR-25 8 CR-06 8 CR-21 8
1970 adult living 22 yr 1970 adult living 22 yr 1970 adult living 22 yr 1967 adult 1967
l living 23 yr unknown
1967 *living 24 yr
unknown 3iving 22 yt 1967 juvenile 3 living 22 yr 1967 unknown ‘living 23 yr
l Through 1990. * Through 1991. 3 Through 1989.
Kleinenberg 1969). Absence of strong demarcations in the first GLG in bone is considered to be due to lack of pronounced fluctuations in growth rates during early life (Klevezal’ and Kleinenberg 1969). Seasonal changes in environmental conditions affecting growth and nutrition beyond the early years have been assumed to result in differential widths of GLs in dugong tusks (Marsh 1980). Hypocalcified dentinal layers in GLGs of dusky dolphins (Lagenorhyncus obscu- rUS) were hypothesized to reflect environmental changes under El Nino conditions (Manzanilla 1989). Other studies, however, indicate that formation of growth- layer groups in marine mammals may be largely based on endogenous rhythms (Hohn et al. 1989). Our finding of well-defined GLGs in periotic bones of specimens from Puerto Rico and Belize (where seasonal climatic changes are more moderate than in Florida) suggests that exogenous effects alone would not fully account for adhesion-line formation in manatees. Examination of manatees from these regions also suggested that age-determination studies using the periotic dome may be extended to the other subspecies.
In bones other than the periotic dome, layering was absent or negligible due to rapid resorption (e.g., pelvic bone mandible) or did not correspond well to age (ribs, forelimb, stapes, tympanic). Previous investigators have speculated that GLGs in some of these other bones (particularly mandibles and ribs) might

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 83
have utility in age-determination studies of sirenians (Odell 1977, Mitchell 1978, Domning and Myrick 1980, Marsh 1980). However, our results indicated that they are less useful for age determination than the periotic dome. GLs in the periotic dome were easier to count and were more consistent with known age. This finding is in accord with the suggestions of Myrick and Kelly (un- published manuscript) and Myrick (1985), who provided initial descriptions suggesting maximum GLG counts would be found in periotic bones. However, these investigators also suggested that periotic bones do not undergo significant resorption, a finding inconsistent with our results based on a larger sample. We also did not see evidence for readily discernible lunar-monthly and daily GLs reported by Myrick and Kelly (unpublished manuscript).
Limitations to the use of GLG counts in the periotic dome for age deter- mination are related to the histomorphological pattern of Haversian substitution and osteon accumulation with increasing age that is common in mammals (Ortner 1975, Martin and Burr 1989). In the process of remodeling, periosteal bone is replaced by secondary osteons and previously formed layers disappear. Bone is an intensely active tissue that plays a major role in mineral homeostasis, and resorption of bone is a complex phenomenon. The efficiency of utilization of ingested calcium also decreases with age (either due to decreased resorption from the gastrointestinal tract or poor retention by tissues), even with large amounts of available calcium (Hansard et al. 1954). Abnormal resorption may also be triggered by pathologic processes or chronic dietary mineral and vitamin im- balance (Maynard and Loosli 1969, Hancox 1972, Underwood 1981). The amount of resorption may also vary with the calcium content of the diet as well as age (Gaster et al. 1967, McLean and Urist 1968). Bone mineral reserves are reduced during periods of curtailed feeding, enforced immobility, hibernation, and hypothermia (Bruce and Wiebers 1969, Steinberg et al. 198 1, Kwiecinski et al. 1987). Pregnancy and lactation cause calcium deprivation regardless of the amount of calcium in the diet and can impose additional stress on miner- alization of bones in females (Atkinson and West 1970, Rasmussen 1977, Pitkin et al. 1979, Kwiecinski et al. 1987).
Variability in the amount of resorption in bones of manatees may be expected to be high, allowing more accurate counts of GLGs in some individuals than in others. In Florida individual manatees are known to differ in migratory patterns (Rathbun et al. 1990, Reid et al. 1991), spend differing amounts of time feeding in areas with markedly different ecological characteristics (Zoodsma 1991), vary in reproductive history (O’Shea and Hartley, in press; Rathbun et al., in press), and have differing histories of exposure to environmental stresses such as cold temperatures, disease, and food deprivation (Buergelt et al. 1984, O’Shea et al. 1985). When adjusted for length and age, results of logistic regression indicated that earbones from female manatees are more prone to resorption than earbones from males. This is consistent with higher reproductive demands for calcium in females.
Compared to other bones, resorption in the periotic dome of manatees tends to occur at a later age, and early layers are less likely to be obliterated. In some cases vestiges of adhesion lines retained between the osteons allowed an ap-

84 MARINE MAMMAL SCIENCE, VOL. 12, NO. 1, 1996
proximate count. In others resorption was extensive, the number of layers was reduced, and accurate age estimates were not possible. Counting GLGs in older animals is also difficult because, as calcium deposition decreases with age, the layers become compressed towards the periosteal edge. This conforms with a general mammalian pattern representing a reduction in growth with age (Kohn 1978, Peters 1983). Furthermore, GLG counts in older animals are often more subjective due to incomplete, closely appressed, or double lines. This was apparent in some of our specimens (e.g., SWFTm-8646B and UF 20773, Fig. 12), where counting each line could have resulted in higher age estimates. We were conservative, however, and omitted counts of secondary adhesion lines as annuli due to evidence (e.g., KDL-8827) that double lines can occur within single years.
All criteria for age determination are subject to error, which may include random variation about some true age, as well as bias due to methodology (Kimura and Lyons 1990). Methods based on annually repeating characteristics tend to produce the best results (Caughley 1977). Although investigators disagree on the consequences of errors in age determination (Caughley 1977, Robinette et al. 1957), it is clear that errors are unavoidable. In addition, age determirnation is a statistical process with an intrinsic level of variability (Kimura and Lyons 1990). Based only on a “within-reader” estimate of error for a single reader of a large sample, we determined that best precision in age estimates was obtained at the lower end of the age scale. This should at least minimize error in the calculation of age-specific reproductive rates for age classes corresponding to sexual maturation (Mansfield 199 1). Factors affecting precision and accuracy in manatee age estimates include the complexity of the depositional pattern in bone, the degree of resorption, and poor preparation in brittle specimens. Despite such limitations, results of this research indicate that GLG counts provide the best means available for age estimation in manatees, and we recommend both application and further development of the technique.
ACKNOWLEDGMENTS
This study was supported by the National Fish and Wildlife Foundation, the US. Marine Mammal Commission (contract T 6223918-l), and the U.S. Fish and Wildlife Service. Marmontel was the recipient of scholarships from the Coordenasb de Aperfei- coamento de Pessoal de Nivel Superior and the Conselho National de Pesquisas, Brazil. Comments on versions of the manuscript were made by J. Eisenberg, D. Laist, L. Lefebvre, R. Reep, M. Sunquist, S. Wright, and anonymous reviewers. J. Shoemaker assisted. with final preparation of figures. Special thanks go to A. Hohn for advice in developing techniques and interpreting an initial series of histological slides, G. Rathbun for foresight in initiating tetracycline administration programs, R. Bonde and C. Beck of the Sirenia Project, and numerous individuals with the National Biological Service, Florida Depart- ment of Environmental Protection, the University of Florida, Sea World of Florida, Miami Seaquarium, and other oceanaria for critical assistance. L. Lefebvre provided institutional support during the final phases of the study. The following museums provided access to skeletal specimens: the Burke Memorial Washington State Museum; the California Acad- emy of Sciences; the Carnegie Museum of Natural History; the Florida Museum of Natural History; the Los Angeles County Museum of Natural History; the Museum of

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 85
Natural and Cultural History, Oklahoma State University; the Museum of Southwestern Biology, University of New Mexico; the North Carolina Museum of Natural History; the Oklahoma Museum of Natural History, University of Oklahoma; the Orman J. Smith Museum of Natural History, Albertson College of Idaho; and the U.S. National Museum of Natural History, Washington, DC. Specimens were collected under authority of U.S. Fish and Wildlife Service endangered species permit PRT-684532.
LITERATURE CITED
AMPRINO, R., AND A. ENGSTRoM. 1952. Studies on x-ray absorption and diffraction of bone tissue. Acta Anatomica 15: l-22.
ATKINSON, R. J., AND R. R. WEST. 1970. Loss of skeletal calcium in lactating females. Journal of Obstetrics and Gynaecology of the British Commonwealth 77:555-560.
BEST, R. C. 1982. A salva@o de uma especie: novas perspectivas para o peixe-boi da Amazonia. Revista IBM, 14 (December), pages not numbered.
BONDE, R. K., T. J. O’SHEA AND C. A. BECK. 1983. Manual of procedures for the salvage and necropsy of carcasses of the West Indian manatee (Trichechus manatus). PB 83-25 5272. National Technical Information Service, Springfield, VA. 175 pp.
BRUCE, P. S., AND J. E. WIEBERS. 1969. Mineral dynamics during hibernation and chronic immobilization: a review. Aerospace Medicine 40:855-861.
BUERGELT, C. D., R. K. BONDE, C. A. BECK AND T. J. O’SHEA. 1984. Pathologic findings in manatees in Florida. The Journal of the American Veterinary Medical Association 185:1331-1334.
CASTAGNET, J. 1974. Etude histologique des marques squelettiques de croissance chez Vipera aspic (L.) (Ophidia Viperidae). Zoologica Scripta 3: 137- 15 1.
CASTAGNET, J. 1980. Quelques remarques sur la methode squelettochronologique chez les vertebres superieurs (oiseaux et mammiferes). Bulletin de la Societe Zoologique de France 105:371-387.
CAUGHLEY, G. 1977. Analysis of vertebrate populations. John Wiley & Sons, Lon- don, UK.
DOMNING, D. P., AND L.-A. C. HAYEK. 1984. Horizontal tooth replacement in the Amazonian manatee (Trichechus inunguis). Mammalia 48: 105-127.
DOMNING, D. P., AND A. C. MYRICK, JR. 1980. Tetracycline marking and the possible layering rate of bone in an Amazonian manatee (Trichechus inunguis). Reports of the International Whaling Commission (Special Issue 3):203-207.
FROST, H. M. 1969. Tetracycline-based histological analysis of bone remodeling. Cal- cified Tissue Research 3:2 11-237.
FROST, H. M. 1983. Bone histomorphometry, choice of marking agent and labeling schedule. Pages 37-52 in R. R. Reeker, ed. Bone histomorphometry: techniques and interpretation. CRC Press, Boca Raton, FL.
FROST, H. M., A. R. VILLANUEVA, H. ROTH AND S. STANISLAVJEVIC. 1961. Tetracycline bone labeling. The Journal of New Drugs (no volume number):206-216.
GASTER, D., E. HAVIVI AND K. GUGGENHEIM. 1967. Interrelations of calcium, fluorine and vitamin D in bone metabolism. British Journal of Nutrition 2 1:413-418.
HANCOX, N. M. 1972. Biology of bone. Cambridge University Press, London, UK. HANSARD, S. L., C. L. COMAR AND M. P. RUMLEE. 1954. The effects of age upon
calcium utilization and maintenance requirements in the bovine. Journal of Animal Science 13:25-36.
HARRELL, F. E., AND K. L. LEE. 1985. The practical value of logistic regression. Pages 1031-1036 in SAS Users Group International, ed. Proceedings of the tenth annual SAS Users Group International Conference. SAS Institute Inc., Cary, NC.
HOHN, A. 1980. Analysis of growth layers in the teeth of Tursiops truncates, using light microscopy, microradiography, and SEM. Reports of the International Whaling Commission (Special Issue 3): 155-160.

86 MARINE MAMMAL SCIENCE. VOL. 12. NO. 1. 1996
HOHN, A., M. D. SCOTT, R. S. WELLS, J. C. SWEENEY AND A. B. IRVINE. 1989. Growth layers in teeth from known-age, free-ranging bottlenose dolphins. Marine Mammal Science 5:315-342.
KIMURA, D. K., AND J. J. LYONS. 1990. Between-reader bias and variability in the age- determination process. Fishery Bulletin 89:53-60.
KLEVEZAL’, G. A. 1980. Layers in the hard tissues of mammals as a record of growth rhythms of individuals. Reports of the International Whaling Commission (Special Issue 3):89-94.
KLEVEZAL’, G. A., AND S. E. KLEINENBERG. 1969. Age determination of mammals from annual layers in teeth and bones. Academy of Sciences of the USSR, Moscow, 1967. IPTS cat. # 5433. Program of Scientific Translations Ltd., Jerusalem, Israel (J. Salkind, translator).
KOHN, R. R. 1978. Principles of mammalian aging. Second ed. Prentice-Hall Foun- dations of Developmental Biology Series. Englewood Cliffs, NJ.
KWIECINSKI, G. G., L. KROOK AND W. A. WIMSATT. 1987. Annual skeletal changes in the little brown bat, Myotis lucifugus lucifugus, with particular reference to pregnancy and lactation. American Journal of Anatomy 178:410-420.
LAWS, R. M. 1952. A new method of age determination for mammals. Nature 169: 972-973.
LAWS, R. M. 1953. A new method of age determination in mammals with special reference to the elephant seal (Mirounga leonina). Falkand Islands Dependencies Survey Scientific Reports 2: l-l 1.
LAWS, R. M. 1958. Growth rate and age of crabeater seals Lobodon carcinophagus. Proceedings of the Zoological Society of London 130:275-288.
LAWS, R. M. 1960. Laminated structure of bones from some marine mammals. Nature 187:338-339.
MANSFIELD, A. W. 1958. The biology of the Atlantic walrus Odobenus rosmarus rosmarus (Linnaeus) in the eastern Canadian Arctic. Fisheries Research Board of Canada, Manuscript Report Series (Biology) 653:1-146.
MANSFIELD, A. W. 1991. Accuracy of age determination in the grey seal Halichoerus grypus of Eastern Canada. Marine Mammal Science 7:44-49.
MANZANILLA, S. R. 1989. The 1982-1983 El Nino event recorded in dentinal growth layers in teeth of Peruvian dusky dolphins (Lagenorhynchus obscurus). Canadian Journal of Zoology 67:2120-2125.
MARMONTEL, M. 1993. Age determination and population biology of the Florida manatee, Trichechus manatus latirostris. Unpublished Ph.D. dissertation, University of Florida, Gainesville, FL. 408 pp.
MARMONTEL, M. In press. Age and reproduction in female Florida manatees. In T. J. O’Shea, B. B. Ackerman and H. F. Percival, eds. Population biology of the Florida manatee. National Biological Service Information and Technology Report 1.
MARMONTEL, M., T. J. O’SHEA AND S. R. HUMPHREY. 1990. An evaluation of bone growth-layer counts as an age-determination technique in Florida manatees. Doc- ument PB-91-103564, National Technical Information Service, Springfield, VA.
MARMONTEL, M., D. K. ODELL AND J. E. REYNOLDS, III. 1992. Reproductive biology of South American manatees. Pages 295-312 in W.C. Hamlett, ed. Repro&&ve biology of South American vertebrates. Springer-Verlag, Berlin.
MARSH, H. 1980. Age determination of the dugong (Dugong dugon) in Northern Australia and its biological implications. Reports of the International Whaling Com- mission (Special issue 3): 18l-201.
MARTIN, R. B., AND D. B. BURR. 1989. Structure, function, and adaptation of compact bone. Raven Press, New York, NY.
MAYNARD, L. A., AND J. K. LOOSLI. 1969. Animal nutrition. Sixth ed. McGraw-Hill Book Co., New York, NY.
MCLEAN, F. C., AND M. R. URIST. 1968. Bone--fundamentals of the physiology of skeletal tissue. Third ed. University of Chicago Press, Chicago, IL.

MARMONTEL ET AL.: MANATEE AGE DETERMINATION 87
MITCHELL, J. 1978. Incremental growth layers in the dentine of dugong incisors (Dugong dugon Muller) and their application to age determination. Zoological Journal of the Linnean Society 62:317-348.
MORRIS, P. 1972. A review of mammalian age determination methods. Mammal Review 2:69-104.
MYRICK, A. 1980. Examination of layered tissues of odontocetes for age determination using polarized light microscopy. Reports of the International Whaling Commission (Special Issue 3):105-112.
MYRICK, A. 1985. Estimating age of young West Indian manatees, Trichechus manatus, from dentinal layers. (Abstract). Fourth International Theriological Congress, Ed- monton, Alberta, Canada.
ODELL, D. K. 1977. Age determination and biology of the manatee. Final report, U.S. Fish and Wildlife Service Contract 14-16-008-930, 125 pp.
ORTNER, D. J. 1975. Aging effects on osteon remodeling. Calcified Tissue Research 18:27-36.
O’SHEA, T. J., AND R. L. REEP. 1990. Encephalization quotients and life history traits in the Sirenia. Journal of Mammalogy 7 1:534-543.
O’SHEA, ‘I’. J., AND W. C. HARTLEY. In press. Reproduction and early age survival of manatees at Blue Spring, Upper St. Johns River, Florida. In T. J. O’Shea, B. B. Ackerman and H. F. Percival, eds. Population biology of the Florida manatee. National Biological Service Information and Technology Report 1.
O’SHEA, T. J., C. A. BECK, R. K. BONDE, H. I. KOCHMAN AND D. K. ODELL. 1985. An analysis of manatee mortality patterns in Florida, 1976-198 1. Journal of Wildlife Management 49: l- 11.
PAYNE, M. R. 1978. Population size and age determination in the Antarctic fur seal Arctocephalus gazella. Mammal Review 8:67-73.
PERRIN, W. F., AND A. C. MYRICK, JR., EDS. 1980. Age determination of toothed whales and sirenians. Reports of the International Whaling Commission (Special Issue 3).
PERRIN, W. F., J. M. COE AND J. R. ZWEIFFEL. 1976. Growth and reproduction of the spotted porpoise Stenella attenuata in the offshore eastern tropical Pacific. Fishery Bulletin 74:229-269.
PERRIN, W. F., D. B. HOLTS AND R. B. MILLER. 1977. Growth and reproduction of the eastern spinner dolphin, a geographical form of Stenella longirostris in the eastern tropical Pacific. Fishery Bulletin 75:725-750.
PETERS, R. H. 1983. The ecological implications of body size. Cambridge University Press, New York, NY.
PITKIN, R. M., W. A. REYNOLDS, G. A. WILLIAMS AND G. K. HARGIS. 1979. Calcium metabolism in normal pregnancy: a longitudinal study. American Journal of Ob- stetrics and Gynecology 133:781-787.
RASMUSSEN, P. 1977. Calcium deficiency, pregnancy and lactation in rats. Some effects on blood chemistry and the skeleton. Calcified Tissue Research 23:87-94.
RATHBUN, G. B., J. P. REID AND G. CAROWAN. 1990. Manatee (Trichechus manatus) distribution and movement patterns in northwestern peninsular Florida. Florida Marine Research Publications 48:1-33.
RATHBUN, G. B., J. P. REID, R. K. BONDE AND J. A. POWELL. In press. Reproduction in free-ranging Florida manatees. In T. J. O’Shea, B. B. Ackerman and H. F. Percival, eds. Population biology of the Florida manatee. National Biological Service Information and Technology Report 1.
REID, J. P., G. B. RATHBUN AND J. R. WILCOX. 1991. Distribution patterns of indi- vidually identifiable West Indian manatees (TrichechUs manatus) in Florida. Marine Mammal Science 7:180-190.
REID, J. P., R. K. BONDE AND T. J. O’SHEA. In press. Reproduction, mortality and tag loss of telemetered and recognizable individual manatees on the Atlantic Coast of Florida. In T. J. O’Shea, B. B. Ackerman and H. F. Percival, eds. Population

88 MARINE MAMMAL SCIENCE, VOL. 12, NO. 1, 1996
biology of the Florida manatee. National Biological Service Information and Tech.- nology Report 1.
ROBINEAU, D. 1969. Morphologie exteRNe du complexe osseux temporal chez les Sir- eniens. Memoirs du Museum National d’Histoire, Serie A, Zoologie 6O:l-32.
ROBINETTE, W. L., D. A. JONES, G. ROGERS AND J. S. GASHWILLER. 1957. Notes on tooth development and wear for Rocky Mountain mule deer. Journal of Wildlife Management 21:134-153.
SAS INSTITUTE INC. 1990. SAS technical report P-200, SAS/STAT software: CALIS and LOGISTIC procedures, release 6.04. SAS Institute, Inc., Gary, NC. 236 pp.
SCHEFFER, V. B. 1950. Growth layers on the teeth of Pinnipedia as an indication of age. Science 112:309-3 11.
SISSONS, H. A. 1949. Intermittent periosteal activity. Nature 163:1001-1002. SISSONS, H. A. 1971. The growth of bone. Pages 145-180 in G. H. Bourne, ed. The
biochemistry and physiology of bone, Volume III. Academic Press, New York, NY. STEINBERG, B., I. J. SINGH AND 0. G. MITCHELL. 1981. The effects of cold-stress,
hibernation, and prolonged inactivity on bone dynamics in the golden hamster, MesocRicetus aUratus. Journal of Morphology 167:43-5 1.
UNDERWOOD, E. J. 198 1. The mineral nutrition of livestock. Second ed. Farnham Royal, Commonwealth Agricultural Bureaux, Slough, England, 180 pp.
WEINMANN, J. P., AND H. SIGHER. 1947. Bone and bones. Fundamentals of bone biology. The C.V. Mosby Company, St. Louis, MO.
ZOODSMA, B. J. 1991. Distribution and behavioral ecology of manatees in southern Georgia. MS. thesis, University of Florida, Gainesville, FL. 202 pp.
Received: 3 May 1994 Accepted: 13 June 1995