Embed Size (px)
Citation preview

Journal ofFish Biology (1991) 39,433-440
Age determination in Pomatoschistus minutus (Pallas) and Pomatoschistus microps (Kreyer) (Pisces: Gobiidae) from the
upper Tagus estuary, Portugal
F. MOREIRA, J. L. COSTA, P. R. ALMEIDA, C. ASSIS AND M. J. COSTA Faculdade de Cigncias de Lisboa, Departamento de Zoologia e Antropologia, Bloco C2.
Camp0 Grande, 1700 Lisboa, Portugal
(Received 28 January 1991, Accepted 1 April 1991)
The use of scale inspection as a means of age-determination in Pomatoschistus microps and Pomatoschistus minutus in the Tagus estuary is confirmed, but annulus formation pattern is different from that described for P. microps in the area of the British Isles (Miller, 1975). In Britain, two annuli are laid down on the scales each year, but in the Tagus, only one annulus is formed. One possible reason for this difference is the fact that in the Tagus the peak breeding season and the lowest water temperatures coincide. Therefore, the two causes of annulus formation coexist at the same time of year.
Population structure in both species is dominated by the 0-group, which appears earlier (April/ May) than reported for other areas. Maximum age seems not to exceed 26 months in P. microps and 32 months in P. minutus.
Key words: Pomatoschistus microps; Pomatoschistus minutus; age determination; Tagus estuary; Portugal.
I. INTRODUCTION
The common goby, Pomatoschistus microps (Krryer), and the sand goby, Pomatoschistus minutus (Pallas), are small gobiid fishes very common in European estuaries (Wiederholm, 1987). In the Tagus estuary, these species are among the most abundant teleosts (Costa, 1980), playing an extremely important role in the estuarine food web (Costa, 1988~).
In the present study, regular monthly sampling of both species over a period of 1 year in the upper Tagus estuary was undertaken to study some aspects of their phenology. The aim of this paper is to test the adequacy of scale use for age-determination in these species.
11. MATERIAL AND METHODS
Samples of P. microps and P. minutus were collected monthly from March 1988 to February 1989, in the estuary of the River Tagus (Fig. 1). This estuary is the largest in western Europe (about 320 km2) (Costa, 19886). Its intertidal area is 41 % of the total area and mean depth is 10.6 m (Bettencourt, 1979).
The main characteristics of the two sampling stations can be seen in Table I and mean monthly temperatures in Fig. 2. All fish were caught at low water with a beam trawl (20 mm mesh in the cod end) and trawl hauls lasted 15 min (constant speed). The common goby and the sand goby were the most numerous fish in the samples; the crustaceans Crangon crangon (Linnaeus) and Carcinus maenas (Linnaeus) were also abundant. All fish were preserved in 70% alcohol immediately after capture and kept in this preservative until examination.
433 0022 - 1 112/91/090433+08 $03.00/0 0 1991 The Fisheries Society ofthe British Isles

434
39"GO'N
38"OO'N
ii
FIG. 1 . Locations of the sampling stations Alcochete and Vila Franca.
Jan Feb Mar Apr May Jun Jul Auq Sep Ocl Nov Dec
FIG. 2. Mean monthly temperatures at the sampling stations. 0, Alcochete; 0, Vila Franca.
In the laboratory, the standard length of all fish was measured to the nearest mm (round down). Each fish was weighed and its sex determined.
The methodology used for ageing the fish was based on Miller (1975). About five scales were taken from the dorsolateral and ventrolateral rows of the caudal peduncule of each individual. After being cleaned of adhering tissue, they were examined in a light microscope (100 x ) and classified into one of the five scale types described by Miller (1975): type a, no

AGE DETERMINATION IN POMA TOSCHISTUS 435
TABLE I. Characteristics of the two sampling stations
Sampling station Alcochete Vila Franca
Mean depth (m) 1.7 3.6 Substrate Mud Muddy-sand Salinity range 8-25 0-14
Temperature range 8-23.4 1 1 4-24 (%o)
("C)
annulus within scale, terminal sclerites wide to very narrow; type b, one annulus, terminal sclerites wide; type c, one annulus, terminal sclerites very narrow or two annuli with terminal sclerites narrow to moderately wide; type d, two annuli, terminal sclerites very narrow; type e, three annuli, terminal sclerites moderately wide.
The width of the four terminal sclerites on the anterior edge of an adequate scale of each individual was determined in arbitrary units (1 unit = 0.01 1 mm).
For statistical analysis parametric tests were used. Therefore terminal sclerite width were log-transformed when appropriate to correct for lack of normality and heteroscedasticity (Sokal& Rohlf, 1969).
111. RESULTS LENGTH-FREQUENCY DISTRIBUTION
The length-frequency distribution for each monthly sample of P. microps and P. minutus shows that the first demersal young fish are found in catches from April, for the former species, and from May, for the latter, with a modal length of 22 and 36 mm, respectively.
Comparing these results with those of other authors (Healey, 1971,1972; Fonds, 1973; Miller, 1975; Hesthagen, 1979; Fouda & Miller, 1981; Claridge et al., 1985; Nellbring, 1985; Hamerlynck et al., 1986; Doornbos & Twisk, 1987), we can see that the young-of-the-year appear earlier in the Tagus estuary, as compared to sites located at higher latitudes.
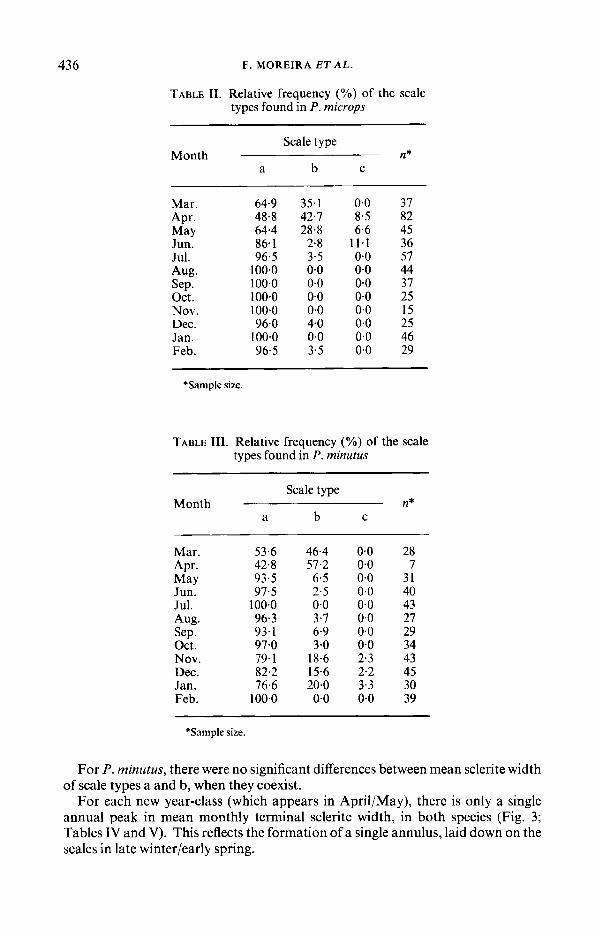
SCALE-READING Only a, b and c scale types were found (Tables I1 and 111). Scale type a was
generally dominant throughout the year, both for P. microps and P. minutus whereas scale type b had its maximum frequency from March to May. Scale type c was rare.
Before calculating the variation of monthly mean terminal sclerite width, we compared this measure in the main scale types (a and b) for the months in which they occurred together.
For P. microps, April was the only month for which the mean terminal sclerite width was significantly different for scale types a and b (t-test, t= 3.35, d.f. = 57, P<O.Ol). This could be due to large sclerite width of the new year-class scales contrasting with narrow sclerites in adults, due to annulus formation. For the other months, no significant differences were found and mean terminal sclerite width are based on pooled mean values.
436 F. MOREIRA ET A L .
TABLE 11. Relative frequency (YO) of the scale types found in P. microps
Scale type
a b C Month n*
Mar. 64.9 35.1 0.0 37 Apr. 48-8 42.7 8.5 82 May 64.4 28.8 6.6 45 Jun. 86.1 2.8 11.1 36 Jul. 96.5 3.5 0.0 57 Aug. 100.0 0.0 0-0 44 Sep. 100.0 0.0 0.0 37 Oct. 100.0 0.0 0.0 25 Nov. 100.0 0.0 0.0 15 Dec. 96.0 4-0 0.0 25 Jan. 100.0 0.0 0.0 46 Feb. 96.5 3.5 0.0 29
*Sample size.
TABLE 111. Relative frequency (%) of the scale types found in P. minutus
Scale type
a b C Month n*
Mar. Apr. May Jun. Jul. Aug. Sep. Oct. Nov. Dec. Jan. Feb.
53.6 42.8 93.5 97.5
100.0 96.3 93.1 97.0 79.1 82.2 76.6
100.0
46.4 0.0 28 57.2 0.0 7 6.5 0.0 31 2.5 0.0 40 0.0 0.0 43 3.7 0.0 27 6.9 0.0 29 3.0 0.0 34
18.6 2.3 43 15-6 2.2 45 20.0 3.3 30 0-0 0.0 39
~~~
*Sample size.
For P. minutus, there were no significant differences between mean sclerite width of scale types a and b, when they coexist.
For each new year-class (which appears in April/May), there is only a single annual peak in mean monthly terminal sclerite width, in both species (Fig. 3; Tables 1V and V). This reflects the formation of a single annulus, laid down on the scales in late winter/early spring.

AGE DETERMINATION IN POMA TOSCHISTUS 431
Mar A p r M a y J u n Jul Auq Sep Oct Nov Dec J a n Feb
FIG. 3. Mean terminal sclerite widths (MTSW) for P. mrcrops(0) and P. minutus (m) (see Tables IV and V).
TABLE IV. Logarithmic transformations of mean terminal sclerite widths (MTSW) in P.
microps (arbitrary units of 0.01 1 mm)
Month MTSW s . E . ~ n t
Mar. Apr. a$ Apr. b$ May Jun. Jul. Aug . Sep. Oct. Nov. Dec. Jan. Feb.
0.7 13 0.859 0.753 0.763 0-794 0.863 0.842 0.789 0.761 0.749 0.740 0.720 0.735
0.022 0.023 0.020 0.024 0.0 19 0.0 13 0.0 12 0.0 13 0.017 0.0 15 0.0 16 0.017 0.02 1
25 33*** 26 40* *(a)$ 31 55** 50 35** 25 15 24 29 27
Significance of difference from previous month:
hE., standard error; n, sample size. SScale types. §Scale type a from April.
**P< 0.01; ***P< 0.001.
In P . microps we could expect annulus formation if we consider that in April mean terminal sclerite width of scale type a is bigger than in May. Nevertheless, this is not reflected in scales (otherwise, scale type b would dominate in the rest of the year).
LONGEVITY Considering that young-of-the-year of P. microps and P. minutus appear in April
and May, respectively, and that the latest date of capture of individuals with scale

43 8 F. MOREIRA E T A L .
TABLE V. Logarithmic transformations of mean terminal sclerite widths (MTSW) in P. minutus
(arbitrary units of 0.01 1 mm) ~~ ~ ~
Month MTSW S.E. n
Mar. 0.367 0,021 23 Apr. 0,757 0.038 7** May 0,923 0.022 27** Jun. 0.862 0.012 33** Jul. 0.764 0-012 41*** Aug. 0.733 0.016 30 Sep. 0.7 16 0.013 29 Oct. 0.705 0,013 34 Nov. 0.679 0.014 43 Dec. 0.628 0.015 44t Jan. 0.632 0.017 30 Feb. 0.610 0.014 39
Significance of difference from previous month: **P<O.OI; ***P<O.OOI. ?Approximate i-test.
type c was June for P . microps and January for P. minutus, the oldest individual P . microps and P . minutus were 26 and 32 months old respectively. This finding for P. microps is similar to Miller’s (1975), although age determination followed different patterns.
MAXlMUM LENGTH For P. microps, the largest female measured 50 mm and the largest male, 46 mm. For P. minutus, both the largest male and female measured 72 mm.
IV. DISCUSSION
The time of appearance of a new year-class and, consequently, the peak breeding season, varies with latitude, suggesting that temperature and photoperiod directly affect the onset of the breeding season in P. microps and P. minutus. Because of this, the young-of-the-year appear earlier in the Tagus estuary, as compared to sites at higher latitudes. The population structure is dominated by the 0-group.
In contrast to Miller’s (1 975) findings in Britain for P . microps, in the Tagus there is only one annual annulus laid down on the scales of both Pomatoschistus species. What could be the cause for this different pattern of annulus formation?
Miller (1975) described the formation of two separate annuli on the scales of P . microps within a period of 6 months. Every year, in March/April, a spring annulus was laid down and towards the end of May, a summer annulus was formed. According to this author, the spring annulus was due to resumption of growth (during the winter, body growth is negligible) and the summer annulus corresponded to growth retardation during the peak breeding season.
In the Tagus estuary, the high summer temperatures (Fig. 2) which cause a decrease in egg survival (Fonds & Van Buurt, 1974) are probably the main reason

AGE D ETERMIN A TI ON I N POMA TOSCHIS TUS 439
for a shift (in relation to other sites) of the peak breeding season of both P. microps and P . minutus towards the cooler temperatures of late winter and early spring (F. Moreira et al., unpublished). Consequently, both reasons for annulus formation (low temperatures and high energetic costs, due to breeding season) occur simul- taneously probably causing the formation of a single annulus in the scales of these species.
It is logical that the relative frequency of scale types b and c should increase in March/April, after annulus formation. Their decrease after was probably due to increasing importance of the new year-class, high mortality of breeding adults of spawning migration to other areas.
If temperature is as important as we suspect, it is predictable that in a latitudinal gradient from north to south, distinction between the spring and summer annuli would become increasingly difficult, as the two rings would come closer, eventually overlapping.
This emphasizes the need for caution in applying specific age determination methods in different geographical areas in these gobies.
We thank Junta Nacional de InvestigaCIo Cientifica e Tecnolbgica for financial support.
References Bettencourt, A. (1979). Estuario do Tejo. Documentopreliminar. Drena: Hidroprojecto. Claridge, P. N., Hardisty, M. W., Potter, I. C. & Williams, C. V. (1985). Abundance, life
history and ligulosis in the Gobies (Teleostei) of the inner Severn estuary. Journal of the Marine Biological Association of the United Kingdom 65,95 1-968.
Costa, M. J. (1980). Fishes of Tejo estuary (Community structure, abundance, and diversity). Arqujvos do Museu Bocage ( 2 ) 7(17), 291-316.
Costa, M. J. (1988~). Ecologie alimentaire des poissons de l’estuaire du Tage. Cybium 12,
Costa, M. J. (19883). The Tagus and Mira estuaries (Portugal) and their role as spawning and nursery areas. Journal of Fish Biology 33 (Suppl. A), 249-250.
Doornbos, G. & Twisk, F. (1987). Density, growth and annual food consumption of gobiid fish in the saline lake Grevelingen, the Netherlands. Netherlands Journal of Sea Research 18,457-479.
Fonds, M. (1973). Sand gobies in the Dutch Wadden Sea (Pomatoschistus, Gobiidae, Pisces). Netherlands Journal of Sea Research 6,417-478.
Fonds, M. & Van Buurt, G. (1974). The influence of temperature and salinity on develop- ment and survival of Goby eggs (Pisces, Gobiidae). Hydrobiological Bulletin 8, 110-1 16.
Fouda, M. M. & Miller, P. J. (1981). Age and growth of the common goby, Pomatoschistus microps, on the south coast of England. Estuarine Coastal and Shelf Science 12,
Hamerlynck, O., Help, C. & Redant, F. (1986). Life history, food consumption and resource partitioning in two sympatric gobies Pomatoschistus minutus and Pomatoschistus lozanoi in Belgian coastal waters. International Council for the Exploration of the Sea C.M. 14 (mimeo).
Healey, M.C. (1 97 I). The distribution and abundance of sand gobies, Gobius minutus in the Ythan estuary. Journal of Zoology London 163,177-229.
Healey, M. C . (1972). On the population ecology of the common goby in the Ythan estuary. Journal of Natural History 6, 133-145.
Hesthagen, I . H. (1979). Migration, breeding and growth in Pomatoschistus minutus (Pallas) (Pisces, Gobiidae) in Oslofjorden, Norway. Sarsia 63, 17-26.
Miller, P. J. (1 975). Age-structure and life-span in the common goby, Pomatoschistus microps. Journal of Zoology, London 177,425448.
30 1-320.
12 1-1 29.

440 F. MOREIRA ET AL.
Nellbring, S. (1985). Abundance, biomass and seasonal variation of fish on shallow soft bottoms in the Ask0 area, Northern Baltic Proper. Sarsia 70,217-225.
Sokal, R. R. & Rohlf, F. J. (1969). Biometry. The Principles and Practice of Statistics in Biological Research. San Francisco: W. H. Freeman.
Wiederholm, A. M. (1987). Distribution of Pomatoschistus minutus and P . microps (Gobiidae, Pisces) in the Bothnian sea: importance of salinity and temperature. Memoranda Societatis pro Fauna et Flora Fennica 6 3 , 5 6 6 2 .













